Introduction
Polycystic ovarian syndrome (PCOS), which has been
renamed metabolic reproductive syndrome, is the most common but
little-known reproductive endocrine dysfunction and metabolic
disorder in women worldwide (1,2).
It is characterized by hyperandrogenism, ovarian polycystic changes
and rarity in ovulation or anovulation, and is often accompanied
with obesity and insulin resistance (3-5).
Diabetes, cardiovascular disease and cancer are its long-term
complications. The prevalence of PCOS in Chinese women aged between
12 and 44 years was reported to be 7.4%, according to the
recommended diagnostic criteria for PCOS by the Androgen Excess
Society (2006), and there is currently a gradual upward trend
(6).
FK-506 binding protein 52 (FKBP52) is a subfamily of
the FK506 binding protein family, and is commonly used as
immunosuppressant. In 1985, FKBP52 was first identified by Tai
et al in the establishment of an antibody against the EC1
epitope of rabbit uterine progesterone receptor complex (7). It was named due to its relative
molecular weight of ~52 kDa, and to date, it has been investigated
in various experimental studies (8). As one of the Hsp90 co-chaperones
that modify steroid hormone receptor activity, including regulation
of receptor maturation, hormone binding and nuclear translocation,
FKBP52 is a positive regulator of androgen receptor (AR) (9). It has been reported to be
overexpressed in prostate cancer cell lines, and FK506 has an
inhibitory influence on androgen-stimulated cell growth (10). Prostate needle biopsies of human
patients were also found to have elevated FKBP52 levels (11). Furthermore, a number of compounds
that suppress the adjustment by FKBP52 on AR function interdicted
androgen-dependent gene expression and cell proliferation in
prostate cancer cells (12).
Preceding studies have also reported that male 52KO mice manifested
phenotypes in accordance with partial androgen insensitivity
(9,13).
There have been few reports on FKBP52 in PCOS. In a
previous study, by comparing normal women of childbearing age with
patients with PCOS using gene chip technology, it was revealed that
FKBP52 protein-coding genes were different, and this may be closely
associated with abnormal androgen PCOS and obesity phenotype
(14). In conclusion, FKBP52 may
be considered a therapeutic target in diseases that rely on AR
signaling pathways, and may be a contributor to endocrine-related
and metabolic diseases, including PCOS.
The mitogen-activated protein kinase
(MAPK)/extracellular signal-regulated kinase (ERK) pathway is a
classic MAPK signal transduction pathway. It is involved in cell
growth, differentiation, environmental stress adaptation, the
inflammatory response and other important cell physiology/pathology
processes. Activated AR in the cytoplasm may interact with several
signaling molecules, which in turn converge on the activation of
MAPK/ERK (15,16). Studies on 5α-dihydrotestost
eroneresponsiveness in prostate cancer cells showed increased
phosphorylation of ERK-1/2 within 5 min, in a dose-dependent
manner, and this response was AR-dependent as no effect was
observed in AR-negative PC-3 prostate cancer cells (17). PCOS with insulin resistance and
PCOS without insulin resistance showed statistically significant
increases than control group (P<0.05) in expression of ERK1/2 in
human luteinized granulosa cells (GCs); there was no significant
difference between the PCOS groups (18).
The present study aimed to investigate the
expression of FKBP52 in the ovarian tissues of PCOS rats. It is
well-known that FKBP4 is the FKBP52-encoding gene. The adenovirus
vectors Ad-Oe-FKBP4-EGFP and Ad-siRNA-FKBP4-EGFP were constructed
to assess how FKBP52 mediates AR through the MAPK/ERK pathway
following GC transfection.
Materials and methods
Animal experiments Experimental
animals
A total of 60 female Sprague-Dawley (SD) rats were
provided by Shanghai Jie Esprit Experimental Animal Co., Ltd.
[Shanghai, China; certificate no. SCXK (Shanghai) 2013-0006]. The
animals were of specific-pathogen-free (SPF) grade and were 21 days
old, with a mean body weight of 58±4 g.
Replication of a PCOS animal model
The 60 21-day-old female SD rats of SPF grade were
randomly divided into three groups following common feeding for 2
days; In the PCOS model (PM) group (n=20), the rats were
administered with dehydroepiandrosterone (DHEA; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at 6 mg/(100 g. d) + 0.2 ml of
injectable soybean oil via hypodermic injection for 35 consecutive
days (19,20); in the oil control (OC) group
(n=20), the rats were administered with 0.2 ml of injectable
soybean oil via hypodermic injection over the same period; in the
normal control (NC) group (n=20), the rats underwent no
specific treatment. All animals were housed with a normal diet
every day under a 12 h light/dark cycle at a temperature of
20-25°C.
The model was successfully established when rats of
the PM group showed loss of their estrous cycle. All rats were
detected for body weight, ovarian weight, organ coefficient and
vaginal smear, and serum was collected and analyzed via an
enzyme-linked immunosorbent assay (ELISA). In addition, four
ovaries of each group were randomly selected for histological
examination of ovarian tissues to further evaluate the efficiency
of the established model.
Specimen collection
Following the final day of modeling, all rats were
weighed and then anesthetized with 2% sodium pentobarbital (30
mg/kg) for laparotomy to collect abdominal aorta blood and ovarian
tissue specimens. When the 60 ovarian tissue specimens were
weighed, the four ovarian specimens of each group were rapidly
fixed in 4% paraformaldehyde fixative, embedded with paraffin, and
cut into sections for hematoxylin and eosin (H&E) staining and
immunohistochemistry (IHC). The rest were stored in a refrigerator
at −80°C, for reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis and western blotting (WB). Serum was
collected following centrifugation of all blood samples at 1000 × g
for 10 min.
H&E and IHC staining
Following 4 h of fixation in 4% paraformaldehyde
fixative, the rat ovarian tissue specimens were subjected to
conventional dehydration followed by paraffin-embedding and thin
sectioning (4-µm thick). H&E staining was performed to
examine the pathological structures of the rat ovary. IHC staining
was conducted to detect the expression of FKBP52 in rat ovaries
with an optical microscope (Leica DM2500, Leica Microsystems GmbH,
Wetzlar, Germany).
ELISA
All serum samples were manipulated according to the
manufacturer’s protocol of every ELISA kit (Elabscience, Wuhan,
China) for follicle stimulating hormone (FSH), luteinizing hormone
(LH), estradiol (E2), progesterone (P) and total
testosterone (T). Experiments were performed in triplicate and
repeated three times.
Fluorescence RT-qPCR analysis
A total of eight ovarian tissue specimens from each
group were milled separately with TRIzol reagent (Beyotime
Institute of Biotechnology, Haimen, China) using an IKA T 10 basic
ULTRA-TURRAX disperser. A NanoDrop 2000 spectrophotometer was used
to measure RNA concentration following extraction of total RNA from
every specimen. Subsequently, the RNA was reverse-transcribed into
cDNA using a reverse transcription kit (Takara Bio, Inc., Otsu,
Japan). Eventually, 1.0 µl cDNA was used as a template for
PCR amplification ChamQ™ SYBR® qPCR Master mix (High ROX
Premixed) was purchased from Vazyme (Nanjing, China). The 10
µl qPCR reaction mixture included the following: cDNA 1
µl, forward primer (10 µM) 0.5 µl, reverse
primer (10 µM) 0.5 µl, SYBR qPCR mix 5 µl,
ddH2O 3 µl. The primer sequences (Sangon Biotech
Co., Ltd., Shanghai, China) used were as follows: GAPDH forward,
5′-GACATGCCGCCTGGAGAAAC-3′ and reverse, 5′-AGCCCAGGATGCCCTTTAGT-3′,
FKBP52 upstream, 5′-CACTACACTGGCTGGCTGCT-3′ and downstream,
5′-TGGTTGCCACAGCAATATCC-3′; AR upstream, 5′-CCTTCACAGCAGCAGTCAGC-3′
and downstream, 5′-CCTGATCTGGAGGAGCTGGT-3′. The amplification
conditions were as follows: Pre-denaturation at 95°C for 10 min,
followed by 40 cycles of denaturation at 95°C for 15 sec and
annealing at 60°C for 1 min. Data were collected and used for
relative quantitative analysis with the 2−∆∆Cq method
(21). The relative mRNA
expression level was obtained by comparing data of the experimental
group with those of the control group. The experiments were
performed in triplicate and repeated three times.
WB
A total of eight ovarian tissue specimens from each
group were milled separately with RIPA lysis buffer using IKA T 10
basic ULTRA-TURRAX. The detection of protein concentration was
performed according to the manufacturer’s protocol of the Enhanced
BCA Protein Assay kit (Beyotime Institute of Biotechnology)
following total protein extraction from every specimen. The
electrophoretic separation of known antigenic proteins were run
using the sodium dodecyl sulfate polyacrylamide gel electrophoresis
method; 50 µg protein were separated by 10% SDS-PAGE and
then transferred onto a polyvinylidene fluoride membrane.
Subsequently, the membranes were blocked in 5% bovine serum albumin
(2 h at room temperature) and then incubated with primary (4°C
overnight) and secondary antibodies (2 h at room temperature).
Anti-FKBP52 antibody (EPR6618; cat. no. ab129097; 1:1,000),
anti-androgen receptor antibody (EP670Y; cat. no. ab52615; 1:1,000)
and anti-ERK1 + ERK2 antibody (EPR17526; cat. no. ab184699;
1:10,000) were purchased from Abcam (Cambridge, UK). Phospho-p44/42
MAPK (Erk1/2) (Thr202/Tyr204) (D13.14.4E) XP® rabbit
monoclonal antibody (cat. no. 4370; 1:2,000), GAPDH (14C10) rabbit
monoclonal antibody (cat. no. 2118; 1:1,000) and anti-rabbit IgG
and HRP-linked antibody (cat. no. 7074; 1:50,000) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). The blots
were developed with Immobilon Western Chemiluminescent HRP
Substrate (Merck KGaA). GAPDH served as an internal control.
Quantitative analysis of protein expression was conducted by using
ImageJ software 1.8.0 (National Institutes of Health, Bethesda, MD,
USA).
Cell experiments
Isolation and culture of GCs
A total of 10 female SD rats were provided by
Shanghai Jie Esprit Experimental Animal Co., Ltd. [certificate no.
SCXK (Shanghai) 2013-0006]. The animals were of SPF grade and were
21 days old, with a mean body weight of 58±4 g. All animals were
housed with a normal diet every day. Every rat was administrated
with pregnant mare serum gonadotropin (20 IU; Sansheng
Pharmaceutical Industry Co., Ltd., Ningbo, China) through
intraperitoneal injection following 2 days of common feeding. After
48 h, the animals were anesthetized with 2% sodium pentobarbital
(30 mg/kg), sacrificed by cervical dislocation, and soaked in 75%
alcohol for 30 min. All ovaries, which were obtained by laparotomy,
were placed into PBS and transferred into DMEM-F12 medium (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) following washing once
again with PBS.
The GCs were harvested by puncturing the follicles
with microsurgical forceps, and ~95% of the GCs were negatively
stained using a Trypan blue assay. The GCs were then centrifuged at
300 × g, 4°C for 5 min, filtered with a cell sieve, and centrifuged
again. The pellet was resuspended at 5×105/ml with
DMEM-F12 medium for the RT-qPCR and WB methods, and at
5×104/ml for H&E, immunocytochemistry (ICC), and
immunofluorescence (IF) techniques in 6-well plates. The GCs were
cultured in the incubator (37°C, 5% CO2) and, after 72
h, the cell adherence rate was ~80%.
Morphology and identification of GCs
H&E staining was performed to observe the
morphology of the GCs with an Olympus inverted microscope (Olympus
Corporation, Tokyo, Japan). As FSH receptor (FSHR) is only
expressed in GCs, ICC and IF were conducted to identify its purity.
It met the requirements of subsequent trials when the positive rate
was >95%.
Adenovirus vector transfection of
GCs
The adenovirus vectors Ad-Oe-FKBP4-EGFP
(2×1010 PFU/ml), Oe negative control virus
(1011 PFU/ml), Ad-siRNA-FKBP4-EGFP (2×1010
PFU/ml) and RNAi negative control virus (5×1010 PFU/ml),
provided by Shanghai GeneChem Co., Ltd. (Shanghai, China) were
constructed to transfect GCs. First, the multiplicity of infection
(MOI) was ascertained by using different diluted concentrations of
adenovirus vectors (MOI = virus titer x virus volume/cell numbers).
The adenovirus vectors were diluted according to a concentration
gradient between 10−2 and 10−7. When cell
adherence rate reached ~80% in 96-well plates, the medium was
discarded. The GCs were then cultured with medium containing
different concentrations of virus vectors. After 12 h, the normal
medium was replaced. The expression of green fluorescence in the
GCs was observed following 48 h of infection (Cell transfection
rate = number of green fluorescent cells/total number of cells
×100%). When the rate reached ~80%, the virus concentration was
considered the optimal dilution concentration.
Ad-Oe-FKBP4-EGFP transfection of GCs
The GCs were divided into three groups: Normal
control group (CO group), EGFP-Oe group (EO group), and
FKBP4-EGFP-Oe group (FO group). When the cell attachment rate
reached ~80%, the medium was discarded. Based on previous
pre-experiment results, DMEM-F12 medium was added to the CO group,
DMEM-F12 medium with Oe negative control virus (2 µl) was
added to the EO group, and DMEM-F12 medium with FKBP4-Oe virus (20
µl) was added to the FO group. After 12 h, the medium was
replaced with normal DMEM-F12 medium, and RNA and protein were
obtained following 48 h of infection.
Ad-siRNA-FKBP4-EGFP transfection of
GCs
The GCs were divided into three groups: Normal
control group (CR group), EGFP-RNAi group (ER group), and
FKBP4-EGFP-RNAi group (FR group). When the cell attachment rate
reached ~80%, the medium was discarded. Based on previous
pre-experiment results, DMEM-F12 medium was added to the CR group,
DMEM-F12 medium with RNAi negative control virus (2 µl) was
added to the ER group, and DMEM-F12 medium with FKBP4-RNAi virus
(20 µl) was added to the FR group. After 12 h, the medium
was then replaced with normal DMEM-F12 medium, and RNA and protein
were obtained following 48 h of infection.
Statistical analysis
The statistical analysis was performed using
GraphPad Prism 7 software (GraphPad Software, Inc., La Jolla, CA,
USA). The measurement data are expressed as the mean ± standard
deviation. A normality test and homogeneity of variance test were
performed prior to making comparison between the groups.
Comparisons of three samples were performed using one-way analysis
of variance at the 0.05 level and multiple comparison between the
groups was performed using Tukey method. Variables that did not
meet a normal distribution were analyzed using a Kruskal-Wallis
test.
Results
PCOS rats show loss of regular estrous
cycle
At 35 days following DHEA treatment, the rats in the
PM group had lost their regular estrous cycles and all of remained
in the estrus period. Microscopy of stained smears of vaginal
secretions showed the presence of large numbers of keratinized
cells, suggesting anovulation. By contrast, the rats in the NC and
OC group had a regular estrous cycle (Fig. 1A).
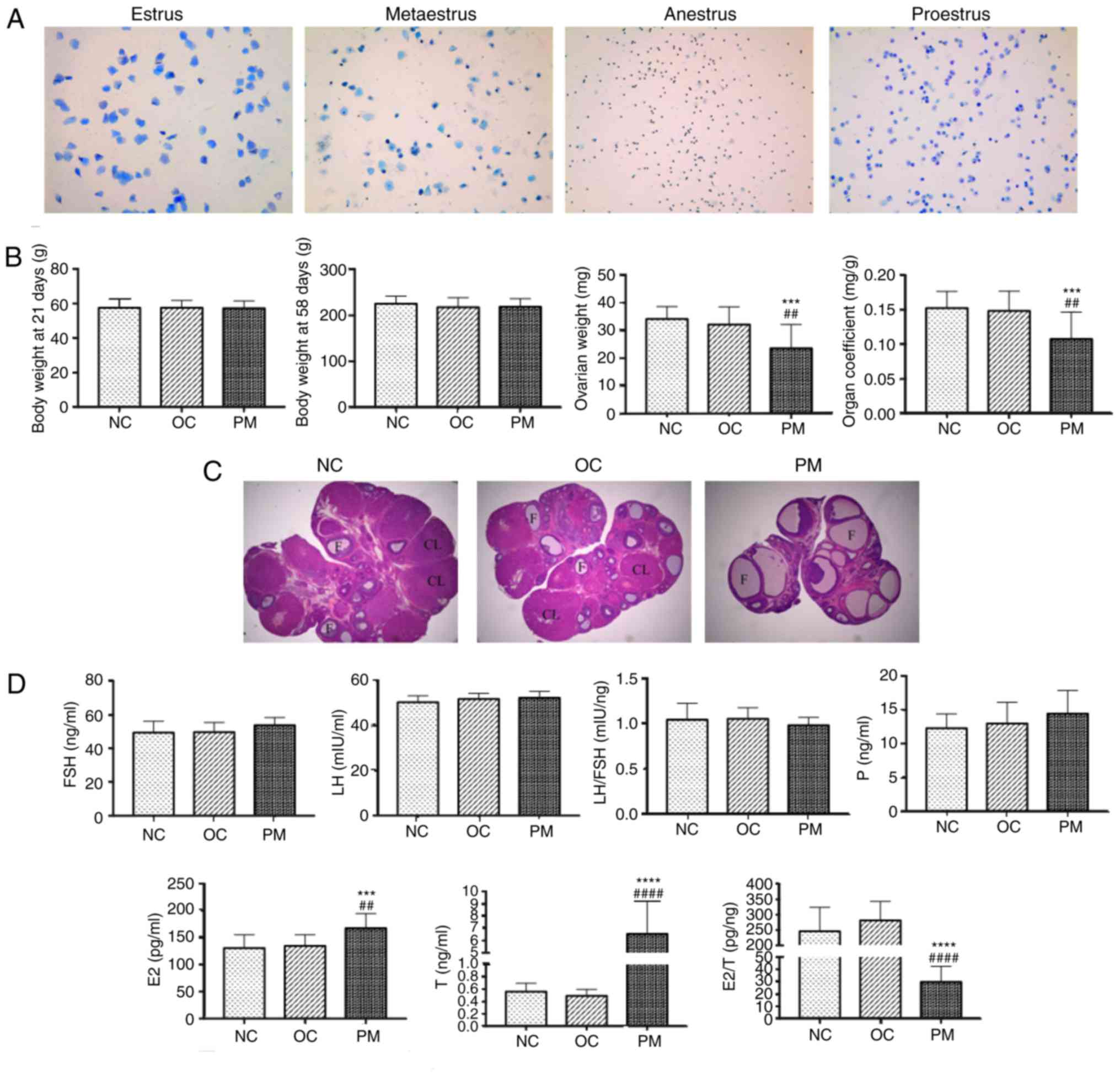 | Figure 1PCOS model evaluation. (A) Microscopy
of stained smears of vaginal secretions (toluidine blue staining;
magnification, ×100; n=20 per group). Estrus (keratinized
epithelial cells); metaestrus (keratinized epithelial cells,
epithelial cells, and white blood cells); anestrus (white blood
cells); proestrus (epithelial cells). NC and OC groups had regular
estrous cycle, however, rats in the PM group remained in the estrus
period and lost their regular estrous cycles, suggesting
anovulation. (B) Comparison of body weight, ovarian weight and
organ coefficient among the three groups (mean ± SD, n=20 per
group). Body weights at 21 and 58 days did not differ significantly
among groups, whereas PM group ovarian weight and organ coefficient
were significantly lower. (C) Comparison of rat ovarian structure
(hematoxylin and eosin staining, ×25 magnification, n=4 per group).
Morphological changes of rat ovarian tissue specimens were examined
by light microscopy. In the NC and OC groups, microscopic
examination revealed the presence of follicles of different
developmental stages and a few corpora lutea; granulosa cells were
orderly arranged with an intact form, mostly in 6-8 layers. In the
PM group, the number of follicles with saccular dilatation
increased, whereas follicles of different developmental stages and
corpora lutea were rare; granulosa cells were arranged loosely in
~2-3 layers, with atresia of some follicles. This result was
consistent with PCOS characteristics. (D) Comparison of FSH, LH,
LH/FSH, P, E2, T and E2/T among groups (mean
± SD, n=20 per group). Sex hormones were measured by enzyme-linked
immunosorbent assay. No significant differences in FSH, LH or
LH/FSH were found. However, E2 and T in the PM group
were significantly higher, and E2/T was significantly
lower. Experiments were performed in triplicate and repeated three
times. ***P<0.0001 and ****P<0.0001 NC
group vs. PM group; ##P<0.01 and
####P<0.0001 OC group vs. PM group. PCOS, polycystic
ovarian syndrome; PM, PCOS model; NC, normal control; OC, oil
control; FSH, follicle stimulating hormone; LH, luteinizing
hormone; E2, estradiol; P, progesterone; T, total
testosterone; F, follicle; CL, corpora lutea; SD, standard
deviation. |
Ovarian weight and organ coefficient are
decreased in PCOS rats
The body weights of the rats at 21 and 58 days among
the three groups showed no statistically significant difference.
However, the ovarian weight and organ coefficient of the PM group
were significantly lower than those in the other two groups
(Fig. 1B).
Pathological structures of rat
ovaries
Morphological changes in the rat ovarian tissue
specimens were examined by light microscopy. In the NC and OC
groups, microscopic examination revealed the presence of follicles
of different developmental stages and a few corpora lutea;
arrangement of the GCs was orderly with an intact form, mostly in
6-8 layers. In the PM group, the number of follicles with saccular
dilatation increased whereas few follicles of different
developmental stages and corpora lutea were observed; the GCs were
arranged loosely in ~2-3 layers, and with atresia of certain
follicles. This result was consistent with PCOS characteristics
(Fig. 1C).
PCOS rats have higher E2 and T
levels and lower E2/T levels
There were no statistically significant differences
in the levels of FSH, LH or LH/FSH in the rats among the three
groups. However, the levels of E2 and T in the PM group
were significantly higher than those in the other two groups, and
E2/T was significantly lower. The experiments were
performed in triplicate and repeated three times (Fig. 1D).
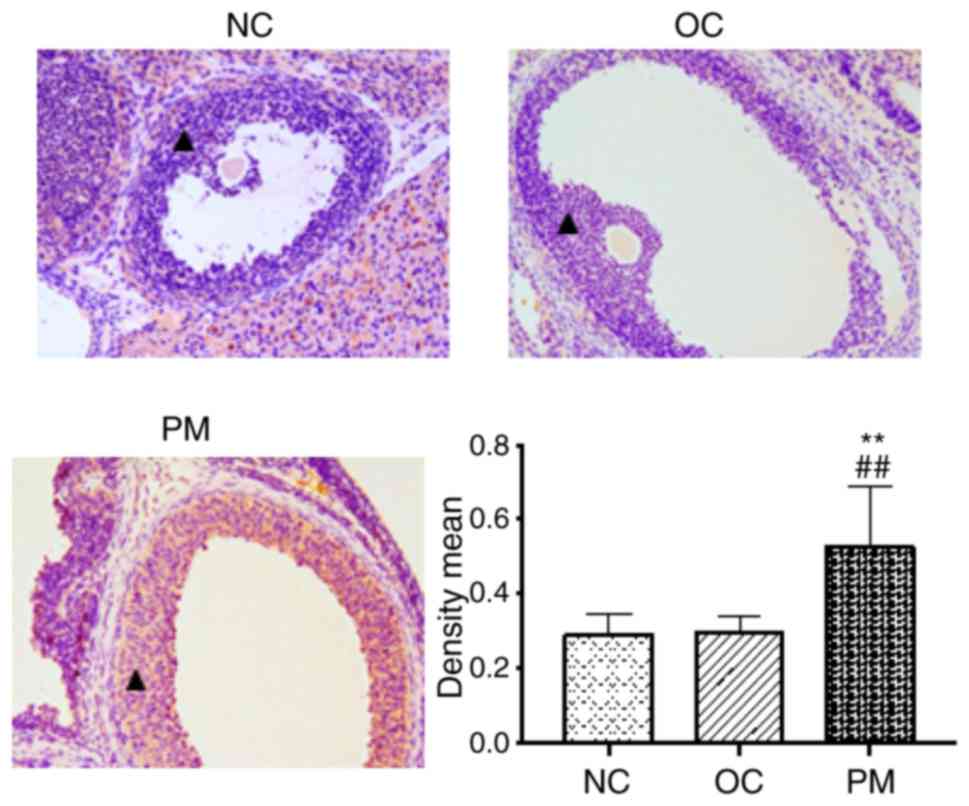
FKBP52 is increased in ovarian sections
of PCOS rats
FKBP52-positive staining (yellow) was present not
only in the nucleus, but also in the cytoplasm among all types of
cell in the rat ovary. In the GCs, the expression of FKBP52 in the
PM group was higher than in the other two groups (Fig. 2).
PCOS rats have higher mRNA expression
levels of FKBP52 and AR
The mRNA expression levels of FKBP52 and AR in the
rat ovarian tissues of the PM group were significantly higher than
those in the NC and OC groups. There was no statistically
significant difference between the NC and OC groups. The
experiments were performed in triplicate and repeated three times
(Fig. 3A).
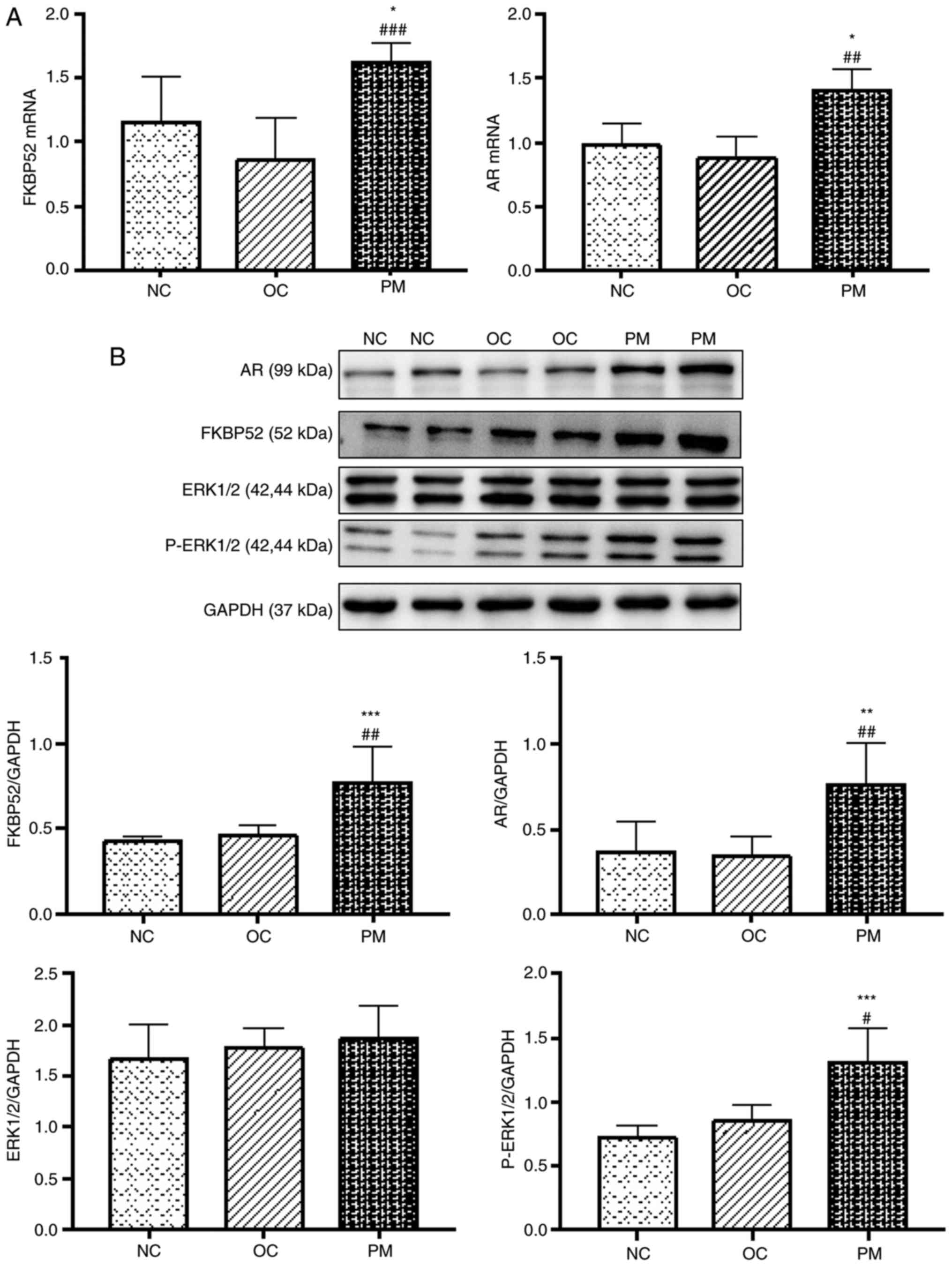 | Figure 3Expression of FKBP52, AR, ERK1/2 and
p-ERK1/2 in rats. (A) Comparison of mRNA expression levels of
FKBP52 and AR among the three groups (mean ± SD, n=8 per group).
The mRNA expression levels of FKBP52 and AR in the rat ovarian
tissues of the PM group were significantly higher than those in the
NC and OC groups. No significant difference was found between the
NC and OC groups (P>0.05). (B) Comparison of protein expression
levels of FKBP52, AR, ERK1/2 and p-ERK1/2 among the three groups
(mean ± SD, n=8 per group). The protein expression levels of
FKBP52, AR and p-ERK1/2 in the rat ovarian tissues of the PM group
were significantly higher than those in the NC and OC groups. No
significant difference was found between the NC and OC groups.
Protein expression of ERK1/2 did not differ significantly among the
groups. Experiments were performed in triplicate and repeated three
times. *P<0.05, **P<0.01 and
***P<0.001 NC group vs. PM group;
#P<0.05, ##P<0.01 and
###P<0.001 OC group vs. PM group. FKBP52, FK-506
binding protein 52; AR, androgen receptor; ERK1/2, extracellular
signal-regulated kinase; p-ERK1/2, phosphorylated ERK1/2; PM,
polycystic ovarian syndrome model; NC, normal control; OC, oil
control; SD, standard deviation. |
PCOS rats have higher protein expression
levels of FKBP52, AR and p-ERK1/2
The protein expression levels of FKBP52, AR and
P-ERK1/2 in the rat ovarian tissues of the PM group were
significantly higher than those of the NC and OC groups. There was
no statistically significant difference between the NC and OC
groups. The protein expression of ERK1/2 did not differ
significantly among the three groups. The experiments were
performed in triplicate and repeated three times (Fig. 3B).
Morphology and identification of GCs
The GCs exhibited a polygonal or cuboidal appearance
under inverted phase contrast microscopy on adhering to the culture
surface, proliferating, and spreading to form a monolayer (Fig. 4A). FSHR-positive staining (yellow)
was present in the nucleus and the cytoplasm of the GCs (Fig. 4B). Therefore, IF staining was
performed. The positive rate was >95%, therefore, the GCs
extracted met the requirements of subsequent trials (Fig. 4C).
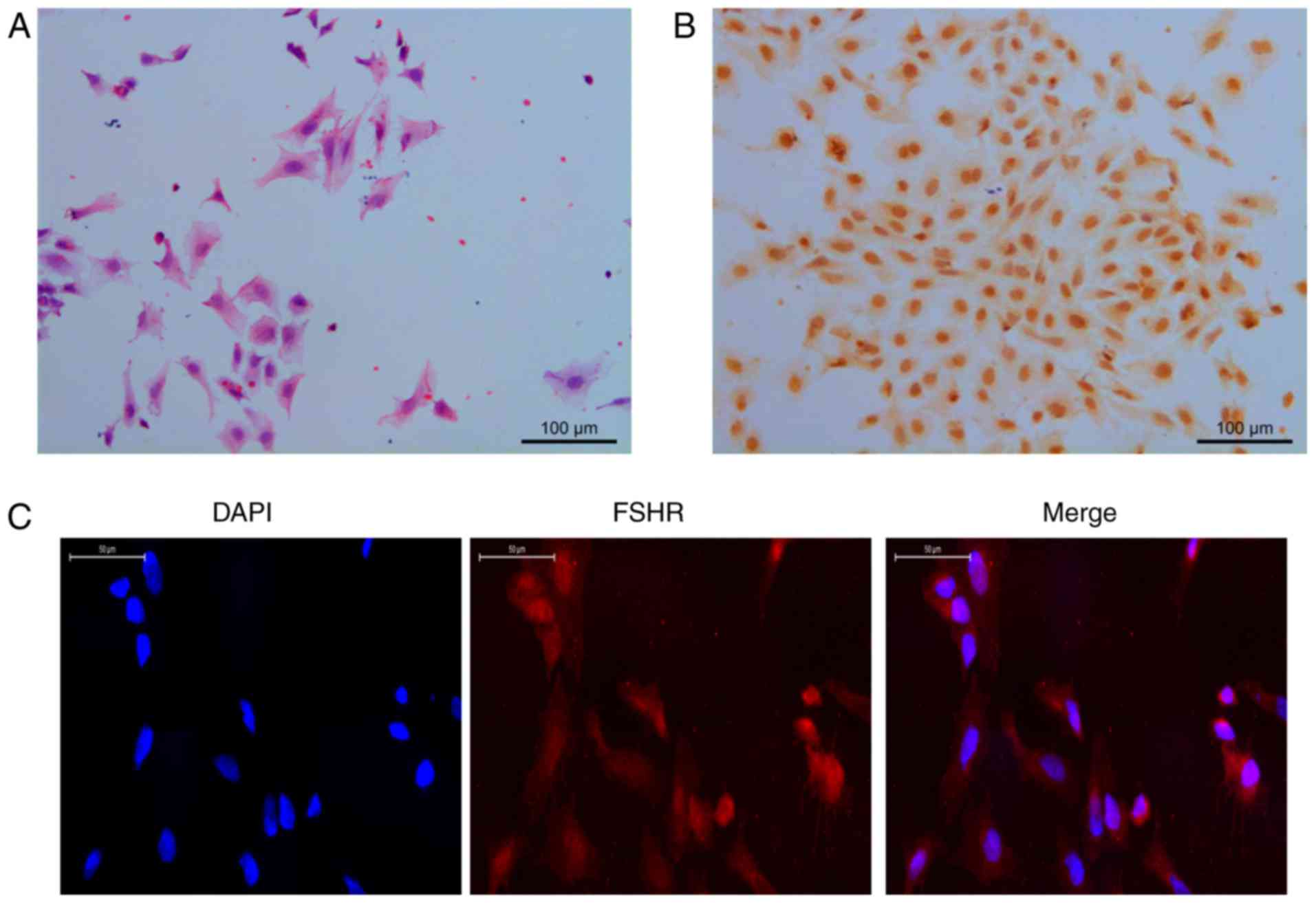 | Figure 4Morphology and identification of GCs
(hematoxylin and eosin staining, magnification, ×200;
immunocytochemistry staining, magnification ×200; IF staining,
magnification ×400). (A) GCs exhibited polygonal or cuboidal
appearance under inverted phase contrast microscopy on adhering to
the culture surface, proliferating, and spreading to form a
monolayer. (B) FSHR-positive staining (yellow) was present in the
nucleus and cytoplasm of GCs. IF staining was performed; (C) the
positive rate was >95%, therefore, the GCs extracted met the
requirements of subsequent trials. GCs, granulosa cells; FSHR,
follicle stimulating hormone receptor; IF, immunofluorescence. |
Expression of green fluorescence in
GCs
Based on the preliminary trial, appropriate results
were obtained following calculation according to the formula of
MOI: For the Oe negative control virus, the optimal dilution
concentration was 10−3, MOI=200; the optimal dilution
concentration of the FKBP4-Oe virus was 10−2, MOI=400;
the optimal dilution concentration of RNAi negative control virus
was 10−3, MOI=100; the optimal dilution concentration of
FKBP4-RNAi virus was 10−2, MOI=400. The cell
transfection rate was >80% (Fig.
5A-D).
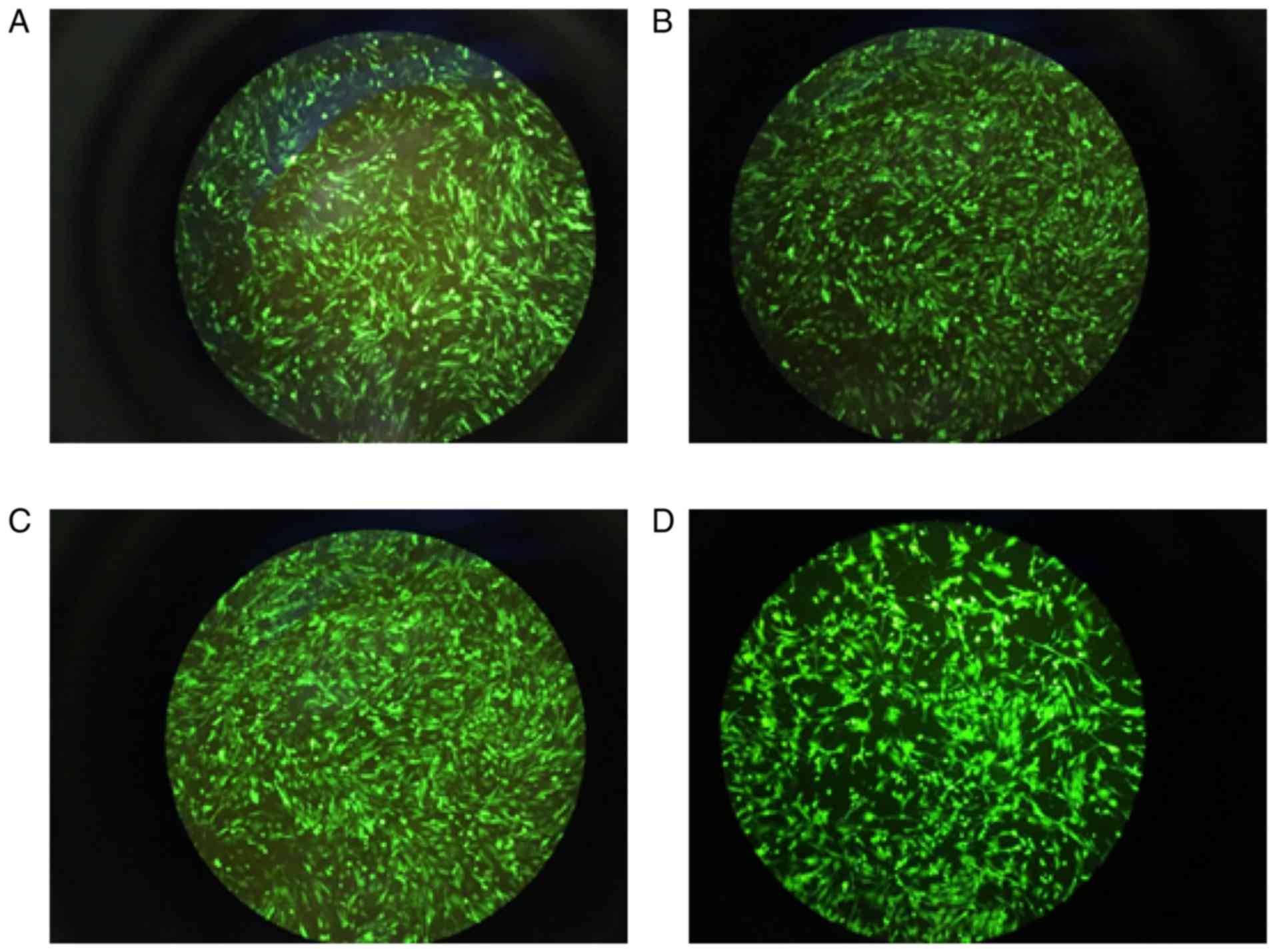 | Figure 5Expression of green fluorescence in
GCs. (A) Oe negative control virus group, 1011 PFU/ml,
MOI=200; (B) FKBP4-Oe virus group, 2×1010 PFU/ml,
MOI=400; (C) RNAi negative control virus group, 5×1010
PFU/ml, MOI=100; (D) FKBP4-RNAi virus group, 2×1010
PFU/ml, MOI=400). The cell transfection rate was >80%.
Magnification, ×100. GCs, granulosa cells; MOI, multiplicity of
infection. |
mRNA expression levels of FKBP52 and AR
increase following overexpression of FKBP4
The mRNA expression levels of FKBP52 and AR in the
FO group were significantly higher than those in the CO group and
EO group. There was no statistically significant difference between
the CO group and EO group. The experiments were performed in
triplicate and repeated three times (Fig. 6A).
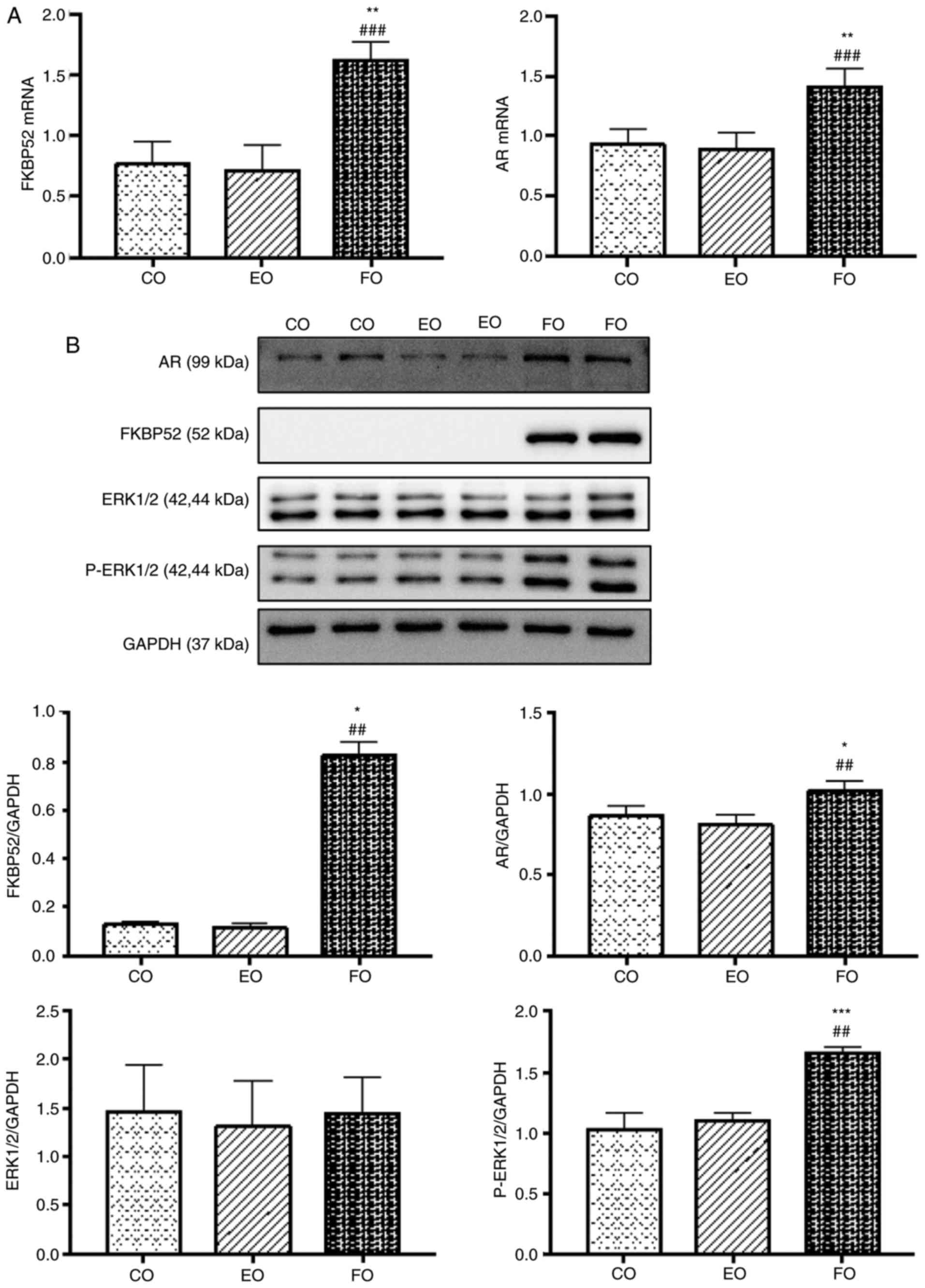 | Figure 6Expression of FKBP52, AR, ERK1/2 and
p-ERK1/2 in GCs. (A) Comparison of the mRNA expression levels of
FKBP52 and AR mRNA (mean ± SD, n=9 per group). mRNA expression
levels of FKBP52 and AR in the FO group were significantly higher
than those in the CO and EO groups. No significant difference was
found between the CO and EO groups (P>0.05). (B) Comparison of
the protein expression levels of FKBP52, AR, ERK1/2 and p-ERK1/2
(mean ± SD, n=6 per group). Protein expression levels of FKBP52, AR
and p-ERK1/2 in the FO group were significantly higher than those
in the CO and EO groups. No significant difference was found
between the CO and EO groups. Protein expression of ERK1/2 did not
differ significantly among the groups. (C) Comparison of the mRNA
expression levels of FKBP52 and AR (mean ± SD, n=9 per group). mRNA
expression levels of FKBP52 and AR in the FR group were
significantly lower than those in the CR and ER groups. No
significant difference was found between the CR and ER groups
(P>0.05). (D) Comparison of the protein expression levels of
FKBP52, AR, ERK1/2 and p-ERK1/2 (mean ± SD, n=6 per group). Protein
expression levels of FKBP52, AR and ERK1/2 in the FR group were
significantly lower than those in the CR and ER groups. p-ERK1/2
showed the opposite result. No significant difference was found
between the CR and ER groups. The experiment was repeated three
times. *P<0.05, **P<0.01 and ***P<0.001 NC group vs. PM
group; #P<0.05, ##P<0.01 and ###P<0.001 OC group vs. PM
group. FKBP52, FK-506 binding protein 52; AR, androgen receptor;
ERK1/2, extracellular signal-regulated kinase; p-ERK1/2,
phosphorylated ERK1/2; PM, polycystic ovarian syndrome model; NC,
normal control; OC, oil control; SD, standard deviation. |
Protein expression levels of FKBP52, AR
and p-ERK following the overexpression of FKBP4
The protein expression levels of FKBP52, AR and
p-ERK1/2 in the FO group were significantly higher than those in
the CO group and EO group. There was no statistically significant
difference between the CO group and EO group. The protein
expression of ERK1/2 did not differ significantly among the three
groups. The experiment was repeated three times (Fig. 6B).
mRNA expression levels of FKBP52 and AR
decrease following FKBP4 silencing
The mRNA expression levels of FKBP52 and AR in the
FR group were significantly lower than those in the CR group and ER
group. There was no statistically significant difference between
the CR group and ER group. The experiments were performed in
triplicate and repeated three times (Fig. 6C).
Protein expression levels of FKBP52, AR
and ERK decrease, and that of p-ERK increases following FKBP4
silencing
The protein expression levels of FKBP52, AR and
ERK1/2 in the FR group were significantly lower than those in the
CR group and ER group, whereas p-ERK showed the opposite result.
There was no statistically significant difference between the CR
group and ER group. The experiment was repeated three times
(Fig. 6D).
Discussion
PCOS is an endocrine disease featuring
hyperandrogenism which has a multifactorial pathogenesis and
heterogeneous clinical manifestations. Among all clinical
manifestations, variant degrees of hyperandrogenism are present in
the majority of patients with PCOS. The hyperandrogenism state of
ovarian tissues in the PCOS population can lead to dysgenesis.
In the present study, PCOS rats had higher
expression levels of FKBP52, AR and p-ERK1/2 in the ovary.
Following adenovirus vector transfection of GCs, the protein
expression levels of AR, ERK1/2 and p-ERK1/2 changed with FKBP4
gene silencing and overexpression.
PCOS model evaluation
At 35 days post-DHEA treatment, the vaginal smear
(Fig. 1A), H&E staining
(Fig. 1C), and ELISA results
(Fig. 1D) revealed that the PCOS
model was successfully established. Differences between rats and
humans in the mechanism of ovulation may explain why the ovarian
weights and ovarian coefficients of the PM group rats were
statistically lower in than the control groups (Fig. 1B).
FKBP52 in reproductive development
Previous studies on two independently derived
FKBP52-deficient (52KO) mouse lines served a vital role in the
reproductive development of mammals. Male 52KO mice exhibit
phenotypes in accordance with partial androgen insensitivity,
incorporating dysgenic prostate and seminal vesicles, ambiguous
external genitalia covering hypospadias and retention of nipples
into adulthood (9,13). They had reduced epididymal sperm
counts and motility (22), and
the sperm showed abnormal morphology. Therefore, gene knock-out
experiments in male mice have revealed FKBP52 as a crucial promoter
of physiological AR activity (9,13).
Female 52KO mice were yield as they exhibited normal
morphology, ovulation and fertilization. The dysgenesis was the
consequence of embryonic implantation and decidualization failure
on account of progesterone insensitivity and uterine defects
(23-25). In addition, FKBP52 may result in
endometriotic lesions, with decreased expression levels of FKBP52
in patients with endometriosis, and increased cell proliferation,
inflammation and angiogenesis resulting from progesterone
resistance in 52KO mice (26).
According to the above-mentioned findings, FKBP52 is important in
reproductive development.
FKBP52, AR, and PCOS
Previous studies have provided evidence to support a
direct pathological role for AR-mediated signaling in the
development of PCOS (27,28). It is an important extraovarian
mediator (29). Certain findings
have indicated how hyperandrogenism modulates AR ubiquitination,
resulting in antral follicle growth arrest in a chronically
androgenized PCOS rat model (30).
Few investigations have been performed on the impact
of FKBP52 on PCOS. In a comparison of normal women of childbearing
age with patients with PCOS using gene chip technology, FKBP52
protein-coding genes were different, and it may be closely
associated with abnormal androgen PCOS and obesity phenotype
(14). High-fat fed diabetic
NONcNZO mice exhibited higher rates of peri- and post-implantation
resorption, and had aberrant expression of uterine interferon-γ and
progesterone receptor (PGR) and its immunophilin co-chaperone
FKBP52 at nidation (30). The
aberrant PGR-targeted gene expression in PCOS-like rats pre- and
post-implantation overlapped with dysregulated expression of
FKBP52, which was linked to endometrial dysfunction and infertility
(31).
The results in the present experiments revealed that
the PM group had higher mRNA and protein levels of FKBP52 (Figs. 2 and 3) and AR (Fig. 3). This suggested that the
overexpression of FKBP52 and AR in the ovaries of PCOS rats may be
attributed to the stimulation of hyperandrogenism.
Morphology and identification of GCs
The H&E staining showed that cells were
polygonal or cuboidal under inverted phase contrast microscopy. As
FSHR is a unique marker of GCs, its positive rate in IHC and IF
staining was >95%. Therefore, the above results indicated that
the GCs extracted met the requirements of subsequent trials
(Fig. 4).
Adenovirus vector transfection of
GCs
GCs have different MOIs when they meet different
viral titers. An appropriate MOI not only guarantees the
transfection rate, but also conserves virus usage. In the
preliminary trial, when the cell transfection rate reached ~80%,
the MOI was calculated using the formula described above. This met
the requirements of subsequent trials (Fig. 5).
Activated AR in the cytoplasm may interact with
several signaling molecules, which in turn converge on MAPK/ERK
activation (15,16). In the present study, the PM rats
had higher expression levels of FKBP52, AR and p-ERK1/2 in the
ovary. Therefore, it was hypothesized that FKBP52 mediates AR
through the MAPK/ERK pathway. The Ad-siRNA-FKBP4-EGFP and
Ad-Oe-FKBP4-EGFP adenovirus vectors were constructed to verify the
association of FKBP52, AR and MAPK/ERK by transfecting GCs.
When FKBP4 was overexpressed, the mRNA and
expression levels of FKBP52 and AR increased and that of p-ERK1/2
increased, whereas ERK1/2 showed no change (Fig. 6A and B). Following FKBP4
silencing, the mRNA and expression levels of FKBP52 and AR
decreased, that of p-ERK1/2 increased and that of ERK1/2 decreased
(Fig. 6C and D). This may be
interpreted as the existence of a compensatory response and this
requires verification in the future. To the best of our knowledge,
this is the first time the above-mentioned results have been
reported. To a certain extent, these results explain the
association of FKBP52, AR and MAPK/ERK, and further verification is
required to confirm this.
Funding
The present study was approved the National Natural
Science Foundation of China (grant no. 81674012).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors’ contributions
SS performed the experiments, participated in
collecting data, performed the statistical analysis and drafted the
manuscript. YT participated in study design and assisted in
drafting the manuscript. Both authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Experimental
Animal Ethics committee of Nanjing University of Chinese Medicine
(Nanjing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
FKBP52
|
FK-506 binding protein 52
|
|
PCOS
|
polycystic ovarian syndrome
|
|
AR
|
androgen receptor
|
|
DHEA
|
dehydroepiandrosterone
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
H&E
|
hematoxylin and eosin
|
|
IHC
|
immunohistochemistry
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
WB
|
western blotting
|
|
GC
|
granulosa cell
|
|
SD
|
Sprague-Dawley
|
|
SPF
|
specific-pathogen-free
|
|
PBS
|
phosphate- buffered saline
|
|
MAPK
|
mitogen-activated protein kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
References
|
1
|
Chiofalo B, Laganà AS, Palmara V, Granese
R, Corrado G, Mancini E, Vitale SG, Ban Frangež H, Vrtačnik-Bokal E
and Triolo O: Fasting as possible complementary approach for
poly-cystic ovary syndrome: Hope or hype. Med Hypotheses. 105:1–3.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Asemi Z, Samimi M, Taghizadeh M and
Esmaillzadeh A: Effects of Ramadan fasting on glucose homeostasis,
lipid profiles, inflammation and oxidative stress in women with
polycystic ovary syndrome in Kashan, Iran. Arch Iran Med.
18:806–810. 2015.PubMed/NCBI
|
|
3
|
Franks S: Controversy in clinical
endocrinology: Diagnosis of polycystic ovarian syndrome: In defense
of the rotterdam criteria. J Clin Endocrinol Metab. 91:786–789.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qu J, Wang Y, Wu X, Gao L, Hou L and
Erkkola R: Insulin resistance directly contributes to androgenic
potential within ovarian theca cells. Fertil Steril. 91:1990–1997.
2009. View Article : Google Scholar
|
|
5
|
Laganà AS and Pizzo A: Authors’ reply to:
‘Empiric’ inositol supplementation in normal-weight non-insulin
resistant women with polycystic ovarian disease: From the absence
of benefit to the potential adverse effects. Arch Gynecol Obstet.
291:959–960. 2015. View Article : Google Scholar
|
|
6
|
Wang FF, Pan JX, Wu Y, Zhu YH, Hardiman PJ
and Qu F: American, European, and Chinese practice guidelines or
consensuses of poly-cystic ovary syndrome: A comparative analysis.
J Zhejiang Univ Sci B. 19:354–363. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tai PK, Maeda Y, Nakao K, Wakim NG,
Duhring JL and Faber LE: A 59-kilodalton protein associated with
progestin, estrogen, androgen, and glucocorticoid receptors.
Biochemistry. 25:5269–5275. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Davies TH and Sánchez ER: FKBP52. Int J
Biochem Cell Biol. 37:42–47. 2005. View Article : Google Scholar
|
|
9
|
Cheung-Flynn J, Prapapanich V, Cox MB,
Riggs DL, Suarez- Quian C and Smith DF: Physiological role for the
cochaperone FKBP52 in androgen receptor signaling. Mol Endocrinol.
19:1654–1666. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Periyasamy S, Warrier M, Tillekeratne MP,
Shou W and Sanchez ER: The immunophilin ligands cyclosporin A and
FK506 suppress prostate cancer cell growth by androgen
receptor-dependent and -independent mechanisms. Endocrinology.
148:4716–4726. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin JF, Xu J, Tian HY, Gao X, Chen QX, Gu
Q, Xu GJ, Song JD and Zhao FK: Identification of candidate prostate
cancer biomarkers in prostate needle biopsy specimens using
proteomic analysis. Int J Cancer. 121:2596–2605. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
De Leon JT, Iwai A, Feau C, Garcia Y,
Balsiger HA, Storer CL, Suro RM, Garza KM, Lee S, Kim YS, et al:
Targeting the regulation of androgen receptor signaling by the heat
shock protein 90 cochaperone FKBP52 in prostate cancer cells. Proc
Natl Acad Sci USA. 108:11878–11883. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yong W, Yang Z, Periyasamy S, Chen H,
Yucel S, Li W, Lin LY, Wolf IM, Cohn MJ, Baskin LS, et al:
Essential role for Co-chaperone FKBP52 but not FKBP51 in androgen
receptor-mediated signaling and physiology. J Biol Chem.
282:5026–5036. 2007. View Article : Google Scholar
|
|
14
|
Ketefian A, Jones MR, Krauss RM, Chen YD,
Legro RS, Azziz R and Goodarzi MO: Association study of androgen
signaling pathway genes in polycystic ovary syndrome. Fertil
Steril. 105:467–473. e42016. View Article : Google Scholar :
|
|
15
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar
|
|
16
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peterziel H, Mink S, Schonert A, Becker M,
Klocker H and Cato AC: Rapid signalling by androgen receptor in
prostate cancer cells. Oncogene. 18:6322–6329. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Belani M, Deo A, Shah P, Banker M, Singal
P and Gupta S: Differential insulin and steroidogenic signaling in
insulin resistant and non-insulin resistant human luteinized
granulosa cells-A study in PCOS patients. J Steroid Biochem Mol
Biol. 178:283–292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Miao ZL, Guo L, Wang YX, Cui R, Yang N,
Huang MQ, Qin WB, Chen J, Li HM, Wang ZN and Wei XC: The
intervention effect of Rosiglitozone in ovarian fibrosis of PCOS
rats. Biomed Environ Sci. 25:46–52. 2012.PubMed/NCBI
|
|
20
|
Anderson E, Lee GY and O’Brien K:
Polycystic ovarian condition in the dehydroepiandrosterone-treated
rat model: Hyperandrogenism and the resumption of meiosis are major
initial events associated with cystogenesis of antral follicles.
Anat Rec. 249:44–53. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Hong J, Kim ST, Tranguch S, Smith DF and
Dey SK: Deficiency of co-chaperone immunophilin FKBP52 compromises
sperm fertilizing capacity. Reproduction. 133:395–403. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tranguch S, Cheung-Flynn J, Daikoku T,
Prapapanich V, Cox MB, Xie H, Wang H, Das SK, Smith DF and Dey SK:
Cochaperone immunophilin FKBP52 is critical to uterine receptivity
for embryo implantation. Proc Natl Acad Sci USA. 102:14326–14331.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tranguch S, Wang H, Daikoku T, Xie H,
Smith DF and Dey SK: FKBP52 deficiency-conferred uterine
progesterone resistance is genetic background and pregnancy stage
specific. J Clin Invest. 117:1824–1834. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Z, Wolf IM, Chen H, Periyasamy S,
Chen Z, Yong W, Shi S, Zhao W, Xu J, Srivastava A, et al:
FK506-binding protein 52 is essential to uterine reproductive
physiology controlled by the progesterone receptor A isoform. Mol
Endocrinol. 20:2682–2694. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hirota Y, Tranguch S, Daikoku T, Hasegawa
A, Osuga Y, Taketani Y and Dey SK: Deficiency of immunophilin
FKBP52 promotes endometriosis. Am J Pathol. 173:1747–1757. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Caldwell AS, Eid S, Kay CR, Jimenez M,
McMahon AC, Desai R, Allan CM, Smith JT, Handelsman DJ and Walters
KA: Haplosufficient genomic androgen receptor signaling is adequate
to protect female mice from induction of polycystic ovary syndrome
features by prenatal hyperandrogenization. Endocrinology.
156:1441–1452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Walters KA: Role of androgens in normal
and pathological ovarian function. Reproduction. 149:R193–R218.
2015. View Article : Google Scholar
|
|
29
|
Caldwell ASL, Edwards MC, Desai R, Jimenez
M, Gilchrist RB, Handelsman DJ and Walters KA: Neuroendocrine
androgen action is a key extraovarian mediator in the development
of polycystic ovary syndrome. Proc Natl Acad Sci USA.
114:E3334–E3343. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lim JJ, Lima PDA, Salehi R, Lee DR and
Tsang BK: Regulation of androgen receptor signaling by
ubiquitination during folliculogenesis and its possible
dysregulation in polycystic ovarian syndrome. Sci Rep. 7:102722017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Albaghdadi AJH and Kan FWK:
Immunosuppression with tacrolimus improved implantation and rescued
expression of uterine progesterone receptor and its co-regulators
FKBP52 and PIASy at nidation in the obese and diabetic mice:
Comparative studies with metformin. Mol Cell Endocrinol. 460:73–84.
2018. View Article : Google Scholar
|