Introduction
Influenza A viruses cause worldwide outbreaks of
influenza and seasonal pandemics, and pose serious risks to public
health (1). Influenza A viruses
belong to the Orthomyxoviridae family, with a negative
single-stranded, segmented RNA genome, and have been categorized
into different subtypes on the basis of their hemagglutinin and
neuraminidase antigens (2). Their
negative-sense RNA genomes have no proof-reading mechanism during
replication, so they are extremely error-prone, giving rise to a
high mutation rate (3).
Therefore, it is a lengthy process to update the matched vaccine
for these quickly modifying viruses, and it is necessary to
continually develop novel effective antiviral drugs against
influenza viruses in order to prepare for the continual seasonal
outbreaks.
Currently available anti-influenza virus drugs
target the viral life cycle, including amantadine, rimantadine,
oseltamivir, zanamivir and peramivir. However, prolonged treatment
and the resulting immuno-compromised status of patients lead to
increases in drug-resistant mutations among influenza viruses
worldwide (4). Previous studies
have indicated that the influenza strains H3N2 and pdmH1N1 are
resistant to adamantanes (5), and
the latest outbreak of the H7N9 virus is also resistant to
oseltamivir (6). On the other
hand, novel antiviral drugs derived from traditional Chinese
medicine (TCM) do not tend to lead to the development of drug
resistance among the viruses so easily. Therefore, numerous studies
have focused on developing anti-influenza drugs using natural
resources, such as traditional medicines, which offer hopeful new
prospects for influenza management (1,7).
Coumarin is a fragrant organic chemical compound of
the benzopyrone chemical class that is a natural substance found in
many plant species, which exhibits a variety of potent
pharmacological activities, including antioxidant, antibacterial,
anti-inflammatory, antitumor and antiviral activities (8-13).
Previous studies by our research group have demonstrated that
eleutheroside B1, a coumarin compound, has a wide spectrum of
anti-human influenza virus efficacy, with an IC50 value
(i.e., the concentration which leads to half-maximal inhibition) of
64-125 µg/ml in vitro, and it inhibited the mRNA
expression of several chemokine genes and the influenza
nucleoprotein (NP) gene, while exhibiting low cytotoxicity
(14).
Over the past few decades, computational chemistry,
molecular biology, pharmacognosy and biotechnology have become
major scientific areas for the research of natural products. Some
modern technologies, including RNA sequencing and molecular
docking, have also been used to identify novel molecules for the
effective treatment of diseases, and to investigate the underlying
mechanisms of action and the specific targets, as well as DNA, RNA,
protein and enzyme interactions, associated with natural products
(15). RNA sequencing is a
genome-wide analytical technology that has been used to analyze the
transcriptome of the host response to human or avian influenza
virus infection (16,17). A previous study has also used this
technology to provide a comprehensive analysis of the
pharmacological effects of natural products (18). The molecular docking approach has
become an increasingly important tool in pharmaceutical research,
and can be used to model the interaction between a small molecule
and a protein at the atomic level, which enables the identification
of potential drug targets, as well as the ability to characterize
the behavior of small molecules in the binding site of target
proteins (19). In the present
study, the pathway profiles of influenza A virus-infected lung
epithelial (A549) cells following eleutheroside B1 treatment were
assessed, which has enabled us to gain a comprehensive
understanding of the mechanism of eleutheroside B1 activity against
influenza A viral infection. From the RNA sequencing results, it
was determined that eleutheroside B1 may exert its pharmacological
effects on multiple targets, including the immune system, the
glycan biosynthesis and metabolism pathways, signaling molecules
and their interactions, transcriptional regulation of the chemokine
signaling pathway, various types of N-glycan biosynthesis, and
cytokine-cytokine receptor interaction. A molecular docking
approach was subsequently used to investigate the potential target
receptor proteins for eleutheroside B1. Finally, a variety of
molecular biology techniques, including reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
assay, bisulfite treatment and DNA methylation analyses, were used
to evaluate the results from the RNA sequencing and molecular
docking experiments.
Materials and methods
Compounds, cells and viruses, and
software
Eleutheroside B1 was purified from Sarcandra
glabra extract (Si Chuan, China) and characterized by
high-resolution mass spectrometry and 1H and
13C nuclear magnetic resonance spectroscopy, as
described previously (13). The
purity of the compound exceeded 98%, according to analysis by
ultra-performance liquid chromatography/time-of-flight mass
spectrometry. Eleutheroside B1 was dissolved in dimethyl sulfoxide
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) as a stock solution
of 50 mg/ml, and stored at −20°C until use. A549 cells were
purchased from the American Tissue Culture Collection (ATCC;
Manassas, VA, USA). The cells were grown in Dulbecco's modified
Eagle's medium with 10% fetal bovine serum under standard
conditions at 37°C in 5% CO2 humidified air. The
influenza virus strain A/PR/8/34 (H1N1) was also purchased from
ATCC. The influenza viruses were propagated in the allantoic
cavities of chicken eggs. ORCID iD of ChemDraw Ultra 8.0 and
SYBYL-X2.1.1 software was used in this study (no.
0000-0003-1628-7416), kindly provided by Dr Xin-An Huang (Tropical
Medicine Institute, Guangzhou University of Chinese Medicine,
Guangzhou, China).
Cell culture, virus infection and sample
preparation
The A549 cells were grown in a monolayer up to 80%
confluency and detached from the flask using 10 mM EDTA (pH 7.4)
and 0.25% trypsin. The cells were harvested, and 6×105
A549 cells were seeded in 6-well tissue culture plates. On the
following day, the cells were washed twice with PBS and infected
with A/PR/8/34 [H1N1; 0.1 multiplicity of infection (MOI)] using
serum-free medium for 2 h at 37°C. The inoculum was removed, and
the cells were treated with or without eleutheroside B1 at a
concentration of 100 µg/ml (14). At 24 h post-infection, the cells
were lysed in TRIzol reagent (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and stored at −80°C.
RNA isolation, cDNA library construction
and sequencing
Total RNA extracts from each sample were obtained
with TRIzol reagent, according to the manufacturer's instructions
(Thermo Fisher Scientific, Inc.). The total RNA quality was
analyzed using agarose electrophoresis (1% gels). The
A260/A280 ratio was determined using a
NanoDrop spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.). RNA integrity was assessed by Agilent 2100
TapeStation analysis (Agilent Technologies, Santa Clara, CA, USA).
An A260/A280 ratio between 1.8 and 2.0 and an
RNA integrity number >7 were considered acceptable parameters
for RNA integrity. RNA sequencing was performed on an Illumina
X-ten RNA-Seq sequence production system (Illumina, Inc., San
Diego, CA, USA).
Pathway analysis of differentially
expressed genes (DEGs)
In order to obtain a list of DEGs, Gene Ontology
(GO) and pathway enrichment analyses were performed. In addition,
GO terms, Interpro (protein sequence analysis and classification)
terms, and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways
that were significantly enriched on our list of genes with altered
expression (P<0.05) were identified. Additionally, the
integrated pathways with statistical values were computed from our
list of DEGs using a reactome pathway analysis (http://www.reactome.org/). For over-representation
analysis, the number of altered genes associated with a pathway was
compared between the treatment and control groups. To determine
pathway perturbation, the significance of a particular gene to a
pathway was considered in determining the overall impact on the
pathway by examining all annotated functions/interactions of the
gene.
Targeting and molecular optimization
Two-dimensional [2D(a)] and three-dimensional
[3D(b)] structural information of the compound were obtained from
the National Centre for Biotechnology Information (NCBI;
(https://pubchem.ncbi.nlm.nih.gov/compound/12302276#section=Top),
and these were saved as mol.2 files. The 3D-formatted molecule was
imported into the Pharmaceutical Target Seeker (PTS; http://www.rcdd.org.cn/PTS/result), and a search
was conducted. Molecular modeling was conducted with the SYBYL
program package (20).
Eleutheroside B1 was used as the template. Its conformation was
searched and identified by energy minimization using the MMFF94
force field with the Powell conjugate gradient minimization
algorithm, and a convergence criterion of 0.005 kcal/(mol·Å). The
MMFF94 charge was used to calculate partial atomic charges. The
parameters were set as defaults.
Protein preparation and docking
Through the PTS, select proteins were identified
that were potentially matched with eleutheroside B1. Given our goal
of attempting to find a potential target of eleutheroside B1 that
would be effective against the influenza virus, and considering our
previous results, Golgi mannosidase α class II member 1 (MAN2A1)
and RNA polymerase II subunit A (POLR2A) were selected as candidate
targets. The four-letter PDB code (3BVT) of Drosophila
melanogaster MAN2A1 (with resolution at 1.3 Å) and the PDB code
(3D9N) of POLR2A (with resolution at 1.60 Å) were retrieved from
the Protein Data Bank (PDB; http://www.pdb.org/pdb/home/home.do). Structural
optimization of the protein was conducted using SYBYL software. The
crystal structure of D. melaogaster MAN2A1 was subjected to
geometry optimization using the Swiss PDB viewer v4.1.0 program.
The compound structure of eleutheroside B1 was prepared for docking
using ChemDraw Ultra 8.0 and SYBYL-X2.1.1 software. The execution
of molecular docking analysis and visualization were conducted
using Surflex-Dock software. The removal of solvent molecules,
hydrogen addition, and the AMBER7 FF99 charges calculation were
performed. Furthermore, all default parameters were used for
docking simulations by SYBYL Vina analysis.
RT-qPCR assay
A549 cells were seeded in 6-well plates at 37°C in
an atmosphere of 5% CO2, then infected with influenza
virus (strain A/PR/8/34; 0.1 MOI) and subsequently treated with
eleutheroside B1 at a diluted concentration. At 24 h
post-infection, the cells were collected for mRNA expression
analysis of the host genes (MAN2A2, POLR2A) and the viral genes
[polymerase acid (PA), polymerase basic (PB) 1 and 2 and
hemagglutinin (HA)] by RT-qPCR. Total RNA was extracted using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Total
RNA (1 µg) was reverse transcribed into cDNA using the
Prime-Script RT-PCR kit (Takara Biotechnology Co., Ltd., Dalian,
China) at 37°C for 15 min, 85°C for 5 sec. The samples were
amplified in SYBR Green qPCR buffer (ABP Biosciences, Jiemei long
biological Co., Ltd., Wuhan, China). qPCR was performed using an
ABI7500 Real-time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with the following thermocycling conditions: 95°C
for 3 min, followed by 40 cycles of 95°C for 5 sec and 60°C for 30
sec (16). Relative gene (PA,
PB1, PB2, HA, POLR2A, MAN2A2 and GAPDH) expression levels were
calculated using the 2−ΔΔCq method (21). Primer sequences are listed in
Table I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene | Primer | Sequence
(5′-3′) |
|---|
| PA | Forward |
ACACTACAGGGGCTGAGAAA |
| Reverse |
TGAACGAGAAAATGTGGATG |
| PB1 | Forward |
AGTTTTGGTGTGTCTGGGA |
| Reverse |
TTCGGGTTTGTATTTGTGTG |
| PB2 | Forward |
ACCCAGATGAAGGCACAG |
| Reverse |
TAGAGTCCCGTTTTCGTTTC |
| POLR2A | Forward |
GATGAACTGAAGCGAATGTCT |
| Reverse |
GTCGTCTCTGGGTATTTGATG |
| HA | Forward |
TGAACAGGGAAAAGGTAGATG |
| Reverse C |
AGGGAGACCAAAAGCAC |
| MAN2A2 | Forward |
GCCCTCATTTTCTGTTTATTG |
| Reverse C |
TGCCCTATTTACCCATCAC |
| GAPDH | Forward |
GCTGAGTATGTTGTGGAGTC |
| Reverse |
GCAGAAGGAGCAGAGATGA |
CpG island (CGI) prediction
A Hidden Markov Model (HMM)-based CGI prediction for
the MAN2A1 and POLR2A gene loci was obtained from the browser
(http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi)
(22).
Bisulfite treatment and DNA methylation
analysis
Genomic DNA was extracted from eleutheroside
B1-treated and non-treated A549 cells following an influenza virus
infection using a DNA Extraction of Cell and Tissue kit (cat. no.
GK0122; Generay Biotech Co., Ltd., Shanghai, China). The bisulfite
conversion was performed with 1 µg genomic DNA using an
EpiTect Fast DNA Bisulfite kit (cat. no. 59824; Qiagen China Co.,
Ltd., Shanghai, China), according to the manufacturer's
instructions. The bisulfite-modified DNA was amplified by PCR with
primers that were designed to cover two regions One primer extended
from −545 bp upstream and −213 bp downstream to the transcription
start site of MAN2A1, whereas the other extended from -199 bp
upstream and 139 bp downstream. For the POLR2A gene, 66 CpG sites
ranging from −606 to +476 bp relative to exon 1 in a CpG island
were selected for subsequent analysis, and for the design of
primers. Subsequently, the first-stage PCR product was amplified
under the following conditions: 95°C for 4 min; 40 cycles of 94°C
for 30 sec, 55°C for 30 sec, and 72°C for 40 sec, followed by a
final extension at 72°C for 5 min. PCR products were purified using
a PCR purification kit (cat. no. GK2043; Generay Biotech Co.,
Ltd.). The PCR products were cloned into the TA cloning vector
(cat. no. GV6021; Generay Biotech Co., Ltd.). Ten clones from each
donor were sequenced using an M13 reverse primer.
Statistical analysis
All data are expressed as the mean ± standard
deviation of at least three separate experiments. Differences
between two groups were analyzed using Student's t-test, whereas
differences between multiple groups were analyzed using one-way
analysis of variance followed by Fisher's Least Significant
Difference post-hoc test. Statistical analyses were performed using
SPSS 18.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
Sequencing and alignment
To investigate the molecular mechanism of
eleutheroside B1 action against influenza A virus infection, RNA
sequencing of influenza A virus-infected human lung epithelial A549
cells, in the presence or absence of eleutheroside B1 treatment,
was performed using an Illumina X-ten RNA-seq sequence production
system. The RNA sequences were subsequently aligned against the
human gene sequences. Three different experimental groups were
established: Lung epithelial A549 cells without infection (A549
group), A549 cells infected with influenza strain A/PR8/34/(H1N1)
alone (PR8 group), and A549 cells both infected with
A/PR8/34/(H1N1) and treated with 100 µg/ml eleutheroside B1
(PR8 + eleu group). At 24 h after the influenza virus infection,
samples were harvested and subjected to library construction and
RNA sequencing. An excess of 13 million raw reads for each sample
were generated from the constructed RNA sequencing libraries using
the Illumina X-ten RNA-seq sequence production system. After
stringent data filtering, 34.75 M (97.11%; A549 group), 33.97 M
(95.93%; PR8 group), or 34.79 M (95.45%; PR8 + eleu group)
sequencing reads could be unambiguously mapped against the
reference genome (Table II),
which confirmed the high quality of the sequencing of these
samples, and a satisfactory variable for subsequent analyses.
| Table IIRNA-seq overview: Reads mapping
quality summary. |
Table II
RNA-seq overview: Reads mapping
quality summary.
| Sample name | Total reads | Total bases | Mapped reads | Mapped rate
(%) | Proper paired
mapped | Singletons | MAPQ≥5 rate
(%) | Discordantly
mapped |
|---|
| A549 | 34753376 | 5.21E+09 | 33748010 | 97.11 | 33748010 | 0 | 95.26 | 0 |
| PR8 | 33971006 | 5.1E+09 | 32589740 | 95.93 | 32589740 | 0 | 94.10 | 0 |
| PR8+eleu | 34797038 | 5.22E+09 | 33215018 | 95.45 | 33215018 | 0 | 93.40 | 0 |
Effect of eleutheroside B1 treatment on
pathway regulation in influenza A virus-infected lung epithelial
A549 cells
The gene lists generated for influenza A
virus-infected human lung epithelial A549 cells in the absence or
presence of eleutheroside B1 treatment revealed large differences
in the number and the type of genes that were transcriptionally
activated. To gain an improved understanding of the host cell
response to influenza virus infection at 24 h, the 1,871 DEGs (958
upregulated and 913 downregulated genes) between the PR8 and the
PR8 + eleu groups were characterized by performing an enrichment
analysis according to biological process, cellular composition and
molecular function. GO enrichment analysis indicated that numerous
DEGs were involved in molecular function and biological processes,
and these results are presented in Table III. The GO enrichment results
revealed that eleutheroside B1 recruited molecular function and
biological process genes in the host to inhibit the infection of
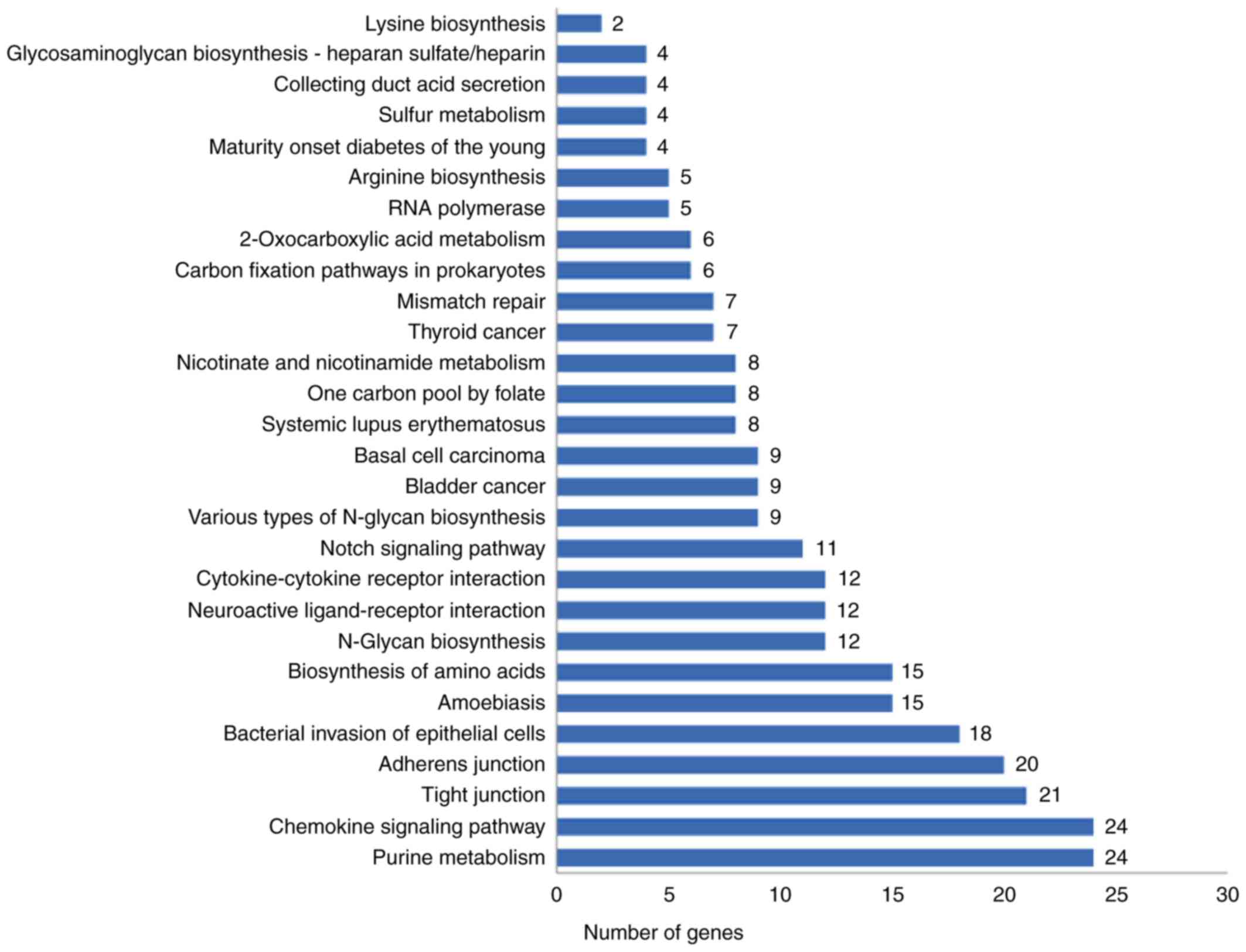
influenza virus. KEGG pathway enrichment analysis suggested that
these DEGs were associated with the immune system, glycan
biosynthesis and metabolism, signaling molecules and their
interaction, transcription (e.g., of the chemokine signaling
pathway), various types of N-glycan biosynthesis, cytokine-cytokine
receptor interaction, and RNA polymerase function, which are
host-response pathways that are activated in response to influenza
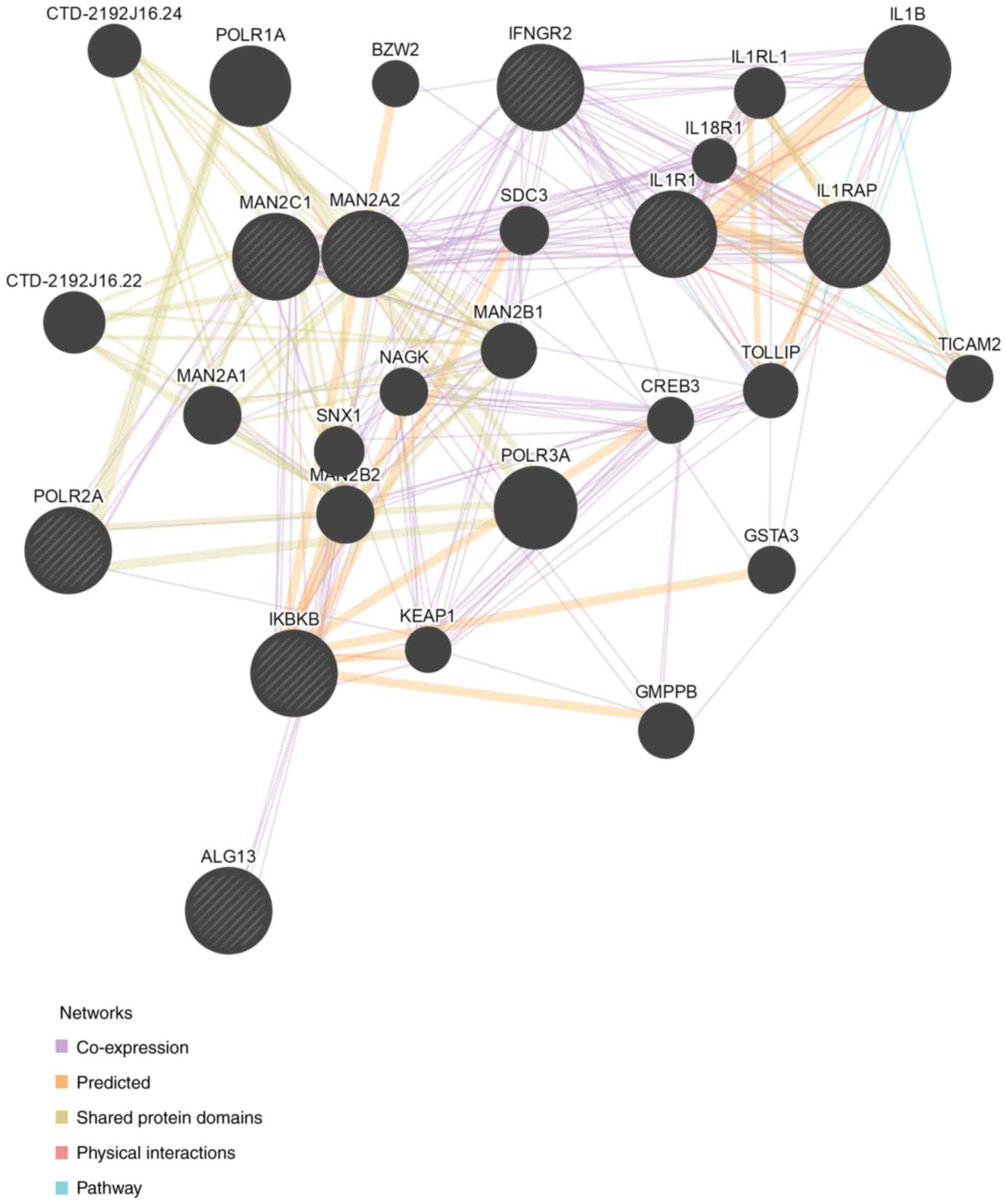
virus infection (Fig. 1). Twelve
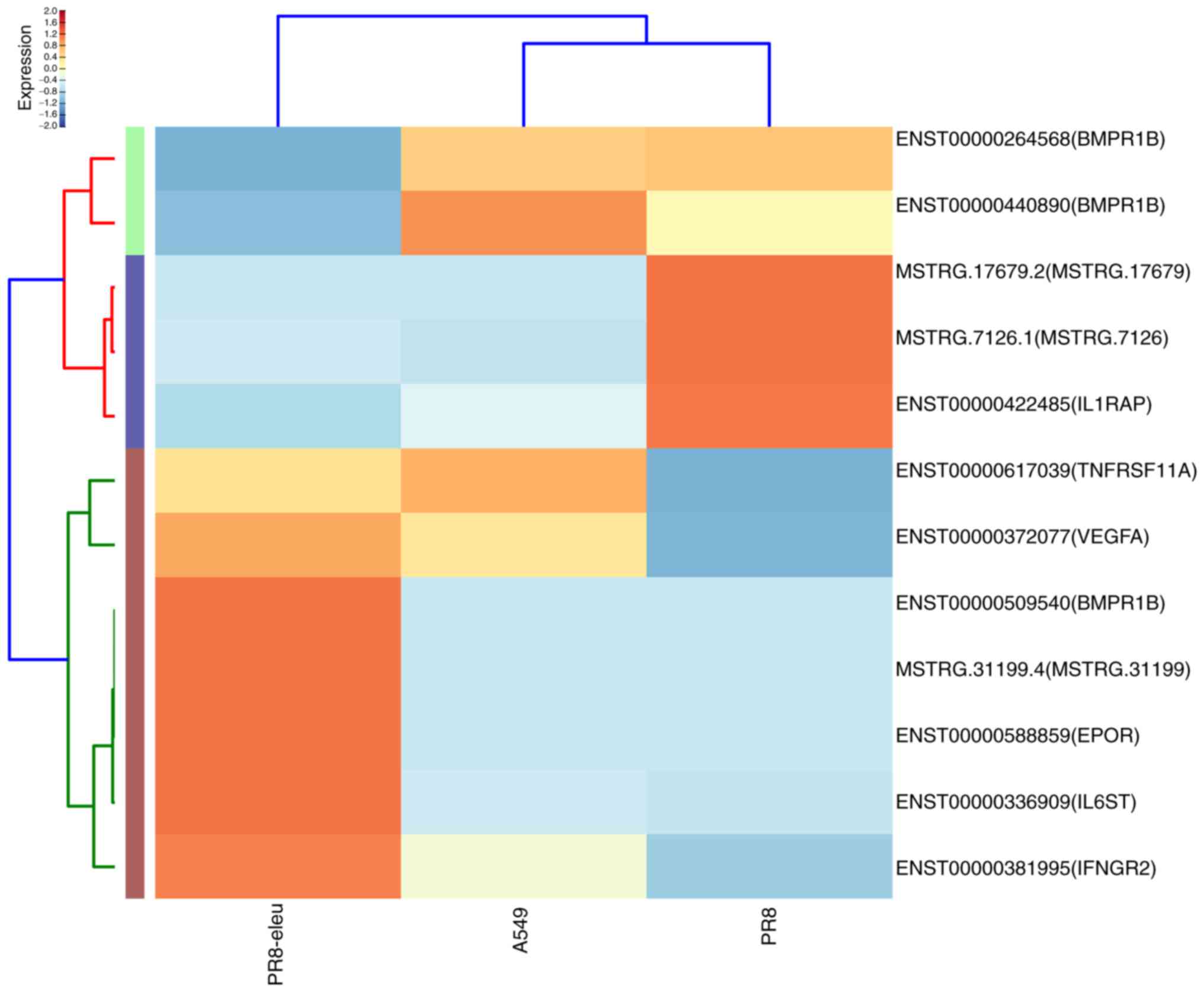
DEGs were enriched in cytokine-cytokine receptor interactions. Of
these DEGs, bone morphogenetic protein receptor type 1B (BMPR1B),
interleukin-6 signal transducer (IL6ST), interferon γ receptor 2
(IFNGR2), interleukin-1 receptor-associated protein (IL1RAP) and
erythropoietin receptor (EPOR) have been reported to be involved in
the host immune response to virus infections (Fig. 2). BMPR1B is a host gene, the
levels of which change during the process of influenza virus
infection (23), and this was
expressed at a 548-fold higher level compared with the PR8 group.
IFNGR2 is the upstream molecule in the interferon γ (IFN-γ)
signaling pathway, which is affected by the influenza virus
(24). Compared with the PR8
group, there was a 1,369-fold upregulation of the IFNGR2 gene in
the PR8 + eleu group. EPOR is crucial for the phosphorylation of
signal transducer and activator of transcription (STAT) 5, which
acts as an important molecule in the Janus kinase (JAK)-STAT
signaling pathway (22).
Influenza A virus directly interferes with IFN-γ-stimulated signal
transduction via this pathway, which enables the virus to inhibit
IFN-γ-inducible gene expression (24). In the present study, levels of the
EPOR gene were found to be upregulated by 255-fold. IL1RAP is a
co-receptor of the IL-1 receptor (IL1R1) and the IL-33 receptor
(ST2), and is required for signaling through both receptor
complexes. The inhibition of IL1RAP is also capable of blocking
IL-1 signaling, which disrupts the activation of nuclear factor
(NF)-κB mediated by IL-1 and IL-33, and may inhibit influenza
virus-induced pro-inflammatory reaction (25).
| Table IIISignificantly enriched GO terms in
response to eleutheroside B1. |
Table III
Significantly enriched GO terms in
response to eleutheroside B1.
| GO description | P-value | Number of
genes |
|---|
| Oxidoreductase
activity, acting on the CH-NH group of donors, NAD or NADP as
acceptor | 0.001378 | 6 |
| Heterocycle
biosynthetic process | 0.001468 | 119 |
| Aromatic compound
biosynthetic process | 0.001584 | 118 |
| Organic cyclic
compound biosynthetic process | 0.001615 | 119 |
| Oxidoreductase
activity, acting on the CH-NH group of donors | 0.002303 | 6 |
| DNA ligase
activity | 0.002306 | 4 |
| DNA ligation | 0.002306 | 4 |
| DNA ligase (ATP)
activity | 0.002306 | 4 |
| Binding | 0.002597 | 754 |
| Molecular
function | 0.002715 | 952 |
| Intra-Golgi
vesicle-mediated transport | 0.003632 | 6 |
| Establishment of
protein localization to Golgi | 0.003993 | 5 |
| Protein targeting
to Golgi | 0.003993 | 5 |
| Retrograde
transport, vesicle recycling within Golgi | 0.003993 | 5 |
| Protein
localization to Golgi apparatus | 0.003993 | 5 |
| Double-stranded
RNA-specific ribonuclease activity | 0.004917 | 4 |
| Ribonuclease III
activity | 0.004917 | 4 |
|
Methylenetetrahydrofolate dehydrogenase
(NADP+) activity | 0.004917 | 4 |
| Negative regulation
of transcription, DNA-templated | 0.005146 | 5 |
| Negative regulation
of gene expression | 0.005146 | 5 |
| Negative regulation
of nucleic acid-templated transcription | 0.006515 | 5 |
| Negative regulation
of RNA metabolic process | 0.006515 | 5 |
| Negative regulation
of RNA biosynthetic process | 0.006515 | 5 |
| Ligase activity,
forming phosphoric ester bonds | 0.006757 | 4 |
|
Nucleobase-containing compound
biosynthetic process | 0.007534 | 110 |
| RNA phosphodiester
bond hydrolysis, endonucleolytic | 0.00998 | 5 |
| Endoribonuclease
activity | 0.00998 | 5 |
| NADP biosynthetic
process | 0.010598 | 3 |
| NAD+ kinase
activity | 0.010598 | 3 |
|
Formate-tetrahydrofolate ligase
activity | 0.010598 | 3 |
| Regulation of
neurotransmitter levels | 0.011107 | 2 |
| Argininosuccinate
synthase activity | 0.011107 | 2 |
|
Methylenetetrahydrofolate reductase
(NAD(P)H) activity | 0.011107 | 2 |
| Gamma-tubulin
binding | 0.011107 | 2 |
|
Pteridine-containing compound biosynthetic
process | 0.011677 | 4 |
| Folic
acid-containing compound biosynthetic process | 0.011677 | 4 |
| Folic
acid-containing compound metabolic process | 0.011677 | 4 |
| DNA topoisomerase
type I activity | 0.015169 | 3 |
| Ligase activity,
forming carbon-nitrogen bonds | 0.015411 | 9 |
| Regulation of Ras
protein signal transduction | 0.016281 | 16 |
| Ras protein signal
transduction | 0.016281 | 16 |
| regulation of
cellular process | 0.016648 | 193 |
| Ion binding | 0.016903 | 192 |
| Regulation of small
GTPase mediated signal transduction | 0.017061 | 18 |
| Regative regulation
of cellular macromolecule biosynthetic process | 0.017259 | 5 |
| Regulation of
developmental process | 0.017259 | 5 |
| Metal ion
binding | 0.017719 | 188 |
| Endoribonuclease
activity, producing 5′-phosphomonoesters | 0.018441 | 4 |
| NADP metabolic
process | 0.018441 | 4 |
| Cation binding | 0.019467 | 188 |
| Regulation of
biological process | 0.019795 | 193 |
| Negative regulation
of biosynthetic process | 0.020303 | 5 |
| Negative regulation
of cellular biosynthetic process | 0.020303 | 5 |
| Negative regulation
of nitrogen compound metabolic process | 0.020303 | 5 |
| Negative regulation
of nucleobase-containing compound metabolic process | 0.020303 | 5 |
| Negative regulation
of macromolecule biosynthetic process | 0.020303 | 5 |
| DNA-directed RNA
polymerase II, core complex | 0.021299 | 2 |
|
Pteridine-containing compound metabolic
process | 0.022568 | 4 |
| Exonuclease
activity | 0.023676 | 5 |
| Biological
regulation | 0.02412 | 201 |
| Heterocycle
metabolic process | 0.024922 | 168 |
| Cellular aromatic
compound metabolic process | 0.025948 | 168 |
| Quanyl-nucleotide
exchange factor activity | 0.02599 | 18 |
| Sulfur compound
transmembrane transporter activity | 0.027145 | 3 |
| Organic cyclic
compound metabolic process | 0.028103 | 168 |
| Cellular nitrogen
compound biosynthetic process | 0.028363 | 129 |
| ARF
guanyl-nucleotide exchange factor activity | 0.031456 | 5 |
| Regulation of ARF
protein signal transduction | 0.031456 | 5 |
| ARF protein signal
transduction | 0.031456 | 5 |
| GDP binding | 0.031599 | 18 |
| Hydrolase activity,
acting on ester bonds | 0.031838 | 40 |
| Regulation of
intracellular signal transduction | 0.031966 | 19 |
| Transmembrane
receptor protein serine/threonine kinase activity | 0.032389 | 4 |
| Regulation of
multicellular organismal development | 0.032389 | 4 |
| Taurine
transmembrane transporter activity | 0.034046 | 2 |
| Negative regulation
of blood vessel morphogenesis | 0.034046 | 2 |
| Calcium-dependent
protein binding | 0.034046 | 2 |
| Taurine:sodium
symporter activity | 0.034046 | 2 |
| Negative regulation
of angiogenesis | 0.034046 | 2 |
| Histamine receptor
activity | 0.034046 | 2 |
| Negative regulation
of vasculature development | 0.034046 | 2 |
| Neurotransmitter
transporter activity | 0.034554 | 3 |
|
Neurotransmitter:sodium symporter
activity | 0.034554 | 3 |
| Mannose metabolic
process | 0.034554 | 3 |
| Organic acid:sodium
symporter activity | 0.034554 | 3 |
| Hormone receptor
binding | 0.0381 | 4 |
| Wnt signaling
pathway | 0.0381 | 4 |
| Nuclear hormone
receptor binding | 0.0381 | 4 |
| Molecular function
regulator | 0.038432 | 32 |
| DNA biosynthetic
process | 0.039752 | 6 |
| Phosphatase
regulator activity | 0.040667 | 5 |
| Intrinsic component
of plasma membrane | 0.040951 | 7 |
| Integral component
of plasma membrane | 0.040951 | 7 |
| DNA topological
change | 0.042894 | 3 |
| Aspartate family
amino acid metabolic process | 0.042894 | 3 |
| Signal
transduction | 0.042957 | 97 |
| Coenzyme
biosynthetic process | 0.044016 | 8 |
| Pyridine nucleotide
biosynthetic process | 0.044349 | 4 |
| Pyridine-containing
compound biosynthetic process | 0.044349 | 4 |
| Nicotinamide
nucleotide biosynthetic process | 0.044349 | 4 |
| Anatomical
structure morphogenesis | 0.044349 | 4 |
| Single organism
signaling | 0.044355 | 97 |
| Cellular response
to stimulus | 0.045164 | 111 |
| Signaling | 0.046516 | 97 |
| Transcription,
DNA-templated | 0.048815 | 90 |
| IMP dehydrogenase
activity | 0.048989 | 2 |
| Negative regulation
of developmental process | 0.048989 | 2 |
| Nucleoside
phosphate biosynthetic process | 0.049767 | 13 |
| Nucleotide
biosynthetic process | 0.049767 | 13 |
Influenza A virus infection in the lung is
associated with a robust host inflammatory response and sustained
elevated levels of inflammatory immune mediators (26), whereas certain natural product
compounds have been demonstrated to exhibit anti-inflammatory
effects (27,28). The DEG data associated with the
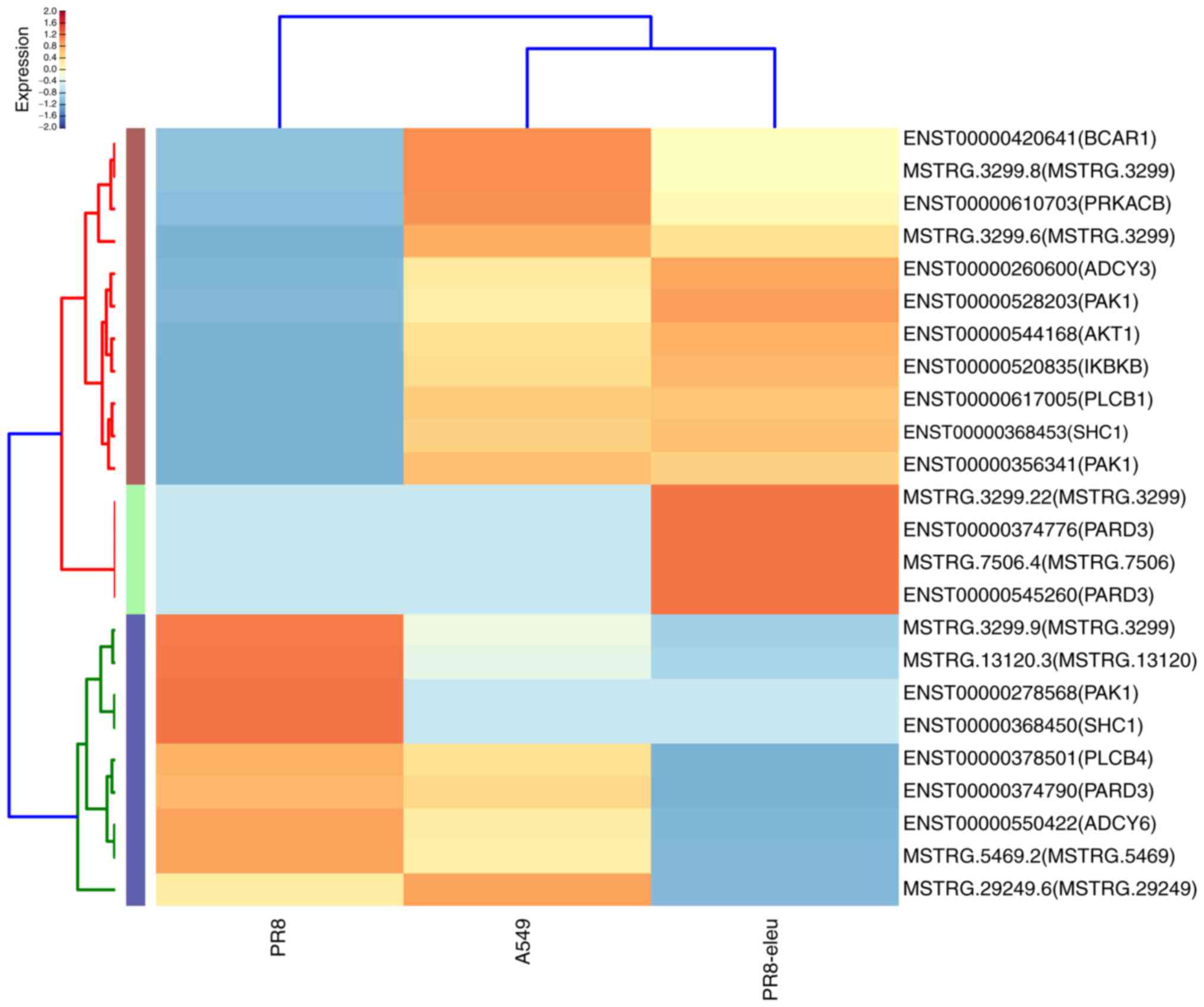
chemokine signaling pathway (Fig.
3) confirmed our previous finding that eleutheroside B1
exhibits anti-inflammatory activity against the influenza virus
(14). The DEGs identified in the
N-glycan biosynthesis pathway suggest other ways in which
eleutheroside B1 may work against the influenza virus. In the
N-glycan biosynthesis pathway, there are only a small number of
glycosyltransferases catalyzing different glycosidic linkages on
different N-glycans that exert a crucial role in cellular activity
(29). The synthesis of influenza
virus hemagglutinins has been reported to be involved in this
process (30). In the present
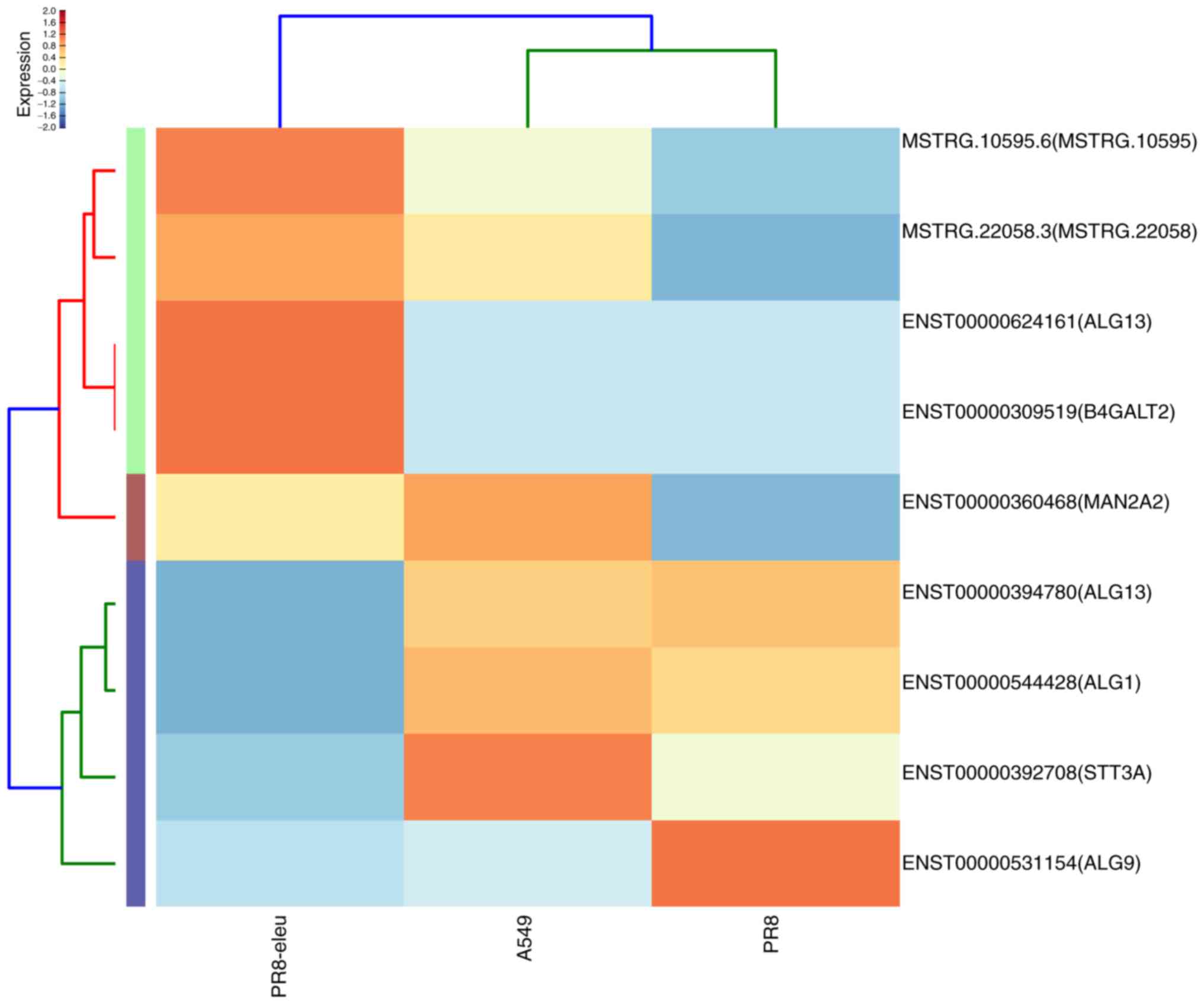
study, 12 enriched DEGs were identified in the major N-glycan
biosynthesis pathway, 9 enriched DEGs were found in various other
types of N-glycan biosynthesis pathways, and 3 genes were found in
other glycan degradation pathways (Fig. 4), all of which are involved in
glycan biosynthesis and metabolism. These included:
UDP-N-acetylglucosaminyltransferase subunit (ALG13),
mannosidase α class 2A member 2 (MAN2A2), mannosidase α class 2C
member 1 (MAN2C1), chitobiosyldiphosphodolichol
β-mannosyltransferase (ALG1), and α-1,2-mannosyltransferase (ALG9).
MAN2C1 has dual functions: One is associated with glycan
catabolism, whereas the other is in apoptotic signaling, and its
downregulation has been reported to delay cancer growth by inducing
either mitotic arrest or apoptosis (31). MAN2C1 overexpression also results
in synthesis of the non-glucosylated Man9GlcNAc2 oligosaccharide
precursor, which is transferred to the protein substrates more
inefficiently, thereby resulting in protein underglycosylation
(32). In the present study,
MAN2C1 was upregulated 9-fold following eleutheroside B1
treatment.
RNA synthesis is a fundamental process in gene
expression, and POLR2A, a housekeeping gene coding for the large
subunit of RNA polymerase II (RNAPII) in RNA biosynthesis (33), is a host factor involved in
RNAPII-dependent transcription and mRNA processing of the influenza
virus (34). The RNA sequencing
results of the present study revealed that POLR2A was downregulated
by 16,384-fold following eleutheroside B1 treatment.
POLR2A may interact with eleutheroside
B1, as determined from the molecular docking assay
Over the course of the last three decades, specific
inhibitors of certain enzymes associated with the N-glycan
biosynthesis pathway have demonstrated their potential for
antiviral and tumor therapy (35). Golgi MAN2A1 is a key enzyme, which
acts late in the N-glycan biosynthesis pathway and is a target in
the development of anticancer and antiviral therapies (36). Mannostatin A effectively blocked
processing of the influenza viral hemagglutinin in cultured MDCK
cells, causing the accumulation of hybrid-type protein-linked
oligosaccharides, a result which is in agreement with blocking
Golgi mannosidase II (36).
MAN2A1 inhibitors are also extracted from plant sources and,
therefore, the present study aimed to investigate whether MAN2A1
may be a potential target of eleutheroside B1. However, we did not
identify the structure of human MAN2A1 and is not available in
databases; therefore, the structure of D. melanogaster
MAN2A1 was used in the present study. In the initial results, D.
melanogaster MAN2A1 was docked with eleutheroside B1. In
addition, the crystal structure of human MAN2A1 was established
through homology modeling, and this predicted structure was used
for docking assays with eleutheroside B1. Furthermore, a previous
study by our research group had shown that eleutheroside B1
inhibited the polymerase activity of influenza virus (14). POLR2A is an important host factor
that is involved in the polymerase activity of influenza virus, and
is downregulated by eleutheroside B1. Therefore, POLR2A was also
selected for further investigation.
Small molecules were docked to these selected
proteins, and the docking scores were analyzed. In general, a
compound that has a high docking score with the protein (Total
Score and CScore) indicates that the compound may have potential
activities against the targeted receptor (37). From the results of the docking
study, it was possible to determine that the selected molecules
exhibited good binding affinity towards the homologous proteins in
the model. The specific scores are listed in Table IV.
| Table IVScore of molecular docking. |
Table IV
Score of molecular docking.
| Protein | Total_Score | CSCORE |
UNIFIED-CECORER |
|---|
| MAN2A1 | 11.3029 | 5 | 3 |
| POLR2A | 9.0133 | 5 | 2 |
To determine its potency, the best-docked
conformation of eleutheroside B1 was analyzed, and this molecular
compound was found to bind to the inside of D. melanogaster
MAN2A1(3BVT) and human POLR2A(3D9N), based upon the hydrogen and
hydrophobic interactions. If such features are identified, it
demonstrates that the compound in question may be responsible for
mediating biological activities. The docked conformations of
eleutheroside B1 in the D. melanogaster MAN2A1(3BVT) and
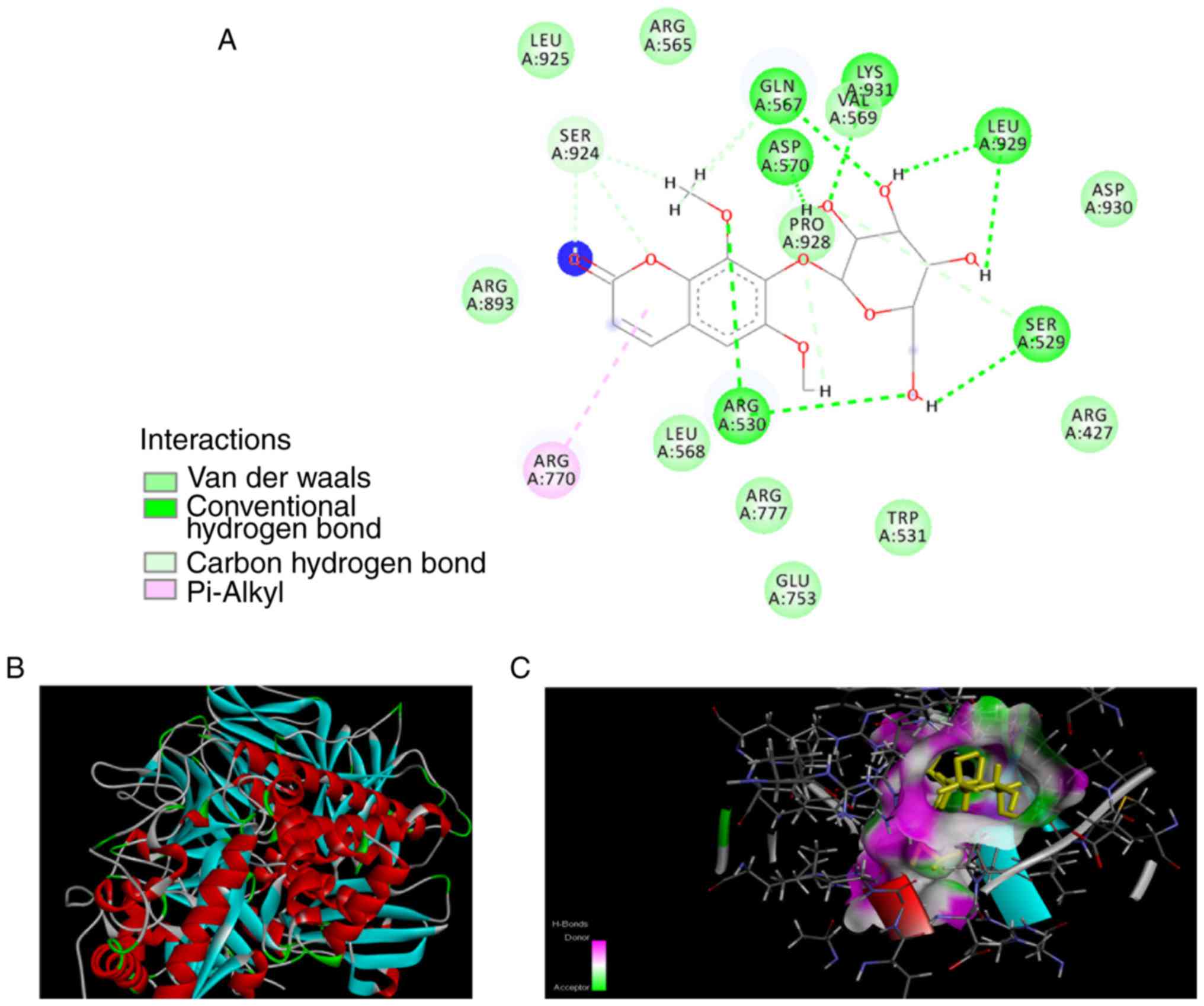
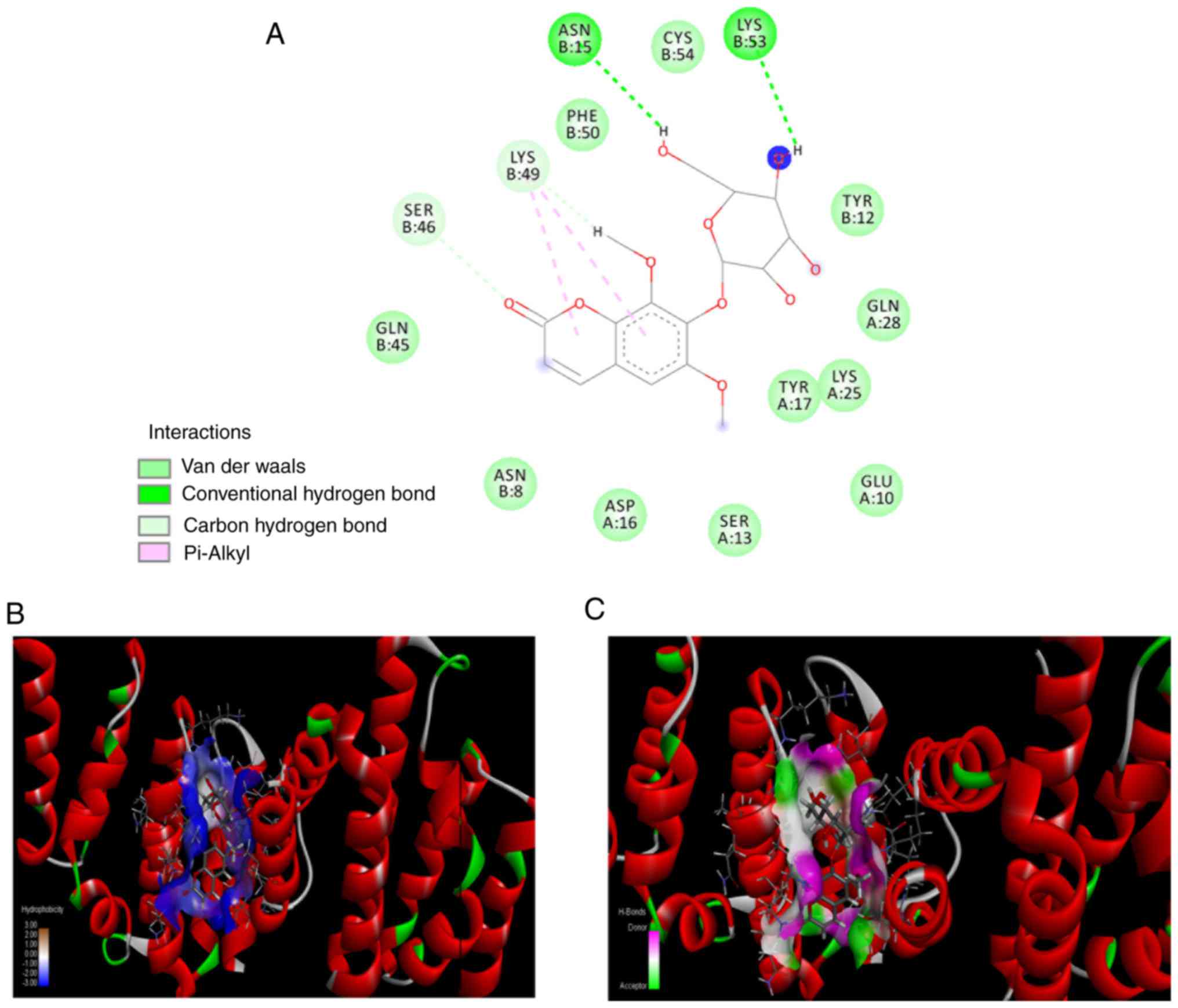
human POLR2A(3D9N) structures are illustrated in Figs. 5 and 6, respectively. The interaction analysis
revealed that there are 13 hydrogen bonds formed by eleutheroside
B1 in the binding site of D. melanogaster MAN2A1(3BVT), and
four hydrogen bonds formed by eleutheroside B1 in the binding site
of human POLR2A(3D9N), together with two carbon-hydrogen bonds and
two conventional hydrogen bonds. Other interactions were also
investigated, including van der Waals' forces, which contact the
surrounding residues in the binding site of bovine serum antigen,
including Arg-893, Leu-925, Arg-565, Leu-568, Arg-777, Glu-753,
Trp-531, Arg-427 and Asp-930. Inspection of the molecular
interactions revealed that hydrogen bonds and van der Waals' forces
were the major factors influencing the stabilities of the
eleutheroside B1-MAN2A1 and the eleutheroside B1-POLR2A complexes,
which are illustrated in Figs. 5
and 6, respectively.
MAN2A1(3BVT) in this docking assay belongs to D.
melanogaster, which has high sequence identity with human
MAN2A1 (41% identity; 61% similarity) (38). A predicted crystal structure for
the human MAN2A1 was also generated through homology modeling with
docked eleu-theroside B1, however this had a lower docking score
(data not shown). Since the protein POLR2A is associated with a
higher molecular docking score and is also of human origin, this
protein was identified as a target protein for eleutheroside B1,
and further experimental verification was performed at the cellular
level.
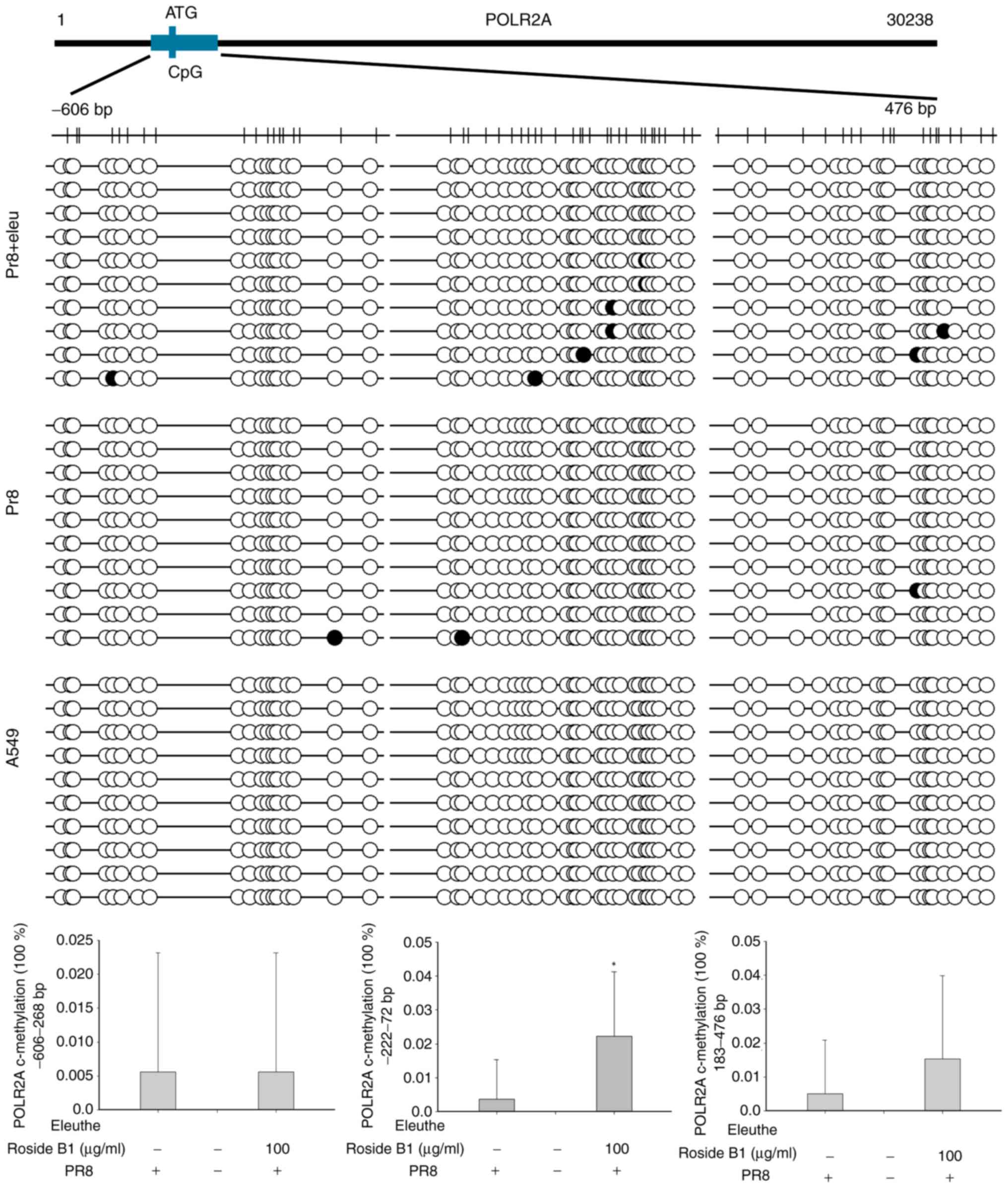
Eleutheroside B1 modifies the CpG
methylation levels of the POLR2A and MAN2A1 genes
Even in the molecular docking assay, human MAN2A1
did not match precisely with eleutheroside B1; however, the
possibility of their biological interaction could not be excluded.
Therefore, MAN2A1 and POLR2A were selected to examine the CpG
methylation levels modified by eleutheroside B1. CpG islands were
identified on the POLR2A and MAN2A1 promoters by sequence analysis
(http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi;
Fig. 7). To investigate whether
eleutheroside B1 modifies DNA methylation of POLR2A and MAN2A1,
bisulfite-sequencing PCR (BSP) was performed to quantitatively
analyze the methylation levels of CpG sites in these genes in A549
cells incubated with or without eleutheroside B1 following
infection with A/PR/8/34 (H1N1). Ten clones were selected for the
analysis of each CpG site. The BSP analysis revealed that the
average proportion of methylated CpGs of the POLR2A gene increased
significantly (P<0.05) in A549 cells treated with eleutheroside
B1 compared with the PR8 group (Fig.
7). However, no significant differences were identified for the
proportion of methylated CpGs of the MAN2A1 gene between A549 cells
with or without eleutheroside B1 treatment following infection with
A/PR/8/34 (H1N1) (data not shown).
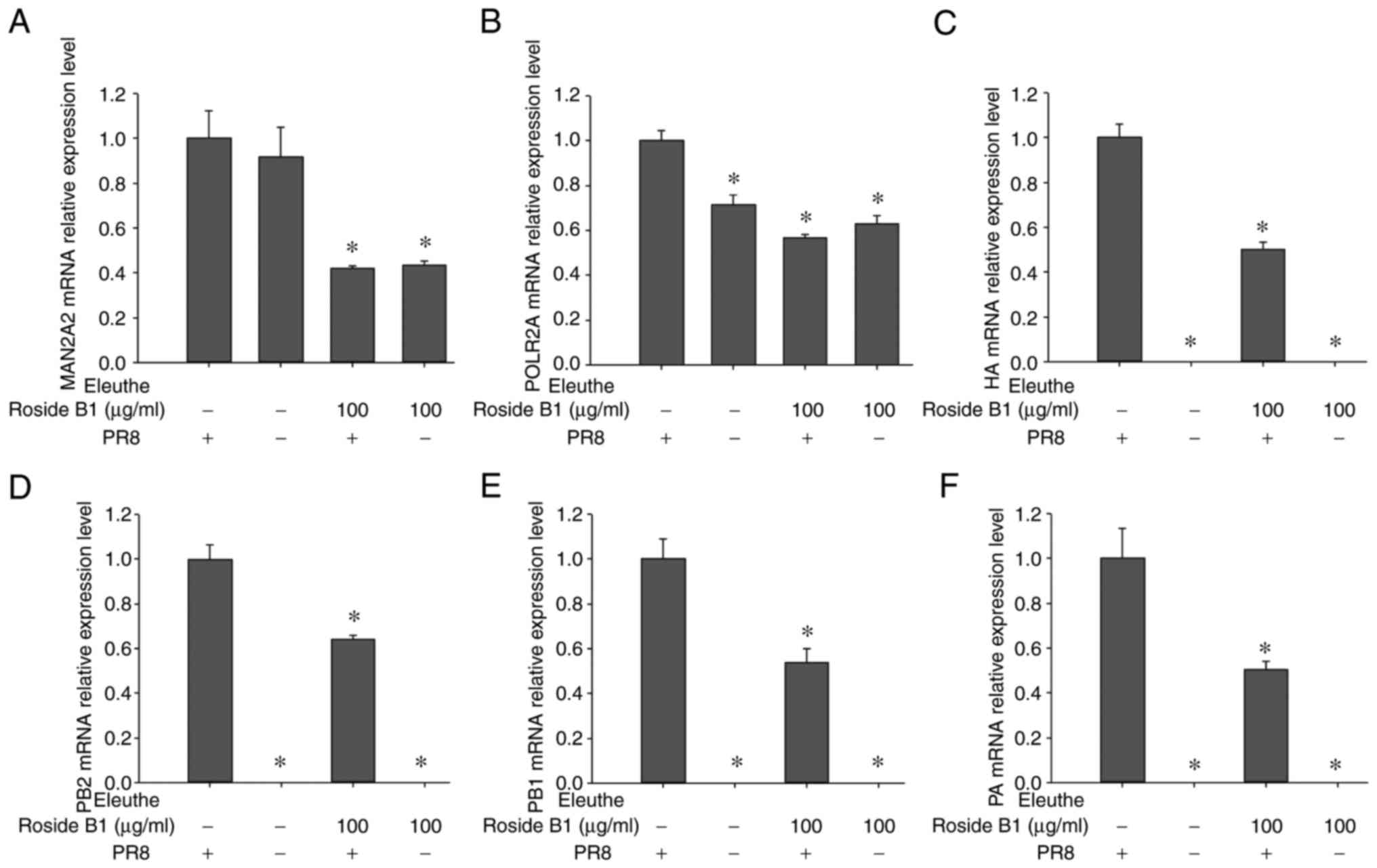
mRNA expression of host genes and virus
genes are influenced by eleutheroside B1 treatment in A549
cells
To identify genes involved in novel pathways, such
as various types of N-glycan biosynthesis and RNA polymerase
function, the expression levels of host genes (MAN2A2, POLR2A) were
detected. In glycan biosynthesis and metabolism, MAN2A1, MAN2A2,
and MAN2C1 were identified as the important targets. MAN2A1 did not
display a potential interaction with eleutheroside B1; MAN2A2,
however, was involved in the various types of N-glycan biosynthesis
pathways, whereas MAN2C1 was involved in other glycan degradation
pathways. POLR2A demonstrated a marked potential interaction with
eleutheroside B1 in the present study, and significant levels of
expression were observed following influenza infection in A549
cells treated with eleutheroside B1, and influenza infection of the
A549 cells in the RNA sequencing studies. Therefore, MAN2A2 and
POLR2A were selected for determination of their expression levels.
In addition, virus genes (PA, PB1, PB2, HA) in A/PR/8 (0.1
MOI)-infected A549 cells were also subjected to mRNA expression
studies. The results demonstrated that the expression of host genes
(MAN2A2, POLR2A) and virus genes (PA, PB1, PB2, HA) were decreased
following eleutheroside B1 treatment at a concentration of 100
µg/ml (Fig. 8). The
expression levels of the influenza virus ribonucleoprotein (RNP)
genes (PA, PB1, PB2) were down-regulated by eleutheroside B1, a
finding which was consistent with the results of a previous study,
showing that eleutheroside B1 may target the influenza virus RNP
and RNA polymerases in the host (14).
Discussion
Since a comprehensive analysis of the mechanisms
underlying specific agents of TCM acting against influenza virus at
the molecular level is difficult, RNA sequencing approaches may
compensate for such draw-backs, and provide a systematic analysis
for the study of TCM pharmacology in order to identify the key
molecular events that are linked with their efficacy (18).
In the RNA sequencing experiments of the present
study, the 1,871 DEGs identified between the PR8 and PR8 + eleu
groups were characterized by performing an enrichment analysis
according to biological processes, cellular composition, and
molecular function. These DEGs were involved in various types of
N-glycan biosynthesis, the chemokine signaling pathway,
cytokine-cytokine receptor interaction, and RNA polymerase
function, which are host responses to influenza viral infection.
The biosynthesis of N-glycans is very complex in mammals, as is the
glycosylation-mediated quality control of protein folding by
N-glycans (39). Glycosylation
has become an area of intensive study in recent years, due to its
ability to impact viral biology. A previous study demonstrated that
N-glycosylation of the influenza hemagglutinin serves an important
role in the life cycle of the influenza virus (35). The present RNA sequencing results
demonstrated that certain DEGs, including ALG13, MAN2A2, MAN2C1,
ALG1, and ALG9, were enriched in glycan biosynthesis and
metabolism. MAN2C1 overexpression also results in the synthesis of
non-glucosylated Man9GlcNAc2 oligosaccharide precursor, which is
transferred to the protein substrates rather inefficiently, thereby
resulting in suboptimal protein glycosylation. In the present
study, MAN2C1 was upregulated by 9-fold following eleutheroside B1
treatment. This finding indicated that eleutheroside B1 may
regulate the N-glycosylation of the influenza hemagglutinin via
this pathway, although further studies are required to confirm this
phenomenon.
Cytokines are soluble extracellular proteins or
glycoproteins that are crucial intercellular regulators and are
involved in innate, as well as adaptive, inflammation of host
defenses, cell growth, differentiation, cell death, angiogenesis,
development and repair processes (40). Cytokines and cytokine receptor
interaction networks are regarded as essential components of
inflammation (41). In the
current RNA sequencing studies, 12 DEGs were enriched in
cytokine-cytokine receptor interactions. These DEGs included
BMPR1B, IL6ST, IFNGR2, IL1RAP, and EPOR, which have been reported
to be involved in host immune response to viral infections. IL-6ST
encodes membrane glycoprotein 130 (gp130). It is the common signal
transducing component of the IL6 cytokine receptor family, which
forms complexes with the ligand-binding receptor subunits of
multiple IL6 cytokine family members, serving as an important
activator of JAK-STAT signaling. IL-6ST has also been reported to
be involved in the host response for the infection of influenza
virus (42,43). These results suggested that
eleutheroside B1 may inhibit influenza virus through cytokine and
cytokine receptor interaction networks, although more evidence is
required to conclusively demonstrate this finding.
Chemokines are small soluble molecules that regulate
cellular homing through molecular gradients (44). A previously published study
reported that the chemokines, C-C motif chemokine ligand 2
(CCL2)/monocyte chemoattractant protein 1 (MCP-1), C-X-C motif
chemokine ligand 8 (CXCL8)/interleukin 8 (IL8), CXCL9/monokine
induced by interferon-γ (MIG), and CXCL10/interferon γ-induced
protein 10 (IP-10), are also associated with mortality caused by
human and avian influenza viral infections (45). In the RNA sequencing results of
the present study, certain DEGs were also identified in the
chemokine signaling pathway, a finding which was consistent with
our previous study showing that eleutheroside B1 exhibited
anti-inflammatory activity against the influenza virus (14).
The influenza virus polymerase performs numerous
functions during the virus' life cycle, suggesting that many
cellular factors interact with this complex and are required for
the viral genome's transcription and replication. POLR2A is one of
the influenza virus polymerase-interacting proteins, and is
required for viral replication and transcriptional activity of the
viral polymerase (34). In the
RNA sequencing results, the levels of POLR2A were downregulated by
21,007-fold [logFC (PR8+eleu/PR8) = −14.3585910971] following
eleutheroside B1 treatment. The influenza virus transcribes and
replicates its genome inside the nucleus of infected cells, a
process that requires viral RNA polymerase, including the three
subunits PA, PB1, and PB2, as well as host cell factors to
transcribe and replicate the viral genome. The cellular
transcription factor POLR2A is required for virus replication and
the transcriptional activity of the viral polymerase. A previous
study demonstrated that the depletion of POLR2A could reduce by
2-fold the viral polymerase transcription of the influenza virus
(34). In the current
transcriptome data, eleutheroside B1 treatment also reduced the
expression of POLR2A, which demonstrated that POLR2A may be a
potential drug target for the inhibition of influenza virus. This
result was consistent with our previous finding that eleutheroside
B1 inhibited RNP mRNA expression and transcriptional activity of
the viral polymerase (14).
Molecular docking methodology explores the behavior
of small molecules in the binding pocket of a target protein, and
calculates the ability of the compounds to act against a particular
target (and their specificity). In modern drug discovery, molecular
docking methodology serves an important role in predicting the
orientation of the ligand, and provides potential leads for
researchers to identify potential drugs and drug targets (46). In the molecular docking results in
the present study, eleutheroside B1 was found to bind to the inside
of the D. melanogaster MAN2A1(3BVT) and human POLR2A, but
this was not the case for the predicted crystal structure of human
MAN2A1. These findings indicated that POLR2A may be the potential
target for the eleutheroside B1 against influenza virus infections
in humans, which was in accordance with the RNA sequencing
results.
MAN2A2 is an important enzyme in the N-glycan
biosynthesis pathway, and its proper functioning is required for
the glycosyltransferases of influenza virus hemagglutinins
(36). POLR2A is a housekeeping
gene in RNA polymerase II-dependent transcription in hosts, and it
is involved in the mRNA processing of influenza virus (33). PA, PB1, PB2, HA are four RNA
segments of the influenza virus genome that are crucial for the
genome-packaging process of influenza virus (47). Similar to human genes, influenza
virus must also pass its genome accurately on to the next
generation to ensure the survival of the species (48). The results of the RT-qPCR assay
revealed that the mRNA expression levels of MAN2A2, POLR2A, PA,
PB1, PB2, and HA, which are principally involved in viral RNP
function and virus replication, were all downregulated following
eleutheroside B1 treatment. Even in the cells receiving treatment
with eleutheroside B1 alone (i.e., no virus), effects on the
expression levels of MAN2A2 and POLR2A were also observed. These
results indicated that eleutheroside B1 may affect certain factors
associated with the N-glycan biosynthesis pathway and mRNA
processing in the host, which leads to the inhibition of the
production of influenza virus proteins.
Through recruiting proteins involved in gene
repression or inhibiting the binding of transcription factors to
DNA, DNA methylation is involved in the regulation of gene
expression, mRNA splicing, and genomic stability (49). In general, DNA methylation may
block the expression of a gene (50). DNA methylation analysis revealed
that eleutheroside B1 increased the average proportion of
methylated CpGs of the POLR2A gene significantly (P<0.05) in
A549 cells with influenza virus infection, compared with the virus
control cells. On the basis of our previous study (14) and the results of the present
study, it can be hypothesized that eleutheroside B1 may affect the
POLR2A gene DNA methylation pattern to inhibit the production of
the influenza virus genome. For the D. melanogaster
MAN2A1(3BVT), the molecular docking results demonstrated that it
was a good match with eleutheroside B1, but this was not true for
the human MAN2A1. Even in the DNA methylation analysis, the DNA
methylation levels of human MAN2A1 gene were not influenced by
eleutheroside B1 treatment. Therefore, the POLR2A gene is more
likely to be the target of eleutheroside B1 compared with the
MAN2A1 gene. However, additional studies are required in order to
confirm the detailed mechanism of action.
In conclusion, using RNA sequencing technology, the
present study demonstrated that eleutheroside B1 may inhibit
influenza virus via the chemokine signaling pathway,
cytokine-cytokine receptor interactions, N-glycan biosynthesis, and
RNA polymerase, and that several DEGs involved in these pathways
are interconnected as a part of a network, as illustrated in
Fig. 9. The changes in the
chemokine signaling pathway and in cytokine-cytokine receptor
interactions revealed in the present study are consistent with our
previous study, which demonstrated that the expression levels of
several cytokines were all downregulated by eleutheroside B1
(14). Previous studies have also
reported that compounds extracted from herbs exert a role in the
inflammatory process, and influence the immune system for the
prevention of infectious disease (51,52). In the present study, it was
observed that eleutheroside B1 targeted the POLR2A gene in order to
affect the expression of viral polymerase. Therefore, eleutheroside
B1 may be a promising agent for the prevention and treatment of
influenza A virus.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. U1502226), the Engineering
Technology Research Center (Development) of Guangdong general
universities (grant no. GCZX-A1408), and a grant from Guangzhou
Municipal Science and Technology Program-Technology Benefiting
Special (grant no. 2014Y2-00031).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
WY, CZ, JH and YW performed the experiments,
analyzed the data, prepared figures and tables; WZ, XH and XL
contributed to analyze data and prepare figures. WY and YW were
involved in the drafting of the manuscript. YW and XW designed the
study and reviewed drafts. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
We would like to thank Dr Richard H. Finnell for his
help at language editing.
References
|
1
|
Wu W, Li R, Li X, He J, Jiang S, Liu S and
Yang J: Quercetin as an antiviral agent inhibits influenza a virus
(IAV) entry. Viruses. 8:E62015. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Webster RG, Bean WJ, Gorman OT, Chambers
TM and Kawaoka Y: Evolution and ecology of influenza A viruses.
Microbiol Rev. 56:152–179. 1992.PubMed/NCBI
|
|
3
|
Yoon SW, Webby RJ and Webster RG:
Evolution and ecology of influenza A viruses. Curr Top Microbiol
Immunol. 385:359–375. 2014.PubMed/NCBI
|
|
4
|
Moss RB, Davey RT, Steigbigel RT and Fang
F: Targeting pandemic influenza: A primer on influenza antivirals
and drug resistance. J Antimicrob Chemother. 65:1086–1093. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Deyde VM, Xu XY, Bright RA, Shaw M, Smith
CB, Zhang Y, Shu Y, Gubareva LV, Cox NJ and Klimov AI: Surveillance
of resistance to adamantanes among influenza A(H3N2) and A(H1N1)
viruses isolated worldwide. J Infect Dis. 196:249–257. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yen HL, McKimm-Breschkin JL, Choy KT, Wong
DD, Cheung PP, Zhou J, Ng IH, Zhu H, Webby RJ, Guan Y, et al:
Resistance to neuraminidase inhibitors conferred by an R292K
mutation in a human influenza virus H7N9 isolate can be masked by a
mixed R/K viral population. mBio. 4:e00396–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin TJ, Lin CF, Chiu CH, Lee MC and Horng
JT: Inhibition of endosomal fusion activity of influenza virus by
Rheum tanguticum (da-huang). Sci Rep. 6:277682016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khan S, Riaz N, Afza N, Malik A,
Aziz-ur-Rehman, Iqbal L and Lateef M: Antioxidant constituents from
Cotoneaster racemiflora. J Asian Nat Prod Res. 11:44–48. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matos MJ, Vazquez-Rodriguez S, Santana L,
Uriarte E, Fuentes-Edfuf C, Santos Y and Muñoz-Crego A: Looking for
new targets: Simple coumarins as antibacterial agents. Med Chem.
8:1140–1145. 2012.PubMed/NCBI
|
|
10
|
Nakamura T, Kodama N, Oda M, Tsuchiya S,
Arai Y, Kumamoto T, Ishikawa T, Ueno K and Yano S: The
structure-activity relationship between oxycoumarin derivatives
showing inhibitory effects on iNOS in mouse macrophage RAW264.7
cells. J Nat Med. 63:15–20. 2009. View Article : Google Scholar
|
|
11
|
Lopez-Gonzalez JS, Prado-Garcia H,
Aguilar-Cazares D, Molina-Guarneros JA, Morales-Fuentes J and
Mandoki JJ: Apoptosis and cell cycle disturbances induced by
coumarin and 7-hydroxycoumarin on human lung carcinoma cell lines.
Lung Cancer. 43:275–283. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McKee TC, Covington CD, Fuller RW, Bokesch
HR, Young S, Cardellina JH II, Kadushin MR, Soejarto DD, Stevens
PF, Cragg GM and Boyd MR: Pyranocoumarins from tropical species of
the genus Calophyllum: A chemotaxonomic study of extracts in the
National Cancer Institute collection. J Nat Prod. 61:1252–1256.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan L, Wang Y, Liang N, Huang XJ, Li MM,
Fan CL, Wu ZL, Li YL and Ye WC: Chemical constituents from the
roots and stems of erycibe obtusifolia and their in vitro antiviral
activity. Planta Med. 79:1558–1564. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Yan W, Chen Q, Huang W, Yang Z, Li
X and Wang X: Inhibition viral RNP and anti-inflammatory activity
of coumarins against influenza virus. Biomed Pharmacother.
87:583–588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brahmachari G: Discovery and development
of antidiabetic agents from natural products. Elsevier Inc;
Waltham, MA: 2016
|
|
16
|
Ranaware PB, Mishra A, Vijayakumar P,
Gandhale PN, Kumar H, Kulkarni DD and Raut AA: Genome wide host
gene expression analysis in chicken lungs infected with avian
influenza viruses. PLoS One. 11:e01536712016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zou W, Chen D, Xiong M, Zhu J, Lin X, Wang
L, Zhang J, Chen L, Zhang H, Chen H, et al: Insights into the
increasing virulence of the swine-origin pandemic H1N1/2009
influenza virus. Sci Rep. 3:16012013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou B, Li J, Liang X, Yang Z and Jiang Z:
Transcriptome profiling of influenza A virus-infected lung
epithelial (A549) cells with lariciresinol-4-β-D-glucopyranoside
treatment. PLoS One. 12:e01730582017. View Article : Google Scholar
|
|
19
|
Meng XY, Zhang HX, Mezei M and Cui M:
Molecular docking: A powerful approach for structure-based drug
discovery. Curr Comput Aided Drug Des. 7:146–157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu L, Zhu H, Li L, Huang J, Sun W, Liu J,
Li H, Luo Z, Wang J, Xue Y, et al: (±) Japonones A and B, two pairs
of new enantiomers with anti-KSHV activities from Hypericum
japonicum. Sci Rep. 6:275882016. View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Toni T and Stumpf MP: Simulation-based
model selection for dynamical systems in systems and population
biology. Bioinformatics. 26:104–110. 2010. View Article : Google Scholar
|
|
23
|
Capitanio S and Wozniak RW: Host cell
factors necessary for influenza a infection: Meta-analysis of
genome wide studies. Quant Biol. 14:2012.
|
|
24
|
Uetani K, Hiroi M, Meguro T, Ogawa H,
Kamisako T, Ohmori Y and Erzurum SC: Influenza A virus abrogates
IFN-gamma response in respiratory epithelial cells by disruption of
the Jak/Stat pathway. Eur J Immunol. 38:1559–1573. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ågerstam H, Karlsson C, Hansen N, Sandén
C, Askmyr M, von Palffy S, Högberg C, Rissler M, Wunderlich M,
Juliusson G, et al: Antibodies targeting human IL1RAP (IL1R3) show
therapeutic effects in xenograft models of acute myeloid leukemia.
Proc Natl Acad Sci USA. 112:10786–10791. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chan RW, Leung CY, Nicholls JM, Peiris JS
and Chan MC: Proinflammatory cytokine response and viral
replication in mouse bone marrow derived macrophages infected with
influenza H1N1 and H5N1 viruses. PLoS One. 7:e510572012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saller R, Büechi S, Meyrat R and
Schmidhauser C: Combined herbal preparation for topical treatment
of Herpes labialis. Forsch Komplementarmed Klass Naturheilkd.
8:373–382. 2001.
|
|
28
|
Amritpal S: Herbal drugs as therapeutic
agents. CRC Press Inc; Boca Raton, FL: pp. 312014
|
|
29
|
Hossler P, Mulukutla BC and Hu WS: Systems
analysis of N-glycan processing in mammalian cells. PLoS One.
2:e7132007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Elbein AD, Dorling PR, Vosbeck K and
Horisberger M: Swainsonine prevents the processing of the
oligosaccharide chains of influenza virus hemagglutinin. J Biol
Chem. 257:1573–1576. 1982.PubMed/NCBI
|
|
31
|
Wang L and Suzuki T: Dual functions for
cytosolic alpha-mannosidase (Man2C1): Its down-regulation causes
mitochondria-dependent apoptosis independently of its
alpha-mannosidase activity. J Biol Chem. 288:11887–11896. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qu L, Ju JY, Chen SL, Shi Y, Xiang ZG,
Zhou YQ, Tian Y, Liu Y and Zhu LP: Inhibition of the
alpha-mannosidase Man2c1 gene expression enhances adhesion of
Jurkat cells. Cell Res. 16:622–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tantale K, Mueller F, Kozulic-Pirher A,
Lesne A, Victor JM, Robert MC, Capozi S, Chouaib R, Bäcker V,
Mateos-Langerak J, et al: A single-molecule view of transcription
reveals convoys of RNA polymerases and multi-scale bursting. Nat
Commun. 7:122482016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tafforeau L, Chantier T, Pradezynski F,
Pellet J, Mangeot PE, Vidalain PO, Andre P, Rabourdin-Combe C and
Lotteau V: Generation and comprehensive analysis of an influenza
virus polymerase cellular interaction network. J Virol.
85:13010–13018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wagner R, Liedtke S, Kretzschmar E, Geyer
H, Geyer R and Klenk HD: Elongation of the N-glycans of fowl plague
virus hemagglutinin expressed in Spodoptera frugiperda (Sf9) cells
by coexpression of human beta 1,2-N-acetylglucosaminyltransferase
I. Glycobiology. 6:165–175. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kobayashi Y and Suzuki Y: Evidence for
N-glycan shielding of antigenic sites during evolution of human
influenza A virus hemagglutinin. J Virol. 86:3446–3451. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lybrand TP: Ligand-protein docking and
rational drug design. Curr Opin Struct Biol. 5:224–228. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van den Elsen JM, Kuntz DA and Rose DR:
Structure of Golgi alpha-mannosidase II: A target for inhibition of
growth and metastasis of cancer cells. EMBO J. 20:3008–3017. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Stanley P, Taniguchi N and Aebi M: Chapter
9: N-Glycans. Essentials of glycobiology. 3. Varki A, Cummings RD,
Esko JD, Stanley P, Hart GW, Aebi M, Darvill AG, Kinoshita T,
Packer NH, Prestegard JH, et al: Cold Spring Harbor Laboratory
Press; Cold Spring Harbor NY: pp. 99–111. 2015
|
|
40
|
Ye MH, Bao H, Meng Y, Guan LL, Stothard P
and Plastow G: Comparative transcriptomic analysis of porcine
peripheral blood reveals differentially expressed genes from the
cytokine-cytokine receptor interaction pathway related to health
status. Genome. 60:1021–1028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dey R, Ji K, Liu Z and Chen L: A
cytokine-cytokine interaction in the assembly of higher-order
structure and activation of the interleukine-3: Receptor complex.
PLoS One. 4:e51882009. View Article : Google Scholar
|
|
42
|
Lin X, Huang C, Shi J, Wang R, Sun X, Liu
X, Zhao L and Jin M: Investigation of pathogenesis of H1N1
influenza virus and swine streptococcus suis Serotype 2
co-infection in pigs by microarray analysis. PLoS One.
10:e01240862015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shinya K, Gao Y, Cilloniz C, Suzuki Y,
Fujie M, Deng G, Zhu Q, Fan S, Makino A, Muramoto Y, et al:
Integrated clinical, pathologic, virologic, and transcriptomic
analysis of H5N1 influenza virus-induced viral pneumonia in the
rhesus macaque. J Virol. 86:6055–6066. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brummer G, Acevedo DS, Hu Q, Portsche M,
Fang WB, Yao M, Zinda B, Myers M, Alvarez N, Fields P, et al:
Chemokine signaling facilitates early-stage breast cancer survival
and invasion through fibroblast-dependent mechanisms. Mol Cancer
Res. 16:296–308. 2018. View Article : Google Scholar
|
|
45
|
Betakova T, Kostrabova A, Lachova V and
Turianova L: Cytokines induced during influenza virus infection.
Curr Pharm Des. 23:2616–2622. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pagadala NS, Syed K and Tuszynski J:
Software for molecular docking: A review. Biophys Rev. 9:91–102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
McCauley JW and Mahy BW: Structure and
function of the influenza virus genome. Biochem J. 211:281–294.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Noda T and Kawaoka Y: Packaging of
influenza virus genome: Robustness of selection. Proc Natl Acad Sci
USA. 109:8797–8798. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hall E, Dekker Nitert M, Volkov P,
Malmgren S, Mulder H, Bacos K and Ling C: The effects of high
glucose exposure on global gene expression and DNA methylation in
human pancreatic islets. Mol Cell Endocrinol. 472:57–67. 2018.
View Article : Google Scholar
|
|
50
|
Deaton AM and Bird A: CpG islands and the
regulation of transcription. Genes Dev. 25:1010–1022. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhou Li J, Li B, Chen C, Wang Q, Li Y,
Chen Z, Yang T, Jiang C, Zhong ZN, et al:
Lariciresinol-4-O-β-D-glucopyranoside from the root of Isatis
indigotica inhibits influenza A virus-induced pro-inflammatory
response. J Ethnopharmacol. 174:379–386. 2015. View Article : Google Scholar
|
|
52
|
Kang EH, Kown TY, Oh GT, Park WF, Park SI,
Park SK and Lee YI: The flavonoid ellagic acid from a medicinal
herb inhibits host immune tolerance induced by the hepatitis B
virus-e antigen. Antiviral Res. 72:100–106. 2006. View Article : Google Scholar : PubMed/NCBI
|