Introduction
Lung cancer is one of the most common aggressive
malignancies and non-small cell lung carcinoma (NSCLC) accounts for
~85% of lung cancer-associated mortalities (1). Metastasis is common in patients with
NSCLC and early metastasis is responsible for a majority that
succumb to the disease (2,3).
Random genetic and epigenetic mutations in cancer cells, combined
with a plastic and responsive microenvironment, support the
metastatic evolution of tumors. Metastasis comprises a series of
complex processes requiring the interaction of different signaling
pathways; it involves the detachment of tumor cells, the
degradation of extracellular matrix (ECM), the invasion, migration
and adhesion of endothelial cells, and the re-establishment of
growth at distant sites (4,5).
Genes associated with the initiation of metastasis and virulence
operate in the early and late stages of invasion and growth, when
located within the primary tumor and in various metastatic
environments, respectively (6). A
previous study suggested that the mammalian target of rapamycin
(mTOR) signaling pathway was involved in the transformation and
neoplastic proliferation of human NSCLC malignancies. Constitutive
activation of the phosphatidylinositol-4,5-bisphosphate 3-kinase
(PI3K)/protein kinase B (Akt)/mTOR signaling pathway occurs in 90%
of NSCLC cell lines (7). The mTOR
signaling pathway primarily regulates growth by affecting ribosome
biogenesis, protein translation and autophagy, and has emerged as a
promising target for therapies against diseases, including cancer
and diabetes (8). It appears to
be a prime strategic target for inhibiting the proliferation,
invasion and migration of thyroid cancer, breast cancer,
glioblastoma and gastric adenocarcinoma (9-12).
Mitogen-activated protein kinase kinase kinase
kinase 3 (MAP4K3), also termed germinal center-like kinase, is a
regulator of cell growth that is required for maximal
mTORC1-dependent S6K/4E-BP1 phosphorylation in cell cultures
(13,14). In addition to promoting the
activation of mTORC1, there is evidence that MAP4K3 is involved in
tumor metastasis, viability and apoptosis. MicroRNA let-7c has been
reported to inhibit the migration and invasion of SKEMS-1 cells by
targeting MAP4K3 (15) and MAP4K3
knockdown almost eradicated breast cancer cell migration (16). MAP4K3 is overexpressed in
pulmonary tissues of patients with NSCLC and its overexpression is
correlated with high recurrence risk and poor recurrence-free
survival rates (17). Therefore,
MAP4K3 may be a prognostic biomarker for NSCLC recurrence and a
promising antimetastatic and antitumor target.
To assist in developing superior anti-NSCLC
treatments, the present study examined a panel of compounds with
anti-MAP4K3 activity and identified two targets, Astragalus
polysaccharide (APS) and 10-hydroxycamptothecin (HCPT). APS is an
active ingredient found in the dried roots of Astragalus
membranaceus, a herb used in numerous traditional Chinese
medicines (18,19), and HCPT, a natural camptothecin
(CPT) derivative, has increased antitumor activity and decreased
toxicity compared with CPT (20).
In the present study, the anti-NSCLC effects of APS and HCPT on
H1299 cells were investigated following single and combination
treatments.
Materials and methods
Reagents, antibodies and plasmid
constructs
pRK5myc-MAP4K3 (M4K3) and AFG kinase-dead (KD)
MAP4K3 were prepared as described previously (13). APS [2-(chlorom
ethyl)-4-(4-nitrophenyl)-1,3-thiazole; ≥98%] was purchased from
ShangHai YuanYe Biotechnology Co., Ltd. (Shanghai, China; CAS no.
89250-26-0; cat. no. C18M6Y1). HCPT (98%) was purchased from
Shanghai Aladdin Biochemical Co., Ltd. (Shanghai, China; CAS no.
19685-09-7; cat. no. H1524105). 3-methyladenine (3-MA) and
chloroquine (CQ) were obtained from Sigma-Aldrich, EMD Millipore
(Billerica, MA, USA). Fetal bovine serum (FBS) was from HyClone, GE
Healthcare Life Sciences (Logan, UT, USA) and penicillin G,
streptomycin and Dulbecco’s modified Eagle’s medium (DMEM) were
from Gibco, Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
p-nitrobenzyl mesylate (PNBM) was from Epitomics (Burlingame, CA,
USA). Myelin basic protein (MBP),
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and isopropyl alcohol were purchased from Sigma-Aldrich, EMD
Millipore. β-actin mouse monoclonal antibody (mAb; cat. no.
ab8226), Bax rabbit antibody (Ab; cat. no. 2772), Bcl-2 rabbit mAb
(cat. no. 3498), caspase-3 rabbit Ab (cat. no. 9662), caspase-9
rabbit Ab (cat. no. 9502S), cytochrome c (D18C7) rabbit mAb
(cat. no. 11940S), p70S6K mouse mAb (cat. no. 611261), phospho-p70
S6K (Thr389) rabbit Ab (cat. no. 9205), MAP4K3 rabbit Ab (cat. no.
PAB3189), anti-myc 9E10 mouse mAb (cat. no. 05-419), thiophosphate
ester rabbit mAb (cat. no. ab92570), microtubule-associated protein
1 light chain 3 (LC3) rabbit Ab (cat. no. 8025) and P62 rabbit mAb
(cat. no. 11940) were purchased from Cell Signaling Technology,
Inc. (Boston, MA, USA), Abcam (Cambridge, UK) or BD Biosciences
(San Diego, CA, USA). Secondary horseradish peroxidase-conjugated
goat anti-rabbit immunoglobulin G (IgG; cat. no. 31460) and
horseradish peroxidase-conjugated goat anti-mouse IgG (cat. no.
31430) antibodies were purchased from Thermo Fisher Scientific,
Inc. All other chemicals were of analytical grade.
Cell culture and transfection
Human H1299 NSCLC cells (H1299), NCI H460 (H460)
cells and 293T cells were obtained from the Chinese Academy of
Sciences (Shanghai Institute of Cell Biology and Biochemistry, and
the Chinese Type Culture Collection, Shanghai, China). The cell
lines were cultured in DMEM supplemented with 10% heat-inactivated
FBS, 100 U/ml penicillin and 100 µg/ml streptomycin. The
cells were maintained at 37°C in a humidified 5% CO2
incubator.
For transfection, the cell lines were plated at 5
×105 cells/well in 6-well plates and incubated overnight
prior to transfection with Lipofectamine® 2000
(Invitrogen, Thermo Fisher Scientific, Inc.), according to the
manufacturer’s protocol. Unless otherwise specified, H1299 cells or
H460 cells were transfected with 0.5 µg pRK5myc Vec, 0.5
µg AFG MAP4K3-KD or 0.5 µg pRK5myc M4K3 and cultured
for 24 h, after which, they were used in subsequent
experiments.
MAP4K3 in vitro kinase activity
assay
Immunoprecipitated myc-tagged MAP4K3 from 293T
cells, produced as previously described (13), were incubated with MBP as a
substrate and the indicated agents. To screen for inhibitors of
MAP4K3 by western blot analysis, ATPγS was used at a concentration
of 1 mM in appropriate kinase buffers (21). The proteins were alkylated with
2.5 mM PNBM for 2 h at room temperature (18-22°C) and the products
were analyzed by western blot analysis.
Cell viability
The H1299 cells were seeded in 96-well cell culture
plates and incubated for 24 h. Following treatment with the agents
at the indicated concentrations and times, cell viability was
evaluated using an MTT assay, which is based on the reduction of a
tetrazolium salt by mitochondrial dehydrogenase by viable cells.
Following treatment, MTT solution (final concentration, 500
µg/ml) was added to each well and incubated for 4 h at 37°C.
The formed formazan crystals were dissolved in DMSO and the
absorbance was measured at 490 nm using a microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Flow cytometry
Cell cycles were analyzed using a propidium iodide
(PI) cell cycle detection kit (BD Biosciences), according to the
manufacturer’s protocol. The cells were trypsinized, washed with
PBS, centrifuged at 190 × g for 5 min at room temperature, and the
concentration was adjusted to 1×106 cells/ml. The cells
were resuspended in solution A at room temperature for 10 min.
Solution B was added for 10 min and the cells were stained with
fluorescein isothiocyanate (FITC)-labeled PI at room temperature
for 15 min under light-protected conditions.
Flow cytometric analysis was performed using a BD
FACSAria II Cell Sorter (BD Biosciences). The percentage of
apoptotic cells was calculated using BD FACSDiva Software v7.0 (BD
Biosciences, Franklin Lakes, NJ, USA). The apoptotic indices were
determined using a FITC-labeled Annexin V/PI apoptosis detection
kit (Sigma-Aldrich, EMD Millipore). Briefly, the cells were
harvested, washed with PBS and centrifuged at 190 × g for 5 min at
room temperature. The cells were adjusted to 1×106
cells/ml, resuspended in binding buffer and stained with
FITC-labeled Annexin V/PI at room temperature for 15 min under
light-protected conditions. Flow cytometric analysis was performed
using a FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes,
NJ, USA).
Wound-healing assay
Wound healing was assayed as previously described
with modifications (22).
Briefly, the H1299 cells and H460 cells were cultured to confluence
in 6-well cell culture plates for 24 h in serum-free medium,
respectively. The medium was replaced with serum-containing medium
followed by the addition of APS or/and HCPT, and cell monolayers
were disrupted by scraping with a 100-µl micropipette tip.
At the indicated times (0 and 24 h without transfection; 0,19 and
60 h with transfection) following scraping, the cells were washed
twice with PBS (pH 7.4) and images were captured using an optical
microscope at ×40 magnification.
In vitro migration and invasion
assay
The chemotactic directional migration of H1299 cells
was measured using 24-well Transwell inserts. Pore filters
(8-µm; Corning Incorporated, Corning, NY, USA) were coated
with gelatin (Sigma-Aldrich, EMD Millipore). Following culture for
24 h in serum-free DMEM, the cells were resuspended in serum-free
medium, seeded in the upper chambers of the Transwell inserts
(1×105 cells/ml) and incubated with APS and/or HCPT.
DMEM containing 10% FBS was placed in the lower chamber. The cells
in each treatment group were incubated for 24 h at 37°C in a
humidified atmosphere with 95% air and 5% CO2. The
filters were stained with 0.1% crystal violet. Cells that had
migrated and adhered to the underside of the filters were counted
and images were captured using an optical microscope at × 40
magnification. The invasiveness of the H1299 cells was measured
using Matrigel-coated Transwell cell culture chambers (8-µm
pore size) as previously described. The method was the same as for
migration analysis. Cells that had penetrated through the Matrigel
coating and adhered to the lower surface of the filters were
counted and images were captured using an optical microscope at ×40
magnification.
Western blot analysis
The cells were washed in 1X PBS and lysed in lysis
buffer as described (13). The
total protein concentrations was determined using the Bradford
method. The total protein or protein fractions (50 µg/lane)
were loaded and separated by 10 or 12% SDS-PAGE and transferred
onto PVDF membranes. The membranes were blocked with 5% skimmed
milk powder for 45 min at room temperature. The membranes were then
incubated with primary antibodies directed against β-actin
(dilution 1:2,000), Bax (dilution 1:1,000), Bcl-2 (dilution
1:1,000), caspase-3 (dilution 1:1,000), caspase-9 (dilution
1:1,000), cytochrome c dilution 1:1,000), p70S6K (dilution
1:500), phospho-p70 S6K (Thr389) (dilution 1:1,000), MAP4K3 rabbit
Ab (dilution 1:500), myc (dilution 1:2,000), thiophosphate ester
(dilution 1:5,000), LC3 (dilution 1:1,000) and P62 (dilution
1:1,000) overnight at 4°C. The membranes were subsequently washed
three times with PBS-0.1% Tween-20 for 10 min and were then
incubated with goat anti-mouse (dilution 1:10,000) or goat
anti-rabbit (dilution 1:5,000) secondary antibodies for 1 h at room
temperature. The expression of individual proteins was detected
with an enhanced chemiluminescence kit (Applygen Technologies,
Inc., Beijing, China). The densitometric values of the bands were
measured using ImageQuant TL software (version 8.1; GE Life
Sciences, Chicago, IL, USA).
Statistical analysis
Data are reported as the mean ± standard error of
the mean. Differences between groups were analyzed using Student’s
t-test with SPSS 14.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of APS, HCPT and their
combination on MAP4K3 and mTOR signaling in H1299 cells
To identify an inhibitor specific for MAP4K3, the
in vitro activity of MAP4K3 was measured by incubating the
kinase with the generic substrate MBP. In the presence of ATPγS,
MAP4K3 kinase activity was markedly inhibited by APS and HCPT
(Fig. 1A).
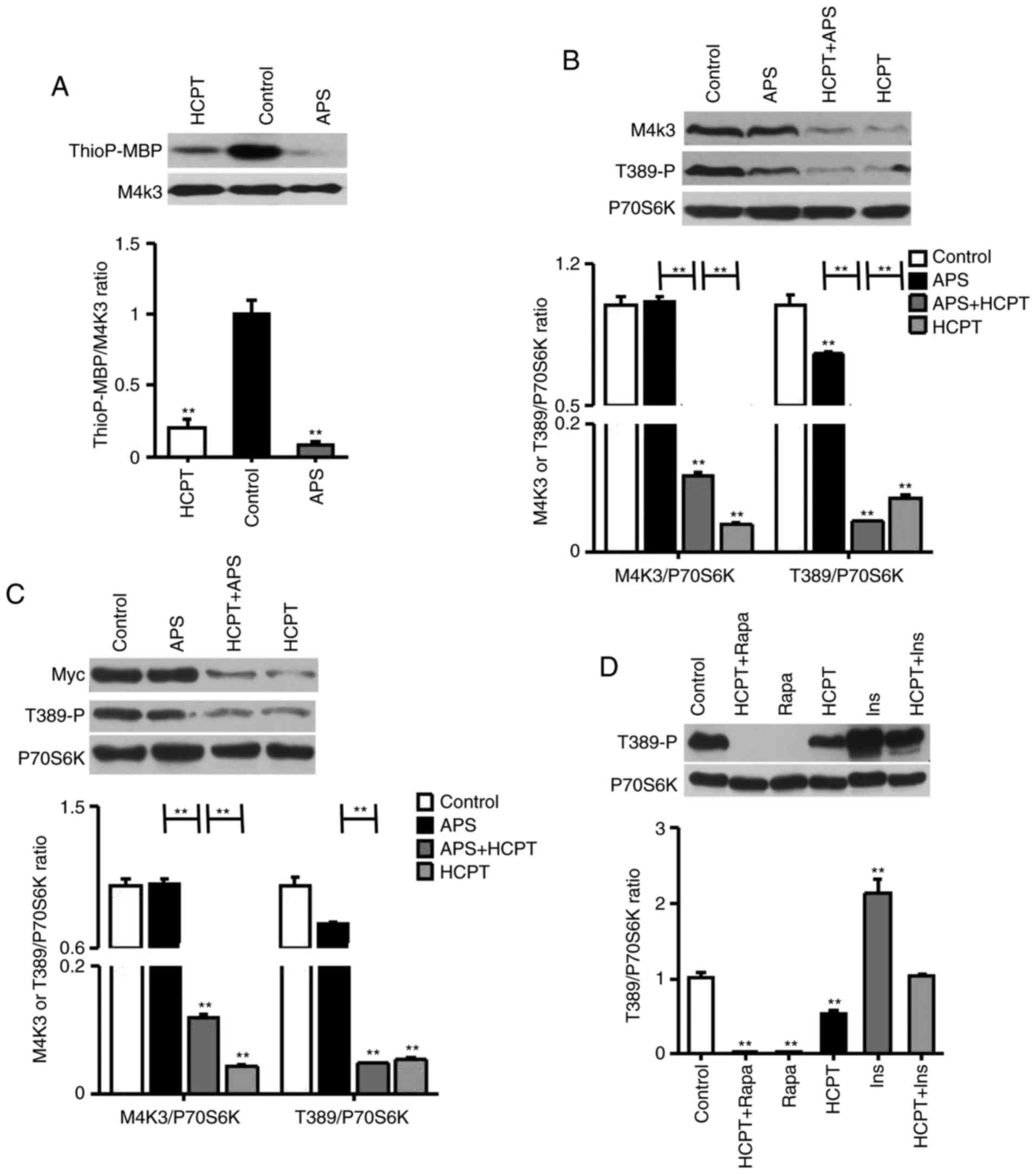 | Figure 1APS, HCPT and the combination of APS
and HCPT inhibit the MAP4K3/mTOR signaling pathway in H1299 cells.
(A) In vitro kinase assays using WT-MAP4K3
immunoprecipitated from 293T cells with myelin basic protein as a
substrate, ATPγS as a phospho-donor and 1 mg/ml APS or 1 µM
HCPT incubated at 30°C for 30 min. (B) Endogenous expression of
MAP4K3 and Thr389-phosphorylation of S6K in H1299 cells, detected
by western blot analysis following incubation with 1 mg/ml APS or 1
µM HCPT, or the combined agents, for 24 h. (C) H1299 cells
were transfected with 0.5 µg pRK5mycMAP4K3, serum-starved
for 16 h and treated for 24 h with 1 mg/ml APS or 1 µM HCPT,
or the combined agents. (D) Thr389-phosphorylation of S6K in H1299
cells was assessed via western blot analysis. Serum-starved cells
were incubated with 1 µM Ins for 30 min or 1 µM HCPT
for 24 h, or Ins followed by HCPT; or, cells were treated with 50
nM Rapa for 1 h or 1 µM HCPT for 24 h, or with Rapa followed
by HCPT. Western blot results are representative of three
independent experiments. APS, Astragalus polysaccharide;
HCPT, 10-hydroxycamptothecin; M4K3, mitogen-activated protein
kinase kinase kinase kinase 3; mTOR, mammalian target of rapamycin;
WT, wild-type; Rapa, rapamycin; Ins, insulin.
**P<0.01, vs. untreated control. |
Subsequently, it was demonstrated that APS and HCPT
inhibited the phosphorylation of S6K. The endogenous expression of
MAP4K3 in H1299 cells was markedly reduced by HCPT, although not by
APS (Fig. 1B). HCPT and the
combined agents inhibited S6K phosphorylation at Thr389 by
10.02±0.01 and 20.89±0.01-fold, respectively, which was increased
compared with APS (1.33±0.02-fold). In addition, the overexpression
of MAP4K3 reduced the inhibition of mTOR signaling in the presence
of APS, although not when treated with HCPT or the combined agents
(Fig. 1C). Unlike rapamycin, a
specific inhibitor of mTOR, HCPT was observed to partially inhibit
insulin-responsive mTOR activation (Fig. 1D). These results suggested that
the two agents impaired the activation of MAP4K3 and act on the
mTOR signaling pathway in a similar manner.
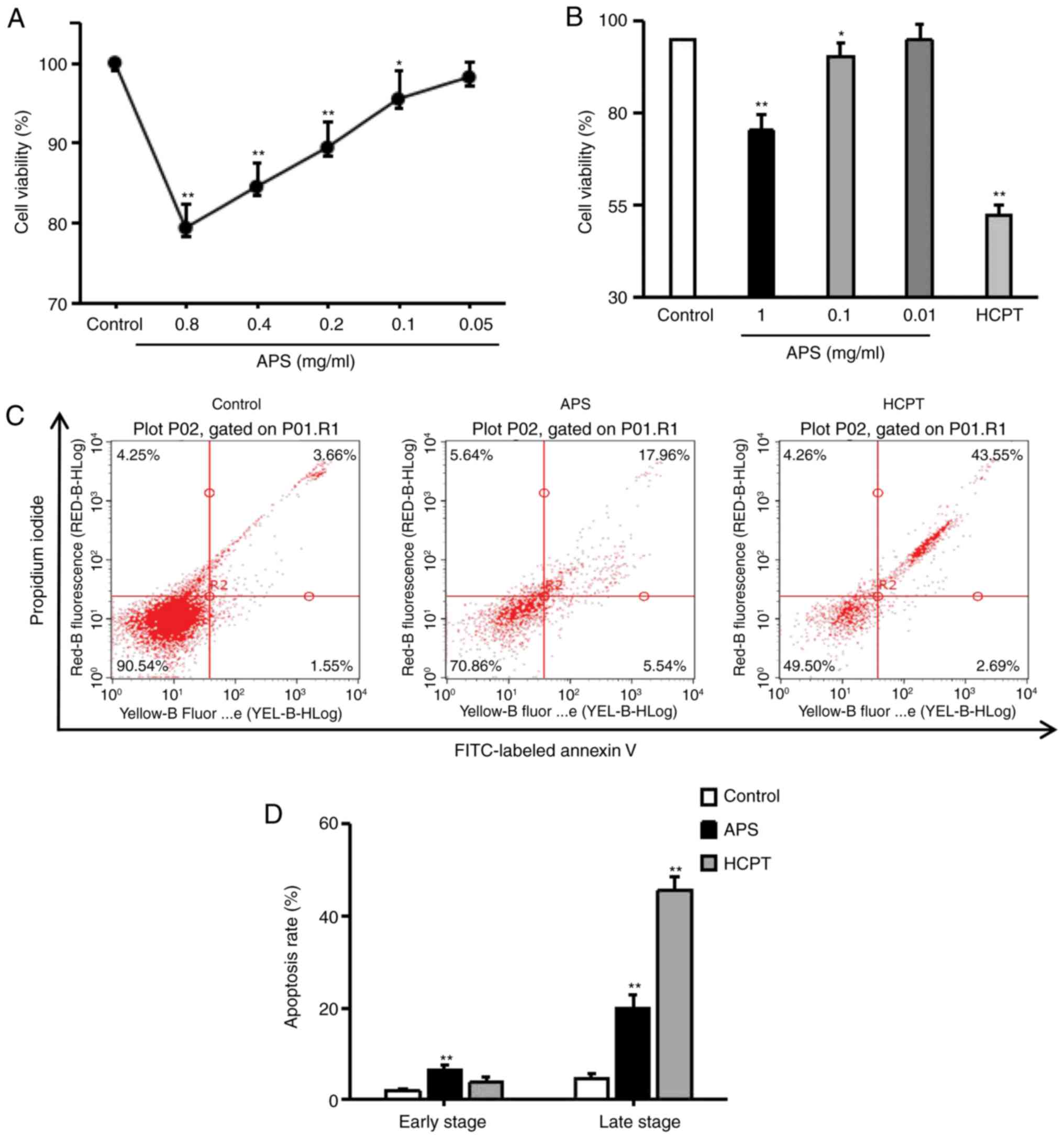
Effects of APS and HCPT on cell viability
and apoptosis
To detect the cytotoxic effects of APS, the H1299
cells were exposed to HCPT and varying of doses of APS for 24 h and
cell viability was examined using an MTT assay. As with HCPT, APS
moderately inhibited the growth and proliferation of H1299 cells
(Fig. 2A and B). The effects of
APS and HCPT on apoptosis were assessed using an Annexin V-FITC/PI
assay. APS significantly enhanced apoptosis in H1299 cells (early
apoptosis, 6.48±1.27%; late apoptosis, 19.88±2.89%) and HCPT
induced late apoptosis, up to 45.59±2.91% (Fig. 2C and D), indicating that APS and
HCPT inhibited cell proliferation by promoting apoptosis in H1299
cells.
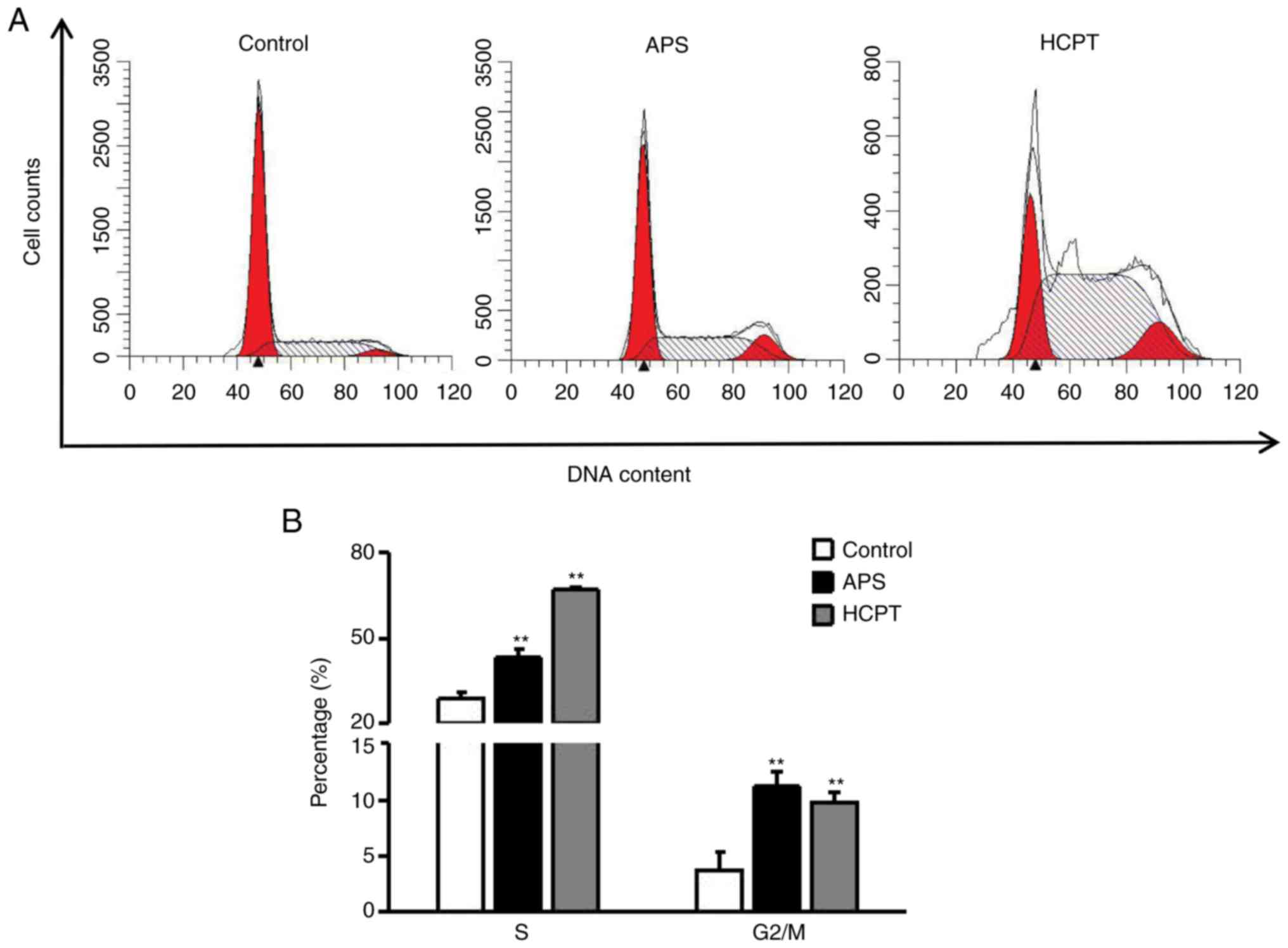
Flow cytometry was used to distinguish between cells
in different phases of the cell cycle. As shown in Fig. 3A and B, following APS treatment,
the percentages of cells in the S and G2/M phases increased to
42.94±3.09 and 11.12±1.32%, respectively, and following HCPT
treatment these proportions were 66.57±2.93 and 9.73±1.39%,
respectively, indicating that HCPT and APS treatment induced S
phase and G2/M phase arrest. These results further suggested that
APS and HCPT induced H1299 cell death by altering the cell cycle
distribution.
APS induces mitochondria-mediated
apoptosis and HCPT induces autophagy in H1299 cells
The upregulation of caspase-3, cytochrome c
and the Bax/Bcl-2 and caspase-9/pro-caspase-9 ratios were observed
following treatment of the H1299 cells with APS (Fig. 4A-D). The results indicated that
APS treatment induced H1299 cell death through the release of
cytochrome c from the mitochondria.
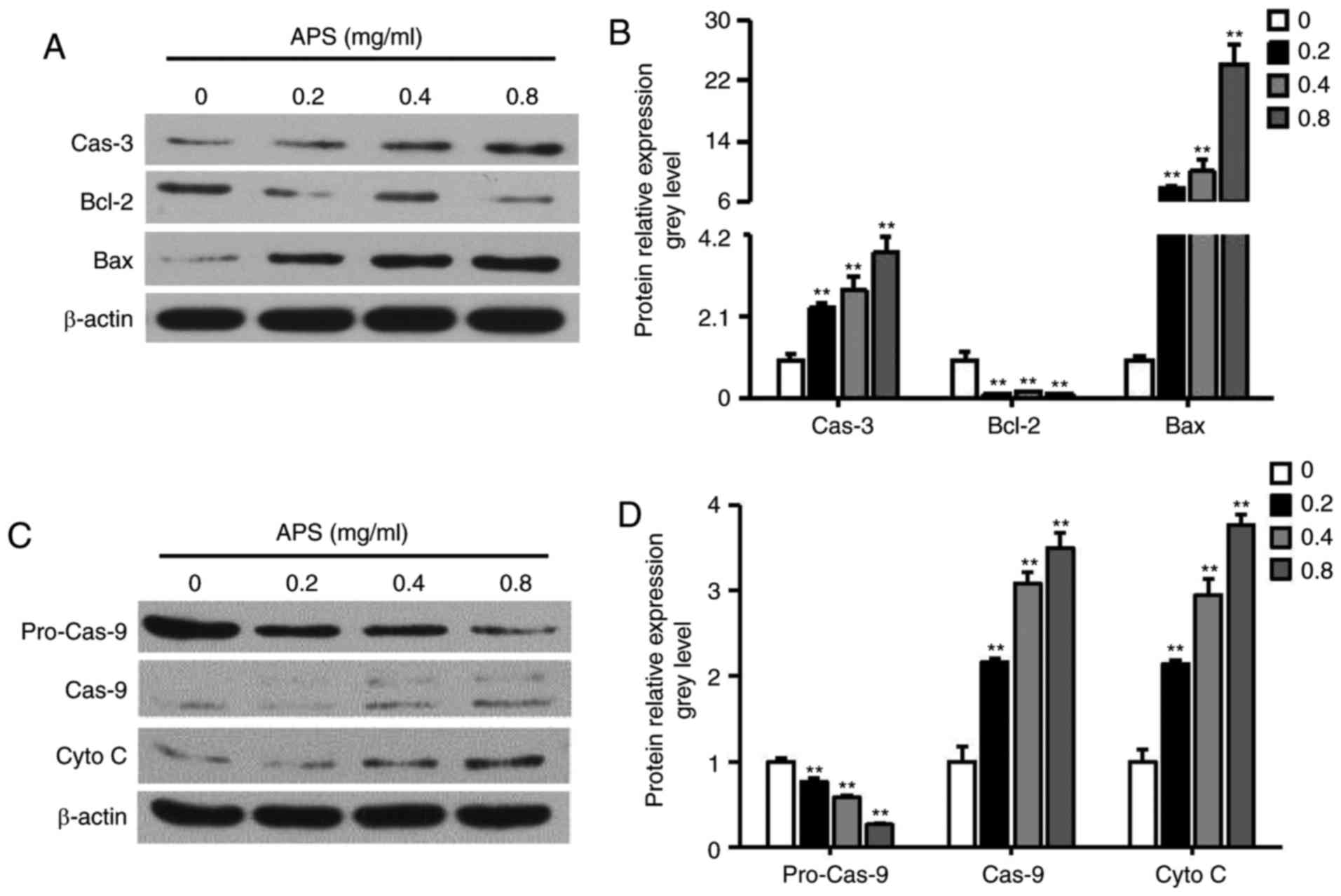 | Figure 4Effects of APS on the mitochondrial
apoptotic pathway in H1299 cells. H1299 cells were incubated with
0.2-0.8 mg/ml APS for 24 h. (A) Western blot analysis was performed
to evaluate levels of Bax, Bcl-2, and Cas-3, with (B)
quantification of results shown in the graph. (C) Western blot
analysis was performed to evaluate levels of pro-Cas-9, Cas-9 and
Cyto C, with (D) quantification of results shown in the graph. Data
are presented as the mean ± standard error of the mean of three
independent experiments. APS, Astragalus polysaccharide;
HCPT, 10-hydroxycamptothecin; Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X protein; Cas, caspase; Cyto C, cytochrome
c. *P<0.05, vs. control. |
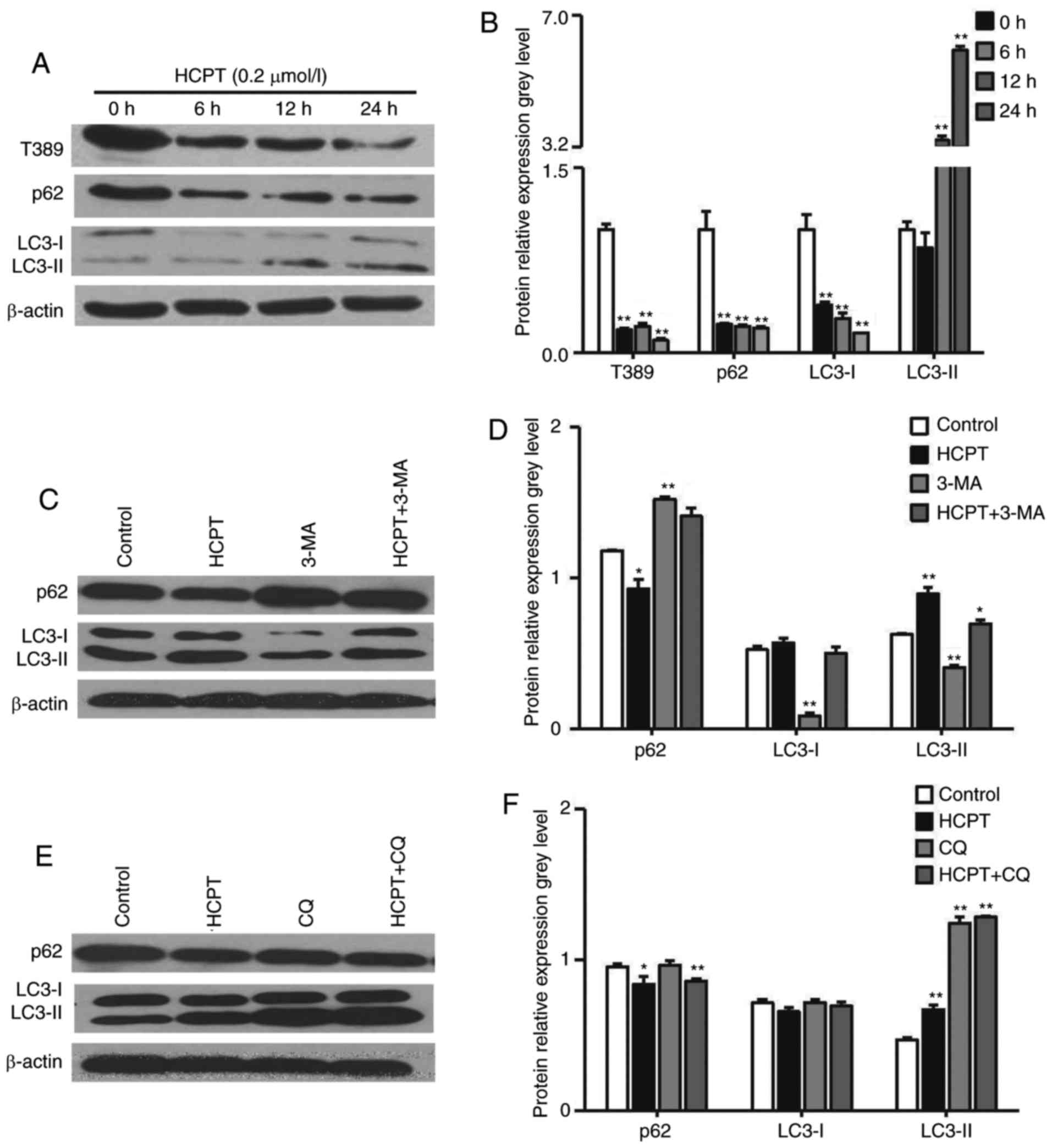
As inhibition of the Akt/mTOR/S6K signaling pathway
is linked to the induction of autophagy (23), the present study examined whether
HCPT induces autophagy by inhibiting this pathway. Autophagy is
activated by the increased expression of LC3-II, which controls the
completion of autophagosome formation. P62 is selectively
incorporated into autophagosomes through directly binding to LC3
and is efficiently degraded during autophagy. Therefore, the total
cellular expression levels of P62 is inversely correlated with the
autophagic activity. In the present study, it was demonstrated that
HCPT induced the formation of LC3-II and decreased the expression
of P62 following treatment with HCPT for 6, 12 and 24 h (Fig. 5A and B). To validate and interpret
this result, western blot analysis was performed with the control
extracts harvested from cells treated with 3-MA, a phosphoinositide
3-kinase inhibitor, which prevents the induction of autophagosomes
and inhibits autophagic flux, and CQ, which inhibits the fusion of
autophagosomes and lysosomes, inhibiting autophagy and leading to
LC3 accumulation. Compared with the H1299 cells incubated with HCPT
alone, pretreatment with 3-MA markedly decreased the levels of
LC3-II and increased the expression of P62 (Fig. 5C and D), whereas pretreatment with
CQ markedly increased the levels of LC3-II (Fig. 5E and F), suggesting that HCPT
treatment induced autophagic flux.
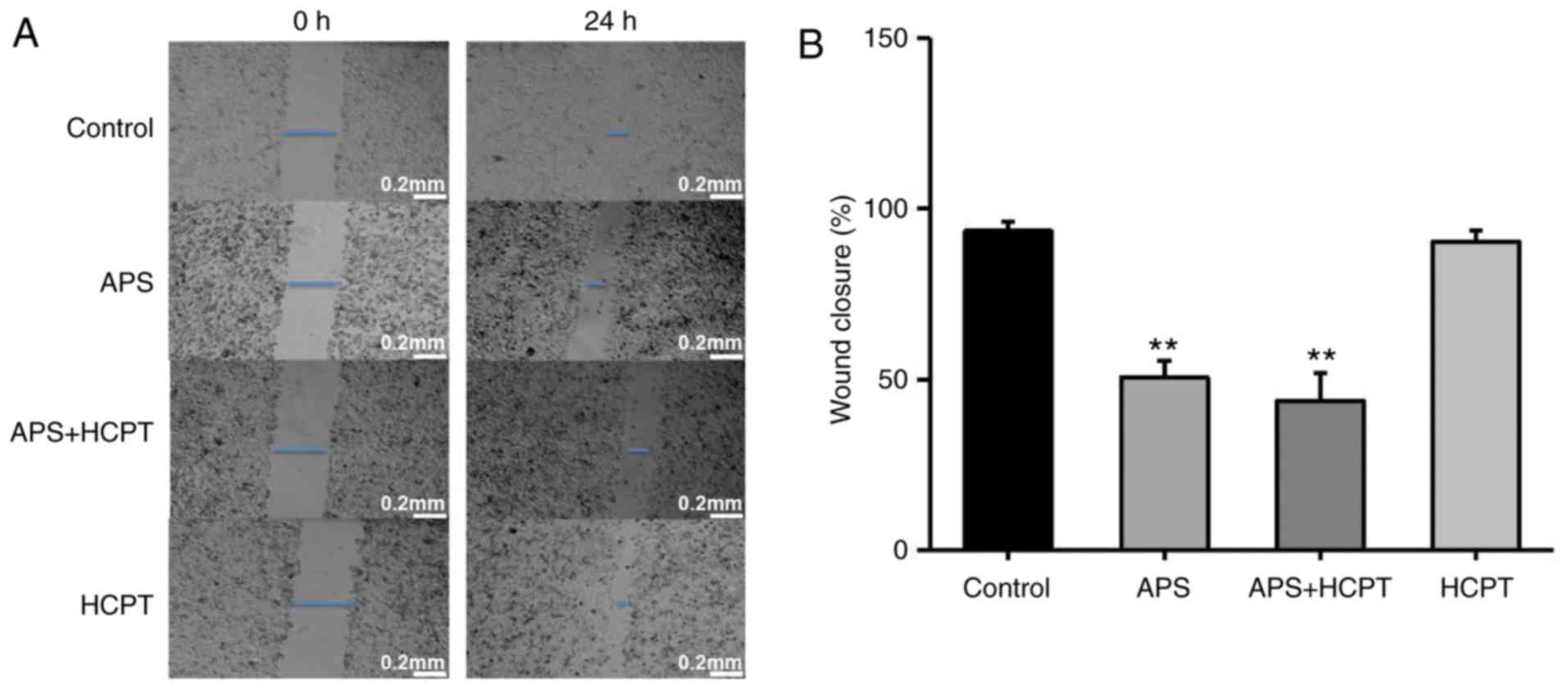
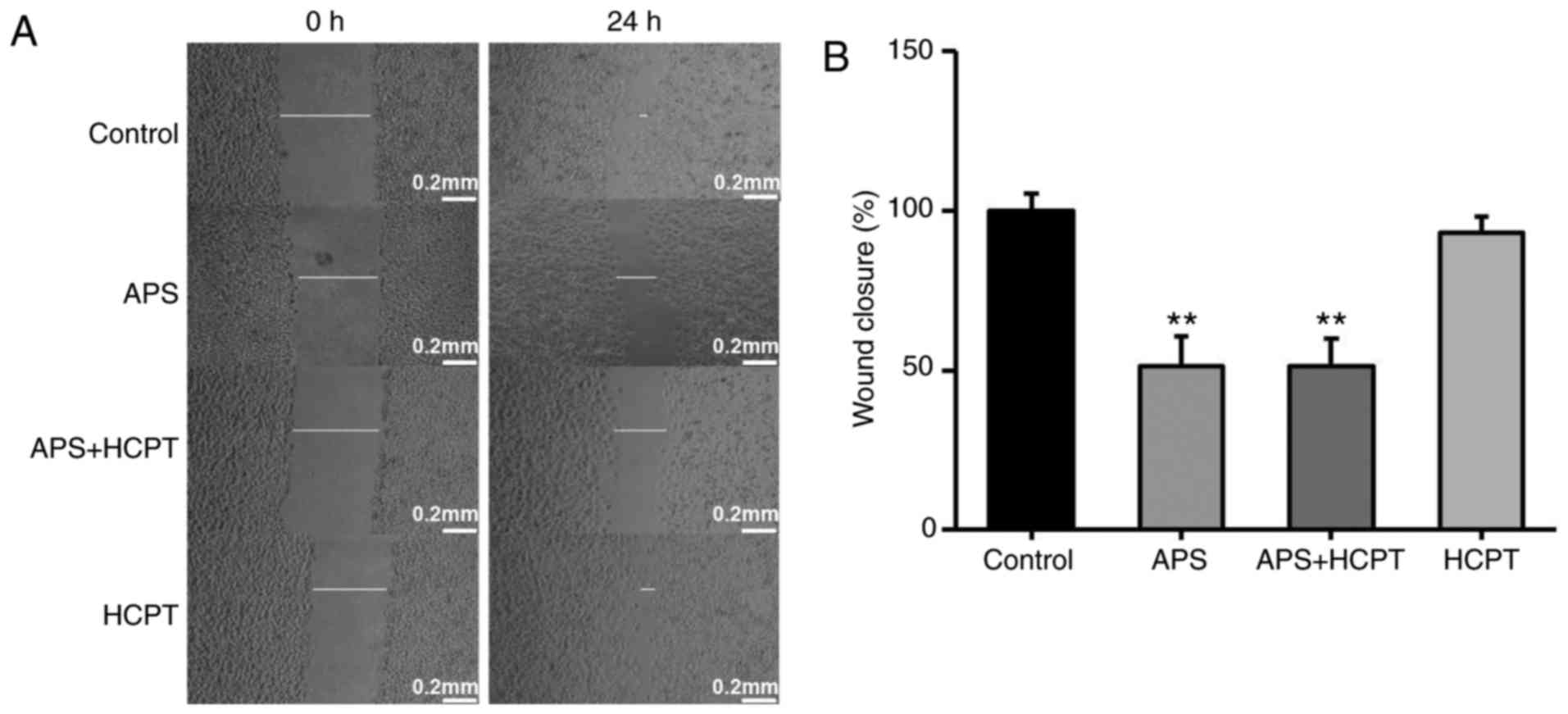
APS and HCPT inhibit the migration and
invasion of H1299 cells
To assess the effect of APS and HCPT on migration
and invasion of H1299 cells, a gradual reduction in the area of
wounding was observed in the control cells incubated with DMEM for
24 h. Treatment of the H1299 cells with APS resulted in significant
inhibition of this migration, whereas HCPT treatment did not affect
migration (Fig. 6A and B). The
present study further assessed the effect of APS and HCPT on the
migration of H460 cells. APS and HCPT affected the migration of
H460 cells similar to that of the H1299 cells (Fig. 7A and B).
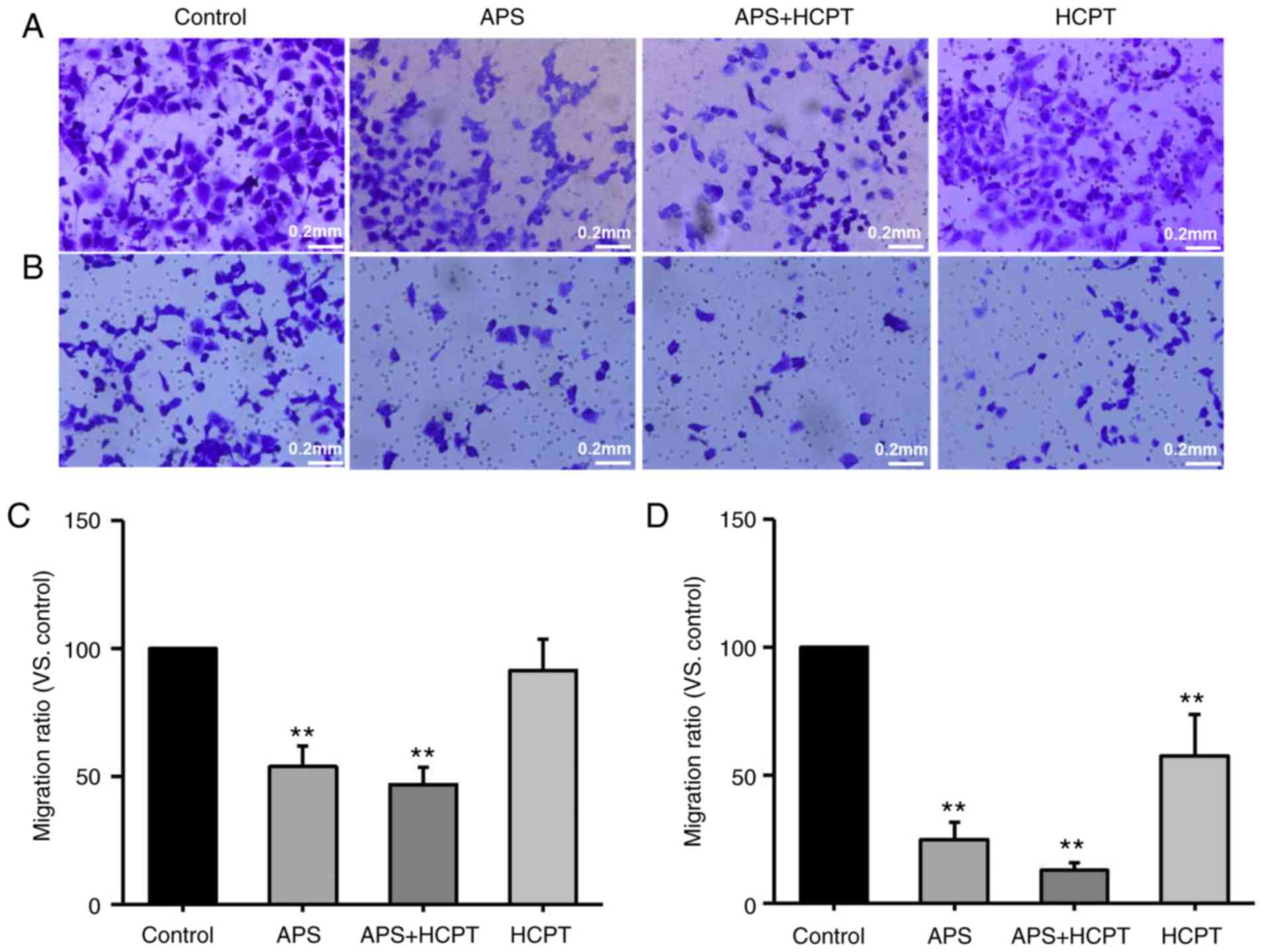
Transwell assays were also performed with
polycarbonate filters to analyze the migration and invasion of
H1299 (Fig. 8A-D). It was
demonstrated that APS and the combination of APS and HCPT inhibited
migration by 46.21±7.93 and 53.09±6.7%, respectively, whereas HCPT
only had a moderate effect on migration, indicating that treatment
with APS and the combined agents inhibited the migration of H1299
cells more effectively than HCPT (Fig. 8A and C). It was also observed that
treatment with APS, the combined agents and HCPT inhibited the
invasion of H1299 cells by 75.13±6.63, 87.22±3.01 and 42.52±16.36%,
respectively, compared with the control, suggesting that the
combination of the agents was more effective in inhibiting H1299
migration and invasion compared with APS treatment alone or HCPT
alone (Fig. 8B and D).
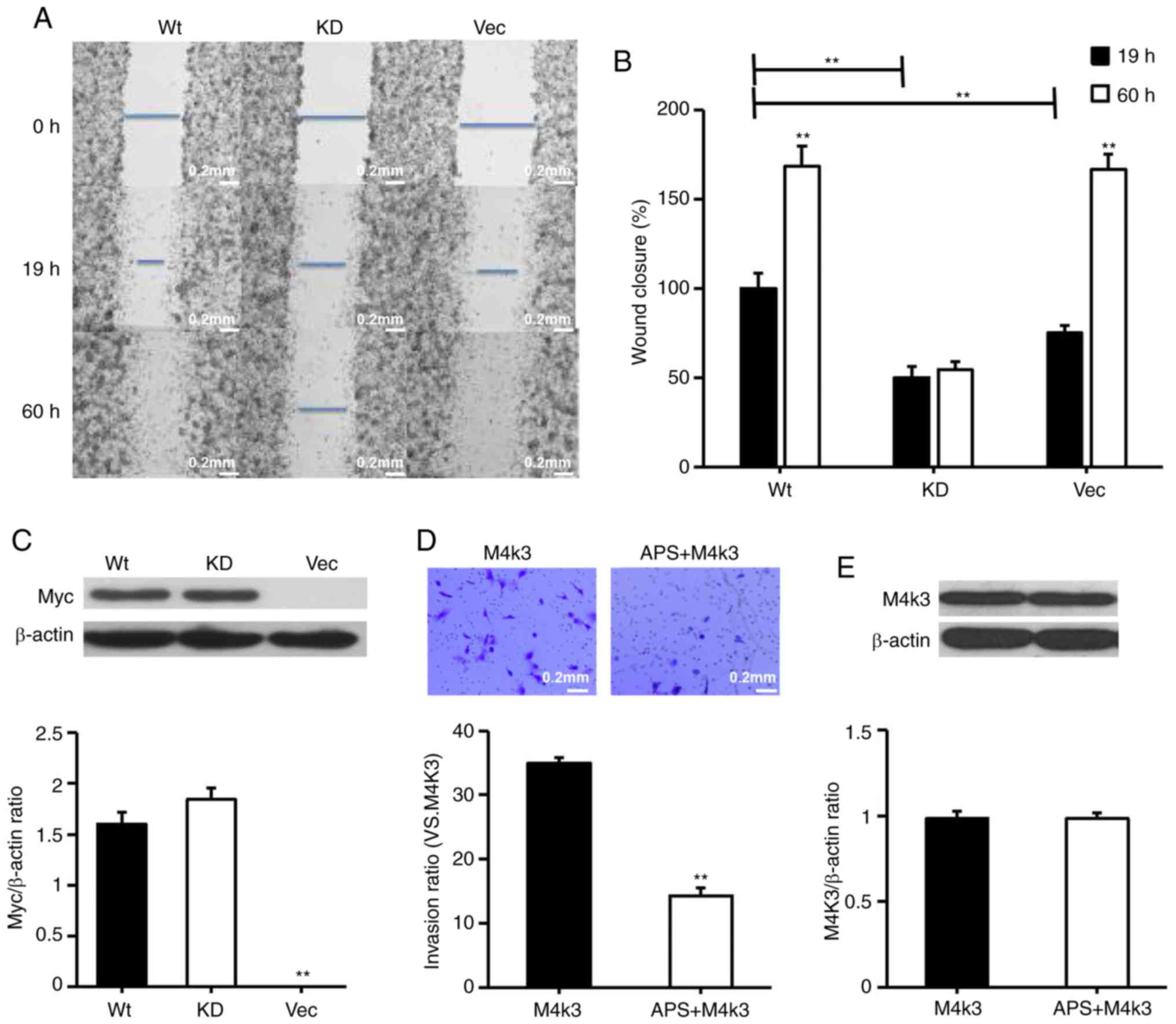
Overexpression of MAP4K3 increases the
migration and invasion of H1299 cells, and these effects are
reversed by APS treatment
The present study considered whether the
overexpression of MAP4K3 stimulates the migration of H1299 cells,
despite H1299 exhibit relatively high background expression of
MAP4K3. The H1299 cells were transfected with wild-type MAP4K3 and
KD MAP4K3, and it was demonstrated that cells transfected with
wild-type MAP4K3 led to more rapid closure of the wounded area
compared with the cells transfected with KD MAP4K3 or the vector
control (Fig. 9A-C). It was also
observed that cells transfected with wild-type MAP4K3 and the
vector control almost fully reversed the inflicted wound within a
60-h incubation period, whereas the cells transfected with the KD
MAP4K3 exhibited decreased migration, suggesting that the KD MAP4K3
behaves like a dominant negative mutation. The results also
suggested that, when cells transfected with wild-type MAP4K3 were
exposed to APS, their invasive properties were reduced (Fig. 9D and E). The overexpression of
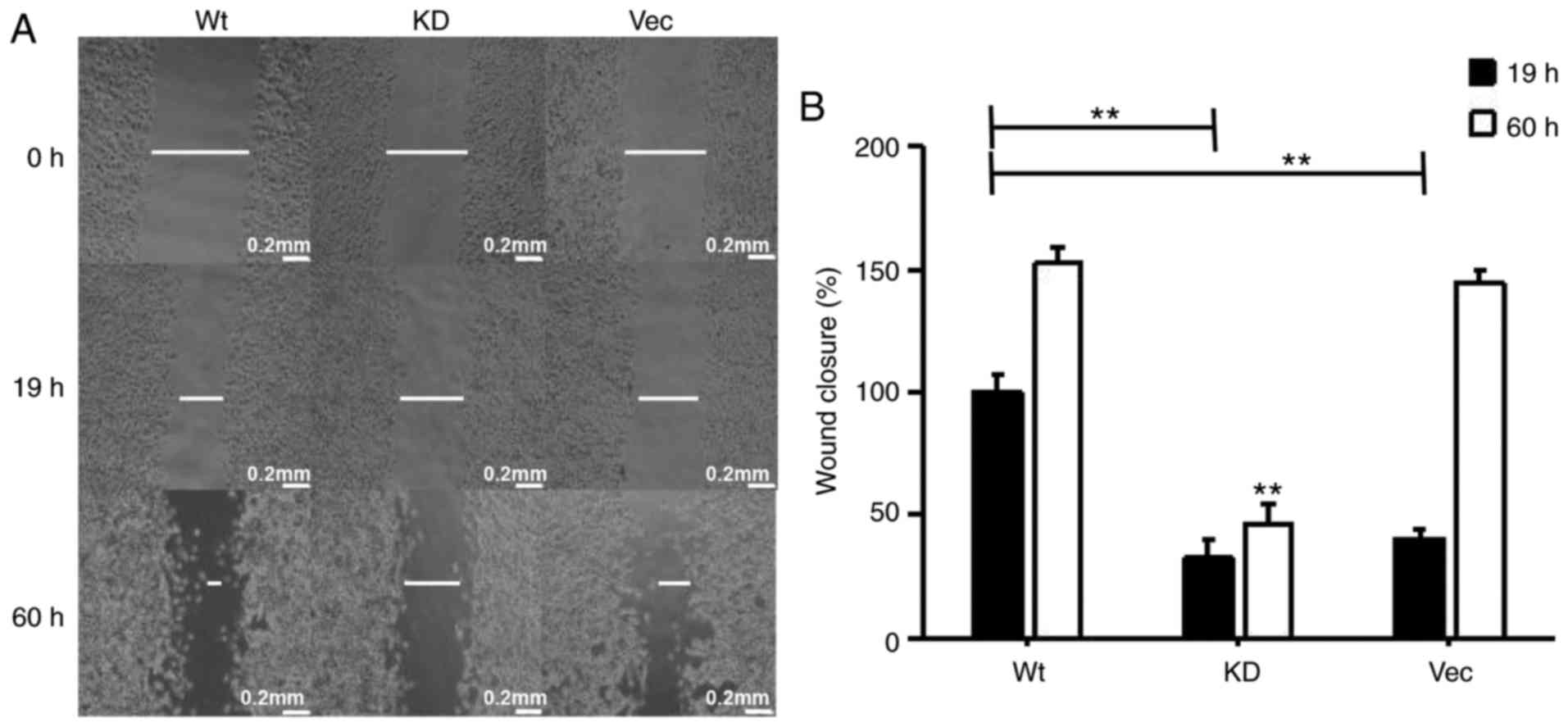
MAP4K3 also increased wound closure in the H460 cell line (Fig. 10A and B).
Discussion
Traditional Chinese medicines have been considered
to be effective in treatment for thousands of years, suggesting
that Chinese herbal medicines may serve as suitable candidates in
drug development. In the present study, the antitumor effects of
commonly used Chinese herbal medicines on H1299 NSCLC cells were
screened using an in vitro kinase assay based on the
activity of MAP4K3. Two candidates, APS and HCPT, were identified.
HCPT was initially used as a positive control in the present study.
In previous experiments, HCPT exhibited significant antitumor
activity against various tumors, including lung cancer cells, human
SMS-KCNR neuroblastoma cells, DU145-TxR prostate cancer cells, and
hepatoma, oral squamous cell carcinoma and breast cancer cells
(24-29). It was found that HCPT inhibited
MAP4K3 kinase activity in vitro and S6K phosphorylation at
Thr389 in H1299 cells, however, whether HCPT was able to induce
autophagy in H1299 cells remained to be elucidated. Therefore, the
expression of autophagy-associated proteins (LC3-I/-II and P62) was
assessed, and it was demonstrated that HCPT induced autophagy in
H1299 cells by the impairment of MAP4K3/mTOR signaling to S6K. As
reported previously for HeLa cells (30), HCPT induced autophagy in H1299
cells.
APS is a mixture of biological macromolecules
extracted from Astragalus spp. that exhibits a number of
biologically-relevant activities, including potent immunoregulatory
properties (31,32), the reduction of chemotoxicity
(33) and adverse
treatment-associated effects (34,35), and antitumor activities (36). APS can increase the expression of
Toll-like receptor 4 to enhance innate immunity during mucosal
bacterial infections in vivo and in vitro (37). It has also been used as an
adjuvant for an avian infectious bronchitis virus vaccine (38), it inhibits cell volume increases
in the myocardium, and has a protective effect on myocardial
ischemia and reperfusion injury/isoprenaline-treated cardiomyocytes
by inhibiting myocardial tissue apoptosis in vivo (39). However, APS treatment inhibited
proliferation and promoted apoptosis in H460 NSCLC cells (36), and combination treatment with
vinorelbine and cisplatin administered to patients with advanced
NSCLC significantly improved quality of life (40). Astragalus and Salvia
miltiorrhiza extract have been shown to inhibit cell migration
and invasion in HepG2 cells (41).
The versatile effects of APS on different cell and
tissue types in vivo and in vitro explain why the
root of Astragalus membranaceus, termed Huang Qi in
Mandarin, is one of the most commonly used Chinese herbal
medicines. The present study reported that APS induced
mitochondria-mediated apoptosis by increasing levels of
pro-apoptotic Bax, caspase-3, caspase-9 and cytochrome c,
and decreasing levels of Bcl-2 and pro-caspase-9 in H1299 cells. It
was also demonstrated that the combination treatment of APS and
HCPT was more effective compared with either APS or HCPT treatment
alone with respect to inhibiting the migration and invasion of
H1299 cells. Furthermore, it was observed that the effects of APS
and HCPT treatment on the migration of H460 were similar to those
on H1299 cells.
MAP4K3 is a member of the Ste20-related family,
named after Sterile 20 involved in pheromone signaling in
Saccharomyces cerevisiae (42). Members of this family are
characterized by a conserved kinase domain and a noncatalytic
region, which enables these proteins to interact with various
signaling molecules and regulatory proteins. MAP4K3 promotes cell
growth in human HeLa cells in culture in a similar manner to Ras
homolog enriched in brain and mTORC1 (43). Therefore, the present study
examined MAP4K3 inhibitors as potential tumor-selective therapeutic
agents by measuring the inhibition of the kinase in vitro.
It was found that APS and HCPT inhibited the activity of MAP4K3
in vitro, particularly HCPT, and the combined agents
exhibited more marked suppression of MAP4K3/mTOR signaling to S6K
in H1299 cells. It was also demonstrated that the overexpression of
wild-type MAP4K3 stimulated wound healing in vitro.
Therefore, MAP4K3 may be a potential therapeutic target for the
treatment of NSCLC.
Future preclinical studies may elucidate and
supplement the findings of the present study. However, target-based
drug screening is a viable approach and mechanism-based combination
regimens, including APS and HCPT, may be beneficial in the
treatment of NSCLC.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 31370764 to LY) and
the Key Project of Natural Science Foundation of Heilongjiang
Province (grant no. ZD2015007 to LY).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
Acquisition of data: TH, WL, YZ, LT and JL; analysis
and interpretation of data: LY, YZ and TH; statistical analysis: YZ
and TH; administrative and technical support: JL and FW; Provision
of reagents: XY; drafting of the manuscript: LY; obtained funding:
LY. LY conceived and supervised the study, planned experiments, and
took responsibility for the integrity of the data and the accuracy
of the data analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr Richard F. Lamb
at the Section of Cell and Molecular Biology, Chester Beatty
Laboratories, Institute of Cancer Research (London, UK), for
providing the pRK5mycMAP4K3 and AFG KD MAP4K3 plasmids.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanzaki R, Higashiyama M, Fujiwara A,
Tokunaga T, Maeda J, Okami J, Kozuka T, Hosoki T, Hasegawa Y,
Takami M, et al: Occult mediastinal lymph node metastasis in NSCLC
patients diagnosed as clinical N01 by preoperative integrated
FDG-PET/CT and CT: Risk factors, pattern, and histopathological
study. Lung Cancer. 71:333–337. 2011. View Article : Google Scholar
|
|
3
|
Spizzo G, Seeber A and Mitterer M: Routine
use of pamidronate in NSCLC patients with bone metastasis: Results
from a retrospective analysis. Anticancer Res. 29:5245–5249.
2009.
|
|
4
|
Fidler IJ, Kim SJ and Langley RR: The role
of the organ micro-environment in the biology and therapy of cancer
metastasis. J Cell Biochem. 101:927–936. 2007. View Article : Google Scholar
|
|
5
|
Shiraga M, Yano S, Yamamoto A, Ogawa H,
Goto H, Miki T, Miki K, Zhang H and Sone S: Organ heterogeneity of
host-derived matrix metalloproteinase expression and its
involvement in multiple-organ metastasis by lung cancer cell lines.
Cancer Res. 62:5967–5973. 2002.PubMed/NCBI
|
|
6
|
Nguyen DX and Massagué J: Genetic
determinants of cancer metastasis. Nat Rev Genet. 8:341–352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen M, Du Y, Qui M, Wang M, Chen K, Huang
Z, Jiang M, Xiong F, Chen J, Zhou J, et al: Ophiopogonin B-induced
autophagy in non-small cell lung cancer cells via inhibition of the
PI3K/Akt signaling pathway. Oncol Rep. 29:430–436. 2013. View Article : Google Scholar :
|
|
8
|
Saxton RA and Sabatini DM: mTOR Signaling
in Growth, Metabolism, and Disease. Cell. 169:361–371. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han B, Cui H, Kang L, Zhang X, Jin Z, Lu L
and Fan Z: Metformin inhibits thyroid cancer cell growth,
migration, and EMT through the mTOR pathway. Tumour Biol.
36:6295–6304. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Zhang HE and Liu Z: MicroRNA-147
suppresses proliferation, invasion and migration through the
AKT/mTOR signaling pathway in breast cancer. Oncol Lett.
11:405–410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang X, Ding Z, Mo J, Sang B, Shi Q, Hu
J, Xie S, Zhan W, Lu D, Yang M, et al: GOLPH3 promotes glioblastoma
cell migration and invasion via the mTOR-YB1 pathway in vitro. Mol
Carcinog. 54:1252–1263. 2015. View
Article : Google Scholar
|
|
12
|
Dai L, Zhuang L and Zhang B, Wang F, Chen
X, Xia C and Zhang B: DAG/PKCδ and IP3/Ca2+/CaMK IIβ operate in
parallel to each other in PLCγ1-driven cell proliferation and
migration of human gastric adenocarcinoma cells, through
Akt/mTOR/S6 pathway. Int J Mol Sci. 16:28510–28522. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Findlay GM, Yan L, Procter J, Mieulet V
and Lamb RF: A MAP4 kinase related to Ste20 is a nutrient-sensitive
regulator of mTOR signalling. Biochem J. 403:13–20. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan L, Mieulet V, Burgess D, Findlay GM,
Sully K, Procter J, Goris J, Janssens V, Morrice NA and Lamb RF:
PP2A T61 epsilon is an inhibitor of MAP4K3 in nutrient signaling to
mTOR. Mol Cell. 37:633–642. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao B, Han H, Chen J, Zhang Z, Li S, Fang
F, Zheng Q, Ma Y, Zhang J, Wu N and Yang Y: MicroRNA let-7c
inhibits migration and invasion of human non-small cell lung cancer
by targeting ITGB3 and MAP4K3. Cancer Lett. 342:43–51. 2014.
View Article : Google Scholar
|
|
16
|
Park J, Lee J and Choi C: Evaluation of
drug-targetable genes by defining modes of abnormality in gene
expression. Sci Rep. 5:135762015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hsu CP, Chuang HC, Lee MC, Tsou HH, Lee
LW, Li JP and Tan TH: GLK/MAP4K3 overexpression associates with
recurrence risk for non-small cell lung cancer. Oncotarget.
7:41748–41757. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fu J, Wang Z, Huang L, Zheng S, Wang D,
Chen S, Zhang H and Yang S: Review of the botanical
characteristics, phytochemistry, and pharmacology of Astragalus
membranaceus (Huangqi). Phytother Res. 28:1275–1283. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Liu F, Yang Y, Li D, Lv J, Ou Y,
Sun F, Chen J, Shi Y and Xia P: Astragalus polysaccharide
ameliorates ionizing radiation-induced oxidative stress in mice.
Int J Biol Macromol. 68:209–214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Han R: Highlight on the studies of
anticancer drugs derived from plants in China. Stem Cells.
12:53–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Allen JJ, Li M, Brinkworth CS, Paulson JL,
Wang D, Hübner A, Chou WH, Davis RJ, Burlingame AL, Messing RO, et
al: A semisynthetic epitope for kinase substrates. Nat Methods.
4:511–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mizushima N and Yoshimori T: How to
interpret LC3 immunoblotting. Autophagy. 3:542–545. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Che X, Yan H, Sun H, Dongol S, Wang Y, Lv
Q and Jiang J: Grifolin induces autophagic cell death by inhibiting
the Akt/mTOR/S6K pathway in human ovarian cancer cells. Oncol Rep.
36:1041–1047. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Z, Zheng Q, Chen W, Wu M, Pan G, Yang
K, Li X, Man S, Teng Y, Yu P and Gao W: Chemosensitizing effect of
Paris Saponin I on Camptothecin and 10-hydroxycamptothecin in lung
cancer cells via p38 MAPK, ERK, and Akt signaling pathways. Eur J
Med Chem. 125:760–769. 2017. View Article : Google Scholar
|
|
25
|
Yuan ZF, Tang YM, Xu XJ, Li SS and Zhang
JY: 10-Hydroxycamptothecin induces apoptosis in human neuroblastoma
SMS-KCNR cells through p53, cytochrome c and caspase 3 pathways.
Neoplasma. 63:72–79. 2016. View Article : Google Scholar
|
|
26
|
Liu Z, Zhu G, Getzenberg RH and Veltri RW:
The upregulation of PI3K/Akt and MAP kinase pathways is associated
with resistance of microtubule-targeting drugs in prostate cancer.
J Cell Biochem. 116:1341–1349. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mallery SR, Shenderova A, Pei P, Begum S,
Ciminieri JR, Wilson RF, Casto BC, Schuller DE and Morse MA:
Effects of 10-hydroxycamptothecin, delivered from locally
injectable poly(lactide-co-glycolide) microspheres, in a murine
human oral squamous cell carcinoma regression model. Anticancer
Res. 21:1713–1722. 2001.PubMed/NCBI
|
|
28
|
Liu W and Zhang R: Upregulation of
p21WAF1/CIP1 in human breast cancer cell lines MCF-7 and MDA-MB-468
undergoing apoptosis induced by natural product anticancer drugs
10-hydroxycamptothecin and camptothecin through p53-dependent and
independent pathways. Int J Oncol. 12:793–804. 1998.PubMed/NCBI
|
|
29
|
Zhang XW, Qing C and Xu B: Apoptosis
induction and cell cycle perturbation in human hepatoma hep G2
cells by 10- hydroxycamptothecin. Anticancer Drugs. 10:569–576.
1999. View Article : Google Scholar
|
|
30
|
Cheng YX, Zhang QF, Pan F, Huang JL, Li
BL, Hu M, Li MQ and Chen Ch: Hydroxycamptothecin shows antitumor
efficacy on HeLa cells via autophagy activation mediated apoptosis
in cervical cancer. Eur J Gynaecol Oncol. 37:238–243.
2016.PubMed/NCBI
|
|
31
|
Yoshida Y, Wang MQ, Liu JN, Shan BE and
Yamashita U: Immunomodulating activity of Chinese medicinal herbs
and Oldenlandia diffusa in particular. Int J Immunopharmacol.
19:359–370. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lai X, Xia W, Wei J and Ding X:
Therapeutic effect of Astragalus polysaccharides on hepatocellular
carcinoma H22-bearing mice. Dose Response. 15:15593258166851822017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cassileth BR, Rizvi N, Deng G, Yeung KS,
Vickers A, Guillen S, Woo D, Coleton M and Kris MG: Safety and
pharmacokinetic trial of docetaxel plus an Astragalus-based herbal
formula for non-small cell lung cancer patients. Cancer Chemother
Pharmacol. 65:67–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Duan P and Wang ZM: Clinical study on
effect of Astragalus in efficacy enhancing and toxicity reducing of
chemotherapy in patients of malignant tumor. Zhongguo Zhong Xi Yi
Jie He Za Zhi. 22:515–517. 2002.In Chinese.
|
|
35
|
Cao Y, Ruan Y, Shen T, Huang X, Li M, Yu
W, Zhu Y, Man Y, Wang S and Li J: Astragalus polysaccharide
suppresses doxorubicin-induced cardiotoxicity by regulating the
PI3k/Akt and p38MAPK pathways. Oxid Med Cell Longev.
2014.674219:2014.
|
|
36
|
Zhang JX, Han YP, Bai C and Li Q: Notch1/3
and p53/p21 are a potential therapeutic target for APS-induced
apoptosis in non-small cell lung carcinoma cell lines. Int J Clin
Exp Med. 8:12539–12547. 2015.PubMed/NCBI
|
|
37
|
Yin X, Chen L, Liu Y, Yang J, Ma C, Yao Z,
Yang L, Wei L and Li M: Enhancement of the innate immune response
of bladder epithelial cells by Astragalus polysaccharides through
up regulation of TLR4 expression. Biochem Biophys Res Commun.
397:232–238. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang P, Wang J, Wang W, Liu X, Liu H, Li
X and Wu X: Astragalus polysaccharides enhance the immune response
to avian infectious bronchitis virus vaccination in chickens.
Microb Pathog. 111:81–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu D, Chen L, Zhao J and Cui K:
Cardioprotection activity and mechanism of Astragalus
polysaccharide in vivo and in vitro. Int J Biol Macromol.
111:947–952. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Guo L, Bai SP, Zhao L and Wang XH:
Astragalus polysaccharide injection integrated with vinorelbine and
cisplatin for patients with advanced non-small cell lung cancer:
Effects on quality of life and survival. Med Oncol. 29:1656–1662.
2012. View Article : Google Scholar
|
|
41
|
Liu X, Yang Y, Zhang X, Xu S, He S, Huang
W and Roberts MS: Compound Astragalus and Salvia miltiorrhiza
extract inhibits cell invasion by modulating transforming growth
factor-beta/Smad in HepG2 cell. J Gastroenterol Hepatol.
25:420–426. 2010. View Article : Google Scholar
|
|
42
|
Leberer E, Dignard D, Harcus D, Thomas DY
and Whiteway M: The protein kinase homologue Ste20p is required to
link the yeast pheromone response G-protein beta gamma subunits to
downstream signalling components. EMBO J. 11:4815–4824. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yan L and Lamb RF: Signalling by amino
acid nutrients. Biochem Soc Trans. 39:443–445. 2011. View Article : Google Scholar : PubMed/NCBI
|