Introduction
The sialyl-Lewis oligosaccharide antigens
(SLea and SLex) are blood group related
antigens which are important in inflammation by initiating the
leukocyte-endothelium interaction (1,2).
To date, it has been found that sialyl-Lewis antigens are
involvedin severalmolecular interactions, including
differentiation, development and malignancy (3-5),
and their expression is markedly enhanced in several types of
carcinoma cells (6,7). It has also been reported that the
expression of sialyl-Lewis oligosaccharide antigens on tumor cells
is closely associated with tumor progression, and statistical
relevance has been confirmed between the postoperative prognosis of
tumor patients and the degree of sialyl-Lewis oligosaccharide
antigen expression on tumor tissues (8,9).
Therefore, sialyl-Lewis oligosaccharide antigens are widely
considered as cancer markers. The present study determined the
expression of three sialyl-Lewis oligosaccharide antigens,
SLea, SLex, and dimeric SLex
(SDLex) on the cell surface of the MHCC97 human
hepatocarcinoma (HCC) cell line. The expression of SLex
was higher in MHCC97 cells compared with its expression in normal
liver cells. The expression of SDLex was also high,
although did not differ significantly between the normal liver
cells and MHCC97 cells. The expression of SLea was only
detected in trace quantities. On using a monoclonal antibody of
sialyl-Lewis antigens, the results indicated that the
anti-SLex antibody KM93 significantly inhibited MHCC97
cell proliferation.
The synthesis of sialyl-Lewis antigens is regulated
by a set of glycosyltransferases. Fucosylation is the final step in
the process of sialyl-Lewis antigen synthesis and it is catalyzed
by α1,3/1,4 fucosyltransferases (α1,3/1,4 FUTs). The α1,3/1,4FUTs
catalyze the connection of fucose in a (1,3)
and a (1,4) linkage to the sialylated precursors
(10-15). Currently, several human α(1,3)
FUT genes (αFUT-3, 4, 5, 6, 7 and 9) have been cloned, identified
and characterized (15-24). Each enzyme has a unique substrate
and tissue specificity (25).
FUT4, FUT9, FUT7 and FUT3 efficiently fucosylate sialylated
acceptors, whereas FUT4 and FUT9 preferentially fucosylate
non-sialylated neutral acceptors (20,24). α1,3/1,4FUT-3 is the only αFUT with
both α1,3 and α1,4 fucosylation activity, leading to the generation
of α1,3-fucosyl-containing Lex and SLex, and
its isomer α1,4-fucosyl-containing SLea, respectively
(7). By contrast, α1,3FUT7 only
catalyzes the synthesis of SLex (NeuAca2/3Galb1/4
(Fuca1-3) GlcNAcb1/3Galb1/R) by adding the distal fucose residue to
α2,3-sialylated lactosamine (9).
FUT7 is mainly expressed in tumor cells, endothelial cells and
leukocytes. There is a rigid acceptor specificity for FUT7, which
can only accept the distal N-acetylglucosamine on α2,3 sialylated
lactosamines to form SLex antigen. Therefore, FUT7 is
the key enzyme of SLex synthesis (9,22,23). It has been widely reported that
the expression levels of α1,3/1,4FUTs are positively correlated
with tumor progression and negatively correlated with patient
prognosis (8,9). For example, tumor growth was
significantly increased following transfection of the FUT3 gene
into PC-3 prostate cancer cells (26). Additionally, cell proliferation
was significantly inhibited following gtransfection of FUT3/6
antisense RNA into colon cancer cells (27). Until now, the role of FUT7 in
cancer cell proliferation and growth has not been fully
elucidated.
In the present study, the effects of the knockdown
of FUT7 on the expression of SLex and human HCC cell
proliferation were examined. The effects of FUT7 small interfering
(si)RNAs on the expression and activation of signaling molecules
were also investigated. It was found that the knockdown of FUT7
decreased the expression of SLex, inhibited the
trans-location and phosphorylation of phospholipase Cγ (PLCγ), and
arrested cell cycle progression, which subsequently led to the
suppression of MHCC97 HCC cell proliferation. These findings
indicated that SLex and FUT7 may be important in human
HCC cell proliferation. Inhibition of the SLex
oligosaccharide antigen may serve as a novel therapeutic method to
treat tumors that express SLex glycoconjugates. In
addition, FUT7 maybe a potential target and FUT7 RNA interference
(RNAi) may be developed as a novel therapeutic approach for
SLex-positive cancer therapy.
Materials and methods
Cell culture and inhibitors
treatment
The THLE-2 normal human liver cell line and HT29
human colorectal adenocarcinoma cell line were purchased from the
American Type Culture Collection (Manassas, VA, USA). The MHCC97
human HCC cell line was purchased from Shanghai Institute of
Biochemistry and Cell Biology, Chinese Academy of Sciences
(Shanghai, China). The THLE-2 cell line was grown in LONZA BEGM
medium (Lonza/Clonetics Corporation, Walkersville, MD, USA). The
MHCC97 cell line was cultured in Dulbecco's modified Eagle's medium
(DMEM; Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% FCS (Invitrogen, Thermo Fisher
Scientific, Inc.). The cells were grown at 37°C in a humidified
incubator under 5% CO2 and passaged when 70-80%
confluent using 0.25% (w/v) trypsin solution in 0.04% (w/v) EDTA.
At 1 h following passage of MHCC97 cells, PLCγ inhibitor (U73122;
10.5 mM; Merck KGaA, Darmstadt, Germany), protein kinase A (PKA)
inhibitor (KT5720; 280 nM; Sigma-Aldrich; Merck KGaA),
phosphoinositide 3-kinase (PI3K) inhibitor (LY294002; 16.5 mM;
Sigma-Aldrich; Merck KGaA), extracellular signal-regulated kinase
(Erk) inhibitor (ErkI; 50 mM; Merck KGaA) and CDC25 inhibitor
(CDC25 inhibitor II; 1.05 mM; Merck KGaA) were added into the
culture medium and the cells were cultured for a further 24 h. Cell
survival was detected through MTT assay.
Detection of Lewis antigen with flow
cytometric analysis
Cell surface Lewis antigen expression was detected
by measuring the indirect immunofluorescence intensity using the
FACS Calibur system (BD Biosciences, San Jose, CA, USA). Cell Quest
Pro software (version 5.1; BD Biosciences) was used for cell
acquisition and analysis. The cells were detached with 2 mM EDTA,
washed twice with cold PBS and then prepared as a single cell
suspension. A suspension of 106 cells was then incubated
with mouse anti-SLex antibody (KM93, cat. no. 11-006,
1:100, GlycoTech, Gaithersburg, MD, USA), anti-SDLex
antibody (FH6, cat. no. 11-007, 1:100, GlycoTech) and
anti-SLea antibody (CA19-9, cat. no. 11-009, 1:100,
GlycoTech) at 4°C for 1 h. Chilled PBS was used to wash the cells
three times to remove unbound antibodies. The cells were then
incubated with R-phycoerythin or peridinin chlorophyll protein
(R-PE or Per-CP)-conjugated rabbit anti-mouse IgM (cat. no. 562033
and 553409, 1:200, BD Biosciences) at 4°C for 30 min in the dark.
Chilled PBS was used to wash and remove unbound secondary
antibodies. The cells were then fixed using 1% formaldehyde (Sigma;
EMD Millipore, Billerica, MA, USA) for 30 min in the dark. A total
of 1×104 cells from each sample tube were acquired for
analysis using FACS Calibur. Cells incubated with secondary
antibody only and unstained cells were used as controls.
Reverse-transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the cells were obtained using ab
RNeasy Mini kit (Qiagen GmbH, Hilden, Germany) following the
manufacturer's protocol. Subsequently, 1 µg of total RNA was
mixed with SYBR-Green PCR Master mix reagents of the PrimeScript RT
Reagent kit (Takara Biotechnology Co., Ltd., Dalian, China) to
synthesize the complementary DNA (cDNA) in the 7900HT real time PCR
system (Applied Bio systems, Thermo Fisher Scientific, Inc.)
according to the standard quantitative PCR protocol (25). Briefly, the RT-qPCR procedure was
performed in a 20-µl reaction volume, including primers
(0.25 µl, 10 pmol/l), 2X Mix SYBR-Green I (10 µl;
Promega, Madison, WI, USA), template DNA (1 µl) and sterile
water. The PCR steps constituted an initial denaturation step (95°C
for 2 min), 50 cycles of 95°C for 15 sec and then 60°C for 45 sec.
The fold change of target gene expression was calculated according
to the following formula: Fold change = 22[[Cq (control) gene X-Cq
(control) actin] [Cq (activated) gene X-Cq (activated) actin]]
(28). The following primer
sequences were used: Human FUT7, forward 5′-CCA CGA TCA CCA TCC
TTG-3′ and reverse 5′-AGG CTT CGG TTG GCA CTC-3′; human
FUT6, forward 5′-CAT TTC TGC TGC CTC AGG-3′ and reverse
5′-GGG CAA GTC AGG CAA CTC-3′; human glyceraldehyde-3-phosphate
dehydrogenase (GAPDH), forward 5′-GAA GGT GAA GGT CGG AGT C-3′ and
reverse 5′-GAA GAT GGT GAT GGG ATT TC-3′. The relative mRNA
expression levels of FUT7 and FUT6 were normalized to the
endogenous mRNA expression of GAPDH.
Western blot analysis
RIPA buffer (Sigma; EMD Millipore) with protease
inhibitor (Sigma; EMD Millipore) was used to lyse cells (26). The protein was collected, and a
BCA Pierce Assay (Thermo Fisher Scientific, Inc.) was used for the
quantification of protein concentration. Subsequently, 50 µg
of protein from each sample was denatured and resolved on a 10%
SDS-PAGE gradient gel (EMD Millipore), and then electro-blotted
onto a PVDF nitrocellulose membrane (EMD Millipore). The PVDF
membranes were then incubated with 5% non-fat milk for 1 h at room
temperature, and then incubated with anti-FUT7 (1:1,000, cat. no.
MAB64091), anti-PLCγ1 (1:500, cat. no. MAB8137) and
anti-phosphorylated PLCγ1 (1:500, cat. no. MAB74541) which were
purchased from Bio-Techne China (Shanghai, China), anti-FUT6
(1:1,000, cat. no. NBP1-57936; Novus Biologicals, LLC, Littleton,
CO, USA), or anti-β-actin antibody (1:1,000, cat. no. MAB8929;
Bio-Techne China) at 4°C overnight. Following washing with TBST,
the membrane was incubated with HRP-conjugated secondary antibodies
(1:3,000, cat. no. HAF008; Bio-Techne China) for 1.5 h at room
temperature. Finally, the signals were developed by enhanced
chemiluminescence (Pierce, Thermo Fisher Scientific, Inc.). Images
of the results were captured and the images were scanned. The
optical density of each protein band was quantified by a scanning
densitometer and Quantity One software, version 4.4.1 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Each lane of protein band
density was normalized with corresponding β-actin density.
The cytosolic protein was isolated from particulate-
conjugated protein using a digitonin separation method (29). The cells were collected and
resuspended in 1 ml saline solution (1 mM EDTA, 150 mMNaCl, 1 mM
PMSF, 2 mM EGTA, 1 µg/ml aprotinin, 10 µg/ml
leupeptin, and 100 µg/ml digitonin) with occasional
agitation for 10 min. The cells were then centrifuged at 13,000 × g
for 5 min at 4°C and the resulting supernatant contained the
cytosolic proteins. The cell pellet was dissolved in 1 ml lysis
buffer (pH 7.4, 1 mM EDTA, 10 mM PBS, 1% Triton X-100, 2 mM EGTA, 1
mM PMSF, 0.1% SDS, 1 µg/ml aprotinin, and 10 µg/ml
leupeptin) and contained the membrane protein
(particulate-conjugated proteins). Subsequently, 80 µg of
protein was separated by SDS-PAGE and transferred onto a PVDF
membrane. The expression levels of PLCγ1 and phosphorylated PLCγ1
were detected by western blot analysis, as described above.
Knockdown of FUT7 in MHCC97 cells by
RNAi
The expression of FUT7 in MHCC97 cells was silenced
using specific siRNAs (Silencer siRNA transfection, Ambion, Thermo
Fisher Scientific, Inc.). Scramble siRNA (siNC; Silencer siRNA
trans-fection, Ambion, Thermo Fisher Scientific, Inc.) was used to
confirm the specificity of FUT7 siRNAs. Untransfected cells were
used as acontrol. The siRNA transfection into MHCC97 cells was
performed following the manufacturer's protocol. A total of
1×105 cells/ml of MHCC97 cells were re-suspended in
DMEM. The transfection complexes were prepared by mixing RNAi MAX
Lipofectamine transfection agent (Ambion; Thermo Fisher Scientific,
Inc.) and siRNA (20 nM) in DMEM. The MHCC97 cells and transfection
complexes were mixed and then incubated for 24 h at 37°C in 6-well
(2×105 cells/well) plates (Nalge Nunc International,
Penfield, NY, USA). The cells were collected at 48 h for RNA
analysis and 72 h for protein analysis. The knockdown of FUT7 was
confirmed by RT-qPCR and western blot analyses.
Cell survival MTT assay
To investigate the effect of FUT7 silencing on cell
proliferation and viability, MTT was added to the MHCC97 cells at
0, 24, 48, 72, 96 and 120 h following transfection with FUT7 siRNA.
In detail, 5×104 cells were seeded into 96-well plates
and, the following day, FUT7 was knocked down as described above.
MTT (20 µl, 5 mg/ml) was added into each well at different
time points. Following culture for 4 h at 37°C the DMEM was gently
aspirated and 100 µl of dimethyl sulfoxide was added into
each well. The absorbance of formazan at a 490 nm wavelength was
measured using a Bio-Rad Microplate Reader 550 (Bio-Rad
Laboratories, Inc.) and a cell growth curve was plotted.
Analysis of cell cycle
Ice-cold PBS was used to wash the collected control
cells and transfected cells, and 1×106 cells were fixed
in 70% ethanol for 24 h at -20°C. The cells were then washed twice
with chilled PBS, and stained withpropidium iodide (PI; 20
µg/ml, Sigma; EMD Millipore) including RNase (100
µg/ml) for 30 min at room temperature. A BD FACScan flow
cytometer was used to analyze the DNA content of cell cycle phases.
A total of 2×104 cells per sample were collected and
analyzed using BD FACSDiva™ analysis software (version 6.2; BD
Biosciences).
Statistical analysis
Statistical analysis was performed using SPSS 20.0
software (IBM SPSS, Armonk, NY, USA). All values are expressed as
the mean ± standard error of the mean and are representative of at
least three independent experiments. The data were analyzed using
one-way analysis of variance. When the results were significant,
post hoc testing of differences between each group was performed
using Bonferroni's correction. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression of sialyl-Lewis
oligosaccharide antigens and FUTs in the MHCC97 human HCC cell
line
The expression of three sialyl-Lewis antigens,
SLea, SDLex and SLex, were
determined and the results are shown in Fig. 1A and B. The expression of
SLex was significantly increased in the MHCC97 human HCC
cells compared with that in THLE-2 normal human liver cells. The
expression of SDLex was high, however, no significant
difference was observed between the MHCC97 cells and THLE-2 cells.
The expression of SLea was absent or negligible. The
high expression of SLea in the HT29 cells confirmed the
high affinity of the anti-Slea antibody (Fig. 1C). RT-qPCR and western blot
methods were used to detect the mRNA and protein levels of FUT7 and
FUT6, respectively. The mRNA and protein expression levels of FUT7
were high in the MHCC97 cells compared with expression levels in
the THLE-2 normal liver cells. FUT6 was also expressed at high
levels, however, no significant difference was found between the
MHCC97 cells and THLE-2 cells (Fig.
1D and E). No FUT3 was detected.
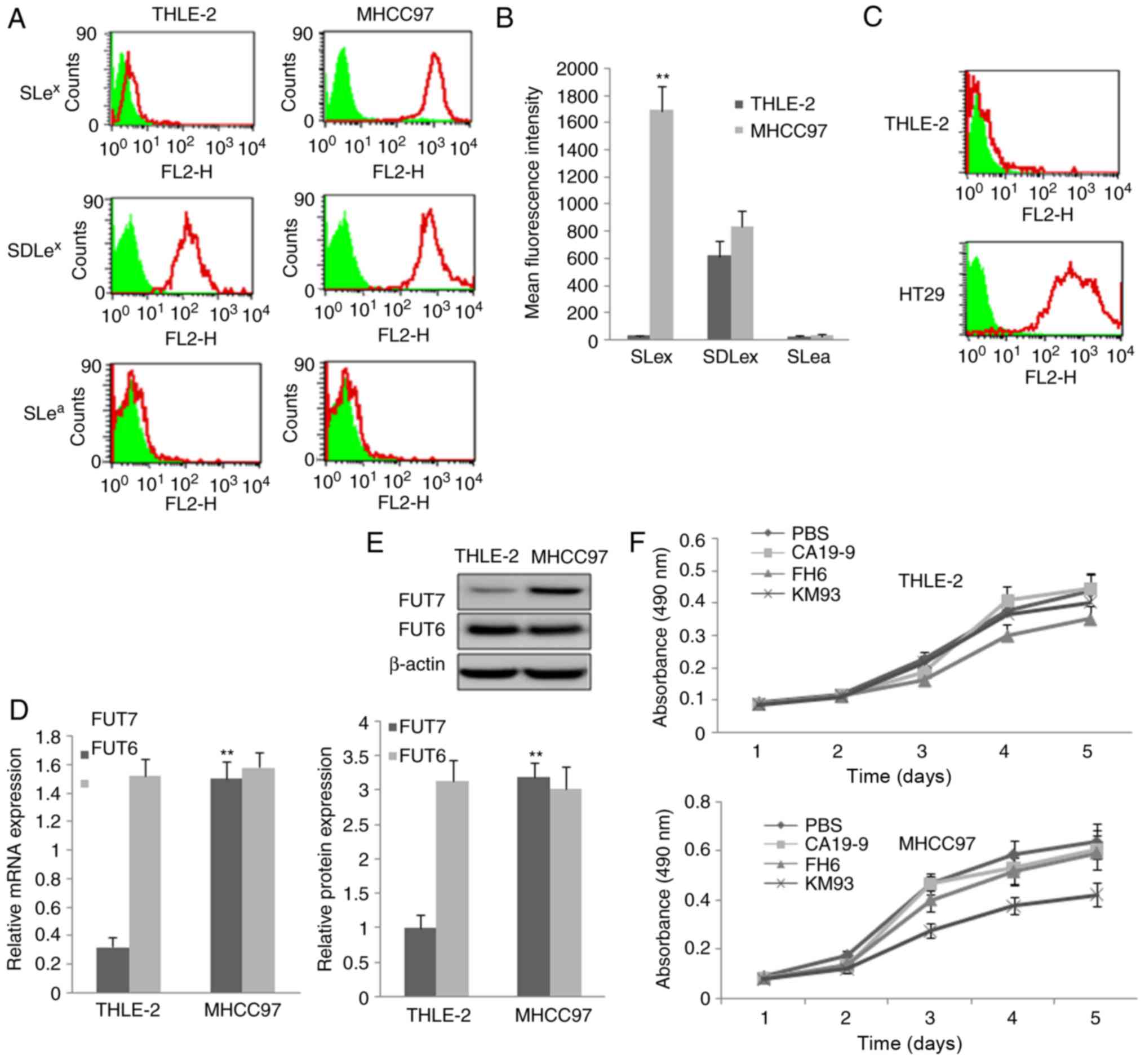 | Figure 1Expression of three sialyl-Lewis
oligosaccharides and relative FUTs and the inhibition effect of
monoclonal antibody on cell proliferation. (A) MHCC97 human
hepatocellular carcinoma cells and THLE-2 normal liver cells were
assayed by flow cytometry using three monoclonal antibodies
targeting sialyl-Lewis antigens. PBS, cells treated with PBS and
without antibodies as a control; KM93, cells treated with mAb
against SLex; FH6, cells treated with mAb against
SDLex; CA19-9, cells treated with mAb against
SLea. In the flow cytometry histograms, the areas in
green show the number of unstained cells and the areas outlined in
red represent cells binding to mAb. (B) Quantitative analysis of
the expression of sialyl-Lewis oligosaccharides by calculation of
mean fluorescence intensity. THLE-2 cells were used as a control.
(C) THLE-2 cells and HT29 cells were assayed by flow cytometry
using monoclonal antibody against SLea antigen to
confirm the high affinity of the SLea antibody. (D) mRNA
expression of FUT7 and FUT6 were detected by reverse
transcription-quantitative polymerase chain reaction analysis. The
relative gene mRNA level was normalized to the respective
glyceraldehyde-3-phosphate dehydrogenase level. THLE-2 cells were
used asa control. (E) Protein expression levels of FUT7 and FUT6
protein were evaluated by western blot analysis (above) and
quantitative analysis of relative protein levels was performed
(below). THLE-2 cells were used as a control.
**P<0.01 (n=3), vs. THLE-2 Control. (F) Effect of
monoclonal antibodies on the proliferation of THLE-2 cells and
MHCC97 cells. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H
tetrazolium bromide assay showed that MHCC97 cell proliferation was
significantly suppressed by KM93, a mAb against SLex. FUT,
fucosyltransferase; SLex, sialyl-Lewis X,
SLea, sialyl-Lewis a; SDLe X, dimeric SLe; mAb,
monoclonal antibody. |
Inhibition of SLex by KM93
monoclonal antibody inhibits MHCC97 cell proliferation
Different monoclonal antibodies of sialyl-Lewis
antigens were added to inhibit the sialyl-Lewis antigens on the
cell surface of MHCC97 cells and normal liver cells, and cell
growth curves were plotted using the data obtained from the MTT
method. It was found that the anti-SLex monoclonal
antibody KM93 significantly suppressed MHCC97 cell proliferation,
but not that of normal liver cells. The anti-SDLex
antibody FH6 had a minimalsuppression effect, which was not
statistically significant. The anti-SLea antibody CA19-9
did not suppress MHCC97 cell proliferation (Fig. 1F).
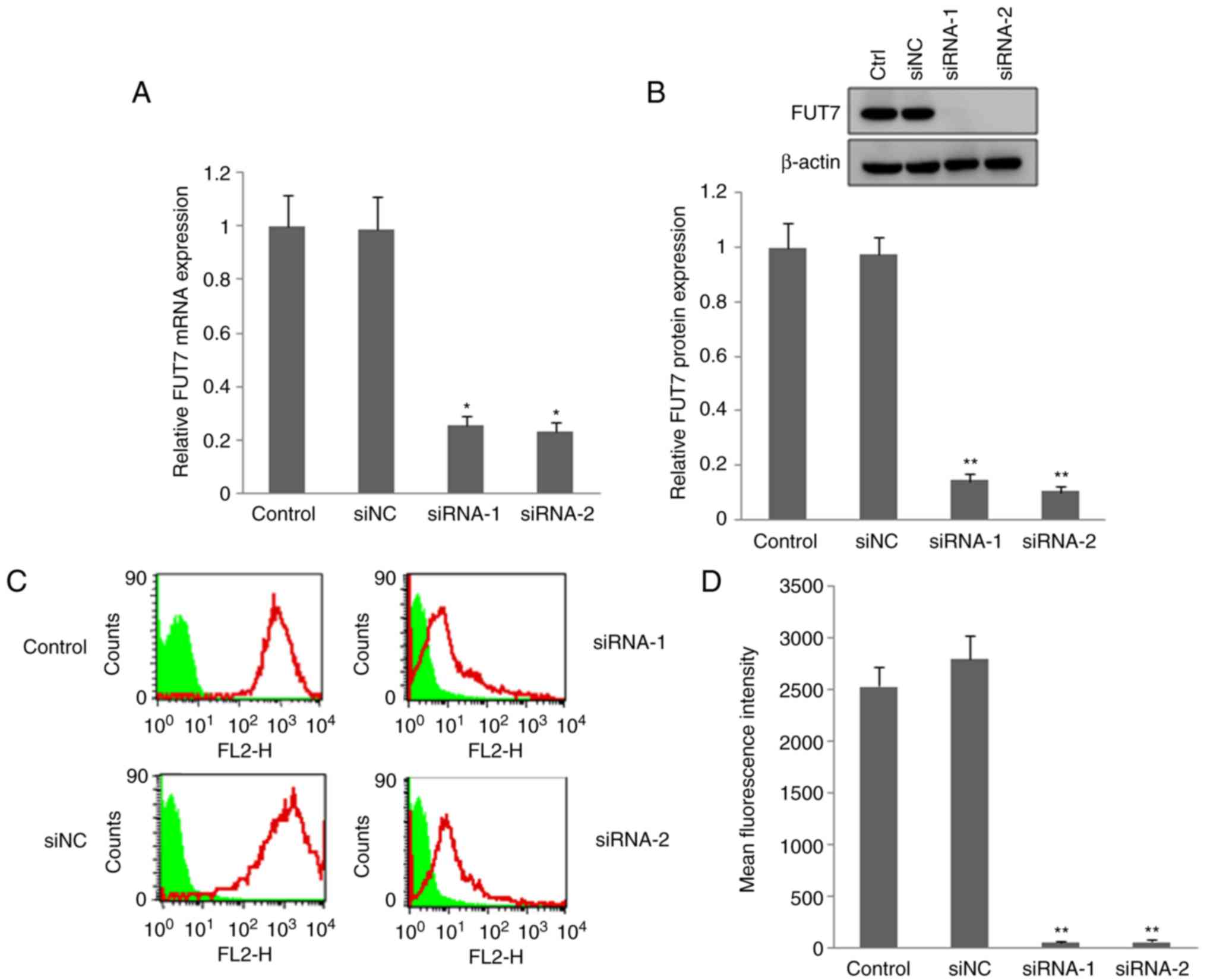
FUT7 siRNAs transfection reduced the
expression of FUT7 and SLex
RT-qPCR, western blot and flow cytometry methods
were used to investigate the effect of FUT7 RNAi on the expression
of FUT7 and SLex. The RT-qPCR and western blot results
showed that the mRNA and protein levels of FUT7 (Fig. 2A and B) were markedly lower in the
FUT7 siRNA-transfected MHCC97 cells, compared with those in the
untransfected wild-type cells (control). The FUT7 siRNAs
effectively inhibited the expression of FUT7. The inhibitory effect
of FUT7 siRNA on the expression of SLex was also
investigated by flow cytometry. The fluorescence intensity of
SLex in the utransfected control cells was more marked
than that in the FUT7-knockdown cells (Fig. 2C and D). siNC-transfected cells
were used to confirm the specificity of FUT7 siRNA. No significant
difference in the expression of FUT7 or SLex was found
between the siNC cells and untransfected control cells. The above
results suggested that siRNAs of FUT7 suppressed the expression of
FUT7 and markedly inhibited the synthesis of SLex
oligosaccharide antigen.
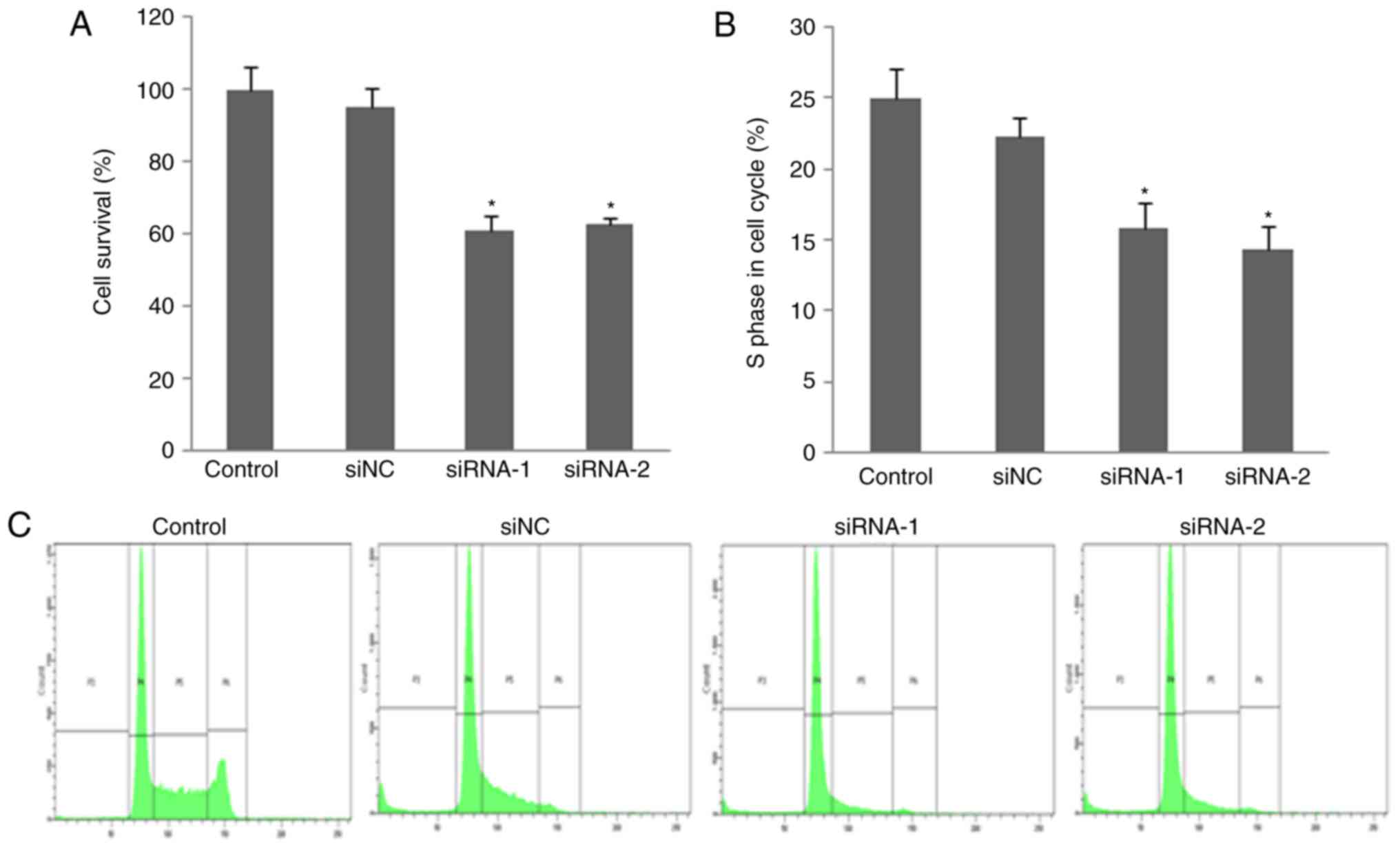
Knockdown of FUT7 inhibits human HCC cell
proliferation
An MTT assay was used to investigate the impact of
FUT7 knockdown in HCC cell proliferation. The number of
proliferated cells was measured by detecting the absorbance at a
wavelength of 490 nm. The MTT assay showed that cell proliferation
of the FUT7-knockdown cells was suppressed compared with that of
control cells. No significant difference was found between the cell
proliferation rate of siNC cells and control cells (Fig. 3A). This indicated that the
silencing of FUT7 may significantly suppress HCC cell
proliferation.
FUT7 knockdown decreases HCC cell
distribution into the S-phase
To further investigate the suppression of cell
proliferation in FUT7-knockdown cells, cell cycle distribution was
analyzed by flow cytometry. As shown in Fig. 3B and C, flow cytometry with
PI-stained cells showed that untransfected control cells and siNC
cells were presented in the G0/G1 (58.78±6.45 and 60.52±6.17%), S
(25.20±2.17 and 22.18±2.87%) and G2/M (2.97±0.60 and 3.10±0.77%)
phases. In the FUT7 siRNA-transfected MHCC97 cells, the S-phase
fraction was lower (16.40±2.10% for FUT7 siRNA-1 and 13.27±2.75%
for FUT7 siRNA-2 cells, P<0.05) and the G0/G1 fraction was
higher (69.62±6.84% for FUT7 siRNA-1 and 67.72±6.53% for FUT7
siRNA-2 cells, respectively, P<0.05). This indicated that the
fractions of cells in the S-phase of cell cycle, which represents a
population of dividing cells, decreased following the knockdown of
FUT7. A concomitant increase was observed in the number of cells at
the G0/G1 phases. These results showed that FUT7 knockdown
suppressed cell proliferation by inhibiting cell cycle progression
into the S-phase. These results further emphasized the importance
of the expression of FUT7 on cell proliferation.
Effects of inhibiting PKA, PI3K/AKT and
PLCγ/Erk signaling pathways on the expression of FUT and MHCC97
cell proliferation
To understand the mechanisms involved, the present
study examined the effects of inhibitors of signaling pathways on
the mRNA levels of FUT7. The RT-qPCR results showed that, in MHCC97
cells, PLCγ inhibitor, PKA inhibitor and PI3K inhibitor
significantly reduced the mRNA levels of FUT7. By contrast, the Erk
inhibitor and CDC25 inhibitor had no effect on the mRNA level of
FUT7 (Fig. 4A). The MTT results
showed that inhibitors of PLCγ and Erk reduced cell survival in
normal liver cells. CDC25, PI3K, PLCγ and Erk inhibitors decreased
the proliferation of MHCC97 cells, with the most marked effect
observed with PLCγ inhibitor (Fig.
4B). These results indicated that the PLCγ inhibitor
significantly reduced the mRNA level of FUT7, and inhibited MHCC97
HCC cell proliferation and survival.
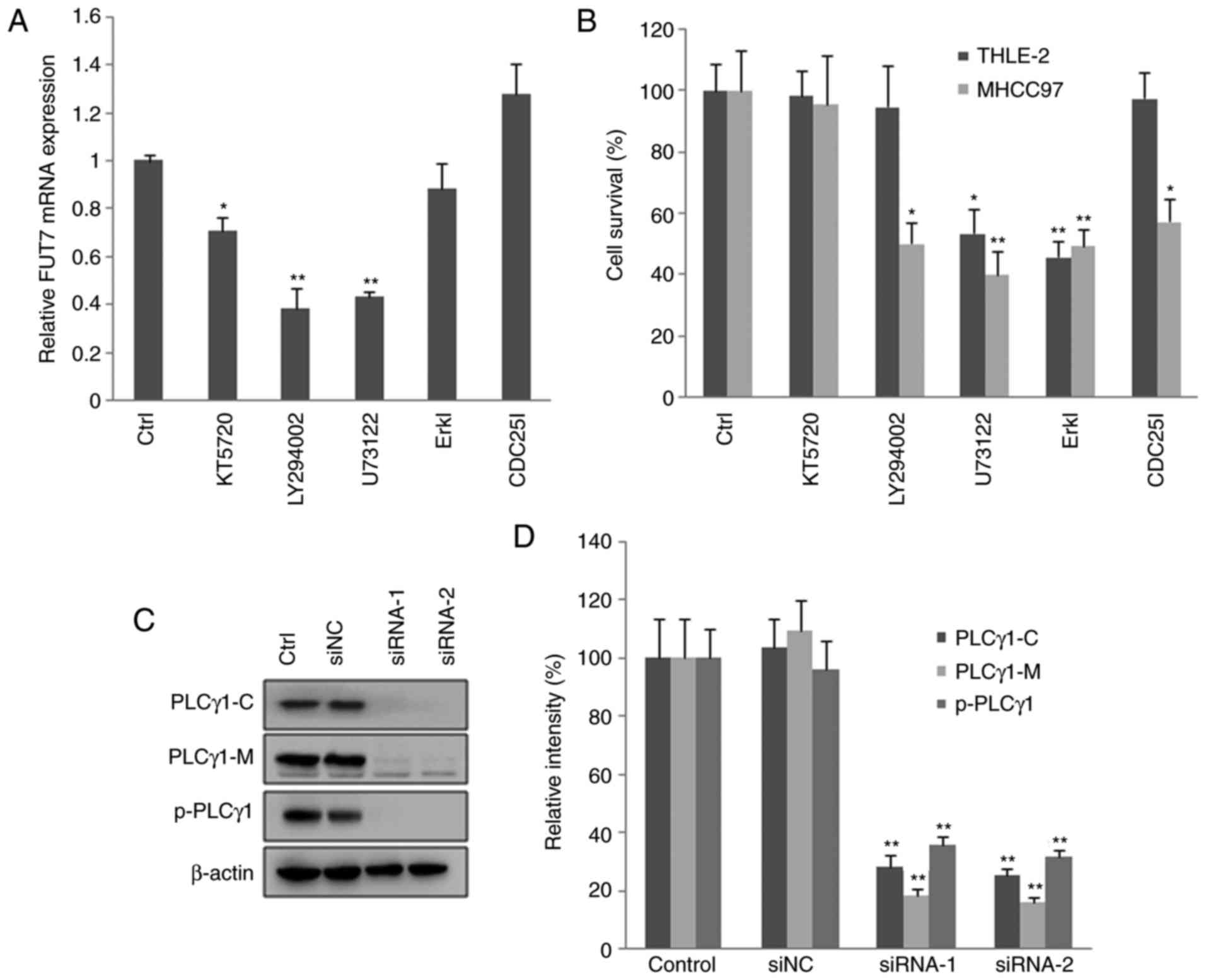 | Figure 4Knocking down FUT7 affects cell
survival via various intracellular signaling mediators. MHCC97
cells were transfected with FUT7 siRNAs. After 1 h, inhibitors of
signaling molecules were added to the culture medium, and the cells
were cultured for a further 48 h for RNA analysis and 72 h for
protein analysis. (A) mRNA levels of FUT7 were quantified by
reverse transcription-quantitative polymerase chain reaction
following treatment with inhibitors. (B) Following treatment with
the inhibitors, cell survival was quantified using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H tetrazolium bromide
assay in the THLE-2 normal liver cells and MHCC97 hepatocarcinoma
cells. (C) Cells were permeabilized and centrifuged to isolate the
cytosolic proteins and particulate-conjugated proteins. Results of
western blot analysis of the target proteins are shown with
profiles. (D) Quantitative analysis of relative protein levels.
*P<0.05 (n=3), vs. Control; **P<0.01
(n=3), vs. Control. FUT, fucosyltransferase; Ctrl, control (no
inhibitor); ErkI, extracellular signal-regulated kinase inhibitor;
siRNA, small interfering RNA; siNC, scramble-siRNA transfected
cells; PLCγ, phospholipase Cγ; PLCγ1-C, PLCγ1 in cytoplasm;
PLCγ1-M, PLCγ1 conjugated with plasma membrane; p-PLCγ1,
phosphorylated PLCγ1 conjugated with plasma membrane; Control,
untransfected cells. |
To observe the effect of FUT7 siRNA transfection on
the activation of PLCγ, the proteins in the cytoplasm were isolated
from particulate-conjugated proteins. The contents of PLCγ1 and
phosphorylated PLCγ1 were detected by western blot analysis. The
results (Fig. 4C and D) showed
that FUT7 siRNA transfection decreased the content of PLCγ1 in the
cytoplasm, and decreased the content of PLCγ1 and phosphorylated
PLCγ1 conjugated on the plasma membrane. The results demonstrated
that FUT7 siRNA transfection inhibited the translocation and
phosphorylation of PLCγ1, and thus inhibited the activation of
PLCγ1. Therefore, the aforementioned results indicated that FUT7
may affect cell proliferation through the activation of PLCγ. As
Erk is the downstream signaling molecule of PLCγ (30), the results indicated that FUT7
modulated cell proliferation via the PLCγ/Erk signaling pathway in
MHCC97 cells.
Discussion
Cell surface glycoconjugates have important
functional roles in cancer cell proliferation, growth, metastasis
and invasiveness. It has been reported that the changing expression
and components of glycoconjugates are correlated with
carcinogenesis in severaltyped of cancer (3-9).
Abnormal elevation in the expression of sialyl-Lewis
oligosaccharide antigens (SLea and SLex) has
been correlated with the malignancy of a wide range of tumors
(6-9). Several antibodies targeting the
sialyl-Lewis oligosaccharide have been developed as potential
therapeutic approaches for the treatment of sialyl-Lewis
oligosaccharide-positive tumors (31-33). However, there has been limited
investigation of methods to effectively inhibit the synthesis of
sialyl-Lewis oligosaccharide and its effects on tumor cell
proliferation and growth.
It has been reported that the SLea
oligosaccharide is mainly expressed in tumor cells derived from
digestive organs, including the pancreas, rectum, biliary tract and
colon (34). In addition, it has
been reported that the SLex oligosaccharide ismainly
expressed in ovarian, breast and pulmonary tumor cells (35). It has also been reported that the
level of SLex oligo-saccharide was significantly higher
in serum of patients with HCC than in serum of normal controls and
patients with liver cirrhosis (36). It has also been shown that the
expression of SLex oligosaccharide is closely associated
with the growth of HCC (37-39). The results of the present study
also indicated that the expression of SLex in HCC
tissues was higher than in adjacent normal liver tissues. In
addition, the findings indicated that the expression of
SLex was significantly increased in MHCC97 human HCC
tumor cells compared with that in normal liver cells. The
expression of SDLex was unchanged and the expression of
SLea was only detected in trace quantities.
FUT7 is the key enzyme for the synthesis of the
SLex oligosaccharide, therefore, the present study
investigated the expression of FUT7 in HCC. The results indicated
that the expression of FUT7 was markedly increased in MHCC97 cells
compared with normal liver cells. Following transfection with FUT7
siRNAs, the expression of SLex was significantly
decreased in the MHCC97 cells. It has been reported that FUT7 has
high substrate specificity and that FUT7 is the only FUT
responsible for the synthesis of SLex Lewis
oligosaccharide antigen (22,23). FUT3/6 is responsible for the
synthesis of SLex and SDLex, and FUT6 is the
major enzyme which is responsible for the synthesis of
SDLex from SLex (20). The results of the present study
indicated that the expression of FUT6 was also high, however, no
statistically significant difference was found between the MHCC97
cells and normal liver cells. The mRNA and protein levels of FUT7
and FUT6 were correlated with the expression of sialyl-Lewis
antigens (SLex and SDLex) in the MHCC97
cells. The expression of FUT7 has been reported to be altered in
several types of tumor (26,27). It has also been documented that
the expression of glycan-related gene in human HCC cells, which is
responsible for the synthesis of N-glycan and glycolipids,
particularly the sialyl-Lewis antigen, was significantly increased
(40). However, the mechanism of
FUT7 in cancer cell growth has not been investigated fully in human
HCC cells.
The results of the present study showed that FUT7 is
not only responsible for SLex synthesis, but is also
involved in cell proliferation. siRNAs of FUT7 suppressed the
expression of FUT7 and markedly inhibited the synthesis of
SLex oligosaccharide antigen and MHCC97 HCC cell
proliferation. The results also indicated that anti-SLex
monoclonal antibody KM93 significantly suppressed MHCC97 HCC cell
proliferation. Therefore, the suppression of cell proliferation
following FUT7 RNAi may be due to the decreased synthesis of
SLex. The detailed functional role of SLex is being
investigated in subsequent investigations.
Following the binding of hepatocyte growth factor
(HGF) ligand to the HGF receptor (receptor tyrosine kinase (MET))
on the liver cell surface, the kinase catalytic activity of MET is
activated, which triggers trans-phosphorylation of the tyrosines of
MET, including Tyr 1234 and Tyr 1235, Tyr 1349 and Tyr 1356. These
tyrosines recruit and engage a number of signaling transducers,
including PI3K, Src homology 2-containing phosphotyrosine
phosphatase2, and PLCγ, therefore initiating a whole spectrum of
biological activities, including cell survival, proliferation,
migration and invasion. The PLC/Erk signal transduction pathway is
one of the most important signaling pathways in regulating cell
proliferation, growth and differentiation (30). PLC can be divided into three
subclasses: PLCβ, PLCγ and PLCδ, according to their different
structures (41). PLCγ is one of
the direct downstream signal transducers of growth factor receptors
and has an important functional role in regulating cell growth and
proliferation. PLCγ1 is widely distributed in various tissues and
cells, whereas PLCγ2 is mainly distributed in the lungs, spleen and
thymus. Therefore, PLCγ1 was selected as the main signaling target
for the present study. The activation of PLCγ1 involves a series of
molecular events (42). First,
ligands conjugate with growth factor receptors that have receptor
protein tyrosine kinase activity. The receptors are then dimerized
and autophosphorylated. The intracellular domain of receptors is
autophosphorylated to form the phosphorylated tyrosine residues,
and these residues of the receptors form the binding site of the
SH2 domain of PLCγ1. Subsequently, PLCγ1 is phosphorylated at
certain tyrosine residues and is activated by receptor kinase. The
activated PLCγ1 then separates from its receptors and trans-locates
to the plasma membrane by binding phospholipid phosphatidylinositol
3,4,5 triphosphate (PIP3) through its PH structure. Subsequently,
PLCγ1 can hydrolyze phospholipid phosphatidylinositol 3,4,5
biphosphate (PIP2) to produce diacylglycerol (DAG) and inositol
1,4,5-trisphosphate (IP3). The phosphorylation and translocation of
PLCγ1 from the cytosol to the plasma membrane are necessary for its
activation and function. Therefore, the activation of PLCγ1 can be
detected by its phosphorylation and translocation. The experimental
results showed that the proliferation of MHCC97 cells was
suppressed following FUT7 siRNA transfection. The relative content
of PLCγ1 conjugated with the plasma membrane of MHCC97 cells
transfected with FUT7 siRNAs was also lower than that of the
control cells, and the phosphorylated PLCγ1 conjugated with the
plasma membrane was also decreased. This indicates that FUT7
knockdown inhibited the phosphorylation and translocation of PLCγ1
from the cytosol to the plasma membrane, and thus, inhibited the
activation of PLCγ1. In addition, siRNA targeting FUT7 reduced cell
proliferation and survival. The PLCγ inhibitor U73122 significantly
reduced the mRNA levels of FUT7 and inhibited the proliferation and
survival of the MHCC97 cells. The experiment using PLCγ inhibitor
supported the hypo thesis that the expression level of FUT7 is
important to sustain MHCC97 HCC cell proliferation and survival.
The mechanistic mechanism of PLCγ/ERK pathway regulating the mRNA
expression of FUT7 remains to be fully elucidated. It was
hypothesized that activated PLCγ1 cleaves the PIP2 into DAG and
IP3. DAG and calcium work together to activate protein kinase C.
This leads to a series of phosphorylation events downstream in the
mitogen-activated protein kinase (MAPK) cascade. First, the
activated PKC activates RAF, and RAF kinase phosphorylates and
activates MAPK kinase. This ultimately results in the
phosphorylation and activation of ERKs. ERK regulates the
activities of several transcription factors, including c-Jun,
activating transcription factor 2, ELK1 and heat shock factor 1. By
altering the levels and activities of transcription factors, ERK
alters the transcription of genes that are involved in regulation
of cell proliferation and cell cycle, including FUT7. The detailed
signaling mechanism and the factors involved in PLCγ1 regulating
the gene transcription of FUT7 remain fully elucidated.
In conclusion, the findings of the present study
revealed that the suppression of PLCγ/Erk signaling pathway
inhibited human HCC cell proliferation, which was, at least
partially, mediated by the decreased expression of FUT7 and its
product, SLex. The results demonstrated that FUT7 siRNAs
downregulated SLex synthesis, inhibited PLCγ activation,
arrested cell cycle progression and effectively reduced MHCC97 HCC
cell proliferation. These results suggest that FUT7 siRNAs maybe a
potential therapeutic agent to treat human SLex-positive
cancer. Therefore, targeting FUT7 for inactivation with RNAi
technology is an attractive approach for controlling HCC cell
proliferation.
Funding
This study was supported by the National Natural
Science Foundation (grant ns 81373173 and 30371359) and the
Research Fund for the Doctoral Program of Higher Education (grant
no. 20132104120002).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DL and HL conceived and designed the experiments;
DL, HS, ZB and GB Performed the experiments; WW, Ml and JL analyzed
the data; DL and HL wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Committee of China Medical University (Shenyang, Liaoning, China).
Written informed consent was obtained from each patient
recruited.
Patient consent for publication
Written informed consent was obtained from each
patient.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Lowe JB: Glycan-dependent leukocyte
adhesion and recruitment in inflammation. Curr Opin Cell Biol.
15:531–538. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Springer TA: Traffic signals for
lymphocyte recirculation and leukocyte emigration: The multistep
paradigm. Cell. 76:301–314. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kannagi R: Molecular mechanism for
cancer-associated induction of sialyl Lewis X and sialyl Lewis A
expression-the Warburg effect revisited. Glycoconj J. 20:353–364.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Roseman S: Reflections on glycobiology. J
Biol Chem. 276:41527–41542. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu YC, Yen HY, Chen CY, Chen CH, Cheng
PF, Juan YH, Chen CH, Khoo KH, Yu CJ, Yang PC, et al: Sialylation
and fucosylation of epidermal growth factor receptor suppress its
dimerization and activation in lung cancer cells. Proc Natl Acad
Sci USA. 108:11332–11337. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kannagi R: Carbohydrate-mediated cell
adhesion involved in hematogenous metastasis of cancer. Glycoconj
J. 14:577–584. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takada A, Ohmori K, Yoneda T, Tsuyuoka K,
Hasegawa A, Kiso M and Kannagi R: Contribution of carbohydrate
antigens sialyl Lewis A and sialyl Lewis X to adhesion of human
cancer cells to vascular endothelium. Cancer Res. 53:354–361.
1993.PubMed/NCBI
|
|
8
|
Ito H, Hiraiwa N, Sawada-Kasugai M,
Akamatsu S, Tachikawa T, Kasai Y, Akiyama S, Ito K, Takagi H and
Kannagi R: Altered mRNA expression of specific molecular species of
fucosyl- and sialyltransferases in human colorectal cancer tissues.
Int J Cancer. 71:5560–5564. 1997. View Article : Google Scholar
|
|
9
|
Ogawa JI, Inoue H and Koide S:
α-2,3-Sialyltransferase type 3N and α-1,3-fucosyltransferase type
VII are related to sialyl Lewis x synthesis and patient survival
from lung carcinoma. Cancer. 79:1678–1685. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuijipers TW: Terminal glycosyltransferase
activity: A selective role in cell adhesion. Blood. 81:873–882.
1993.
|
|
11
|
Candelier JJ, Mollicone R, Mennesson B,
Bergemer AM, Henry S, Couillin P and Oriol R:
Alpha-3-fucosyltransferases and their glycoconjugate antigen
products in the developing human kidney. Lab Invest. 69:449–459.
1993.PubMed/NCBI
|
|
12
|
Stroup GB, Anumula KR, Kline TF and
Caltabiano MM: Identification and characterization of two distinct
a-1-3-L-fucosyltransferase activities in human colon carcinoma.
Cancer Res. 50:6787–6792. 1990.PubMed/NCBI
|
|
13
|
Mollicone R, Candelier JJ, Mennesson B,
Couillin P, Venot AP and Oriol R: Five specificity patterns of
(1→3)-α-L-fucosyltransferase activity defined by use of synthetic
oligosaccharide acceptors. Differential expression of the enzymes
during human embryonic development and in adult tissues. Carbohydr
Res. 228:265–276. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Macher BA, Holmes EH, Swiedler SJ, Stults
CM and Srnka CA: Human alpha 1-3 fucosyltransferases. Glycobiology.
6:577–584. 1991. View Article : Google Scholar
|
|
15
|
Kukowska-Latallo JF, Larsen RD, Nair RP
and Lowe JB: A cloned human cDNA determines expression of a mouse
stage-specific embryonic antigen and the Lewis blood group alpha
(1,3/1,4) fucosyltransferase. Genes Dev. 4:1288–1303. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goelz SE, Hession C, Goff D, Griffiths B,
Tizaed R, Newman B, Chi-Rosso G and Lobb R: ELFT: A gene that
directs the expression of an ELAM-1 ligand. Cell. 63:1349–1356.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lowe JB, Kukowska-Latallo JF, Nair RP,
Larsen RD, Marks RM, Macher BA, Kelly R and Ernst LK: Molecular
cloning of a human fucosyltransferase gene that determines
expression of the Lewis x and VIM-2 epitopes but not
ELAM-1-dependent cell adhesion. J Biol Chem. 266:17467–17477.
1991.PubMed/NCBI
|
|
18
|
Kumar R, Potvin B, Muller WA and Stanley
P: Cloning of a human alpha(1,3)-fucosyltransferase gene that
encodes ELFT but does not confer ELAM-1 recognition on Chinese
hamster ovary cell transfectants. J Biol Chem. 266:21777–21783.
1991.PubMed/NCBI
|
|
19
|
Weston BW, Nair RP, Larsen RD and Lowe JB:
Isolation of a novel human alpha (1,3)fucosyltransferase gene and
molecular comparison to the human Lewis blood group alpha (1,3/1,4)
fucosyltransferase gene. Syntenic, homologous, nonallelic genes
encoding enzymes with distinct acceptor substrate specificities. J
Biol Chem. 267:4152–4160. 1992.PubMed/NCBI
|
|
20
|
Weston BW, Smith PL, Kelly RJ and Lowe JB:
Molecular cloning of a fourth member of a human alpha (1,3)
fucosyltransferase gene family Multiple homologous sequences that
determine expression of the Lewis x, sialyl Lewis x, and difucosyl
sialyl Lewis x epitopes. J Biol Chem. 267:24575–24584.
1992.PubMed/NCBI
|
|
21
|
Koszdin KL and Bowen BR: The cloning and
expression of a human alpha-1,3 fucosyltransferase capable of
forming the E-selectin ligand. Biochem Biophys Res Commun.
187:152–157. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sasaki K, Kurata K, Funayama K, Nagata M,
Watanabe E, Ohta S, Hanai N and Nishi T: Expression cloning of a
novel alpha 1,3-fucosyltransferase that is involved in biosynthesis
of the sialyl Lewis x carbohydrate determinants in leukocytes. J
Biol Chem. 269:14730–14737. 1994.PubMed/NCBI
|
|
23
|
Natsuka S, Gersten KM, Zenita K, Kannagi R
and Lowe JB: Molecular cloning of a cDNA encoding a novel human
leukocyte alpha-1,3-fucosyltransferase capable of synthesizing the
sialyl Lewis x determinant. J Biol Chem. 269:16789–16794.
1994.PubMed/NCBI
|
|
24
|
Kudo K, Ikehara Y, Togayachi A, Kaneko M,
Hiraga T, Sasaki K and Narimatsu H: Expression, cloning and
characterization of a novel murine alpha1,3-fucosyltransferase,
mFuc-IX, that synthesis the Lewis X (CD15) epitope in brain and
kidney. J Biol Chem. 273:26729–26738. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Narimatsu H: Human fucosyltransferases:
Tissue distribution of blood group antigens, cancer associated
antigens and fucosyltransferase. Tanpakushitsu Kakusan Koso.
43(Suppl 16): S2394–S2403. 1998.In Japanese.
|
|
26
|
Albrethsen J, Bøgebo R, Gammeltoft S,
Olsen J, Winther B and Raskov H: Upregulated expression of human
neutrophil peptides 12 and 3 (HNP13) in colon cancer serum and
tumours: A biomarker study. BMC Cancer. 5:82005. View Article : Google Scholar
|
|
27
|
Hiller KM, Mayben JP, Bendt KM, Manousos
GA, Senqer K, Cameron HS and Weston BW: Transfection of alpha (1,3)
fucosyltransferase antisense sequences impairs the proliferative
and tumorigenic ability of human colon carcinoma cells. Mol
Carcinog. 27:280–288. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods.
25:402–408. 2001. View Article : Google Scholar
|
|
29
|
Cordeiro C and Freire AP: Digiton in
permeabilization of Saccharomyces cerevisiae cells for in situ
enzyme assay. Anal Biochem. 229:145–148. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berridge MJ: Inositol triphosphate and
diacylycerol as second messenger. Biochem J. 220:345–360. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Klinger M, Farhan H, Just H, Drobny H,
Himmler G, Loibner H, Mudde GC, Freissmuth M and Sexl V: Antibodies
directed against Lewis -Y antigen inhibit signaling of Lewis-Y
modified ErbB receptors. Cancer Res. 64:1087–1093. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Farhan H, Schuster C, Klinger M, Weisz E,
Waxenecker G, Schuster M, Sexl V, Mudde GC, Freissmuth M and
Kircheis R: Inhibition of xenograft tumor growth and
down-regulation of ErbB receptors by an antibody directed against
Lewis Y antigen. J Pharmacol Exp Ther. 319:1459–1466. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Scott AM, Geleick D, Rubira M, Clarke K,
Nice EC, Smyth FE, Stockert E, Richards EC, Carr FJ, Harris WJ, et
al: Construction, production, and characterization of humanized
anti-Lewis Y monoclonal antibody 3S193 for targeted immunotherapy
of solid tumors. Cancer Res. 60:3254–3261. 2000.PubMed/NCBI
|
|
34
|
Kannagi R: Carbohydrate antigen sialyl
Lewis a-its pathophysiological significance and induction mechanism
in cancer progression. Chang Gung Med J. 30:189–209.
2007.PubMed/NCBI
|
|
35
|
Kannagi R, Izawa M, Koike T, Miyazaki K
and Kimura N: Carbohydrate-mediated cell adhesion in cancer
metastasis and angiogenesis. Cancer Sci. 95:377–384. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sakai N, Okada Y and Tsuji T: Increased
serum levels of the carrier molecules of the carbohydrate antigen
sialyl Lewis X in liver diseases. Acta Med Okayama. 51:79–85.
1997.PubMed/NCBI
|
|
37
|
Fujiwara Y, Shimada M, Takenaka K,
Kajiyama K, Shirabe K and Sugimachi K: The Sialyl Lewis X
expression in hepatocarcinogenesis: Potential predictor for the
emergence of hepatocellular carcinoma. Hepatogastroenterology.
49:213–217. 2002.PubMed/NCBI
|
|
38
|
Okada Y, Jin-no K, Ikeda H, Sakai N,
Sotozono M, Yonei T, Nakanishi S, Moriwaki S and Tsuji T: Changes
in the expression of sialyl-Lewisx, a hepatic
necroinflammation-associated carbohydrate neoantigen, in human
hepatocellular carcinomas. Cancer. 73:1811–1816. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Torii A, Nakayama A, Harada A, Nakao A,
Nonami T, Sakamoto J, Watanabe T, Ito M and Takagi H: Expression of
the CD15 antigen in hepatocellular carcinoma. Cancer. 71:3864–3867.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kang X, Wang N, Pei C, Sun L, Sun R, Chen
J and Liu Y: Glycan-related gene expression signatures in human
metastatic hepatocellular carcinoma cells. Exp Ther Med. 3:415–422.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Delmas P, Crest M and Brown DA: Functional
organization of PLC signaling microdomains in neurons. Trends
Neurosci. 27:41–47. 2004. View Article : Google Scholar
|
|
42
|
Liu B and Wu D: Analysis of G
protein-mediated activation of phospholipase C in cultured cells.
Methods Mol Biol. 237:99–102. 2004.
|