Introduction
Alzheimer’s disease (AD), a progressive degenerative
disease of the nervous system, is characterized by a substantial
decline in cognitive function and poor prognosis. Intracellular
neurofibrillary tangles and extracellular senile plaques,
particularly aggregated amyloid-β titanium and hyper-phosphorylated
tau, are considered typical markers of AD histopathology (1). Although the exact molecular
mechanism of neurodegeneration in AD remains unclear, through basic
and clinical studies (2,3), oxidative stress and neuronal
apoptosis have been reported to serve a pivotal function in the
process of AD. Oxidative stress, resulting in an imbalance of
anti-oxidation and pro-oxidation, may promote amyloid β (Aβ)
aggregation accompanied by the over-accumulation of inflammatory
reactive oxygen species (ROS) (4). Additionally, hyper-levels of ROS
tend to enhance the dissipation of the mitochondrial transmembrane
potential (MMP). In a short feedback loop, the increased
permeability of mitochondria further enhances the over-releasing of
ROS and other factors, including cytochrome c, into the
cytoplasm, which triggers the cell apoptosis program (5,6).
Abnormally high levels of neurons damaged by glutamate, a
central-nervous neurotransmitter, are closely associated with ROS
accumulation (7).
The global impact of AD is rapidly expanding
(8), as >40 million patients
globally have been reported, which is expected to increase to 70
million by 2030 (8). Therefore,
the prevention and therapization of AD is an urgent challenge.
Unfortunately, clinical trials of AD drugs have a high failure
rate, and the majority of them exert various adverse effects
including loss of appetite, gastrointestinal discomfort, difficulty
sleeping and muscle spasms (9).
Herbal compounds have been considered as potential agents for the
prevention of AD (10,11). Crocin, the main component of
Crocus sativus L. extract, is a yellow carotenoid that has
been reported to exert anti-inflammatory (12), anti-depressant (13), memory improvement (14) and anti-apoptotic properties
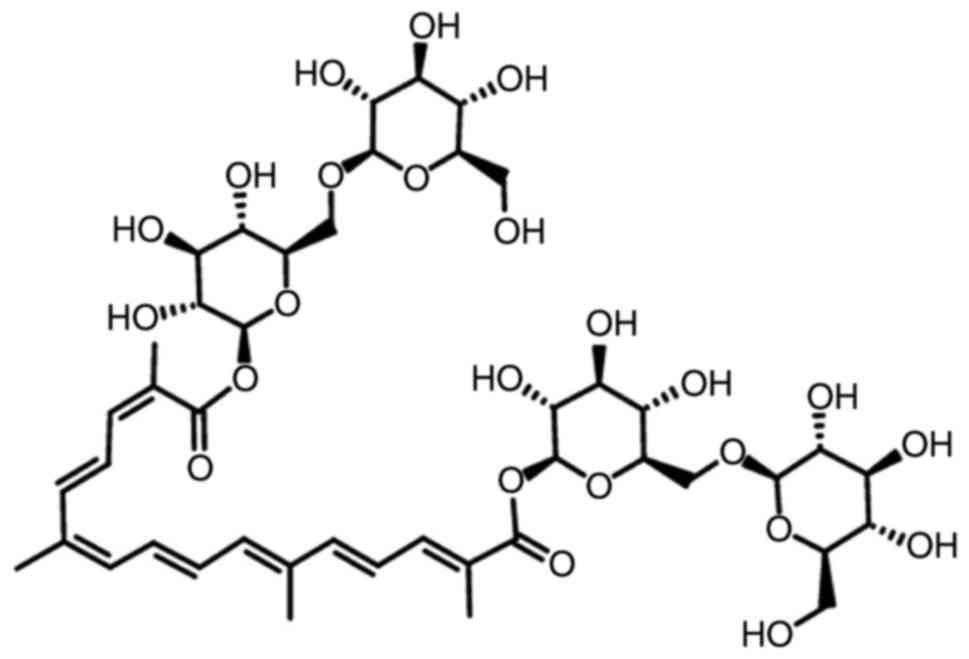
(15). The structure of crocin is
presented in Fig. 1. In PC12
cells, crocin prevents cell apoptosis via increasing glutathione
synthesis (16); meanwhile, in
aging Wistar rats, crocin enhances their memory abilities, mainly
through its anti-oxidant activities (17). In rats administered tramadol
hydrochloride, crocin improves their learning and reduces memory
impairment (18). However, no
systematic study of the effects of crocin on AD, and its underlying
mechanisms, has been reported.
In the present study, the neuroprotective effects of
crocin against AD were investigated in l-glutamate (L-Glu)-induced HT22
apoptotic cells, and in aluminum trichloride (AlCl3) and
d-galactose
(d-gal)-induced AD
mice. The present study aimed to provide experimental evidence that
crocin may be a candidate agent for the adjuvant treatment of
AD.
Materials and methods
Cell culture
HT22 cells (cat. no. 337709; BeNa Culture
Collection, Beijing, China), a mouse hippocampal neuronal cell
line, were cultured in Dulbecco’s modified Eagle’s medium (DMEM)
containing 10% fetal bovine serum, 1% 100 µg/ml streptomycin
and 100 units/ml penicillin (all Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in a 5% CO2
incubator to provide a humidified atmosphere (Thermo Fisher
Scientific, Inc.).
MTT assay
HT22 cells cultured in 96-well plates, were
pretreated with crocin (cat. no. B21336; purity ≥98%; Shanghai
Yuanye Biotechnology Co., Ltd., Shanghai, China) at doses of 0.5,
1, 2 and 4 µM for 3 h at 37°C and then incubated with 25 mM
l-Glu for another 24
h at 37°C. The control cells were not treated with 25 mM
l-Glu and crocin, and
incubated for 24 h. A total of 5 mg/ml MTT (Merck KGaA, Darmstadt,
Germany) was added for 4 h at 37°C, and then 100 µl dimethyl
sulfoxide (Merck KGaA, Darmstadt, Germany) was added. Absorbance
was analyzed using a SynergyTM4 Microplate Reader at 490
nm (BioTek Instruments, Inc., Winooski, VT, USA).
Cell apoptosis assay
HT22 cells cultured in 6-well plates were pretreated
with crocin at doses of 0.5 and 2 µM for 3 h at 37°C and
then incubated with 25 mM l-Glu for 24 h at 37°C. Collect
cells and resuspend in 100 µl 1% FBS, then incubated with
Annexin V & Dead Cell Reagent (cat. no. 4700-1485, 100
tests/bottle) for 20 min at 25°C in the dark, and detected using
the Muse™ Cell Analyzer (EMD Millipore, Billerica, MA, USA).
MMP, intracellular ROS and
Ca2+ measurement
HT22 cells were pretreated with crocin at doses of
0.5 and 2 µM for 3 h and then incubated with 25 mM
l-Glu for 12 h. Cells
were further incubated with 2 µM
5,5′,6,6′-TetrAchloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide
staining (EMD Millipore, Billerica, MA, USA), 10 µM
2′-7′-dichlorodihydrofluorescein diacetate (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) or 2 µM Fluo-4-AM (Molecular
Probes; Thermo Fisher Scientific, Inc.) for 20 min at 25°C in the
dark to investigate the changes in MMP and the levels of
intracellular of ROS and Ca2+, respectively. All
fluorochrome were dissolve in serum-free DMEM. Subsequent to
washing, the fluorescence intensities were detected using
fluorescence microscopy (CCD camera, TE2000; Nikon Corporation,
Tokyo, Japan).
Western blot analysis
HT22 cells were pretreated with crocin at doses of
0.5 and 2 µM for 3 h and then incubated with 25 mM
l-GLU for 24 h. Cells
were lysed for 0.5 h at 0°C with radio immunoprecipitation assay
buffer containing 2% phenylmethanesulfonyl fluoride (Sigma-Aldrich;
Merck KGaA) and 1% protease inhibitor cocktail (Sigma-Aldrich;
Merck KGaA). Subsequent to the detection of the protein
concentration using the BCA kit (Merck KGaA) 40 µg lysates
were separated using 12% dodecyl sulfate, sodium salt
poly-vinylidene fluoride membranes (0.45 µm; Merck KGaA).
The membranes were blocked with 5% bovine serum albumin for 2 h at
4°C, and then incubated with primary antibodies as follows: B-cell
lymphoma-extra large (Bcl-xL; 26 kDa; cat. no. ab32370), Bcl2
associated X, apoptosis regulator (Bax; 21 kDa; cat. no. ab32503),
Bcl2 associated agonist of cell death (Bad; 23 kDa; cat. no.
ab32445), cleaved caspase-3 (32 kDa; cat. no. ab2302),
phosphorylated (P-) protein kinase B (Akt; phospho S473; 60 kDa
S473; cat. no. ab18206), total (T)-Akt (60 kDa; cat. no. ab106693),
P-mammalian target of rapamycin (mTOR; phospho S2448; 289 kDa
S2448; cat. no. ab109268), T-mTOR (289 kDa; cat. no. ab83495) and
GAPDH (36 kDa; cat. no. ab181602), which were purchased from Abcam
(Cambridge, MA, USA) at 4°C overnight at a dilution of 1:2,000.
Subsequent to three washes with TBST (consists of 500 ml TBS and
0.5 ml Tween-20), TBS (20X) is composed of 48.4 g Tris, 584 g NaCl
and 900 ml distilled water (Beijing Chemical Works Beijing, China),
the membranes were incubated with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit immunoglobulin G antibody (cat.
no. SH-0032; Bejing Dingguo Changsheng Biotechnology Co., Ltd.,
Beijing, China) at 4°C for 4 h at a dilution of 1:2,000. An
enhanced chemiluminesence kit (Merck KGaA) was used to visualize
the protein bands under an imaging system (BioSpectrum600).
Quantitation of the results was accomplished by ImageJ (×64) 1.48u
software (National Institutes of Health, Bethesda, MD,
USA).
AD mice model establishment and
administration
The experiment was ethically approved by the
Institution Animal Ethics Committee of Jilin University (Changchun,
China; license no. 20160409). A total of 100 BALB/c mice (6-8
weeks; 18-20 g) were maintained at a standard 12:12 h light/dark
cycle at 23±1°C. The mice were fed autoclaved standard chow ad
libitum.
As described in a previous study (19), a total of 60 mice were randomly
intraperitoneally injected with 120 mg/kg d-gal (Sigma-Aldrich; Merck KGaA)
and intragastrically administrated with 20 mg/kg AlCl3
(Sigma-Aldrich; Merck KGaA) once a day for 8 weeks, as the process
of the establishment of the AD mice model. From the 5th week
onwards, crocin-treated mice were intragastrically treated with 5
or 20 mg/kg crocin everyday (n=20/group); meanwhile, model mice
were intragastrically administrated with saline (n=20). A total of
20 mice injected intraperitoneal and intragastrical with saline for
8 weeks served as the control group. Another 20 mice were
intraperitoneally and intragastrically injected with saline
throughout the whole experiment and intragastric treatment with 20
mg/kg crocin from the 5th week, which served as the crocin single
treated mice group. If any mouse exhibited endpoint signs,
including rapidly reduced bodyweight (loss of >20% bodyweight),
they were euthanized using an intraperitoneal injection of 200
mg/kg pentobarbital. During the whole experiment period, the
maximum body weight loss was <8.3% amongst all experimental
mice.
Animal behavioral detection Morris water
maze test
As described in a previous study (19), a circular pool filled with 10 cm
(depth) water (22-24°C) containing 1 l milk was used to perform the
water maze test. A total of 5 days prior to the form test, mice
were trained for 5 min per day. The latency from the immersion of
each mouse into the pool until they escaped onto the hidden
platform was recorded. On the test day, mice were subjected to the
pool in the same quadrant to identify the platform, and the time
spent to locate the platform within 120 sec was recorded.
Open field test
A soundproof dark box was set with a bottom area of
30×30 cm, with a central region of 15×15 cm as the central area,
and the remaining area as the surrounding area. On the test day,
mice were placed in the fixed central position in the box, and
their moving track and the time of exploring the central area
within 5 min were observed and recorded.
Sample collection and biochemical
detection
Blood of all experimental mice were collected from a
caudal vein subsequent to the behavioral tests. All mice were
euthanized by an intraperitoneal injection of 200 mg/kg
pentobarbital. Following this, the brains and hypothalamus were
immediately collected. The cerebral cortex, separated from the
brains (n=20/group) and the hypothalamus (n=20/group) were
homogenized in ice-cold phosphate buffered saline (PBS).
Subsequent to detecting the protein concentration
using a BCA assay kit (Merck KGaA), the levels of acetyl-choline
(Ach; cat. no. CK-E20536), acetylcholinesterase (AchE; cat. no.
CK-E93899), choline acetyltransferase (ChAT; cat. no. CK-E94456),
super oxidase dismutase (SOD; cat. no. CK-E20348), glutathione
peroxidase (GSH-Px; cat. no. CK-E92669) and ROS (cat. no.
CK-E91516) in serum, the cerebral cortex and the hypothalamus, and
Aβ1-42 (cat. no. CK-E94157) in the serum and cerebral cortex were
detected using enzyme-linked immunosorbent assay kits according to
the manufacturers protocols (Shanghai Yuanye Biotechnology Co.,
Ltd.).
Histological examination
The fixed whole brain hemisphere, kidney and spleen
were washed, gradient dehydration using 30, 50, 70, 80, 95 and 100%
ethanol in sequence and finally embedded in paraffin at 25°C for 12
h, which were further sliced into 4 µm sections. Then, a
hydrating gradient using 100, 95, 80, 70 and 50% ethanol, and
distilled water were applied in sequence and sections were stained
using hematoxylin-eosin (H&E) as previously described (20).
Immunohistochemistry
Brain tissues (n=10/group) were fixed with 4%
formalin solution at 25°C for 24 h, gradient dehydration using 30,
50, 70, 80, 95 and 100% ethanol in sequence, washed in xylene and
embedded in paraffin, and then sliced into 5 µm-thick
sections. Slides were dewaxed, hydrated and boiled for 10 min in 10
mM sodium citrate buffer (pH 6) and cooled at 25°C for 30 min.
Subsequent to incubation for 10 min in 3% hydrogen peroxide,
sections were blocked with normal 10% goat serum for 30 min at 25°C
and then incubated overnight at 4°C with primary antibodies against
Aβ (1:200; cat. no. ab32136; Abcam, Cambridge, UK). Subsequent to
washing with PBS, the sections were incubated with horseradish
peroxidase-conjugated anti-rabbit secondary antibody (cat. no.
sc-3836; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 1 h
at 25°C and then incubated with streptavidin-organism HRP complex
(Shanghai BestBio Science, China) for 60 min at 25°C. The
peroxidase conjugate was counterstained using 5% diaminobenzidine
tetrahydrochloride solution as the chromogen and hematoxylin at
25°C for 5 min. Immunoperoxidase staining of Aβ was observed using
light microscopy (magnification ×100; Olympus Corporation, Tokyo,
Japan).
Statistical analysis
Data were expressed as the mean ± the standard error
of the mean. One-way analysis of variance followed by post-hoc
multiple comparisons (Dunn’s test) was used to determine
statistical significance using SPSS 16.0 software (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Crocin protected HT22 cells against
l-Glu-induced
mitochondrial apoptosis
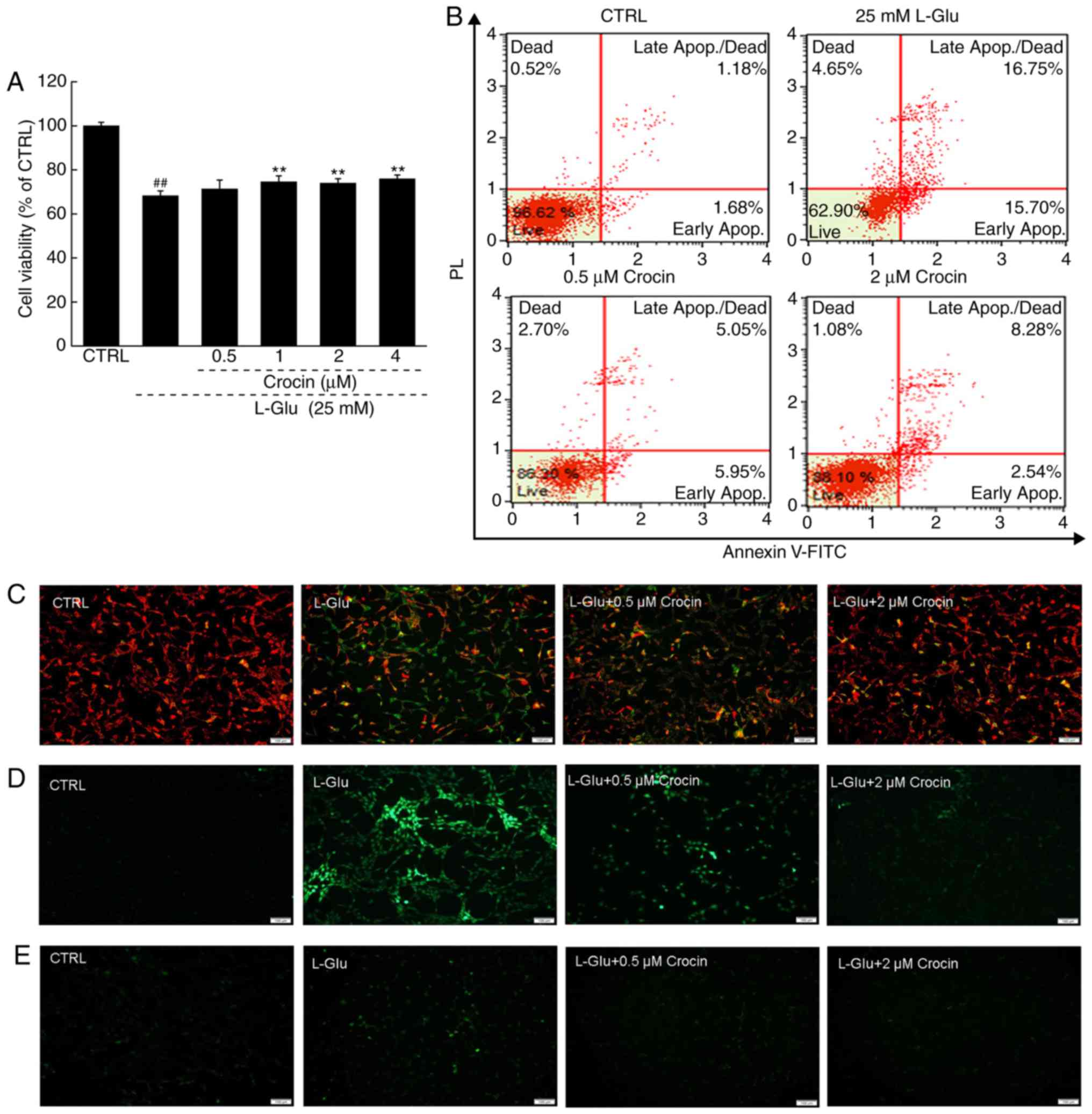
Crocin significantly increased cell viability in the
cells exposed to l-Glu for 24 h compared with
cells exposed to l-Glu alone (P<0.01; Fig. 2A). Among the HT22 cells exposed to
l-Glu for 24 h, 32.5%
were revealed to be apoptotic; in contrast, 3 h pre-incubation with
crocin reduced the apoptosis rate to 10.8% (Fig. 2B).
An imbalance in MMP characterizes the early stage of
mitochondrial injury (21).
Compared with the l-Glu-damaged HT22 cells
subsequent to 12 h of exposure, 3 h of pre-incubation with crocin
strongly restored the MMP dissipation, as indicated by the
increased red fluorescence intensity and reduced green fluorescence
intensity (Fig. 2C).
Crocin also strongly suppressed the hyper-levels of
intracellular ROS compared with the l-Glu-damaged HT22 cells, as
indicated by the reduced green fluorescence intensity (Fig. 2D); and intracellular ROS serves as
an important mediator of cell damage (22).
The overload of Ca2+ caused by
l-Glu is another
factor reportedly associated with mitochondrial function (23). In the 12 h l-Glu-exposed HT22 cells, crocin
strongly inhibited the overload of Ca2+ compared with
the l-Glu-damaged
HT22 cells, as indicated by the reduced green fluorescence
intensity (Fig. 2E).
The expression of Bcl-2 family members directly
influence mitochondrial function (24). In l-Glu-damaged HT22 cells, the low
expression of Bcl-xL and high expression of Bax, Bad and cleaved
caspase-3 compared with control cells were identified (Fig. 3A). Comparatively, 3 h of
pre-incubation with crocin substantially reversed these changes to
anti- and pro-apoptotic protein levels, and even restored them to
their standard levels (Fig.
3A).
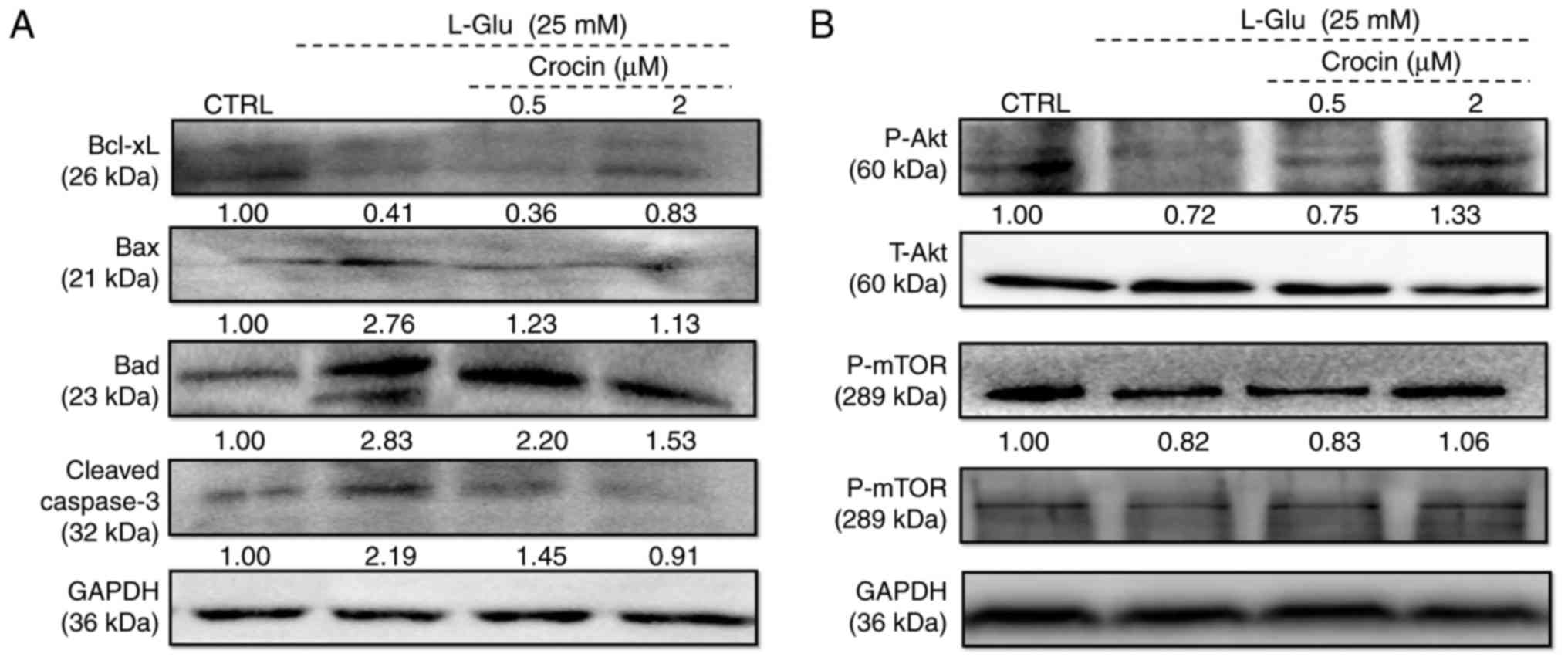 | Figure 3Crocin regulated the expression of
anti- and pro-apoptotic proteins in HT22 cells exposed to
l-Glu for 24 h.
Expression of (A) Bcl-xL, Bax, Bad, cleaved caspase-3, (B) P-Akt,
T-Akt, P-mTOR and T-mTOR were examined. Crocin increased the
expression of Bcl-xL, P-Akt and P-mTOR, and reduced the expression
of Bax, Bad and cleaved caspase-3. Quantification data were
normalized using GAPDH and corresponding total proteins, and the
mean fold of band intensity compared with the CTRL group was noted
(n=6). l-Glu,
l-glutamate; CTRL,
control; Bcl-xL, B-cell lymphoma-extra large; Bax, Bcl2 associated
X, apoptosis regulator; Bad, Bcl2 associated agonist of cell death;
P-, phosphorylated; T-, total; Akt, protein kinase B; mTOR,
mechanistic target of rapamycin. |
Akt/mTOR signaling is involved in the pro-survival
and anti-apoptosis of neuronal protection (25). Following 24 h of exposure in HT22
cells, 25 mM of l-Glu
substantially reduced the phosphorylation of Akt and mTOR compared
with the control cells (Fig. 3B),
and these reductions were markedly reversed by 3 h of
pre-incubation of crocin (Fig.
3B).
Crocin improved cognitive abilities of AD
mice, and reduced Aβ deposition in their brains
In the present study, mice with AD induced by
AlCl3 and d-gal administration were studied
to investigate the beneficial effects of crocin in animal models.
In the open field test, the AD mice demonstrated significantly more
chaotic movements, without purpose, around the central field area
compared with the control group (P<0.001; Fig. 4A). The most notable feature of AD
is memory loss, with diminishing spatial discernment. The Morris
water maze, an useful tool for the evaluation of anti-AD agents,
has been widely used to assess learning and memory abilities
(26). Herein, in the Morris
water maze test, the AD mice required significantly more time to
locate the platform hidden in the water, and demonstrated chaotic
movements compared with the control group (P<0.05; Fig. 4B). The crocin-treated mice
circulated around the periphery significantly more compared with
the center in the open field test (P<0.05; Fig. 4A), and required significantly less
time to locate the platform in the Morris water maze test
(P<0.05; Fig. 4B) compared
with the non-treated AD mice. Crocin-alone treatment caused no
significant changes to healthy mice behaviors in the open field
(Fig. 4A) and Morris water maze
tests (Fig. 4B).
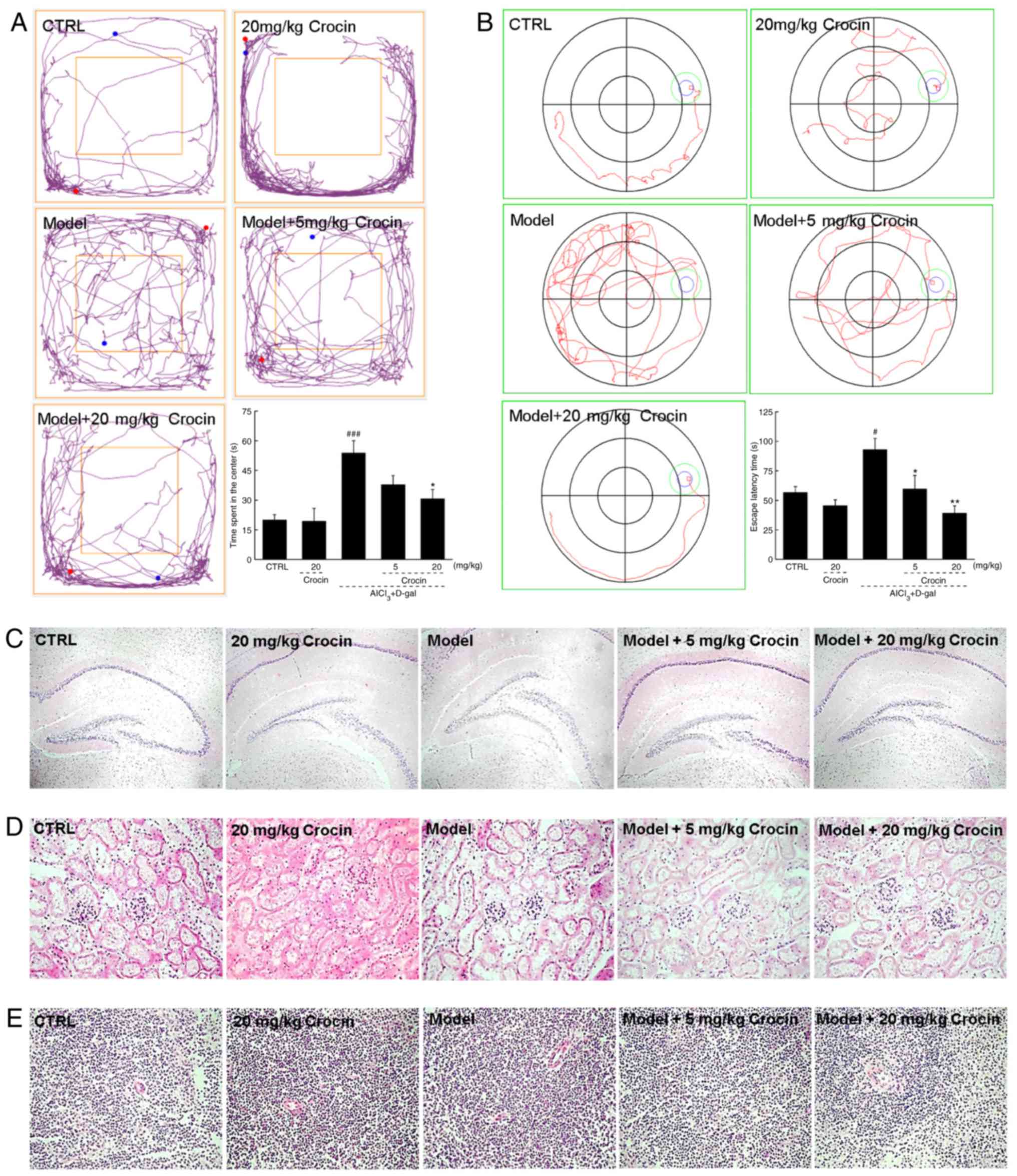 | Figure 4Crocin improves AD-like behaviors in
AlCl3 and d-gal developed AD mice. Compared
with non-treated AD mice, crocin administration reduced (A) the
time spent in the center of the open-yield test, and (B) the time
spent locating the platform in Morris water maze. Data expressed as
mean ± the standard error of the mean (n=20). #P<0.05
and ###P<0.001 vs. control mice,
*P<0.05 and **P<0.01 vs. AD mice. (C)
No substantial changes in brain tissue were noted amongst all
experimental mice detected by H&E staining (magnification, ×40;
n=10). No substantial changes in the (D) kidney and (E) spleen were
noted amongst all experimental mice detected by H&E staining
(magnification, ×200; n=10). AD, Alzheimer’s disease;
AlCl3, aluminum trichloride; d-gal, d-galactose; CTRL, control;
H&E, haemotoxylin and eosin. |
The data obtained from H&E staining demonstrated
that crocin displayed no adverse effects on the brains (Fig. 4C), kidneys (Fig. 4D) or spleens of the mice (Fig. 4E).
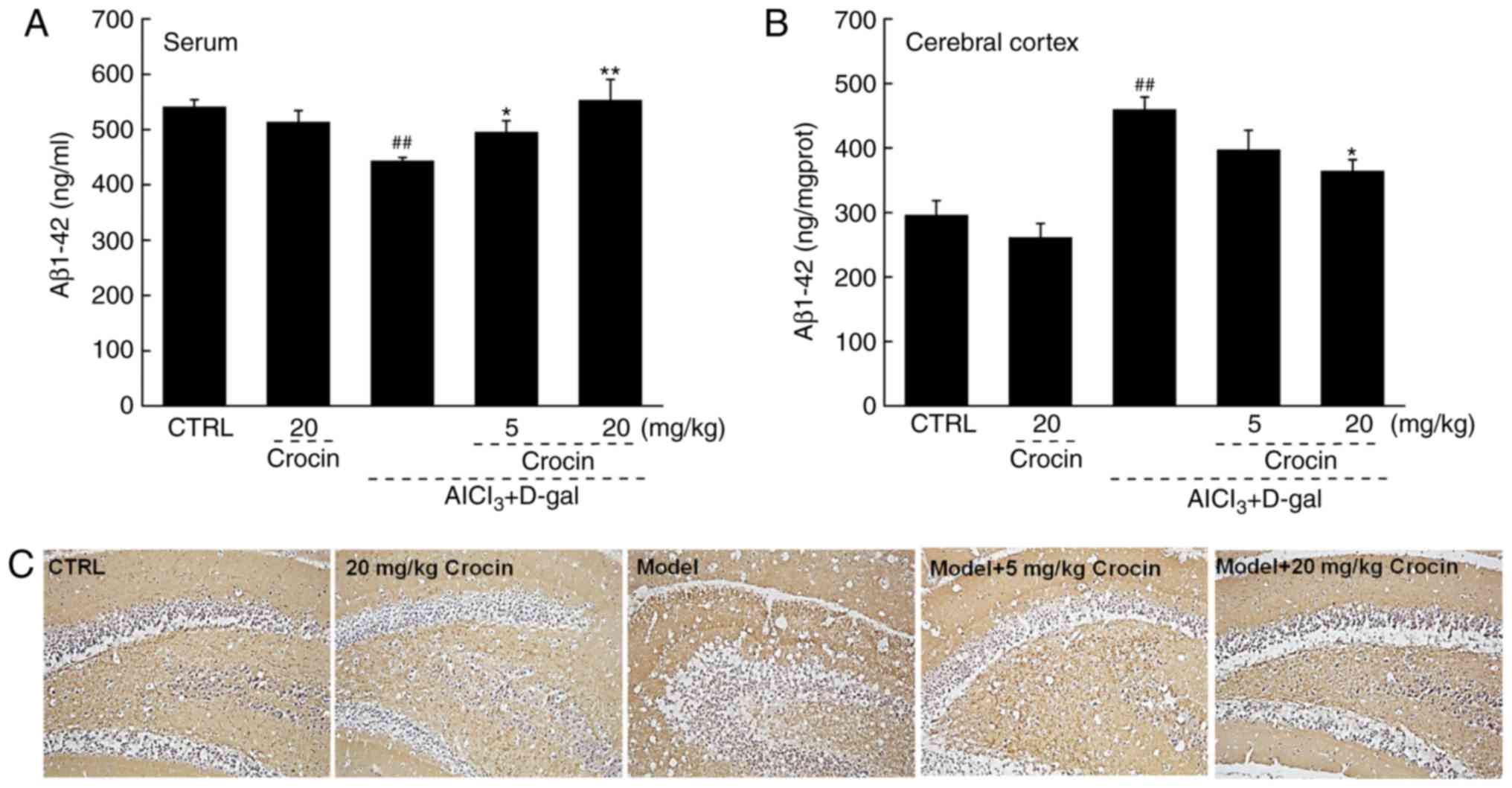
Aβ is the principal constituent of neuritic plaques,
which have been proposed, mainly on genetic grounds, to be central
to the pathogenesis of this form of dementia (27). In the AD mice, significantly lower
levels of Aβ1-42 in serum and significantly higher levels of Aβ1-42
in the cerebral cortex were observed compared with the control
(P<0.01; Fig. 5A and B).
Comparatively, crocin resulted in a >24.8% increment in the
serum levels of Aβ1-42 (P<0.05; Fig. 5A), and a >20.8% reduction in
the cerebral cortex levels of Aβ1-42 compared with the AD untreated
mice (P<0.05; Fig. 5B). The
results of the immunohistochemical assay revealed that the
deposition of Aβ1-42 in the hippocampal area was strongly
suppressed following 4 weeks of crocin administration (Fig. 5C).
Crocin regulated the levels of
cholinergic neurotransmitters in the serum, cerebral cortex and
hypothalamus of AD mice
The characteristic memory impairment of AD is
closely associated with the lack of cholinergic neurotransmitters
(28). Significantly lower levels
of Ach and ChAT and higher levels of AchE were observed in the
serum, cerebral cortex and hypothalamus of the AD mice compared
with the control groups, indicating injury to the cholinergic
system in the AD mice (P<0.05; Table I). Four-week crocin administration
signficantly restored the pathological alterations of the levels of
Ach, AchE and ChAT in the serum, cerebral cortex and hypothalamus
of the AD mice, suggesting the ability of crocin to mitigate
cholinergic dysfunction (P<0.05; Table I).
 | Table ICrocin regulated the levels of Ach,
AchE and ChAT in the serum, cerebral cortex and hypothalamus. |
Table I
Crocin regulated the levels of Ach,
AchE and ChAT in the serum, cerebral cortex and hypothalamus.
| Control | Crocin (20
mg/kg) | AlCl3 +
d-gal | Crocin (mg/kg)
|
|---|
| 5 | 20 |
|---|
| Serum | | | | | |
| Ach
(µg/ml) | 911.9±19.9 | 1057.0±24.4b | 768.8±13.4b | 826.3±15.7 | 1022.5±35.4f |
| AchE (nmol/l) | 138.9±2.2 | 150.9±6.0 | 179.3±5.0b | 121.8±8.2e | 113.2±8.5e |
| ChAT (pmol/l) | 284.7±10.6 | 283.8±8.6 | 233.2±3.9b | 273.5±9.1d | 281.6±2.9e |
| Cerebral
cortex | | | | | |
| Ach
(µg/mgprot) | 735.7±56.4 | 741.8±22.1 | 509.6±8.4a | 629.7±61.8 | 743.2±50.5d |
| AchE
(nmol/gprot) | 56.6±4.7 | 55.9±1.8 | 105.1±4.8c | 97.7±5.4 | 40.6±2.3f |
| ChAT
(pmol/gprot) | 229.3±15.6 | 207.2±11.3 | 167.6±3.3a | 252.2±25.3d | 221.6±1.5 |
| Hypothalamus | | | | | |
| Ach
(µg/mgprot) | 262.7±13.0 | 292.0±27.1 | 161.3±15.1c | 210.1±10.2 | 239.2±16.0d |
| AchE
(nmol/gprot) | 53.4±3.1 | 55.9±1.8 | 77.5±4.8b | 60.4±1.6d | 55.7±3.8e |
| ChAT
(pmol/gprot) | 144.3±5.4 | 165.8±6.5 | 80.9±6.5b | 109.6±3.3e | 115.0±5.2e |
Crocin regulated the levels of pro- and
anti-oxidative factors in the serum, cerebral cortex and
hypothalamus of AD mice
The brain is sensitive to oxidative stress, and
hyper-levels of ROS may arise in response to β-amyloid precursor
proteins and mitochondrial DNA, resulting in neuronal apoptosis
(29). In the AD-induced mice,
the levels of ROS were increased, and the levels of SOD and GSH-Px
were decreased significantly in the serum, cerebral cortex and
hypothalamus compared with the control mice (P<0.05; Table II). Encouragingly, crocin
significantly suppressed ROS levels, and enhanced the SOD and
GSH-Px levels in the serum, cerebral cortex and hypothalamus of the
AD mice compared with the AD alone mice (P<0.05; Table II).
| Table IICrocin regulated the levels of ROS,
SOD and GSH-Px in the serum, cerebral cortex and hypothalamus. |
Table II
Crocin regulated the levels of ROS,
SOD and GSH-Px in the serum, cerebral cortex and hypothalamus.
| Control | Crocin (20
mg/kg) | AlCl3 +
d-gal | Crocin (mg/kg)
|
|---|
| 5 | 20 |
|---|
| Serum | | | | | |
| ROS (U/ml) | 450.8±4.5 | 475.5±6.3 | 540.6±5.9b | 532.5±6.5 | 491.3±18.0d |
| SOD (U/ml) | 331.4±7.5 | 317.5±13.4 | 277.5±5.0b | 309.2±7.7d | 336.3±13.7e |
| GSH-Px (U/ml) | 780.0±21.4 | 756.3±17.7 | 632.8±20.8b | 685±14.9 | 775±24.4e |
| Cerebral
cortex | | | | | |
| ROS
(U/mgprot) | 286.3±24.8 | 323.8±2.1 | 402.5±23.8c | 389.2±35.3 | 303.3±7.3e |
| SOD
(U/mgprot) | 175.3±13.5 | 162.0±16.8 | 128.8±5.3a | 166.1±13.9d | 195.7±19.2e |
| GSH-Px
(U/mgprot) | 322.4±20.9 | 282.2±12.0 | 269.2±1.9a | 358.2±28.0d | 412.7±36.5e |
| Hypothalamus | | | | | |
| ROS
(U/mgprot) | 126.0±11.8 | 138.8±8.9 | 176.6±12.6b | 144.2±9.9d | 131.9±8.2d |
| SOD
(U/mgprot) | 132.7±10.9 | 118.6±4.3 | 68.2±3.7c | 92.6±6.5e | 107.4±9.8e |
| GSH-Px
(U/mgprot) | 212.8±20.7 | 220.1±7.7 | 121.9±10.2b | 199.0±18.6d | 211.4±15.9e |
Discussion
Although AD is the most prevalent neurodegenerative
disease, affecting over 47 million people globally, no satisfactory
treatment has been reported so far (30). Neuronal apoptosis is believed to
be one of the key events precipitating AD (31). In the present study, crocin
mitigated cell apoptosis and mitochondrial dysfunction in HT22
cells induced by l-Glu. A rise in extracellular
glutamate concentration may result in neuronal cell death by means
of oxidative stress, which is associated with mitochondrial
disorder and ROS accumulation (32). Mitochondria are involved in the
apoptosis and regulation of intracellular Ca2+
homeostasis (33). The overload
of intracellular Ca2+ results in mitochondrial
depolarization, which eventually intensifies oxidative
stress-induced cell apoptosis (34,35). ROS, produced by aerobic cells
during metabolism, is the foremost cause of oxidative stress
(36), resulting in neuronal
degeneration and cellular damage in AD (37). Cumulative oxygen damage may cause
mitochondrial dysfunction and simultaneously increase ROS
production (31). In the present
study, crocin inhibited the high expression of cleaved caspase-3,
Bad and Bax, and upregulated the low expression of Bcl-xL in
l-Glu-exposed HT22
cells. Bcl-2 family proteins are directly responsible for
mitochondrial apoptosis, and their ratio is considered to be a
biomarker indicating mitochondrial function (37,38). Consistent with a previous study,
upregulated Bax and Bad expression accelerates cell apoptosis by
increasing mitochondrial permeability; meanwhile, enhanced levels
of Bcl-xL help to inhibit cell apoptosis (38). Conversely, MMP dissipation caused
by oxidative damage sparks caspase-3, which is cleaved, eventually
resulting in apoptosis (39). It
has been previously demonstrated that glycyrrhizic acid and
Sparassis crispa polysaccharides exert neuroprotective
effects on differentiated PC12 cells, protecting those cells
against the toxicity of l-Glu by adjusting their MMP
levels (40,41). Taken together, the evidence
demonstrates that crocin protects HT22 cells against l-Glu-induced apoptosis mainly
through modulating mitochondrial apoptotic changes.
Furthermore, crocin upregulated the phosphorylation
levels of Akt and mTOR in 24-h l-Glu-exposed cells. Akt serves a
central role during apoptosis, and helps to regulate the levels of
Bcl-2 family members in AD to exert a neuro-protective effect
(30). mTOR, a serine/threonine
kinase, is highly conserved in evolution (42), and may be phosphorylated by
activated Akt. mTOR is responsible for regulating cell metabolism
via the mitochondrial-associated pathway (43). As reported, the activation of Akt
signaling may be controlled by ROS levels (44). These data suggest that Akt/mTOR
signaling is involved, at least partially, in crocin-mediated
neuroprotection of l-Glu-damaged HT22 cells.
Mice with AD, induced by chronic treatment with
AlCl3 and d-gal, exhibited AD-like
behaviors in the present study, particularly cognitive disorders
and dysmnesia, which were strongly improved by crocin treatment.
The AD-like behaviors associated with brain damage are mainly
caused by oxidative stress (45,46), cholinergic system impairment
(47,48) and the aggregation of Aβ (49). Abnormally high levels of
intracellular ROS accelerate AD progression (29); meanwhile, SOD and GSH-Px,
representative endogenous antioxidants, have been reported as the
first-line defense against oxidative stress (50). Encouragingly, crocin enhanced the
levels of SOD and GSH-Px, and reduced the levels of ROS in the
serum, cerebral cortex and hypothalamus of the AD model mice. Aβ
plaque deposition in the brain is closely associated with AD
(51), and may trigger a series
of cascade reactions including mitochondrial dysfunction and
oxidative stress (30,37). Aβ is normally cleared from the
brain into the periphery, while changes in its production or
clearance may result in its accumulation in the brain, accompanied
by reduced peripheral levels and the clinicopathological
manifestations of AD (52).
Herein, crocin modulated the concentrations of Aβ1-42 in the serum
and the brain. Its anti-oxidative activities were key to crocin’s
ability to improve the cognitive competence of the experimental AD
mice.
The central cholinergic system of patients with AD
is severely damaged (53). As a
major modulator of learning and memory abilities, Ach is
dynamically controlled by the terminating enzyme AchE and the
synthesizing enzyme ChAT (54).
As the level of AchE increases, the cognitive function of the
nervous system gradually decreases (28). ChAT is the most suitable factor
for monitoring cholinergic neurons (55). Oxidative stress may enhance the
expression of AchE (56), which
helps to promote the formation of Aβ and its aggregation into Aβ
plaques (57). The cholinergic
system in AD models induced by Aβ aggregation is severely damaged
(58). Altogether, the present
data have revealed that crocin regulates the levels of Ach, AchE
and ChAT in the serum, cerebral cortex and hypothalamus of AD mice,
suggesting the importance of cholinergic function.
The neuroprotective effects of crocin were
successfully confirmed in l-Glu-damaged HT22 cells, and the
ability of crocin to improve memory abilities and cognitive
functions was verified in AlCl3 and d-gal-induced AD mice. For the
first time to the best of our knowledge, the present study verified
the roles of oxidative stress-mediated apoptosis and cholinergic
functions in the neuroprotective mechanism of crocin.
Funding
The present study is supported by the Science
Foundation in Jilin Province of China (grant no. 20180101098JC) and
the Special Projects of Cooperation between Jilin University and
Jilin Province in China (grant no. SXGJSF2017-1).
Availability of data and materials
All data generated and analyzed during the present
study are included in this published article.
Authors’ contributions
DW and XCh designed the experiments. CW, XCa, WH, ZL
and FK performed the experiments. CW, XCa and WH processed data. DW
and CW wrote the paper. DW and XCh revised the paper.
Ethics approval and consent to
participate
Institution Animal Ethics Committee of Jilin
University approved the experimental protocol (approval no.
20160409).
Patient consent for publication
Not applicable.
Competing interests
The authors have declared that there are no
competing interests.
Acknowledgments
Not applicable.
Abbreviations:
|
Ach
|
acetylcholine
|
|
AchE
|
acetylcholinesterase
|
|
Akt
|
protein kinase B
|
|
AlC13
|
aluminum trichloride
|
|
Aβ
|
amyloid β
|
|
Bcl-xL
|
B-cell lymphoma-extra large
|
|
ChAT
|
choline acetyltransferase
|
|
d-gal
|
d-galactose
|
|
DMEM
|
Dulbecco’s modified Eagle’s medium
|
|
Fluo-4,AM
|
1-[2-Amino-5-(2,7-difluoro-6-hydroxy-3-oxo-9-xanthenyl)
phenoxy]-2-(2-amino-5-methylphenoxy) ethane-N,N,N′,N′-tetraacetic
acid, pentaacetoxymethyl ester
|
|
GSH-Px
|
glutathione peroxidase
|
|
HRP
|
horseradish peroxidase
|
|
HT22
|
Hippocampal neuronal cell line
|
|
l-Glu
|
l-glutamate
|
|
MMP
|
mitochondrial transmembrane
potential
|
|
mTOR
|
mammalian target of rapamycin
|
|
MTT
|
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium
bromide
|
|
ROS
|
reactive oxygen species
|
|
SOD
|
superoxide dismutase
|
|
TBS
|
t-butyldimethylsilyl
|
|
TBST
|
TBS + Tween
|
References
|
1
|
Holtzman DM, Morris JC and Goate AM:
Alzheimer’s disease: The challenge of the second century. Sci
Transl Med. 3:77sr712011. View Article : Google Scholar
|
|
2
|
Chen Z and Zhong C: Oxidative stress in
Alzheimer’s disease. Neurosci Bull. 30:271–281. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mines MA, Beurel E and Jope RS: Regulation
of cell survival mechanisms in Alzheimer’s disease by glycogen
synthase kinase-3. Int J Alzheimers Dis. 2011:8610722011.
|
|
4
|
Daulatzai MA: Cerebral hypoperfusion and
glucose hypo-metabolism: Key pathophysiological modulators promote
neurodegeneration, cognitive impairment, and Alzheimer’s disease. J
Neurosci Res. 95:943–972. 2017. View Article : Google Scholar
|
|
5
|
Tyagi N, Ovechkin AV, Lominadze D, Moshal
KS and Tyagi SC: Mitochondrial mechanism of microvascular
endothelial cells apoptosis in hyperhomocysteinemia. J Cell
Biochem. 98:1150–1162. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu X, Wang J, Lu C, Zhu C, Qian B, Li Z,
Liu C, Shao J and Yan J: The role of lysosomes in BDE 47-mediated
activation of mitochondrial apoptotic pathway in HepG2 cells.
Chemosphere. 124:10–21. 2015. View Article : Google Scholar
|
|
7
|
Luo P, Fei F, Zhang L, Qu Y and Fei Z: The
role of glutamate receptors in traumatic brain injury: Implications
for postsynaptic density in pathophysiology. Brain Res Bull.
85:313–320. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McDade E and Bateman RJ: Stop alzheimer’s
before it starts. Nature. 547:153–155. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Querfurth HW and LaFerla FM: Alzheimer’s
disease. N Engl J Med. 362:329–344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jesky R and Hailong C: Are herbal
compounds the next frontier for alleviating learning and memory
impairments? An integrative look at memory, dementia and the
promising therapeutics of traditional chinese medicines. Phytother
Res. 25:1105–1118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Man SC, Chan KW, Lu JH, Durairajan SS, Liu
LF and Li M: Systematic review on the efficacy and safety of herbal
medicines for vascular dementia. Evid Based Complement Alternat
Med. 2012:4262152012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ochiai T, Soeda S, Ohno S, Tanaka H,
Shoyama Y and Shimeno H: Crocin prevents the death of PC-12 cells
through sphingomyelinase-ceramide signaling by increasing
glutathione synthesis. Neurochem Int. 44:321–330. 2004. View Article : Google Scholar
|
|
13
|
Vahdati Hassani F, Naseri V, Razavi BM,
Mehri S, Abnous K and Hosseinzadeh H: Antidepressant effects of
crocin and its effects on transcript and protein levels of CREB,
BDNF, and VGF in rat hippocampus. Daru. 22:16–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Essa MM, Vijayan RK, Castellano-Gonzalez
G, Memon MA, Braidy N and Guillemin GJ: Neuroprotective effect of
natural products against Alzheimer’s disease. Neurochem Res.
37:1829–1842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qi Y, Chen L, Zhang L, Liu WB, Chen XY and
Yang XG: Crocin prevents retinal ischaemia/reperfusion
injury-induced apoptosis in retinal ganglion cells through the
PI3K/AKT signalling pathway. Exp Eye Res. 107:44–51. 2013.
View Article : Google Scholar
|
|
16
|
Ochiai T, Ohno S, Soeda S, Tanaka H,
Shoyama Y and Shimeno H: Crocin prevents the death of rat
pheochromyctoma (PC-12) cells by its antioxidant effects stronger
than those of alpha-tocopherol. Neurosci Lett. 362:61–64. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Heidari S, Mehri S and Hosseinzadeh H:
Memory enhancement and protective effects of crocin against
d-galactose aging model in the hippocampus of Wistar rats. Iran J
Basic Med Sci. 20:1250–1259. 2017.
|
|
18
|
Baghishani F, Mohammadipour A,
Hosseinzadeh H, Hosseini M and Ebrahimzadeh-Bideskan A: The effects
of tramadol administration on hippocampal cell apoptosis, learning
and memory in adult rats and neuroprotective effects of crocin.
Metab Brain Dis. 33:907–916. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang D, Li S, Chen J, Liu L and Zhu X: The
effects of astilbin on cognitive impairments in a transgenic mouse
model of Alzheimer’s disease. Cell Mol Neurobiol. 37:695–706. 2017.
View Article : Google Scholar
|
|
20
|
Zhang X, Chen Y, Cai G, Li X and Wang D:
Carnosic acid induces apoptosis of hepatocellular carcinoma cells
via ROS-mediated mitochondrial pathway. Chem Biol Interact.
277:91–100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ravindran S, Swaminathan K, Ramesh A and
Kurian GA: Nicorandil attenuates neuronal mitochondrial dysfunction
and oxidative stress associated with murine model of vascular
calcification. Acta Neurobiol Exp (Wars). 77:57–67. 2017.
|
|
22
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar
|
|
23
|
Kritis AA, Stamoula EG, Paniskaki KA and
Vavilis TD: Researching glutamate-induced cytotoxicity in different
cell lines: A comparative/collective analysis/study. Front Cell
Neurosci. 9:912015. View Article : Google Scholar
|
|
24
|
Peña-Blanco A and García-Sáez AJ: Bax, Bak
and beyond-mitochondrial performance in apoptosis. FEBS J.
285:416–431. 2018. View Article : Google Scholar
|
|
25
|
Heras-Sandoval D, Pérez-Rojas JM,
Hernández-Damián J and Pedraza-Chaverri J: The role of
PI3K/AKT/mTOR pathway in the modulation of autophagy and the
clearance of protein aggregates in neurodegeneration. Cell Signal.
26:2694–2701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brandeis R, Brandys Y and Yehuda S: The
use of the Morris Water Maze in the study of memory and learning.
Int J Neurosci. 48:29–69. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Butterfield DA, Drake J, Pocernich C and
Castegna A: Evidence of oxidative damage in Alzheimer’s disease
brain: Central role for amyloid beta-peptide. Trends Mol Med.
7:548–554. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yamini P, Ray RS and Chopra K: Vitamin D3
attenuates cognitive deficits and neuroinflammatory responses in
ICV-STZ induced sporadic Alzheimer’s disease. Inflammopharmacology.
26:39–55. 2018. View Article : Google Scholar
|
|
29
|
Tönnies E and Trushina E: Oxidative
stress, synaptic dysfunction, and Alzheimer’s disease. J Alzheimers
Dis. 57:1105–1121. 2017. View Article : Google Scholar
|
|
30
|
Xu T, Niu C, Zhang X and Dong M:
β-Ecdysterone protects SH-SY5Y cells against beta-amyloid-induced
apoptosis via c-Jun N-terminal kinase- and Akt-associated
complementary pathways. Lab Invest. 98:489–499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Koh CH, Whiteman M, Li QX, Halliwell B,
Jenner AM, Wong BS, Laughton KM, Wenk M, Masters CL, Beart PM, et
al: Chronic exposure to U18666A is associated with oxidative stress
in cultured murine cortical neurons. J Neurochem. 98:1278–1289.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Prasansuklab A, Meemon K, Sobhon P and
Tencomnao T: Ethanolic extract of Streblus asper leaves protects
against glutamate-induced toxicity in HT22 hippocampal neuronal
cells and extends lifespan of Caenorhabditis elegans. BMC
Complement Altern Med. 17:5512017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hung CH, Cheng SS, Cheung YT, Wuwongse S,
Zhang NQ, Ho YS, Lee SM and Chang RC: A reciprocal relationship
between reactive oxygen species and mitochondrial dynamics in
neurodegeneration. Redox Biol. 14:7–19. 2018. View Article : Google Scholar
|
|
34
|
Park HS, Cho HS and Kim TW: Physical
exercise promotes memory capability by enhancing hippocampal
mitochondrial functions and inhibiting apoptosis in obesity-induced
insulin resistance by high fat diet. Metab Brain Dis. 33:283–292.
2018. View Article : Google Scholar
|
|
35
|
Xu D and Peng Y: Apolipoprotein E 4
triggers multiple pathway-mediated Ca2+ overload, causes CaMK II
phosphorylation abnormity and aggravates oxidative stress caused
cerebral cortical neuron damage. Eur Rev Med Pharmaco.
21:5717–5728. 2017.
|
|
36
|
Fan LF, He PY, Peng YC, Du QH Ma YJ, Jin
JX, Xu HZ, Li JR, Wang ZJ, Cao SL, et al: Mdivi-1 ameliorates early
brain injury after subarachnoid hemorrhage via the suppression of
inflammation-related blood-brain barrier disruption and endoplasmic
reticulum stress-based apoptosis. Free Radic Biol Med. 112:336–349.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tong YN, Bai L, Gong R, Chuan JL, Duan XM
and Zhu YX: Shikonin protects PC12 cells against β-amyloid
peptide-induced cell injury through antioxidant and antiapoptotic
activities. Sci Rep. 8:262018. View Article : Google Scholar
|
|
38
|
Wang X, Wu J, Yu C, Tang Y, Liu J, Chen H,
Jin B, Mei Q, Cao S and Qin D: Lychee seed saponins improve
cognitive function and prevent neuronal injury via inhibiting
neuronal apoptosis in a rat model of Alzheimer’s disease.
Nutrients. 9:E1052017. View Article : Google Scholar
|
|
39
|
Chen JX and Yan SD: Pathogenic role of
mitochondrial [correction of mitochondral] amyloid-beta peptide.
Expert Rev Neurother. 7:1517–1525. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang D, Guo TQ, Wang ZY, Lu JH, Liu DP,
Meng QF, Xie J, Zhang XL, Liu Y and Teng LS: ERKs and
mitochondria-related pathways are essential for glycyrrhizic
acid-mediated neuropro-tection against glutamate-induced toxicity
in differentiated PC12 cells. Braz J Med Biol Res. 47:773–779.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu S, Wang D, Zhang J, Du M, Cheng Y, Liu
Y, Zhang N, Wang D and Wu Y: Mitochondria related pathway is
essential for polysaccharides purified from Sparassis crispa
mediated neuro-protection against glutamate-induced toxicity in
differentiated PC12 cells. Int J Mol Sci. 17:E1332016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pourtalebi Jahromi L, Sasanipour Z and
Azadi A: Promising horizon to alleviate Alzeheimer’s disease
pathological hallmarks via inhibiting mTOR signaling pathway: A new
application for a commonplace analgesic. Med Hypotheses.
110:120–124. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Laplante M and Sabatini DM: mTOR signaling
at a glance. J Cell Sci. 122:3589–3594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang J, Wang X, Vikash V, Ye Q, Wu D, Liu
Y and Dong W: ROS and ROS-mediated cellular signaling. Oxid Med
Cell Longev. 2016:43509652016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lu J, Zheng YL, Luo L, Wu DM, Sun DX and
Feng YJ: Quercetin reverses d-galactose induced neurotoxicity in
mouse brain. Behav Brain Res. 171:251–260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Praticò D, Uryu K, Sung S, Tang S,
Trojanowski JQ and Lee VM: Aluminum modulates brain amyloidosis
through oxidative stress in APP transgenic mice. FASEB J.
16:1138–1140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao F, Li XG, Zhang XY, Hou JD, Lin LF,
Gao Q and Luo HM: Combined administration of d-galactose and
aluminium induces Alzheimer-like lesions in brain. Neurosci Bull.
27:143–155. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kumar A, Dogra S and Prakash A: Protective
effect of curcumin (Curcuma longa), against aluminium toxicity:
Possible behavioral and biochemical alterations in rats. Behav
Brain Res. 205:384–390. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Banks WA, Niehoff ML, Drago D and Zatta P:
Aluminum complexing enhances amyloid beta protein penetration of
blood-brain barrier. Brain Res. 1116:215–221. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Islam MT: Oxidative stress and
mitochondrial dysfunction-linked neurodegenerative disorders.
Neurol Res. 39:73–82. 2017. View Article : Google Scholar
|
|
51
|
Dey A, Bhattacharya R, Mukherjee A and
Pandey DK: Natural products against Alzheimer’s disease:
Pharmaco-therapeutics and biotechnological interventions.
Biotechnol Adv. 35:178–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fei M, Jianghua W, Rujuan M, Wei Z and
Qian W: The relationship of plasma Abeta levels to dementia in
aging individuals with mild cognitive impairment. J Neurol Sci.
305:92–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Karthivashan G, Ganesan P, Park SY, Kim JS
and Choi DK: Therapeutic strategies and nano-drug delivery
applications in management of ageing Alzheimer’s disease. Drug
Deliv. 25:307–320. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ferreira-Vieira TH, Guimaraes IM, Silva FR
and Ribeiro FM: Alzheimer’s disease: Targeting the Cholinergic
System. Curr Neuropharmacol. 14:101–115. 2016. View Article : Google Scholar :
|
|
55
|
Fu AL, Li Q, Dong ZH, Huang SJ, Wang YX
and Sun MJ: Alternative therapy of Alzheimer’s disease via
supplementation with choline acetyltransferase. Neurosci Lett.
368:258–262. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang JY, Jiang H, Gao W, Wu J, Peng K,
Shi YF and Zhang XJ: The JNK/AP1/ATF2 pathway is involved in
H2O2-induced acetylcholinesterase expression during apoptosis. Cell
Mol Life Sci. 65:1435–1445. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lushchekina SV, Kots ED, Novichkova DA,
Petrov KA and Masson P: Role of Acetylcholinesterase in β-Amyloid
aggregation studied by accelerated molecular dynamics.
Bionanoscience. 7:396–402. 2017. View Article : Google Scholar
|
|
58
|
Nitta A, Itoh A, Hasegawa T and Nabeshima
T: beta-Amyloid protein-induced Alzheimer’s disease animal model.
Neurosci Lett. 170:63–66. 1994. View Article : Google Scholar : PubMed/NCBI
|