Introduction
Angiogenesis, an essential process in which new
blood vessels are formed, not only serves a crucial role in
physiological processes, including embryonic development and wound
repair, but also in pathological conditions, including chronic
inflammation, cancer, heart disease and diabetic retinopathy
(1,2). It exerts an important effect in
promoting aggressive tumor activity, including tumor growth,
metastasis and invasion (3).
Progression of cancer is characterized by the stimulation of
pro-angiogenic factors, including hypoxia, vascular endothelial
growth factor (VEGF), cytokine interleukin-6 and
metalloproteinases, in the growing endothelial cells (4). VEGF has been demonstrated to be one
of the most important angiogenesis growth factors that induces
permeability, proliferation, migration and tube formation (5). Numerous stimuli, including hypoxia,
cytokines and oxidative stress, can increase VEGF expression
(2,6). The inhibition of neoangiogenesis is
considered to be an important potential strategy for efficient and
effective antitumor agents that prevent cancer proliferation and
metastasis (7,8).
Reactive oxygen species (ROS), including
H2O2 and superoxide anion radicals, are a
family of reactive molecules that serve a pivotal role in numerous
cellular processes, including metabolism, differentiation,
proliferation and cell death, by regulating critical signaling
pathways (9). ROS are generated
under various physiological and pathological conditions, including
inflammation (10) and ischemia
(11). Excessive levels of ROS
result in an imbalance in the metabolism of these reactive
intermediates as a result of oxidative stress, and is associated
with various diseases, including cancer and cardiovascular and
neurodegenerative diseases (12).
The hypoxia inducible factor (HIF)-1α response to hypoxia is
associated with hypoxia-induced production of ROS (13). Accumulating evidence indicates
that VEGF and ROS exert a critical role in vascular pathophysiology
(14,15). However, the interaction between
VEGF and ROS remains controversial. Certain studies have
demonstrated that VEGF could take advantage of ROS as messenger
intermediates downstream of VEGF receptor 2, thus affecting
cellular proliferation and tube formation (16,17). Other studies indicated that ROS
trigger the induction of VEGF, but the underlying interaction
remains ambiguous (18-20). KC7F2, as an exogenous HIF-1α
translation inhibitor, is involved in cancer-associated
angiogenesis, and can be evaluated in terms of its
anti-proliferative actions and effects on the metabolism of cancer
cells (21). Furthermore, KC7F2
can also be used as a HIF-1α inhibitor (22).
Melatonin (MLT), a well-known natural hormone
secreted primarily in the pineal gland, has attracted attention due
to its various effects in numerous critical physiological,
including sleep, the circadian rhythm and antioxidant cell
protection against free radicals, and pathological, including tumor
progression, processes (23).
Accumulating evidence indicates that MLT is capable of directly
scavenging radicals and radical-associated reactants, as well as
indirectly stimulating anti-oxidative enzymes (24,25). It has been reported that MLT can
inhibit the development of a variety of cancer types (26), including liver (27-29), lung (30,31), breast (32-34), pancreatic and brain cancer
(35). However, the underlying
mechanism of MLT varies in different cancer types (36). Additionally, reports indicated
that MLT decreases the expression of HIF-1α and VEGF caused by
different factors in various cultivated cells, particularly
hypoxia-induced accumulation of HIF-1α protein and expression of
VEGF (37-39). Accumulating evidence indicates
that the antitumor effect of MLT is associated with the inhibition
of angiogenesis (24,40,41). It has been reported that ROS
produced under hypoxic conditions inactivate the PHD2 enzyme and
thus prevent its degradation (42). Therefore, ROS stabilizes HIF-1α,
allowing it to act as a transcription factor. However, the specific
mechanism of anti-angiogenesis activity of MLT has not been
systematically elucidated. Therefore, the aim of the present study
was to investigate the effect of MLT on HUVECs and the associated
underlying mechanism.
Materials and methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
obtained from the Cell Bank of the Chinese Academy of Science
(Shanghai, China) and cultured in a mixture containing RPMI-1640,
10% fetal bovine serum (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA), 100 IU/ml penicillin and 100 μg/ml
streptomycin (Sangon Biotech Co., Ltd., Shanghai, China). The cells
were incubated at 37°C in a humidified atmosphere which was
maintained at 5% CO2. All experiments were performed
with HUVECs that had been cultured for ≤6 passages.
Hypoxia treatment
Control cells were incubated for 6 or 24 h under
normoxic conditions (21% O2, 5% CO2 and 74%
N2 at 37°C) in a humidified incubator. According to the
manufacturer’s protocols, hypoxic conditions (termed hypoxia) were
induced using an airtight modular incubator chamber
(Billups-Rothenberg, Inc., San Diego, CA, USA). Briefly, the cells
[1×104 cells/well in cell viability assay;
2×104 cells/well in tube formation assay; and
1×105 cells/well in western blotting, ELISA and flow
cytometry (FCM) assays] were sealed in the modular incubator
chamber with a sterile 1X PBS reserve to maintain humidity, and
then purged with a reduced O2 gas mixture (1%
O2, 5% CO2 and 94% N2) at 37°C for
6 h or 24 h.
Cell viability assay
HUVECs were seeded into 96-well plates at a density
of 1×104 cells/well. The conditioned medium was
aspirated and 100 μl fresh Cell Counting kit 8 (CCK8)
solution (Dojindo Molecular Technologies, Inc., Kunamoto, Japan)
with serum-free RPMI-1640 medium was carefully added to each well.
According to the manufacturer’s protocols, the plates were then
incubated at 37°C for 0.5-4 h in the dark. The absorbance was
periodically detected using a microplate reader at 450 nm.
Detection of ROS and VEGF by flow
cytometry (FCM)
After treatment with or without MLT
(1×10-5 M) under normoxia or hypoxia (according to the
aforementioned conditions) at 37°C for 6 h, or pretreatment with
H2O2 (50 μM; based on its
cyto-toxicity, a dose of 50 μM H2O2
was selected for subsequent experiments) or VEGF (5 ng/ml) for 4 h,
followed by MLT (1×10-5 M) at 37°C for 6 h, and then the
release of ROS of HUVECs was detected by the Reactive Oxygen
Species Assay kit (cat. no. S0033; Beyotime Institute of
Biotechnology, Haimen, China). The levels of intracellular ROS
generation were determined by incubating the cells in serum-free
RPMI-1640 supplemented with 10 mM 2,7-dichlorofluorescein diacetate
(DCFH-DA; Beyotime Institute of Biotechnology) or PBS (as the blank
control) in the dark at 37°C for 30 min. DCFH-DA can be converted
to the fluorescent dichlorofluorescein by ROS. Briefly, the cells
were pretreated with different compounds (MLT,
H2O2, VEGF, KC7F2, MLT plus VEGF, or MLT plus
KC7F2; the concentrations of these compounds were the same as
aforementioned) at 37°C for 6 h and/or conditions (hypoxia and
normoxia, according to the aforementioned conditions). The cells
were then rinsed and washed with cold PBS, followed by incubation
with 1 mmol/l DCHF-DA in the dark at 37°C for 30 min. The cells
were then trypsinized and washed with PBS again and resuspended in
serum-free RPMI-1640 medium (1×106 cells/1 ml medium)
for the FCM assay. FCM was performed on a Beckman Cyan flow
cytometer (Beckman Coulter, Inc., Brea, CA, USA) using CellQuest
software (version 7.1; Beckman Coulter, Inc.). At least 15,000
events were analyzed. All experiments were performed with
biological triplicates and data are representative of at least
three independent experiments.
Additionally, the median fluorescence intensity of
VEGF was measured with a FCM assay. Cells were washed with PBS once
and then trypsinized and collected by gently centrifugation at 350
× g at 4°C for 6 min. A commercial kit, eBioscience™ Intracellular
Fixation/Perm Buffer (cat. no. 88-8824-00; eBio-science; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), was used to fix and
rupture the membrane according to the manufacturer’s protocols.
After centrifugation at 150 × g for 10 min at room temperature, the
precipitate was resuspended in 1 ml 0.9% physiological saline and
centrifuged at 150 × g for 10 min at room temperature. The
precipitate was then resuspended in 150 μl 0.9%
physiological saline and blocked with human AB serum
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 4°C for 30 min.
Subsequently, a VEGF antibody [human VEGF allophycocyanin
(APC)-conjugated antibody; 5 μl; 1:20; cat. no. IC2931A;
R&D Systems, Inc., Minneapolis, MN, USA] or isotype antibody
(mouse IgG2A APC-conjugated antibody; 5 μl; 1:20; cat. no.
IC003A; R&D Systems, Inc.) was then incubated with the cells at
room temperature in the dark for 30 min. Cells were centrifuged 350
× g at 4°C for 6 min and washed with PBS twice to remove the
non-specific binding antibody. Analysis was performed on a Beckman
flow cytometer with CellQuest software.
Cell treatment
To examine the effect of different reagents, HUVECs
were cultivated with serum-free RPMI-1640 medium containing
1×10-5 M MLT (Sigma-Aldrich; Merck KGaA) alone at 37°C
for 6 h, and/or pretreated with H2O2 (50
μM; Sigma-Aldrich; Merck KGaA), KC7F2 (20 μM;
Sigma-Aldrich; Merck KGaA) and VEGF (5 ng/ml; R&D Systems,
Inc.) at 37°C for 4 h. The vehicles (1% PBS for VEGF group and 1%
DMSO for other groups) were used as the controls. The cell culture
supernatant was collected following MLT exposure at 37°C for 6 h
and stored at -80°C for further study.
Tube formation assay
Basement membrane extracellular matrix (Matrigel; BD
Biosciences, San Jose, CA, USA) was thawed at 4°C overnight.
Pipette tips (200 μl) and a 96-well plate were also kept at
4°C overnight, and the plate and tips were placed on ice during the
entire experiment. Primary Matrigel (60 μl) was loaded in
each well, and the plate was incubated at 37°C for 30 min to allow
the matrix to polymerize. The pretreated HUVECs were resuspended
and recounted to achieve the appropriate cell density
(2×104 cells/well). The plate was kept at room
temperature for 15 min and then transferred to the incubator at
37°C. After 4-6 h incubation, the capillary-like tube formation was
quantified by counting numbers of junctions/enclosed circles in 5
randomly selected optical fields using an Olympus BX51+DP70
fluorescence microscope (Olympus Corporation, Tokyo, Japan;
magnification, ×40 or ×100).
Assessment of the secretion levels of
VEGF
The VEGF protein secreted into the conditioned
medium by HUVECs was measured with a commercially available human
VEGF ELISA kit (cat. no. 1117342; DAKEWE, Inc., Shenzhen, China;
http://www.biocity.net/). The conditioned medium
was collected and centrifuged at 350 × g at 4°C for 15 min to
remove cellular debris, and then recollected and stored at -80°C
until the ELISA assay was performed.
Western blot analysis for HIF-1α and
VEGF
HUVECs were seeded (2×105 cells/well) in
a 6-well plate and incubated at 37°C for 24 h, then treated with
MLT (1×10-5 M) or left untreated under normoxic or
hypoxic conditions (according to the aforementioned conditions) at
37°C for 6 h prior to protein isolation. The cells in culture were
rinsed twice with ice-cold PBS and treated with
radioimmunoprecipitation assay lysis buffer supplemented with 1 mM
phenylmethylsulfonyl fluoride and protease inhibitor cocktail (all
from Beyotime Institute of Biotechnology) on ice for 30 min, then
centrifuged at 13,400 × g at 4°C for 25 min. The supernatant was
collected. Protein concentration was measured by bicinchoninic acid
protein assay (Beyotime Institute of Biotechnology). Equal amounts
of total protein (15 μg) were segregated by 10% SDS-PAGE
(Epizyme Biotechnology, Shanghai, China) and transferred onto
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). The membranes were blocked with 10% non-fat milk in TBS
containing 0.1% Tween-20 (TBST) for 1 h at room temperature.
Subsequently, the membranes were probed overnight at 4°C with
primary antibodies against HIF-1α (rabbit; 1:500 dilution; cat. no.
36169S), VEGF (rabbit; 1:1,000 dilution; cat. no. 2463S) or β-actin
(rabbit; 1:3,000 dilution; cat. no. 4970S; Cell Signaling
Technology, Inc., Danvers, MA, USA). This was followed by washing
with 1X TBST buffer four times and incubation with the
corresponding horseradish peroxidase-conjugated secondary antibody
(1:5,000 dilution; cat. no. 2357; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at room temperature for 2 h. Immunoreactive bands
were visualized using a Western Blotting Luminol Reagent (ECL) kit
(Pierce; Thermo Fisher Scientific, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Analysis of HIF-1α (HIF1A) and VEGF
mRNA expression in HUVECs was conducted via RT-qPCR following
incubation of cells with 1 mM MLT and/or hypoxic conditions
(according to the aforementioned conditions) at 37°C for 6 h. Total
RNA was isolated from HUVECs using TRIzol® reagent
(Takara Bio, Inc., Otsu, Japan), according to the manufacturer’s
protocol. The A260/A280 nm absorbance ratio was maintained at
1.8-2.0. For cDNA synthesis, RT was performed using PrimeScript RT
Master mix (Takara Bio, Inc.), according to the manufacturer’s
instructions. qPCR was performed with an ABI 7900HT system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with SYBR®
Green PCR Master mix (Takara Bio, Inc.), according to the
manufacturer’s instructions. The thermocycling conditions were:
95°C for 30 min for 1 cycle; 94°C for 5 sec and 60°C for 34 sec for
40 cycles; finally, an extension step at 72°C for 10 min.
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were used for
normalization, and the relative expression levels of HIF1A
and VEGF were calculated using the 2-ΔΔCq method
(43). The primer sequences were
as follows: HIF1A forward, 5’-GAA CGT CGA AAA GAA AAG TCT
CG-3’ and reverse, 5’-CCT TAT CAA GAT GCG AAC TCA CA; VEGF
forward, 5’-GGG CAG AAT CAT CAC GA A GT-3, and reverse, 5’-AAA TGC
TTT CTC CGC TCT GA-3; and GAPDH forward, 5’-GGA GCG AGA TCC
CTC CAA AAT-3 and reverse, 5’-GGC TGT TGT CAT ACT TCT CAT GG-3.
Statistical analysis
All data are presented as the mean ± standard error
of the mean. Student’s t-test for two groups comparisons or one-way
ANOVA with the Bonferroni’s post-hoc test for multiple comparisons
was performed using GraphPad Prism 6 software (GraphPad Software,
Inc., La Jolla CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
MLT suppresses the viability and
angiogenesis of HUVECs
To investigate the influence of MLT on the
proliferation and angiogenesis of HUVECs in vitro, CCK8 and
tube formation assays were performed. As depicted in Fig. 1A, MLT treatment suppressed the
viability of HUVECs in a dose-dependent manner (P<0.001 for Ctrl
vs. 10-5 M MLT or P<0.0001 for Ctrl vs.
10-3 M MLT). Additionally, MLT (1×10-5 M)
resulted in a significantly reduced level of tube formation of
HUVECs, compared with the control (Fig. 1B and C; P<0.001). In addition,
hypoxia condition promoted the tube formation of HUVECs, and this
effect was reversed by MLT (Fig. 1B
and C; P<0.05 for Nor Ctrl vs. Nor MLT or P<0.001 for Hyp
Ctrl vs. Hyp MLT). These data indicate that MLT restricts the
viability and angiogenesis of HUVECs in vitro.
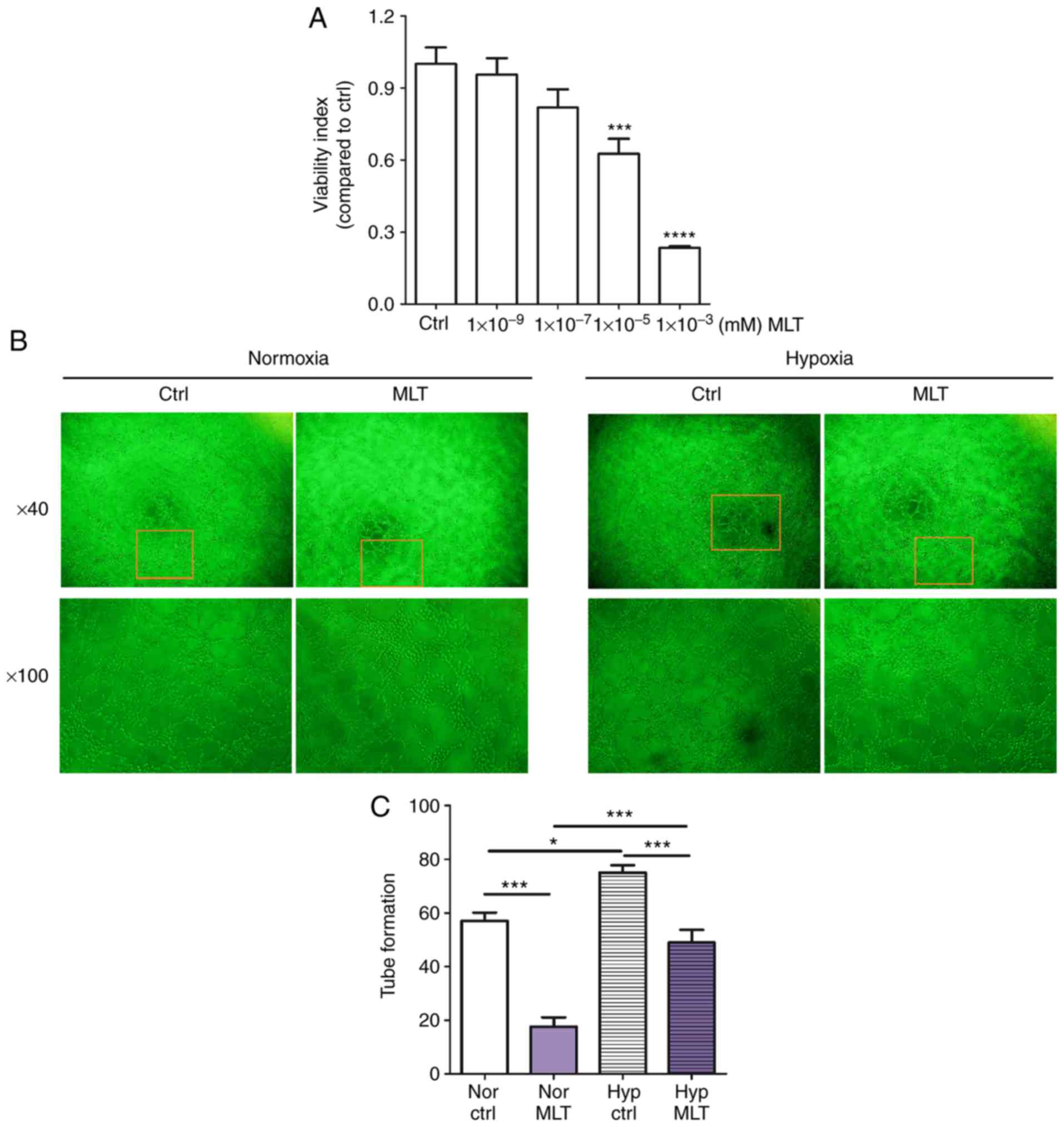 | Figure 1MLT suppresses the viability and
angiogenesis of HUVECs. (A) HUVECs were treated with MLT at various
concentrations (1×10-9, 1×10-7,
1×10-5 or 1×10-3 mM) or left untreated for 24
h. The viability of HUVECs was determined using a Cell Counting kit
8 viability assay. (B) HUVECs were treated with MLT
(1×10-5 M) under normoxia or hypoxia conditions, and
imaged under an Olympus microscope. Magnification, ×40 or ×100. (C)
The statistics histogram for. (B) Data are presented as the mean ±
standard error of the mean. *P<0.05,
***P<0.001 or ****P<0.0001, compared
with the Ctrl group. Ctrl, control; MLT, melatonin; HUVECs, human
umbilical vein endothelial cells; Nor Ctrl, Ctrl HUVECs under
normoxia condition; Nor MLT, MLT-treated HUVECs under normoxia
condition; Hyp Ctrl, Ctrl HUVECs under hypoxia condition; Hyp MLT,
MLT-treated HUVECs under hypoxia condition. |
MLT obstructs hypoxia-induced VEGF
production of HUVECs
It is widely acknowledged that VEGF is an important
angiogenic factor that stimulates the formation of new blood
vessels (5). To elucidate the
molecular mechanisms of MLT associated with tube formation, the
expression of VEGF following treatment with MLT was determined. It
was observed that MLT significantly restricted the expression of
VEGF at the mRNA and protein levels (Fig. 2A-C; P<0.0001, compared with the
Ctrl groups). Additional treatment with recombinant human VEGF
protein dose-dependently increased the viability of HUVECs
(Fig. 2D; P<0.01 for Ctrl vs.
1 ng/ml VEGF, P<0.001 for Ctrl vs. 2.5 ng/ml VEGF or P<0.0001
for Ctrl vs. 5 ng/ml VEGF), while this effect was weakened by MLT
(Fig. 2E; P<0.01 for Ctrl vs.
MLT, P<0.001 for VEGF vs. MLT+VEGF). The same inhibition
occurred in the tube formation assay, with MLT counteracting
VEGF-stimulated tubular network formation (Fig. 2F; P<0.05 for Ctrl vs. MLT or
P<0.001 for VEGF vs. MLT+VEGF). These data indicate that the
inhibitory effect of MLT on viability and angiogenesis of HUVECs is
dependent on VEGF.
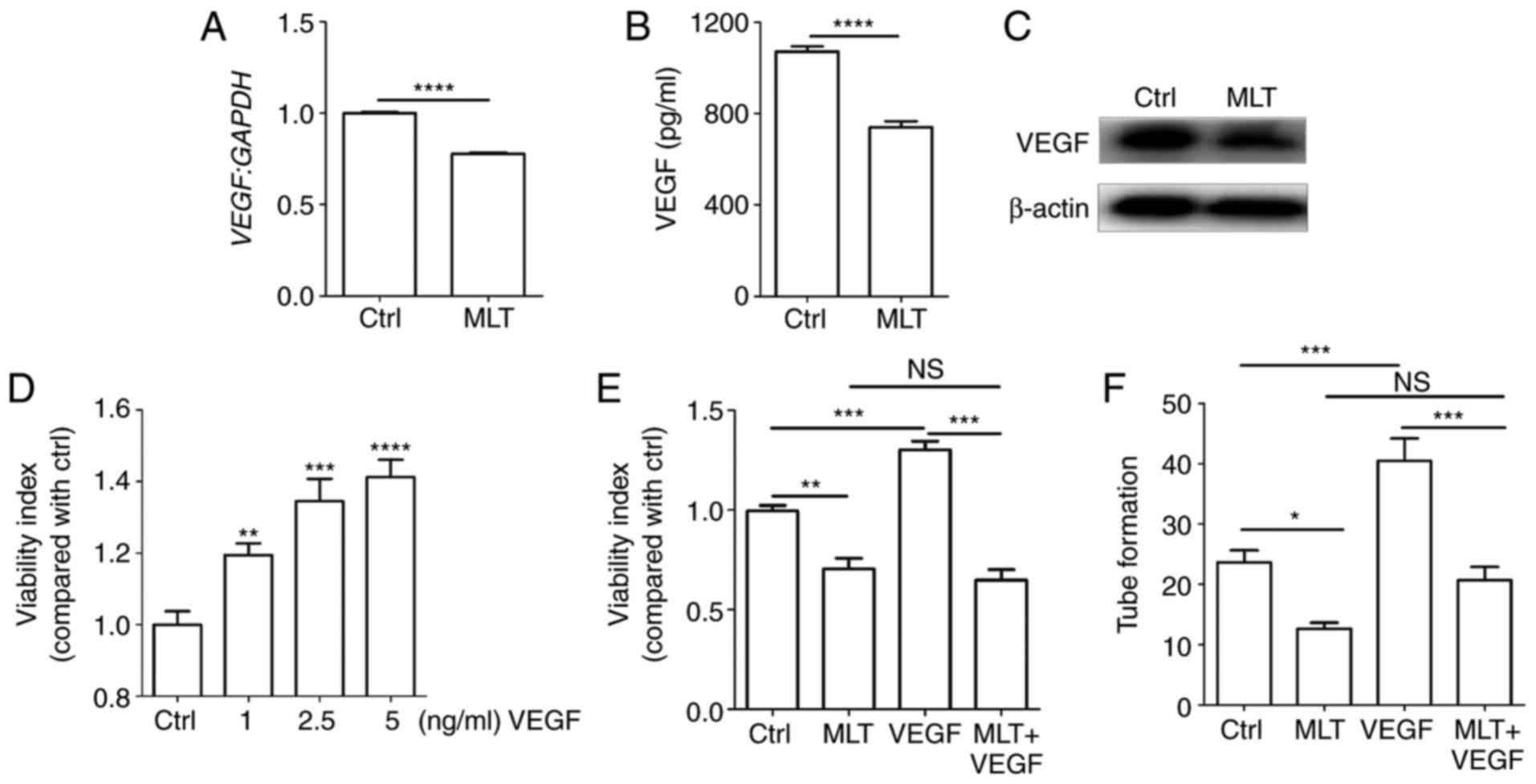 | Figure 2The inhibitory effect of MLT on the
viability and angiogenesis of HUVECs is dependent on VEGF. HUVECs
were treated with MLT (1×10-5 M) for 24 h, then the expression of
VEGF was determined by (A) reverse transcription-quantitative
polymerase chain reaction, (B) ELISA and (C) western blotting. (D)
HUVECs were stimulated with or without recombinant human VEGF (1-5
ng/ml), then cell viability was detected by CCK8 assay. Cells were
pretreated with VEGF (5 ng/ml) for 4 h followed by MLT for a
further 2 h, then (E) cell viability and (F) tube formation were
analyzed via CCK8 and tube formation assays, respectively. Data are
presented as the mean ± standard error of the mean.
*P<0.05, **P<0.01,
***P<0.001 or ****P<0.0001, compared
with the Ctrl group. NS, no significant difference; VEGF, vascular
endothelial growth factor; HUVECs, human umbilical vein endothelial
cells; CCK8, Cell Counting kit 8; Ctrl, control; MLT,
melatonin. |
MLT suppresses the expression of VEGF
induced by hypoxia
Hypoxia stimulates the formation of new capillary
vessels to counteract low oxygen tension. HUVECs were cultured
under normoxic or hypoxic conditions. As depicted in Fig. 3A, hypoxia significantly enhanced
HIF1A mRNA levels (P<0.05, compared with the Ctrl group).
By contrast, MLT exerted a significant inhibitory effect on the
transcription of HIF1A (Fig.
3B; P<0.01, compared with the Ctrl group). Subsequently, it
was determined that the mRNA and protein level of VEGF were
significantly upregulated in HUVECs under hypoxia condition
(Fig. 3C and D; P<0.01 for
normoxia group vs. hypoxia group or P<0.001 for normoxia group
vs. hypoxia group). Additionally, the results of western blotting
demonstrated that MLT notably suppressed the expression of HIF-1α
and VEGF that was upregulated due to hypoxia (Fig. 3E-G). These results indicate that
MLT can inhibit hypoxia-induced VEGF expression and further
obstruct VEGF-induced angiogenesis of HUVECs.
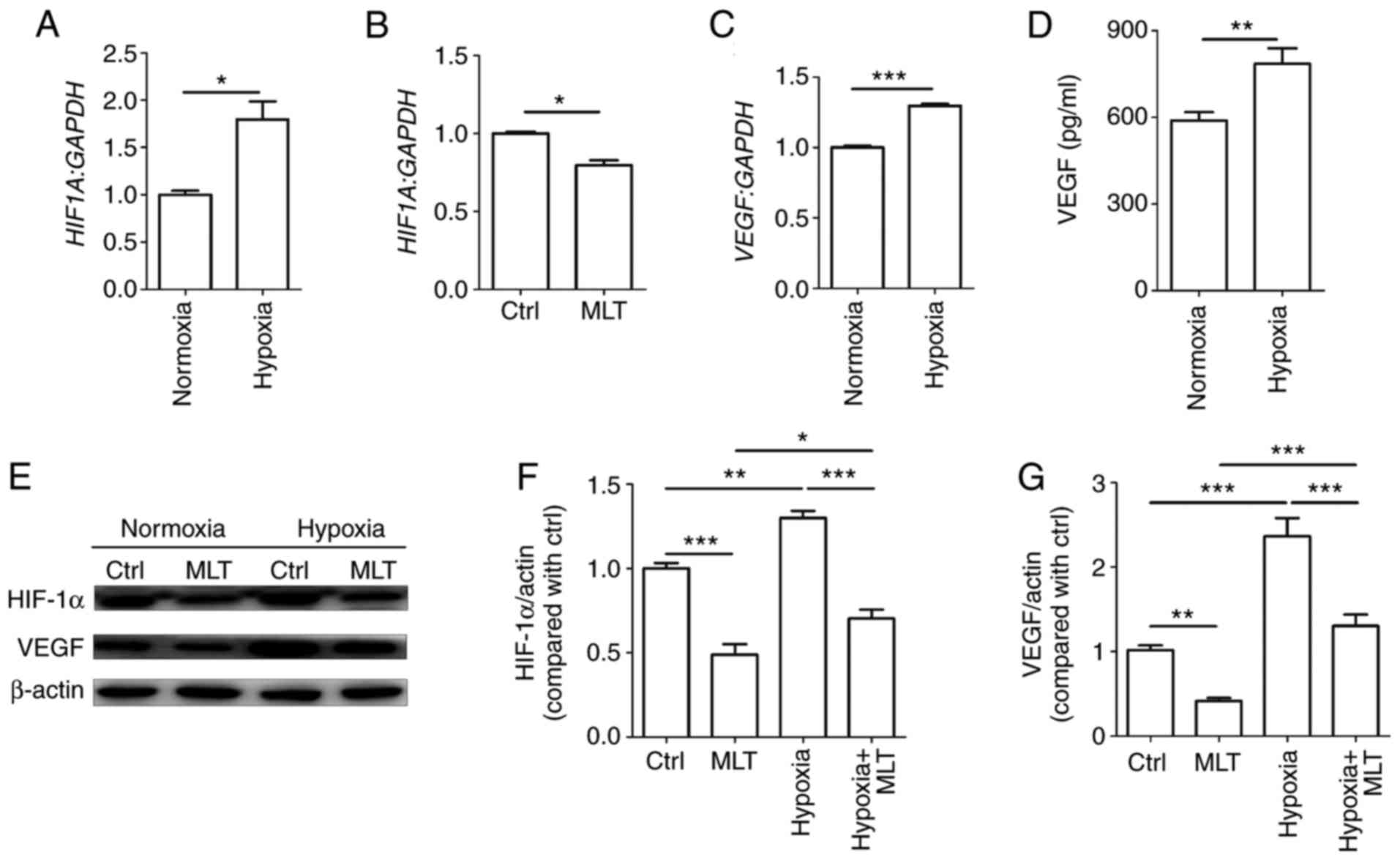 | Figure 3MLT obstructs the expression of VEGF
induced by hypoxia. HUVECs were (A) cultured under normoxia or
hypoxia conditions, (B) or treated with or without MLT
(1×10-5 M) for 24 h. The mRNA expression of HIF1A
was detected by RT-qPCR. (C) HUVECs were cultured under normoxia or
hypoxia conditions for 24 h, and the mRNA expression of VEGF
was detected by RT-qPCR. (D) The secretion of VEGF was detected by
ELISA. (E) HUVECs were treated with or without MLT
(1×10-5 M) under the condition of normoxia or hypoxia
for 24 h, and then the expression of HIF-1α and VEGF was measured
by western blotting. (F) The statistics histogram of HIF-1α. (G)
The statistics histogram of VEGF. Data are presented as the mean ±
standard error of the mean. *P<0.05,
**P<0.01 or ***P<0.001. VEGF, vascular
endothelial growth factor; HUVECs, human umbilical vein endothelial
cells; Ctrl, control; MLT, melatonin; HIF-1α, hypoxia inducible
factor-1α; HIF1A, HIF-1α; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
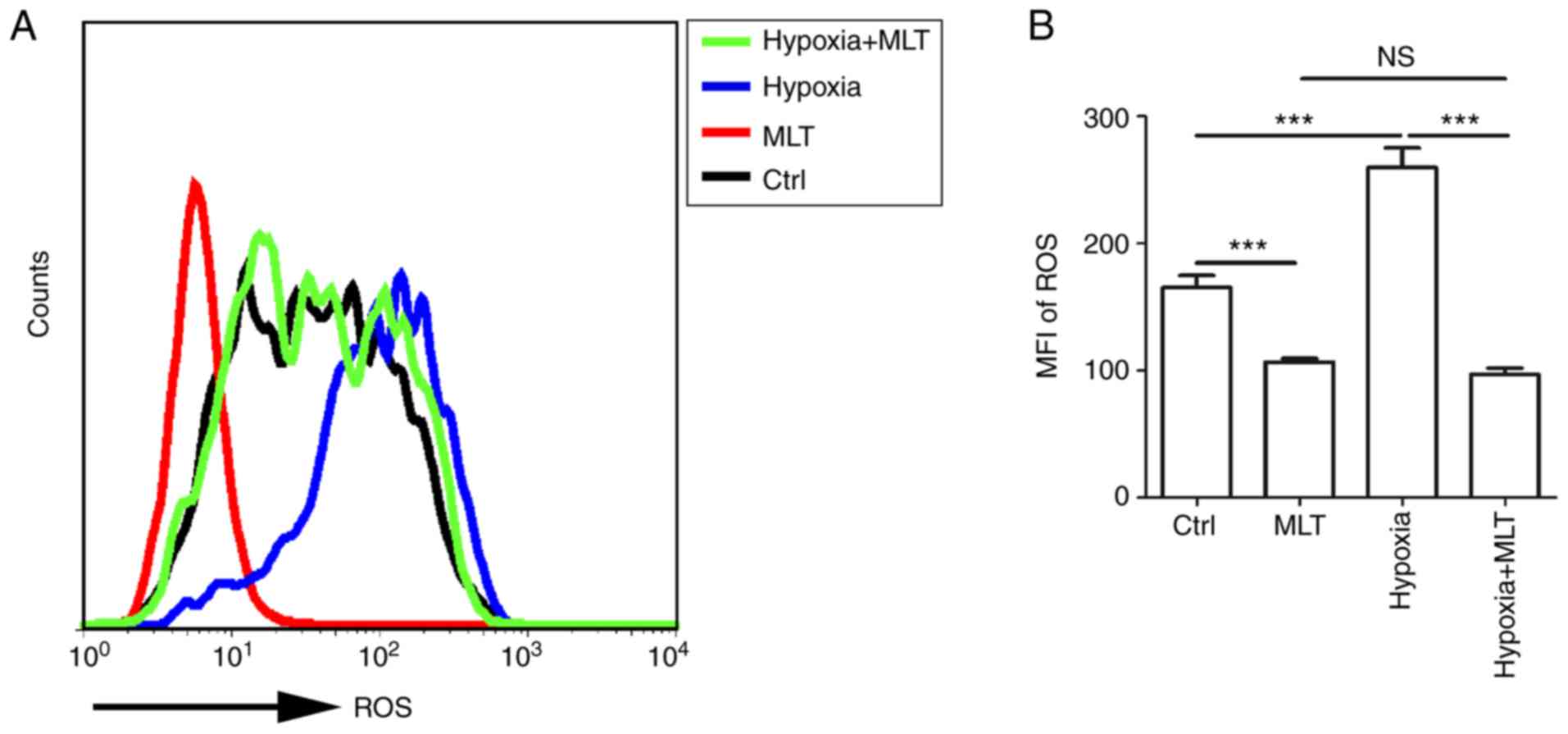
MLT suppresses the release of ROS,
particularly under the condition of hypoxia
ROS can act as signaling molecules in a variety of
cellular processes, including regulating gene transcription, cell
growth, differentiation, apoptosis and metabolism (12,44). Furthermore, emerging evidence
indicated that ROS acts as a key element in stabilizing HIF-1α. To
gain an insight into the association between hypoxia and ROS,
HUVECs were treated with or without MLT under the condition of
normoxia or hypoxia. MLT markedly suppressed the release of ROS by
HUVECs (Fig. 4A and B;
P<0.001, compared with the Ctrl group). ROS release was
increased under the condition of hypoxia, and this effect was
reversed by MLT (Fig. 4A and B;
P<0.001 for hypoxia vs. hypoxia+MLT).
Positive feedback between ROS and VEGF in
HUVECs
To identify the association between ROS and VEGF,
HUVECs were stimulated with different concentrations of recombinant
human VEGF. Exogenous VEGF promoted the release of ROS in a
dose-dependent manner (Fig. 5A;
P<0.05 for Ctrl vs. 2.5 ng/ml VEGF or P<0.01 for Ctrl vs. 5
ng/ml VEGF). H2O2 is frequently applied as
the representative ROS in modeling and inducing oxidative stress,
thus, H2O2 was used as an inducer of ROS
(13,14). Treatment with
H2O2 enhanced the production of ROS to a
certain extent (Fig. 5B;
P<0.05 for Ctrl vs. 100 μM H2O2 or
Ctrl vs. 200 μM H2O2; or P<0.001
for Ctrl vs. 500 μM H2O2 or Ctrl
vs.1,000 μM H2O2). Based on its
cytotoxicity, a dose of 50 μM H2O2 was
selected for subsequent experiments. As indicated in Fig. 5C, the expression of VEGF was
significantly upregulated by H2O2 (P<0.01
compared to ctrl group).
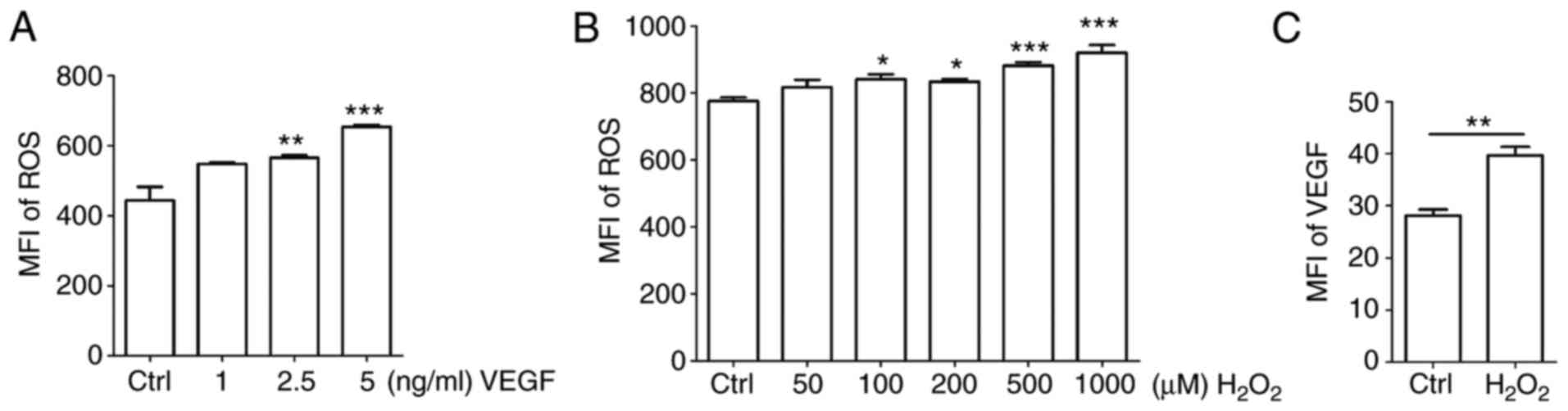 | Figure 5Positive feedback between ROS and
VEGF in HUVECs. (A and B) HUVECs were treated with (A) recombinant
human VEGF (0-5 ng/ml) or (B) H2O2 (0-1,000
μM) for 4 h, and then ROS release of cells was detected by
FCM. (C) HUVECs were treated with H2O2 (50
μM) for 4 h, and then the expression of VEGF in HUVECs was
detected by FCM. Data are presented as the mean ± standard error of
the mean, *P<0.05, **P<0.01 or
***P<0.001, compared with the Ctrl. MFI, mean
fluorescence intensity; ROS, reactive oxygen species; Ctrl,
control; FCM, flow cytometry; VEGF, vascular endothelial growth
factor; HUVECs, human umbilical vein endothelial cells. |
MLT inhibits the viability and
angiogenesis of HUVECs by disrupting the crosstalk between ROS and
VEGF
As indicated by the aforementioned results,
exogenous H2O2 stimulates the expression of
ROS and VEGF in HUVECs. HUVECs were pretreated with
H2O2 (50 μM) or VEGF (5 ng/ml) for 4
h, and then cultured with or without MLT for 2 h. It was determined
that the release of ROS and the induction of VEGF upregulated by
H2O2 were inhibited by MLT (Fig. 6A-D; P<0.01 for Ctrl vs. MLT or
P<0.001 for H2O2 vs.
MLT+H2O2). Furthermore, MLT reversed the
stimulatory effect of H2O2 on VEGF expression
in HUVECs (Fig. 6E; P<0.001
for H2O2 vs. MLT+H2O2).
To examine the effects of H2O2 treatment on
HUVECs, cell viability in response to H2O2
stimulation was detected. H2O2 was observed
to promote the viability of HUVECs (Fig. 6F; P<0.001 for Ctrl vs.
H2O2). However, treatment with additional MLT
significantly weakened this effect (Fig. 6F; P<0.01 for
H2O2 vs. MLT+H2O2).
Additionally, MLT suppressed the tube formation of HUVECs that was
induced by H2O2 or VEGF (Fig. 6G and H; P<0.05 for
H2O2 vs. MLT+H2O2 or
P<0.001 for VEGF vs. MLT+VEGF).
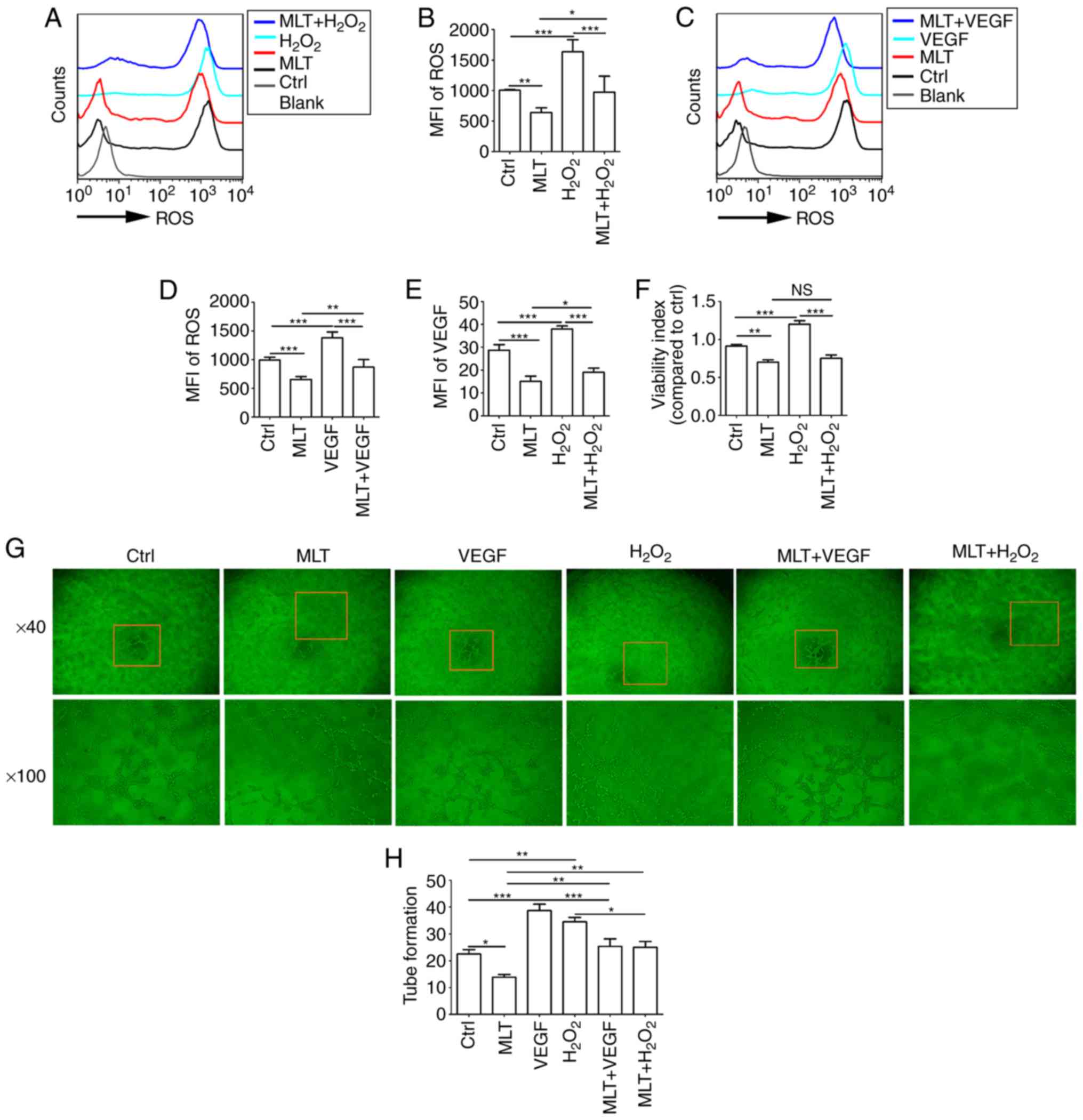 | Figure 6MLT inhibits the viability and
angiogenesis of HUVECs by disrupting the crosstalk between ROS and
VEGF. (A) HUVECs were pretreated with H2O2
(50 μM) for 4 h, followed by MLT (1×10-5 M) for 6
h, and then the release of ROS was detected by flow cytometry. (B)
The statistics histogram for (A). (C) HUVECs were pretreated with
VEGF (5 ng/ml) for 4 h, followed by MLT (1×10-5 M) for 6
h, and then the release of ROS was detected by flow cytometry. (D)
The statistics histogram for. (C) Following treatment with
H2O2 for 4 h and MLT for 6 h, (E) the
expression of VEGF and (F) viability of HUVECs was detected by flow
cytometry and a Cell Counting kit 8 assay, respectively. (G)
Following stimulation with MLT, H2O2 or VEGF
alone, or combined treatment with MLT. Tube formation of HUVECs was
measured with a tube formation assay. (H) The statistics histogram
for (G). Data are presented as the mean ± standard error of the
mean. *P<0.05, **P<0.01 or
***P<0.001. Blank, blank control (PBS treatment);
MFI, mean fluorescence intensity; ROS, reactive oxygen species;
Ctrl, control; VEGF, vascular endothelial growth factor; HUVECs,
human umbilical vein endothelial cells; Ctrl, control; MLT,
melatonin. |
MLT combined with KC7F2 severely disrupts
the growth and angiogenesis of HUVECs by targeting the
hypoxia/ROS/VEGF axis
KC7F2 has been reported to have an inhibitory effect
on the expression of HIF-1α. KC7F2 decreased cell viability in a
dose-dependent manner (Fig. 7A;
P<0.01 for Ctrl vs. 20 μM KC7F2 or P<0.001 for Ctrl
vs. 200 μM KC7F2, compared with the Ctrl group). It was also
identified that a dose of 20 μM KC7F2 suppressed the release
of ROS (Fig. 7B; P<0.001 for
Ctrl vs. KC7F2), production of VEGF (Fig. 7C; P<0.001 for Ctrl vs. KC7F2),
cell viability (Fig. 7D;
P<0.01 for Ctrl vs. KC7F2) and tube formation (Fig. 7E and F; P<0.01 for Ctrl vs.
KC7F2) in HUVECs. Combined treatment with KC7F2 and MLT resulted in
reduced ROS release, VEGF production, cell viability and
angiogenesis of HUVECs, compared with KC7F2 alone (Fig. 7B-F; P<0.05 for KC7F2 vs.
MLT+KC7F2 or P<0.01 for KC7F2 vs. MLT+KC7F2). These results
indicate that MLT combined with KC7F2 suppresses the growth and
angiogenesis of HUVECs by targeting the hypoxia/ROS/VEGF axis.
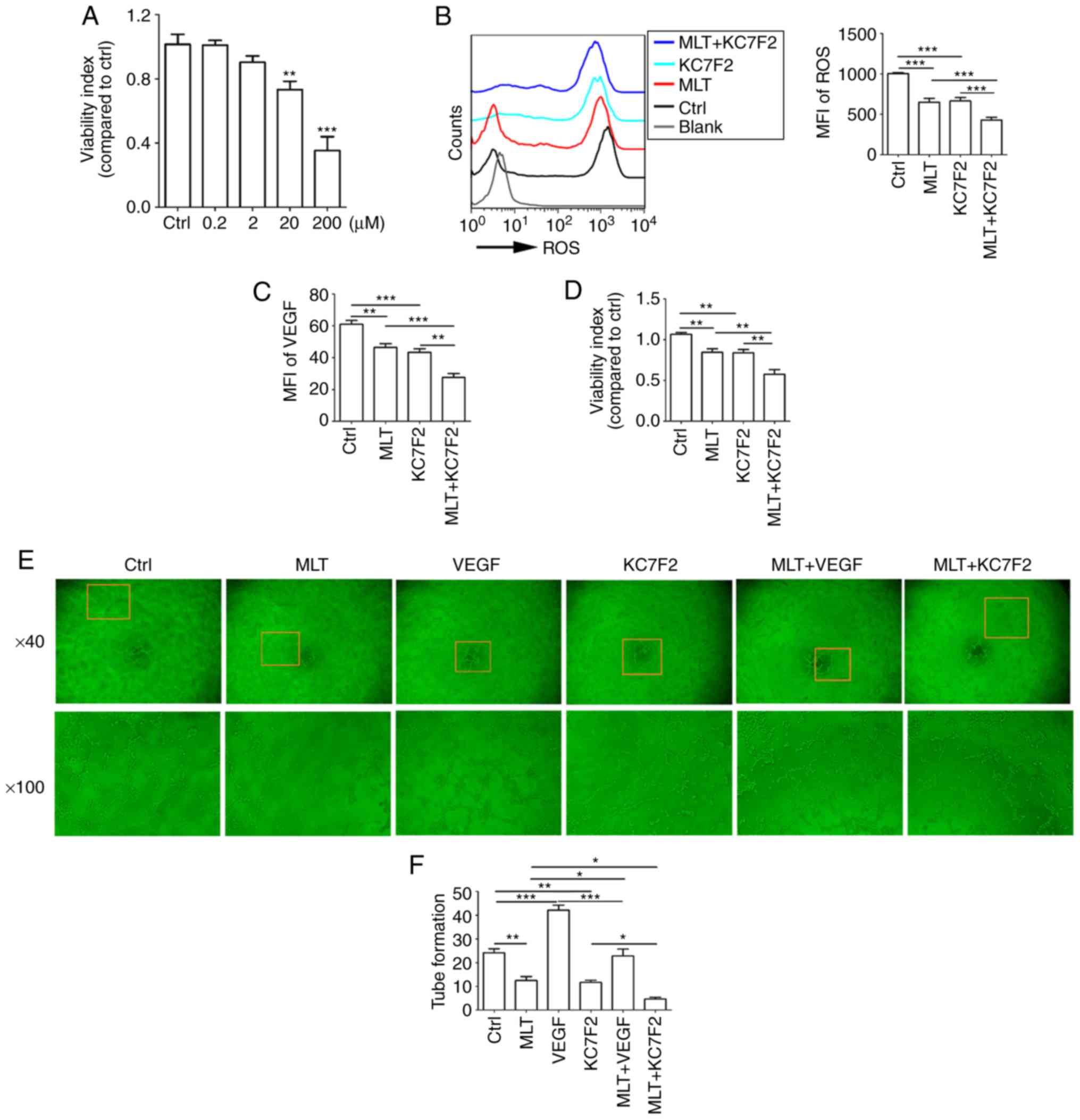 | Figure 7MLT combined with KC7F2 severely
disrupts the growth and angiogenesis of HUVECs by targeting the
hypoxia/ROS/VEGF axis. (A) HUVECs were treated with KC7F2 (0, 0.2,
2, 20 or 200 μM) for 4 h, and then cell viability was
analyzed with a CCK8 assay. HUVECs were pretreated with KC7F2 (20
μM) or VEGF (5 ng/ml) for 4 h, then treated with MLT
(1×10-5 M) for 6 h. (B) The release of ROS, (C) the
expression of VEGF, (D) cell viability and (E) tube formation were
analyzed by (B and C) flow cytometry, (D) CCK8 and (E) tube
formation assays (E). (F) The statistics histogram of tube
formation. Data are presented as the mean ± standard error of the
mean. *P<0.05, **P<0.01 or
***P<0.001. Blank, blank control (PBS treatment);
MFI, mean fluorescence intensity; ROS, reactive oxygen species;
CCK8, Cell Counting kit 8; Ctrl, control; VEGF, vascular
endothelial growth factor; HUVECs, human umbilical vein endothelial
cells; MLT, melatonin. |
Discussion
Numerous studies in recent years indicated that MLT
markedly inhibits the proliferation and metastasis of cancer cells
(26,27,30,32,36), and therefore could suppress the
progression and development of cancer, reduce the risk of mortality
and adverse events, and even improve prognosis and quality of life.
Tumor growth, development and metastasis are associated with
angiogenesis, particularly for solid tumor types (2,6,8,36,45,46). Endothelial cells in the tumor
microenvironment serve a pivotal role in the development and
progression of cancer through modulating angiogenesis (8,45).
Previous studies demonstrated that pharmacological concentrations
of MLT have a direct anti-angiogenic effect though the suppression
of proliferation in vascular endothelial cells, as well as an
indirect effect via inhibition of pro-angiogenesis cytokines,
including VEGF, epidermal growth factor and insulin-like growth
factor (37,41,47). VEGF is one of the most potent
pro-angiogenic cytokines, which specifically triggers the
proliferation of endothelial cells and increases permeability
(48). However, the underlying
mechanisms of this remain poorly elucidated. Previous studies in
this area have only been preliminary and the effect of MLT on VEGF
expression and the underlying mechanisms remain ambiguous (37,41,47). In the present study, MLT markedly
restrained viability and disrupted tube formation in HUVECs, and
this effect was dependent on VEGF. Additionally, 1×10-5
M MLT decreased the cell viability. However, 1×10-3 M
MLT significantly suppressed cell viability to 30%, and these
HUVECs would be dead. Therefore, in the present study
1×10-5 M was selected in the subsequent trials, and the
effect of MLT on the apoptosis and death of HUVECs should be
studied further.
Tumor angiogenesis is a key process for delivering
oxygen and nutrients to growing tumors, and therefore is considered
an essential characteristic of the microenvironment in solid tumor
types (49). The production of
certain angiogenic cytokines and growth factors is regulated by
hypoxia, while tumor angiogenesis and subsequent rapid tumor growth
also further accelerate tumor hypoxia (37,38). This feedback mechanism is a
critical factor resulting in pathogenic features of cancer,
including poor treatment prognosis, and progression to malignancies
and metastatic disease (48). In
the present study, it was determined that MLT could inhibit
hypoxia-induced HIF-1α expression in HUVECs at mRNA level and
protein level. However, the role of MLT on HIF1A at
transcriptional level in nuclear extracts and the detailed
mechanism require further study. Additionally, it was observed that
hypoxia stimulated VEGF production. Inhibitor of HIF-1α
significantly downregulated VEGF expression. However, whether the
regulation of hypoxia on VEGF depends on the specific
transcriptional activation of HIF1A on VEGF remains to be
further studied.
Chronic hypoxia is the primary cause of high
concentration ROS formation within tumor cells (50). ROS, which are produced by numerous
physiological oxidative activities and stress reactions in the
body, are also associated with numerous physiological, including
cell autophagy and immunoregulation, and pathophysiological,
including tumor progression, processes (51-54). At low levels, ROS exhibit
beneficial effects, serving as signaling molecules by regulating
intracellular signals and maintaining homeostasis, including in the
processes of proliferation, differentiation, adhesion, migration,
invasion and apoptosis; however, at high levels, ROS are
deleterious to cells, resulting in damage to proteins, lipids and
DNA, and eventually resulting in autophagy and programmed cell
death (55,56). An increased level of ROS has been
demonstrated to promote cell proliferation, cell migration, cell
survival and epithelial-mesenchymal transition through activating
the mitogen-activated protein kinase and Ras-extracellular
signal-regulated kinases pathway (57,58), finally resulting in oxidative
stress, which initiates carcinogenesis (59). Previous evidence indicates that
ROS may act as second messengers in response to hypoxia, and serve
an important role in stabilizing HIF-1α protein and inducing the
production of angiogenic factors. Furthermore, chemical
antioxidants suppress HIF-1α accumulation and inhibit the
transcription of VEGF via a mechanism that involves ROS (55).
In the present study, it was determined that
hypoxia enhances the release of ROS and VEGF expression.
Additionally, there is a positive feedback mechanism between ROS
and VEGF, which contributes to the high viability and angiogenesis
of HUVECs. Notably, MLT could effectively suppress these effects,
particularly in combination with KC7F2 (a novel small molecule
HIF-1α translation inhibitor). Collectively, these results indicate
that MLT inhibits the viability and angiogenesis of HUVECs by
targeting the HIF-1α/VEGF/ROS axis. Wang et al (60) demonstrated that MLT downregulates
the MLT nuclear receptor RZR/RAR related orphan receptor (ROR)γ
expression causing growth-inhibitory and anti-angiogenesis activity
in human gastric cancer cells in vitro and in vivo.
However, whether RZR/RORγ is involved in the effect of MLT on the
HIF-1α/VEGF/ROS axis requires further investigation. Additionally,
ROS can regulate the B-cell lymphoma 2 family via direct and
indirect mechanisms (61,62), indicating that tumor growth is
also inversely associated with the level of ROS. Therefore, the
effect of KC7F2 and ROS on the apoptosis of HUVECs should be
studied further.
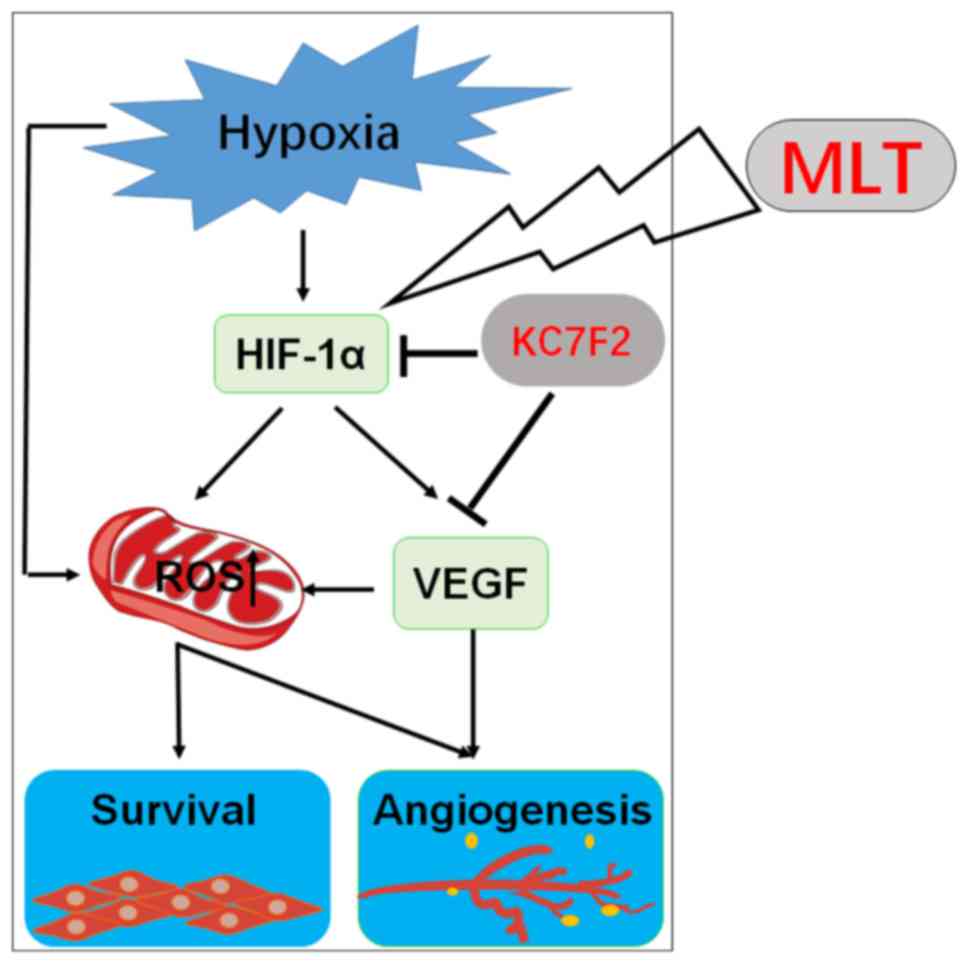
In conclusion, as depicted in Fig. 8, the present observations
demonstrate that hypoxia upregulates the level of ROS and VEGF in a
direct manner and HIF-1α-dependent manner. ROS production results
in an increase in the viability and angiogenesis of HUVECs. As an
important angiogenesis growth factor, VEGF can directly promote
angiogenesis of HUVECs. Furthermore, VEGF increases ROS production
and amplifies the stimulatory effect of ROS on the viability and
angiogenesis of HUVECs. MLT serves a dual role in the inhibition of
angiogenesis, acting directly as a growth-associated inhibitor, or
indirectly as an antioxidant and free radical scavenging agent. MLT
suppresses the viability and angiogenesis of HUVECs through
downregulating the hypoxia/HIF-1α/ROS/VEGF pathway in vitro.
As an inhibitor of HIF-1α, KC7F2 can exert an anti-angiogenesis
effect synergistically with MLT in vitro. In summary, the
present data indicate that MLT may be a potential anticancer agent
in solid tumors with abundant blood vessels, particularly in
combination with KC7F2. However, further in vivo studies and
trials are required.
Funding
The present study was supported by the Major
Research Program of National Natural Science Foundation of China
(grant nos. 91542108, 81471513, 31671200, 81571509 and 81401273),
the Shanghai Rising-Star Program (grant no. 16QA1400800), the
Development Fund of Shanghai Talents (grant no. 201557), the
Oriented Project of Science and Technology Innovation from the Key
Laboratory of Reproduction Regulation of NPFPC (grant no. CX2017-2)
and the Program for Zhuoxue of Fudan University, the Program of
Jiangsu Key Youth Medical Talents (grant no. QNRC2016244) and the
Introduction Project of Suzhou Clinical Medicine Expert Team (grant
no. SZYJTD201708).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors’ contributions
JC conducted all experiments and prepared the
figures and the manuscript. HLY, CJG, and YKL assisted with flow
cytometry analysis. JS, RZ, YYH and XYZ assisted in the study
design and critically revised the manuscript for important
intellectual content. MQL designed the study, supervised the
project and edited the manuscript. All the authors were involved in
writing the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ushio-Fukai M and Alexander RW: Reactive
oxygen species as mediators of angiogenesis signaling-Role of
NAD(P)H oxidase. Mol Cell Biochem. 264:85–97. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gacche RN and Meshram RJ: Targeting tumor
micro-environment for design and development of novel
anti-angiogenic agents arresting tumor growth. Prog Biophys Mol
Biol. 113:333–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park JH, Yoon J and Park B: Pomolic acid
suppresses HIF1 α/VEGF-mediated angiogenesis by targeting p38-MAPK
and mTOR signaling cascades. Phytomedicine. 23:1716–1726. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ushio-Fukai M and Nakamura Y: Reactive
oxygen species and angiogenesis: NADPH oxidase as target for cancer
therapy. Cancer Lett. 266:37–52. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Frezzetti D, Gallo M, Maiello MR,
D’Alessio A, Esposito C, Chicchinelli N, Normanno N and De Luca A:
VEGF as a potential target in lung cancer. Expert Opin Ther
Targets. 21:959–966. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Z, Dabrosin C, Yin X, Fuster MM,
Arreola A, Rathmell WK, Generali D, Nagaraju GP, El-Rayes B,
Ribatti D, et al: Broad targeting of angiogenesis for cancer
prevention and therapy. Semin Cancer Biol. 35(Suppl): S224–S243.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee SH, Jeong D, Han YS and Baek MJ:
Pivotal role of vascular endothelial growth factor pathway in tumor
angiogenesis. Ann Surg Treat Res. 89:1–8. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Delli Carpini J, Karam AK and Montgomery
L: Vascular endothelial growth factor and its relationship to the
prognosis and treatment of breast, ovarian, and cervical cancer.
Angiogenesis. 13:43–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park WH: Effects of antioxidants and MAPK
inhibitors on cell death and reactive oxygen species levels in
H2O2-treated human pulmonary fibroblasts. Oncol Lett. 5:1633–1638.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blaser H, Dostert C, Mak TW and Brenner D:
TNF and ROS crosstalk in inflammation. Trends Cell Biol.
26:249–261. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tapeinos C, Larranaga A, Sarasua JR and
Pandit A: Functionalised collagen spheres reduce H2O2 mediated
apoptosis by scavenging overexpressed ROS. Nanomedicine.
14:2397–2405. 2018. View Article : Google Scholar
|
|
13
|
Chandel NS, Maltepe E, Goldwasser E,
Mathieu CE, Simon MC and Schumacker PT: Mitochondrial reactive
oxygen species trigger hypoxia-induced transcription. Proc Natl
Acad Sci USA. 95:11715–11720. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pearlstein DP, Ali MH, Mungai PT, Hynes
KL, Gewertz BL and Schumacker PT: Role of mitochondrial oxidant
generation in endothelial cell responses to hypoxia. Arterioscler
Thromb Vasc Biol. 22:566–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maraldi T, Prata C, Caliceti C, Vieceli
Dalla, Sega F, Zambonin L, Fiorentini D and Hakim G: VEGF-induced
ROS generation from NAD(P)H oxidases protects human leukemic cells
from apoptosis. Int J Oncol. 36:1581–1589. 2010.PubMed/NCBI
|
|
16
|
Colavitti R, Pani G, Bedogni B, Anzevino
R, Borrello S, Waltenberger J and Galeotti T: Reactive oxygen
species as downstream mediators of angiogenic signaling by vascular
endothelial growth factor receptor-2/KDR. J Biol Chem.
277:3101–3108. 2002. View Article : Google Scholar
|
|
17
|
Oshikawa J, Kim SJ, Furuta E, Caliceti C,
Chen GF, McKinney RD, Kuhr F, Levitan I, Fukai T and Ushio-Fukai M:
Novel role of p66Shc in ROS-dependent VEGF signaling and
angiogenesis in endothelial cells. Am J Physiol Heart Circ Physiol.
302:H724–H732. 2012. View Article : Google Scholar :
|
|
18
|
Ushio-Fukai M: VEGF signaling through
NADPH oxidase-derived ROS. Antioxid Redox Signal. 9:731–739. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen S, Zhou Y, Zhou L, Guan Y, Zhang Y
and Han X: Anti-neovascularization effects of DMBT in age-related
macular degeneration by inhibition of VEGF secretion through
ROS-dependent signaling pathway. Mol Cell Biochem. 448:225–235.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jing Y, Liu LZ, Jiang Y, Zhu Y, Guo NL,
Barnett J, Rojanasakul Y, Agani F and Jiang BH: Cadmium increases
HIF-1 and VEGF expression through ROS, ERK, and AKT signaling
pathways and induces malignant transformation of human bronchial
epithelial cells. Toxicol Sci. 125:10–19. 2012. View Article : Google Scholar
|
|
21
|
Narita T, Yin S, Gelin CF, Moreno CS,
Yepes M, Nicolaou KC and Van Meir EG: Identification of a novel
small molecule HIF-1alpha translation inhibitor. Clin Cancer Res.
15:6128–6136. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guan G, Zhang Y, Lu Y, Liu L, Shi D, Wen
Y, Yang L, Ma Q, Liu T, Zhu X, et al: The HIF-1α/CXCR4 pathway
supports hypoxia-induced metastasis of human osteosarcoma cells.
Cancer Lett. 357:254–264. 2015. View Article : Google Scholar
|
|
23
|
Zhou H, Chen J, Lu X, Shen C, Zeng J, Chen
L and Pei Z: Melatonin protects against rotenone-induced cell
injury via inhibition of Omi and Bax-mediated autophagy in Hela
cells. J Pineal Res. 52:120–127. 2012. View Article : Google Scholar
|
|
24
|
Goradel NH, Asghari MH, Moloudizargari M,
Negahdari B, Haghi-Aminjan H and Abdollahi M: Melatonin as an
angiogenesis inhibitor to combat cancer: Mechanistic evidence.
Toxicol Appl Pharmacol. 335:56–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Su SC, Hsieh MJ, Yang WE, Chung WH, Reiter
RJ and Yang SF: Cancer metastasis: Mechanisms of inhibition by
melatonin. J Pineal Res. 62:e123702017. View Article : Google Scholar
|
|
26
|
Reiter RJ, Rosales-Corral SA, Tan DX,
Acuna-Castroviejo D, Qin L, Yang SF and Xu K: Melatonin, a full
service anti-cancer agent: Inhibition of initiation, progression
and metastasis. Int J Mol Sci. 18:E8432017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carbajo-Pescador S, Ordonez R, Benet M,
Jover R, García-Palomo A, Mauriz JL and González-Gallego J:
Inhibition of VEGF expression through blockade of Hif1alpha and
STAT3 signalling mediates the anti-angiogenic effect of melatonin
in HepG2 liver cancer cells. Br J Cancer. 109:83–91. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang TH, Wu CH, Yeh CT, Su SC, Hsia SM,
Liang KH, Chen CC, Hsueh C and Chen CY: Melatonin suppresses
hepato-cellular carcinoma progression via lncRNA-CPS1-IT-mediated
HIF-1 α inactivation. Oncotarget. 8:82280–82293. 2017.PubMed/NCBI
|
|
29
|
Xu Pi H, Reiter S, Guo RJ, Zhang P, Li L,
Li Y, Cao M, Tian Z, Xie LJ, et al: SIRT3-SOD2-mROS-dependent
autophagy in cadmium-induced hepatotoxicity and salvage by
melatonin. Autophagy. 11:1037–1051. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma Z, Yang Y, Fan C, Han J, Wang D, Di S,
Hu W, Liu D, Li X, Reiter RJ and Yan X: Melatonin as a potential
anticarcinogen for non-small-cell lung cancer. Oncotarget.
7:46768–46784. 2016.PubMed/NCBI
|
|
31
|
Plaimee P, Weerapreeyakul N, Barusrux S
and Johns NP: Melatonin potentiates cisplatin-induced apoptosis and
cell cycle arrest in human lung adenocarcinoma cells. Cell Prolif.
48:67–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Proietti S, Cucina A, Reiter RJ and
Bizzarri M: Molecular mechanisms of melatonin’s inhibitory actions
on breast cancers. Cell Mol Life Sci. 70:2139–2157. 2013.
View Article : Google Scholar
|
|
33
|
Proietti S, Catizone A, Masiello MG,
Dinicola S, Fabrizi G, Minini M, Ricci G, Verna R, Reiter RJ,
Cucina A and Bizzarri M: Increase in motility and invasiveness of
MCF7 cancer cells induced by nicotine is abolished by melatonin
through inhibition of ERK phosphorylation. J Pineal Res.
64:e124672018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gonzalez-Gonzalez A, Gonzalez A,
Alonso-Gonzalez C, Menéndez-Menéndez J, Martínez-Campa C and Cos S:
Complementary actions of melatonin on angiogenic factors, the
angiopoietin/Tie2 axis and VEGF, in co-cultures of human
endothelial and breast cancer cells. Oncol Rep. 39:433–441.
2018.
|
|
35
|
Lee H, Lee HJ, Jung JH, Shin EA and Kim
SH: Melatonin disturbs SUMOylation-mediated crosstalk between c-Myc
and nestin via MT1 activation and promotes the sensitivity of
paclitaxel in brain cancer stem cells. J Pineal Res. 65:e124962018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hill SM, Frasch T, Xiang S, Yuan L,
Duplessis T and Mao L: Molecular mechanisms of melatonin anticancer
effects. Integr Cancer Ther. 8:337–346. 2009. View Article : Google Scholar
|
|
37
|
Lai YH, Hu DN, Rosen R, Sassoon J, Chuang
LY, Wu KY and Wu WC: Hypoxia-induced vascular endothelial growth
factor secretion by retinal pigment epithelial cells is inhibited
by melatonin via decreased accumulation of hypoxia-inducible
factors-1α protein. Clin Exp Ophthalmol. 45:182–191. 2017.
View Article : Google Scholar
|
|
38
|
Park SY, Jang WJ, Yi EY, Jang JY, Jung Y,
Jeong JW and Kim YJ: Melatonin suppresses tumor angiogenesis by
inhibiting HIF-1 alpha stabilization under hypoxia. J Pineal Res.
48:178–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park JW, Hwang MS, Suh SI and Baek WK:
Melatonin downregulates HIF-1 alpha expression through inhibition
of protein translation in prostate cancer cells. J Pineal Res.
46:415–421. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cui P, Luo Z, Zhang H, Su Y, Li A, Li H,
Zhang J, Yang Z and Xiu R: Effect and mechanism of melatonin’s
action on the proliferation of human umbilical vein endothelial
cells. J Pineal Res. 41:358–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
González A, González-González A,
Alonso-González C, Menéndez-Menéndez J, Martínez-Campa C and Cos S:
Melatonin inhibits angiogenesis in SH-SY5Y human neuroblastoma
cells by downregulation of VEGF. Oncol Rep. 37:2433–2440. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vriend J and Reiter RJ: Melatonin and the
von Hippel-Lindau/HIF-1 oxygen sensing mechanism: A review. Biochim
Biophys Acta. 1865:176–183. 2016.PubMed/NCBI
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
44
|
Pan B, Zhong W, Deng Z, Lai C, Chu J, Jiao
G, Liu J and Zhou Q: Inhibition of prostate cancer growth by
solanine requires the suppression of cell cycle proteins and the
activation of ROS/P38 signaling pathway. Cancer Med. 5:3214–3222.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Alvarez-García V, González A,
Alonso-González C, Martínez-Campa C and Cos S: Antiangiogenic
effects of melatonin in endothelial cell cultures. Microvasc Res.
87:25–33. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Vijayalaxmi, Thomas CR Jr, Reiter RJ and
Herman TS: Melatonin: From basic research to cancer treatment
clinics. J Clin Oncol. 20:2575–2601. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dai M, Cui P, Yu M, Han J, Li H and Xiu R:
Melatonin modulates the expression of VEGF and HIF-1 alpha induced
by CoCl2 in cultured cancer cells. J Pineal Res. 44:121–126. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Colombo J, Maciel JM, Ferreira LC, Da
Silva RF and Zuccari DA: Effects of melatonin on HIF-1 α and VEGF
expression and on the invasive properties of hepatocarcinoma cells.
Oncol Lett. 12:231–237. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Y, Liu Q, Wang F, Ling EA, Liu S,
Wang L, Yang Y, Yao L, Chen X, Wang F, et al: Melatonin antagonizes
hypoxia-mediated glioblastoma cell migration and invasion via
inhibition of HIF-1 α. J Pineal Res. 55:21–130. 2013. View Article : Google Scholar
|
|
50
|
Guzy RD and Humacker PT: Oxygen sensing by
mitochondria at complex III: The paradox of increased reactive
oxygen species during hypoxia. Exp Physiol. 91:807–819. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Scherz-Shouval R and Elazar Z: Regulation
of autophagy by ROS: Physiology and pathology. Trends Biochem Sci.
36:30–38. 2011. View Article : Google Scholar
|
|
52
|
D’Autréaux B and Toledano MB: ROS as
signalling molecules: Mechanisms that generate specificity in ROS
homeostasis. Nat Rev Mol Cell Biol. 8:813–824. 2007. View Article : Google Scholar
|
|
53
|
Zhang Y, Choksi S, Chen K, Pobezinskaya Y,
Linnoila I and Liu ZG: ROS play a critical role in the
differentiation of alternatively activated macrophages and the
occurrence of tumor-associated macrophages. Cell Res. 23:898–914.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Diebold L and Chandel NS: Mitochondrial
ROS regulation of proliferating cells. Free Radic Biol Med.
100:86–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xia C, Meng Q, Lin LZ, Rojanasakul Y, Wang
XR and Jiang BH: Reactive oxygen species regulate angiogenesis and
tumor growth through vascular endothelial growth factor. Cancer
Res. 67:10823–10830. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li Z, Xu X, Leng X, He M, Wang J, Cheng S
and Wu H: Roles of reactive oxygen species in cell signaling
pathways and immune responses to viral infections. Arch Virol.
162:603–610. 2017. View Article : Google Scholar
|
|
57
|
Morgan MJ and Liu ZG: Crosstalk of
reactive oxygen species and NF-κB signaling. Cell Res. 21:103–115.
2011. View Article : Google Scholar
|
|
58
|
Liu J, Chang F, Li F, Fu H, Wang J, Zhang
S, Zhao J and Yin D: Palmitate promotes autophagy and apoptosis
through ROS-dependent JNK and p38 MAPK. Biochem Biophys Res Commun.
463:262–267. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Prasad S, Gupta SC and Tyagi AK: Reactive
oxygen species (ROS) and cancer: Role of antioxidative
nutraceuticals. Cancer Lett. 387:95–105. 2017. View Article : Google Scholar
|
|
60
|
Wang RX, Liu H, Xu L, Zhang H and Zhou RX:
Melatonin downregulates nuclear receptor RZR/RORγ expression
causing growth-inhibitory and anti-angiogenesis activity in human
gastric cancer cells in vitro an in vivo. Oncol Lett. 12:897–903.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
You BR, Shin HR and Park WH: PX-12
inhibits the growth of A549 lung cancer cells via G2/M phase arrest
and ROS-dependent apoptosis. Int J Oncol. 44:301–308. 2014.
View Article : Google Scholar
|
|
62
|
Bauer G: Central signaling elements of
intercellular reactive oxygen/nitrogen species-dependent induction
of apoptosis in malignant cells. Anticancer Res. 37:499–513. 2017.
View Article : Google Scholar : PubMed/NCBI
|