Introduction
Gastric cancer is a common form of malignant tumor
in gastrointestinal tract cancer. In recent years, gastric cancer
ranks as the third highest for mortality rate and fourth highest
for morbidity rate of all types of cancer in the world (1). In less developed countries in
particular, it is the third most frequently diagnosed cancer in men
and the leading cause of cancer-associated mortality (2). Although a decreasing trend in
gastric cancer incidence and mortality rates has been observed in
several countries, it is not effectively treated currently due to
its complex etiology (3). Surgery
is the optimal treatment approach for gastric cancer at present,
however, the 5-year-survival rate remains low. The majority of
patients relapse following surgery (4). Systemic and partial chemotherapy is
necessary for patients with advanced gastric cancer (5). However, gastric cancer cells show
high drug resistance to most common chemotherapeutic agents
(6,7). Consequently, identifying novel
effective anticancer drugs is crucial for cancer therapy.
Corilagin
[β-1-O-galloyl-3,6-(R)-hexahydroxydiphenoyl-D-glucose] is a unique
component of the tannin family. The molecular weight of corilagin
is 634.45 (8). Corilagin has been
identified in several medicinal plants, including Longan (9), Lumnitzera racemose (10), Terminalia catappa L
(11) and Phyllanthus
species (12). Previous studies
have shown that corilagin has extensive pharmacological actions,
including anti-inflammatory (13), antioxidative (11), antiviral (12), hepatoprotective (10), antiatherogenic (8) and antitumor activities, and low
adverse effects. A study by Guo et al (14) demonstrated that corilagin can
protect against herpes simplex virus-1 (HSV-1) encephalitis through
inhibiting the Toll-like receptor (TLR)2 signaling pathways. In
their study, it was found that corilagin markedly prevented an
increase in the levels of TLR2 and its downstream mediators
following HSV-1 challenge. In addition, it was shown that corilagin
directly inhibited inflammatory cytokines, including tumor necrosis
factor (TNF)-α and interleukin (IL)-6 proteins. The effect of
corilagin on hepatoprotective properties has been reported; the
underlying hepatoprotective mechanism of corilagin was examined in
a trauma-hemorrhagic shock rodent model and it was found that the
drug markedly alleviated pro-inflammatory cytokine and neutrophil
accumulation via the AKT pathway (15). Similarly, Du et al
(16) indicated that corilagin
effectively relieved hepatic fibrosis by inhibiting the expression
of molecules associated with the IL-13/signal transducer and
activator of transcription 6 signaling pathway. Furthermore,
studies have confirmed that corilagin has notable antitumor effects
on a number of tumor cells, including hepatoma (17), ovarian cancer (18), cholangiocarcinoma (19) and glioblastoma (20). Studies have shown that corilagin
can markedly inhibit the growth of ovarian cancer cells in
vitro and in vivo by increasing cell cycle arrest at the
G2/M stage, enhancing apoptosis and inhibiting the TGF-β signaling
pathways (18,21,22). However, the mechanism involved has
not been fully elucidated in gastric cancer. Therefore, the present
study was designed to investigate the effect of corilagin on the
apoptosis, autophagy and necroptosis of SGC7901 and BGC823 human
gastric cancer cells.
Cell apoptosis, controlled by a large number of
genes, acts as one of the most vital processes in the regulation of
carcinogenesis (23). It has been
well documented that signaling pathways leading to apoptosis
involve the sequential activation of cysteine proteases, known as
caspases (24). In the initial
step of the apoptotic process, it triggers the activation of an
apoptotic signaling program, which leads to cell death rather than
killing the cell directly (25).
Autophagy, commonly referred to as ‘self-eating’, is
sensitized by various types of intracellular stress, for example,
DNA damage and low nutrient levels. Autophagy is mostly a
protective process involving the capture and digestion of cellular
constituents within lysosomes. However, the hyperactivation of
autophagy can cause autophagic cell death (26).
Necroptosis is a more recently described form of
programmed cell death, which differs from apoptosis and has similar
morphological characteristics to necrosis, including cell swelling,
rupture of the plasma membrane and condensation of the chromatin.
In recent years, necroptosis has attracted wide attention due to
its specific function in physiological and pathological processes.
Receptor interaction protein 3 (RIP3), a serine/threonine kinase,
is required for activation of the necrotic cell death pathway.
However, RIP3 deficiency has been found in the majority of cancer
cell lines. Therefore, RIP3 may be important in cancer progression
(27,28).
Reactive oxygen species (ROS), a cellular
metabolite, is important in the development of cancer (29). Oxidative stress is an imbalance
between ROS and the antioxidant defense system. Excessive ROS
production at certain levels act as signal molecules to stimulate
cell apoptosis and DNA damage (30). Accordingly, it is recognized that
ROS are involved in antitumor function.
In the present study, the effects of
corilagin-induced growth inhibition and apoptosis were first
evaluated in gastric cancer cells using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenytetrazolium bromide (MTT)
assay, EdU proliferation assay, lactate dehydrogenase (LDH) release
assay, ROS generation assay, Hoechst 33342 staining detection, flow
cytometric analysis and western blot analysis. Subsequent
investigation focused on the ability of corilagin to induce
autophagy in human gastric cancer cells and whether the inhibition
of autophagy can enhance the effect of corilagin. Finally,
experiments were performed to investigate whether necroptosis
occurs in gastric cancer cells following corilagin treatment.
Materials and methods
Reagents and antibodies
Corilagin with a purity of 99.36% was purchased from
Qingyun Biology (Nanjing, China). Acridine orange (AO), Earle's
balanced salt solution (EBSS), dimethyl sulfoxide (DMSO),
N-acetyl-L-cysteine (NAC), MTT, anti-β-actin antibody (cat. no.
A1978) and Hoechst 33342 were purchased from Sigma-Aldrich; Merck
KGaA (Darmstadt, Germany). Fetal bovine serum (FBS), horseradish
peroxidase-conjugated goat anti-rabbit antibody (cat. no. A24537),
RPMI-1640 medium and Dulbecco's modified Eagle's medium (DMEM) were
purchased from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). Penicillin-streptomycin was obtained from PAA; GE Healthcare
(Chicago, IL, USA). Chloroquine (CQ) and necrosulfonamide (NSA)
were purchased from Tocris Biosciences (Bristol, UK). Pancreatin
was purchased from Solarbio Science & Technology Co., Ltd.
(Beijing, China). Z-VAD-FMK and necrostatin-1 (Nec-1) were obtained
from Selleck Chemicals (Houston, TX, USA). Anti-LC3 antibody (cat.
no. NB100-2220) was purchased from Novus Biologicals, LLC
(Littleton, CO, USA) and anti-RIP3 antibody (cat. no. GTX107574)
was obtained from Genetex Inc. (Irvine, CA, USA). The lactate
dehydrogenase (LDH) cyto-toxicity assay kit, anti-caspase-3
antibody (cat. no. AC030), anti-caspase-8 antibody (cat. no.
AC056), anti-caspase-9 antibody (cat. no. AC062) and anti-poly
ADP-ribose polymerase (PARP) antibody (cat. no. AP102) were
purchased from Beyotime Institute of Biotechnology (Haimen, China).
The Annexin V-FITC/propidium iodide (PI) apoptosis detection kit
and ROS detection kit were obtained from Nanjing KeyGen Biotech
Co., Ltd. (Nanjing, China). All other chemicals were of analytical
grade available from commercial sources.
Cell culture
The SGC7901 and BGC823 human gastric cancer cell
lines were obtained from the Chinese Academy of Sciences (Shanghai,
China). The GES-1 human gastric mucosal epithelial cell line, HeLa
human cervical cancer cell line and HT-29 human colorectal
carcinoma cell line were donated by the School of Life Sciences,
Xiamen University (Xiamen, China). The HT-29, SGC7901 and BGC823
cells were maintained in RPMI-1640 medium, the HeLa and GES-1 cells
were cultured in DMEM. The media were supplemented with 10% FBS and
100 U/ml penicillin/streptomycin. The cells were cultured at 37°C
in a humidified atmosphere of 95% air and 5% CO2.
Cell viability assay
The cell growth was measured using an MTT assay. The
cells were seeded into 96-well plates at a density of
1×104 cells/well. After 24 h of incubation, the cells
were treated with corilagin at serial concentrations (0, 10, 20,
30, 40 and 50 µM) for 24 h. Furthermore, for synergistic
effect analysis, corilagin (30 µM) combined with Z-VAD-FMK
(CQ, NAC, NSA or Nec-1) were added to the cells and incubated for
24 h. At the end of treatment, 20 µl of MTT was added to
each well (final concentration: 0.5 mg/ml) and incubated for 4 h.
The medium was then replaced with 150 µl of DMSO. The
absorbance was measured with a microplate reader at a wavelength of
490 nm.
EdU proliferation assay
The cells (density, 1×104 cells/well)
were seeded into a 96-well plate and then treated with corilagin
for 24 h. Cell proliferation was assessed using the EdU
proliferation assay kit (Guangzhou RiboBio Co., Ltd., Guangzhou,
China). The stained cells were observed under a fluorescence
microscope (Olympus IX51; Olympus Corporation, Tokyo, Japan).
Cell morphological analysis using Hoechst
33342 staining
The cells (density, 1.5×105 cells/well)
were cultured in 12-well plates and treated with corilagin (0, 10,
20 and 30 µM) for 24 h. Following treatment, the cells were
washed twice with PBS and fixed with 4% paraformaldehyde
(Sigma-Aldrich; Merck KGaA) in PBS for 10 min at room temperature.
The cells were then washed twice with PBS and stained with 10
µg/ml Hoechst 33342 for 15 min at 37°C. The morphological
changes were detected using a fluorescence microscope (Olympus
Corporation). This assay was performed following the procedures as
described by Lee et al (31).
LDH release assay
The cells (density, 1×104 cells/well)
were seeded into 96-well plates and then treated with corilagin at
different concentrations (0, 10, 20 and 30 µM). The LDH
assay was then performed according to the steps described by the
manufacturer's protocol for the LDH cytotoxicity assay kit
(Beyotime Institute of Biotechnology).
Annexin V-FITC/PI double staining
apoptosis detection
The Annexin V-FITC/PI kit was used to detect the
apoptosis of cells by flow cytometry. Briefly, the cells were
cultured and treated with corilagin (0, 10, 20 and 30 µM).
After 24 h, the cells were collected and harvested. The cells were
then washed with cold PBS, adjusted to 1×106 cells/ml in
1X binding buffer and stained with Annexin V-FITC and PI solution
for 15 min at room temperature in the dark. Finally, the stained
cells were analyzed by flow cytometry (Beckman Coulter, Inc., Brea,
CA, USA).
Acridine orange (AO) staining
analysis
The SGC7901 and BGC823 cells were collected and
plated into 12-well plates. After 12 h of incubation, the cells
were treated with corilagin for 24 h. The cells were then stained
with 1 µM AO at 37°C for 15 min. Finally, the acidic
autophagic vacuoles were visualized using a fluorescence
microscope.
Detection of ROS generation assay
The ROS generation assay was performed as previously
described (32). The SGC7901 and
BGC823 cells were exposed to various concentrations (0, 10, 20 and
30 µM) of corilagin with or without NAC for 24 h, washed
with PBS, and resuspended in culture medium (without serum)
containing 10 µM DCFH-DA. As positive control, the cells
were treated with Rosup (50 µg/ml) for 30 min. Subsequently,
ROS production was detected by flow cytometry.
Reverse transcription-polymerase chain
reaction (RT-PCR) assay
Total RNA from the gastric cancer cells was
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), following which cDNA was synthesized with 1
µg of total RNA at 37°C for 15 min using the Primescript RT
reagent kit (Takara Biotechnology Co., Ltd., Dalian, China). The
RT-PCR assay was performed using the Roche
LightCycler®96 Real-time PCR system with the SYBR Premix
EX Taq Ⅱ kit (Takara, Biotechnology Co., Ltd.). The reaction volume
was 20 µl, including SYBR Premix EX Taq II (10 µl),
Forward Primer (0.8 µl), Reverse Primer (0.8 µl),
cDNA (2 µl), Sterilized Purified Water (6.4 µl). The
amplification conditions were the following: Initial
pre-degeneration at 95°C for 2 min, followed by 40 cycles of
denaturation at 95°C for 10 sec and annealing/extension at 60°C for
20 sec. The relative mRNA levels were determined with the
2−ΔΔCq method (33).
GAPDH was used as an RIP3 mRNA internal control. The RIP3 primers
were forward, 5'-ACT CCC GGC TTA GAA GGA CT-3' and reverse, 5'-GCC
CTG CTC CTC TTG GTA AG-3'. The GAPDH primers were forward, 5'-TGC
ACC ACC AAC TGC TTA GC-3' and reverse, 5'-GGC ATG GAC TGT GGT CAT
GAG-3'.
Western blot analysis
Following corilagin treatment, the cells were
collected and lysed in RIPA lysis buffer. The protein quantity was
analyzed using a Bicinchoninic acid protein assay kit (Thermo
Fisher Scientific, Inc.). The protein samples (20 µg) were
separated using SDS-PAGE (8-12%) and transferred onto PVDF
membranes. Subsequently, these membranes were blocked using 5%
non-fat milk, and incubated with the appropriate primary antibodies
against LC3 (cat. no. NB100-2220), RIP3 (cat. no. GTX107574),
caspase-3 (cat. no. AC030), caspase-8 (cat. no. AC056), caspase-9
(cat. no. AC062) and PARP (cat. no. AP102; all 1:1,000) at 4°C
overnight. The membranes were then incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (1:5,000;
cat. no. A24537) for 1 h at room temperature. Finally, the protein
bands were visualized using the ECL detection system (Pierce;
Thermo Fisher Scientific, Inc.).
Statistical analysis
All data are presented as the mean ± standard
deviation of three independent experiments. Statistical analysis
was performed using Student's t-test with GraphPad Prism 6 software
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
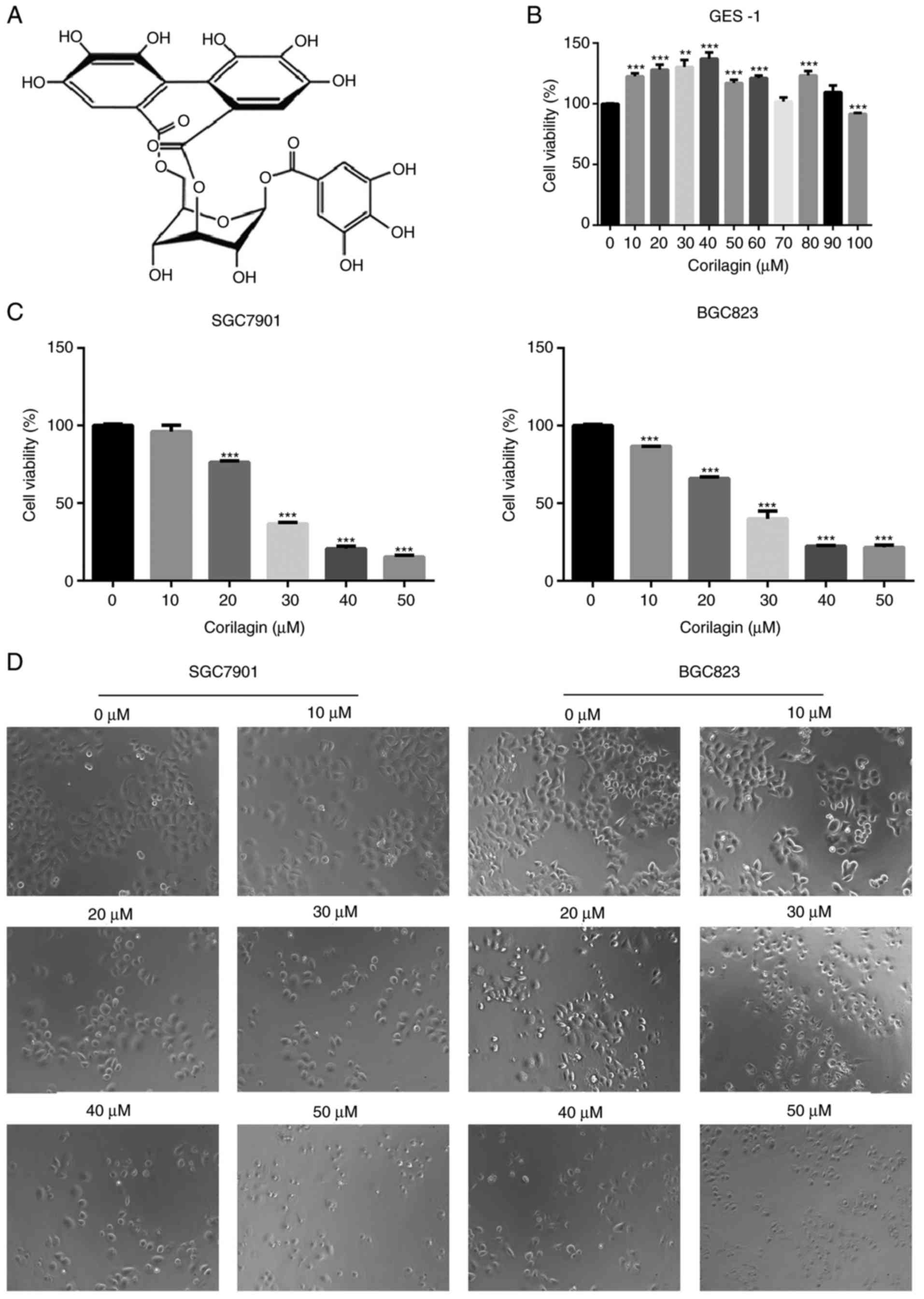
Corilagin inhibits human gastric cancer
cell growth but has no effect on normal cells
To evaluate the growth inhibitory effect of
corilagin (Fig. 1A), the effects
of corilagin on the proliferative activities of the cells were
investigated. The effect of corilagin on the growth of GES-1 human
gastric mucosal epithelial cells was examined using an MTT assay.
As shown in Fig. 1B, the cell
viability in the corilagin-treated groups did not decrease. The
effects on SGC7901 and BGC823 cell lines were also investigated
using the MTT assay. As presented in Fig. 1C, corilagin significantly
inhibited cell proliferation of the two cancer cell lines in a
concentration-dependent manner compared with the control group. The
cytotoxic effect of corilagin on SGC7901 and BGC823 cells was also
assessed by microscopic observations. The results (Fig. 1D) indicated that the cells exposed
in corilagin for 24 h exhibited morphological changes compared with
the untreated cells. Specifically, the cell number was effectively
decreased and the cells became rounded following corilagin
treatment, and this occurred in in a dose-dependent manner. The
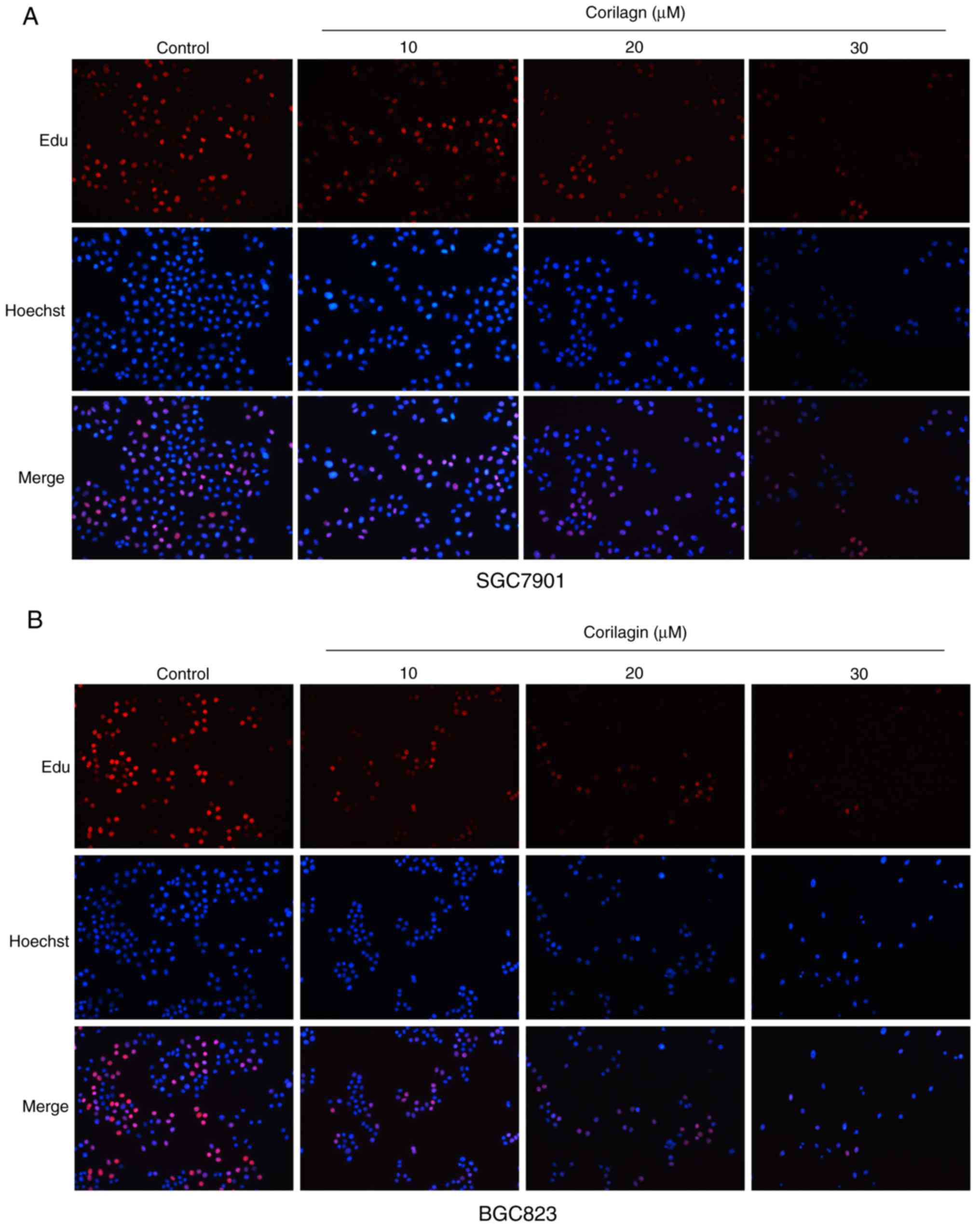
results of the EdU assay (Fig. 2A and
B) confirmed the effects on the proliferation of SGC7901 and
BGC823 cells. Based on these preliminary results, 0, 10, 20 and 30
µM were selected as the appropriate concentration ranges for
SGC7901 and BGC823 cells in the following experiments.
Corilagin induces the apoptosis of
SGC7901 and BGC823 cells
To determine whether the efficacy of corilagin on
cell growth inhibition was a result of apoptosis. Annexin V/PI and
Hoechst 33342 staining assays were combined to assess apoptotic
cell death. First, the morphological characteristics were observed
in apoptotic cells and the results are shown in Fig. 3A. The untreated control cells
manifested weak homogeneous blue fluorescence and a normal nuclear
structure. The corilagin-treated cells exhibited chromosomal
condensation, formation of apoptotic bodies and brighter granular
blue fluorescence, which are characteristics of apoptotic-like
morphological changes. The Annexin V/PI staining assay was
performed to further detect the effect of corilagin on apoptosis.
As shown in Fig. 3B, corilagin
treatment induced apoptosis of the gastric cancer cells in a
dose-dependent manner. Additionally, the number of early and late
stage apoptotic cells were significantly increased following
exposure to corilagin.
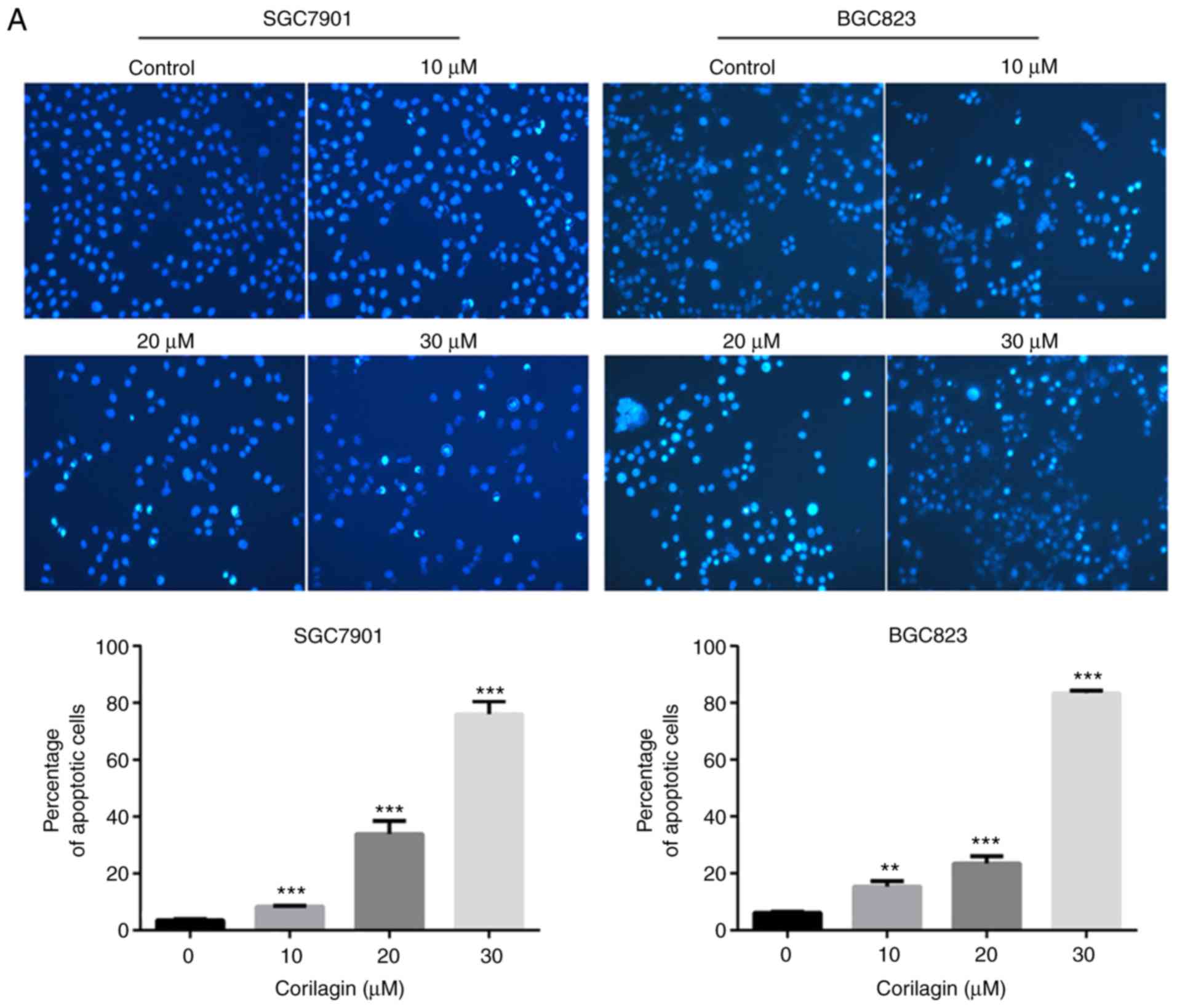 | Figure 3Corilagin induces apoptosis of human
gastric cancer cells. (A) Cells were treated with various
concentrations of corilagin (0, 10, 20 and 30 µM) for 24 h,
and then stained with Hoechst 33342. Images were captured by
fluorescence microscopy at magnification, ×200. Percentages of
apoptotic cells are reported as the mean ± SD (n=3). (B)
Corilagin-treated cells were analyzed by flow cytometry following
staining with Annexin V-FITC and PI. The percentages of cells in
survival, early and late apoptotic stages calculated and are
expressed as the mean ± SD (n=3). (C) LDH release from SGC7901 and
BGC823 cells treated with corilagin for 24 h. Values are reported
as the mean ± SD (n=3). (D) Cells were pretreated with 20 µM
Z-VAD-FMK for 2 h and then incubated with corilagin (30 µM)
for 24 h. Cell viability was analyzed using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenytetrazolium bromide assay.
Data are reported as the mean ± SD (n≥3) of three replicate
experiments. *P<0.05, **P<0.01 and
***P<0.001 vs. control group (detected using
Student's t-test). SD, standard deviation; PI, propidium
iodide. |
The release of LDH into culture media is one of the
indicators of cell death (34).
Therefore, the cytotoxic activity of corilagin on human gastric
cancer cell lines was assessed using an LDH release assay. The
results showed that the percentage of LDH release markedly
increased in a concentration-dependent manner (Fig. 3C) and suggested that cell damage
and cell death had occurred. To further investigate whether
caspase-dependent apoptotic cell death had was present, Z-VAD-FMK,
a pan caspase inhibitor, was used to detect cell viability using an
MTT assay. As shown in Fig. 3D,
Z-VAD-FMK markedly restored cell viability in the SGC7901 and
BGC823 cell lines.
These results indicated that corilagin induced
apoptotic cell death and that the caspase-dependent apoptotic
pathway may be key in the growth inhibitory effect of corilagin on
gastric cancer cells.
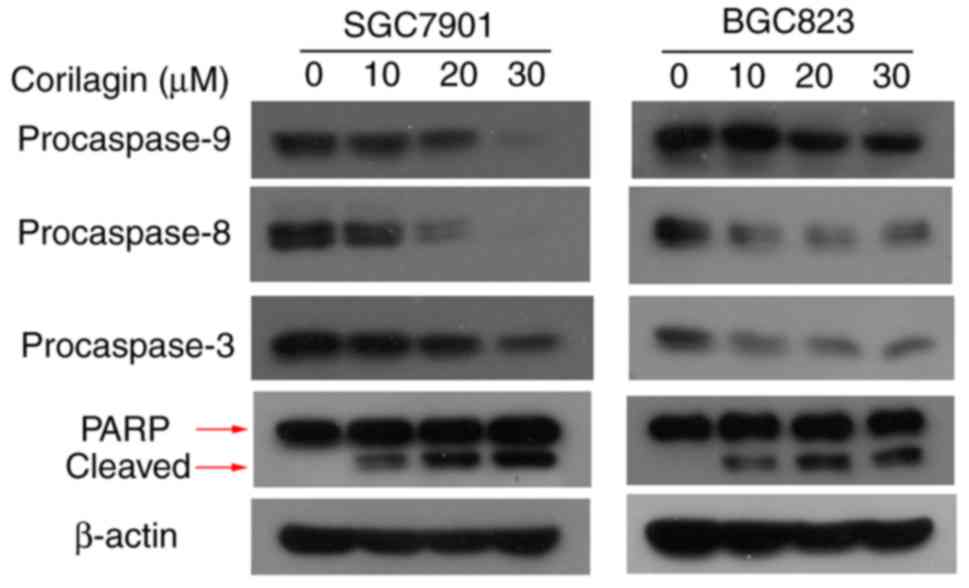
Effect of corilagin on the expression of
apoptosis-related proteins in human gastric cancer cells
To verify the mechanisms of apoptosis of human
gastric cancer cells induced by corilagin, the expression of key
proteins involved in cell apoptosis were detected by western blot
analysis. As shown in Fig. 4, it
was found that corilagin treatment decreased the protein levels of
procaspase-8, -9 and -3 in a dose-dependent manner. Furthermore,
corilagin increased the level of cleaved PARP in a
concentration-dependent manner. These results suggested that
corilagin induced caspase-dependent apoptosis of human gastric
cancer cells.
Corilagin induces autophagy, which has a
cytoprotective effect in human gastric cancer cells
To examine whether corilagin triggers the occurrence
of autophagy, the formation of acidic autophagic vacuoles in
gastric cancer cells was detected using AO dye. Under a
fluorescence microscope, acidic vesicles exhibit bright red
fluorescence and cell nuclei show green fluorescence. As shown in
Fig. 5A, treatment with corilagin
caused enhancement of acidic vesicles in SGC7901 and BGC823 cells
compared with the control group (EBSS, as a positive control). It
is known that LC3 protein is a key marker for autophagy. Therefore,
LC3 conversion from LC3I to LC3II was assessed by western blot
analysis. The results (Fig. 5B)
showed that corilagin markedly increased the level of LC3II. These
findings suggested that corilagin treatment induced autophagy of
human gastric cancer cells.
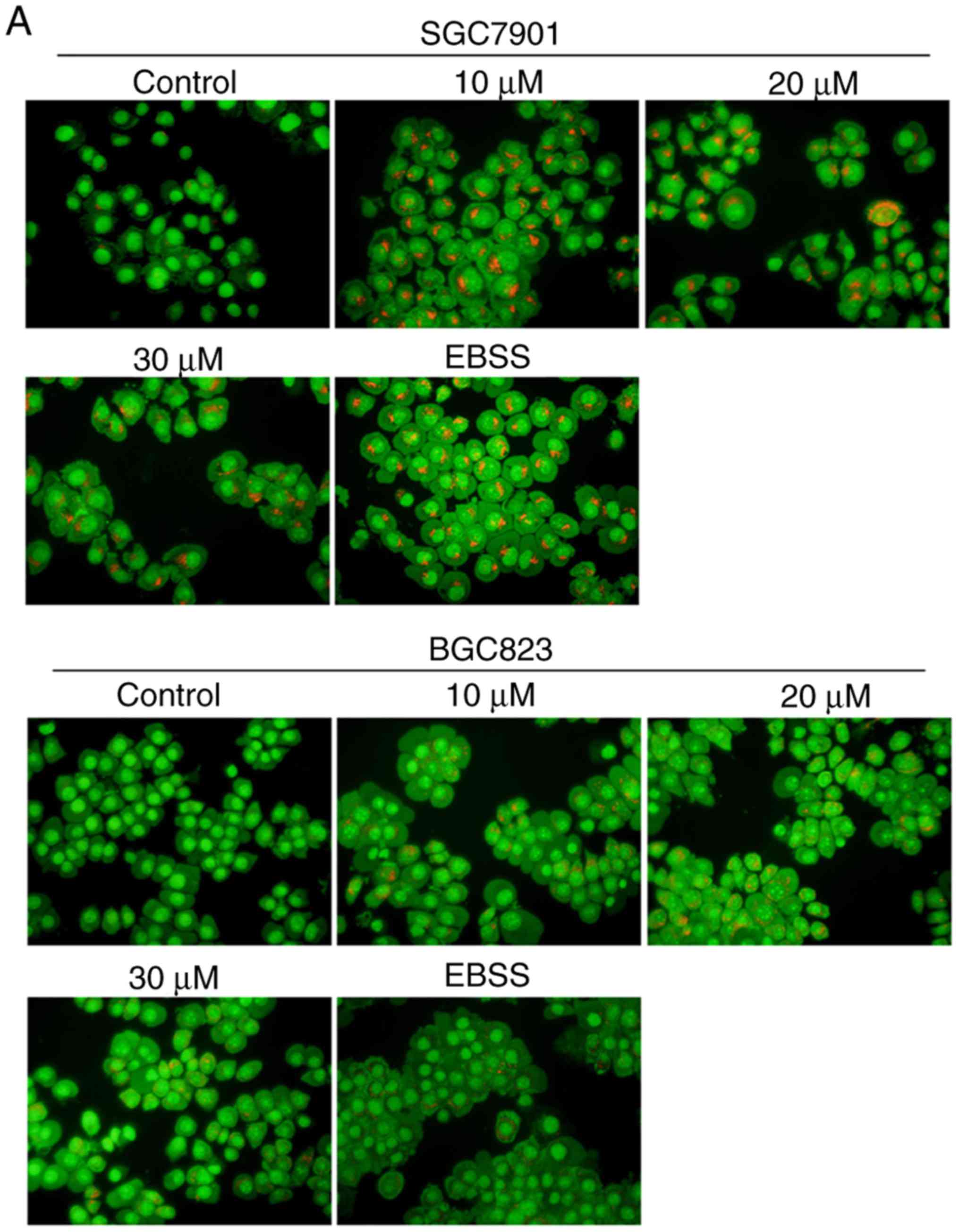 | Figure 5Corilagin triggers autophagy and
inhibition of autophagy increases cell growth suppression in
SGC7901 and BGC823 cells. (A) Observation of corilagin-induced
acidic autophagic vesicles in SGC7901 and BGC823 cells using a
fluorescence microscope. SGC7901 and BGC823 cells were treated with
corilagin (0, 10, 20 and 30 µM) for 24 h, followed by
acridine orange dye (EBSS as positive control), and images were
captured under a fluorescence microscope (magnification, ×200). (B)
Cells were incubated with corilagin at different concentrations (0,
10, 20 and 30 µM) for 24 h, following which LC3I and LC3II
levels were detected via western blot analysis. (C) SGC7901 and
BGC823 cells were pretreated with CQ (20 µM) for 2 h, then
incubated with corilagin (30 µM) for 24 h, and expression of
LC3 was evaluated by western blot analysis. (D) Cells were
pretreated with CQ (20 µM) for 2 h, then treated with
corilagin (30 µM) for 24 h, and cell viability was measured
using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenytetrazolium bromide
assay. Results are expressed as the mean ± standard deviation (n≥3)
from three independent experiments. **P<0.01 and
***P<0.001 vs. control group (detected using
Student's t-test). CQ, chloroquine. |
To further corroborate whether autophagy has as
protective effect on cancer cells, CQ, a late autophagy inhibitor,
and corilagin were used to co-treat SGC7901 and BGC823 cells. The
results from the western blot and MTT assays, respectively, showed
that the expression of LC3II was markedly enhanced and cell
viability was significantly decreased compared with the SGC7901 and
BGC823 cells incubated in corilagin only (Fig. 5C and D). These data indicated that
corilagin induced autophagy in gastric cancer cells and that the
inhibition of autophagy improved the effect of corilagin on the
suppression of cell proliferation.
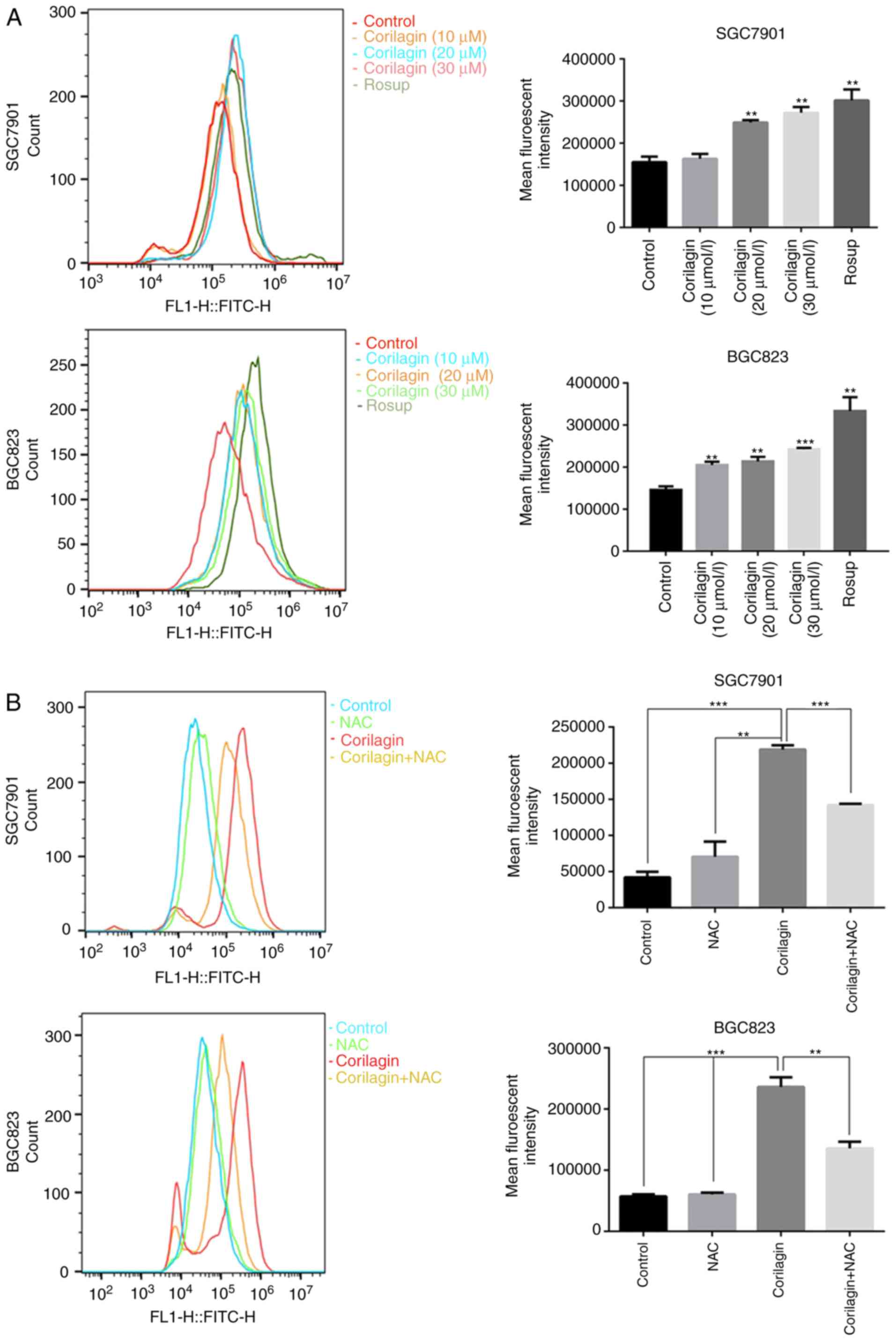
Corilagin induces ROS generation in human
gastric cancer cells
ROS, as significant signaling molecules, are
involved in signal transduction and sustaining cellular redox
homeostasis in aerobic organisms (35). ROS are not only supporters of
tumor cells, but are also efficient therapeutic tools in treating
cancer (29). In the present
study, intracellular ROS production was detected using a DCFH-DA
probe in gastric cancer cells. Following treatment with corilagin
for 24 h, the flow cytometry revealed that corilagin significantly
increased ROS generation in a dose-dependent manner (Fig. 6A). To further analyze the role of
ROS, the ROS scavenger NAC was used to inhibit the accumulation of
intracellular ROS (Fig. 6B). In
addition, an MTT assay was performed to examine cell viability, as
shown in Fig. 6C. Corilagin
induced cell growth inhibition, which was markedly attenuated by
pre-treatment with NAC. These data suggested that there is an
increase of cell viability when the level of ROS was reduced with
NAC.
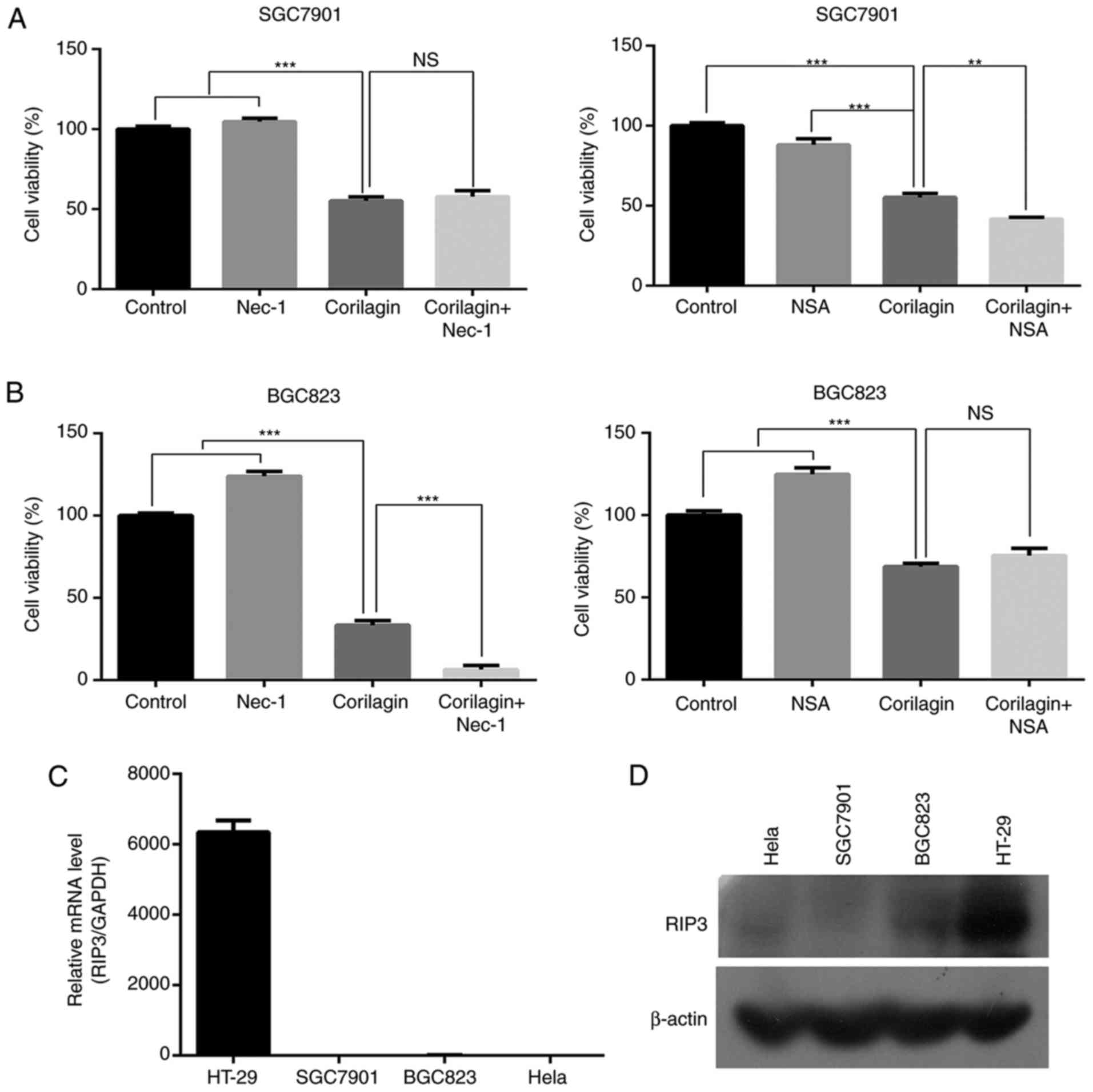
Corilagin cannot activate necroptosis in
SGC7901 and BGC823 cells
To identify whether corilagin induces necroptosis in
gastric cancer cells, an MTT assay was used to examine the cell
growth inhibitory effect of corilagin. First, the SGC7901 and
BGC823 cells were pre-treated with necroptosis inhibitor Nec-1 or
NSA for 2 h, and then treated with corilagin for 24 h. As shown in
Fig. 7A and B, neither Nec-1 nor
NSA significantly restored cell viability in the corilagin-treated
SGC7901 and BGC823 cells. It is known that RIP3 is required for
activation of the necroptosis signaling pathway. Therefore, RT-PCR
and western blot analyses were performed to examine the expression
of RIP3. The results, as shown in Fig. 7C and D, revealed that RIP3 was not
expressed in the SGC7901 or BGC823 cells. HeLa cells were used as a
negative control and HT-29 cells were used as a positive control.
These results demonstrated that corilagin cannot induce necroptosis
in SGC7901 and BGC823 cell lines.
Discussion
As far as we know, although certain pharmacological
and biochemical effects of corilagin have been reported, the
anticancer effect of corilagin has received limited investigation.
Attar et al (18)
investigated the apoptotic and genomic effects of corilagin on
SKOV3 ovarian carcinoma cells. Their results indicated that
corilagin increased apoptosis and altered the genomic expression
levels in SKOV3 cells in a time- and dose-dependent manner.
However, there have been no reports on the effect of corilagin in
gastric carcinoma cells. In the present study, it was first
exhibited that corilagin markedly inhibited cell proliferation in a
concentration-dependent manner in SGC7901 and BGC823 cells.
Furthermore, it was demonstrated that corilagin showed less
toxicity towards normal cells, for example, GES-1 human gastric
mucosal epithelial cells. These results indicated that corilagin
has potent antitumor activity against gastric cancer cells with
less toxicity towards normal cells.
To the best of our knowledge, apoptosis is a
specific type of programmed cell death that is key in the antitumor
activity of several natural products. In the present study, it was
shown that the growth inhibitory properties of corilagin were due
to apoptotic cell death, as detected by Annexin V/PI staining.
Additionally, it was also found that the gastric cancer cells
exhibited the characteristics of apoptotic-like morphological
changes, determined using a Hoechst 33342 staining assay. These
data suggested that corilagin induces the apoptosis of human
gastric cancer cells. Caspases have been widely investigated in the
past (36); they are key in the
cascade that result in apoptosis. Caspases are classified either as
initiators or executors of apoptosis, depending on their time of
entry into the cell death process. Initiator caspases, including
caspases -2, -8, -9 and -10, enter the apoptotic cascade in the
early stage and are responsible for activating the executor
caspases -3, -6 or -7 (37). In
the present study, it was shown that the protein levels of
procaspase -8, -9 and -3 were decreased following corilagin
treatment. Simultaneously, PARP, an indicator of caspase
activation, was cleaved by corilagin incubation. The expression
level of cleaved PARP was increased in a dose-dependent manner in
SGC7901 and BGC823 cells. It was also found that Z-VAD-FMK markedly
restored cell viability. Taken together, these data certified that
corilagin induced caspase-dependent apoptotic cell death in human
gastric cancer.
As a specific cell death pathway, autophagy can be
triggered by various intracellular stimuli and it has conflicting
effects in tumor therapy, as it can have tumor-promoting or
tumor-inhibiting effects. The role of autophagy in diverse
therapies is cell line- and activator-dependent (38). In the present study, the
activation of autophagy was observed in corilagin-treated SGC7901
and BGC823 cells. The autophagy inhibitor, CQ, was used to detect
whether autophagy has a protective effect on corilagin-exposed
gastric cancer cells. The results showed that CQ and corilagin
co-treatment further decreased cell viability compared with that in
cells treated with corilagin alone. Accordingly, it was
hypothesized that corilagin induces autophagy, which has a
cytoprotective role in human gastric cancer cells. However,
additional data are required to confirm this conclusion.
ROS is a collective noun for the reactive forms of
oxygen, and acts as a second messenger that is involved in
signaling cascades (39). The
excessive generation of ROS can result in oxidative stress, DNA
damage and cell death through apoptosis or necrosis (40). For example, Su et al
(41) found that sonodynamic
treatment significantly increased intracellular ROS generation and
induced the apoptosis of K562 human leukemia cells in a
ROS-dependent manner. Similarly, in the present study, it was shown
that corilagin markedly increased ROS production in the SGC7901 and
BGC823 cells. The present study also demonstrated that the cell
growth inhibition induced by corilagin was effectively attenuated
by pre-treatment with NAC. These results suggested that ROS may be
vital in the inhibition of human gastric cancer cell growth caused
by corilagin treatment.
Based on the above results, it was confirmed that
corilagin can induce autophagy, apoptosis and ROS accumulation in
gastric cancer cells. The specific associations among these factors
have attracted increased attention. A study by Srivastava et
al (42) reported that neem
oil limonoids triggered p53-independent apoptosis and autophagy in
cancer cells; furthermore, they found the existence of cross-talk
between them. Numerous studies have demonstrated that autophagy has
as a protective role in several types of cancer cell and the
inhibition of autophagy can increase anticancer drug-induced
apoptosis (43,44). In addition, ROS have been shown to
be important in the pathways of cellular apoptosis and autophagy
(45). For example, Xu et
al (46) demonstrated that
the inhibition of autophagy increased ROS-mediated apoptosis in
mesangial cells. They revealed that apoptosis and autophagy were
dependent on ROS production. In the present study, it was suggested
that the inhibition of autophagy enhanced the effect of corilagin
on gastric cancer cells. Therefore, autophagy may inhibit apoptosis
in corilagin-treated SGC7901 and BGC823 cells. In addition,
corilagin markedly enhanced ROS generation, which is vital in
inhibiting the proliferation of human gastric cancer cells.
Therefore, it is possible that ROS accumulation promoted apoptosis
of the corilagin-exposed gastric cancer cells. However, further
examination is required to analyze the associations of apoptosis,
autophagy and ROS accumulation in subsequent investigations.
Necroptosis, namely programmed necrosis, is
identified as an emerging form of programmed cell death distinct
from apoptosis. Necroptosis can be activated by several
inducements, for example one of which stimulates TNF-α and is a
typical response. In TNF-α-induced necroptosis, RIP3 is necessary
as a key regulator (47). In the
present study, it was initially verified that cell viability was
not enhanced via pre-treatment with necroptosis inhibitor Nec-1 or
NSA prior to corilagin treatment in SGC7901 and BGC823 cells. It
was then shown that RIP3 was not expressed in the SGC7901 and
BGC823 cell lines. Therefore, these findings suggest that corilagin
cannot induce necroptosis in human gastric cancer cells, but
further investigations are required to confirm this.
In conclusion, corilagin can distinctly inhibit the
proliferation of SGC7901 and BGC823 cells in vitro, while
showing low toxicity towards normal cells. Furthermore,
corilagin-treated cells showed induction of apoptotic cell death by
activating caspase -8, -9, -3 and PARP proteins. Alternatively,
corilagin-induced the autophagy of gastric cancer cells and the
inhibition of autophagy improved the effects of corilagin on cell
growth suppression. The excessive production of ROS may be
important in inhibiting the growth of gastric cancer cells
following corilagin treatment. The novel cell death pathway,
necroptosis, was not induced by corilagin-incubation in the SGC7901
and BGC823 cell lines. Therefore, the natural product, corilagin
may be a promising novel drug for the treatment of human gastric
cancer.
Funding
The present study was supported by the grants from
the National Natural Science Foundation of China (grant nos.
81872045 and 81470793), the Special Fund for Public Welfare
Research Institutes of Fujian Province (grant no. 2017R1036-1 and
2018R1036-4) and the Major Technology Program of
Industry-University-Research Cooperation in Fujian Province (grant
no. 2016Y4011).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JX conceived and performed the experiments and wrote
the manuscript. JX, GZ, YT and JY analysed the data. YL and GS
designed the study and provided funding. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Professor Tianhui Hu
(Cancer Research Center, Medical College of Xiamen University,
Xiamen, China) for supporting this study. The authors would also
like to thank the valuable suggestions and comments on this article
received from editor and reviewers.
References
|
1
|
Wang C, Zhang J, Cai M, Zhu Z, Gu W, Yu Y
and Zhang X: DBGC: A database of human gastric cancer. PLoS One.
10:e1425912015.
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu C, Song G, Zhang B, Liu Z, Chen R,
Zhang H and Hu T: Intestinal metabolite compound K of panaxoside
inhibits the growth of gastric carcinoma by augmenting apoptosis
via Bid-mediated mitochondrial pathway. J Cell Mol Med. 16:96–106.
2012. View Article : Google Scholar
|
|
4
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kodera Y, Fujiwara M, Koike M and Nakao A:
Chemotherapy as a component of multimodal therapy for gastric
carcinoma. World J Gastroenterol. 12:2000–2005. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ji YB, Qu ZY and Zou X: Juglone-induced
apoptosis in human gastric cancer SGC-7901 cells via the
mitochondrial pathway. Exp Toxicol Pathol. 63:69–78. 2011.
View Article : Google Scholar
|
|
7
|
Dai ZJ, Gao J, Ji ZZ, Wang XJ, Ren HT, Liu
XX, Wu WY, Kang HF and Guan HT: Matrine induces apoptosis in
gastric carcinoma cells via alteration of Fas/FasL and activation
of caspase-3. J Ethnopharmacol. 123:91–96. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Duan W, Yu Y and Zhang L: Antiatherogenic
effects of phyllanthus emblica associated with corilagin and its
analogue. Yakugaku Zasshi. 125:587–591. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rangkadilok N, Worasuttayangkurn L,
Bennett RN and Satayavivad J: Identification and quantification of
polyphenolic compounds in Longan (Euphoria longana Lam.) fruit. J
Agric Food Chem. 53:1387–1392. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Darwish AG, Samy MN, Sugimoto S, Otsuka H,
Abdel-Salam H and Matsunami K: Effects of hepatoprotective
compounds from the leaves of Lumnitzera racemosa on
acetaminophen-induced liver damage in vitro. Chem Pharm Bull
(Tokyo). 64:360–365. 2016. View Article : Google Scholar
|
|
11
|
Kinoshita S, Inoue Y, Nakama S, Ichiba T
and Aniya Y: Antioxidant and hepatoprotective actions of medicinal
herb, Terminalia catappa L. from Okinawa Island and its tannin
corilagin. Phytomedicine. 14:755–762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Notka F, Meier GR and Wagner R: Inhibition
of wild-type human immunodeficiency virus and reverse transcriptase
inhibitor-resistant variants by Phyllanthus amarus. Antiviral Res.
58:175–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao L, Zhang SL, Tao JY, Pang R, Jin F,
Guo YJ, Dong JH, Ye P, Zhao HY and Zheng GH: Preliminary
exploration on anti-inflammatory mechanism of Corilagin
(beta-1-O-gal-loyl-3,6-(R)-hexahydroxydiphenoyl-D-glucose) in
vitro. Int Immunopharmacol. 8:1059–1064. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo YJ, Luo T, Wu F, Liu H, Li HR, Mei YW,
Zhang SL, Tao JY, Dong JH, Fang Y and Zhao L: Corilagin protects
against HSV1 encephalitis through inhibiting the TLR2 signaling
pathways in vivo and in vitro. Mol Neurobiol. 52:1547–1560. 2015.
View Article : Google Scholar
|
|
15
|
Liu FC, Chaudry IH and Yu HP:
Hepatoprotective effects of Corilagin following hemorrhagic shock
are through Akt-dependent pathway. Shock. 47:346–351. 2017.
View Article : Google Scholar :
|
|
16
|
Du P, Ma Q, Zhu ZD, Li G, Wang Y, Li QQ,
Chen YF, Shang ZZ, Zhang J and Zhao L: Mechanism of Corilagin
interference with IL-13/STAT6 signaling pathways in hepatic
alternative activation macrophages in schistosomiasis-induced liver
fibrosis in mouse model. Eur J Pharmacol. 793:119–126. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ming Y, Zheng Z, Chen L, Zheng G, Liu S,
Yu Y and Tong Q: Corilagin inhibits hepatocellular carcinoma cell
proliferation by inducing G2/M phase arrest. Cell Biol Int.
37:1046–1054. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Attar R, Cincin ZB, Bireller ES and
Cakmakoglu B: Apoptotic and genomic effects of corilagin on SKOV3
ovarian cancer cell line. Onco Targets Ther. 10:1941–1946. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu Y, Xiao L, Ming Y, Zheng Z and Li W:
Corilagin suppresses cholangiocarcinoma progression through Notch
signaling pathway in vitro an in vivo. Int J Oncol. 48:1868–1876.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Milani R, Brognara E, Fabbri E, Finotti A,
Borgatti M, Lampronti I, Marzaro G, Chilin A, Lee KK, Kok SH, et
al: Corilagin induces high levels of apoptosis in the
Temozolomide-resistant T98G glioma cell line. Oncol Res. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jia L, Jin H, Zhou J, Chen L, Lu Y, Ming Y
and Yu Y: A potential anti-tumor herbal medicine, Corilagin,
inhibits ovarian cancer cell growth through blocking the TGF-β
signaling pathways. BMC Complement Altern Med. 13:332013.
View Article : Google Scholar
|
|
22
|
Li X, Deng Y, Zheng Z, Huang W, Chen L,
Tong Q and Ming Y: Corilagin, a promising medicinal herbal agent.
Biomed Pharmacother. 99:43–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hikita H, Kodama T, Shimizu S, Li W,
Shigekawa M, Tanaka S, Hosui A, Miyagi T, Tatsumi T, Kanto T, et
al: Bak deficiency inhibits liver carcinogenesis: A causal link
between apoptosis and carcinogenesis. J Hepatol. 57:92–100. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahamed M, Akhtar MJ, Siddiqui MA, Ahmad J,
Musarrat J, Al-Khedhairy AA, AlSalhi MS and Alrokayan SA: Oxidative
stress mediated apoptosis induced by nickel ferrite nanoparticles
in cultured A549 cells. Toxicology. 283:101–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Li T, Zhang L, Wang X, Dong H, Li
L, Fu D, Li Y, Zi X, Liu HM, et al: A novel chalcone derivative S17
induces apoptosis through ROS dependent DR5 up-regulation in
gastric cancer cells. Sci Rep. 7:98732017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shimizu S, Yoshida T, Tsujioka M and
Arakawa S: Autophagic cell death and cancer. Int J Mol Sci.
15:3145–3153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu B, Xu M, Tian Y, Yu Q, Zhao Y, Chen X,
Mi P, Cao H, Zhang B, Song G, et al: Matrine induces RIP3-dependent
necroptosis in cholangiocarcinoma cells. Cell Death Discov.
3:160962017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu W, Liu P and Li J: Necroptosis: An
emerging form of programmed cell death. Crit Rev Oncol Hematol.
82:249–258. 2012. View Article : Google Scholar
|
|
29
|
Manda G, Isvoranu G, Comanescu MV, Manea
A, Debelec Butuner B and Korkmaz KS: The redox biology network in
cancer pathophysiology and therapeutics. Redox Biol. 5:347–357.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen H, Li Y, Zhu Y, Wu L, Meng J, Lin N,
Yang D, Li M, Ding W, Tong X and Su Q: Advanced glycation end
products promote ChREBP expression and cell proliferation in liver
cancer cells by increasing reactive oxygen species. Medicine
(Baltimore). 96:e74562017. View Article : Google Scholar
|
|
31
|
Lee Y, Sung B, Kang YJ, Kim DH, Jang JY,
Hwang SY, Kim M, Lim HS, Yoon JH, Chung HY and Kim ND:
Apigenin-induced apoptosis is enhanced by inhibition of autophagy
formation in HCT116 human colon cancer cells. Int J Oncol.
44:1599–1606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang P, Zhang YH, Zheng XW, Liu YJ, Zhang
H, Fang L, Zhang YW, Yang C, Islam K, Wang C and Naranmandura H:
Phenylarsine oxide (PAO) induces apoptosis in HepG2 cells via
ROS-mediated mitochondria and ER-stress dependent signaling
pathways. Metallomics. 9:1756–1764. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Attarde SS and Pandit SV: Cytotoxic
activity of NN-32 toxin from Indian spectacled cobra venom on human
breast cancer cell lines. BMC Complement Altern Med. 17:5032017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Briehl MM: Oxygen in human health from
life to death-An approach to teaching redox biology and signaling
to graduate and medical students. Redox Biol. 5:124–139. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shalini S, Dorstyn L, Dawar S and Kumar S:
Old, new and emerging functions of caspases. Cell Death Differ.
22:526–539. 2015. View Article : Google Scholar :
|
|
37
|
MacKenzie SH and Clark AC: Death by
caspase dimerization. Adv Exp Med Biol. 747:55–73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao C, She T, Wang L, Su Y, Qu L, Gao Y,
Xu S, Cai S and Shou C: Daucosterol inhibits cancer cell
proliferation by inducing autophagy through reactive oxygen
species-dependent manner. Life Sci. 137:37–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Forman HJ, Maiorino M and Ursini F:
Signaling functions of reactive oxygen species. Biochemistry.
49:835–842. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Eckert A, Keil U, Marques CA, Bonert A,
Frey C, Schüssel K and Müller WE: Mitochondrial dysfunction,
apoptotic cell death, and Alzheimer's disease. Biochem Pharmacol.
66:1627–1634. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Su X, Wang P, Yang S, Zhang K, Liu Q and
Wang X: Sonodynamic therapy induces the interplay between apoptosis
and autophagy in K562 cells through ROS. Int J Biochem Cell Biol.
60:82–92. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Srivastava P, Yadav N, Lella R, Schneider
A, Jones A, Marlowe T, Lovett G, O'Loughlin K, Minderman H, Gogada
R and Chandra D: Neem oil limonoids induces p53-independent
apoptosis and autophagy. Carcinogenesis. 33:2199–2207. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Masud AM, Kariya R, Kawaguchi A, Matsuda
K, Kudo E and Okada S: Inhibition of autophagy by chloroquine
induces apoptosis in primary effusion lymphoma in vitro and in vivo
through induction of endoplasmic reticulum stress. Apoptosis.
21:1191–1201. 2016. View Article : Google Scholar
|
|
44
|
Wang X, Wang P, Zhang K, Su X, Hou J and
Liu Q: Initiation of autophagy and apoptosis by sonodynamic therapy
in murine leukemia L1210 cells. Toxicol In Vitro. 27:1247–1259.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Datta K, Babbar P, Srivastava T, Sinha S
and Chattopadhyay P: p53 dependent apoptosis in glioma cell lines
in response to hydrogen peroxide induced oxidative stress. Int J
Biochem Cell Biol. 34:148–157. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xu L, Fan Q, Wang X, Zhao X and Wang L:
Inhibition of autophagy increased AGE/ROS-mediated apoptosis in
mesangial cells. Cell Death Dis. 7:e24452016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu X, Li Y, Peng S, Yu X, Li W, Shi F,
Luo X, Tang M, Tan Z, Bode AM and Cao Y: Epstein-Barr virus encoded
latent membrane protein 1 suppresses necroptosis through targeting
RIPK1/3 ubiquitination. Cell Death Dis. 9:532018. View Article : Google Scholar : PubMed/NCBI
|