Introduction
Allergic asthma, a chronic airway inflammatory
disease, is a serious public health issue, and the prevalence of
asthma has increased considerably worldwide (1). Generally, the major characteristics
of allergic asthma are an airway inflammatory response, mucus
overproduction, obstruction and airway remodeling, which are
closely associated with high levels of Th2-type cytokines,
including interleukin (IL)-5/IL-13, eosinophil influx and serum
immunoglobulin E (IgE) production (2-4).
The increased level of monocyte chemoattractant protein (MCP-1) is
closely associated with inflammatory cell influx in the
pathogenesis of allergic asthma (5-7).
Nuclear factor-κB (NF-κB) is critical for the regulation of Th2
cytokine production, Th2 cell differentiation and mucus
overproduction (8). It is also
well documented that mitogen-activated protein kinases (MAPKs) are
important in the activation, proliferation and migration of
inflammatory cells, and the activation of MAPKs is significantly
higher in the lungs of allergic asthma animals compared with those
in normal controls (9,10). Heme oxygenase-1 (HO-1) is an
antioxidant protein that has anti-inflammatory properties, and
there is considerable evidence for its protective effect against
ovalbumin (OVA)-induced airway inflammation (11).
Natural compounds have attracted attention due to
their potent anti-inflammatory effects and minimal side-effects for
the treatment of chronic inflammatory diseases, including allergic
asthma (12,13). Cape gooseberry [Physalis
peruviana L. (PP)] is a species within the Solanaceae family,
which has potent antioxidant activity and has a variety of
biological effects, including antimycobacterial, anticancer and
anti-inflammatory activities (14-16). The levels of nitric oxide (NO) in
lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages were found
to be effectively downregulated by total extract from the calyces
of PP (17). In our previous
study, a methanol extract of PP markedly reduced the degree of
inflammatory cell recruitment, including inflammatory cytokines and
chemokines, which are considered important indicators of the
progression of airway inflammatory in chronic obstructive pulmonary
disease (COPD)-like models in animals (18). Therefore, the results from
previous studies suggest the possibility that treatment with PP may
effectively attenuate the inflammatory response in the lung tissues
of allergic asthma animal models. However, to the best of our
knowledge, no previous studies have investigated the
anti-inflammatory activity of PP in a mouse model of OVA-induced
allergic asthma. Therefore, in the present study, the ability of PP
to ameliorate pathological phenotypes, including airway
inflammation and mucus hypersecretion, was evaluated in an
OVA-induced asthma model.
Materials and methods
Preparation of PP
The fresh P. peruviana plant was collected
from the forest hills of the Katu Village, Lore Lindu National Park
(Central Sulawesi, Indonesia). The collected plant sample was
identified by the Center for Pharmaceutical and Medical Technology
(Tangerang, Indonesia), and authentication was confirmed by the
Herbarium Bogoriense (Bogor, Indonesia). Voucher specimens were
recorded as KRIB 0049496 and PMT 1884, which have been deposited in
the herbarium of the Korea Research Institute of Bioscience and
Biotechnology (Cheongju, Korea) and at the Center for
Pharmaceutical and Medical Technology and Herbarium Bogoriense
(18). Following drying and
grinding of the leaves of the plant. A total of 150 g of powder was
added to 150 ml of methanol, and extraction was performed by
maceration at room temperature for 18 h. The extract was filtered
and concentrated using a rotary evaporator (Laborota 4000;
Heidolph, Jakarta, Indonesia) under reduced pressure, thereby
obtaining 7.05 g of PP methanolic extract. In the following
experiments, the extract was dissolved in dimethyl sulfoxide (DMSO)
to a concentration of 20 mg/ml and then diluted to various
concentrations prior to use.
Mouse model of airway inflammation
induced by OVA
The experimental procedure was performed according
to the methods of Jeon et al (19). Briefly, healthy female BALB/c mice
(n=24; 6-weeks old; body weight, 16-18 g) were obtained from
Koatech Co. (Pyeongtaek-si, Korea) and were acclimated to a
specific pathogen-free condition (22-23°C and 55-60% humidity) with
free access to food and water at least 1 week prior to the
experiments. Mice were immunized twice intraperitoneally on days 0
and 14 with 0.2 ml of a vehicle solution containing 30 µg
OVA and 3 mg Alums (cat. no. 77161; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). At 7 days following the second immunization, OVA
inhalation was performed on three consecutive days using a
nebulizer (NE-U12; OMRON Corp., Tokyo, Japan) (1% OVA: alum-free
saline solution, 60 min/day, day 21-23). The oral administration of
PP or dexamethasone (DEX) was administered for six consecutive days
(day 18-23). The mice were randomly divided into four groups as
follows: Normal control group (NC), OVA (OVA
sensitization/challenge) group, DEX (OVA sensitization/challenge +
oral gavage of 1 mg/kg of DEX) group, and PP5 (OVA
sensitization/challenge + oral gavage of 5 mg/kg of PP) group. The
experimental protocol is presented in Fig. 1. The experimental procedures of
the present study were performed in accordance with procedures
approved by the Institutional Animal Care and Use Committee of the
Korea Research Institute of Bioscience and Biotechnology
(KRIBB-AEC-18054) and in compliance with the National Institutes of
Health Guidelines for the Care and Use of Laboratory Animals and
Korean National Laws for Animal Welfare.
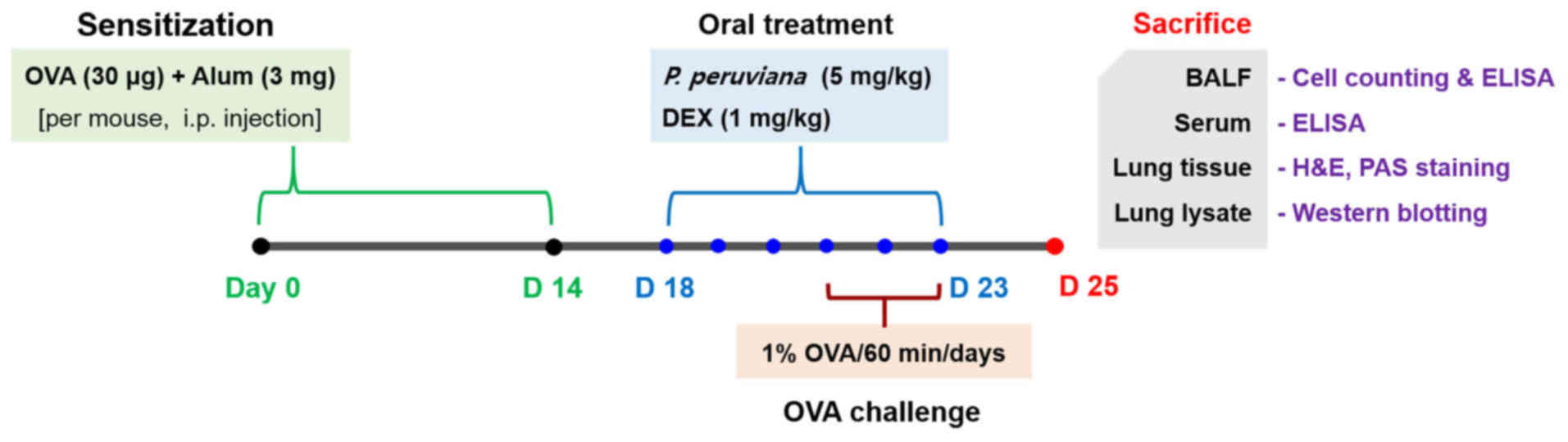 | Figure 1Experimental procedure for the
allergic asthma model and administration of PP or DEX. BALB/c mice
were divided into four groups (n=6 in each group). The mice were
orally administered with PP (5 mg/kg) or DEX (1 mg/kg) between days
18 and 23. On days 21-23, the mice were exposed to 1% OVA using a
nebulizer for 1 h each day. On day 25, the mice were sacrificed,
and the BALF, serum and lung tissues were acquired. PP, Physalis
peruviana L.; DEX, dexamethasone; OVA, ovalbumin; BALF,
bronchoalveolar lavage fluid; H&E, hematoxylin and eosin; PAS,
periodic acid-Schiff; D, day. |
Determination of inflammatory cell
counts
To count the numbers of inflammatory cells,
bronchoalveolar lavage fluid (BALF) collection was performed
according to the protocol described by Lee et al (20). Briefly, the collection of BALF was
performed 48 h following the final administration of DEX and PP by
tracheal cannulation and infusion with 700 µl of ice-cold
PBS (total volume, 1.4 ml). To distinguish the different cells, 0.1
ml BALF was centrifuged at 264 × g for 5 min at room temperature,
and the numbers of eosinophils and macrophages were counted using a
Diff-Quik® staining kit according to the manufacturer's
protocol (IMEB, Inc., Deerfield, IL, USA) and light microscope
(magnification, ×400).
Measurement of inflammatory cytokines in
the BALF and IgE in the serum
The levels of inflammatory mediators in the BALF,
including IL-4, IL-5 and IL-13, were evaluated using ELISA kits
according to the manufacturer's protocol. Blood samples were
collected from the mice 48 h following the final administration of
PP and DEX, and serum was prepared. Total IgE levels in the serum
were determined using an ELISA kit according to the manufacturer's
protocol (IL-4, IL-5 and IL-13 ELISA kits, cat. nos. M4000B, M5000
and M1300CB; R&D Systems, Inc., Minneapolis, MN, USA; and IgE
ELISA kit, cat. no. 432404; BioLegend, Inc., San Diego, CA,
USA).
Western blot analysis
The lung tissues were homogenized in CelLytic™ MT
cell lysis reagent (cat. no. c3228; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) prior to protein quantification. The protein
concentration was determined with the Pierce BCA Protein assay kit
(cat. no. 23225; Thermo Fisher Scientific, Inc.). Total protein was
separated by 10-12% SDS-PAGE (protein quantity, 20-50 µg),
and the samples were transferred onto PVDF membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked with 5%
skim milk solution for 1 h and incubated overnight at 4°C with
primary antibodies. The primary antibodies and dilution rates were
as follows: Anti-phosphorylated (p)-p38 (cat. no. 9211),
anti-p-extracellular signal-regulated kinase (ERK; cat. no. 4370),
anti-p-NF-κB p65 (cat. no. 3033) and anti-β-actin (cat. no. 4967)
at 1:1,000 dilutions (Cell Signaling Technology, Inc., Danvers, MA,
USA), anti-MCP-1 (cat. no. sc-28879), anti-p38 (cat. no. sc-7149),
anti-ERK (cat. no. sc-154), anti-p-c-Jun N-terminal kinase (JNK;
cat. no. sc-6254), anti-JNK (cat. no. sc-474), anti-NF-κB p65 (cat.
no. sc-372), anti-inhibitor of NF-κB (IκB-α; cat. no. sc-1643) and
anti-KEN-5 (cat. no. sc-59373) at 1:1,000 dilutions (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), and anti-HO-1 (cat. no.
PA5-27338; Thermo Fisher Scientific, Inc.) at 1:1,000 dilution. The
membranes were washed four times with TBST for 10 min and incubated
with HRP-conjugated secondary antibodies (goat anti-mouse, cat. no.
115-035-003; goat anti-rabbit, cat. no. 111-035-003: 1:2,000
dilution; Jackson ImmunoResearch Laboratories, Inc., West Grove,
PA, USA) at room temperature for 2 h. Finally, the membranes were
developed using an enhanced chemiluminescence kit (Thermo Fisher
Scientific, Inc.). The LAS-4000 luminescent image analyzer was used
to visualize all bands, and Fuji Multi Gauge version 3.0 (both
Fujifilm, Tokyo, Japan) was used to assess the band density.
Histological analysis
Following collection of the BALF, the lung tissues
were removed and fixed in 10% (v/v) neutral-buffered formalin
solution. For histological examination, the lung tissues were
embedded in paraffin and sectioned at a thickness of 4 µm.
The lung sections were then stained with hematoxylin (cat. no.
3580; BBC Biochemical Inc., Mount Vernon, WA, USA) and eosin (cat.
no. 6766007; Thermo Fisher Scientific, Inc.) solutions (H&E)
and a periodic acid-Schiff (PAS) stain solution kit (cat. no.
K7308; IMEB, Inc., San Marcos, CA, USA). The degree of inflammatory
cell influx and mucus production was visualized using a light
microscope (Leica Microsystems, Ltd., Milton Keynes, UK;
magnification, ×100 and ×200).
Cell culture
RAW264.7 mouse macrophages were obtained from the
American Type Culture Collection (Manassas, VA, USA) and grown in
DMEM supplemented with 10% heat-inactivated fetal bovine serum
(FBS) and a 1% (w/v) antibiotic-antimycotic solution (Invitrogen;
Thermo Fisher Scientific, Inc.) and incubated in a 5%
CO2 incubator at 37°C. To determine the levels of MCP-1,
the RAW 264.7 cells were seeded at 2.5×105 cells per well in a
6-well plate and incubated with 10% DMEM for 24 h. Following
treatment with PP (5, 10, 20 and 40 µg/ml) for 1 h, the
cells were incubated with LPS (200 ng/ml) for 20 h in a 5%
CO2 incubator at 37°C. The quantity of MCP-1 in the
supernatants was determined using an ELISA kit (BD Biosciences,
Franklin Lakes, NJ, USA).
Statistical analysis
The values are expressed as the mean ± standard
deviation. The statistical significance was determined by one-way
analysis of variance followed by a multiple comparison test with
Dunnett's adjustment using SPSS ver. 12.0 (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significance difference.
Results
Treatment with PP reduces the number of
eosinophils in the BALF of an OVA-induced airway inflammation
animal model
The numbers of inflammatory cells, including
eosinophils and macrophages, were significantly upregulated in the
BALF of the OVA-sensitized/challenged mice compared with those in
the NC group (Fig. 2A and B).
However, these levels were markedly downregulated in the PP-treated
mice compared with those in the OVA group. The mice treated with
DEX, which was used a positive control, also exhibited a decrease
in the numbers of eosinophils and macrophages in the BALF compared
with those in the OVA-challenged mice. The effects of PP were
similar to those of DEX treatment at 1 mg/kg.
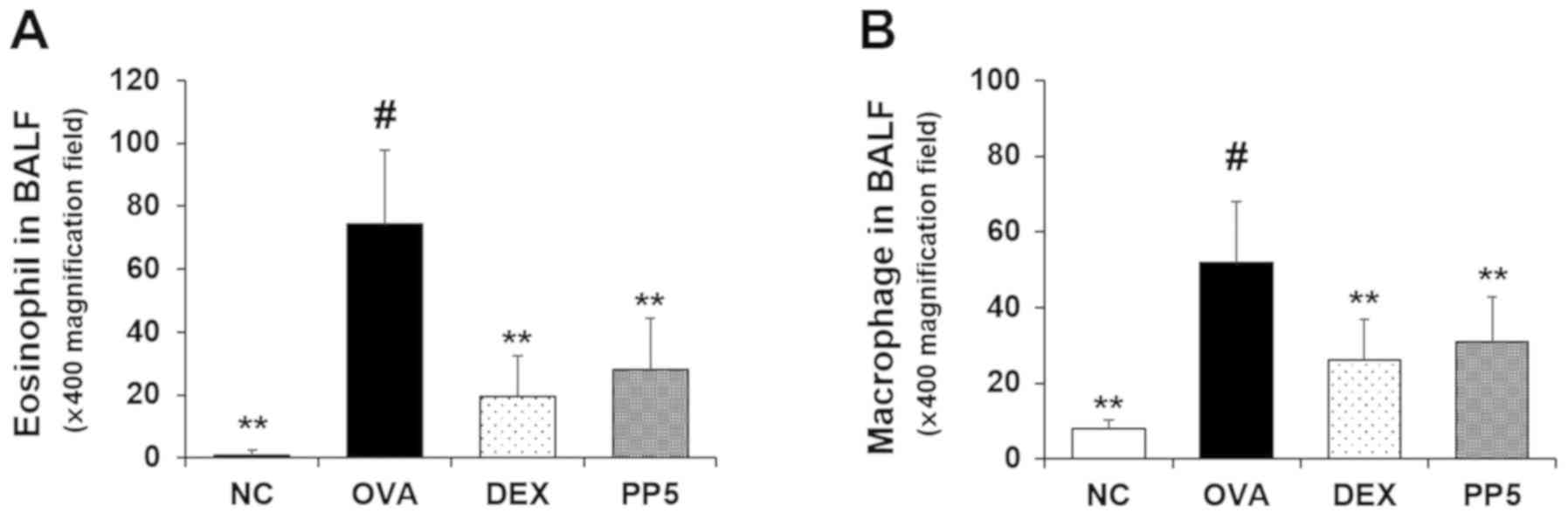 | Figure 2PP decreases the numbers of
inflammatory cells in the BALF of mice with OVA-induced airway
inflammation. Diff-Quik® staining was used to determine
the different cells in the BALF. (A) Eosinophil and (B) macrophage
counts were determined. Data are expressed as the mean ± standard
deviation (n=6). NC, normal control mice; OVA, mice administered
with OVA; DEX, mice administered with DEX (1 mg/kg) + OVA; PP5,
mice administered with PP (5 mg/kg) + OVA. #P<0.05,
vs. NC group; **P<0.01, vs. OVA group. PP,
Physalis peruviana L.; NC, normal control group; OVA,
ovalbumin; DEX, dexamethasone + OVA; PP5, 5 mg/kg PP + OVA; BALF,
bronchoalveolar lavage fluid. |
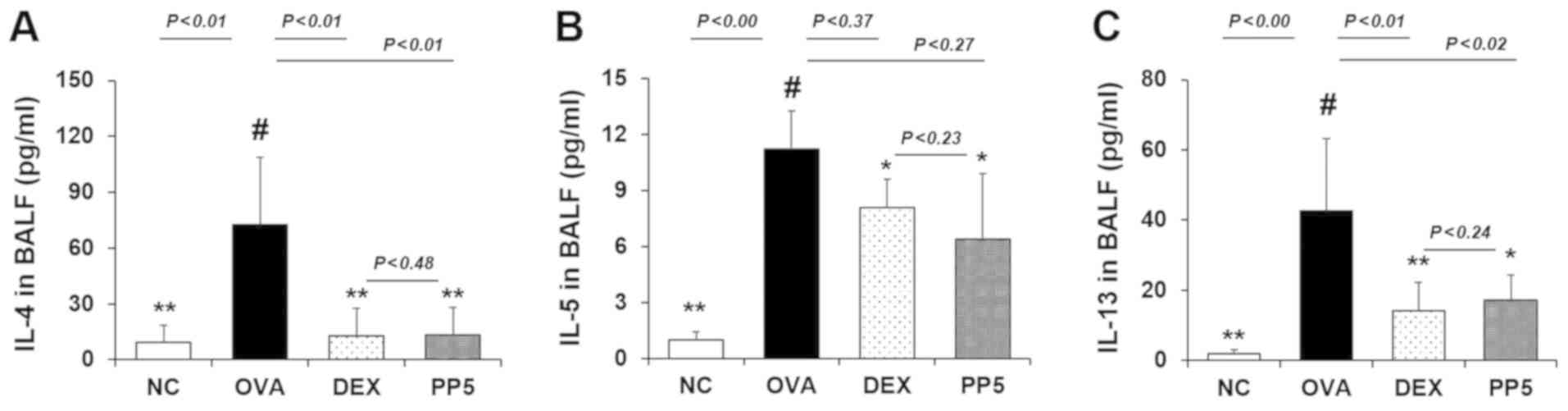
Treatment with PP decreases the levels of
Th2 cytokines in the BALF
As Th2 cytokines are involved in the airway
inflammatory response in allergic asthma (21-23), the production of Th2 cytokines,
including IL-4, IL-5 and IL-13, was examined by ELISA. The
OVA-challenged mice exhibited a significant increase in IL-4 and
IL-5 production compared with the NC mice. (Fig. 3A and B). However, this level was
significantly downregulated in the PP treatment group. Similar to
the results obtained for IL-4 and IL-5, the PP-treated mice
exhibited a marked reduction in the level of IL-13 compared with
the OVA-challenged mice (Fig.
3C). The inhibitory rates of IL-4 were 82.1% (DEX, 1 mg/kg) and
81.4% (PP, 5 mg/kg). The inhibitory rates of IL-5 were 31.3 % (DEX,
1 mg/kg) and 43.2% (PP, 5 mg/kg). The inhibitory rates of IL-13
were 66.7% (DEX, 1 mg/kg) and 59.6% (PP, 5 mg/kg). The inhibitory
effect of 5 mg/kg PP on Th2 cytokines was similar to that of 1
mg/kg DEX.
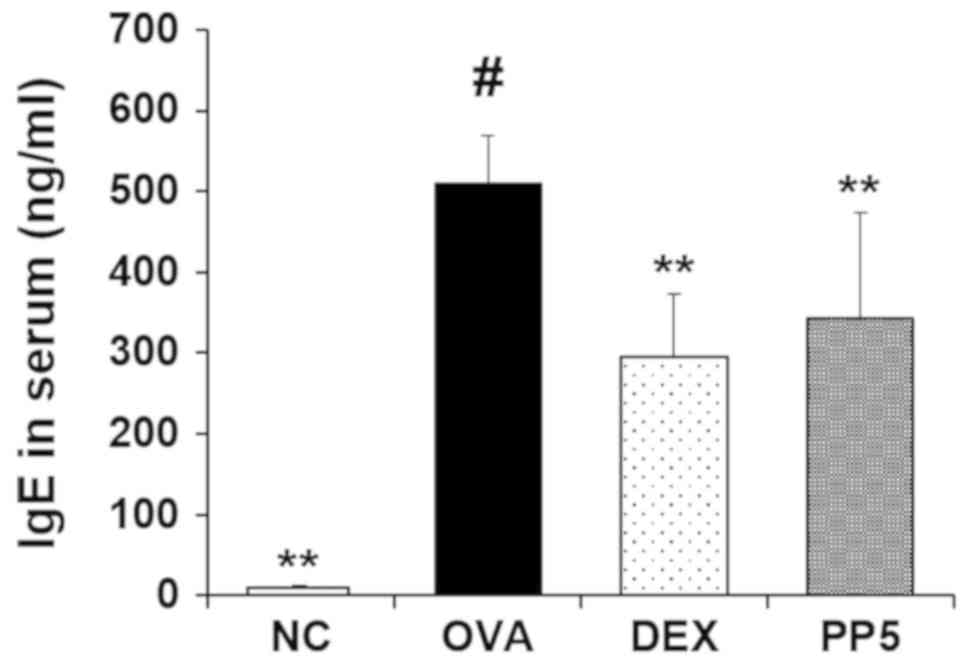
Treatment with PP attenuates the
production of IgE in the serum
ELISA was performed to evaluate the effect of PP on
the production of IgE. The results revealed that the production of
IgE in the serum was higher in the OVA-challenged mice compared
with that in the NC mice (Fig.
4). However, the increased level of IgE was significantly
reduced in the PP-treated mice compared with that in the
OVA-challenged mice.
Treatment with PP inhibits the levels of
inflammatory cell influx and mucus hypersecretion in the lungs
H&E staining was performed to determine whether
PP affects the inflammatory cell recruitment induced by OVA. In the
OVA-challenged mice, cell infiltration into the peribronchial
lesions in the lungs was markedly increased (Fig. 5A). However, this cell infiltration
was notably downregulated in the PP-treated mice. In order to
assess the effect of PP on mucus production, PAS staining was
performed. The results revealed the presence of mucus
hypersecretion in the bronchial airways of the OVA-challenged mice
(Fig. 5B). Notably, this level
was reduced in the lungs of mice in the PP treatment group compared
with that in the lungs of the OVA-challenged mice. The effect of 5
mg/kg PP was similar to that of treatment with 1 mg/kg DEX. Western
blot analysis was performed to determine the expression levels of
MCP-1 and KEN-5 in the different groups. The expression levels of
MCP-1 and KEN-5 were markedly enhanced in the OVA-challenged mice,
whereas these levels were lower in the PP group, as in the DEX
reference drug group (Fig. 5C-E).
These results indicate that PP may be useful in airway inflammation
via the suppression of inflammatory cell recruitment, chemokine
production and mucus production.
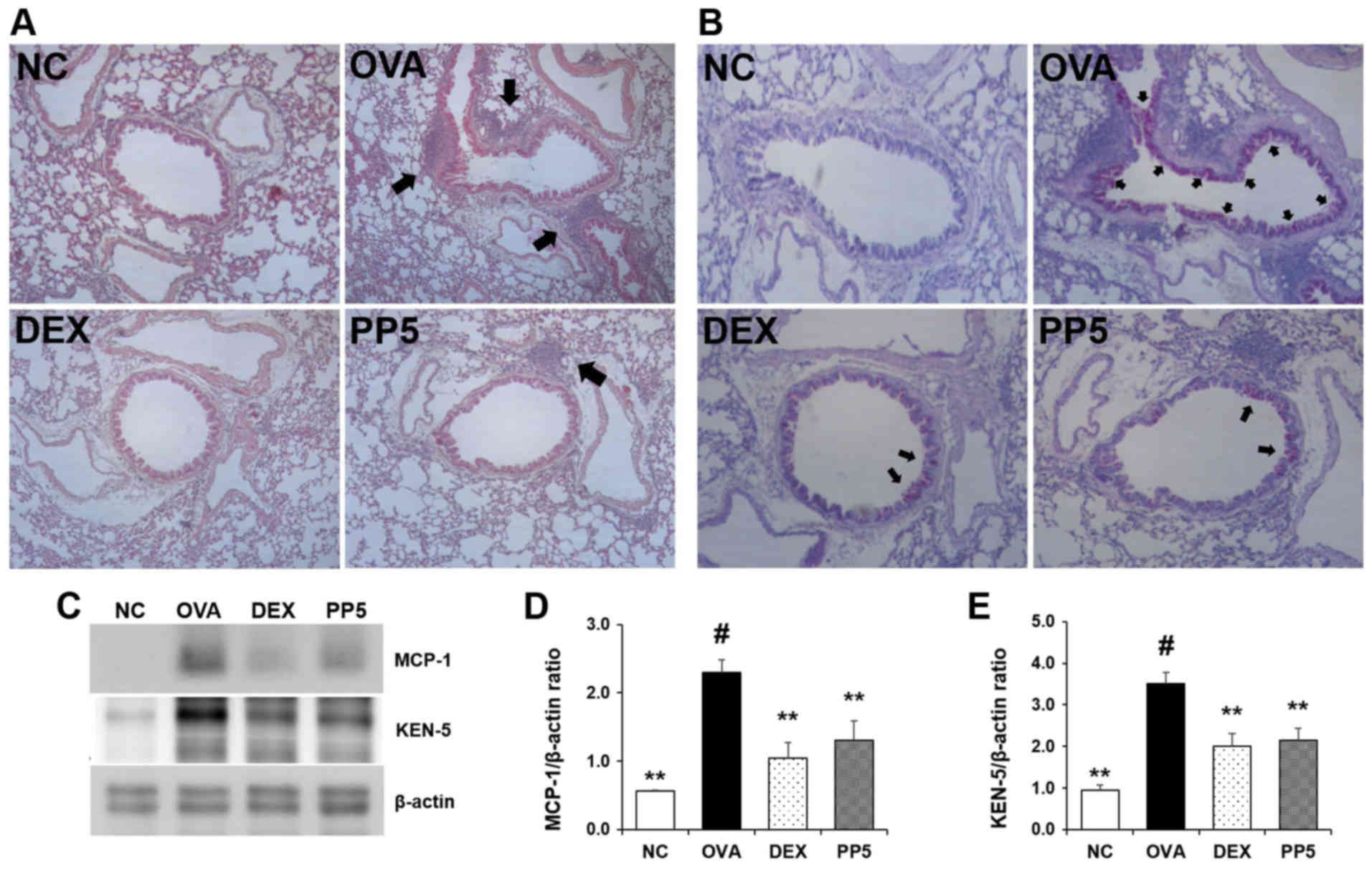 | Figure 5PP reduces the recruitment of
inflammatory cells and the production of mucus in the lungs of mice
with OVA-induced airway inflammation. (A) Levels of inflammatory
cell influx were confirmed with hematoxylin and eosin staining
(peribronchial lesion, magnification, ×100). The arrow indicates
the influx of inflammatory cells (B) Periodic acid-Schiff staining
was used to assess mucus production (peribronchial lesion,
magnification, ×200). The arrow indicates the production of mucus.
(C) Western blot analysis was used to determine the protein
expression levels of MCP-1 and KEN-5 in the lung tissue samples.
Quantitative analysis of (D) MCP-1 and (E) KEN-5 was performed by
densitometric analysis. #P<0.05, vs. NC group;
**P<0.01 vs. OVA group. PP, Physalis peruviana
L.; NC, normal control group; OVA, ovalbumin; DEX, dexamethasone +
OVA; PP5, 5 mg/kg PP + OVA; MCP-1, monocyte chemoattractant
protein-1. |
Treatment with PP reduces the activation
of NF-κB in the lungs
It has been reported that NF-κB is activated
following the activation of its inhibitor IκB (24,25). Upregulation in the phosphorylation
of IκB and NF-κB has been identified in the lung samples of
OVA-induced allergic asthma mice (26). To evaluate the effect of PP on the
OVA-induced NF-κB signaling pathway, the level of IκB
phosphorylation was investigated using western blot analysis. As
shown in Fig. 6A and B, the level
of IκB phosphorylation was significantly increased compared with
that in the NC mice. However, treatment with PP effectively
downregulated this level. The increase in OVA-induced NF-κB
phosphorylation was significantly suppressed by PP treatment
(Fig. 6A and C).
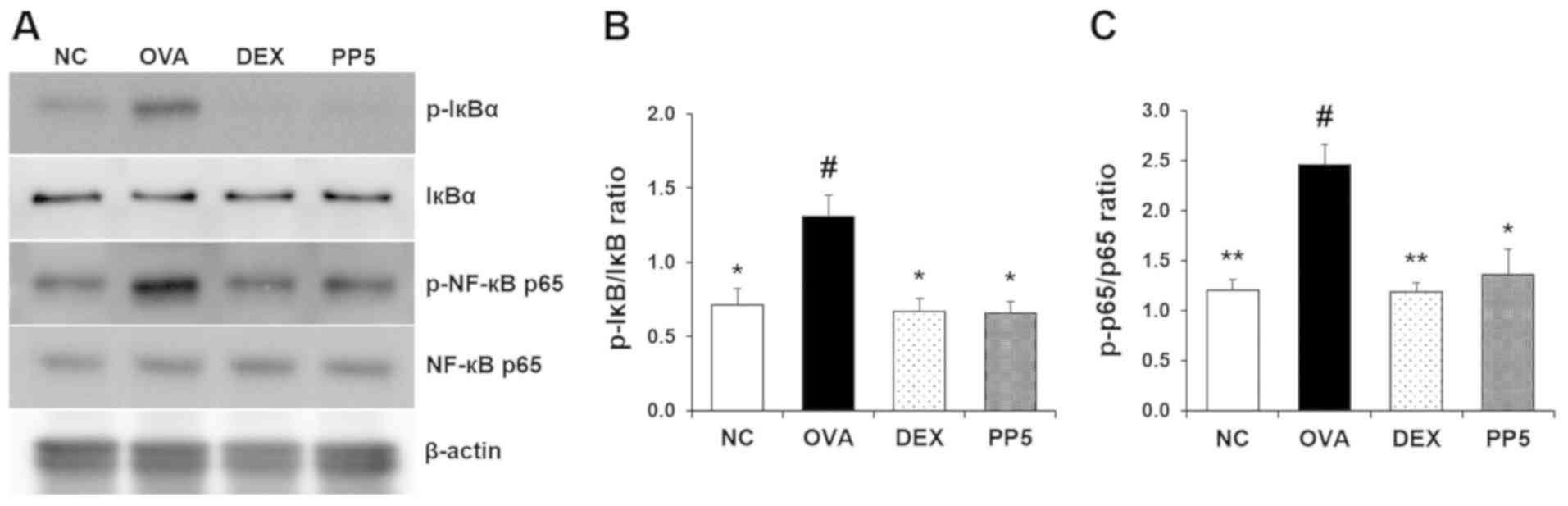 | Figure 6PP inhibits the activation of NF-κB
in the lungs of mice with OVA-induced airway inflammation. (A)
Levels of NF-κB and IκB activation were determined by western blot
analysis. Quantitative analysis of (B) p-IκB and (C) p-p65 was
performed by densitometric analysis. #P<0.05, vs. NC
group; *P<0.05 and **P<0.01, vs. OVA
group. PP, Physalis peruviana L.; NF-κB, nuclear factor-κB;
IκB, inhibitor of NF-κB; p-, phosphorylated; NC, normal control
group; OVA, ovalbumin; DEX, dexamethasone + OVA; PP5, 5 mg/kg PP +
OVA. |
Treatment with PP inhibits the activation
of p38 and JNK in the lungs
The levels of MAPK activation were subsequently
evaluated, and it was confirmed that the activation of p38, JNK and
ERK were upregulated in the lungs of the OVA-challenged mice
(Fig. 7A). Notably, treatment
with PP effectively inhibited the activation of p38 and JNK induced
by OVA in mice. By contrast, the activation of ERK was not
significantly affected by PP treatment.
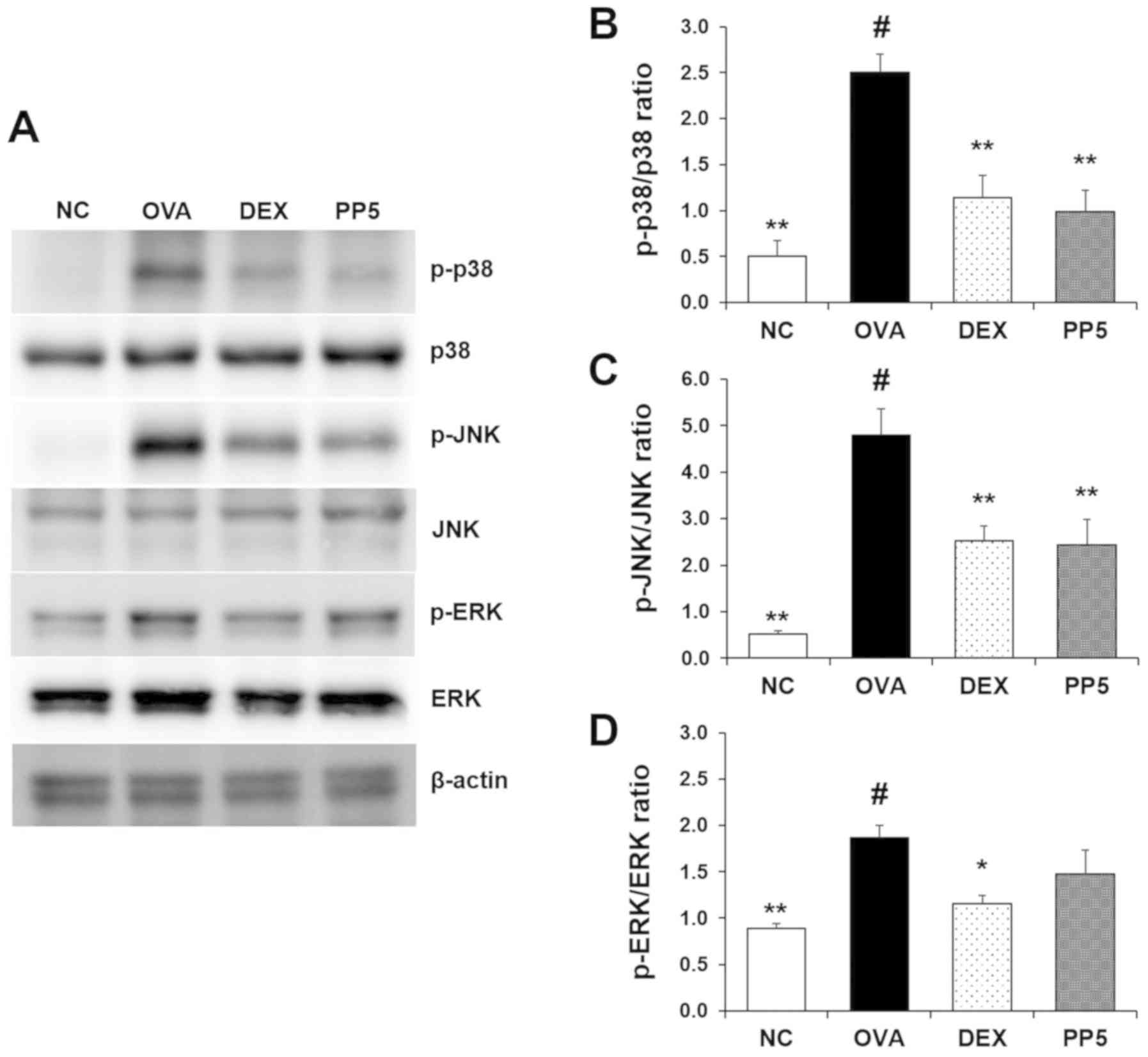 | Figure 7PP downregulates the activation of
p38 mitogen-activated protein kinases in the lungs of mice with
OVA-induced airway inflammation. (A) Levels of p38, ERK and JNK
activation were determined by western blot analysis. Quantitative
analysis of (B) p-p38, (C) p-JNK and (D) p-ERK was performed by
densitometric analysis. #P<0.05, vs. NC group;
*P<0.05 and **P<0.01, vs. OVA group.
PP, Physalis peruviana L.; ERK, extracellular
signal-regulated kinase; JNK, c-Jun N-terminal kinase; p-,
phosphorylated; NC, normal control group; OVA, ovalbumin; DEX,
dexamethasone + OVA; PP5, 5 mg/kg PP + OVA. |
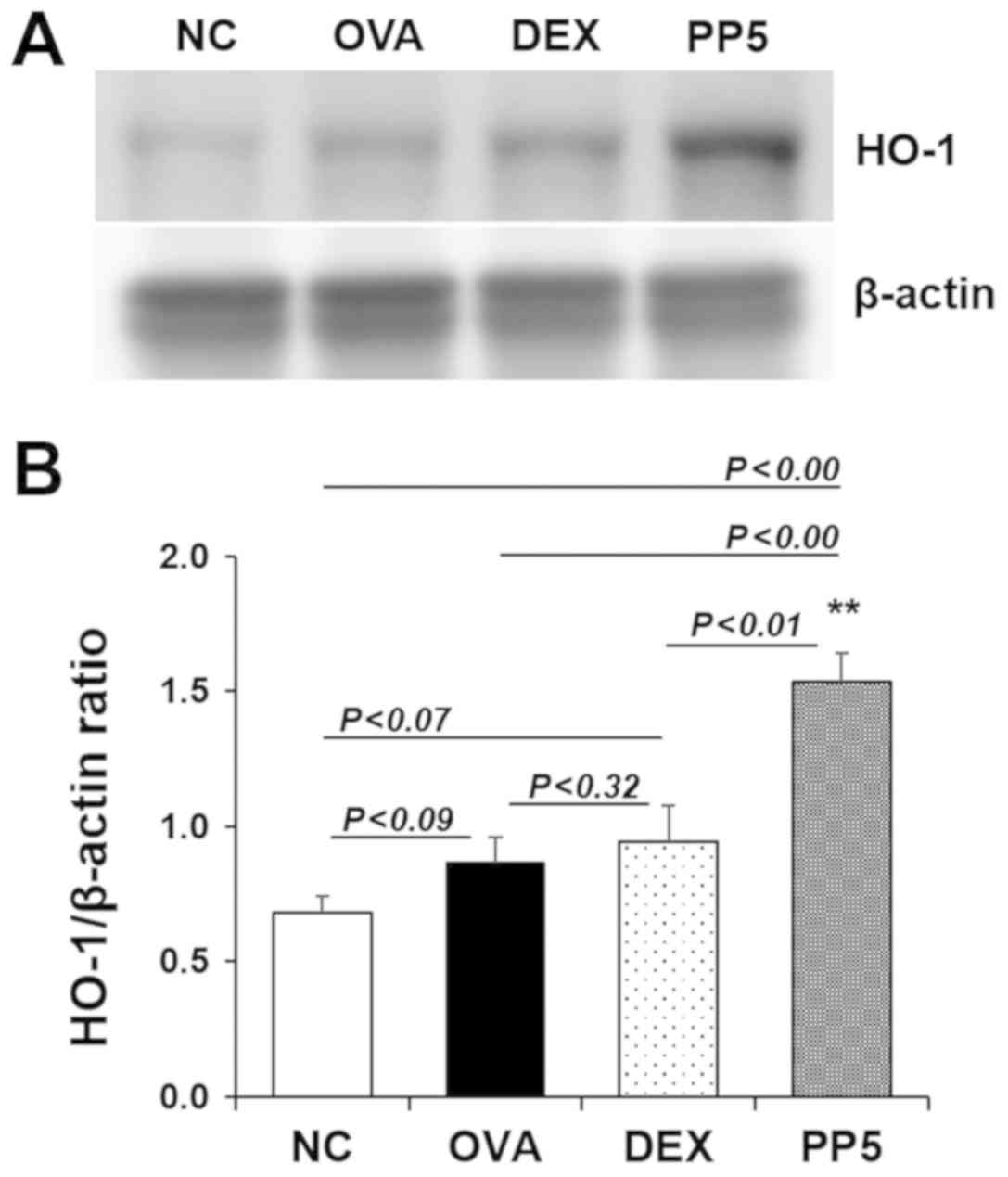
Treatment with PP leads to the induction
of HO-1 in the lungs
Given the importance of HO-1 in inflammatory lung
diseases (27,28), the present study evaluated the
effect of PP on the induction of HO-1. As shown in Fig. 8A and B, marginal increases in the
expression of HO-1 were observed in the OVA and DEX groups compared
with that in the NC group. These increases were not statistically
significant (NC, vs. OVA group, P<0.09; NC, vs. DEX group,
P<0.07). No significant difference was observed between the OVA
and DEX group (P<0.32). Of note, the expression of HO-1 was
significantly upregulated in the PP-treated group compared with
that in the NC, OVA and DEX groups (NC, vs. PP group, P<0.01;
OVA, vs. PP group, P<0.01; DEX, vs. PP group, P<0.01).
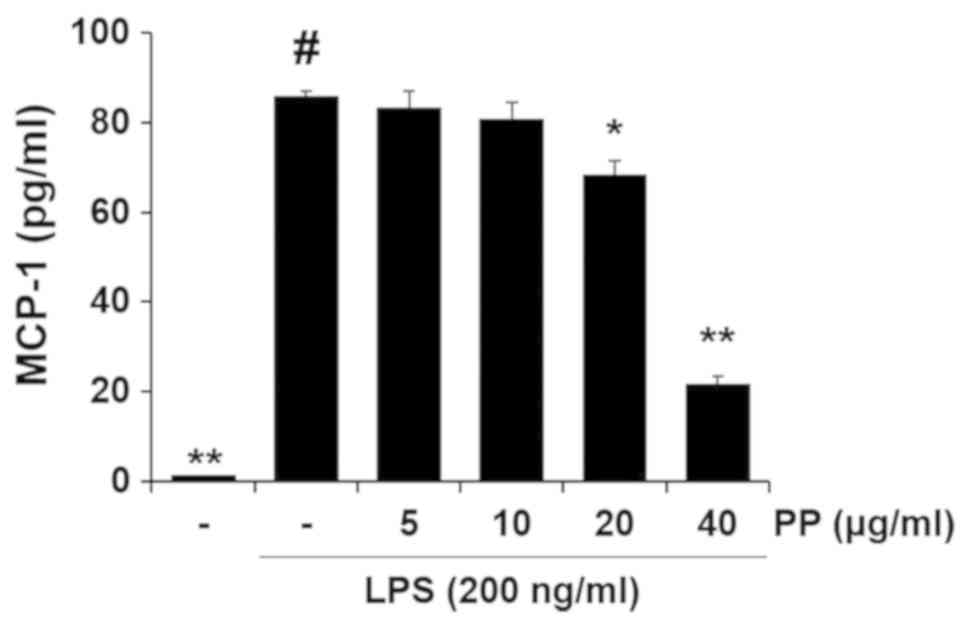
Inhibitory effect of PP on LPS-stimulated
inflammatory molecule in RAW264.7 macrophages
It was confirmed that PP can regulate the
recruitment of macrophages and the production of MCP-1 in an
OVA-induced animal model (Fig. 2B
and 5C). Therefore, the present
study subsequently evaluated the effect of PP on the production of
MCP-1 in activated macrophages. As shown in Fig. 9, there was a significant increase
in MCP-1 in LPS-stimulated RAW264.7 macrophages, whereas
pretreatment with PP effectively decreased the levels of MCP-1.
Discussion
Airway inflammation and mucus hypersecretion are the
major characteristic of allergic asthma. Accordingly, the aim of
the present study was to evaluate the protective effects of PP
against the progression of these characteristics in an experimental
animal model of OVA-induced allergic asthma. The results
demonstrated that PP ameliorated airway inflammation and the
overproduction of mucus, and indicated that the improvement effects
were correlated with the inactivation of NF-κB/p38/pJNK and the
induction of HO-1.
Airway inflammation is regarded as a typical feature
of inflammatory lung diseases, including allergic asthma (29). Eosinophil accumulation is the
prominent characteristic of the airway inflammatory response and
eosinophil-derived inflammatory mediators and cytokines lead to
mucus hypersecretion and bronchoconstriction (30-32). Macrophages are important in the
airway inflammatory response and contribute to the process of
tissue remodeling in allergic asthma (33,34). The increased levels of eosinophils
and macrophages in the BALF are the main features of the
OVA-induced allergic asthma animal model (8). Based on this, the present study
investigated the inhibitory effects of PP on the OVA-induced
eosinophil and macrophage recruitment. The results showed that PP
had regulatory effects on the recruitment of inflammatory cells,
including eosinophils and macrophages, in the allergic asthma
animal model (Fig. 2A and B).
The abnormal Th2 type response is also a key feature
of the allergic reaction (4,35).
Th2-derived IL-5 is responsible for the maturation, survival and
recruitment of eosinophils (21).
It is well known that IL-13 stimulates the B lymphocyte-derived
production of IgE (22) and has a
primary stimulatory action in the pathophysiology of allergic
asthma by causing mucus hypersecretion (36). Therefore, regulation of the
abnormal production of Th2-derived IL-5 and IL-13 are important in
the alleviation of the allergic response. In the present study, it
was confirmed that PP treatment effectively inhibits the
OVA-induced production of IL-5 and IL-13 (Fig. 3). These findings suggest that PP
has an immunoregulatory effect in an allergic asthma model.
It is well documented that IgE amplifies the
allergic response, and its immunomodulatory functions include the
activation of mast cells and the increase of B cell-derived
allergen uptake for antigen presentation (37). IgE also stimulates inflammatory
cells to produce the inflammatory molecules histamine and
leukotrienes, which cause inflammatory cell influx and mucus
hypersecretion (38). Therefore,
it is important to control the levels of IgE to prevent or treat
allergic asthma. The present study focused on the inhibitory effect
of PP on the overproduction of IgE. The ELISA results show that PP
can act as a potent inhibitor of IgE (Fig. 4).
Macrophage activation is not only associated with
the airway inflammatory response but also with airway remodeling
(39), and cell-derived MCP-1
acts as a potent eosinophil chemoattractant (40,41). Therefore, the regulation of
macrophage activation and overproduction of MCP-1 is an alternative
therapeutic strategy for treating allergic asthma. In a previous
study, PP was shown to exert anti-inflammatory effects on
LPS-stimulated macrophage cells by reducing NO (17). In our previous study, PP was shown
to inhibit the production of MCP-1in a COPD animal model (18). In the present study, it was
confirmed that PP reduced the number of inflammatory cells,
including eosinophils and macrophages, in the BALF of OVA-induced
allergic asthma mice (Fig. 2).
Based on previous results and those of the present study, it was
expected that PP exerts an inhibitory effect on OVA-induced
inflammatory cell influx and the overexpression of MCP-1. As
expected, PP treatment decreased the levels of inflammatory cell
influx, T cell marker expression and the expression of MCP-1
compared with the OVA group (Fig. 5A
and C) and effectively downregulated the production of MCP-1 in
LPS-stimulated RAW264.7 macrophages (Fig. 9), indicating that PP may be a
potent inhibitor of MCP-1 in airway inflammatory diseases,
including allergic asthma.
The original function of mucus is to protect the
lungs against foreign particles (42). However, the overproduction of
mucus interrupts the airflow and leads to airway obstruction
(43). Goblet cell hyperplasia is
responsible for the overproduction of mucus and airway
hyper-responsiveness (AHR), which are the major hallmarks of
allergic asthma (44). In the
present study, the PAS staining results revealed that treatment
with PP ameliorated OVA-induced mucus hypersecretion (Fig. 5B).
In the inflammatory response, IκBα is phosphorylated
and rapidly degraded, and this event leads to the phosphorylation
of NF-κB and its nuclear translocation, and eventually induces the
production of inflammatory molecules (4,45,46). Downregulation of the activation of
NF-κB is closely associated with the amelioration of airway
inflammation by suppressing inflammatory cell recruitment,
expression of inflammatory molecules and AHR in OVA-challenged
lungs (1,4,45).
Therefore, NF-κB is a promising molecular target in the treatment
of allergic asthma. Castro et al reported that PP leads to
the downregulation of NF-κB-dependent inflammatory molecules,
including tumor necrosis factor-α, cyclooxygenase-2 (COX-2), and
inducible nitric oxide synthase (iNOS) in an in vivo model
(17). In our previous study, the
regulatory effect of PP on NF-κB-dependent molecules, including
iNOS, COX-2 and MCP-1, were observed in a COPD-like animal model
(18). In present study, the
regulatory effect of PP on the OVA-induced expression of MCP-1 was
confirmed (Fig. 4C). Therefore,
it was expected that PP can regulate the activation of NF-κB. In
the present study, it was found that the NF-κB signaling pathway
was activated in the OVA group, leading to the phosphorylation of
IκB and NF-κB. However, treatment with PP significantly suppressed
the phosphorylation of IκB and NF-κB (Fig. 6). These results suggest that
suppression of the NF-κB signaling pathway is closely associated
with the protective effect of PP in an allergic asthma animal
model.
The MAPK signaling pathway is also considered to be
linked with the inflammatory response (47). It has been shown that MAPKs are
crucial in the pathophysiology of asthma by controlling
inflammatory molecules; their activation has been detected in in
vitro and in vivo asthma models, and inhibition of their
activation inhibits the airway inflammatory response (6,48).
The results of the present study revealed that treatment with PP
decreased the OVA-induced activation of MAPKs (Fig. 7). In particular, the levels of p38
and JNK activation were markedly downregulated following
administration with PP in the lungs of the OVA-induced mice. These
findings suggest that PP treatment disrupts the activity of p38 and
JNK.
Previously in vivo experiments demonstrated
that treatment with antioxidants ameliorate allergic airway
inflammation by upregulating the expression of HO-1 in a mouse
model of asthma, and increased levels of HO-1 in the group treated
with OVA and antioxidant were higher compared with those in normal
control or OVA-treated groups in a concentration-dependent manner
(1,27,28). Based on these results, the present
study examined the effect of PP on the induction of HO-1. The
results showed that the effects on the expression of HO-1 in the
NC, OVA and OVA + PP administration group were similar to those of
previous studies (1,27,28). In particular, treatment with 5
mg/kg PP markedly increased the expression of HO-1 compared with
that in the NC, OVA or DEX groups. This result suggests that the
upregulation of HO-1 may be associated with an improved airway
inflammatory response in the OVA-induced allergic asthma model
(Fig. 8). Therefore, PP may be
considered as an essential herbal plant for the prevention of
airway inflammation.
In conclusion, the results of the present study
demonstrated a significant reduction in the eosinophil count of
BALF samples from the PP-treated group. Similarly, PP suppressed
the lung infiltration of inflammatory cells. The results also
showed significant alleviation in the levels of IL-4, IL-5 and
IL-13 in the BALF and in the level of IgE in serum samples of the
PP-treated group. Furthermore, PP downregulated the levels of mucus
hypersecretion. These effects were associated with the inhibited
activation of NF-κB/p38-pJNK MAPK and upregulation of the
expression of HO-1. These findings provide evidence that PP
possesses potent anti-inflammatory effects and exerts protective
effects against OVA-induced asthmatic symptoms.
Funding
The present study was supported by a grant from the
Korea Research Institute of Bioscience and Biotechnology Research
Initiative Program (grant no. KGM5521911) and the Ministry of
Health and Welfare (grant no. HI14C1277) of the Republic of
Korea.
Availability of data and materials
All data generated and/or analyzed during the
present study are included in this published article.
Authors' contributions
HAP and OKK performed the experiments and
contributed to the interpretation of the results. HWR contributed
to the extraction of P. peruviana L. JHM, MWP and MHP
contributed to the acquisition of data. JHP, SC, IP and PY made
substantive intellectual contribution to the published study. SRO
provided conceptual advice. JWL designed the study and wrote the
manuscript. KSA supervised the project and was involved in revising
it critically for important intellectual content. All authors
discussed the results and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Institutional
Animal Care and Use Committee of the Korea Research Institute of
Bioscience and Biotechnology.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
OVA
|
ovalbumin
|
|
BALF
|
bronchoalveolar lavage fluid
|
|
IL-5
|
interleukin 5
|
|
IL-13
|
interleukin 13
|
|
IgE
|
immunoglobulin E
|
|
MCP-1
|
monocyte chemoattractant protein-1
|
|
AHR
|
airway hyperresponsiveness
|
|
NF-κB
|
nuclear factor-κB
|
|
IκB
|
inhibitor of NF-κB
|
|
MAPKs
|
mitogen-activated protein kinases
|
|
HO-1
|
heme oxygenase-1
|
|
PP
|
Physalis peruviana L
|
References
|
1
|
Qian J, Ma X, Xun Y and Pan L: Protective
effect of forsythiaside A on OVA-induced asthma in mice. Eur J
Pharmacol. 812:250–255. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu J, Wei Y, Luo Q, Xu F, Zhao Z, Zhang
H, Lu L, Sun J, Liu F, Du X, et al: Baicalin attenuates
inflammation in mice with OVA-induced asthma by inhibiting NF-κB
and suppressing CCR7/CCL19/CCL21. Int J Mol Med. 38:1541–1548.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ntontsi P, Papathanassiou E, Loukides S,
Bakakos P and Hillas G: Targeted anti-IL-13 therapies in asthma:
Current data and future perspectives. Expert Opin Investig Drugs.
27:179–186. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gu X, Zhang Q, Du Q, Shen H and Zhu Z:
Pinocembrin attenuates allergic airway inflammation via inhibition
of NF-κB pathway in mice. Int Immunopharmacol. 53:90–95. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ji P, Hu H, Yang X, Wei X, Zhu C, Liu J,
Feng Y, Yang F, Okanurak K, Li N, et al: AcCystatin, an
immunoregulatory molecule from Angiostrongylus cantonensis,
ameliorates the asthmatic response in an aluminium
hydroxide/ovalbumin-induced rat model of asthma. Parasitol Res.
114:613–624. 2015. View Article : Google Scholar
|
|
6
|
Rajajendram R, Tham CL, Akhtar MN,
Sulaiman MR and Israf DA: Inhibition of epithelial CC-Family
chemokine synthesis by the synthetic chalcone DMPF-1 via disruption
of NF-κB nuclear translocation and suppression of experimental
asthma in mice. Mediators Inflamm. 2015.176926:2015.
|
|
7
|
Lian Q, Jiang W, Cheng Y, Cao H, Liu M,
Wang J, Li Y, Song X and Wang F: A novel pentapeptide originated
from calf thymus named TIPP shows an inhibitory effect on lung
allergic inflammation. Int Immunopharmacol. 24:256–266. 2015.
View Article : Google Scholar
|
|
8
|
Zhang Q, Wang L, Chen B, Zhuo Q, Bao C and
Lin L: Propofol inhibits NF-κB activation to ameliorate airway
inflammation in ovalbumin (OVA)-induced allergic asthma mice. Int
Immunopharmacol. 51:158–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duan W, Chan JH, Wong CH, Leung BP and
Wong WS: Anti-inflammatory effects of mitogen-activated protein
kinase kinase inhibitor U0126 in an asthma mouse model. J Immunol.
172:7053–7059. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Duan W and Wong WS: Targeting
mitogen-activated protein kinases for asthma. Curr Drug Targets.
7:691–698. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye P, Yang XL, Chen X and Shi C:
Hyperoside attenuates OVA-induced allergic airway inflammation by
activating Nrf2. Int Immunopharmacol. 44:168–173. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shen ML, Wang CH, Lin CH, Zhou N, Kao ST
and Wu DC: Luteolin Attenuates Airway Mucus Overproduction via
Inhibition of the GABAergic System. Sci Rep. 6:327562016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shin NR, Ryu HW, Ko JW, Park SH, Yuk HJ,
Kim HJ, Kim JC, Jeong SH and Shin IS: Artemisia argyi attenuates
airway inflammation in ovalbumin-induced asthmatic animals. J
Ethnopharmacol. 209:108–115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Osorio-Guarín JA, Enciso-Rodríguez FE,
González C, Fernández-Pozo N, Mueller LA and Barrero LS:
Association analysis for disease resistance to Fusarium oxysporum
in cape gooseberry (Physalis peruviana L). BMC Genomics.
17:2482016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu SJ, Tsai JY, Chang SP, Lin DL, Wang SS,
Huang SN and Ng LT: Supercritical carbon dioxide extract exhibits
enhanced antioxidant and anti-inflammatory activities of Physalis
peruviana. J Ethnopharmacol. 108:407–413. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Franco LA, Matiz GE, Calle J, Pinzon R and
Ospina LF: Antiinflammatory activity of extracts and fractions
obtained from Physalis peruviana L. calyces. Biomedica. 27:110–115.
2007.in Spanish. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Castro J, Ocampo Y and Franco L: Cape
Gooseberry [Physalis peruviana L.] Calyces Ameliorate TNBS
Acid-induced Colitis in Rats. J Crohn's Colitis. 9:1004–1015. 2015.
View Article : Google Scholar
|
|
18
|
Park HA, Lee JW, Kwon OK, Lee G, Lim Y,
Kim JH, Paik JH, Choi S, Paryanto I, Yuniato P, et al: Physalis
peruviana L. inhibits airway inflammation induced by cigarette
smoke and lipopolysaccharide through inhibition of extracellular
signal-regulated kinase and induction of heme oxygenase-1. Int J
Mol Med. 40:1557–1565. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jeon CM, Shin IS, Shin NR, Hong JM, Kwon
OK, Kim HS, Oh SR, Myung PK and Ahn KS: Siegesbeckia glabrescens
attenuates allergic airway inflammation in LPS-stimulated RAW 264.7
cells and OVA induced asthma murine model. In t. Immunopharmacol.
22:414–419. 2014. View Article : Google Scholar
|
|
20
|
Lee JW, Park JW, Shin NR, Park SY, Kwon
OK, Park HA, Lim Y, Ryu HW, Yuk HJ, Kim JH, et al: Picrasma
quassiodes (D. Don) Benn. attenuates lipopolysaccharide
(LPS)-induced acute lung injury. Int J Mol Med. 38:834–844. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kupczyk M and Kuna P: Benralizumab: An
anti-IL-5 receptor α monoclonal antibody in the treatment of
asthma. Immunotherapy. 10:349–359. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Deo SS, Mistry KJ, Kakade AM and Niphadkar
PV: Role played by Th2 type cytokines in IgE mediated allergy and
asthma. Lung India. 27:66–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu W, Hu M, Zhang Q, Yu J and Su W:
Effects of anthraquinones from Cassia occidentalis L. on
ovalbumin-induced airways inflammation in a mouse model of allergic
asthma. J Ethnopharmacol. 221:1–9. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma Y, Tang T, Sheng L, Wang Z, Tao H,
Zhang Q, Zhang Y and Qi Z: Aloin suppresses lipopolysaccharide
induced inflammation by inhibiting JAK1 STAT1/3 activation and ROS
production in RAW264.7 cells. Int J Mol Med. 42:1925–1934.
2018.PubMed/NCBI
|
|
25
|
Lee JW, Seo KH, Ryu HW, Yuk HJ, Park HA,
Lim Y, Ahn KS and Oh SR: Anti-inflammatory effect of stem bark of
Paulownia tomentosa Steud. in lipopolysaccharide (LPS)-stimulated
RAW264.7 macrophages and LPS-induced murine model of acute lung
injury. J Ethnopharmacol. 210:23–30. 2018. View Article : Google Scholar
|
|
26
|
Gao Y, Zhaoyu L, Xiangming F, Chunyi L,
Jiayu P, Lu S, Jitao C, Liangcai C and Jifang L: Abietic acid
attenuates allergic airway inflammation in a mouse allergic asthma
model. Int Immunopharmacol. 38:261–266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu J, Zhu YT, Wang GZ, Han D, Wu YY, Zhang
DX, Liu Y, Zhang YH, Xie XM, Li SJ, et al: The PPARγ agonist,
rosiglitazone, attenuates airway inflammation and remodeling via
heme oxygenase-1 in murine model of asthma. Acta Pharmacol Sin.
36:171–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang C, Choi YH, Xian Z, Zheng M, Piao H
and Yan G: Aloperine suppresses allergic airway inflammation
through NF-κB, MAPK, and Nrf2/HO-1 signaling pathways in mice. Int
Immunopharmacol. 65:571–579. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim YH, Choi YJ, Kang MK, Park SH, Antika
LD, Lee EJ, Kim DY and Kang YH: Astragalin inhibits allergic
inflammation and airway thickening in ovalbumin-challenged mice. J
Agric Food Chem. 65:836–845. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang P, Shi HY, Wu XM, Zhang YH, Zhong YJ,
Deng WJ, Zhang YP and Xie M: Targeted inhibition of GATA-6
attenuates airway inflammation and remodeling by regulating
caveolin-1 through TLR2/MyD88/NF-κB in murine model of asthma. Mol
Immunol. 75:144–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wardlaw AJ, Brightling C, Green R,
Woltmann G and Pavord I: Eosinophils in asthma and other allergic
diseases. Br Med Bull. 56:985–1003. 2000. View Article : Google Scholar
|
|
32
|
Venturini CL, Macho A, Arunachalam K, de
Almeida DAT, Rosa SIG, Pavan E, Balogun SO, Damazo AS and Martins
DTO: Vitexin inhibits inflammation in murine ovalbumin-induced
allergic asthma. Biomed Pharmacother. 97:143–151. 2018. View Article : Google Scholar
|
|
33
|
Byrne AJ, Jones CP, Gowers K, Rankin SM
and Lloyd CM: Lung macrophages contribute to house dust mite driven
airway remodeling via HIF-1α. PLoS One. 8:e692462013. View Article : Google Scholar
|
|
34
|
Poston RN, Chanez P, Lacoste JY,
Litchfield T, Lee TH and Bousquet J: Immunohistochemical
characterization of the cellular infiltration in asthmatic bronchi.
Am Rev Respir Dis. 145:918–921. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li R, Wang J, Zhu F, Li R, Liu B, Xu W, He
G, Cao H, Wang Y and Yang J: HMGB1 regulates T helper 2 and T
helper17 cell differentiation both directly and indirectly in
asthmatic mice. Mol Immunol. 97:45–55. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hao W, Wang J, Zhang Y, Wang Y, Sun L and
Han W: Leptin positively regulates MUC5AC production and secretion
induced by interleukin-13 in human bronchial epithelial cells.
Biochem Biophys Res Commun. 493:979–984. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Licari A, Castagnoli R, Panfili E,
Marseglia A, Brambilla I and Marseglia GL: An update on anti-IgE
therapy in pediatric respiratory diseases. Curr Respir Med Rev.
13:22–29. 2017. View Article : Google Scholar :
|
|
38
|
Bax HJ, Keeble AH and Gould HJ:
Cytokinergic IgE action in mast cell activation. Front Immunol.
3:2292012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song YD, Li XZ, Wu YX, Shen Y, Liu FF, Gao
PP, Sun L and Qian F: Emodin alleviates alternatively activated
macrophage and asthmatic airway inflammation in a murine asthma
model. Acta Pharmacol Sin. 39:1317–1325. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schneider D, Hong JY, Bowman ER, Chung Y,
Nagarkar DR, McHenry CL, Goldsmith AM, Bentley JK, Lewis TC and
Hershenson MB: Macrophage/epithelial cell CCL2 contributes to
rhinovirus-induced hyperresponsiveness and inflammation in a mouse
model of allergic airways disease. Am J Physiol Lung Cell Mol
Physiol. 304:L162–L169. 2013. View Article : Google Scholar :
|
|
41
|
Nguyen TH, Maltby S, Simpson JL, Eyers F,
Baines KJ, Gibson PG, Foster PS and Yang M: TNF-α and macrophages
are critical for respiratory syncytial virus-induced exacerbations
in a mouse model of allergic airways disease. J Immunol.
196:3547–3558. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lillehoj ER and Kim KC: Airway mucus: Its
components and function. Arch Pharm Res. 25:770–780. 2002.
View Article : Google Scholar
|
|
43
|
Evans CM, Kim K, Tuvim MJ and Dickey BF:
Mucus hypersecretion in asthma: Causes and effects. Curr Opin Pulm
Med. 15:4–11. 2009. View Article : Google Scholar :
|
|
44
|
Xiong YY, Wang JS, Wu FH, Li J and Kong
LY: The effects of (±)-Praeruptorin A on airway inflammation,
remodeling and transforming growth factor-β1/Smad signaling pathway
in a murine model of allergic asthma. Int Immunopharmacol.
14:392–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang W, Li ML, Xia MY and Shao JY:
Fisetin-treatment alleviates airway inflammation through inhbition
of MyD88/NF-κB signaling pathway. Int J Mol Med. 42:208–218.
2018.PubMed/NCBI
|
|
46
|
Wei DZ, Guo XY, Lin LN, Lin MX, Gong YQ,
Ying BY and Huang MY: Effects of Angelicin on Ovalbumin
(OVA)-Induced Airway Inflammation in a Mouse Model of Asthma.
Inflammation. 39:1876–1882. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yoon SC, Je IG, Cui X, Park HR, Khang D,
Park JS, Kim SH and Shin TY: Anti-allergic and anti-inflammatory
effects of aqueous extract of Pogostemon cablin. Int J Mol Med.
37:217–224. 2016. View Article : Google Scholar
|
|
48
|
Wijerathne CUB, Seo CS, Song JW, Park HS,
Moon OS, Won YS, Kwon HJ and Son HY: Isoimperatorin attenuates
airway inflammation and mucus hypersecretion in an
ovalbumin-induced murine model of asthma. Int Immunopharmacol.
49:67–76. 2017. View Article : Google Scholar : PubMed/NCBI
|