Introduction
Primary colon adenocarcinoma is one of the leading
causes of cancer-associated mortality worldwide (1). Age, sex, diet and genetic factors
have implications in the development of colon adenocarcinoma
(2). Although, a number of
treatment approaches, including surgery and chemotherapy, are used
for the management of this type of cancer, the five-year survival
rates remain poor (3). The late
diagnosis and unavailability of reliable biomarkers and therapeutic
targets form a major obstacle in the treatment of colon
adenocarcinoma (4). MicroRNAs are
small RNA molecules, which do not code for any protein and have
been found to be involved in a number of cellular processes
(5). MicroRNAs have also been
implicated in the development of a number of diseases, which
include, but are not limited to, cancer (6). The expression of several microRNAs
has been reported to be dysregulated in cancer cells and microRNAs
are being considered as important therapeutic targets/agents
(7). MicroRNA (miR)-145 is one
such microRNA, which has been found to be dysregulated in a number
of cancer types. For example, the expression of miR-145 has been
reported to be significantly downregulated in gastric and prostate
cancer (1,8). Furthermore, miR-145 has been shown
to be involved in the regulation of proliferation, migration and
invasion of cancer cells; for example, Sachdeva and Mo reported
that miR-145 inhibits the proliferation and metastasis of breast
cancer cells (9). However, the
role and therapeutic potential of miR-145 have not been
investigated in primary colon adenocarcinoma cells. In the present
study, the expression of miR-145 in normal and primary colon
adenocarcinoma cells was examined. It was found that the expression
of miR-145 was significantly downregulated in all colon
adenocarcinoma cell lines. Furthermore, the ectopic expression of
miR-145 in colon adenocarcinoma cells inhibited their proliferation
via the induction of apoptosis and suppression of migration and
invasion by targeting mitogen-activated protein kinase 1 (MAPK1).
The expression of MAPK1 has been reported to be upregulated in a
number of types of cancer and several anticancer agents have been
shown to target MAPK1 (10). In
the present study, the effects of the overexpression of miR-145
were also evaluated in xenografted mice and it was observed that
miR-145 inhibited tumor growth in vivo. Taken together, the
results suggested that miR-145 may prove to be an important
therapeutic target/agent for the management of colon
adenocarcinoma.
Materials and methods
Cell lines and culture conditions
The normal colon cell line (CCD-18Co) and primary
colon adenocarcinoma cell lines (Hs255.T, SW480, Hs257.T, HT29,
LS174T, SNU-C1, and DLD-1) were procured from American Type Culture
Collection (Manassas, VA, USA). All cell lines were maintained in
Dulbecco’s modified Eagle’s medium supplemented with 10% fetal
bovine serum (Thermo Fisher Scientific, Inc., Waltham, MA, USA),
antibiotics (100 U/ml penicillin and 100 µg/ml streptomycin), and 2
mM glutamine. The cells were cultured in a CO2 incubator
(Thermo Fisher Scientific, Inc.) at 37°C with 98% humidity and 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The total RNA was extracted from the primary colon
adenocarcinoma cell lines (Hs255.T, SW480, Hs257.T, HT29, LS174T,
SNU-C1, DLD-1) and normal cell line (CCD-18Co) with the assistance
of RNeasy kits (Qiagen, Inc., Valencia, CA, USA). To reverse
transcribe the cDNA, the Omniscript RT (Qiagen, Inc.) was employed
using 1 µg of the extracted RNA. The cDNA was then used as a
template for RT-qPCR analysis with the assistance of the Taq PCR
Master Mix kit (Qiagen, Inc.) according to the manufacturer’s
protocol. The reaction mixture consisted of 20 µl containing
1.5 mM MgCl2, 2.5 units Taq DNA Polymerase, 200
µM dNTP, 0.2 µM of each primer and 0.5 µg DNA.
The cycling conditions were as follows: 95°C for 20 sec, followed
by 40 cycles of 95°C for 15 sec, and 58°C for 1 min. GAPDH was used
as an internal control and the relative quantification
(2−ΔΔCq) method was used to evaluate the quantitative
variation between the samples as described previously (11). The primers used in the present
study are presented in Table
I.
 | Table IList of primers used for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
List of primers used for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Primer sequence |
|---|
| MAPK1 | F:
5′-TGGATTCCCTGGTTCTCTCTAAAG-3′ |
| R:
5′-GGGTCTGTTTTCCGAGGATGA-3′ |
| miR-145 | F:
5′-ACACTCCAGCTGGGCAGGTCAAAAGGGTCC-3′ |
| R:
5′-TGCGGGTGCTCGCTTCGGCAGC-3′ |
| GAPDH | F:
5′-GAAGGTGAAGGTCGGAGTC-3′ |
| R:
5′-GAAGATGGTGATGGGATTTC-3′ |
Cell transfection
When the primary colon adenocarcinoma SW480 cells
reached 80% confluence, they were transfected with mimics-negative
control (NC; 5′-GUAGGAGUAGUGAAAGGCC-3′), miR-145 mimics
(5′-GUCCAGUUUUCCCAGGAAUCCCU-3′), or miR-145 inhibitor
(5′-AAGGGAUUCCUGGGAAACUGGAC-3′), from Shanghai GenePharma
(Shanghai, China; 10 pmol), small interfering (si)-negative control
(si-NC) (5′-CGAACUCACUGGUCUGACC-3′), siRNA-MAPK1
(5′-AGUUCGAGUAUACUUCAAGUU-3′) and pcDNA-MAPK1 (2 µg, Taijin
Saier Biotechnology, China) with Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) as per the manufacturer’s
protocol.
Cell viability and colony formation
assays
The cell viability of the primary SW480
adenocarcinoma cells was assessed using theWST-1 colorimetric
assay. Briefly, the SW480 primary adenocarcinoma cells were seeded
in 96-well plates at the density of 2×105 cells/well.
The cells were then incubated with WST-1 at 37°C for 4 h. The
absorbance at 450 nm was then measured using a microplate reader to
determine the viability of primary adenocarcinoma cells. For
assessment of the colony formation potential of the SW480 cells,
the cells were collected at the exponential phase of growth and
were then counted using a hemocytometer. The platting of the
transfected cells was performed at 200 cells/well. The plates were
then maintained at 37°C for 6 days. Following incubation of the
cells for 6 days, they were subjected to washing with PBS and
fixation with methanol. The SW480 cells were then stained with
crystal violet, and analyzed by light microscopy (Olympus
Corporation, Tokyo, Japan).
Detection of apoptosis
The nuclear morphology of the SW480 primary
adenocarcinoma cells was assessed by fluorescence microscopy
following subjecting the cells to DAPI, AO/EB, Annexin V/PI
staining for determination of the apoptotic cell populations, as
described previously (12,13).
Target identification and dual luciferase
assay
To identify the target, miR-145 was subjected to the
online software TargetScan version 7.2 (http://www.targetscan.org). For the luciferase
reporter assay, the binding sites of the wild-type (WT) and the
mutated (MUT) MAPK1 3′-UTR were cloned into the downstream region
of the luciferase gene in the PGL3-REPORT luciferase vector
(Invitrogen; Thermo Fisher Scientific, Inc.). The cells were then
subjected to co-transfection with the WT and MUT PGL3-MAPK1-3′ UTR
vectors and the miR-145 mimics. Finally, the luciferase activity
was determined using a Luciferase Reporter Assay kit (Promega
Corporation, Madison, WI, USA) as per the manufacturer’s protocol,
and Renilla luciferase activity was used to normalize the data.
Cell migration and invasion assay
The cell migration and invasion of the SW480 cells
were investigated using the method of Wang et al, as
described previously (14).
Western blot analysis
The primary colon adenocarcinoma cells were
transfected with different constructs and then centrifuged at 400 g
for 5 min at 4°C, subjected to PBS washing and then subsequently
lysed in RIPA buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM
NaCl, 1% Triton X-100, 0.1% SDS, 5 mM EDTA, 30 mM
Na2HPO4, 50 mM NaF, 0.5 mM NaVO4,
2 mM phenyl methylsulfonyl fluoride, and 10% protease cocktail
inhibitor. This was followed by incubation for 30 min and the
supernatant containing proteins were harvested by centrifugation at
13,000 g for 15 min at 4°C. Protein concentration was determined by
the Bradford method (15) and
stored at −80°C. For western blot analysis, equal quantities of
proteins (50 µg) were loaded and resolved on an
SDS-polyacrylamide gel (10%). Following gel electrophoresis, the
proteins were transferred onto polyvinylidene difluoride membranes.
The membranes were blocked in blocking buffer (10 mM Tris-HCl, 150
mM NaCl, 0.1% Tween-20) containing 5% non-fat milk for 1 h at room
temperature and blotted with MAPK1 primary antibodies (1:1,000;
anti-MAPK1, cat. no. sc-136288, Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) overnight at 4°C. The blots were washed in TBS,
incubated with horseradish peroxidase-conjugated secondary antibody
(1:1,000; cat. no. sc-516087, Santa Cruz Biotechnology, Inc.) for 1
h at 4°C, washed again three times with TBS and chemiluminescence
was captured on hyperfilm following incubating the blots in ECL
plus solution.
Mice xenografts
The 4-week-old male BALB/c nude mice weighing 18±2 g
were obtained from the animal holding capacity of the Shanghai
Hospital of Traditional Chinese Medicine and maintained per the
National Institutes of Health standards for the care and use of
laboratory animals. The study was approved and supervised by the
Ethics Committee of Shanghai Hospital of Traditional Chinese
Medicine (Shanghai, China) under approval number ECSHT-AS345/2017.
The animals were provided free access to a pellet diet, and water
was freely available. The animals were maintained in
well-ventilated rooms with a controlled environment of a day: night
light/dark cycle and temperature of 28±2°C. The mice were randomly
divided into two groups (n=15 per group). SW480 cell clones
(~1.0×107 cells/mouse), were subcutaneously injected
with miR-145 mimics or miR-NC into the back of the mice. The mice
were sacrificed by deep anesthesia with isoflurane at different
time points and sacrifice was verified by monitoring breathing and
heartbeat. The tumors were extracted and tumor weight and volume
were measured. The tumor volume was determined using the following
formula: V = (W x W x L)/2; where ‘V’ is the volume, ‘W’ is the
width and ‘L’ is the length of the tumor. RNA and proteins were
extracted from the tumor tissues for further experimentation. Each
group included five mice and experiments were repeated three
times.
Statistical analysis
The experiments were performed three times and the
values presented as the mean ± standard deviation of the three
replicates. Student’s t-test (for comparison between two samples)
and one-way analysis of variance followed by Tukey’s test (for
comparison between more than two samples) were used for statistical
analysis using GraphPad Prism software (version 7; GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression analysis of miR-145 in primary
colon adenocarcinoma cells
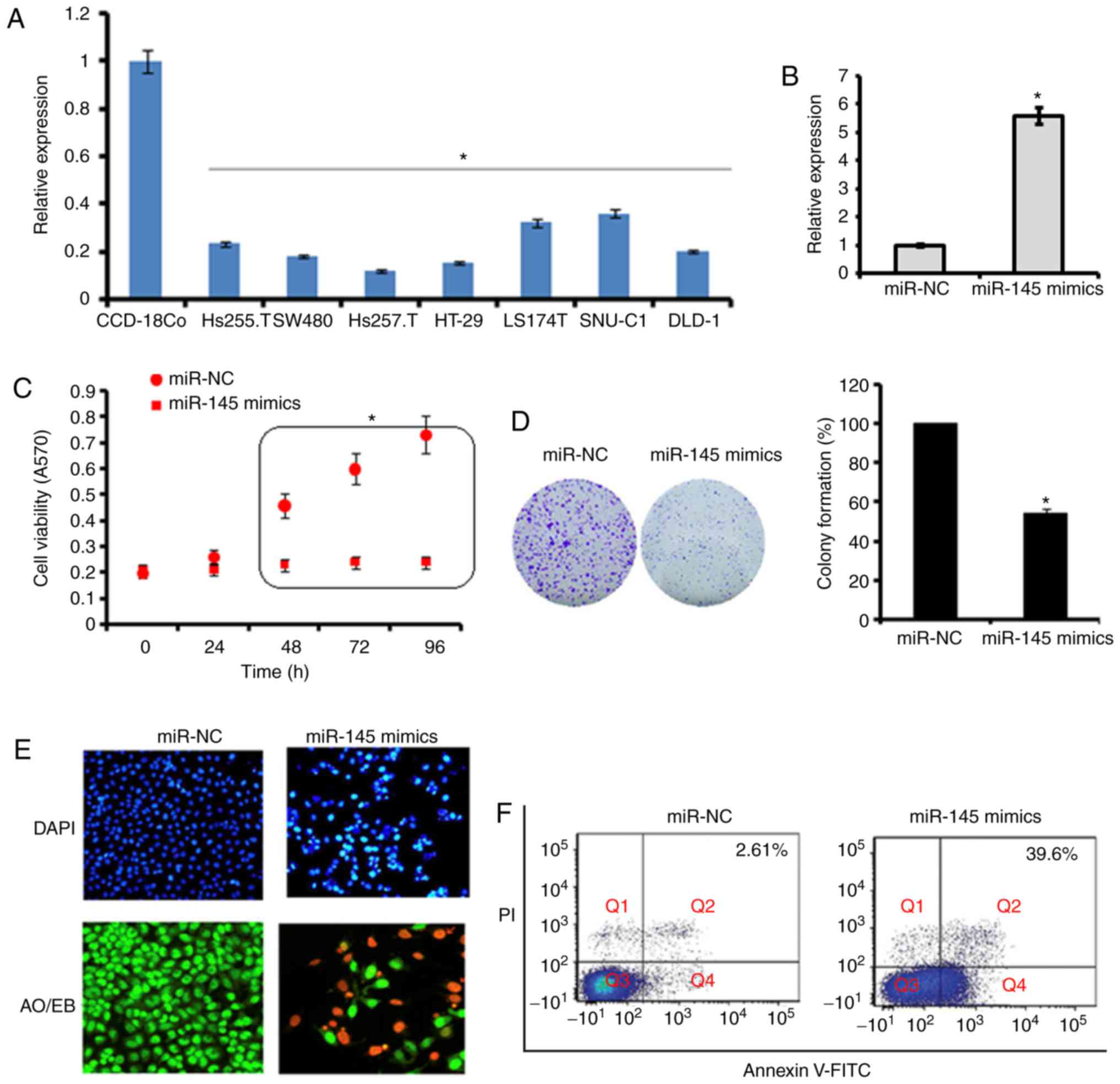
The expression of miR-145 was examined in seven
primary colon adenocarcinoma cell lines (Hs255.T, SW480, Hs257.T,
HT29, LS174T, SNU-C1, DLD-1) and one normal cell line (CCD-18Co) by
RT-qPCR analysis. The results revealed that the expression of
miR-145 was significantly downregulated in all colon adenocarcinoma
cell lines (Hs255.T, SW480, Hs257.T, HT29, LS174T, SNU-C1, and
DLD-1). The expression of miR-145 was up to 10-fold lower in the
Hs255.T, SW480, Hs257.T, HT29, LS174T, SNU-C1 and DLD-1
adenocarcinoma cell lines, in comparison to the normal CCD-18Co
cell line. The most marked downregulation was found in the Hs257.T
cell line, whereas the least downregulated expression of miR-145
was found in the SNU-C1 cell line (Fig. 1A).
Ectopic expression of miR-145 inhibits
the proliferation of SW480 cells
Due to the low expression of miR-145 in all the
colon adenocarcinoma cell lines, the SW480 cell line was selected
for the overexpression experiments to examine the role of miR-145
on cell proliferation. The overexpression of miR-145 was induced by
transfection of the SW480 cells with miR-145 mimics, which resulted
in a 5.6-fold increase in the expression of miR-145 as revealed by
the RT-qPCR analysis (Fig. 1B).
Furthermore, it was found that the overexpression of miR145 caused
significant (P<0.05) inhibition of the proliferation of the
SW480 cells (Fig. 1C). These
results were validated by the colony formation assay, which showed
that the overexpression of miR-145 suppressed the colony formation
potential of the SW840 cells (Fig.
1D). To investigate the underlying mechanism, the cells were
subjected to DAPI and AO/EB staining. It was found that the ectopic
expression of miR-145 in cells triggered apoptotic death of the
SW480 primary colon adenocarcinoma cells (Fig. 1E and F). The percentage of the
apoptotic cell populations was estimated by Annexin V/PI staining
and it was found that, compared with 2.61% apoptotic cells in the
miR-NC-transfected SW480 cells, the apoptotic cell percentage was
39.6% in the miR-145 mimics-transfected cells. Taken together these
results indicated that the ectopic expression of miR-145 inhibited
the proliferation of SW480 colon adenocarcinoma cells by triggering
apoptotic cell death.
Ectopic expression of miR-145 suppresses
the migration and invasion of SW480 cells
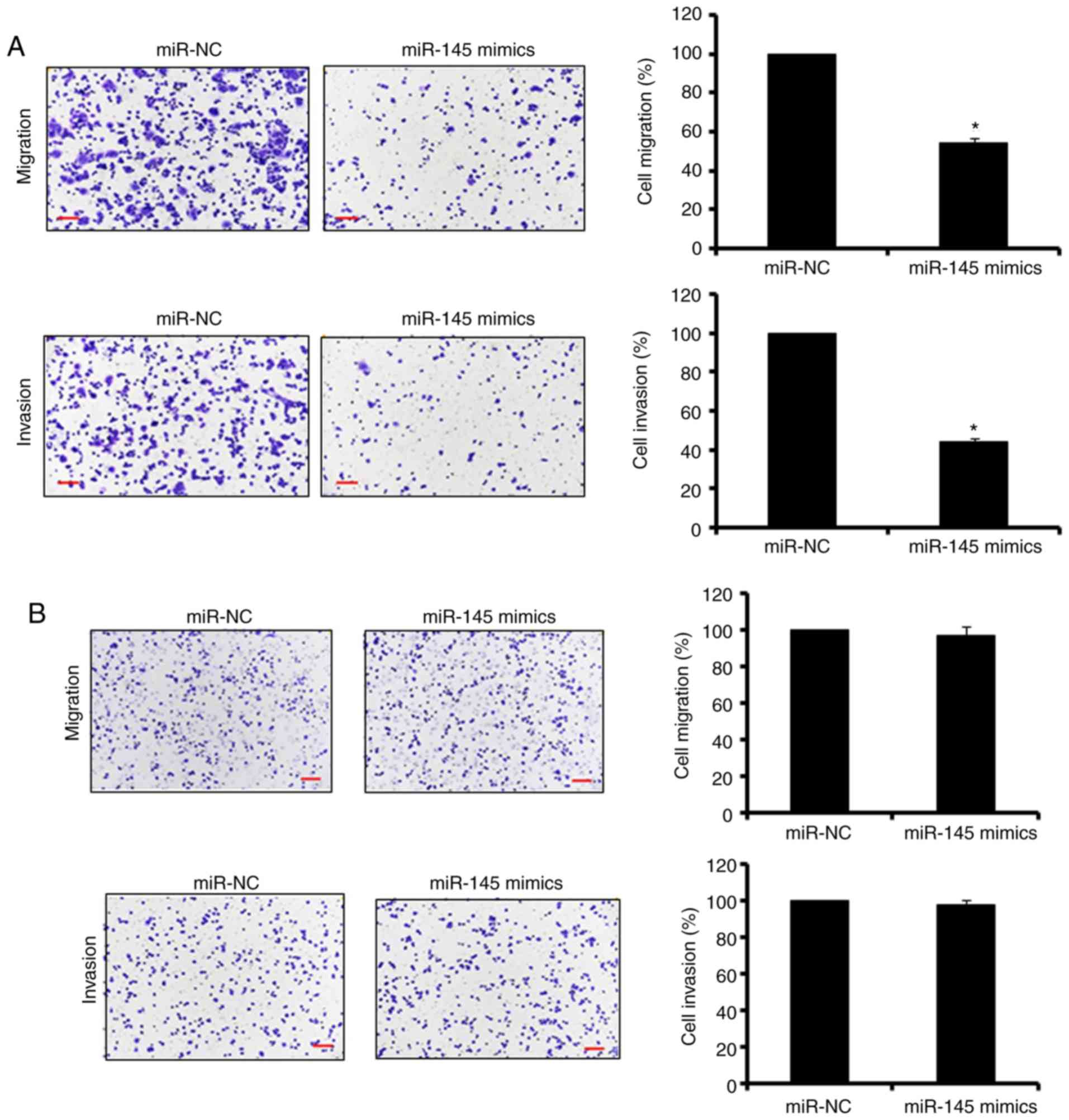
The Transwell cell assays were used to assess the
effect of the overexpression of miR-145 on the migration and
invasion of the SW480 colon adenocarcinoma cells. The results
revealed that the ectopic expression of miR-145 significantly
(P<0.05) suppressed the migration and invasion potential of the
SW480 colon adenocarcinoma cells (up to 60%; Fig. 2A). To determine whether the
effects of the overexpression of miR-145 on the migration and
invasion were specific to the cancer cells, miR-145 was also
overexpressed in the normal CCD-18Co cells. The results indicated
that the overexpression of miR-145 had no significant effect on the
migration and invasion of the normal CCD-18Co cells (Fig. 2B). Taken together, these results
suggested that miR-145 specifically inhibited the migration and
invasion of the colon adenocarcinoma cells.
miR-145 exerts its effects on SW480 cells
by targeting MAPK1
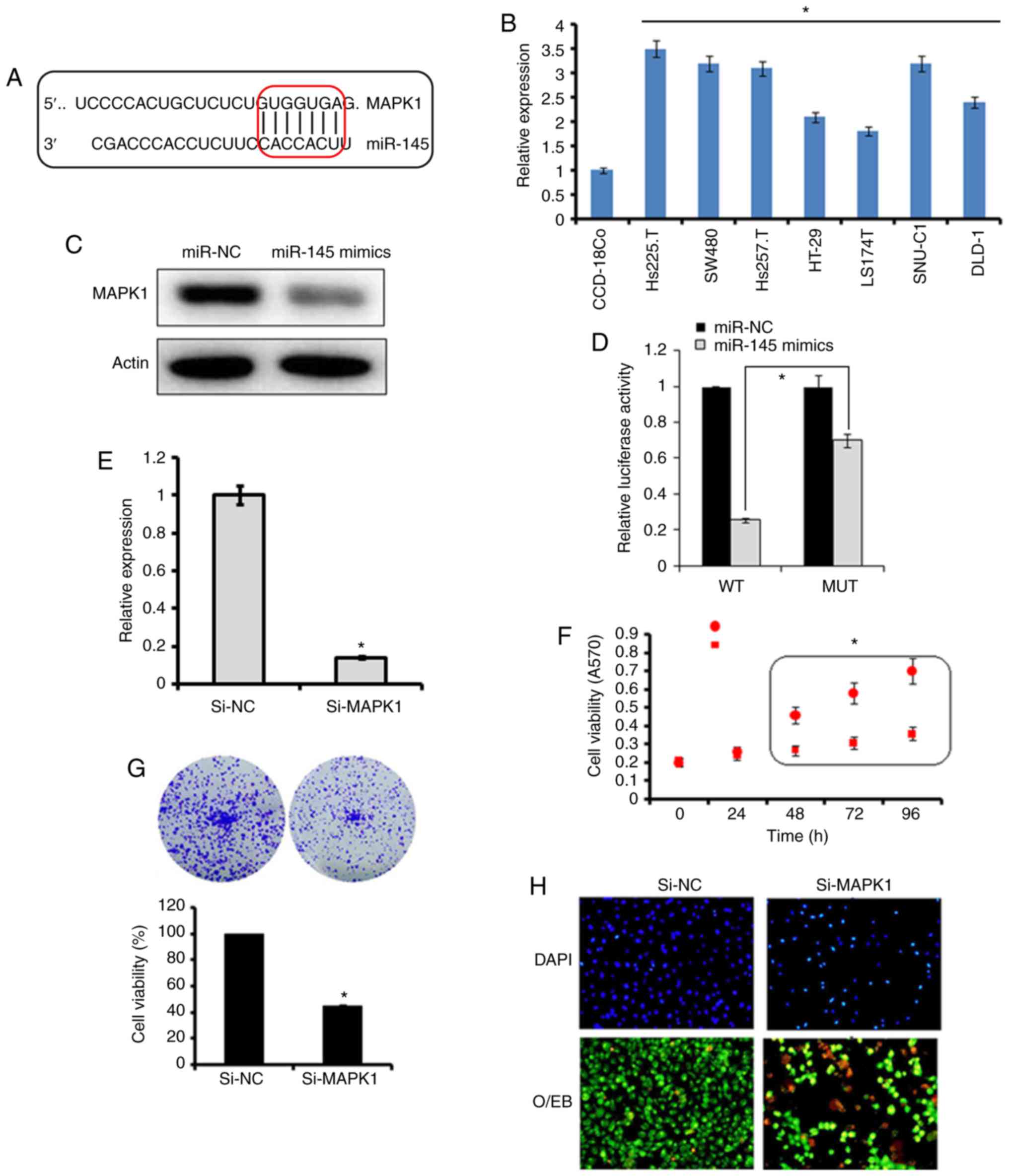
The bioinformatics analysis with TargetScan revealed
several targets for miR-145, which included MAPK1, signal
transducer and activator of transcription, mucin 1, mucin 13, and
insulin receptor substrate 1 (data not shown). However, no studies
have investigated MAPK1 as the target of miR-145, therefore, MAPK1
was selected for further investigation in the present study
(Fig. 3A). Firstly, the
expression of MAPK1 was investigated in the Hs255.T, SW480,
Hs257.T, HT29, LS174T, SNU-C1, and DLD-1colon adenocarcinoma cell
lines and the CCD-18Co normal cell line. It was found that the
expression of MAPK1 was significantly upregulated in the colon
adenocarcinoma cell lines (Fig.
3B). However, the ectopic expression of miR-145 in SW480 colon
adenocarcinoma cells, caused marked downregulation of the
expression of MAPK1 (Fig. 3C).
The dual luciferase reporter assay further confirmed MAPK1 to be
the target of miR-145 (Fig. 3D).
To further investigate the effect of MAPK1 on the proliferation,
migration and invasion of the SW480 cells, its expression was
silenced by transfection of the SW480 cells with si-MAPK1. The
reduction in the expression of the si-MAPK1-transfected SW480 cells
was confirmed by RT-qPCR analysis, which showed a 7.2-fold decrease
in the expression of MAPK1 in the SW480 cells (Fig. 3E). The effect of MAPK1 suppression
on the proliferation of SW480 cells was also investigated. It was
found that the silencing of MAPK1 also inhibited the proliferation
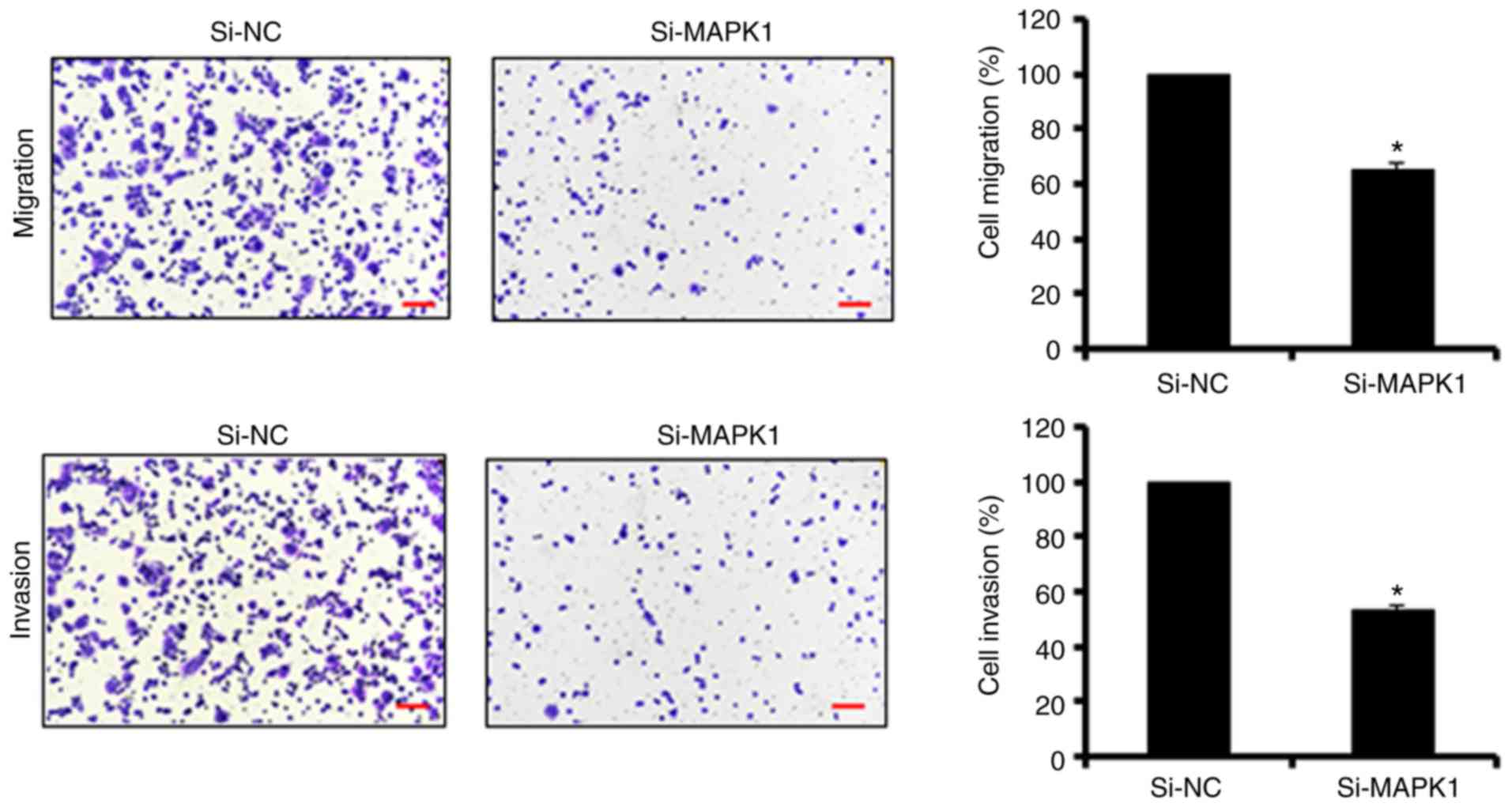
of SW480 cells by triggering apoptosis (Fig. 3F-H). Similar to the overexpression
of miR-145, MAPK1 silencing suppressed the migration and invasion
of the SW480 colon adenocarcinoma cells, further confirming MAPK1
as the target of miR-145 (Fig.
4). Subsequently, to overrule the possibility of the
involvement of any other target in the miR-145-induced effects, the
present study investigated whether co-transfection of the SW480
cells with si-MAPK1 and miR-145 inhibitor, was able to rescue the
effects of MAPK1 silencing on SW480 cell proliferation and
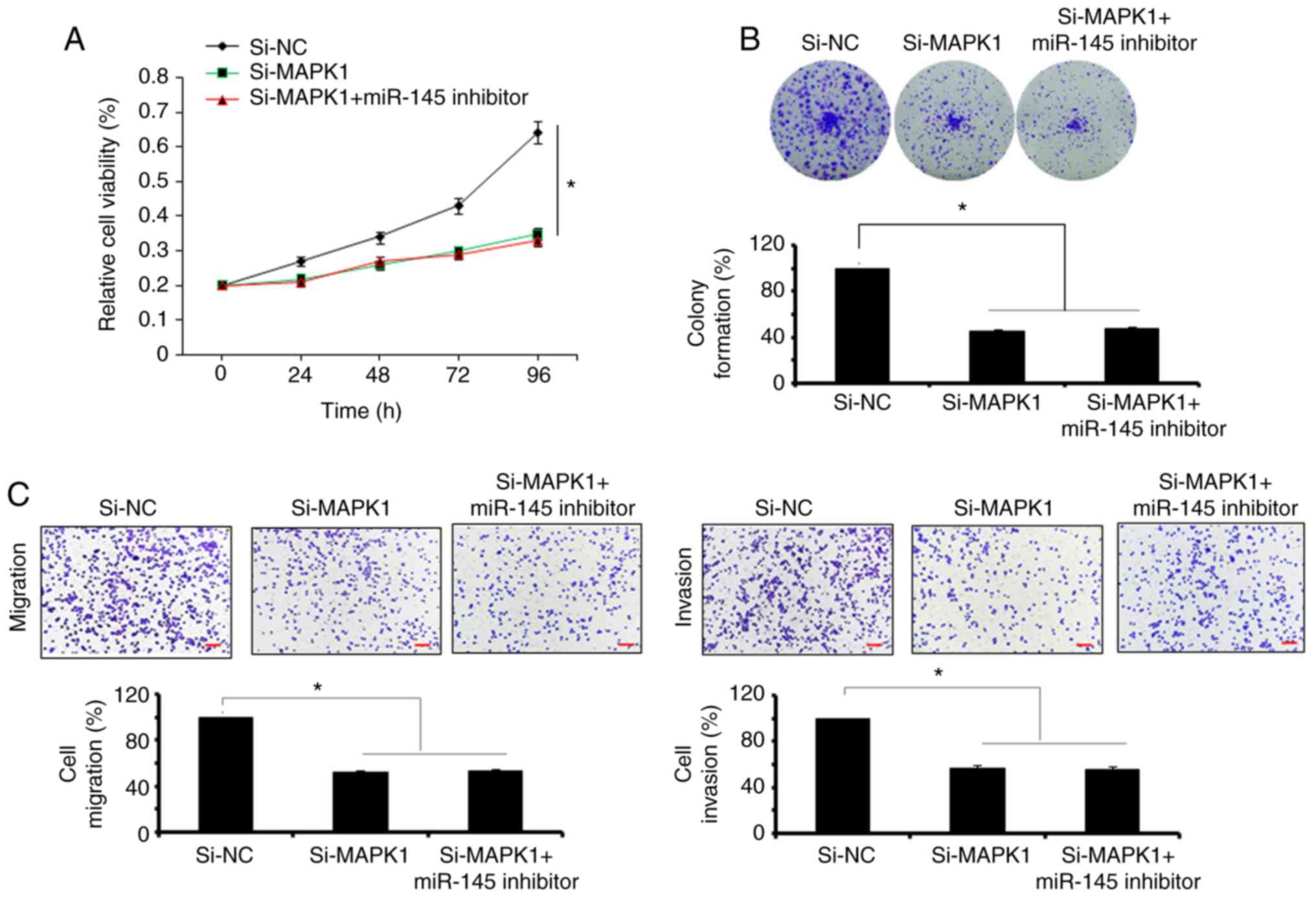
migration. It was observed that the inhibition of miR-145 did not
reverse the effects of MAPK1 silencing on the proliferation,
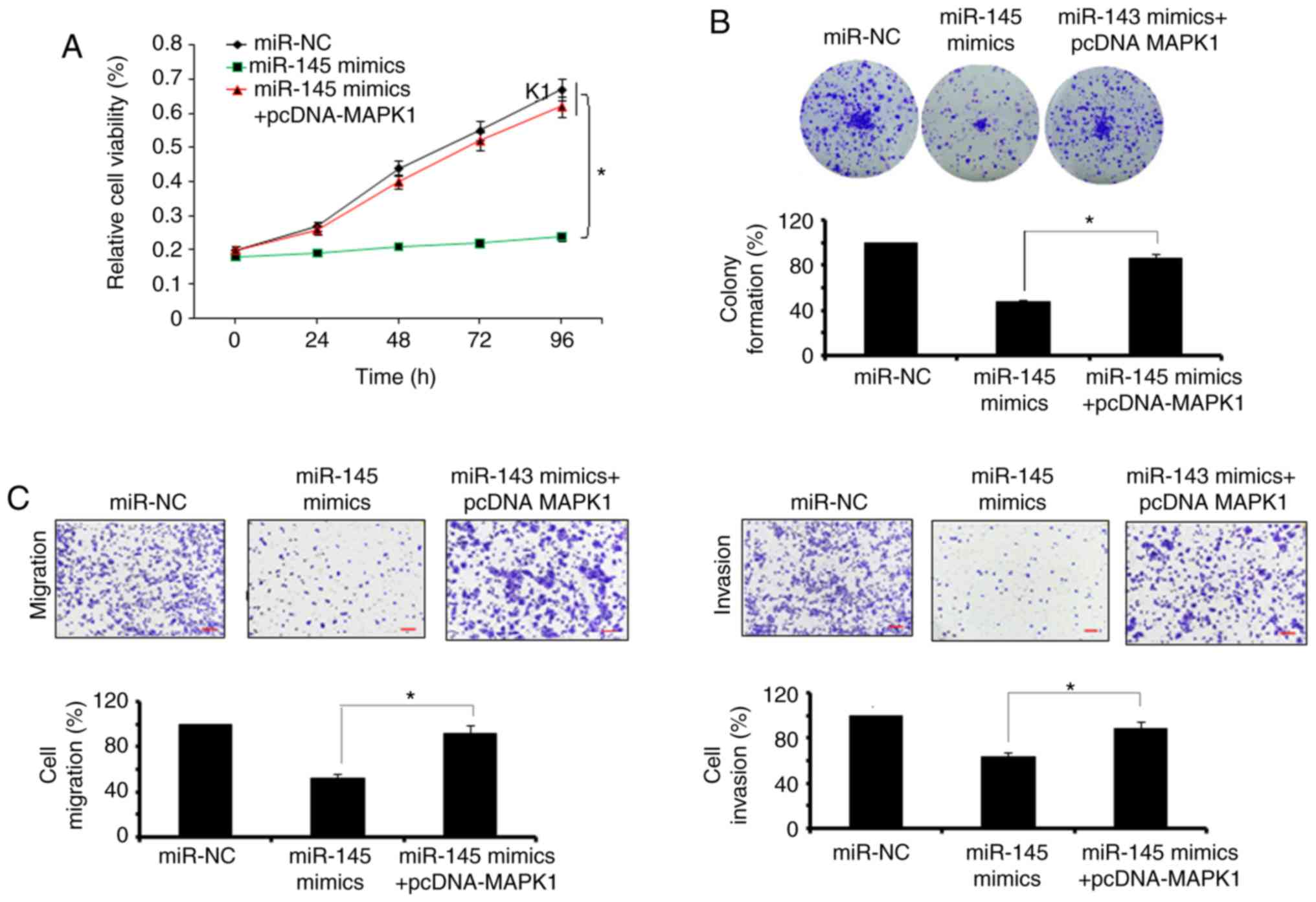
migration and invasion of SW480 cells (Fig. 5A-C). However, the overexpression
of MAPK1 reversed the effects of the miR-145 mimics on the
proliferation and migration and invasion of the SW480 cells
(Fig. 6A-C).
miR-145 inhibits tumor growth in
vivo
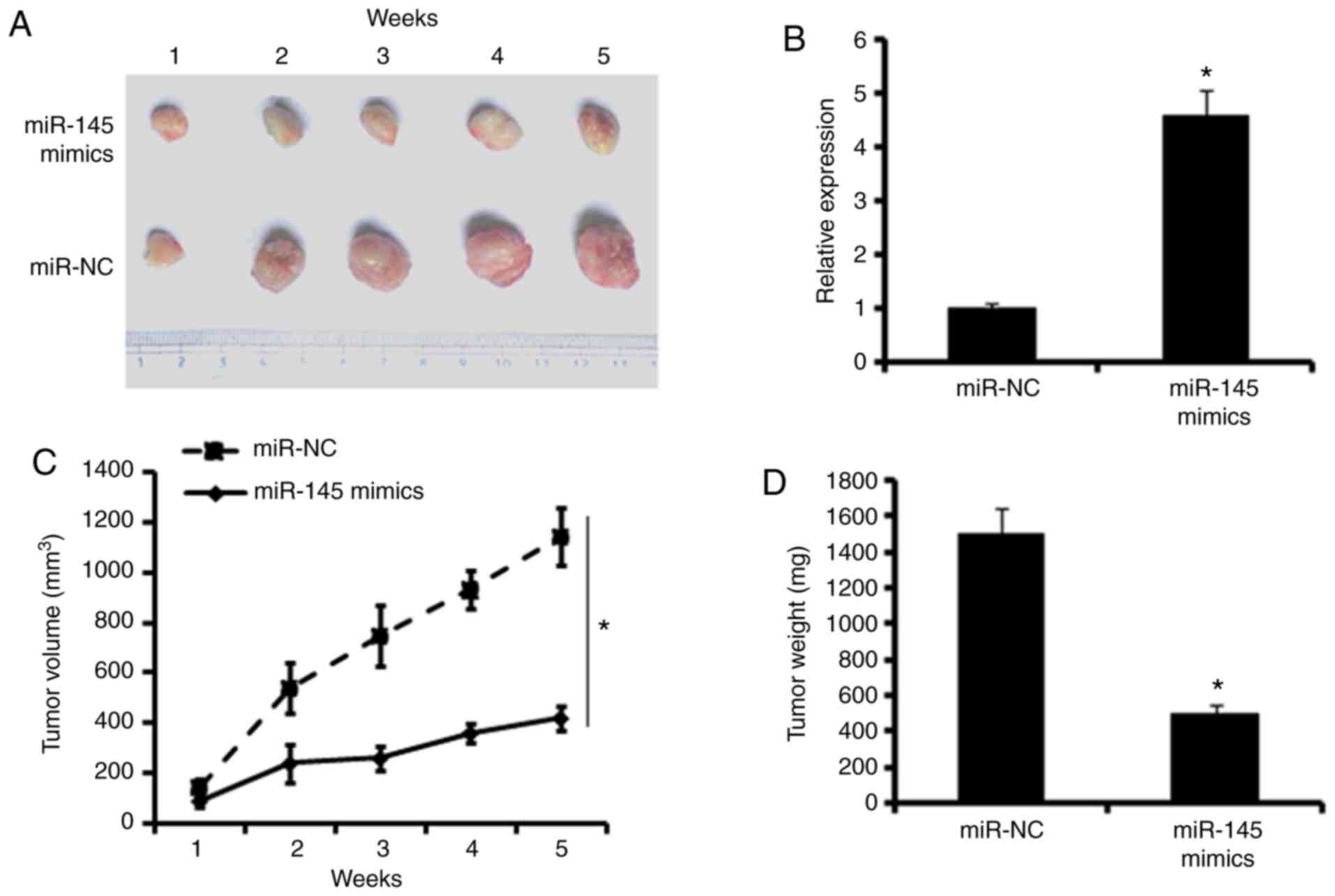
To examine the effects of miR-145 effects in
vivo, a colon adenocarcinoma xenograft model using BALB/c nude
mice was developed. The mice were sacrificed and the tumors were
harvested (Fig. 7A). The RT-PCR
assays showed that xenografts of the miR-145 mimic group exhibited
higher expression of miR-145 compared with the miR-NC group,
indicating the successful overexpression of miR-145 in the colon
adenocarcinoma xenograft (Fig.
7B). In addition, the monitoring of tumor volume and the
measurement of tumor weights revealed that the overexpression of
miR-145 significantly inhibited the growth of tumor xenografts,
leading to decreased tumor volumes and weights (Fig. 7C and D).
Discussion
Colon adenocarcinoma has a high rate of mortality
owing to it late diagnosis and lack of reliable biomarkers and
therapeutic targets (16). miRNAs
are ~20 nucleotide long non-coding RNA molecules, which have been
found to have vital functions in almost all biological pathways
(17). Therefore, miRNAs have
effects on a wide array of cancer-related processes, which include,
but are not limited to, proliferation, metastasis and cell cycle
(18). There is substantial
evidence relating to the implications of miRNAs in the development
of cancer (7). It has been
reported that miRNAs are often downregulated in tumors due to a
number of factors (8).
Consistently, in the present study, it was observed that the
expression of miR-145 was significantly downregulated in primary
colon adenocarcinoma cells. Furthermore, the ectopic expression of
miR-145 inhibited the proliferation of colon adenocarcinoma cells
by the induction of apoptosis. The overexpression of miR-145 also
inhibited the migration and invasion of SW480 colon adenocarcinoma
cells. These results are supported by the literature. For example,
miR-145 was reported to suppress the proliferation and metastasis
of primary colon adenocarcinoma (19). Similarly, Yin et al
reported that miR-145 inhibited the growth of lung adenocarcinoma
by targeting octamer-binding transcription factor 4 (20). Furthermore, miRNA expression
studies have revealed the role of miR-145 in the metastasis of
colorectal cancer (21). In the
present study, the overexpression of miR-145 had no apparent
effects on the migration and invasion of non-cancerous cells,
indicating that the effects of miR-145 are cancer-specific.
Investigation of the underlying mechanism revealed that miR-145
exerted its effects by targeting MAPK1, and silencing MAPK1 had
similar effects on the proliferation, migration and invasion of
primary colon adenocarcinoma cells as the overexpression of
miR-145. Although the inhibition of miR-145 did not rescue the
effects of MAPK1silencing on the proliferation, migration and
invasion of SW480 cells, the overexpression of MAPK1 reversed the
effects of miR-145 mimics on the proliferation, migration and
invasion of SW480 cells. MAPK1 has also been previously reported to
regulate the growth and apoptosis of cancer cells, confirming the
results of the present study (22). In addition, Li et al
reported that silencing MAPK1 inhibited the proliferation of HeLa
cells (23). Furthermore, MAPK1
has been reported to regulate the apoptotic cell death of cancer
cells (24), and MAPK1 has been
shown inhibit the expression of matrix metalloproteinase (MMP)-2
and MMP-9 in cervical cancer cells, ultimately inhibiting their
migration and invasion (25).
Finally, the in vivo evaluation in the present study
revealed the growth inhibitory effects of the overexpression of
miR-145 on the xenografted tumors. Taken together, it was concluded
that miR-145 inhibited the proliferation, migration and invasion of
primary colon adenocarcinoma cells by targeting MAPK1; therefore,
miR-145 may offer therapeutic potential for the management of colon
adenocarcinoma.
Funding
The study was supported by Shanghai Hospital of
Traditional Chinese Medicine (Shanghai, China) (grant no.
TCM-1156/2016).
Availability of data and materials
The datasets used and/or analyzed in the present
study are available from the corresponding author on reasonable
request.
Authors’ contributions
YY, XJL and PL performed all the experiments. YY and
XJL collected the materials and provided instrumental suggestion
for the present study. The present study was designed and
supervised by XTG. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved and supervised by the Ethics
Committee of Shanghai Hospital of Traditional Chinese Medicine
under approval number ECSHT-AS345/2017.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Schetter AJ, Leung SY, Sohn JJ, Zanetti
KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, et
al: MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. Jama. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hanif R, Qiao L, Shiff SJ and Rigas B:
Curcumin, a natural plant phenolic food additive, inhibits cell
proliferation and induces cell cycle changes in colon
adenocarcinoma cell lines by a prostaglandin-independent pathway. J
Lab Clin Med. 130:576–584. 1997. View Article : Google Scholar
|
|
3
|
Hong J, Lu H, Meng X, Ryu JH, Hara Y and
Yang CS: Stability, cellular uptake, biotransformation, and efflux
of tea polyphenol (-)-epigallocatechin-3-gallate in HT-29 human
colon adenocarcinoma cells. Cancer Res. 62:7241–7246.
2002.PubMed/NCBI
|
|
4
|
Tsukuda K, Tanino M, Soga H, Shimizu N and
Shimizu K: A novel activating mutation of the K-ras gene in human
primary colon adenocarcinoma. Biochem Biophys Res Commun.
278:653–658. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji W, Sun B and Su C: Targeting microRNAs
in cancer gene therapy. Genes (Basel). 8. pii; pp. E212017,
View Article : Google Scholar
|
|
9
|
Sachdeva M and Mo YY: miR-145-mediated
suppression of cell growth, invasion and metastasis. Am J Transl
Res. 2:170–180. 2010.PubMed/NCBI
|
|
10
|
Fei B and Wu H: MiR-378 inhibits
progression of human gastric cancer MGC-803 cells by targeting MAPK
in vitro. Oncol Res. 20:557–564. 2013. View Article : Google Scholar
|
|
11
|
Steibel JP, Poletto R, Coussens PM and
Rosa GJ: A powerful and flexible linear mixed model framework for
the analysis of relative quantification RT-PCR data. Genomics.
94:146–152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vidya Priyadarsini R, Senthil Murugan R,
Maitreyi S, Ramalingam K, Karunagaran D and Nagini S: The flavonoid
quercetin induces cell cycle arrest and mitochondria-mediated
apoptosis in human cervical cancer (HeLa) cells through p53
induction and NF-κB inhibition. Eur J Pharmacol. 649:84–91. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kasibhatla S, Amarante-Mendes GP, Finucane
D, Brunner T, Bossy-Wetzel E and Green DR: Acridine orange/ethidium
bromide (AO/EB) staining to detect apoptosis. CSH Protoc.
2006:pii:pdb-rot44932006.
|
|
14
|
Wang Y, Zhao W and Fu Q: miR-335
suppresses migration and invasion by targeting ROCK1 in
osteosarcoma cells. Mol Cell Biochem. 384:105–111. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gotham SM, Fryer PJ and Paterson WR: The
measurement of insoluble proteins using a modified Bradford assay.
Anal Biochem. 173:353–358. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fhaner CJ, Liu S, Ji H, Simpson RJ and
Reid GE: Comprehensive lipidome profiling of isogenic primary and
metastatic colon adenocarcinoma cell lines. Anal Chem.
84:8917–8926. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alvarez-Garcia I and Miska EA: MicroRNA
functions in animal development and human disease. Development.
132:4653–4662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao S, Wang P, Hua Y, Xi H, Meng Z, Liu T,
Chen Z and Liu L: ROR functions as a ceRNA to regulate Nanog
expression by sponging miR-145 and predicts poor prognosis in
pancreatic cancer. Oncotarget. 7:1608–1618. 2016.
|
|
20
|
Yin R, Zhang S, Wu Y, Fan X, Jiang F,
Zhang Z, Feng D, Guo X and Xu L: microRNA-145 suppresses lung
adenocarcinoma-initiating cell proliferation by targeting OCT4.
Oncol Rep. 25:1747–1754. 2011.PubMed/NCBI
|
|
21
|
Arndt GM, Dossey L, Cullen LM, Lai A,
Druker R, Eisbacher M, Zhang C, Tran N, Fan H, Retzlaff K, et al:
Characterization of global microRNA expression reveals oncogenic
potential of miR-145 in metastatic colorectal cancer. BMC Cancer.
9:3742009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang JY and Richardson BC: The MAPK
signalling pathways and colorectal cancer. Lancet Oncol. 6:322–327.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li XW, Tuergan M and Abulizi G: Expression
of MAPK1 in cervical cancer and effect of MAPK1 gene silencing on
epithelial-mesenchymal transition, invasion and metastasis. Asian
Pac J Trop Med. 8:937–943. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lwin WW, Park K, Wauson M, Gao Q, Finn PW,
Perkins D and Khanna A: Systems biology approach to transplant
tolerance: Proof of concept experiments using RNA interference
(RNAi) to knock down hub genes in Jurkat and HeLa cells in vitro. J
Surg Res. 176:e41–e462012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li XW, Tuergan M and Abulizi G: Expression
of MAPK1 in cervical cancer and effect of MAPK1 gene silencing on
epithelial mesenchymal transition, invasion and metastasis. Asian
Pac J Trop Med. 8:937–943. 2015. View Article : Google Scholar : PubMed/NCBI
|