Introduction
Cutaneous melanoma is transformed from melanocytes,
and the predominant cause of melanoma is believed to be long-term
UV irradiation (1). Melanoma has
a highly invasive nature; even a primary tumor with a diameter of
2.5-4 mm can metastasize to multiple organs in the whole body,
leading to a very poor prognosis (2,3).
In 2014, there were >76,000 new cases of melanoma in the United
States and >9,000 patients succumbed to melanoma (4). Sadly, at present, the 5-year overall
survival (OS) rate for patients with stage IV melanoma is <15%
(5,6) Furthermore, patients suffering from
metastatic melanoma have a poor prognosis (1,3).
Thus, identifying novel biomarkers related to the prognosis and
progression of melanoma may improve the treatment and outcomes for
patients with melanoma.
The human cell division cycle associated 8 (CDCA8)
protein is an integral part of the vertebrate chromosomal passenger
complex (CPC) (7). The CPC
consists of at least four proteins: Aurora B, inner centromere
protein (INCENP), survivin, and CDCA8, overexpression of which is
associated with tumorigenesis of human cancers (8,9).
CDCA8 is a putative oncogene that is upregulated in multiple types
of cancer and is necessary for the survival and malignant nature of
various cancer cells (10). Its
overexpression is involved in the progression of lung cancer and
breast cancer (11), and can
result in poor prognosis of gastric cancer and lung cancer
(12). Thus, CDCA8 is considered
to be a promising target for the development of novel therapeutics
and diagnostics. Despite these previous studies, the role of CDCA8
on cutaneous melanoma remains unexplored.
Rho-associated coiled-coil-containing protein kinase
(ROCK) is a serine threonine kinase modulating several critical
cellular processes, including actin cytoskeleton organization,
apoptosis, reactive oxygen species formation, cell migration and
adhesion (13-15). In mammalians, two highly
homologous isoforms, ROCK1 and ROCK2 has been identified. ROCK has
several phosphorylation substrates, including myosin light chain
(MLC), myosin light chain phosphatase (MLCP), and LIM kinase (LIMK)
(16), and recent studies have
identified ROCK as a promising candidate for the therapy of highly
metastatic cancer (17).
The present study aimed to investigate the role of
CDCA8 in cutaneous melanoma and its effect on the ROCK signaling
pathway. First, the mRNA expression of CDCA8 in cutaneous melanoma
tissues and the association between CDCA8 expression and cutaneous
melanoma patient survival were examined, using the ONCOMINE and
Gene Expression Omnibus (GEO) databases. Second, the effects and
functional mechanism of CDCA8 on cutaneous melanoma cell lines were
investigated in vitro.
Methods and materials
Database analysis
The data of CDCA8 expression in cutaneous melanoma
were downloaded from the ONCOMINE (http://www.oncomine.org) database, which contained 45
cutaneous melanoma samples and 7 normal samples. In addition, CDCA8
expression data (GSE8401) for melanoma metastases were downloaded
from the public GEO (https://www.ncbi.nlm.nih.gov/geo/) database, which
contained 31 primary melanoma tumor samples and 52 metastatic
melanoma tumor samples.
The relationship of CDCA8 expression with clinical
features in cutaneous melanoma patients was analyzed using data
downloaded from The Cancer Genome Atlas (TCGA; https://cancergenome.nih.gov/) database. Based on
these data, a statistical analysis of the effect of CDCA8 on the
prognosis of cutaneous melanoma was performed.
Cell culture
The human cutaneous melanoma cell lines A375 and
MV3, and the normal human cells Hacat were purchased from the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China). Cells were routinely cultured in RPMI-1640 culture medium
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in a 5%
CO2 atmosphere, supplemented with 100 U/ml penicillin,
0.1 mg/ml streptomycin (both from Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and a fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) concentration of 10%. The cells were washed 3
times with PBS and digested with trypsin following growth to the
logarithmic phase. A single cell suspension was then seeded into
six-well plates for following experiments.
Cell transfection
When cell density in the six-well plates reached
~80%, according to the manufacturer’s instructions, the cells were
transfected with small interfering (si)RNA targeting CDCA8
(si-CDCA8, 5′-GUG GAA AUA CGA AUC AAG C-3′), or non-specific
control si-RNA (si-con; 5′-UUC UCC GAA CGU GUC ACG U-3′) using
Lipofectamine 2000 transfection reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for 24 h. Afterwards, the effect of the siRNA
transfection was verified by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR), and the cells were used for
subsequent experiments.
RT-qPCR assay
Following transfection for 24 h, total RNA was
extracted from cells by 1.0 ml TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA was synthesized into cDNA with the
reverse transcriptase M-MLV kit (Takara Biotechnology Co., Ltd.,
Dalian, China). The expression levels of CDCA8 were detected by
qPCR using the SYBR Premix Ex Taq kit (Applied Biosystems, Foster
City; Thermo Fisher Scientific, Inc.). The primers were: GAPDH
forward, 5′-GGA GCG AGA TCC CTC CAA AAT-3′ and reverse, 5′-GGC TGT
TGT CAT ACT TCT CAT GG-3′; and CDCA8 forward, 5′-GTT CCA AGG AGA
AGC CCA CA-3′ and reverse, 5′-CCG GTC CAC ATT CTC TGG AA-3′. GAPDH
was used as an internal control. The specific thermo-cycling
conditions were as follows: 5 min at 95°C, followed by 40 cycles of
95°C for 30 sec and 60°C for 45 sec, and a final step of 72°C for
30 min. The relative quantification of CDCA8 expression was
determined by the 2-ΔΔCq method (18) and each sample was repeated three
times independently.
Cell proliferation assay
At 24 h post-transfection, cells were counted and
100 µl cell suspensions (1,000 cells/well) were seeded into
96-well plates. According to the manufacturer’s introductions, the
viability of A375 and MV3 cells was detected every 24 h (at 24, 48,
72 and 96 h) using the Cell Counting kit-8 assay (CCK-8; Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China), after
10 µl CCK-8 reagent was added in each well. A proliferation
curve was plotted based on optical density (OD) values measured at
450 nm by a micro-plate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Every sample was assayed three times.
Colony formation assay
Cell suspensions (400 cells/plate) were seeded into
60 mm plates containing 5 ml culture medium at 37°C, following
transfection for 24 h. After 2 weeks, visible clones could be
observed in the culture plates. Subsequently, colonies were fixed
and stained for 30 min, following washing 2 times with PBS. The
number of colonies in the two groups were counted and compared
under a microscope in 5 random optical fields. Each sample was
repeated in triplicate.
Matrigel and wound healing assay
Matrigel and the wound healing assay were used to
detect cell invasion and migration, respectively. For cell
invasion, 100 µl Matrigel (BD Biosciences, Franklin Lakes,
NJ, USA; serum-free medium diluted 1:6) was added to the upper
chamber of a 24-well plate Transwell chamber and then placed in a
37°C 5% CO2 incubator for 4-6 h to form a gel.
Subsequently, 100 µl transfected cell suspension
(1×105) was added in the upper chamber, while 500
µl complete medium was added into the bottom chamber.
Following overnight culture so that the cells invade to the lower
surface of the filter, cells were washed and fixed with 4%
paraformaldehyde for 30 min. Finally, invaded cells were stained
with 0.1% crystal violet for 20 min and 5 visual fields were
selected randomly using a BX51 inverted microscope (Olympus
Corporation, Tokyo, Japan) at magnification, ×100.
For the wound healing assay, cells were cultured in
RPMI-1640 medium in six-well plates (5×105 cells/ml) for
24 h, then a wound was made in each plate with a 100 µl
plastic pipette tip. After being washed three times with PBS, the
cells were cultured for another 24 h. The wound width was then
observed in each well using an Olympus BX51 microscope (Olympus
Corporation) at magnification, ×200 in five random fields. Each
sample was performed in triplicate.
Western blot analysis
Following transfection for 24 h, total proteins were
extracted from the cells with the Radio Immunoprecipitation Assay
(RIPA) lysis buffer (Beyotime Institute of Biotechnology, Shanghai,
China). Then the protein concentrations were quantified with a BCA
Protein Assay kit (Beyotime Institute of Biotechnology). Total
protein samples (20 µg/lane) were separated by SDS-PAGE in a
10% polyacrylamide sequencing gel and transferred onto a
polyvinylidene fluoride (PVDF) membrane. Following blocking with 5%
non-fat dry milk for 1 h, the membranes were incubated with primary
antibodies overnight at 4°C. After washing for 3-5 min, the
membranes were incubated with secondary antibodies at room
temperature for 1 h. The antibodies were as follows: ROCK1 Rabbit
Polyclonal antibody (cat. no. 21850-1-AP; 1:1,000 dilution),
β-Actin Rabbit Polyclonal antibody (cat. no. 20536-1-AP; 1:1,000
dilution), MLC 2 Rabbit Polyclonal antibody (cat. no. 55462-1-AP;
1:2,000 dilution) and secondary horseradish peroxidase-labelled
goat anti-rabbit antibody (cat. no. 10545-2-AP; 1:5,000 dilution),
Rabbit anti-human tubulin (cat. no. 10068-1-AP; 1:5,000 dilution;
all ProteinTech Group, Inc., Chicago, IL, USA) was used as the
internal control and the relative expression of the target protein
was calculated relative to the tubulin expression, following
detection with an electro-chemiluminescence imaging analysis system
(Thermo Fisher Scientific, Inc.). The bands were scanned and
analyzed with the Quantity One software (Bio-Rad Laboratories,
Inc.).
Statistical analysis
The Kaplan-Meier’s estimator and the Log rank test
were used to detect the prognostic value of CDCA8 expression.
Comparisons between two groups were performed with the Student’s
t-test, while comparisons between multiple groups were performed
with one-way analysis of variance. Comparison between the groups
was made by a Student-Newman-Keuls test. All statistical analyses
were performed with SPSS 22.0 (IBM Corp., Armonk, NY, USA)
software. Data were presented as mean ± standard deviation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CDCA8 is upregulated in cutaneous
melanoma tissues
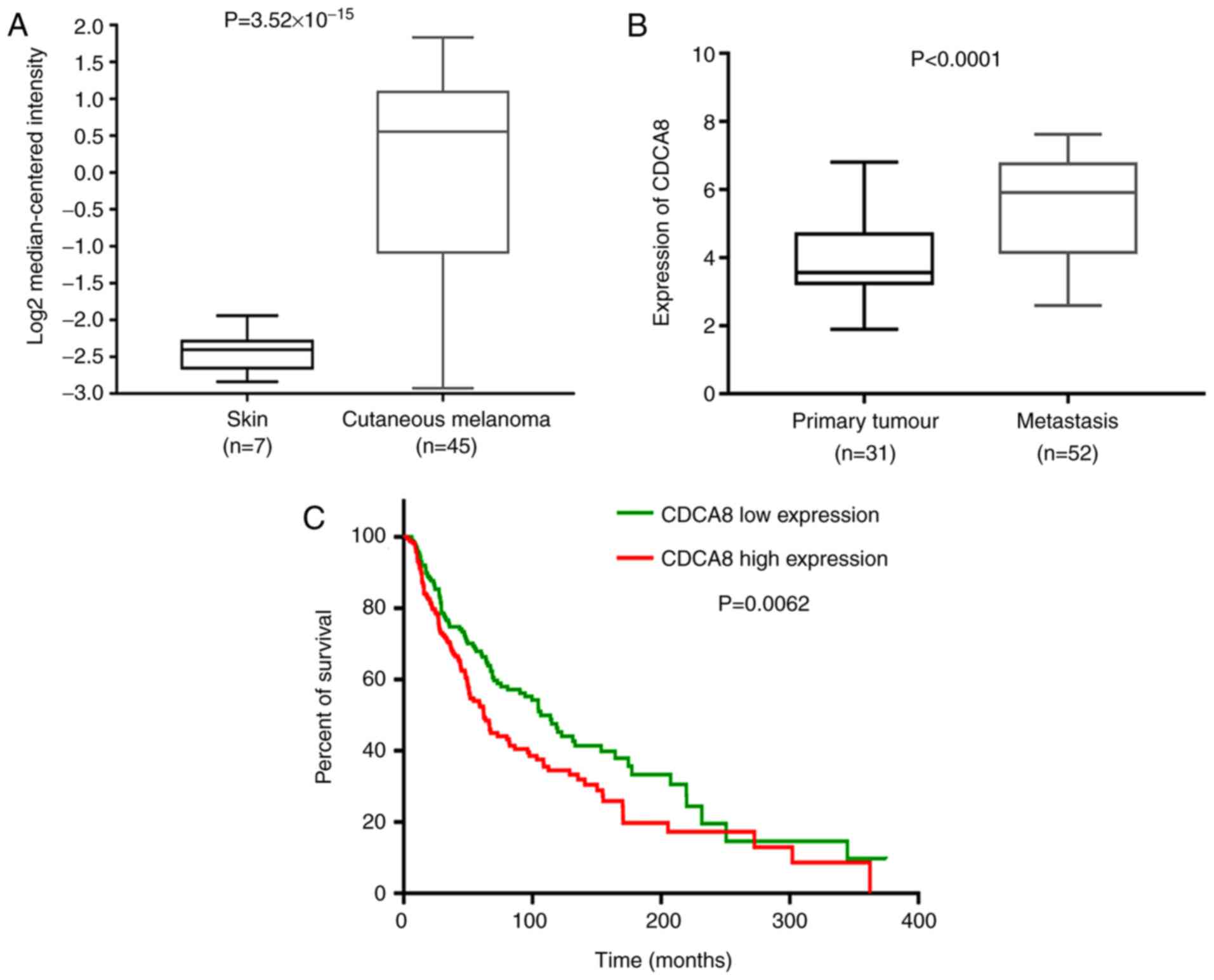
Firstly, based on the ONCOMINE and GEO data, CDCA8
expression levels were demonstrated to be significantly increased
in cutaneous melanoma tissues compared with normal skin tissues
(Fig. 1A). In addition, CDCA8
levels in metastatic melanoma tumors were significantly higher
compared with primary melanoma tumors (Fig. 1B).
Correlation of CDCA8 expression and
clinical characteristics of patients with cutaneous melanoma
In order to further explore the impact of CDCA8 on
cutaneous melanoma progression, RNA-Sequencing data of CDCA8
expression from TCGA database were downloaded, and the correlation
was analyzed using Perl language package and Chi-square test. As
presented in Table I, the
expression levels of CDCA8 were significantly associated with lymph
node metastasis in cutaneous melanoma patients (P=0.039). By
contrast, age, sex, clinical-stage, primary tumor and metastasis
stage were not relevant to CDCA8 expression (all P>0.05). These
results indicated that CDCA8 was associated with the progression of
cutaneous melanoma.
 | Table IClinicopathological parameters of
patients with cutaneous melanoma in relation to CDCA8
expression. |
Table I
Clinicopathological parameters of
patients with cutaneous melanoma in relation to CDCA8
expression.
| Characteristic | Expression of CDCA8
| P-value |
|---|
| Low | High |
|---|
| Age, years | | | 0.112 |
| <60 | 128 | 111 | |
| ≥60 | 101 | 118 | |
| Sex | | | 0.178 |
| Female | 94 | 80 | |
| Male | 135 | 149 | |
| Clinical stage | | | 0.091 |
| I+II | 99 | 116 | |
| III+IV | 104 | 87 | |
| Pathologic-T | | | 0.849 |
| T1+T2 | 72 | 68 | |
| T3+T4 | 121 | 119 | |
| Pathologic-M | | | 0.264 |
| M0 | 200 | 209 | |
| M1 | 14 | 9 | |
| Pathologic-N | | | 0.039a |
| N0 | 102 | 126 | |
| N1 | 97 | 79 | |
High CDCA8 expression is associated with
poor prognosis in cutaneous melanoma patients
Next, the correlation between the prognosis of
cutaneous melanoma patients and CDCA8 expression levels was
explored using Kaplan-Meier analysis and Log-rank test. As
illustrated in Fig. 1C, the
survival rates of the CDCA8 high expression group and the low
expression group were significantly different (P=0.0062). The
overall survival time of patients with high CDCA8 expression was
significantly shorter, suggesting that CDCA8 expression might be a
prognostic marker in cutaneous melanoma patients.
In order to further confirm these results, cox
proportional hazards analysis was performed (Table II). Significant univariate
associations of survival were observed for CDCA8 expression,
clinical stage, pathologic-T, pathologic-N stage and age (all
P<0.01; Table II).
Multivariate analysis further identified two of these factors to be
significant negative predictors of survival: CDCA8 expression
[hazard ratio (HR)=1.615, P=0.003)] and pathologic-T stage
(HR=1.923, P=0.000). These results demonstrated that CDCA8 was an
independent predictor for prognosis of cutaneous melanoma
patients.
| Table IIPredictors of cutaneous
melanoma-specific survival in patients. |
Table II
Predictors of cutaneous
melanoma-specific survival in patients.
| Variable | Univariate analysis
| Multivariate
analysis
|
|---|
| P-value | HR | 95% CI | P-value | HR | 95% CI |
|---|
| CDCA8 expression
(high/low) | 0.006a | 1.462 | 1.112-1.922 | 0.003a | 1.615 | 1.173-2.223 |
| Clinical stage
(I+II/III+IV) | 0.002a | 1.607 | 1.187-2.175 | 0.657 | 0.795 | 0.289-2.186 |
| Pathologic-T
(T1+T2/T3+T4) | <0.001a | 1.967 | 1.429-2.707 | 0.000a | 1.923 | 1.364-2.710 |
| Pathologic-M
(M0/M1) | 0.161 | 1.617 | 0.826-3.169 | | | |
| Pathologic-N
(N0/N1+N2+N3) | <0.001a | 1.739 | 1.284-2.356 | 0.092 | 2.395 | 0.867-6.611 |
| Age, years
(60/≥60) | 0.001a | 1.650 | 1.244-2.189 | 0.543 | 1.109 | 0.795-1.547 |
| Sex
(female/male) | 0.240 | 1.191 | 0.890-1.596 | | | |
CDCA8 is overexpressed in cutaneous
melanoma cells
To characterize the prognosis-associated CDCA8
activity in vitro, the expression levels of CDCA8 were
examined in cutaneous melanoma cells A375 and MV3, and in the
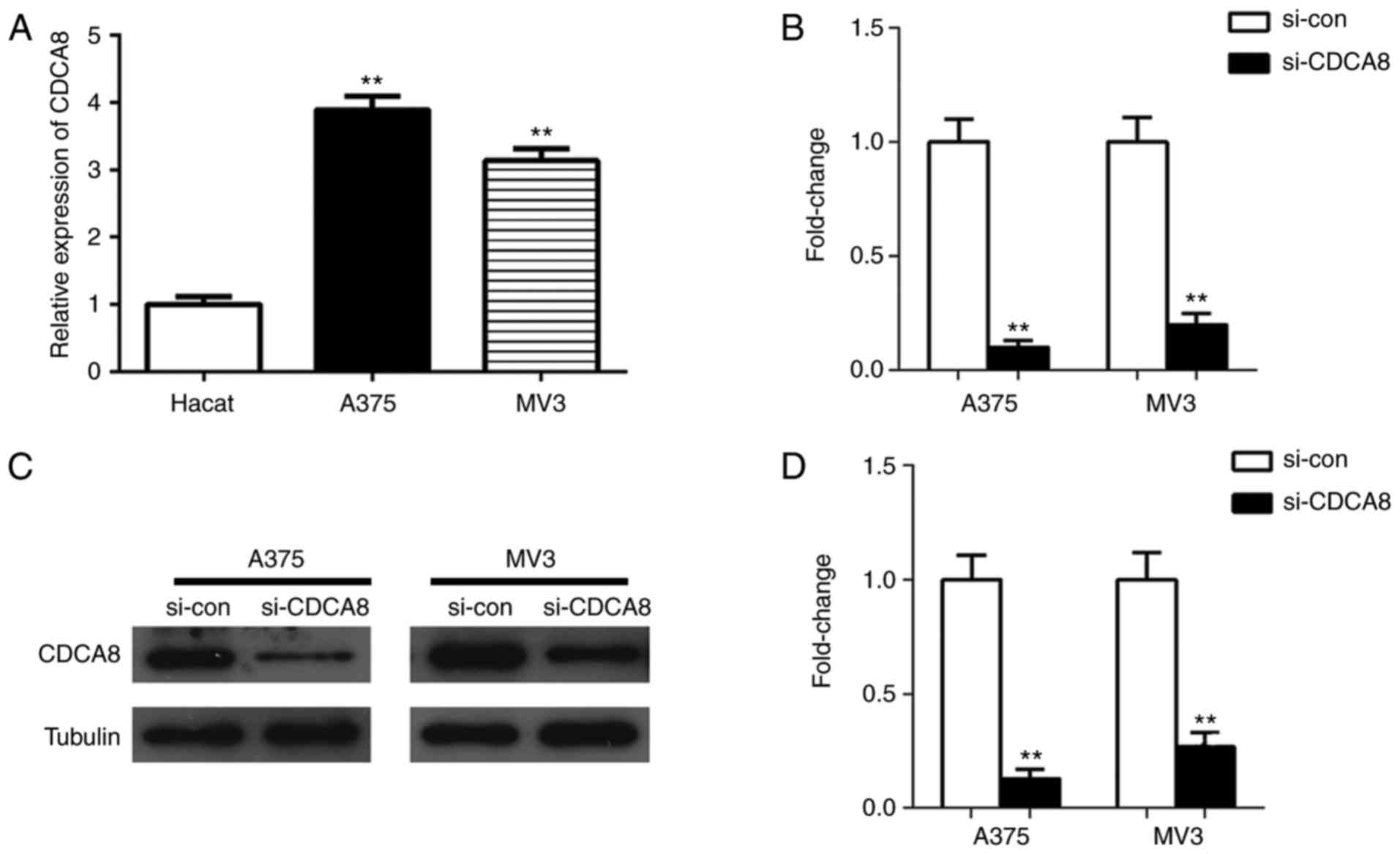
normal Hacat cells, by RT-qPCR analysis. As illustrated in Fig. 2A, CDCA8 expression was
significantly increased in A375 and MV3 cells compared with Hacat
cells (both P<0.01). These results suggested that CDCA8 was
upregulated in cutaneous melanoma cells.
CDCA8 knockdown in A375 and MV3
cells
Subsequently, a knockout vector carrying siRNA
targeting CDCA8 (si-CDCA8) was transfected into A375 and MV3 cells.
A non-targeting siRNA sequence (si-con) was used as a control.
Total RNA and total protein was extracted from the cells at 24 h
post-transfection and examined by RT-qPCR and western blot
analysis, respectively. The results demonstrated that si-CDCA8
could significantly reduce the mRNA and protein expression levels
of CDCA8 in A375 and MV3 cells, and the knockdown efficiency was
>70% (Fig. 2B-D; all
P<0.01).
CDCA8 knockdown results in decreased
proliferation in cutaneous melanoma cells
The function of CDCA8 was explored next in cutaneous
melanoma cells. Firstly, cell proliferation activity was examined
following CDCA8 knockdown in A375 and MV3 cells using CCK-8 and
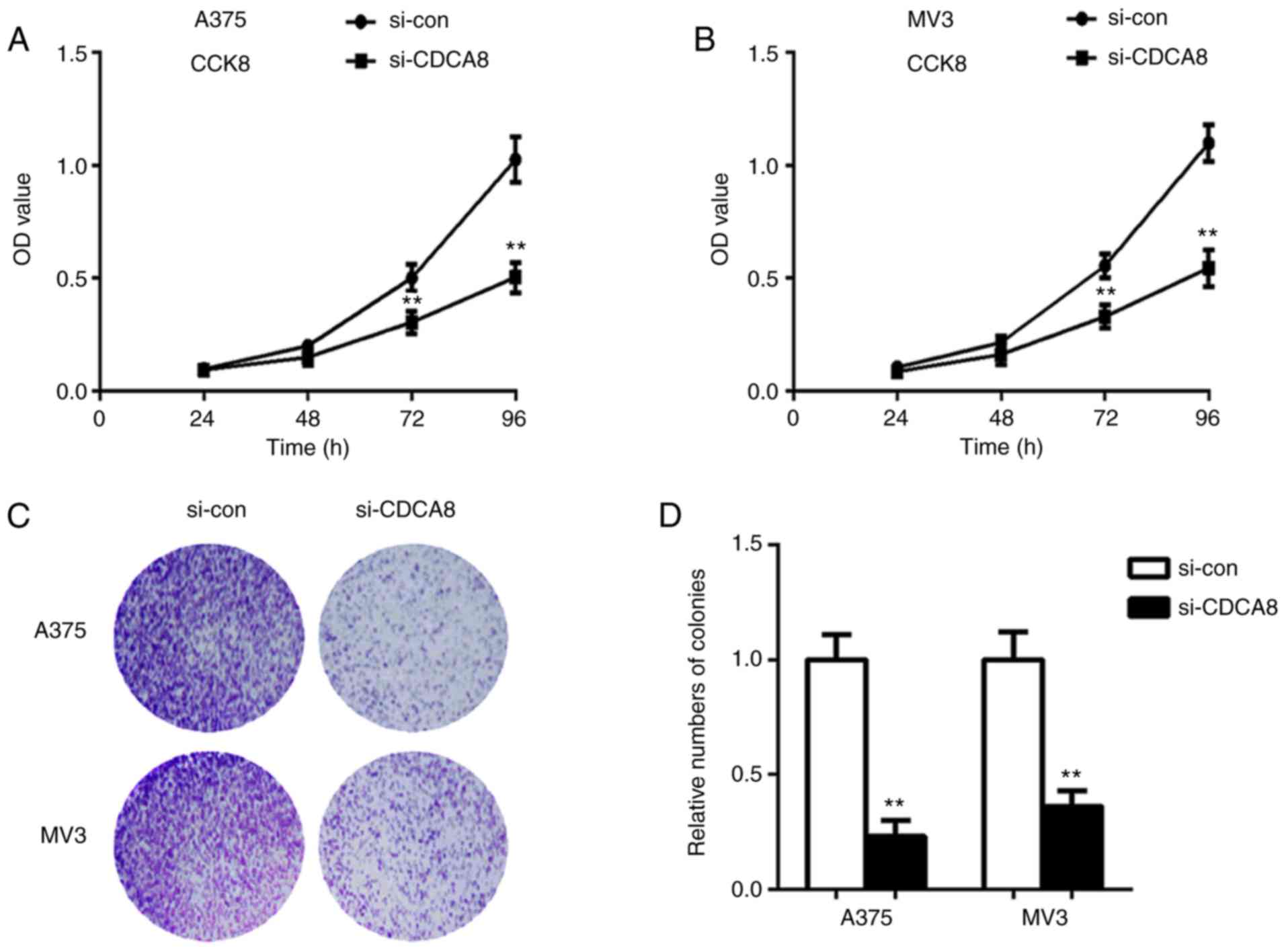
colony formation assays. In A375 cells, the OD value in si-CDCA8
group was significantly decreased compared with the si-con group at
72 and 96 h (Fig. 3A; P<0.01).
Similar results were observed in MV3 cells (Fig. 3B; P<0.01). In addition, the
colony formation rate of cells treated with si-CDCA8 was
significantly decreased compared with the control cells, in both
A375 and MV3 cell lines (Fig. 3C and
D; P<0.01). The present results demonstrated that knockdown
of CDCA8 suppressed the proliferation of cutaneous melanoma
cells.
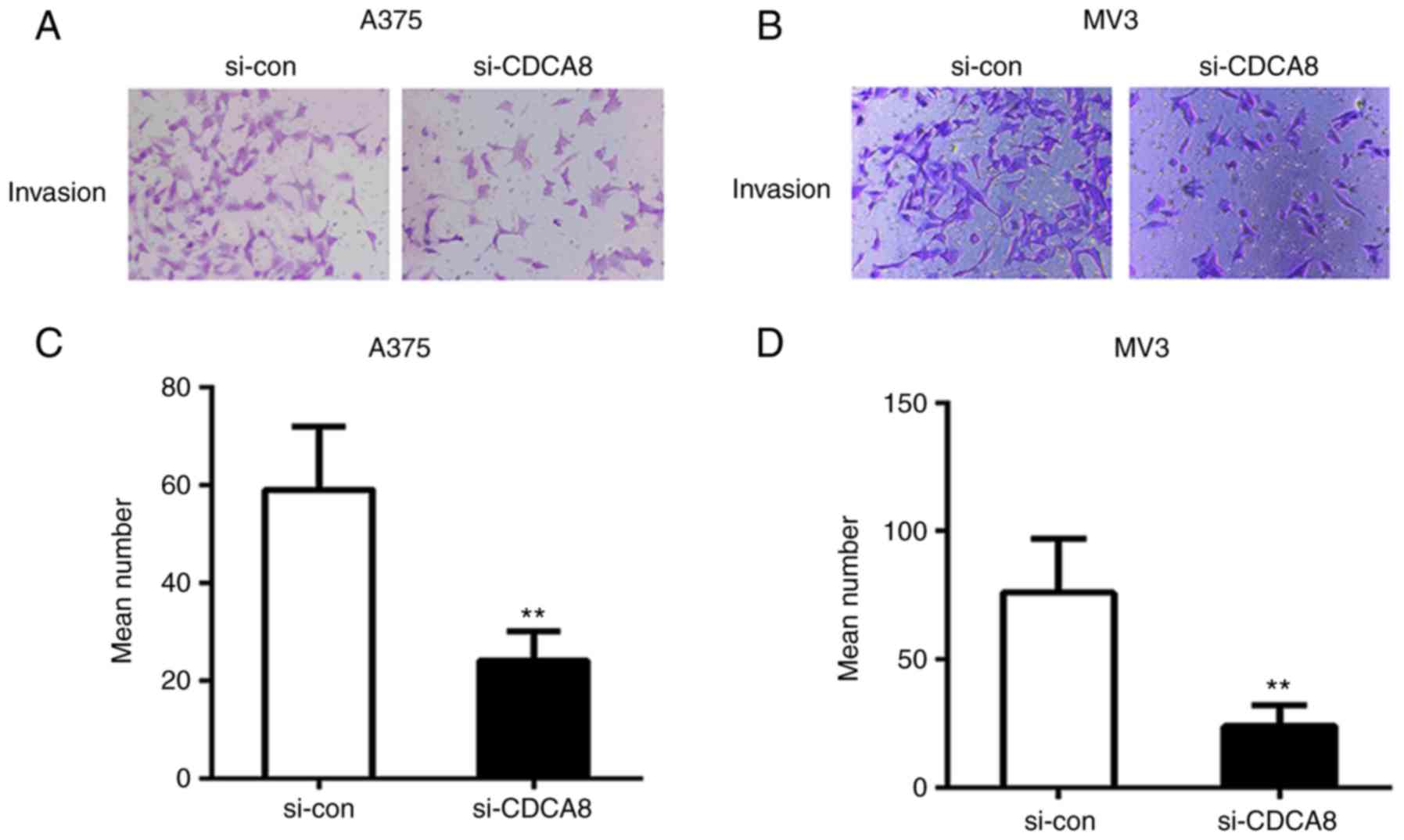
CDCA8 knockdown inhibits cutaneous
melanoma cell migration and invasion
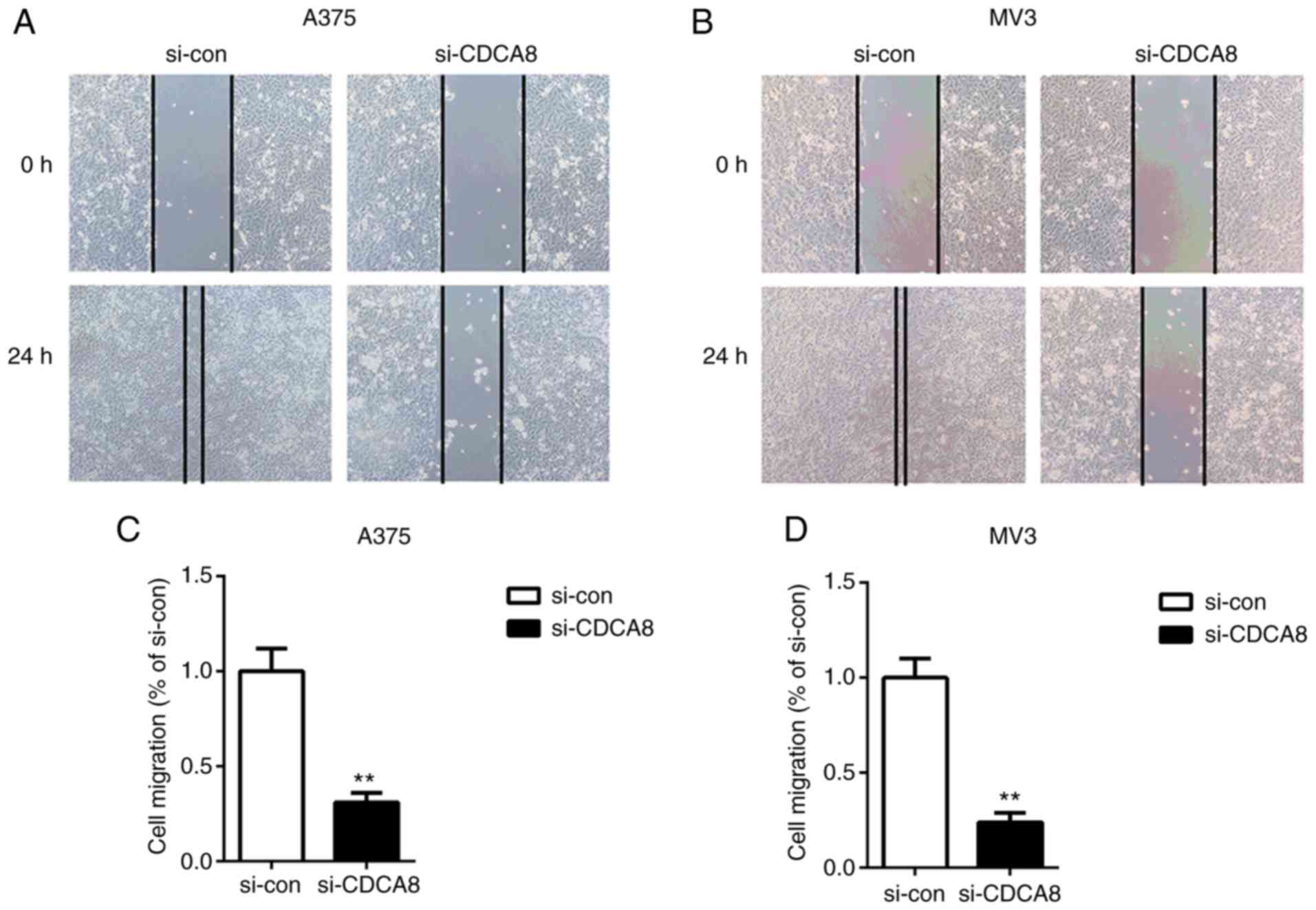
Next, wound healing was performed to examine the
migration speed of A375 and MV3 cells following CDCA8 knockdown. As
illustrated in Fig. 4, cell
migration was markedly decreased in CDCA8-silenced cells compared
with the si-con group (P<0.01). Furthermore, the results from
Matrigel invasion assay demonstrated that the number of invaded
cells in the si-CDCA8 group was significantly reduced compared with
the si-con group (Fig. 5;
P<0.01). These findings indicated that CDCA8 knockdown inhibited
migration and invasion in cutaneous melanoma cells.
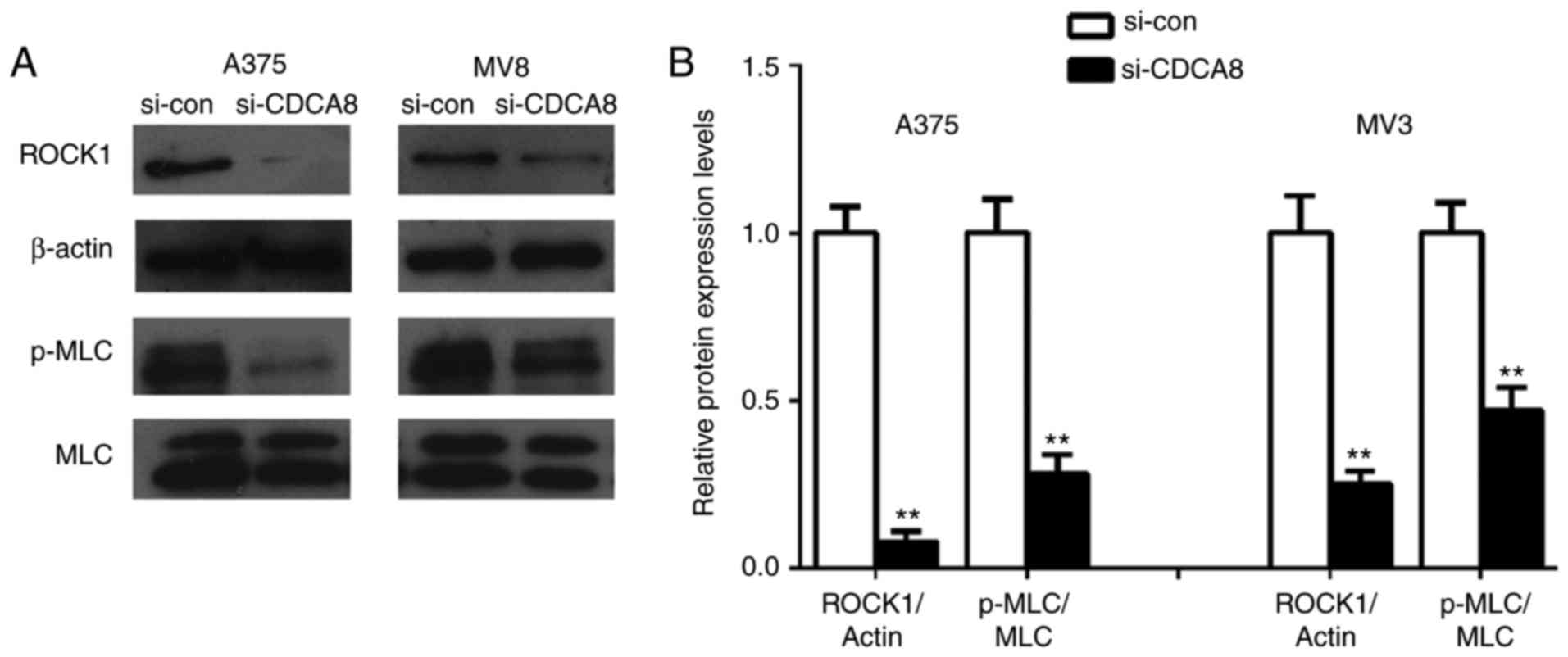
CDCA8 knockdown inhibits the activation
of ROCK signaling in cutaneous melanoma cells
Given that the ROCK signaling pathway has an
important role in cancer and, particularly, in cell motility,
metastasis and angiogenesis, the hypothesis that CDCA8 may regulate
the ROCK pathway was examined in vitro. The western blot
results demonstrated that CDCA8 knockdown in A375 and MV3 cells
reduced the expression levels of ROCK1 and the phosphorylation
levels of MLC, two downstream effector proteins of the ROCK pathway
(Fig. 6; all P<0.01). The
above results indicated that the ROCK signaling pathway might be
involved in cutaneous melanoma cell proliferation and invasion
induced by CDCA8.
Discussion
To our knowledge, there has been no study exploring
the function of CDCA8 in melanoma, including in vitro
studies in cutaneous melanoma cells. In the present study, for the
first time, we identified a close association between cutaneous
melanoma cells and CDCA8 expression. Based on ONCOMINE and GEO
data, CDCA8 expression levels were demonstrated to be overexpressed
in cutaneous melanoma tissues compared with normal tissues, and
high CDCA8 expression was associated with poor prognosis in
cutaneous melanoma patients. The in vitro results
demonstrated that the cutaneous melanoma cell lines A375 and MV3
had increased CDCA8 expression compared with normal cells. Notably,
CDCA8 knockdown inhibited cell proliferation, migration and
invasion in both cutaneous melanoma cell lines. Furthermore, the
regulation of CDCA8 expression and function was strongly associated
with the ROCK pathway.
As a mitotic regulatory gene, the activation of
CDCA8 transcription should conduce to the rapid cell growth
(19). In fact, CDCA8 has been
demonstrated to be indispensable for the growth of lung cancer
cells, which was significantly inhibited by siRNA against CDCA8
(7,19,20). In addition, proliferation of human
embryonic stem cells (hESCs) was also reduced by CDCA8 knockdown
(21). These studies are
consistent with the present findings that CDCA8 knockdown inhibited
the A375 and MV3 cell proliferation, migration and invasion.
Furthermore, the present analysis revealed that the expression of
CDCA8 was significantly associated with lymph node metastasis.
Regional lymph node metastases constitute the most common mode of
initial presentation with metastatic melanoma (22-24). The updated 2009 AJCC melanoma
staging system reported that, in the absence of nodal metastases,
patients with intralymphatic metastases have a 5-year survival rate
of only 69% (25), which may be
one of the reasons CDCA8 affects the prognosis of cutaneous
melanoma patients. In addition to being associated with poor
prognosis in cutaneous melanoma patients, CDCA8 was also an
independent prognostic factor, similar to the role of CDCA8
observed previously in breast cancer (12). These results indicate that CDCA8
has a crucial role in the progression of cutaneous melanoma.
In the present study, CDCA8 knockdown inhibited ROCK
signaling in cutaneous melanoma cells. The ROCK signaling pathway
is associated with cell proliferation and differentiation,
apoptosis, cell cycle, cell polarity, the cytoskeleton and
vasoconstriction (13,14,26). The upstream protein Rho GTPase
exists in all eukaryotic organisms and has roles in cell migration,
movement, proliferation and differentiation (27). Activated RhoA binds directly to
the C-terminus of ROCK and activates it. Activated ROCK
phosphorylates myosin and its regulatory proteins to regulate
changes of and contract the cytoskeleton (28). Therefore, ROCK signaling is
important for cytoskeleton reorganization, cell migration,
movement, contraction and proliferation. ROCK1, one of the two ROCK
isoforms, is a major downstream effector of the small GTPase RhoA
(29,30). ROCK1 has a role in cancer,
especially cell motility, metastasis, and angiogenesis (27,31,32). Furthermore, ROCK directly
phosphorylates MLC, conducing to the actin-myosin force generation
that is required for membrane blebbing (15), cell contraction (26,33) and the formation of apoptotic
bodies (29). The present results
suggested that CDCA8 knockdown reduced the expression levels of
ROCK1 and phosphorylated MLC in A375 and MV3 cells. Taken together,
these findings suggest that CDCA8 knockdown inhibited cutaneous
melanoma cell proliferation and invasion potentially via the ROCK
signaling pathway.
In summary, the present study used bioinformatics
analysis and in vitro cell experiments to demonstrate that
CDCA8 is a facilitator of the malignant progression of cutaneous
melanoma cells via the ROCK pathway. Therefore, CDCA8 might be a
promising prognostic factor and a potential therapeutic target for
cutaneous melanoma patients. However, there are several limitations
in this study. CDCA8 was suggested to be phosphorylated in
vitro by aurora kinase B (AURKB), and co-transactivation of
CDCA8 and AURKB was observed in several cancers. Consequently, the
AURKB expression in cutaneous melanoma samples needs to be
evaluated in synergy with CDCA8 in vitro. In addition, a
decrease in ROCK1 expression may cause an increase in
caspase-3/cleaved-caspase-8, which is important in cell apoptosis.
Further studies to investigate the cell apoptosis following CDCA8
knockdown are necessary. Finally, further investigations with
overexpression of CDCA8 and/or CDCA8 inhibitors are warranted in
subsequent studies.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors’ contributions
CC, BT and DL conceived and designed the project. DL
and WL provided administrative support. DQ, XJ, XQ collected data.
XQ, LC and WD analyzed and interpreted the results. All authors
contributed to writing, and all authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest
References
|
1
|
Matthews NH, Li WQ, Qureshi AA, Weinstock
MA and Cho E: Epidemiology of melanoma. Environmental and molecular
mutagenesis. 2017.
|
|
2
|
Goel N, Ward WH, Yu JQ and Farma JM:
Short-term and long-term management of melanoma. Cutaneous
melanoma: Etiology and therapy. Ward WH and Farma JM: Codon
Publications; Brisbane (AU): 2017, View Article : Google Scholar
|
|
3
|
Joyce KM: Surgical management of melanoma.
Cutaneous Melanoma: Etiology and therapy. Ward WH and Farma JM:
Codon Publications; Brisbane (AU): 2017, View Article : Google Scholar
|
|
4
|
Norval M and Wright CY: The Epidemiology
of cutaneous melanoma in the white and black African population
groups in South Africa. Cutaneous melanoma: Etiology and therapy.
Ward WH and Farma JM: Codon Publications; Brisbane (AU): 2017,
View Article : Google Scholar
|
|
5
|
Rausch MP and Hastings KT: Immune
checkpoint inhibitors in the treatment of melanoma: From basic
science to clinical application. Cutaneous melanoma: Etiology and
therapy. Ward WH and Farma JM: Codon Publications; Brisbane (AU):
2017, View Article : Google Scholar
|
|
6
|
G Lben K, Berberoglu U, Altinyollar H,
Kinas V and Turanli S: Sentinel lymph node status affects long-term
survival in patients with intermediate-thickness melanoma. J Cancer
Res Ther. 12:840–844. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Phan NN, Wang CY, Li KL, Chen CF, Chiao
CC, Yu HG, Huang PL and Lin YC: Distinct expression of CDCA3,
CDCA5, and CDCA8 leads to shorter relapse free survival in breast
cancer patient. Oncotarget. 9:6977–6992. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gu Y, Lu L, Wu L, Chen H, Zhu W and He Y:
Identification of prognostic genes in kidney renal clear cell
carcinoma by RNAseq data analysis. Mol Med Rep. 15:1661–1667. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yan H, Li Z, Shen Q, Wang Q, Tian J, Jiang
Q and Gao L: Aberrant expression of cell cycle and material
metabolism related genes contributes to hepatocellular carcinoma
occurrence. Pathol Res Pract. 213:316–321. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li S, Liu X, Liu T, Meng X, Yin X, Fang C,
Huang D, Cao Y, Weng H, Zeng X and Wang X: Identification of
biomarkers correlated with the TNM staging and overall survival of
patients with bladder cancer. Front Physiol. 8:9472017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun L, Li J and Yan B: Gene expression
profiling analysis of osteosarcoma cell lines. Mol Med Rep.
12:4266–4272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiao DC, Lu ZD, Qiao JH, Yan M, Cui SD and
Liu ZZ: Expression of CDCA8 correlates closely with FOXM1 in breast
cancer: Public microarray data analysis and immunohistochemical
study. Neoplasma. 62:464–469. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Çarçak N, Yavuz M, Eryiğit Karamahmutoğlu
T, Kurt AH, Urhan Küçük M, Onat FY and Büyükafsar K: Suppressive
effect of Rho-kinase inhibitors Y-27632 and fasudil on
spike-and-wave discharges in genetic absence epilepsy rats from
Strasbourg (GAERS). Naunyn-Schmiedebergs Arch Pharmacol.
391:1275–1283. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cho SY, Sung CO, Chae J, Lee J, Na D, Kang
W, Kang J, Min S, Lee A, Kwak E, et al: Alterations in the Rho
pathway contribute to Epstein-Barr virus-induced lymphomagenesis in
immunosup-pressed environments. Blood. 131:1931–1941. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dos Santos AA, López-Granero C, Farina M,
Rocha JBT, Bowman AB and Aschner M: Oxidative stress, caspase-3
activation and cleavage of ROCK-1 play an essential role in
MeHg-induced cell death in primary astroglial cells. Food Chem
Toxicol. 113:328–336. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Honjo M and Tanihara H: Impact of the
clinical use of ROCK inhibitor on the pathogenesis and treatment of
glaucoma. Jap J Ophthalmol. 62:109–126. 2018. View Article : Google Scholar
|
|
17
|
Li N, Chen J, Zhao J and Wang T:
MicroRNA-3188 targets ETS-domain protein 4 and participates in
RhoA/ROCK pathway to regulate the development of atherosclerosis.
Pharmazie. 72:687–693. 2017.
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Dai C, Miao CX, Xu XM, Liu LJ, Gu YF, Zhou
D, Chen LS, Lin G and Lu GX: Transcriptional activation of human
CDCA8 gene regulated by transcription factor NF-Y in embryonic stem
cells and cancer cells. J Biol Chem. 290:22423–22434. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Papini D, Langemeyer L, Abad MA, Kerr A,
Samejima I, Eyers PA, Jeyaprakash AA, Higgins JM, Barr FA and
Earnshaw WC: TD-60 links RalA GTPase function to the CPC in
mitosis. Nat Commun. 6:76782015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Marko NF, Weil RJ, Schroeder JL, Lang FF,
Suki D and Sawaya RE: Extent of resection of glioblastoma
revisited: Personalized survival modeling facilitates more accurate
survival prediction and supports a maximum-safe-resection approach
to surgery. J Clin Oncol Mar. 32:774–82. 2014. View Article : Google Scholar
|
|
22
|
Huang SK and Hoon DS: Liquid biopsy
utility for the surveillance of cutaneous malignant melanoma
patients. Mol Oncol. 10:450–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pan Y, Haydon AM, McLean CA, McDonald PB
and Kelly JW: Prognosis associated with cutaneous melanoma
metastases. Australas J Dermatol. 56:25–28. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jaber DF, Jallad MN and Abdelnoor AM: The
effect of ciprofloxacin on the growth of B16F10 melanoma cells. J
Cancer Res Ther. 13:956–960. 2017.PubMed/NCBI
|
|
25
|
Weiss SA, Hanniford D, Hernando E and
Osman I: Revisiting determinants of prognosis in cutanous melanoma.
Cancer. 121:4108–4123. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCarthy CG, Wenceslau CF, Ogbi S, Szasz T
and Webb RC: Toll-like receptor 9-dependent AMPKalpha activation
occurs via TAK1 and contributes to RhoA/ROCK signaling and actin
polymerization in vascular smooth muscle cells. J Pharmacol Exp
Ther. 365:60–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takaoka M, Ito S, Miki Y and Nakanishi A:
FKBP51 regulates cell motility and invasion via RhoA signaling.
Cancer Sci. 108:380–389. 2017. View Article : Google Scholar :
|
|
28
|
Pich C, Teiti I, Sarrabayrouse G, Gallardo
F, Gence R and Tilkin-Mariame AF: Melanoma expressed-CD70 is
regulated by RhoA and MAPK pathways without affecting vemurafenib
treatment activity. PLoS One. 11:e01480952016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sahin L, Cevik OS, Koyuncu DD and
Buyukafsar K: Role of rho-kinase (ROCK) in tonic but not phasic
contraction in the frog stomach smooth muscle. Life Sci. 198:46–55.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zeng Y, Xie H, Qiao Y, Wang J, Zhu X, He
G, Li Y, Ren X, Wang F, Liang L and Ding Y: Formin-like2 regulates
Rho/ROCK pathway to promote actin assembly and cell invasion of
colorectal cancer. Cancer Sci. 106:1385–1393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen K, Wang Y, Zhang Y, Zhou H, Song Y,
Cao Z, Kou J and Yu B: Cocktail of four active components derived
from Sheng Mai San inhibits hydrogen peroxide-induced PC12 cell
apoptosis linked with the caspase-3/ROCK1/MLC pathway. Rejuvenation
Res. 18:517–527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Voorneveld PW, Kodach LL, Jacobs RJ, Liv
N, Zonnevylle AC, Hoogenboom JP, Biemond I, Verspaget HW, Hommes
DW, de Rooij K, et al: Loss of SMAD4 alters BMP signaling to
promote colorectal cancer cell metastasis via activation of Rho and
ROCK. Gastroenterology. 147:196–208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mu D, Zhou G, Li J, Su B and Guo H:
Ursolic acid activates the apoptosis of prostate cancer via
ROCK/PTEN mediated mitochondrial translocation of cofilin-1. Oncol
Lett. 15:3202–3206. 2018.PubMed/NCBI
|