Introduction
Pulmonary fibrosis is a chronic and progressive
disease characterized by alveolar epithelial injury and abnormal
collagen production (1,2). Patients with pulmonary fibrosis
often exhibit these effects, and pulmonary function is irreversibly
lost (3). Clinical research has
shown that patients with pulmonary fibrosis have a median survival
of only 2.5-3.5 years after diagnosis. Though the current treatment
options (pirfenidone and nintedanib) have improved, the prognosis
of pulmonary fibrosis has not improved (4,5).
Therefore, safer drugs with improved efficacy are needed to treat
pulmonary fibrosis.
Pulmonary fibrosis is caused by abnormal
proliferation of myofibroblasts and fibroblasts, which secrete
excessive extracellular matrix (ECM) proteins (4,5).
Epithelial-mesenchymal transition (EMT) is a process in which
polarized immotile epithelial cells are converted to motile
mesenchymal cells. This is characterized by the loss of proteins
associated with the polarized epithelial phenotype, such as
E-cadherin, and an increase in mesenchymal markers such as vimentin
and fibronectin (6,7). In addition, EMT has been reported to
be involved in myofibroblast formation (8). During pulmonary fibrosis, alveolar
epithelial cells (AECs) contribute to the formation of
myofibroblasts through EMT (9).
Thus, an approach targeting EMT might also be promising in the
treatment of pulmonary fibrosis (10).
The endoplasmic reticulum (ER), an important
organelle, serves as a key role in biological synthesis, including
correct protein folding, secretion and membrane protein
post-translational modifications (11). Endoplasmic reticulum stress (ERS)
triggers the activation of the unfolded protein response (UPR). The
UPR and the endoplasmic reticulum response have been recognized to
be activated in response to injury stress, oxidative stress,
inflammation, infection and gene mutations, and are involved in
conditions such as chronic liver fibrosis (12), pulmonary fibrosis (13-16), renal fibrosis (17) and myocardial fibrosis (18,19).
Isorhamnetin (Isor; molecular formula,
C16H12O7), a flavonol aglycone
isolated from the plant Hippophae rhamnoides L, is
frequently used in traditional Chinese medicine to prevent and
treat various diseases. Isor has been demonstrated to exert a
variety of effects, including antioxidant, anti-inflammatory,
antitumor, antiviral, anti-ERS and neurodegenerative injury
protection effects (20,21). Recent research has revealed that
Isor attenuates liver fibrosis by inhibiting transforming growth
factor (TGF)β/SMAD signaling and relieving oxidative stress
(22). Furthermore, Isor has been
reported to alleviate lipopolysaccharide-induced acute lung injury
in mice (23). Lung injury can
lead to pulmonary fibrosis (24).
Therefore, the present study speculated that Isor may have a key
role in pulmonary fibrosis. However, the function and mechanism
have not yet been clarified.
In the present study, the effect of Isor on
bleomycin (BLM)-induced pulmonary fibrosis was investigated. The
results demonstrated that Isor mitigated pulmonary fibrosis induced
by BLM. Mechanistically, the results revealed that Isor-mediated
ERS prevention was partially dependent on the regulation of EMT
progression. Based on the present findings, Isor might serve as a
potential therapeutic strategy for the treatment of pulmonary
fibrosis.
Materials and methods
Reagents and antibodies
Recombinant human TGFβ1 was purchased from
PeproTech, Inc. (Rocky Hill, NJ, USA). Isor was purchased from
Baomanbio (Shanghai, China). BLM was purchased from Hisun Company
(Zhejiang, China). Antibodies targeting collagen I (dilution,
1:6,000; cat. no. ab138492), α-smooth muscle actin (α-SMA;
dilution, 1:300; ab32575) and 78 kDa glucose-regulating protein
(GRP78)/binding immunoglobulin protein (BiP; dilution, 1:1,200;
cat. no. BM0134) were obtained from Abcam (Cambridge, UK).
Antibodies targeting TGFβ1 (dilution, 1:1,000; cat. no. AM4195),
protein kinase R-like endoplasmic reticulum kinase (PERK; dilution,
1:1,200; cat. no. BM0524) and E-cadherin (dilution, 1:1,200; cat
no. BM0537) were obtained from Abzoom Biolabs, Inc. (Dallas, TX,
USA). Antibodies targeting vimentin (dilution, 1:1,200; YT4880),
phosphorylated (p)-PERK (dilution, 1:2,000; cat. no. YP1055), DNA
damage-inducible transcript 3 (DDIT3; also known as CHOP; dilution,
1:1,200; cat. no. YT0911), eukaryotic translation initiation factor
2 subunit α (eIF2α; dilution, 1:1,200; cat. no. YT1507) and p-eIF2α
(dilution, 1:1,000; cat. no. YP0093) were obtained from ImmunoWay
Biotechnology, Plano, TX, USA. Horseradish peroxidase (HRP)-coupled
sheep anti-rat (dilution, 1:15,000; cat. no. SA001) or sheep
anti-rabbit (dilution, 1:15,000; cat. no. SA009) secondary
antibodies were obtained from Auragene Technology, Co., Inc.
(Changsha, China).
BLM-induced pulmonary fibrosis and
treatment
A total of 15 male 4-week old C57 mice (20-25 g in
weight; SLRC Laboratory Animal Company, Changsha, China) were
housed in rooms with a 12-h light/dark cycle at 25°C and 40-70%
humidity for 1 week prior to the experiment. Mice were fasted for
12 h and had ad libitum access to fresh tap water up until
the beginning of the experiment. During the experiment, the mice
had ad libitum access to food and water. The mice were then
randomly assigned to the Isor treatment group, the BLM group or the
control group. Mice in the Isor and BLM groups were
intraperitoneally injected with BLM (3.5 U/kg; Hisun Company,
Zhejiang, China), while mice in the control group were injected
with normal saline. The Isor treatment group was divided into two
subgroups: High dose (30 mg/kg) and low dose (10 mg/kg). Each
subgroup was treated with Isor by intragastric administration once
a day. Mice in the control group and BLM group were administered
the same volume of distilled water by gavage. After 28 days, the
mice were euthanized by pentobarbitone overdose. Lung tissues were
collected and used for hematoxylin and eosin (H&E) and Sirius
red staining and western blot analyses. All experiments involving
animals were approved by the Ethics Committee of Hunan Normal
University Medical College (Changsha, China).
Cell culture
Human A549 cells and human bronchial epithelial
cells (HBECs) were obtained from the Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China) and cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
fetal bovine serum (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at 37°C with 5% CO2. These cells were divided into
five groups: The control group, the TGFβ1 group, and three TGFβ1 +
Isor groups. Cells in the TGFβ1 group were stimulated with TGFβ1
alone (5 ng/ml) for 48 h. Cells in the TGFβ1 + Isor groups were
treated with 5 ng/ml TGFβ1 and 25, 50 or 100 µM Isor for 48
h.
Histological and immunohistochemical
analyses
The lung specimens were fixed in 4% paraformaldehyde
at room temperature for 48 h, dehydrated in a graded alcohol series
and embedded in paraffin blocks. Sections were blocked with 3%
hydrogen peroxide for 15 min at room temperature. Five-micron-thick
sections were then stained with H&E for 10 min for routine
examination, or with Sirius red for 8 min to visualize collagen
deposition at room temperature. For immunohistochemistry staining,
sections were stained with the aforementioned antibodies, developed
with 3,3′-diaminobenzidine, and counterstained with hematoxylin.
Samples were viewed using a light microscope (magnification for
H&E staining, ×100 and ×400; magnification for
immunohistochemistry staining, ×200 and ×400).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted with TRIzol reagent (Thermo
Fisher Scientific, Inc.). A total of 2 µg RNA was subjected
to reverse transcription using the PrimeScript Reverse
Transcription Reagent kit (Thermo Fisher Scientific, Inc.).
Quantitative analysis of the change in expression levels was
performed using SYBR Premix Ex Taq (Dongsheng Biotech, Guangzhou,
China) on an ABI 7300 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows: An
initial incubation at 95°C for 10 sec, followed by 40 cycles of
denaturation at 95°C for 10 sec and annealing at 60°C for 30 sec.
The sequences of the primers used were: GRP78, sense,
5′-GAAGGTTACCCATGCAGTT-3′ and antisense, 5′-AGCAATAGTTCCAGCGTCT-3′;
CHOP, sense, 5′-CCTGAAAGCAGCACCAAA-3′ and anti-sense,
5′-ACAAGCTCCATGTAGCAAAC-3′; and β-actin, sense,
5′-AGGGGCCGGACTCGTCATACT-3′ and antisense,
5′-GGCGGCACCACCATGTACCCT-3′. Data analysis was performed by the
comparative Cq method using ABI software (25). mRNA expression was normalized to
that of β-actin.
Western blot analysis
Protein was extracted from lung tissues and cells
with RIPA lysis buffer (Auragene Technology, Co., Inc., Shenzen,
China). Protein concentrations were determined with bicinchoninic
acid protein assay reagents (Auragene Technology, Co., Inc.)
according to the manufacturer’s instructions. Equal amounts of
protein (10 µg) from each sample were separated by 10%
SDS-PAGE and transferred to polyvinylidene fluoride membranes.
Membranes were blocked with 3% BSA-TBST at room temperature for 60
min. The membranes were probed with primary antibodies overnight at
4°C and then incubated with a HRP-coupled sheep anti-rat (dilution,
1:15,000; cat. no. SA001) or sheep anti-rabbit (dilution, 1:15,000;
cat. no. SA009; both from Auragene Technology, Co., Inc.) secondary
antibody at room temperature for 40 min. Signals were visualized
with an enhanced chemiluminescence detection kit (Auragene
Technology, Co., Inc.). Densitometric analysis was performed using
Image-Pro Plus v. 6.0 (Media Cybernetics, Silver Spring, MD,
USA).
Immunofluorescence cytochemistry
Cells were fixed in 4% paraformaldehyde for 30 min
at room temperature. After three 5-min washes with PBS, the cells
were blocked in PBS containing 3% goat serum (OriGene Technologies,
Inc., Beijing, China) and 0.5% Triton X-100 for 1 h at room
temperature. Afterwards, the cells were incubated with the
indicated primary antibodies at 4°C overnight and then incubated
with Cy3-conjugated goat anti-rabbit IgG(H+L) secondary antibody
(1:500) for 1 h at room temperature (catalog no. SA012; Auragene
Technology, Co., Inc.). The coverslips were counter-stained with
DAPI and imaged with a confocal laser scanning microscope (AE31;
Motic Group, Co., Ltd., Xiamen, China).
Statistical analysis
Data from three independent experiments were
expressed as the mean ± standard deviation and processed using SPSS
17.0 statistical software (SPSS, Inc., Chicago, IL, USA).
Differences between groups were evaluated by one-way analysis of
variance followed by Tukey’s test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Isor alleviates BLM-induced pulmonary
fibrosis in mice
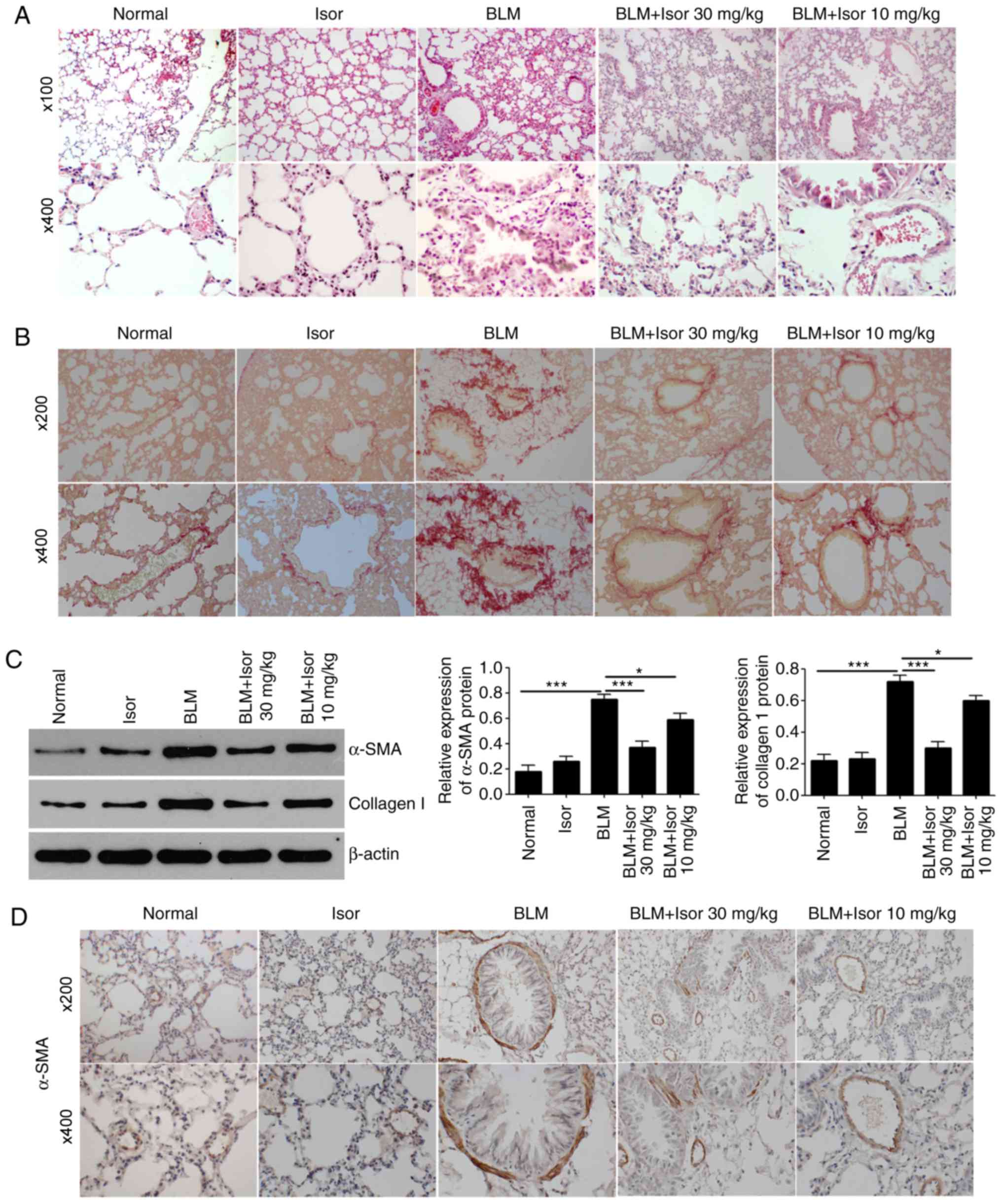
In accordance with previous studies (26,27), histological examination of lung
tissues demonstrated that four weeks of exposure to bleomycin in
mice led to significant pulmonary fibrotic changes, including
disturbed alveolar structure, marked thickening of the
interalveolar septa and dense interstitial infiltration by
inflammatory cells and fibroblasts (Fig. 1A). However, Isor treatment (10 and
30 mg/kg) significantly attenuated the fibrotic changes induced by
BLM (Fig. 1A), and the higher
concentration exhibited better inhibitory effect on fibrosis, with
the histological morphology being similar to the control mice. The
hallmark characteristic of BLM-induced pulmonary fibrosis is the
excessive deposition of extracellular matrix proteins, such as
collagen. Collagen deposition was clearly observed in the lungs of
BLM-treated mice by Sirius red staining (Fig. 1B). As expected, Isor treatment
significantly attenuated matrix protein deposition in the lungs
(Fig. 1B). In addition, α-SMA is
a hallmark of myofibroblasts and a marker of pulmonary fibrosis. As
illustrated in Fig. 1C, the
protein expression levels of α-SMA and collagen I were markedly
upregulated in the lungs of BLM-treated mice. Isor treatment
significantly reduced the production of collagen I and α-SMA in
BLM-treated lungs (Fig. 1C). The
expression of α-SMA was also monitored by immunohistochemical
analysis. BLM treatment resulted in markedly increased expression
of α-SMA in the area of pulmonary fibrosis, while this effect was
largely reversed in the Isor-treated groups (Fig. 1D).
Isor inhibits BLM-induced pulmonary
ERS
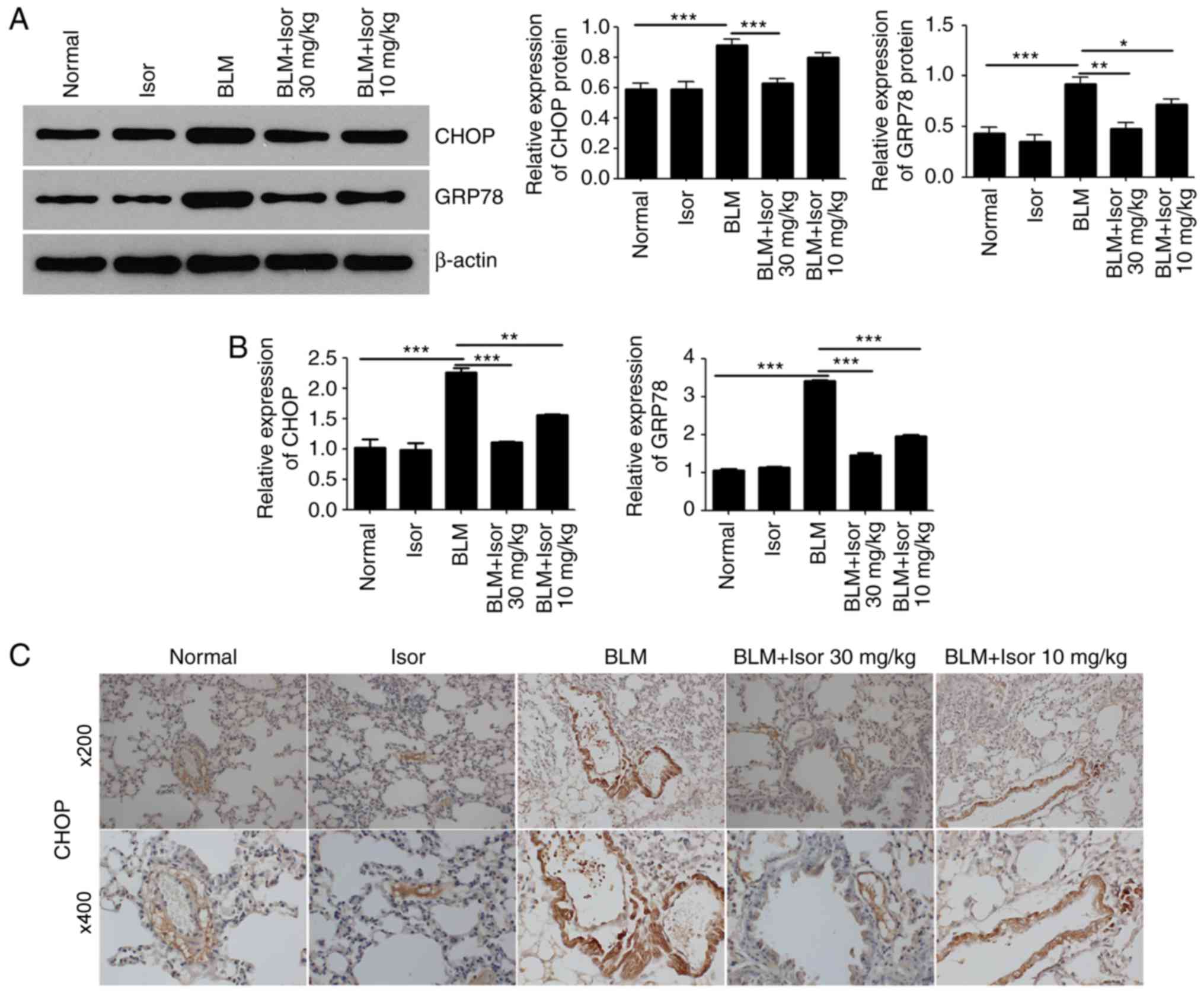
Recent studies have demonstrated that pulmonary ERS
is involved in the pathogenesis of BLM-induced pulmonary fibrosis
(14-16). Therefore, the effects of Isor on
BLM-induced pulmonary stress were analyzed. The protein expression
levels of GRP78 and CHOP were examined, as both proteins are widely
used as markers for ERS. As illustrated in Fig. 2A and B, the expression of GRP78
and CHOP was upregulated in the BLM-treated mice compared with the
control mice, at both the protein and mRNA level. Isor treatment
significantly attenuated the BLM-induced increase in GRP78 and CHOP
expression, at both the transcriptional and translational level
(Fig. 2A and B). These findings
suggested that Isor treatment inhibited ERS, which was in
accordance with a previous study (28). Immunohistochemistry staining for
CHOP was performed in lung tissues for further confirmation. CHOP
expression was decreased in Isor-treated BLM-induced lung tissues
(10 and 30 mg/kg) compared with BLM-induced lung tissues (Fig. 2C).
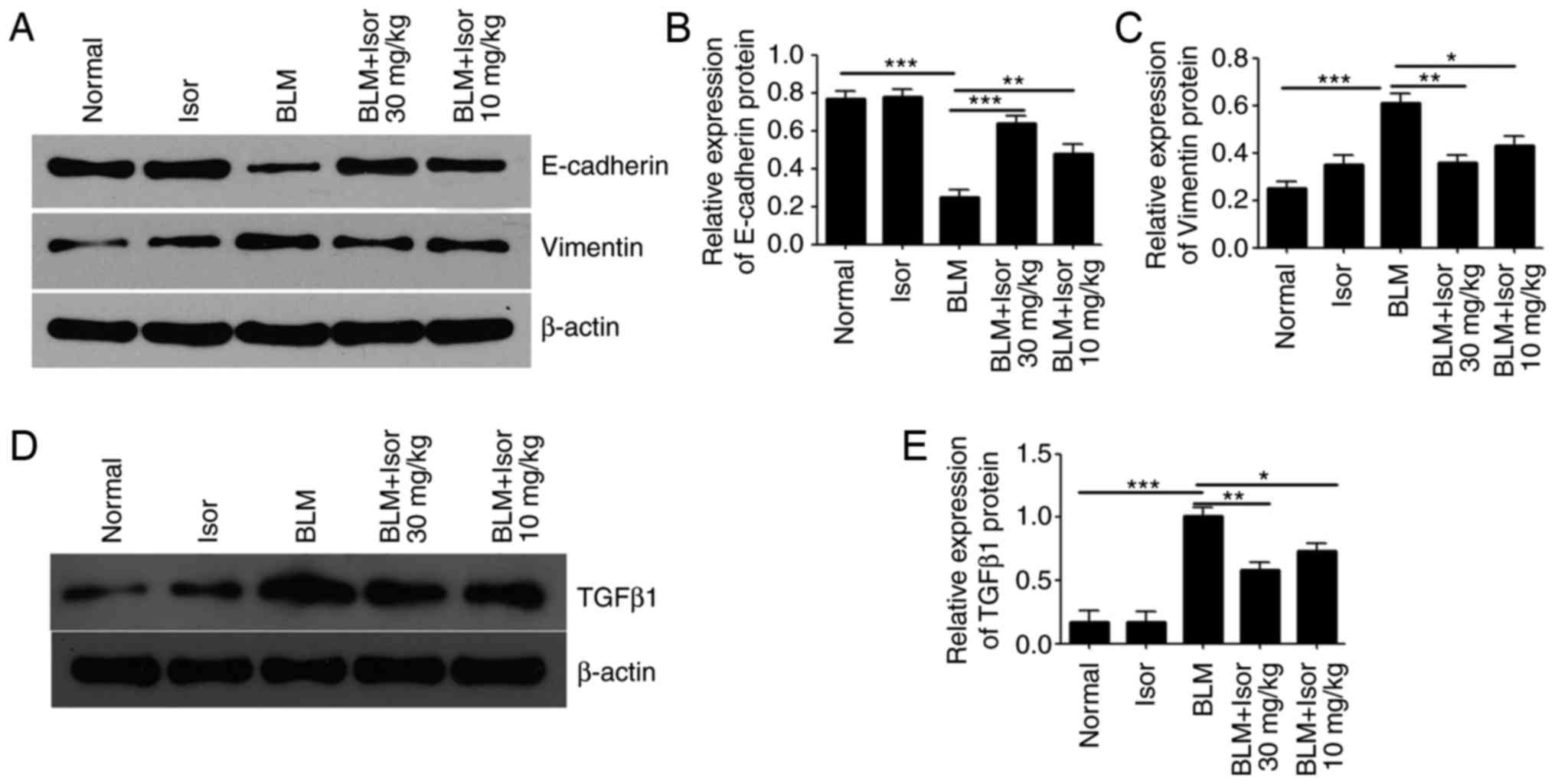
Isor alleviates ERS-mediated EMT
ERS-induced EMT contributes to fibrotic remodeling
in the lungs (15,29). Isor obviously inhibited
ERS-induced EMT of myofibroblasts in the lungs, as evidenced by
repression of pulmonary α-SMA expression described above. For
further confirmation, expression of the EMT-related proteins
E-cadherin, Vimentin and TGFβ1 was evaluated by western blotting.
The results demonstrated a significant upregulation of Vimentin and
TGFβ1 expression and downregulation of E-cadherin expression in the
BLM-treated group compared with the control group (Fig. 3A-E). However, Isor treatment
alleviated the increase in Vimentin and TGFβ1 expression and the
decrease in E-cadherin expression, compared with the BLM-treated
group (Fig. 3A-E). These findings
indicated that Isor treatment alleviated ERS-mediated EMT in
BLM-induced pulmonary fibrosis in vivo.
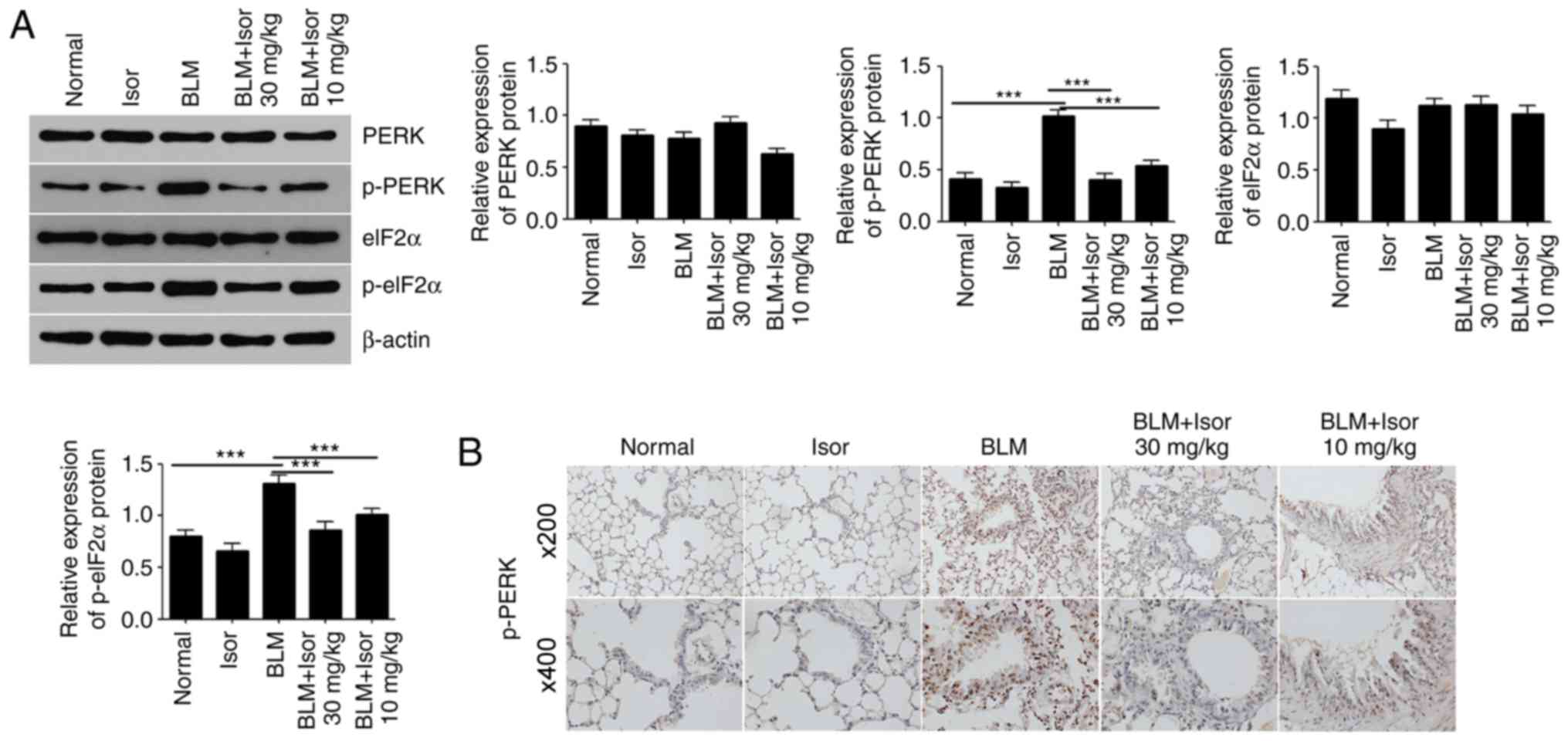
Isor inhibits ERS by downregulating
p-PERK
To investigate the possible pathways by which Isor
inhibits ERS, the expression levels of total and phosphorylated
PERK were determined by western blotting. PERK phosphorylation
levels significantly increased in the BLM-induced group compared
with the control group, while a dramatic decrease in p-PERK was
observed in the Isor-treated BLM groups (Fig. 4A). Immunohistochemistry staining
results for p-PERK in lung tissue sections supported the western
blot analysis results (Fig. 4B).
eIF2α is a downstream target of the PERK pathway. As illustrated in
Fig. 4A, consistent with PERK
phosphorylation, the levels of phosphorylated eIF2α in the lungs of
BLM-treated mice were elevated dramatically. Isor treatment
significantly attenuated BLM-induced eIF2α phosphorylation in the
lungs (Fig. 4A). Thus, Isor
exhibited an antifibrotic effect on BLM-induced pulmonary fibrosis
in vivo.
Effects of Isor on the expression of
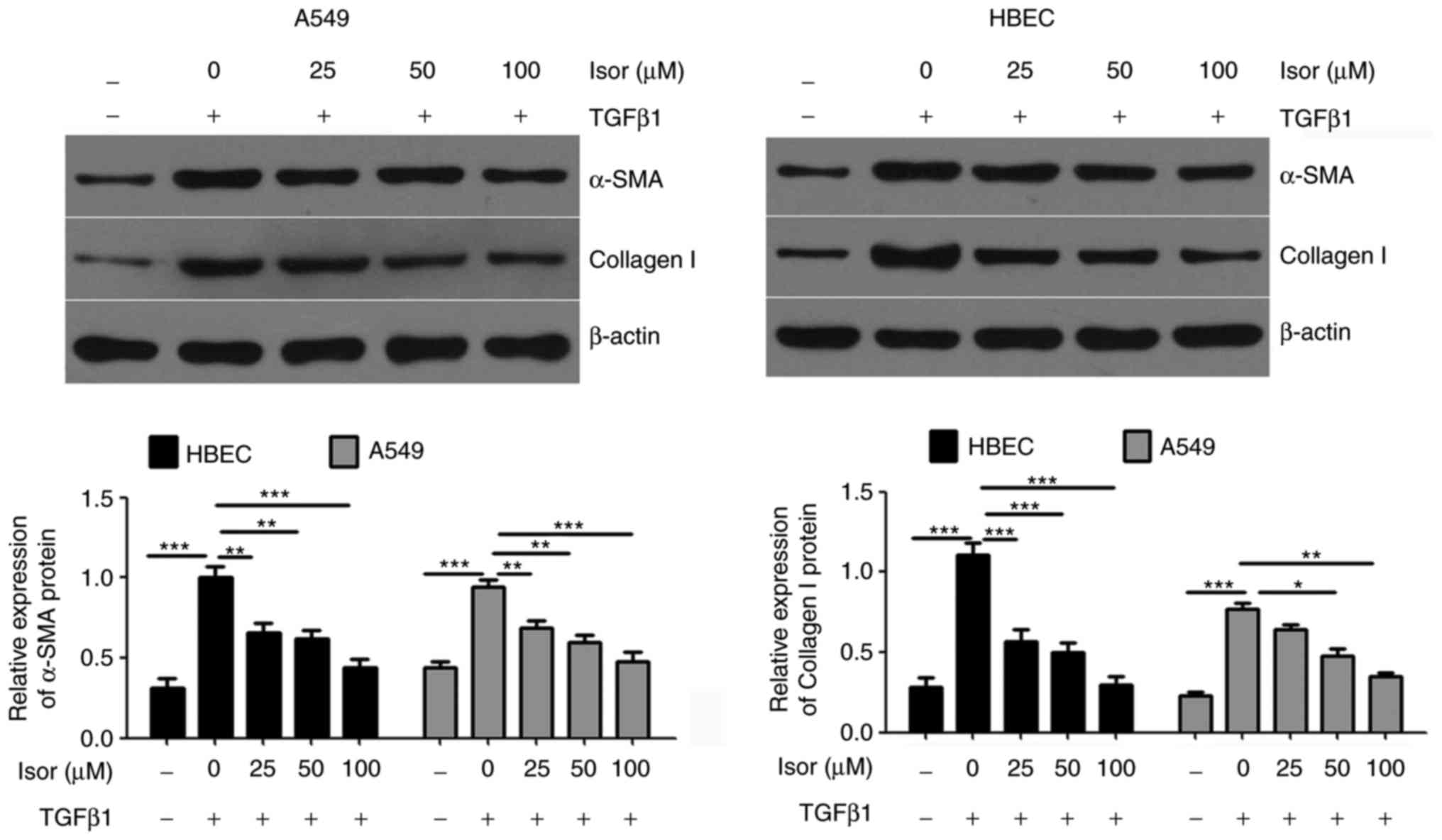
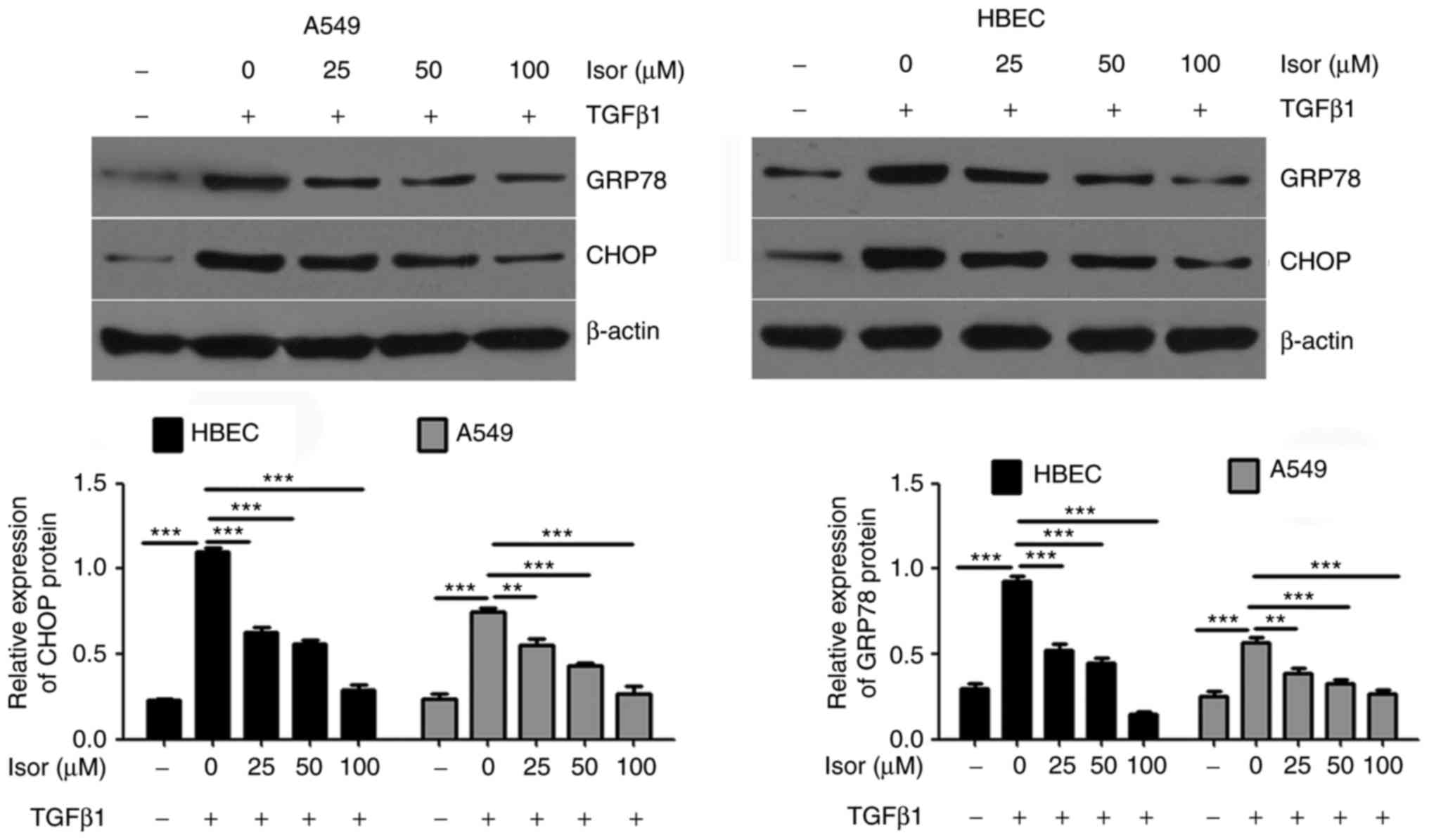
fibrosis-associated factors and ERS markers in A549 cells and HBECs
induced by TGFβ1
To further validate the antifibrotic effect of Isor,
in vitro studies were conducted in A549 cells and HBECs
simulated with TGFβ1. Results obtained in vitro were similar
to those observed in vivo. Treatment of cells with TGFβ1
increased the expression of fibrosis-related factors (collagen I
and α-SMA; Fig. 5) and ER markers
(GRP78 and CHOP; Fig. 6).
However, Isor suppressed the upregulated expression of collagen I,
α-SMA, GRP78 and CHOP in the presence of TGFβ1 in a dose-dependent
manner (Figs. 5 and 6).
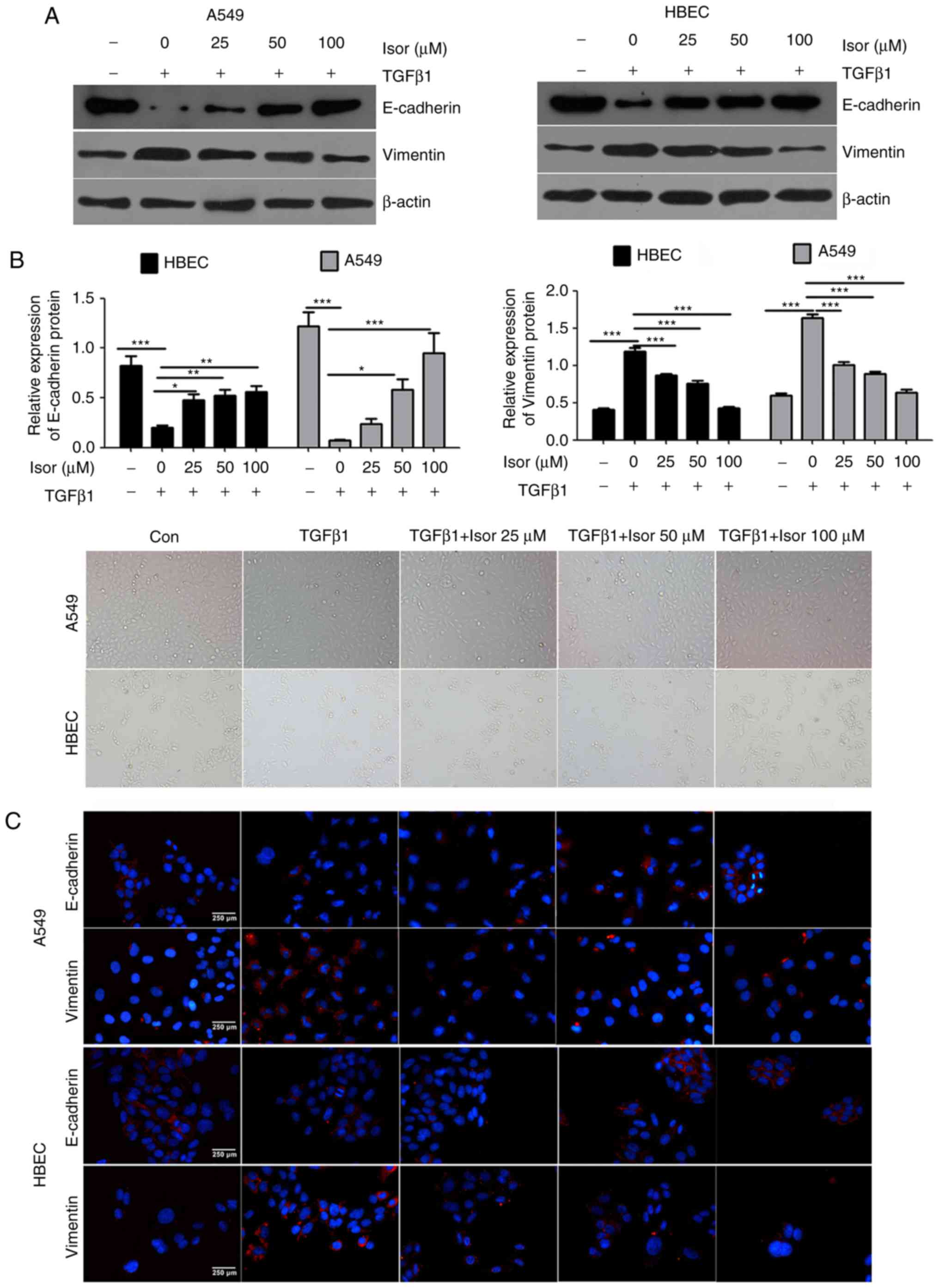
Effects of Isor on TGFβ1-induced EMT in
A549 cells and HBECs
Exposure to TGFβ1 caused A549 cells and HBECs to
undergo EMT, during which the morphology of A549 cells changed from
polygonal or cobblestone-like to a spindle-like shape, and that of
HBECs changed from a short fusiform shape to a long fusiform shape
(Fig. 7A). Aside from the
morphological changes, the expression levels of E-cadherin were
decreased, while the expression levels of Vimentin were increased
(Fig. 7B); these results were
also confirmed by immunostaining analysis (Fig. 7C). By contrast, treating cells
with Isor reversed the TGFβ1-induced EMT in a dose-dependent
manner, as illustrated by phenotypic cellular alterations (Fig. 7C) and the expression profiles of
the EMT markers E-cadherin and Vimentin (Fig. 7B). Taken together, the in
vivo and in vitro data suggest that Isor protects
against BLM-induced pulmonary fibrosis by inhibiting ERS-mediated
EMT in the lungs.
Discussion
The present study demonstrated for the first time
that Isor could inhibit pulmonary fibrosis in vivo and in
vitro. The results also demonstrated that BLM and TGFβ1
upregulated EMT and ERS in mice with pulmonary fibrosis and in
cells, respectively. Isor, a natural antioxidant polyphenol, was
capable of attenuating the effect of BLM-induced EMT and ERS in
vivo and in vitro. In addition, it suppressed
BLM-induced ERS by suppressing the PERK signaling pathway.
Increasing evidence has shown that EMT is involved
in the formation of myofibroblasts and serves an important role in
the pathogenesis of pulmonary fibrosis. In addition, inhibition of
EMT could prevent epithelial cells from transforming into
myofibroblasts, alleviating pulmonary fibrosis. For example, NLR
family pyrin domain containing 3 (NLRP3) modulates EMT through the
TGFβ1 signaling pathway, inhibiting pulmonary fibrosis (30). GHK inhibits BLM-induced fibrosis
progression, the inflammatory response and EMT via the
TGFβ1/Smad2/3 and insulin-like growth factor-1 (IGF-1) pathway
(31). In the present study, the
results demonstrated that Isor has the same function as NLRP3 and
GHK, as it inhibited BLM- and TGFβ1-induced upregulation of
N-cadherin and downregulation of E-cadherin in mice and cells.
Taken together, these results revealed that Isor inhibits
BLM-induced pulmonary fibrosis through its inhibition of EMT in the
lungs.
Multiple studies have confirmed that ERS and UPR
signaling activation are involved in the pathogenesis of idiopathic
pulmonary fibrosis, which includes elevated expression of GRP78,
activating transcription factor 6 (ATF6), PERK, and
inositol-requiring enzyme-1 (IRE-1) (15,32). Previous studies have demonstrated
that silencing of synoviolin and calcium/calmodulin-dependent
protein kinase I (CaMKI) suppress ERS and alveolar epithelial cell
apoptosis, protecting against pulmonary fibrosis (33), which indicates that ERS has an
important role in pulmonary fibrosis. Wang et al (34) also reported that chlorogenic acid
could inhibit pulmonary fibrosis through inhibition of ERS in
vivo and in vitro. These previous findings support the
current results. The present study investigated the effect of Isor
on ERS, which was promoted by BLM-induced pulmonary fibrosis. The
results demonstrated that the expression of ER chaperones (CHOP and
GRP78) and members of the ERS signaling pathway (p-PERK and
p-eIF2α) was markedly increased in mice with BLM-induced pulmonary
fibrosis, and this effect was reversed by Isor treatment.
Furthermore, it was demonstrated that Isor inhibited the ERS
induced by TGFβ1 in A549 cells and HBECs in vitro. In
addition, it is reported that Isor can inhibit ERS in obesity
(27), and Isor inhibited
ERS-induced injury through protein kinase C in N2a cells, which is
in accordance with the present results. Taken together, these
results suggest that Isor inhibited BLM-induced pulmonary fibrosis
through inhibition of ERS in the lung.
According to recent reports, ERS can induce EMT and
the progression of pulmonary fibrosis. Inhibition of ERS could
alleviate pulmonary fibrosis (7,8).
For example, melatonin inhibits ERS and EMT in BLM-induced
pulmonary fibrosis (35).
Therefore, Isor may inhibit EMT in pulmonary fibrosis via its
inhibition of ERS in the lung. The target proteins involved in this
process require further investigation.
In summary, the present study revealed that Isor
inhibited EMT and ERS in BLM-induced pulmonary fibrosis via the
PERK pathway. These findings may provide novel insights into the
pathogenesis of pulmonary fibrosis. Additional investigations are
necessary to elucidate the full antifibrotic potential of Isor as a
concomitant therapy for patients with lung fibrosis, including that
produced during BLM treatment, especially with regard to timing of
its administration.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Science and
Technology Innovation Program of Hunan Province (grant no.
2017SK50515).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors’ contributions
YY designed the study and obtained funding. QZ
performed the animal experiments and tests. QZ, MT, BO, CL and CH
performed the cell function assay. MT and YY provided major
contributions to the writing of the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All experiments involving animals were approved by
the Ethics Committee of Hunan Normal University Medical College
(Changsha, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
King TE Jr, Pardo A and Selman M:
Idiopathic pulmonary fibrosis. Lancet. 378:1949–1961. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wolters PJ, Collard HR and Jones KD:
Pathogenesis of idiopathic pulmonary fibrosis. Annu Rev Pathol.
9:157–179. 2014. View Article : Google Scholar :
|
|
3
|
Raghu G, Collard HR, Egan JJ, Martinez FJ,
Behr J, Brown KK, Colby TV, Cordier JF, Flaherty KR, Lasky JA, et
al: An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary
fibrosis: Evidence-based guidelines for diagnosis and management.
Am J Respir Crit Care Med. 183:788–824. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raghu G, Rochwerg B, Zhang Y, Garcia CA,
Azuma A, Behr J, Brozek JL, Collard HR, Cunningham W, Homma S, et
al: An official ATS/ERS/JRS/ALAT clinical practice guideline:
Treatment of idiopathic pulmonary fibrosis. An update of the 2011
clinical practice guideline. Am J Respir Crit Care Med.
192:e3-e192015. View Article : Google Scholar
|
|
5
|
Walter N, Collard HR and King TE Jr:
Current perspectives on the treatment of idiopathic pulmonary
fibrosis. Proc Am Thorac Soc. 3:330–338. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Borensztajn K, Crestani B and Kolb M:
Idiopathic pulmonary fibrosis: From epithelial injury to
biomarkers-insights from the bench side. Respiration. 86:441–452.
2013. View Article : Google Scholar
|
|
7
|
Li T, Yang X, Xin S, Cao Y and Wang N:
Paraquat poisoning induced pulmonary epithelial mesenchymal
transition through Notch1 pathway. Sci Rep. 7:9242017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang YC, Liu JS, Tang HK, Nie J, Zhu JX,
Wen LL and Guo QL: miR-221 targets HMGA2 to inhibit
bleomycin-induced pulmonary fibrosis by regulating
TGF-β1/Smad3-induced EMT. Int J Mol Med. 38:1208–1216. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang M, Wang YP, Zhu LQ, Cai Q, Li H and
Yang HF: MAPK pathway mediates epithelial-mesenchymal transition
induced by paraquat in alveolar epithelial cells. Environ Toxicol.
31:1407–1414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takemasa A, Ishii Y and Fukuda T: A
neutrophil elastase inhibitor prevents bleomycin-induced pulmonary
fibrosis in mice. Eur Respir J. 40:1475–1482. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Back SH, Lee K, Vink E and Kaufman RJ:
Cytoplasmic IRE1alpha-mediated XBP1 mRNA splicing in the absence of
nuclear processing and endoplasmic reticulum stress. J Biol Chem.
281:18691–18706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Galligan JJ, Smathers RL, Shearn CT, Fritz
KS, Backos DS, Jiang H, Franklin CC, Orlicky DJ, Maclean KN and
Petersen DR: Oxidative stress and the ER stress response in a
murine model for early-stage alcoholic liver disease. J Toxicol.
2012.207594:2012.
|
|
13
|
Baek HA, Kim DS, Park HS, Jang KY, Kang
MJ, Lee DG, Moon WS, Chae HJ and Chung MJ: Involvement of
endoplasmic reticulum stress in myofibroblastic differentiation of
lung fibroblasts. Am J Respir Cell Mol Biol. 46:731–739. 2012.
View Article : Google Scholar
|
|
14
|
Tanjore H, Lawson WE and Blackwell TS:
Endoplasmic reticulum stress as a pro-fibrotic stimulus. Biochim
Biophys Acta. 1832.940–947. 2013.
|
|
15
|
Lawson WE, Cheng DS, Degryse AL, Tanjore
H, Polosukhin VV, Xu XC, Newcomb DC, Jones BR, Roldan J, Lane KB,
et al: Endoplasmic reticulum stress enhances fibrotic remodeling in
the lungs. Proc Natl Acad Sci USA. 108:10562–10567. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lawson WE, Crossno PF, Polosukhin VV,
Roldan J, Cheng DS, Lane KB, Blackwell TR, Xu C, Markin C, Ware LB,
et al: Endoplasmic reticulum stress in alveolar epithelial cells is
prominent in IPF: Association with altered surfactant protein
processing and herpesvirus infection. Am J Physiol Lung Cell Mol
Physiol. 294:L1119–L1126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chiang CK, Hsu SP, Wu CT, Huang JW, Cheng
HT, Chang YW, Hung KY, Wu KD and Liu SH: Endoplasmic reticulum
stress implicated in the development of renal fibrosis. Mol Med.
17:1295–1305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ayala P, Montenegro J, Vivar R, Letelier
A, Urroz PA, Copaja M, Pivet D, Humeres C, Troncoso R, Vicencio JM,
et al: Attenuation of endoplasmic reticulum stress using the
chemical chaperone 4-phenylbutyric acid prevents cardiac fibrosis
induced by isoproterenol. Exp Mol Pathol. 92:97–104. 2012.
View Article : Google Scholar
|
|
19
|
Spitler KM and Webb RC: Endoplasmic
reticulum stress contributes to aortic stiffening via proapoptotic
and fibrotic signaling mechanisms. Hypertension. 63:e40-e452014.
View Article : Google Scholar
|
|
20
|
Ahn H and Lee GS: Isorhamnetin and
hyperoside derived from water dropwort inhibits inflammasome
activation. Phytomedicine. 24:77–86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao L, Yao R, Liu Y, Wang Z, Huang Z, Du
B, Zhang D, Wu L, Xiao L and Zhang Y: Isorhamnetin protects against
cardiac hypertrophy through blocking PI3K-AKT pathway. Mol Cell
Biochem. 429:167–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang JH, Kim SC, Kim KM, Jang CH, Cho SS,
Kim SJ, Ku SK, Cho IJ and Ki SH: Isorhamnetin attenuates liver
fibrosis by inhibiting TGF-β/Smad signaling and relieving oxidative
stress. Eur J Pharmacol. 783:92–102. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang B, Li XP, Ni YF, Du HY, Wang R, Li
MJ, Wang WC, Li MM, Wang XH, Li L, et al: Protective effect of
isorhamnetin on lipopolysaccharide-induced acute lung injury in
mice. Inflammation. 39:129–137. 2016. View Article : Google Scholar
|
|
24
|
Knudsen L, Ruppert C and Ochs M: Tissue
remodelling in pulmonary fibrosis. Cell Tissue Res. 367:607–626.
2017. View Article : Google Scholar
|
|
25
|
Higashiyama H, Yoshimoto D, Okamoto Y,
Kikkawa H, Asano S and Kinoshita M: Receptor-activated Smad
localisation in bleomycin-induced pulmonary fibrosis. J Clin
Pathol. 60:283–289. 2007. View Article : Google Scholar
|
|
26
|
Zhuang Y, Dai J, Wang Y, Zhang H, Li X,
Wang C, Cao M, Liu Y, Ding J, Cai H, et al: MiR-338* targeting
smoothened to inhibit pulmonary fibrosis by epithelial-mesenchymal
transition. Am J Transl Res. 8:3206–3213. 2016.
|
|
27
|
Rodríguez-Rodríguez C, Torres N,
Gutiérrez-Uribe JA, Noriega LG, Torre-Villalvazo I, Leal-Díaz AM,
Antunes-Ricardo M, Márquez-Mota C, Ordaz G, Chavez-Santoscoy RA, et
al: The effect of isorhamnetin glycosides extracted from Opuntia
ficus-indica in a mouse model of diet induced obesity. Food Funct.
6:805–815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao H, Qin HY, Cao LF, Chen YH, Tan ZX,
Zhang C and Xu DX: Phenylbutyric acid inhibits
epithelial-mesenchymal transition during bleomycin-induced lung
fibrosis. Toxicol Lett. 232:213–220. 2015. View Article : Google Scholar
|
|
29
|
Tian R, Zhu Y, Yao J, Meng X, Wang J, Xie
H and Wang R: NLRP3 participates in the regulation of EMT in
bleomycin-induced pulmonary fibrosis. Exp Cell Res. 357:328–334.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou XM, Wang GL, Wang XB 2, Liu L, Zhang
Q, Yin Y, Wang QY, Kang J and Hou G: GHK peptide inhibits
bleo-mycin-induced pulmonary fibrosis in mice by suppressing
TGFbeta1/Smad-mediated epithelial-to-mesenchymal transition. Front
Pharmacol. 8:9042017. View Article : Google Scholar
|
|
31
|
Wynn TA and Ramalingam TR: Mechanisms of
fibrosis: Therapeutic translation for fibrotic disease. Nat Med.
18:1028–1040. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Uhal BD, Nguyen H, Dang M, Gopallawa I,
Jiang J, Dang V, Ono S and Morimoto K: Abrogation of ER
stress-induced apoptosis of alveolar epithelial cells by
angiotensin 1–7. Am J Physiol Lung Cell Mol Physiol. 305:L33–L41.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Winters CJ, Koval O, Murthy S, Allamargot
C, Sebag SC, Paschke JD, Jaffer OA, Carter AB and Grumbach IM:
CaMKII inhibition in type II pneumocytes protects from
bleomycin-induced pulmonary fibrosis by preventing
Ca2+-dependent apoptosis. Am J Physiol Lung Cell Mol
Physiol. 310:L86–L94. 2016. View Article : Google Scholar
|
|
34
|
Wang YC, Dong J, Nie J, Zhu JX, Wang H,
Chen Q, Chen JY, Xia JM and Shuai W: Amelioration of
bleomycin-induced pulmonary fibrosis by chlorogenic acid through
endoplasmic reticulum stress inhibition. Apoptosis. 22:1147–1156.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
PLOS ONE Staff: Correction: Melatonin
inhibits endoplasmic reticulum stress and epithelial-mesenchymal
transition during bleomycin-induced pulmonary fibrosis in mice.
PLoS One. 10:e1193812015.
|