Introduction
Post-stroke depression (PSD) and post-stroke anxiety
(PSA) are serious and often interrelated complications of stroke.
PSD is the most common psychiatric disorder following ischemic
stroke and occurs in ~1/3 of all stroke survivors (1); PSA prevalence is ~20-25% (2). Comorbidity of PSD and PSA was
reported to be 12.3% of stroke survivors at 3 months post-stroke
(3). PSD is positively correlated
with PSA (2), and comorbid
depression and anxiety worsen the prognosis and severity of
depressive symptoms (4). PSD and
PSA can compromise rehabilitation outcomes, lead to poorer
cognitive and physical outcomes and can negatively impact patient
quality of life following a stroke (2,5),
making this a serious and costly public health issue, warranting
investigations into antidepressant preventive and curative
therapies.
Pre-stroke social isolation gives rise to cognitive
impairment of functional recovery in humans and has been associated
with an increased risk of PSD and PSA (6-9).
An experimental study provided direct evidence that isolating mice
immediately following stroke leads to PSD and PSA, which may
contribute to decreased general locomotor activity and histological
outcomes in the stroke hemisphere (10). Several socially isolated patients
are not clinically identified until they seek medical attention
following a stroke, and stroke survivors experience and perceive
higher levels of social isolation compared with age-matched healthy
individuals (11). Therefore, it
is essential to investigate treatment strategies for post-stroke
isolation-mediated PSD and PSA, subsequent neurogenesis, and
neurological and cognitive dysfunction. Translational efforts
targeting socio-emotional factors in post-stroke functional
recovery may widen the intervention options for social support and
may be an interesting addition to current therapeutic strategies
for acute stroke. However, the role and underlying mechanisms of
post-stroke social isolation in hippocampal neurogenesis and
post-stroke memory deficits remain to be fully elucidated.
Pharmacological treatment with antidepressants
initiated timely following a stroke can prevent the development of
PSD (12-14). It has also been reported that
antidepressant pharmacotherapy promotes long-term functional
recovery following a stroke, including everyday activities, and
cognitive and executive functioning (15-17). Studies have identified hyperforin
as the major active compound in St. John’s Wort, a plant used in
self-medicating mild to moderate forms of depression (18,19). A previous study reported that
14-day delayed treatment with hyperforin in the recovery phase of a
stroke promotes angiogenesis and improves functional recovery
following an ischemic stroke (20). Hippocampal neurogenesis is closely
associated with cognitive function and mood following a stroke
(21). It remains to be
elucidated whether delayed 7-day administration of the natural
antidepressant hyperforin to post-stroke socially isolated mice
during stroke recovery can promote hippocampal neurogenesis and
restore memory function via the inhibition of PSD and PSA.
Although there is controversy regarding the level of
transforming growth factor-β (TGF-β) in depression, studies have
demonstrated that patients with major depression disorder (MDD)
exhibit reduced serum levels of TGF-β (22,23) and a decrease in TGF-β
network-associated gene transcripts in the choroid plexus (24). A 6-week period of treatment with
the antidepressants fluoxetine, venlafaxine or paroxetine increases
the levels of TGF-β1 (25). TGF-β
members promote the proliferation of neuroepithelial stem cells,
neurite outgrowth and synapse formation (26). Whether the inhibition of TGF-β is
involved in post-stroke social isolation-mediated PSD and PSA and
the attenuation of hippocampal neurogenesis and memory function
remains to be elucidated.
The purpose of the present study was to investigate
whether delayed 7-day treatment with hyperforin to post-stroke
socially isolated mice can inhibit PSD and PSA via TGF-β, enhance
neurogenesis in the hippocampal dentate gyrus (DG) and consequently
improve impaired memory following ischemic stroke.
Materials and methods
Experimental animals
Adult male C57BL/6J mice (age, 8-10 weeks; weight,
23-25 g) were provided by Beijing Vital River Laboratory Animal
Technology Co., Ltd. (Beijing, China). The animals were maintained
at temperature of 23±11°C with 50±10% relative humidity in specific
pathogen-free conditions and had access to food and water ad
libitum. All procedures were performed in accordance with the
National Institute of Health Guide for the Care and Use of
Laboratory Animals (publication no. 80-23) revised 1996, and were
approved by the Animal Care and Use Committee for experimental
animals of Tongji Medical College (Wuhan, China).
Middle cerebral artery occlusion (MCAO)
procedure and housing conditions
Focal cerebral ischemia was induced by MCAO as
previously reported (27). The
mice were anesthetized by intraperitoneal injection of ketamine
(100 mg/kg) and xylazine (8 mg/kg). Under an operating microscope,
the right external carotid artery was incised and a 6-0
silicone-coated nylon monofilament with a rounded tip was inserted
into the internal carotid artery until it reached the bifurcation
of the MCA for occlusion for 60 min, followed by reperfusion. The
sham-operated mice underwent an identical surgical procedure
without ischemia. A laser-Doppler probe (Periflux system 5000;
Perimed AB, Stockholm, Sweden) was placed on the skull (5 mm
lateral and 2 mm posterior to the bregma) to monitor the regional
cerebral blood flow (rCBF) during MCAO. Mice with <25% rCBF
reduction following occlusion and ~80% rCBF increase upon
reperfusion were included in further analyses.
During the MCAO, the rectal temperature was
maintained at 37±0.5°C. Immediately following surgery and on a
daily basis thereafter, the MCAO mice in all housing groups
received a subcutaneous injection of normal saline (NS; 1%
v/w).
Each male mouse was randomly allocated to housing
groups in standard clear plastic cages (27×17×12.5 cm) either
individually (socially isolated, SI) or with an ovariectomized
female mouse (pair-housed, PH) immediately following ischemia for
14 continuous days.
Experimental groups and drug
administration
Experiment 1: The mice were randomly divided into
six groups (Fig. 1A): Group 1)
Sham operated SI mice (sham + SI; n=12 for behavioral assessment);
group 2) sham operated PH mice (sham + PH; n=12 for behavioral
assessment); group 3) post-stroke SI mice [MCAO + SI; n=12 for
behavioral assessment; n=6 for reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis; n=6 for western
blotting (WB); and n=6 for immunofluorescence (IF)]; group 4)
post-stroke PH mice (MCAO + PH; n=12 for behavioral assessment; n=6
for RT-qPCR; n=6 for WB; and n=6 for IF); group 5) post-stroke SI
mice treated with recombinant mouse (r) TGF-β (MCAO + SI + rTGF-β;
n=12 for behavioral assessment and n=6 for IF); and group 6)
post-stroke SI mice treated with PBS (MCAO + SI + PBS; n=12 for
behavioral assessment and n=6 for IF). Immediately following
surgery, a cannula was inserted into the left lateral ventricle
(0.1 mm posteriorly, 0.9 mm lateral to the midline and 3.1 mm deep
from the bregma) in groups 5 and 6 using a stereotaxic instrument
(RWD Life Science Co., Ltd., Shenzhen, China) for
intracerebroventricular (i.c.v.) injection (28). rTGF-β (2.5 µg/kg; Sino
Biological, Inc., Beijing, China) or sterile PBS was administrated
i.c.v. into the left lateral ventricle of the SI mice using a
sterile 26-G Hamilton microsyringe (80330; Hamilton Company, Reno,
NV, USA) every 24 h for 7 continuous days commencing at 7 days
post-ischemia (dpi).
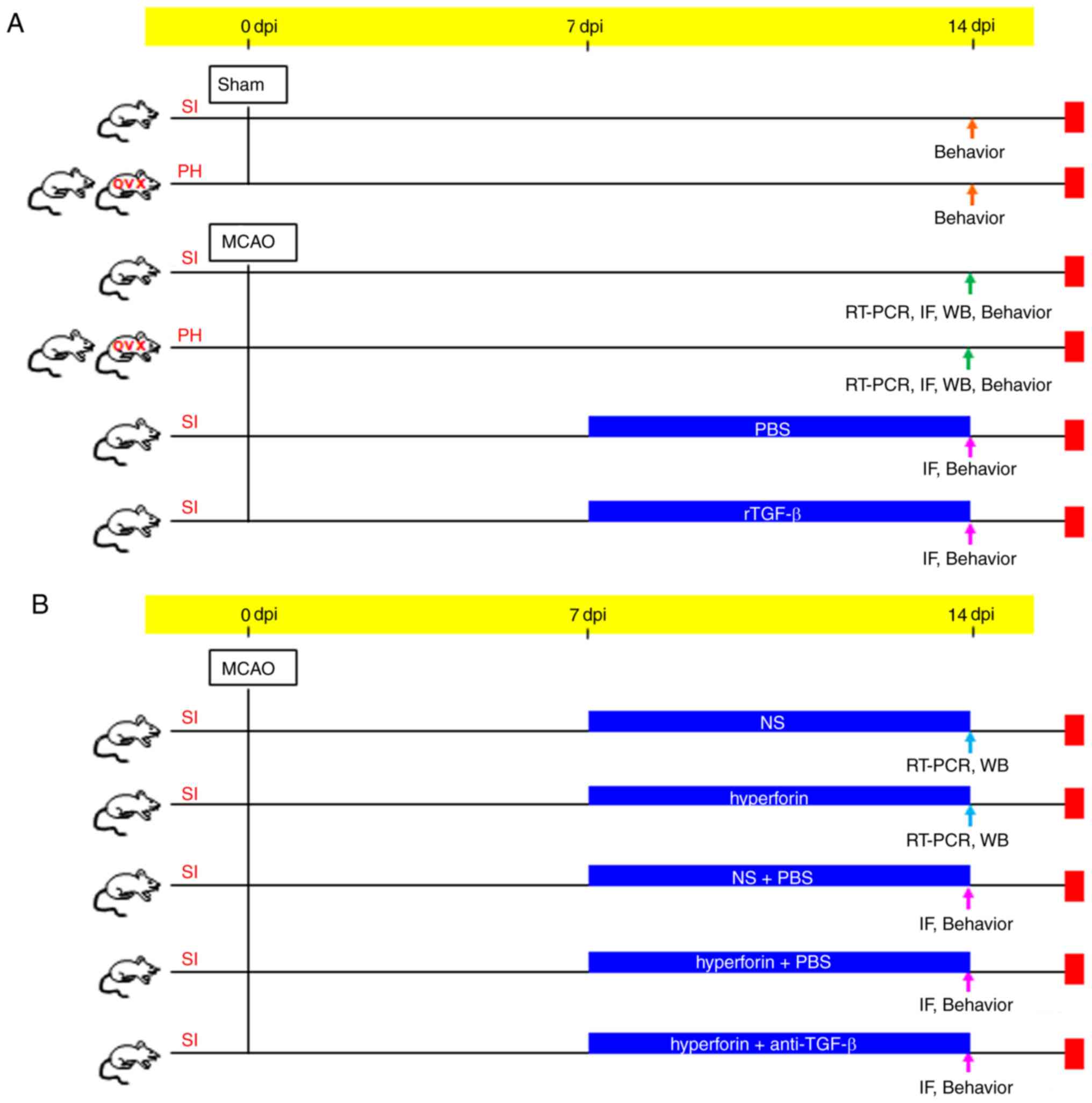 | Figure 1Schematic representation of the
animal grouping. Transient focal cerebral ischemia was induced by a
filament occlusion of the right middle cerebral artery. (A)
Experiment 1: Mice were PH or SI housed for 14 days immediately
following stroke. mRNA expression (RT-qPCR), protein expression
(IF, WB) and behavioral analyses were performed to examine the
effects of post-stroke social isolation on post-stroke depression
and anxiety, hippocampal neurogenesis and memory function. Mediated
effects of TGF-β on post-stroke social isolation were also
investigated. (B) Experiment 2: RT-qPCR, IF, WB and behavioral
analyses were performed to evaluate the effects of hyperforin on
the treatment of post-stroke social isolation-induced post-stroke
depression and anxiety, reduced hippocampal neurogenesis and memory
deficits. Effects of TGF-β on hyperforin-treated post-stroke social
isolation were also investigated. dpi, days post-ischemia; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; WB,
western blotting; IF, immunofluorescence; TGF-β, transforming
growth factor-β; anti-TGF-β, TGF-β-neutralizing antibody; rTGF-β,
recombinant mouse TGF-β; OVX, ovariectomized female mice; PH,
pair-housed male and OVX; SI, socially isolated. |
Experiment 2: The mice were randomly divided into
five groups (Fig. 1B): Group 1)
Post-stroke SI mice treated with hyperforin (MCAO + SI +
hyperforin; n=6 for RT-qPCR and n=6 for WB); group 2) post-stroke
SI mice treated with NS (MCAO + SI + NS; n=6 for RT-qPCR and n=6
for WB); group 3) post-stroke SI mice treated with NS plus PBS
(MCAO + SI + NS + PBS; n=12 for functional assays and n=6 for IF);
group 4) post-stroke SI mice treated with hyperforin plus
TGF-β-neutralizing antibody (MCAO + SI + hyperforin + anti-TGF-β;
n=12 for functional assays and n=6 for IF); and group 5)
post-stroke SI mice treated with hyperforin plus PBS (MCAO + SI +
hyperforin + PBS; n=12 for functional assays and n=6 for IF).
Hyperforin (Cayman Chemical Company, Ann Arbor, MI, USA) was
dissolved in ethanol at 20 µg/µl. The stock solution
was diluted with NS to 0.5 µg/µl and intranasally
administered to alternating nostrils with a 2 min interval between
each application, every 24 h for 7 days (starting at 7 dpi). Drops
(2 µl) were administered to the surface of each nare,
allowing the mice to inhale each drop into the nasal cavity
(29). A total of 10 µl
treatment was delivered over 5 min. The animals in groups 2 and 3
were treated accordingly using NS. TGF-β-neutralizing antibody (1
µg; Abcam, Cambridge, UK) was dissolved in PBS and i.c.v.
injected into the left lateral ventricle 30 min prior to the
application of hyperforin.
5-Bromo-2′-deoxyuridine (BrdU)
labeling
The mice were injected intraperitoneally twice with
S-phase marker BrdU (50 µg/g body weight in NS;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) with 8 h between
injections at 13 dpi. The mice were anesthetized and transcardially
perfused 1 day following the final injection to analyze
BrdU-labeling of the recently proliferated cells by
immunofluorescence staining and observed under a fluorescence
microscope (BX51; Olympus Corporation, Tokyo, Japan).
Behavioral assessments Forced swim task
(FST)
Depression-type behavior in the mice (n = 12/group)
was assessed using the FST at 14 dpi. The mice were placed into an
opaque cylinder tank (diameter, 24 cm; depth, 53 cm), containing 30
cm3 of 25°C water and the duration of immobility was
scored over 6 min. Immobility was assessed during the final 4 min
as previously described (10).
Quantification of floating, vs. swim time was analyzed using
Observer software (version 5; Exeter Software, Setauket, NY,
USA).
Sucrose consumption test (SCT)
To examine depressive-type behavior, the SCT was
performed at 14 dpi. Two identical 10-ml vials were placed on a
custom-made wire cage top. At 3 days prior to MCAO, the
individually-housed mice were provided with two 10-ml vials of
water for 12 h and then were returned to their original housing
condition and provided with their normal drinking water and cage
tops. After 12 h, the mice were provided with two different 10-ml
vials of 3% sucrose for 12 h. Following the completion of
habituation, all mice were returned to their original housing
condition and provided with their normal drinking water and cage
tops. The mice were deprived of water for 12 h prior to the test
and were then given access to 3% sucrose solution over 12 h. The
volume of 3% sucrose consumed was recorded.
Open field test (OFT)
Anxiety-type behavior in the mice (n = 12/group) was
assessed using the OFT at 14 dpi. The mice were placed in an open
field apparatus (40×40×37.5 cm) and allowed to explore for 20 min.
Locomotor activity was quantified using a photobeam activity system
(FlexField; San Diego Instruments, San Diego, CA, USA). The
relative level of activity occurring in the periphery, vs. center
squares was recorded.
Y-maze task
Spatial reference memory was assessed using the
continuous variant of the Y-maze spontaneous alternation procedure.
The Y-maze task was used with three identical open arms (50×16×32
cm). The mice were placed in the center of the maze and allowed to
explore the three maze arms freely for 8 min. The mice made correct
alternations when they sequentially visited the three arms without
repeated entry into a previously visited arm. The total arm entries
during the session were counted. Spontaneous alternation was
calculated as follows: Spontaneous alternation (%) =
[alternations/(total arm entries - 2)] × 100.
Novel object recognition task (NORT)
To assess recognition memory, the mice were placed
in a chamber (35×25×35 cm) made of gray plastic for 30 min 2 days
prior to the NORT. During the training sessions, two identical
objects were placed symmetrically in the center of the chamber and
the mouse was allowed to explore for 10 min. The time spent
exploring each object was recorded. During the retention tests, the
mice were placed in the chamber, in which one of the familiar
objects was replaced with a novel object, and the mice were allowed
to explore freely for 5 min. The mice were considered to be
exploring an object when they faced the object at a distance of ≤1
cm. A discrimination index was determined as the ratio of the time
spent exploring any one object (training session) or the novel
object (retention session) over the total time spent exploring both
objects.
Step-through passive avoidance task
To assess the contextual memory impairment, a
step-through passive avoidance apparatus containing light (14×10×25
cm) and dark (25×25×25 cm) compartments equipped with an electric
grid floor was used. The mice were allowed to adjust to the
apparatus the day preceding the trials. For the training trials, a
mouse was placed in the light compartment. When the mouse crossed
to the dark compartment with all four paws, the door closed and the
mouse was punished by intermittent electric shocks (2 sec;
intensity, 0.8 mA). Following 30 sec, the mouse was removed from
the apparatus. The retention tests were conducted 24 h following
the training session (without electric shock) and observed for ≤300
sec.
Western blotting
Brain tissues of the ischemic hemisphere was lysed
in RIPA lysis buffer containing protease and phosphatase inhibitors
(Nanjing KeyGen Biotech Co., Ltd., Nanjing, China). An equal
quantity of protein (30-50 µg/lane) was resolved on a 10%
SDS-PAGE gel (Beyotime Institute of Biotechnology, Shanghai, China)
and blotted onto a PVDF membrane (Millipore; Merck KGaA) using an
electrophoresis apparatus (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The membranes were blocked with 5% non-fat milk/TBST to
reduce nonspecific binding and subsequently incubated with specific
rabbit anti-TGF-β monoclonal antibody (Abcam; cat. no. ab64715) at
1:1,000 dilution and anti-β-actin (1:2,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA; cat. no. sc-10731) overnight
at 4°C. Following extensive washing with TBST, the membranes were
incubated with an appropriate peroxidase-conjugated secondary
antibody (1:3,000; Wuhan Sanying Biotechnology, Wuhan, China; cat.
no. LP1001b) for 2 h at room temperature. Following three washes
with TBST, chemiluminescent signals were visualized using
electrochemiluminescence western blotting detection reagents
(Millipore; Merck KGaA) and bands were captured using an UVP gel
documentation system (UVP, LLC, Phoenix, AZ, USA). The band
intensity was quantified using ImageJ software version 1.41
(National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence staining
The mice were anesthetized and transcardially
perfused with NS and 4% paraformaldehyde in PBS (pH 7.4). Following
blocking in normal goat serum (Sigma-Aldrich; Merck KGaA) to reduce
nonspecific binding, paraffin-embedded brain sections (thickness,
4-µm) were incubated overnight at 4°C with diluted
monoclonal primary antibodies (Cell Signaling Technology, Inc.,
Danvers, MA, USA) as follows: Rabbit Ki67 (1:100; cat. no. 9129),
rabbit phosphorylated-histone H3 (PH3; 1:50; cat. no. 9706), mouse
BrdU (1:100; cat. no. 5292), rabbit doublecortin (DCX; 1:50; cat.
no. 14802) and rabbit NeuN (1:200; cat. no. 12943). Following
washing with PBS, the slides were incubated with DyLight
549-conjugated goat anti-rabbit (cat. no. A23320) or DyLight
488-conjugated goat anti-mouse secondary antibodies (cat. no.
A23210) at a 1:200 dilution (Abbkine Scientific Co., Ltd., Wuhan,
China) at 37°C for 2 h. The antibodies were diluted in 1% bovine
serum albumin (Sigma-Aldrich; Merck KGaA) in PBS. Nuclei were
stained with DAPI and slides were observed using a fluorescence
microscope (BX51; Olympus Corporation). In the hippocampal DG,
positive cells (Ki67+, PH3+ and
BrdU+/DCX+) were counted in the granule cell
layer of the dorsal DG of hippocampi (bregma from -2.3 to -4.5
mm).
RT-qPCR measurements
Total RNA from the post-ischemic hemisphere was
isolated using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer’s
protocol. cDNA was obtained using Taqman reverse transcriptase
(Applied Biosystems; Thermo Fisher Scientific, Inc.). TGF-β and
β-actin cDNA were amplified using Power SYBR-Green (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Two-step qPCR was
performed (95°C for 15 sec, 60°C for 60 sec for 40 cycles) with
specific primers for TGF-β (forward,
5′-GTGTGGAGCAACATGTGGAACTCTA-3′ and reverse,
5′-TTGGTTCAGCCACTGCCGTA-3′) and β-actin (forward,
5′-AAGGCCAACCGTGAAAAGAT-3′ and reverse,
5′-GTGGTACGACCAGAGGCATAC-3′). The relative quantitation value is
expressed as 2−ΔΔCq, where ΔCq is the difference between
the mean ΔCq value of duplicate measurements of the sample and
β-actin control (30).
Statistical analysis
Data from the behavioral assessments (FST, SCT and
OFT), RT-qPCR analysis and western blotting were analyzed by
two-way analysis of variance (ANOVA) with surgery and housing
condition as between subject factors. Other data were analyzed
using one-way ANOVA to compare several groups, followed by Fisher’s
Protected Least Significant Difference test for pairwise comparison
of groups. The results are presented as the mean ± standard error
of the mean. P<0.05 was considered to indicate a statistically
significant difference. Statistical analysis was performed using
GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
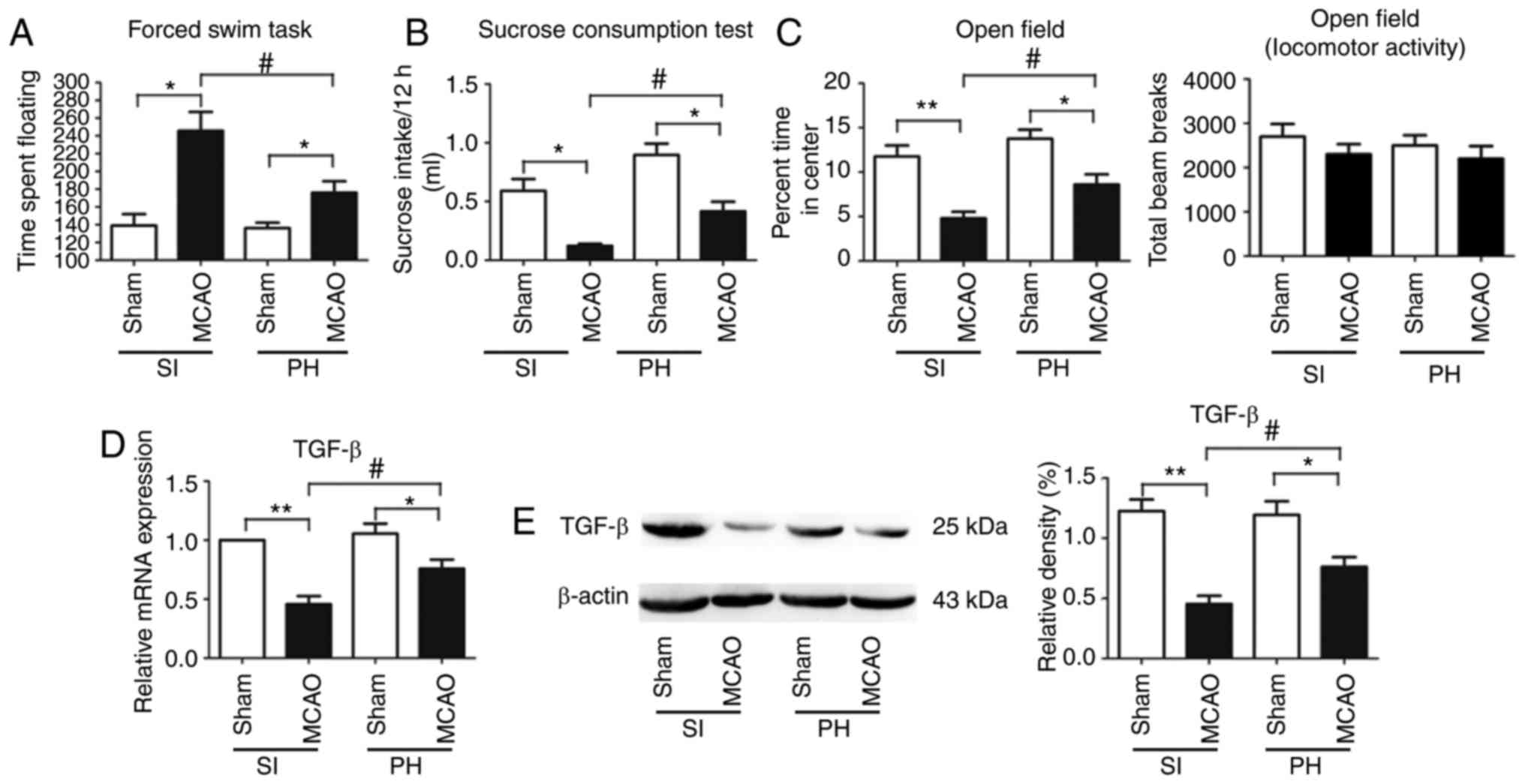
Post-stroke isolation leads to PSD and
PSA
Depression-type behavior was evaluated using FST and
SCT. The floating time in the FST was significantly increased
(P<0.05; Fig. 2A) and
consumption of sucrose was significantly decreased (P<0.05;
Fig. 2B) in the post-stroke SI
mice compared with the sham-operated SI mice at 14 dpi.
Furthermore, compared with the sham-operated PH mice at 14 dpi,
floating times in the post-stroke PH mice were significantly
increased (P<0.05; Fig. 2A)
and sucrose consumption was significantly decreased (P<0.05;
Fig. 2B). The post-stroke PH mice
exhibited a significant reduction in floating times (P<0.05;
Fig. 2A) and a significant
increase in sucrose consumption (P<0.05; Fig. 2B) compared with the post-stroke SI
mice at 14 dpi.
Anxiety-type behavior was measured by the OFT. The
post-stroke SI mice spent significantly less time in the center of
the open field chamber than the sham-operated SI mice at 14 dpi
(P<0.01; Fig. 2C). The
percentage of time spent in the center of the open field chamber
was further significantly reduced in the post-stroke PH mice
compared with the sham-operated PH mice at 14 dpi (P<0.05;
Fig. 2C). Post-stroke PH had a
significant effect on OFT; the post-stroke PH mice spent a
significantly increased duration in the center of the open field
chamber compared with the post-stroke SI mice (P<0.05; Fig. 2C). No significant difference was
observed in locomotor activity or exploration among the four groups
in the OFT (P>0.05; Fig.
2C).
Post-stroke isolation decreases the
expression of TGF-β in the ischemic hippocampus during stroke
recovery
The mRNA and protein expression levels of TGF-β in
the ischemic hippocampi were significantly decreased in the
post-stroke SI mice compared with those in the post-stroke PH mice
at 14 dpi (P<0.05; Fig. 2D and
E).
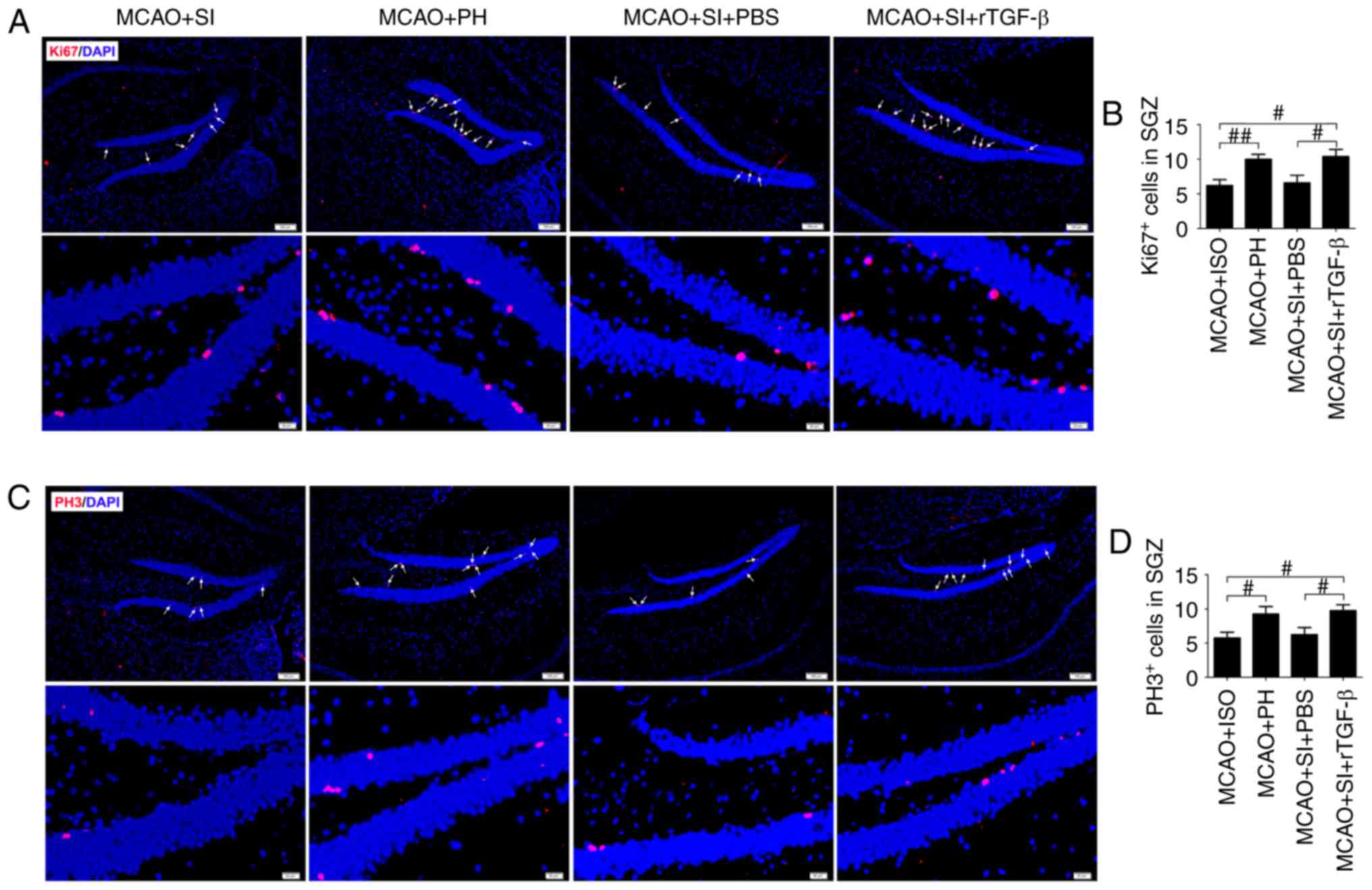
rTGF-β treatment restores the reduced
hippocampal neurogenesis induced by social isolation during stroke
recovery
The numbers of general proliferation marker
Ki67+ and M-phase-specific marker PH3+ cells
in the ischemic hippocampi were significantly decreased in the
post-stroke SI mice compared with those in the post-stroke PH mice
at 14 dpi (P<0.01 and P<0.05, respectively; Fig. 3A-D). The administration of rTGF-β
to the post-stroke SI mice significantly increased the numbers of
Ki67+ and PH3+ cells in the ischemic
hippocampus compared with those in the post-stroke SI mice treated
with PBS at 14 dpi (P<0.05; Fig.
3A-D). The post-stroke SI mice treated with rTGF-β exhibited
similar numbers of Ki67+ and PH3+ cells in
the ischemic hippocampi as the post-stroke PH mice (P>0.05;
Fig. 3A-D).
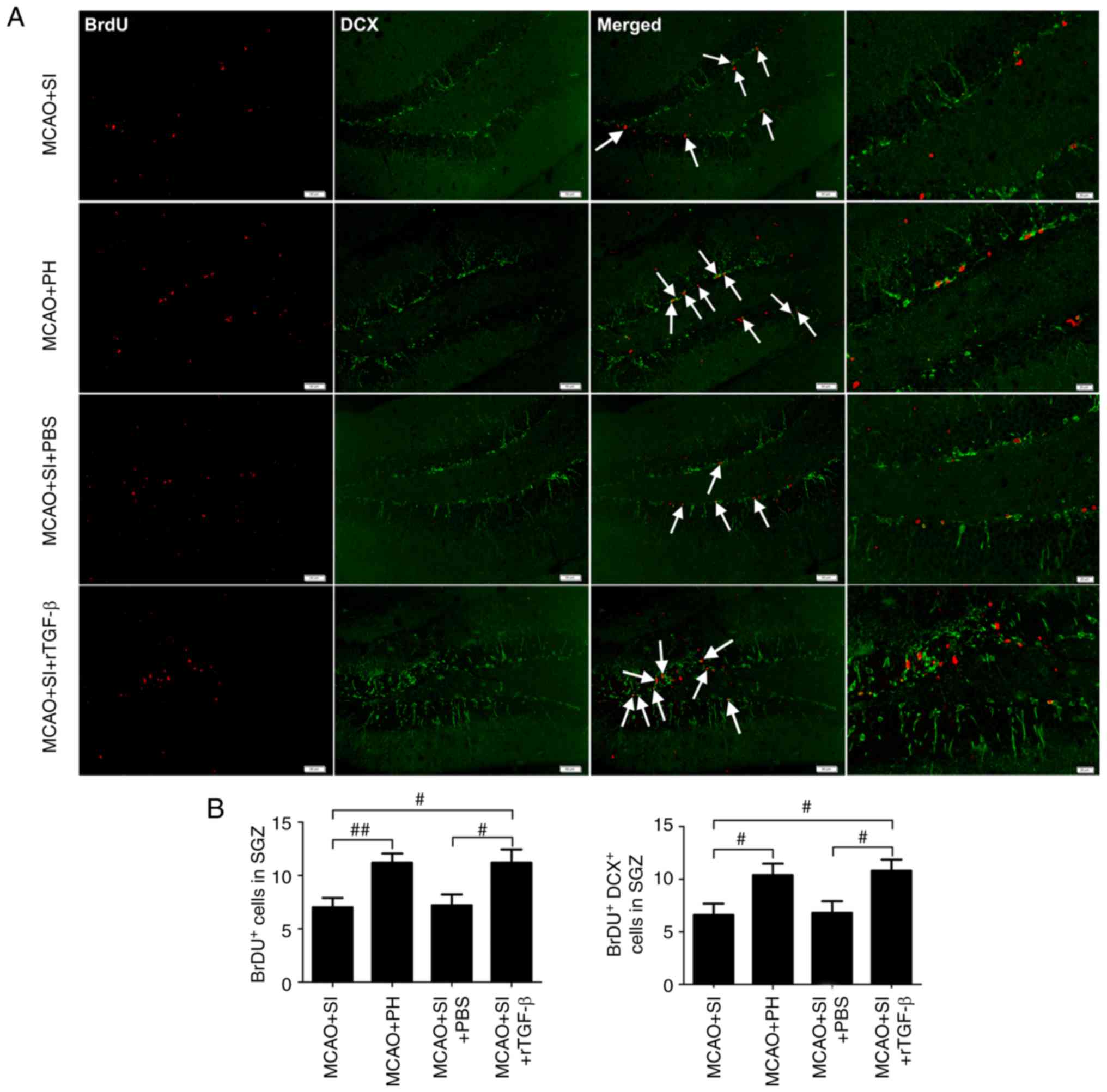
Cell proliferation was assessed at 13 dpi following
treatment of the mice with BrdU to label S-phase cells; on 14 dpi,
the dividing cells were analyzed. It was observed that the
post-stroke SI mice exhibited a significantly decreased number of
BrdU+ cells in the ischemic hippocampus compared with
the post-stroke PH mice (P<0.01; Fig. 4A and B). The post-stroke SI mice
administered with PBS had significantly fewer BrdU+
cells in the ischemic hippocampus compared with the post-stroke SI
mice administered with rTGF-β (P<0.05), which themselves had a
similar number of BrdU+ cells in the ischemic
hippocampus as the post-stroke PH mice (P>0.05; Fig. 4A and B).
DCX, a neuron-specific microtubule-associated
protein, expresses specifically in neuroblasts and functions as a
marker for neuronal precursors and neurogenesis. Quantitative
determination of BrdU/DCX double-labeled cells was performed to
assess adult hippocampal neurogenesis. Significantly lower numbers
of BrdU+/DCX+ cells in the ischemic
hippocampus were observed in the post-stroke SI mice compared with
the post-stroke PH mice at 14 dpi (P<0.05; Fig. 4A and B). Furthermore,
significantly lower numbers of BrdU+/DCX+
cells were observed in the ischemic hippocampus of the PBS-treated
post-stroke SI mice compared with the rTGF-β-treated post-stroke SI
mice (P<0.05), which themselves exhibited a similar number of
BrdU+/DCX+ cells in the ischemic hippo-campus
as the post-stroke PH mice (P>0.05; Fig. 4A and B).
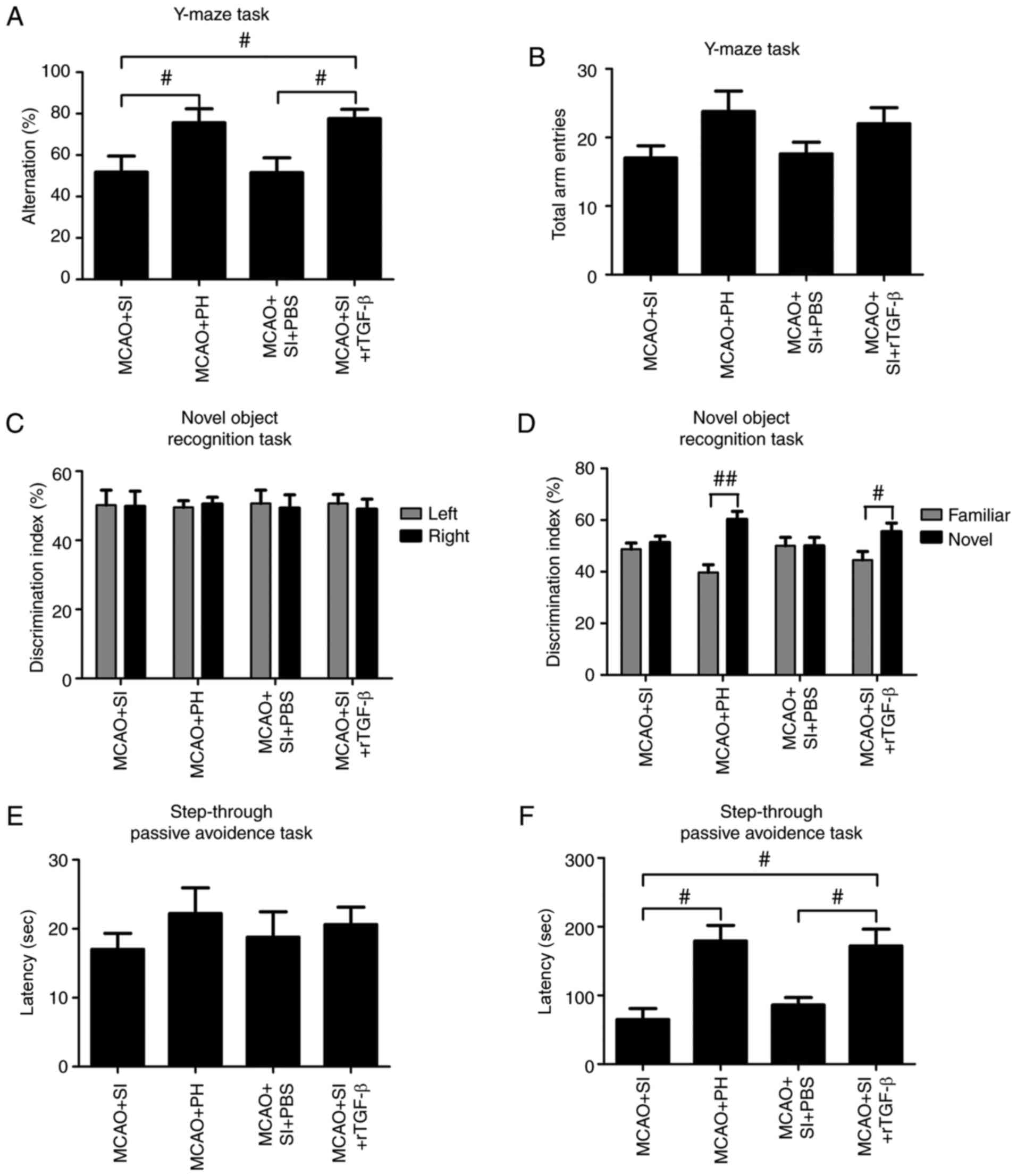
rTGF-β improves the social
isolation-induced exaggeration of impaired memory-associated
behaviors following a stroke
The percentage of correct alternations in the Y-maze
was significantly lower in the post-stroke SI mice compared with
the post-stroke PH mice at 14 dpi (P<0.05; Fig. 5A); no significant change in the
number of total arm entries was observed (P>0.05; Fig. 5B). The post-stroke SI mice treated
with rTGF-β exhibited a significant increase in the percentage of
correct alternations compared with the PBS-treated post-stroke SI
mice (P<0.05; Fig. 5A); no
significant change in the number of total arm entries was observed
(P>0.05; Fig. 5B).
In the NORT, the mice spent a comparable duration
exploring each object, yielding a discrimination index of ~50% in
all groups during the training session (Fig. 5C). During the retention trial
performed 1 h following training, the post-stroke PH mice spent a
significantly longer duration exploring the novel object compared
with the familiar object (P<0.01; Fig. 5D), whereas the post-stroke SI mice
explored the objects equally (P>0.05; Fig. 5D). The post-stroke SI mice treated
with rTGF-β exhibited a significant preference for the novel object
compared with the PBS-treated post-stroke SI mice (P<0.05;
Fig. 5D).
In the passive-avoidance task, no significant
differences were observed in step-through latency times without
shocks among groups (P>0.05; Fig.
5E). Post hoc analysis of the animals assessed following
training revealed that the post-stroke PH mice exhibited
significant higher step-through latency times 24 h following shocks
compared with the post-stroke SI mice at 14 dpi (P<0.05;
Fig. 5F). Treatment with rTGF-β
significantly restored the reduced step-through latency times in
the post-stroke SI mice during the retention test (P<0.05;
Fig. 5F).
Hyperforin improves the social
isolation-induced exaggeration of PSD and PSA via TGF-β
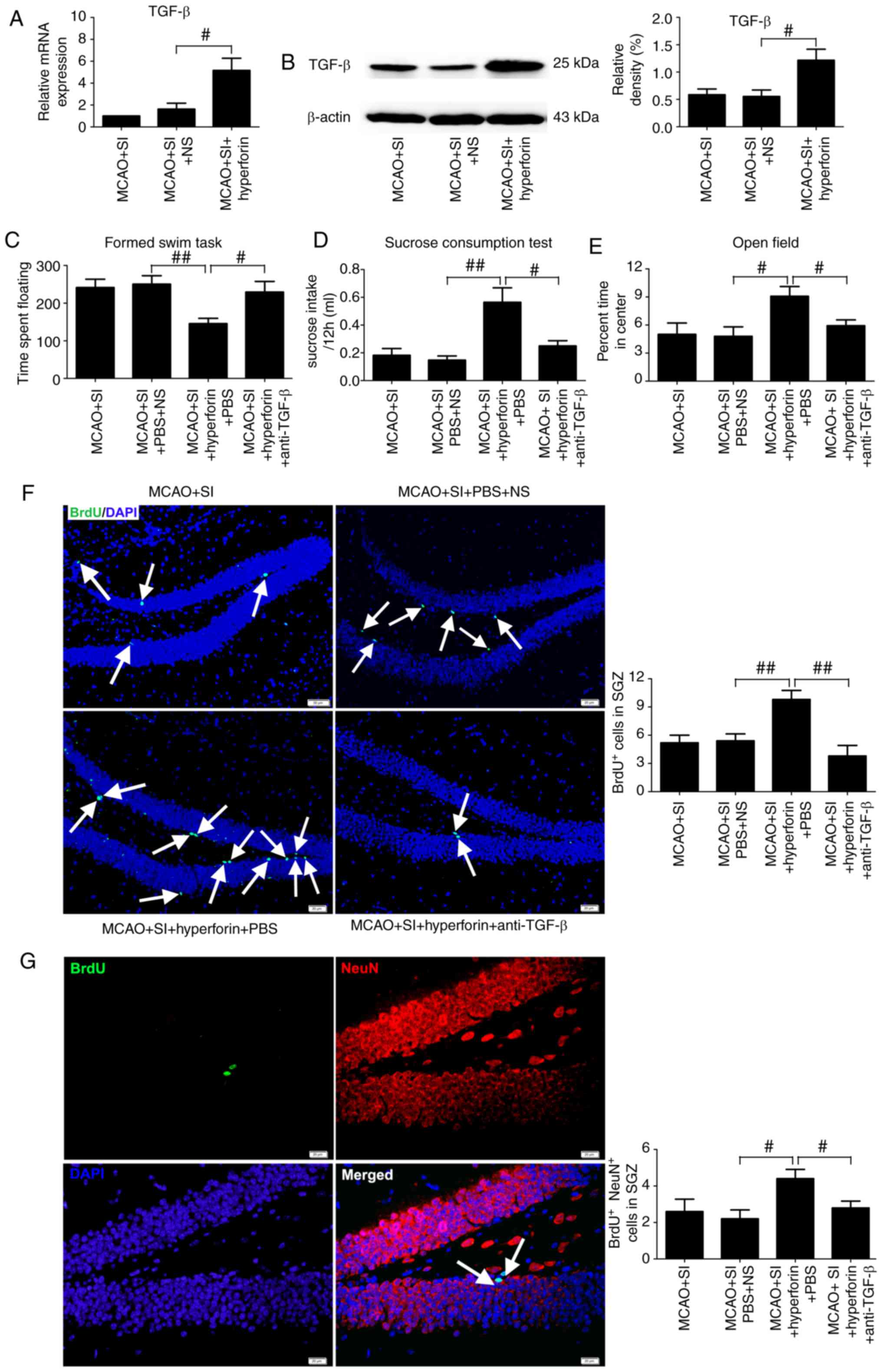
Intranasal administration of hyperforin to the
post-stroke SI mice significantly increased the mRNA and protein
levels of TGF-β in the ischemic hippocampi compared with those in
the NS-treated post-stroke SI mice at 14 dpi (P<0.05; Fig. 6A and B).
The floating times in the FST were significantly
decreased (P<0.01; Fig. 6C)
and sucrose consumption was significantly increased (P<0.01;
Fig. 6D) in the
hyperforin-treated post-stroke SI mice compared with the NS-treated
post-stroke SI mice at 14 dpi. Application of a TGF-β-neutralizing
antibody significantly reversed the hyperforin-mediated decrease in
floating times (P<0.05; Fig.
6C) and increase in sucrose consumption (P<0.05; Fig. 6D) in the post-stroke SI mice.
The percentage of time spent in the center of the
open field chamber was significantly higher in the
hyperforin-treated post-stroke SI mice compared with that in the
NS-treated post-stroke SI mice at 14 dpi (P<0.05; Fig. 6E). TGF-β-neutralizing antibodies
reversed the hyperforin-mediated increase in the time spent in the
center of the open field chamber in the post-stroke SI mice
(P<0.05; Fig. 6E).
Hyperforin promotes decreased hippocampal
neurogenesis in social isolation following stroke via TGF-β
Intranasal administration of hyperforin to the
post-stroke SI mice led to a significant increase in the number of
BrdU+ and BrdU+/NeuN+ cells in
ischemic hippocampi compared with that in the NS-treated
post-stroke SI mice at 14 dpi (P<0.01 and P<0.05,
respectively; Fig. 6F and G).
TGF-β-neutralizing antibody treatment reversed the
hyperforin-mediated increase in BrdU+ and
BrdU+/NeuN+ cells in the ischemic hippocampi
of post-stroke SI mice at 14 dpi (P<0.01 and P<0.05,
respectively; Fig. 6F and G).
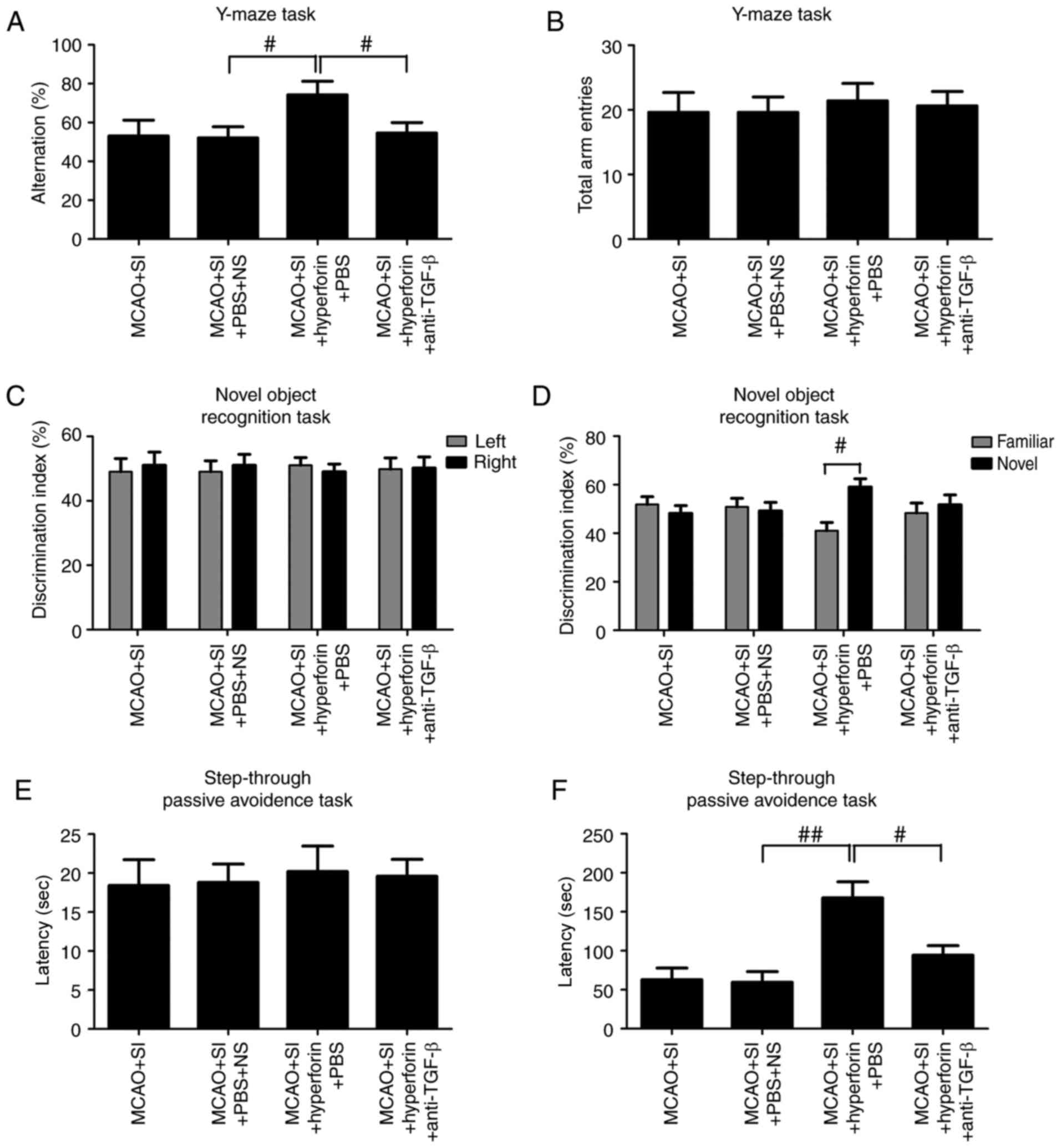
Hyperforin improves impaired
memory-associated behaviors in social isolation following a stroke
via TGF-β
The percentage of correct alternations in the Y-maze
was significantly higher in the hyperforin-treated post-stroke SI
mice compared with the NS-treated post-stroke SI mice at 14 dpi
(P<0.05; Fig. 7A) and no
significant change in the number of total arm entries was observed
(P>0.05; Fig. 7B).
TGF-β-neutralizing antibody treatment significantly reversed the
hyperforin-mediated increase in the percentage of correct
alternations in the post-stroke SI mice at 14 dpi (P<0.05;
Fig. 7A), without significant
changes to the number of total arm entries (P>0.05; Fig. 7B).
During the training session in the NORT, mice spent
a comparable time exploring the same objects in all groups
(P>0.05; Fig. 7C). During the
retention trial performed 1 h following training, the
hyperforin-treated post-stroke SI mice spent significantly more
time exploring the novel object compared with the familiar object
(P<0.05; Fig. 7D) and the
NS-treated post-stroke SI mice explored the objects equally
(P>0.05; Fig. 7D). The
hyperforin-treated post-stroke SI mice treated with
TGF-β-neutralizing antibody exhibited significantly impaired
discrimination between familiar and novel objects at 14 dpi
(P<0.05; Fig. 7D).
In the passive-avoidance task, no significant
differences in step-through latency times without shock were
observed among groups (P>0.05; Fig. 7E). Post hoc analysis following
training revealed that hyperforin treatment significantly increased
step-through latency times compared with those in the NS-treated
post-stroke SI group at 14 dpi (P<0.01; Fig. 7F). TGF-β-neutralizing antibody
treatment significantly reversed this hyperforin-mediated increase
in step-through latency times (P<0.05; Fig. 7F).
Discussion
The results of the present study indicated that
daily nasal administration of hyperforin during stroke recovery
ameliorates the post-stroke isolation-mediated exaggeration of
reduced hippocampal neurogenesis and impaired memory-associated
behaviors following ischemic stroke. Several critical observations
were made: i) Post-stroke social isolation-mediated PSD and PSA may
reduce hippocampal neurogenesis and impaired memory function via
TGF-β; ii) nasal administration of hyperforin improved PSD and PSA
via TGF-β in post-stroke SI mice; iii) reduced hippocampal
neurogenesis and impaired memory function in post-stroke SI mice
was restored following hyperforin treatment via TGF-β.
Increasing evidence has demonstrated that social
interaction is the most recognized mental and psychological factor
influencing post-stroke mortality rate and functional recovery
(31-33). Social interaction exerts prominent
beneficial effects on infarct volume and functional recovery in
both genders (34). Stroke
survivors are at increased risk of experiencing social isolation,
often due to mobility limitations. Considering the fact that many
socially isolated patients are not clinically identified until they
seek medical attention following a stroke, assessing the effects of
post-stroke social isolation compared with pre-stroke social
isolation on clinical outcomes is critical to developing therapies
targeting socio-emotional factors in neural regeneration and
functional recovery. Pre-stroke social isolation increases the
volume of infarcts following ischemic stroke (34), and previous studies have
demonstrated that infarct volume during the acute phase of a stroke
significantly influences neurogenesis during stroke recovery.
Infarct size is positively associated with neurogenesis following
ischemic stroke (35). To avoid
the effects of infarct volume on post-stroke neurogenesis, social
isolation was implemented immediately following a stroke rather
than pre-stroke. In the present study, a PSD and PSA model was
successfully established by MCAO using male C57BL/6 mice combined
with post-stroke isolated-housing initiated immediately following
ischemia. The data obtained are consistent with findings from a
previous study demonstrating that mice isolated immediately
following a stroke exhibited PSD and PSA (10). Of note, no significant differences
in PSD or PSA were observed between the sham groups in the SI or PH
mice.
In the adult central nervous system (CNS), cell
proliferation occurs throughout adulthood in two main neurogenic
niches that contain neural stem cells: The subventricular zone of
the lateral cerebral ventricles and the DG subgranular zone. These
neurogenic regions harbor adult neural progenitor cells (36). Hippocampal neurogenesis is closely
associated with cognitive function and mood following a stroke, and
impaired hippocampal neurogenesis is accompanied by poor learning
and memory (21). Evidence
supports that PSD and PSA are associated with reduced neurogenesis
following ischemic stroke (37,38). In addition, clinical studies have
implicated the detrimental effect of PSD on the recovery of
function and cognitive abilities (39,40). The present study demonstrated for
the first time, to the best of our knowledge that post-stroke
isolation inhibited hippocampal neurogenesis leading to memory
deficits. These findings suggest that induced PSD and PSA may have
important effects on hippocampal neurogenesis and memory function
in post-stroke isolation.
Increasing evidence indicates that members of the
TGF-β family, in addition to exerting well-established neurotrophic
and neuroprotective effects in the CNS, are important in
neuropsychiatric diseases, including anxiety, depression and other
neuropsychiatric disorders (26).
TGF-β network-associated gene transcripts in the choroid plexus and
levels of TGF-β in the serum are markedly decreased in patients
with MDD (22-24). In the present study, it was
demonstrated that SI mice exhibited decreased TGF-β levels in their
hippocampus during the stroke recovery phase. rTGF-β administration
restored the reduced hippocampal neurogenesis and impaired memory
function in the post-stroke SI mice. These observations suggested
that the social isolation-induced attenuation of TGF-β following a
stroke may be critical in reducing neurogenesis in the hippocampi
and memory function, and inducing anxiety and depression.
Hyperforin, the main active ingredient of St John’s
wort extract, exhibits antidepressant properties (18,19). A previous study demonstrated that,
in addition to its neuroprotective role in ischemic injury in the
acute phase following an ischemic stroke (41), hyperforin increases angiogenesis
and improves functional recovery (20). In the present study, it was
observed for the first time, to the best of our knowledge, that the
nasal administration of hyperforin inhibited PSD and PSA in SI mice
via TGF-β. Furthermore, during stroke recovery, these mice
exhibited increased hippocampal neurogenesis and memory function,
which was mediated by TGF-β. Taken together, these results
demonstrated the important modulatory effects of TGF-β on the
hyperforin-mediated inhibition of PSD and PSA, and its promoting
effects on hippocampal neurogenesis and memory function in SI mice.
Intranasal administration of hyperforin was selected in the present
study due to its noninvasive, rapid onset, safe and effective
characteristics (42,43). The treatment of CNS disorders is
often restricted due to drug transport across the blood-brain
barrier (BBB) and subsequently to the brain. The BBB provides a
major obstacle in the delivery of drugs to the CNS, with ~100% of
macromolecular drugs and >98% of small molecular drugs failing
to cross and enter the brain at pharmacologically significant
levels (44). Other than
essential nutrients, only small lipid-soluble molecules with
molecular weights <500 Da can effectively be transported across
the BBB and reach efficacious concentrations in the brain (45). Although invasive approaches for
delivering therapeutics to the brain, including i.c.v., have been
applied to bypass the BBB, these approaches are not clinically
practical due to their inconvenience, questionable safety and high
cost. Non-invasive intranasal delivery has been demonstrated to
circumvent the BBB and allows the direct and rapid delivery of
large and/or charged therapeutics to the CNS (43,46). Non-invasive intranasal
administration as an alternative to invasive delivery methods
represents the most promising and novel method for delivering
therapeutics directly to the brain (43). Therefore, the beneficial effects
of hyperforin administration via a nasal route may be considered in
developing therapeutic approaches for post-stroke social
isolation-mediated PSD and PSA with reduced hippocampal
neurogenesis and memory deficits.
In conclusion, the presented data demonstrate that
post-stroke social isolation-mediated PSD and PSA may reduce
hippocampal neurogenesis and impair memory function via TGF-β. The
intranasal delivery of hyperforin during stroke recovery activated
TGF-β, which may inhibit PSD and PSA and improve reduced
hippocampal neurogenesis and memory deficits in SI mice. Therefore,
hyperforin treatment via intranasal routes may be useful in
treating post-stroke social isolation-mediated decreased memory
function.
Acknowledgments
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81601156
and 81571286).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
SY and JZ contributed to the experimental design.
YZ, PY and HL established and performed the stroke model,
functional assays and western blotting. HY performed
immunocytochemistry and RT-qPCR analysis. JZ handled the imaging
tools. SY and JZ analyzed and interpreted the data. JZ prepared the
manuscript. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All procedures were performed in accordance with the
National Institute of Health Guide for the Care and Use of
Laboratory Animals (NIH publication no. 80-23) revised 1996, and
were approved by the Animal Care and Use Committee of experimental
animals of Tongji Medical College (Wuhan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hackett ML and Pickles K: Part I:
Frequency of depression after stroke: An updated systematic review
and meta-analysis of observational studies. Int J Stroke.
9:1017–1025. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Campbell Burton CA, Murray J, Holmes J,
Astin F, Greenwood D and Knapp P: Frequency of anxiety after
stroke: A systematic review and meta-analysis of observational
studies. Int J Stroke. 8:545–559. 2013. View Article : Google Scholar
|
|
3
|
Barker-Collo SL: Depression and anxiety 3
months post stroke: Prevalence and correlates. Arch Clin
Neuropsychol. 22:519–531. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morrison V, Pollard B, Johnston M and
MacWalter R: Anxiety and depression 3 years following stroke:
Demographic, clinical, and psychological predictors. J Psychosom
Res. 59:209–213. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ayerbe L, Ayis S, Wolfe CD and Rudd AG:
Natural history, predictors and outcomes of depression after
stroke: Systematic review and meta-analysis. Br J Psychiatry.
202:14–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Boden-Albala B, Litwak E, Elkind MS,
Rundek T and Sacco RL: Social isolation and outcomes post stroke.
Neurology. 64:1888–1892. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Whyte EM, Mulsant BH, Vanderbilt J, Dodge
HH and Ganguli M: Depression after stroke: A prospective
epidemiological study. J Am Geriatr Soc. 52:774–778. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ouimet MA, Primeau F and Cole MG:
Psychosocial risk factors in poststroke depression: A systematic
review. Can J Psychiatry. 46:819–828. 2001. View Article : Google Scholar
|
|
9
|
Meng C, Zhang JC, Shi RL, Zhang SH and
Yuan SY: Inhibition of interleukin-6 abolishes the promoting
effects of pair housing on post-stroke neurogenesis. Neuroscience.
307:160–170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
O’Keefe LM, Doran SJ, Mwilambwe-Tshilobo
L, Conti LH, Venna VR and McCullough LD: Social isolation after
stroke leads to depressive-like behavior and decreased BDNF levels
in mice. Behav Brain Res. 260:162–170. 2014. View Article : Google Scholar
|
|
11
|
Ebrahim S, Barer D and Nouri F: Use of the
nottingham health profile with patients after a stroke. J Epidemiol
Community Health. 40:166–169. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rasmussen A, Lunde M, Poulsen DL, Sorensen
K, Qvitzau S and Bech P: A double-blind, placebo-controlled study
of sertraline in the prevention of depression in stroke patients.
Psychosomatics. 44:216–221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Niedermaier N, Bohrer E, Schulte K,
Schlattmann P and Heuser I: Prevention and treatment of poststroke
depression with mirtazapine in patients with acute stroke. J Clin
Psychiatry. 65:1619–1623. 2004. View Article : Google Scholar
|
|
14
|
Robinson RG, Jorge RE, Moser DJ, Acion L,
Solodkin A, Small SL, Fonzetti P, Hegel M and Arndt S: Escitalopram
and problem-solving therapy for prevention of poststroke
depression: A randomized controlled trial. JAMA. 299:2391–2400.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Narushima K, Paradiso S, Moser DJ, Jorge R
and Robinson RG: Effect of antidepressant therapy on executive
function after stroke. Br J Psychiatry. 190:260–265. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Acler M, Robol E, Fiaschi A and Manganotti
P: A double blind placebo RCT to investigate the effects of
serotonergic modulation on brain excitability and motor recovery in
stroke patients. J Neurol. 256:1152–1158. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jorge RE, Acion L, Moser D, Adams HJ Jr
and Robinson RG: Escitalopram and enhancement of cognitive recovery
following stroke. Arch Gen Psychiatry. 67:187–196. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cervo L, Rozio M, Ekalle-Soppo CB, Guiso
G, Morazzoni P and Caccia S: Role of hyperforin in the
antidepressant-like activity of Hypericum perforatum extracts.
Psychopharmacology (Berl). 164:423–428. 2002. View Article : Google Scholar
|
|
19
|
Chatterjee S, Filippov V, Lishko P,
Maximyuk O, Noldner M and Krishtal O: Hyperforin attenuates various
ionic conductance mechanisms in the isolated hippocampal neurons of
rat. Life Sci. 65:2395–2405. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J, Yao C, Chen J, Zhang Y, Yuan S
and Lin Y: Hyperforin promotes post-stroke functional recovery
through interleukin (IL)-17A-mediated angiogenesis. Brain Res.
1646:504–513. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shetty AK: Hippocampal injury-induced
cognitive and mood dysfunction, altered neurogenesis, and epilepsy:
Can early neural stem cell grafting intervention provide
protection. Epilepsy Behav. 38:117–124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Musil R, Schwarz MJ, Riedel M, Dehning S,
Cerovecki A, Spellmann I, Arolt V and Muller N: Elevated macrophage
migration inhibitory factor and decreased transforming growth
factor-beta levels in major depression-no influence of celecoxib
treatment. J Affect Disord. 134:217–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sutcigil L, Oktenli C, Musabak U, Bozkurt
A, Cansever A, Uzun O, Sanisoglu SY, Yesilova Z, Ozmenler N,
Ozsahin A and Sengul A: Pro- and anti-inflammatory cytokine balance
in major depression: Effect of sertraline therapy. Clin Dev
Immunol. 2007.76396:2007.
|
|
24
|
Turner CA, Thompson RC, Bunney WE,
Schatzberg AF, Barchas JD, Myers RM, Akil H and Watson SJ: Altered
choroid plexus gene expression in major depressive disorder. Front
Hum Neurosci. 8:2382014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee KM and Kim YK: The role of IL-12 and
TGF-beta1 in the pathophysiology of major depressive disorder. Int
Immunopharmacol. 6:1298–1304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krieglstein K, Zheng F, Unsicker K and
Alzheimer C: More than being protective: Functional roles for
TGF-β/activin signaling pathways at central synapses. Trends
Neurosci. 34:421–429. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Mao X, Zhou T, Cheng X and Lin Y:
IL-17A contributes to brain ischemia reperfusion injury through
calpain-TRPC6 pathway in mice. Neuroscience. 274:419–428. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu J, Wu DM, Zheng YL, Hu B, Cheng W,
Zhang ZF and Shan Q: Ursolic acid improves high fat diet-induced
cognitive impairments by blocking endoplasmic reticulum stress and
IκB kinase β/nuclear factor-κB-mediated inflammatory pathways in
mice. Brain Behav Immun. 25:1658–1667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma M, Ma Y, Yi X, Guo R, Zhu W, Fan X, Xu
G, Frey WH II and Liu X: Intranasal delivery of transforming growth
factor-beta1 in mice after stroke reduces infarct volume and
increases neurogenesis in the subventricular zone. BMC Neurosci.
9:1172008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Hinojosa R, Haun J, Hinojosa MS and
Rittman M: Social isolation poststroke: Relationship between
race/ethnicity, depression, and functional independence. Top Stroke
Rehabil. 18:79–86. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Steptoe A, Shankar A, Demakakos P and
Wardle J: Social isolation, loneliness, and all-cause mortality in
older men and women. Proc Natl Acad Sci USA. 110:5797–5801. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Morris PL, Robinson RG and Raphael B:
Prevalence and course of depressive disorders in hospitalized
stroke patients. Int J Psychiatry Med. 20:349–364. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Venna VR, Weston G, Benashski SE,
Tarabishy S, Liu F, Li J, Conti LH and McCullough LD: NF-κB
contributes to the detrimental effects of social isolation after
experimental stroke. Acta Neuropathol. 124:425–438. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Moraga A, Pradillo JM, Cuartero MI,
Hernandez-Jimenez M, Oses M, Moro MA and Lizasoain I: Toll-like
receptor 4 modulates cell migration and cortical neurogenesis after
focal cerebral ischemia. FASEB J. 28:4710–4718. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gross CG: Neurogenesis in the adult brain:
Death of adogma. Nat Rev Neurosci. 1:67–73. 2000. View Article : Google Scholar
|
|
37
|
Wang SH, Zhang ZJ, Guo YJ, Teng GJ and
Chen BA: Hippocampal neurogenesis and behavioural studies on adult
ischemic rat response to chronic mild stress. Behav Brain Res.
189:9–16. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Loubinoux I, Kronenberg G, Endres M,
Schumann-Bard P, Freret T, Filipkowski RK, Kaczmarek L and
Popa-Wagner A: Post-stroke depression: Mechanisms, translation and
therapy. J Cell Mol Med. 16:1961–1969. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeong YJ, Kim WC, Kim YS, Choi KW, Son SY
and Jeong YG: The relationship between rehabilitation and changes
in depression in stroke patients. J Phys Ther Sci. 26:1263–1266.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mpembi MN, Miezi SM, Nzuzi TM, Massamba
VK, Henrard S, De Partz MP, Peeters A, Macq J, Dubois V and
Constant E: Clinical profile of post-cerebrovascular depression:
Descriptive cross-sectional study in the rehabilitation center for
people with disabilities of kinshasa (DR Congo). Pan Afr Med J.
17:1092014.In French.
|
|
41
|
Lin Y, Zhang JC, Fu J, Chen F, Wang J, Wu
ZL and Yuan SY: Hyperforin attenuates brain damage induced by
transient middle cerebral artery occlusion (MCAO) in rats via
inhibition of TRPC6 channels degradation. J Cereb Blood Flow Metab.
33:253–262. 2013. View Article : Google Scholar :
|
|
42
|
Chapman CD, Frey WH II, Craft S, Danielyan
L, Hallschmid M, Schioth HB and Benedict C: Intranasal treatment of
central nervous system dysfunction in humans. Pharm Res.
30:2475–2484. 2013. View Article : Google Scholar :
|
|
43
|
Illum L: Transport of drugs from the nasal
cavity to the central nervous system. Eur J Pharm Sci. 11:1–18.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pardridge WM: Drug targeting to the brain.
Pharm Res. 24:1733–1744. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Abbott NJ: Blood-brain barrier structure
and function and the challenges for CNS drug delivery. J Inherit
Metab Dis. 36:437–449. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dhuria SV, Hanson LR and Frey WH II:
Intranasal delivery to the central nervous system: Mechanisms and
experimental considerations. J Pharm Sci. 99:1654–1673. 2010.
View Article : Google Scholar
|