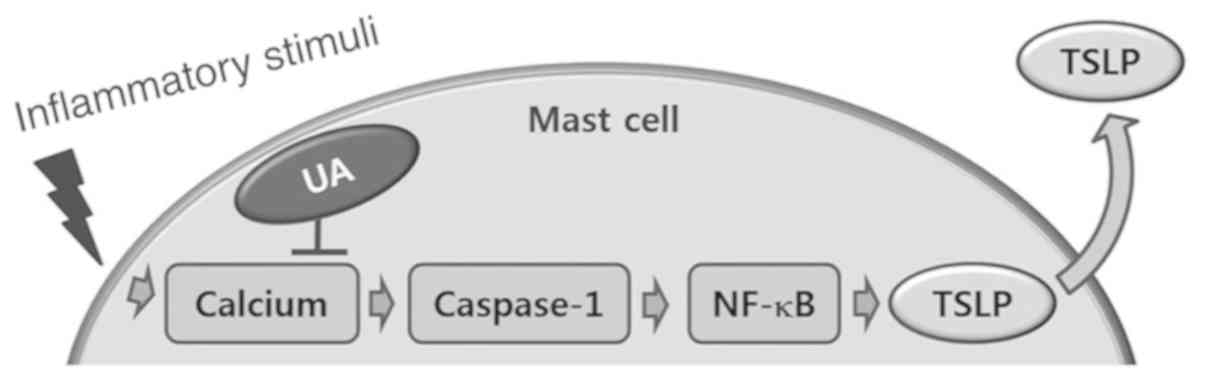
Introduction
Atopic dermatitis (AD) is a recurrent and chronic
inflammatory skin disorder that affects children as well as adults
(1,2). The prevalence of AD is ~1-3% in
adults and 25% in children of industrialized countries (3), and it has increased in the urban
areas of such countries over the past decade (4). As a result of the increasing
prevalence of AD, the burden of AD-associated medical costs has
increased in industrialized countries (5). In addition, the recurrent eczema
lesions, itching, lack of sleep and restricted diet may compromise
the quality of life of AD patients (5).
Thymic stromal lymphopoietin (TSLP) is considered as
a pivotal factor in the pathogenesis of allergic diseases, such as
AD and asthma. In patients with AD, TSLP gene expression increased
by epicutaneous house dust mite injections (6). The protein and mRNA expression
levels of TSLP in skin lesions of AD patients are higher compared
with those in healthy controls (7). In addition to epithelial cells and
keratinocytes, mast cells also play an important role in atopic
diseases (8). The increases in
the population and activation of mast cells in AD models reported
by several researchers (9-11)
indicate the significance of mast cells in AD. HMC-1 is a human
mast cell line (12). The effects
of UA on the HMC-1 cell line were examined, as similar levels of
TSLP are produced by HMC-1 and bone marrow-derived mast cells
(13).
In general, protease caspases play critical roles in
apoptosis, whereas caspase-1 is implicated in inflammatory
responses (14-16). Deficiency of caspase-1 ameliorates
dextran sulfate sodium-induced intestinal inflammation (17). In addition, TSLP expression and
production were found to be mediated by caspase-1 and nuclear
factor (NF)-κB signaling in HMC-1 cells in a previous study
(18). Additionally, caspase-1
inhibitor treatment may decrease NF-κB activation, suggesting that
caspase-1 acts as an upstream regulator of NF-κB (18).
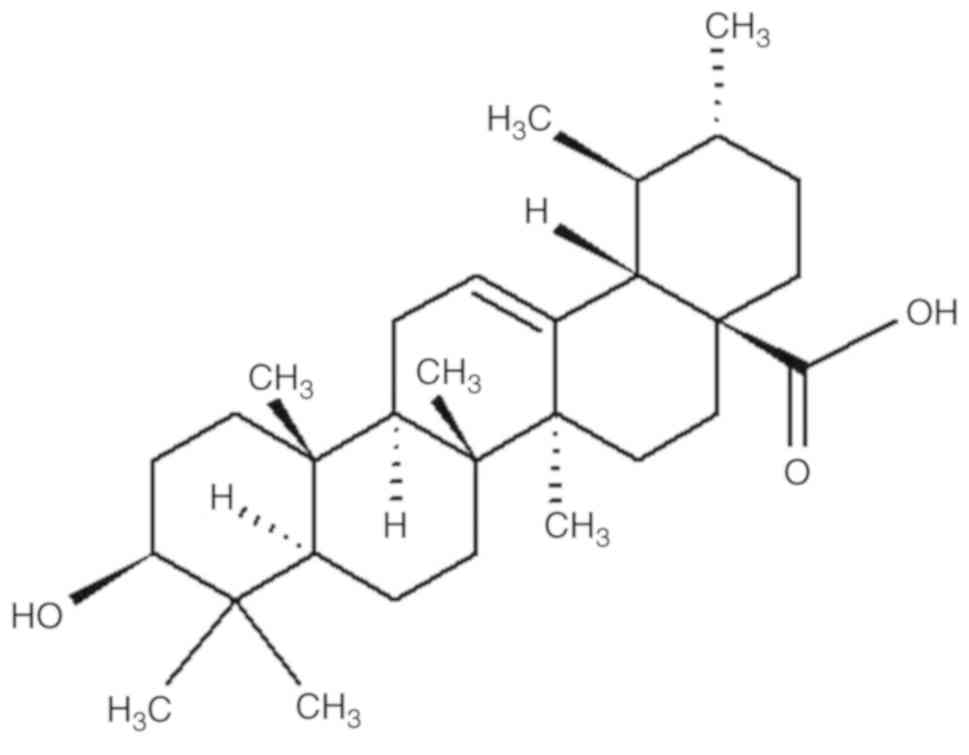
Ursolic acid (UA; Fig.
1), a pentacyclic triterpenoid found in holy basil and apple
peels (19), has various
pharmacological properties, such as antioxidant, anti-inflammatory
and anticancer (20,21). Recently, Gan et al
(22) reported that UA
ameliorates CCl4-induced liver fibrosis. However, the
regulatory effect of UA on TSLP production by mast cells has not
been fully elucidated. The aim of the present study was to
investigate the effects of UA on the HMC-1 human mast cell line and
determine whether UA can regulate TSLP production in mast
cells.
Materials and methods
Materials
UA, phorbol myristate acetate (PMA), calcium
ionophore A23187, avidin-peroxidase and dimethyl sulfoxide were
obtained from Sigma-Aldrich; Merck KGaA. TMB substrate and tumor
necrosis factor (TNF)-α antibodies were purchased from Pharmingen.
Power SYBR®-Green PCR master mix was purchased from Applied
Biosystems; Thermo Fisher Scientific, Inc. TSLP antibodies and
caspase-1 assay kit were obtained from R&D Systems, Inc.
Finally, IKKβ, PARP, GAPDH, IκBα and NF-κB p65 antibodies were
obtained from Santa Cruz Biotechnology, Inc.
Cell culture
HMC-1 cells were cultured in IMDM with
heat-inactivated fetal bovine serum (10%), streptomycin (100
μg/ml) and penicillin (100 U/ml) at 37°C with 5%
CO2.
MTT assay
MTT assay was performed to measure cytotoxicity, as
described previously (23). HMC-1
cells (3×105) were incubated with UA (0.002-0.2
μg/ml) in 24-well plates, which were subsequently incubated
with MTT solution (5 mg/ml) for 4 h. To dissolve the MTT formazan,
1 ml of dimethyl sulfoxide was added and 200 μl of
supernatant were removed and transferred to a 96-well microplate.
Finally, each well was read at 540 nm.
Cytokine assay
HMC-1 cells (3×105) were pretreated with
UA (0.002-0.2 μg/ml) for 1 h prior to stimulation with 0.05
μM PMA plus 1 μM calcium ionophore (PMACI), and then
incubated for 7 h. TSLP and TNF-α levels were assessed in the
culture supernatants using ELISA, as described previously (24).
Quantitative polymerase chain reaction
(qPCR) analysis
HMC-1 cells (1×106) were pretreated with
UA (0.002-0.2 μg/ml) for 1 h prior to PMACI stimulation, and
then incubated for 5 h. qPCR was carried out using the Power
SYBR-Green PCR master mix. mRNA detection was performed with the
ABI StepOne real-time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) as described previously (25). PCR analysis was conducted using
the following primers: TSLP, forward 5′-CCCAGGCTATTCGGAAACTCAG-3′
and reverse 5′-CGCCACAATCCTTGTAATTGTG-3′; GAPDH, forward
5′-TCGACAGTCAGCCGCATCTTCTTT-3′ and reverse
5′-ACCAAATCCGTTGACTCCGACCTT-3′.
Caspase-1 assay
HMC-1 cells (5×106) were pretreated with
UA (0.002-0.2 μg/ml) for 1 h prior to PMACI stimulation, and
then incubated for 1 h. Caspase-1 activation was evaluated using a
caspase-1 assay kit, as described previously (26).
Nuclear and cytoplasmic extracts
HMC-1 cells (5×106) were pretreated with
UA (0.002-0.2 μg/ml) for 1 h prior to PMACI stimulation, and
then incubated for 2 h. Isolation of nuclear and cytoplasmic
proteins was carried out as described previously (27). In brief, the cells were washed in
ice-cold phosphate-buffered saline (PBS) and centrifuged at 15,000
× g for 1 min. The cells were resuspended in 40 μl of a cold
hypotonic buffer (10 mM HEPES/KOH, 2 mM MgCl2, 0.1 mM
EDTA, 10 mM KCl, 1 mM DTT, and 0.5 mM PMSF, pH 7.9). Next, the
cells were swollen on ice for 15 min, lysed gently with 2.5
μl 10% Nonidet P-40 and centrifuged at 15,000 × g for 3 min
at 4°C. The supernatant was then collected and used as the
cytoplasmic extract. The pellets of nuclei were gently resuspended
in 40 μl cold saline buffer (50 mM HEPES/KOH, 50 mM KCl, 300
mM NaCl, 0.1 mM EDTA, 10% glycerol, 1 mM DTT and 0.5 mM PMSF, pH
7.9) and placed on ice for 20 min. After centrifugation at 15,000 ×
g for 15 min at 4°C, the aliquots of supernatant containing the
nuclear proteins were frozen in liquid nitrogen and stored at −70°C
until analysis. Finally, the bicinchoninic acid protein assay
(Sigma-Aldrich; Merck KGaA) was used to measure the protein
concentrations.
Western blot analysis
HMC-1 cells (5×106) were pretreated with
UA (0.002-0.2 μg/ml) for 1 h prior to PMACI stimulation.
Proteins of obtained lysates were separated and transferred to
nitrocellulose paper, as described previously (28). In brief, the cell lysates were
prepared in a sample buffer containing sodium dodecyl sulfate
(SDS). The samples were heated at 95°C for 5 min and briefly cooled
on ice. Following centrifugation at 15,000 × g for 5 min, the
proteins in the cell lysates were separated by 10%
SDS-polyacrylamide gel electrophoresis and transferred to
nitrocellulose paper. The membrane was blocked with 5% skimmed milk
in PBS-Tween-20 for 1 h at room temperature and then incubated with
primary antibodies (1:500 dilution for all primary antibodies;
NF-κB; cat. no. sc-8008; IκBα; cat. no. sc-847; IKKβ; cat. no.
sc-7607; PARP; sc-8007; GAPDH; cat. no. sc-32233; all purchased
from Santa Cruz Biotechnology, Inc.) overnight at 4°C and secondary
(mouse anti-rabbit IgG-HRP; 1:5,000; cat. no. sc-2357; goat
anti-mouse IgG-HRP, 1:5,000; cat. no. sc-2005; all purchased from
Santa Cruz Biotechnology, Inc.) antibodies for 1 h at room
temperature. Finally, the protein bands were visualized by an
enhanced chemiluminesence solution (Amersham; GE Healthcare)
following the manufacturer's instructions. All protein expression
levels were quantitated using ImageJ software (National Institutes
of Health).
Fluorescent measurements of the
intracellular calcium level
HMC-1 cells (1×105) were pretreated with
Fura-2/AM for 30 min. After washing twice with medium containing
the extracellular calcium chelator EGTA (0.5 mM), the cell
suspension (1×105) was seeded into a 96-well plate and
pretreated with UA (0.002-0.2 μg/ml) for 20 min. Next, the
cells were stimulated with PMACI for 5 min. Plate fluorescence was
measured at 440 nm (excitation, 360 nm) in a spectrofluorometer
(29).
Statistical analysis
IBM SPSS 23.0 (IBM Corp.) was used to statistically
analyze the results. Statistical analyses included performing
independent t-tests and analysis of variance with Tukey's post hoc
test. The differences were considered statistically significant at
P<0.05, and the results are presented as mean ± standard error
of the mean.
Results
Effect of UA on the production of
TSLP
To evaluate the effect of UA on the production of
TSLP in HMC-1 cells, HMC-1 cells were exposed to PMACI for 7 h. The
levels of TSLP were evaluated with ELISA. Exposure to PMACI
elevated the production of TSLP in HMC-1 cells (Fig. 2A); however, the elevated TSLP
production was significantly lowered by UA (0.02 and 0.2
μg/ml; Fig. 2A). The TSLP
production levels at concentrations of 0.002, 0.02 and 0.2
μg/ml were 0.127±0.006, 0.111±0.003 and 0.101±0.004,
respectively. The levels of TSLP production in the blank and
control groups were 0.079±0.002 and 0.135±0.008, respectively.
Treatment with UA (0.2 μg/ml) reduced the TSLP production up
to 61.442±6.947%. However, UA alone did not notably change the
level of TSLP production from that in the blank (PBS-treated cells)
group (data not shown). When HMC-1 cells were treated with UA at
concentrations of 0.002-0.2 μg/ml, cell viability did not
change (Fig. 2C). Higher
concentrations of UA (2 and 20 μg/ml) did not achieve
further TSLP inhibition (Fig.
2A). In addition, prolonged UA pretreatment did not achieve
further TSLP inhibition (Fig.
2D). A 1-h pretreatment with UA inhibited TSLP production by up
to ~60%, whereas a 24-h UA pretreatment produced a ~40% TSLP
inhibition (Fig. 2D). This may be
due to spontaneously released TSLP. Furthermore, when UA was added
1 h after PMACI stimulation, it did not significantly inhibit TSLP
production (Fig. 2E).
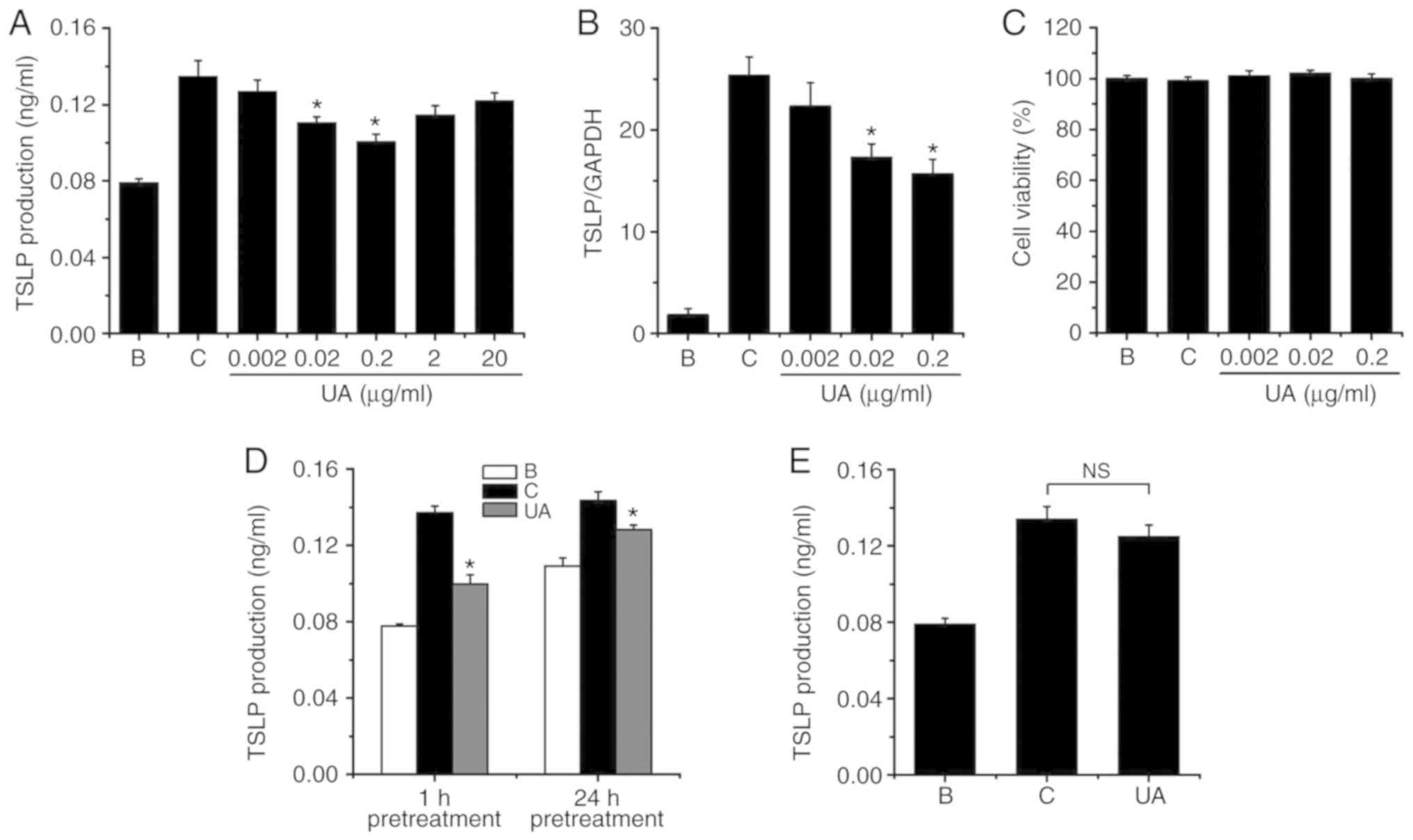 | Figure 2Effects of UA on the regulation of
TSLP production and mRNA expression. (A) HMC-1 cells
(3×105) were treated with various concentrations of UA
(0.002-20 μg/ml) for 1 h, after which time the HMC-1 cells
were stimulated with PMACI for 7 h. The TSLP levels were determined
by ELISA. (B) HMC-1 cells (1×106) were exposed to PMACI
for 5 h. The level of TSLP mRNA expression was evaluated with qPCR.
(C) Various concentrations of UA (0.002-0.2 μg/ml) were
applied to HMC-1 cells (3×105) for 1 h, and the HMC-1
cells were then stimulated with PMACI for 7 h. Cytotoxicity was
analyzed by the MTT assay. (D) UA (0.2 μg/ml) pretreatment
was applied to HMC-1 cells (3×105) for 24 or 1 h, after
which time the HMC-1 cells were stimulated with PMACI for 7 h. The
TSLP levels were determined by ELISA. (E) HMC-1 cells
(3×105) were stimulated with PMACI for 7 h and UA (0.2
μg/ml) was added to the HMC-1 cells 1 h after PMACI
stimulation. The TSLP levels were determined by ELISA. B,
PBS-treated cells; C, PBS + PMACI-treated cells. Data are presented
as mean ± standard error of the mean from three independent
experiments. *P<0.05 vs. the PBS + PMACI-treated
cells. NS, not significant. UA, ursolic acid; TSLP, thymic stromal
lymphopoietin; PMACI, phorbol myristate acetate and calcium
ionophore; HMC, human mast cell; qPCR, quantitative polymerase
chain reaction; PBS, phosphate-buffered saline. |
Effect of UA on mRNA expression of
TSLP
To determine the regulatory effect of UA on the TSLP
mRNA expression, a variety of concentrations (0.002-0.2
μg/ml) of UA were added as pretreatment for 1 h prior to the
exposure of HMC-1 cells to PMACI. Exposure to PMACI elevated TSLP
mRNA expression, whereas the elevated TSLP mRNA expression was
lowered by UA treatment (Fig.
2B). The TSLP mRNA expression values at concentrations of
0.002, 0.02 and 0.2 μg/ml were 22.667±2.333, 17.333±1.333
and 15.667±1.453, respectively. The relative expression levels of
TSLP mRNA in the blank and control groups were 1.800±0.640 and
25.333±1.856, respectively.
Effects of UA on activation of NF-κB and
degradation of IκBα
To investigate whether the regulatory effect of UA
is mediated by NF-κB/IκBα signaling, the activation of NF-κB p65
and degradation of IκBα were assessed by western blot analysis.
Exposure to PMACI elevated NF-κB activation in the nuclear extract,
whereas the elevated NF-κB activation was lowered by UA treatment
(Fig. 3). The relative intensity
values for NF-κB activation in the blank, control and UA groups
(0.002, 0.02 and 0.2 μg/ml) were 0.408±0.025, 0.629±0.039,
0.533±0.034, 0.455±0.028 and 0.424±0.053, respectively. However, UA
treatment did not produce a significant change in cytoplasmic NF-κB
protein levels (Fig. 3).
Proteolytic degradation of IκBα results in activation of NF-κB
(30,31); thus, we investigated whether the
regulatory effect of UA is due to IκBα degradation. Exposure to
PMACI elevated IκBα degradation in the cytoplasmic extract;
however, the elevated IκBα degradation was lowered by UA treatment
(Fig. 3). The relative intensity
values of IκBα in the blank, control and UA groups were
0.438±0.019, 0.288±0.010, 0.304±0.016, 0.363±0.006 and 0.384±0.012,
respectively. Phosphorylation and degradation of IκBα is due to IκB
kinase (IKK) complex activation, and the IKK complex consists of
three core subunits (IKKα, IKKβ and IKKγ), among which IKKβ is
predominant (32); thus, we
investigated whether IκBα degradation by UA is due to IKKβ.
Exposure to PMACI elevated the IKKβ protein levels; however, the
elevated IKKβ protein levels were reduced by UA treatment (Fig. 3). The relative intensity values of
IKKβ in the blank, control and UA groups were 0.223±0.019,
0.298±0.004, 0.287±0.003, 0.253±0.006 and 0.251±0.007,
respectively.
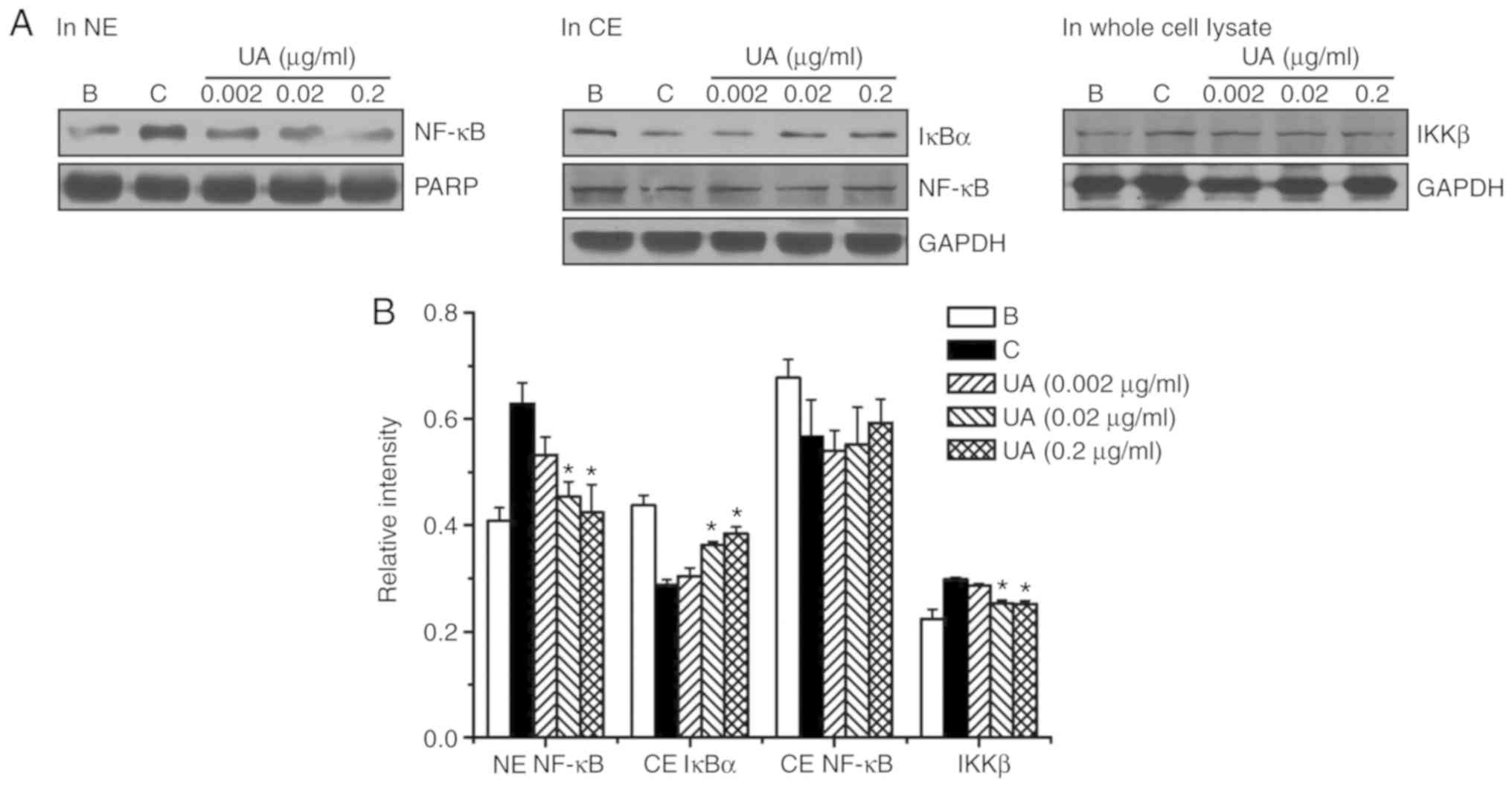 | Figure 3Effects of UA on the regulation of
NF-κB p65 activation, IκBα degradation, and IKKβ activation. (A) UA
(0.002-0.2 μg/ml) was added to HMC-1 cells
(5×106) for 1 h, after which time the HMC-1 cells were
stimulated with PMACI for 2 h. (B) The protein expression levels
were quantitated by densitometry. B, PBS-treated cells; C, PBS +
PMACI-treated cells. Data are presented as mean ± standard error of
the mean from three independent experiments. *P<0.05
vs. PBS + PMACI-treated cells. UA, ursolic acid; NF-κB, nuclear
factor-κB; HMC, human mast cell; PMACI, phorbol myristate acetate
and calcium ionophore; NE, nuclear extract; CE, cytoplasmic
extract; PARP, poly(ADP-ribose) polymerase. |
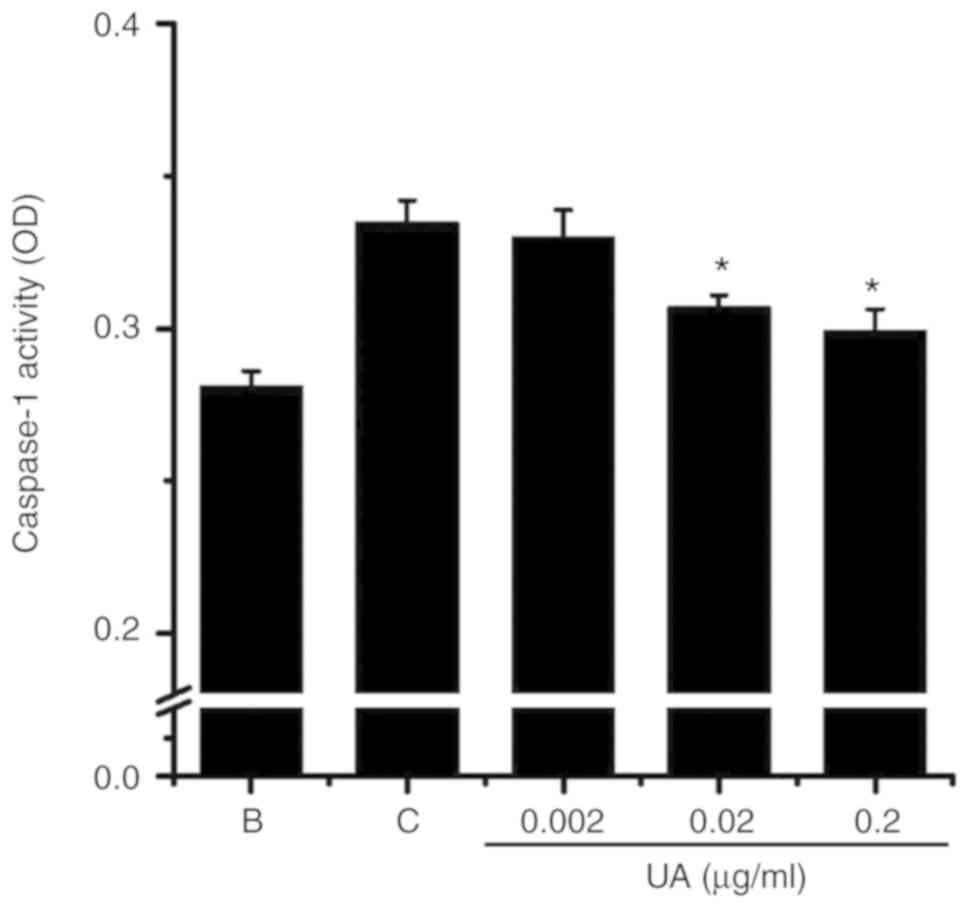
Effect of UA on the activation of
caspase-1
The level of caspase-1 activation was evaluated with
a caspase-1 assay kit to examine whether the effect of UA was
mediated through caspase-1 activation. Exposure to PMACI increased
the levels of caspase-1 activation, whereas the elevated caspase-1
activation was lowered by UA treatment (Fig. 4). The levels of caspase-1
activation in the blank, control and UA groups were 0.281±0.005,
0.335±0.007, 0.330±0.009, 0.307±0.004 and 0.299±0.007,
respectively.
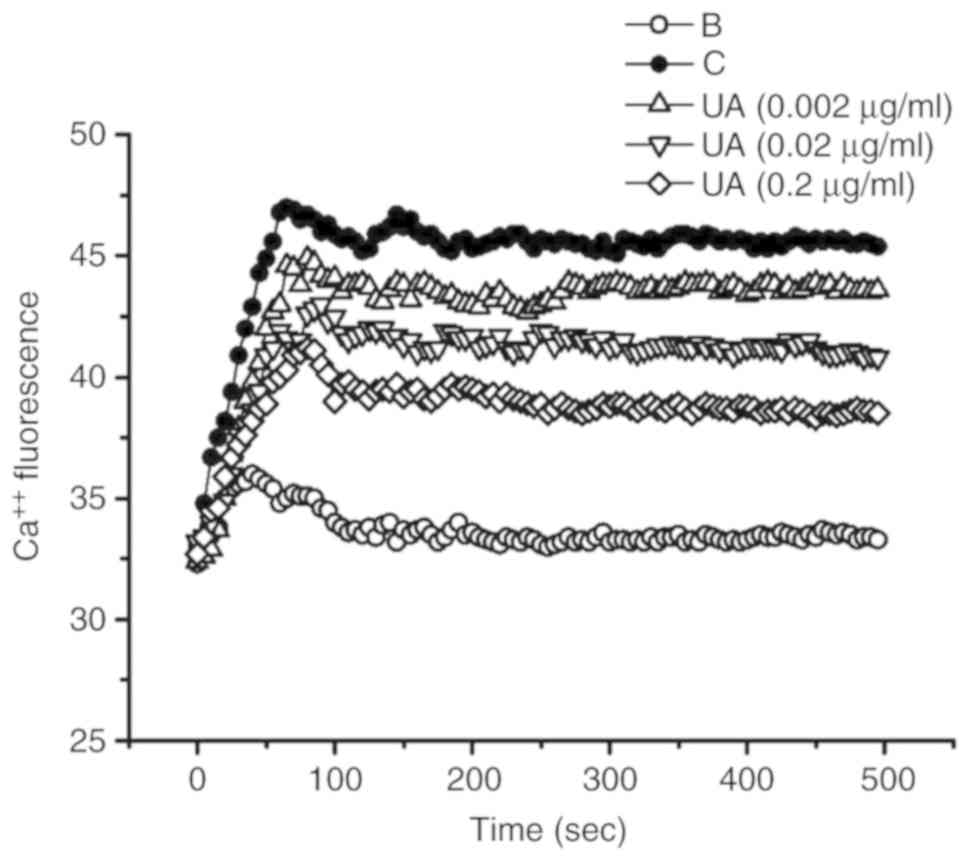
Effect of UA on calcium level
An increase in the intracellular calcium levels has
been reported to enhance caspase-1 activation (33). Thus, the regulatory effect of UA
on intracellular calcium levels was examined in HMC-1 cells.
Exposure to PMACI increased intracellular calcium levels; however,
this increase was prevented by UA treatment (Fig. 5).
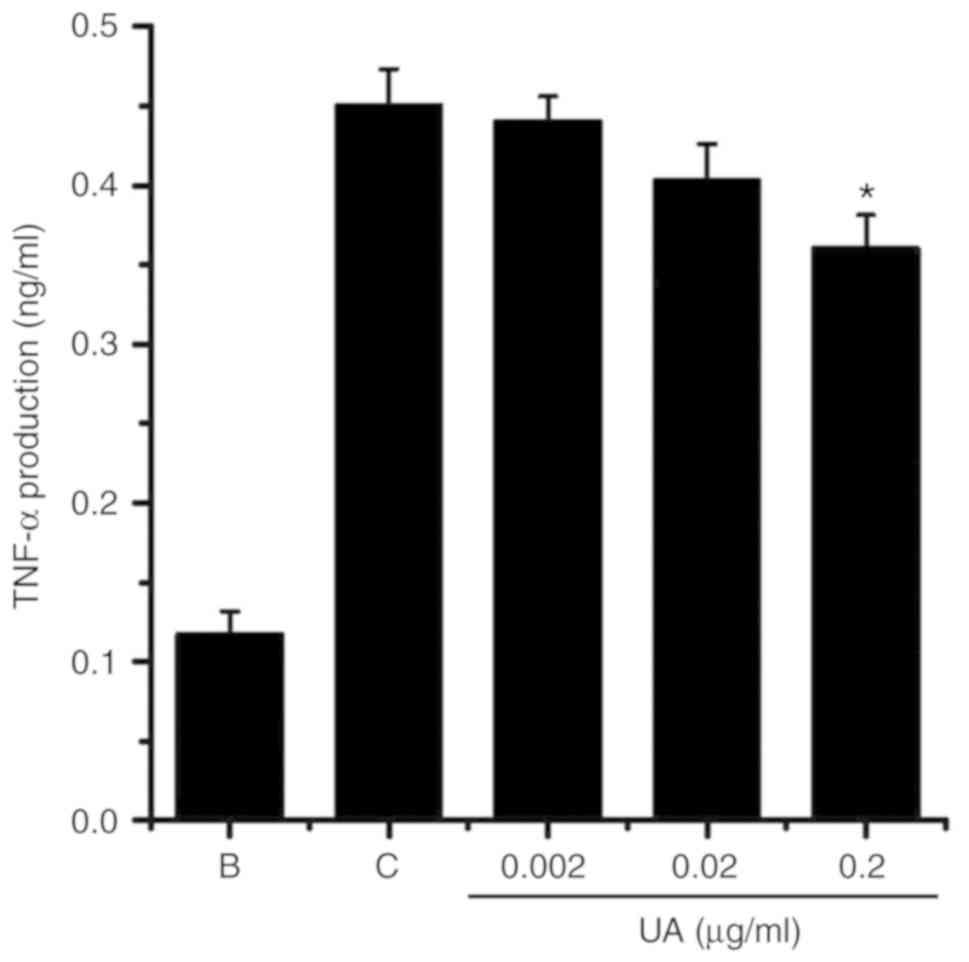
Effect of UA on pro-inflammatory cytokine
levels in HMC-1 cells
The pro-inflammatory cytokine tumor necrosis factor
(TNF)-α is overexpressed in AD (34). To substantiate the presence of UA
effects in AD, the levels of TNF-α were measured in HMC-1 cells.
Exposure to PMACI increased TNF-α production in HMC-1 cells
(Fig. 6); however, this increase
in TNF-α production was markedly reduced by treatment with 0.2
μg/ml UA (Fig. 6).
Discussion
In the present study, UA was shown to suppress the
production and mRNA expression of TSLP in HMC-1 cells. In addition,
UA reduced NF-κB activation, IκBα degradation and caspase-1
activity in HMC-1 cells. Finally, it was demonstrated that UA
downregulated intracellular calcium levels in HMC-1 cells.
When mast cells are activated, there are increases
in the activation of protein kinase C (PKC) and intracellular
calcium levels (35). To
replicate this condition in the present study, PMA was used to
activate PKC, and calcium ionophore to increase the levels of
intracellular calcium. Exposure to PMACI was reported to increase
the production and mRNA expression of TSLP in HMC-1 cells (18), and high levels of TSLP have been
detected in the skin lesions of AD patients (36). Moreover, it has been suggested
that TSLP enhances skin inflammatory responses in a murine AD model
(37), whereas dexamethasone, an
anti-inflammatory drug, inhibits expression of TSLP in a murine
model of AD (38). The results of
this study revealed that the production and mRNA expression of TSLP
were reduced by UA treatment in HMC-1 cells (Fig. 2). Therefore, UA appears to be
helpful in the treatment of atopic and inflammatory disorders.
Lee and Ziegler (39) suggested that TSLP expression is
mediated by NF-κB. Our previous report clarified that TSLP
production and mRNA expression were regulated via NF-κB signaling
in mast cells (18). The present
study demonstrated that NF-κB activation and IκBα degradation were
lowered by UA treatment (Fig. 3).
Furthermore, Shen et al (40) reported that NF-κB is a critical
transcription factor for the production of TSLP. Thus, it may be
hypothesized that UA reduces TSLP levels via blockade of NF-κB
signaling in HMC-1 cells.
Pro-inflammatory stimuli generally activate
caspase-1 (41). Several reports
have demonstrated that caspase-1 activation results from
pro-inflammatory stimulation, such as exposure to PMACI (42,43). In the present study, caspase-1
activation was found to be lowered by UA treatment (Fig. 4). Thus, it may be hypothesized
that UA suppresses the production and mRNA expression of TSLP by
blocking caspase-1 activation in HMC-1 cells.
The endoplasmic reticulum (ER) is an intracellular
calcium store in mast cells (44), and mitochondria are involved in
the modulation of intracellular calcium levels (45). Tang et al (45) reported that UA inhibits
mitochondrial calcium release. In the present study, UA treatment
prevented an increase in intracellular calcium, suggesting that UA
contributes to the prevention of ER and mitochondrial calcium
release (Fig. 5). An increase in
intracellular calcium levels promotes the activation of caspase-1
(33), whereas caspase-1
activation in HMC-1 cells may be reduced by treatment with the
calcium chelator BAPTA-AM (46).
The results of the present study revealed that an increase in
intracellular calcium may be prevented by UA treatment (Fig. 5). Thus, UA appears to decrease
TSLP levels via blockade of calcium/caspase-1/NF-κB signaling in
HMC-1 cells (Fig. 7). When UA was
added following PMACI stimulation, it did not produce a significant
change in TSLP inhibition (Fig.
2E). Thus, UA may exert a preventive rather than a therapeutic
effect on AD. Finally, UA significantly attenuated the effects of
PMACI; however, the levels of TNF-α, calcium fluorescence,
TSLP/GAPDH and NF-κB/GAPDH did not return to those observed in
non-PMACI-treated cells. High-fat diet exacerbated AD-like skin
lesions in NC/Nga mice and increased TSLP levels in skin lesions,
whereas pro-inflammatory cytokines, such as interleukin (IL)-4,
IL-13, interferon-γ and TNF-α did not exhibit significant changes
(47). To confirm the role of
TSLP in AD-like skin lesions, when Moon et al (47) prepared TSLP knockout NC/Nga mice,
TSLP deficiency markedly decreased AD-like skin lesions. Although
the present conditions differ from those in the previous report
(47), TSLP inhibition by UA may
contribute significantly to the amelioration of the symptoms of
allergic and atopic disorders.
In conclusion, the present study demonstrated that
UA inhibits the production and mRNA expression of TSLP in HMC-1
cells. Moreover, UA decreased the activation of NF-κB, degradation
of IκBα and activation of caspase-1. Furthermore, UA downregulated
the levels of intracellular calcium. Therefore, these results
suggest that UA, through its ability to downregulate TSLP, may be a
valuable agent for the treatment and/or prevention of atopic and
inflammatory diseases.
Funding
This study was supported by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education, Science and Technology
(NRF-2017R1D1A1B03035976).
Availability of data and materials
The data generated and analyzed during the present
study are available from the corresponding authors on reasonable
request.
Authors' contributions
PDM and NRH wrote the manuscript and conducted all
experiments. JSL analyzed the data. HMK and HJJ designed the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Guttman-Yassky E, Hanifin JM, Boguniewicz
M, Wollenberg A, Bissonnette R, Purohit V, Kilty I, Tallman AM and
Zielinski MA: The role of phosphodiesterase 4 in the
pathophysiology of atopic dermatitis and the perspective for its
inhibition. Exp Dermatol. 28:3–10. 2019.
|
|
2
|
Yu JH, Jin M, Choi YA, Jeong NH, Park JS,
Shin TY and Kim SH: Suppressive effect of an aqueous extract of
Diospyros kaki calyx on dust mite
extract/2,4-dinitrochlorobenzene-induced atopic dermatitis-like
skin lesions. Int J Mol Med. 40:505–511. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Löwa A, Jevtić M, Gorreja F and Hedtrich
S: Alternatives to animal testing in basic and preclinical research
of atopic dermatitis. Exp Dermatol. 27:476–483. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bieber T: Atopic dermatitis. Ann Dermatol.
22:125–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Plötz SG and Ring J: What's new in atopic
eczema? Expert Opin Emerg Drugs. 15:249–267. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Landheer J, Giovannone B, Mattson JD,
Tjabringa S, Bruijnzeel-Koomen CA, McClanahan T, de Waal Malefyt R,
Knol E and Hijnen D: Epicutaneous application of house dust mite
induces thymic stromal lymphopoietin in nonlesional skin of
patients with atopic dermatitis. J Allergy Clin Immunol.
132:1252–1254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo Y, Zhou B, Zhao M, Tang J and Lu Q:
Promoter demethylation contributes to TSLP overexpression in skin
lesions of patients with atopic dermatitis. Clin Exp Dermatol.
39:48–53. 2014. View Article : Google Scholar
|
|
8
|
Zhu Y, Pan WH, Wang XR, Liu Y, Chen M, Xu
XG, Liao WQ and Hu JH: Tryptase and protease-activated receptor-2
stimulate scratching behavior in a murine model of
ovalbumin-induced atopic-like dermatitis. Int Immunopharmacol.
28:507–512. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schneider C, Döcke WD, Zollner TM and Röse
L: Chronic mouse model of TMA-induced contact hypersensitivity. J
Invest Dermatol. 129:899–907. 2009. View Article : Google Scholar
|
|
10
|
Han NR, Oh HA, Nam SY, Moon PD, Kim DW,
Kim HM and Jeong HJ: TSLP induces mast cell development and
aggravates allergic reactions through the activation of MDM2 and
STAT6. J Invest Dermatol. 134:2521–2530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han NR, Moon PD, Yoou MS, Chang TS, Kim HM
and Jeong HJ: Effect of massage therapy by VOSKIN 125+ painkiller®
on inflammatory skin lesions. Dermatol Ther. 31:e126282018.
View Article : Google Scholar
|
|
12
|
Gauchat JF, Henchoz S, Mazzei G, Aubry JP,
Brunner T, Blasey H, Life P, Talabot D, Flores-Romo L, Thompson J,
et al: Induction of human IgE synthesis in B cells by mast cells
and basophils. Nature. 365:340–343. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moon PD, Choi IH and Kim HM: Berberine
inhibits the production of thymic stromal lymphopoietin by the
blockade of caspase-1/NF-κB pathway in mast cells. Int
Immunopharmacol. 11:1954–1959. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schneider KS, Groß CJ, Dreier RF, Saller
BS, Mishra R, Gorka O, Heilig R, Meunier E, Dick MS, Ćiković T, et
al: The inflammasome drives GSDMD-independent secondary pyroptosis
and IL-1 release in the absence of caspase-1 protease activity.
Cell Rep. 21:3846–3859. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han NR, Moon PD, Kim NR, Kim HY, Jeong HJ
and Kim HM: Schisandra chinensis and its main constituent
schizandrin attenuate allergic reactions by down-regulating
caspase-1 in ovalbumin-sensitized mice. Am J Chin Med. 45:159–172.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo L, Kong Q, Dong Z, Dong W, Fu X, Su L
and Tan X: NLRC3 promotes host resistance against Pseudomonas
aeruginosa-induced keratitis by promoting the degradation of IRAK1.
Int J Mol Med. 40:898–906. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Błażejewski AJ, Thiemann S, Schenk A, Pils
MC, Gálvez EJC, Roy U, Heise U, de Zoete MR, Flavell RA and Strowig
T: Microbiota normalization reveals that canonical caspase-1
activation exacerbates chemically induced intestinal inflammation.
Cell Rep. 19:2319–2330. 2017. View Article : Google Scholar
|
|
18
|
Moon PD and Kim HM: Thymic stromal
lymphopoietin is expressed and produced by caspase-1/NF-κB pathway
in mast cells. Cytokine. 54:239–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thakur R, Sharma A, Lingaraju MC, Begum J
and Kumar D, Mathesh K, Kumar P, Singh TU and Kumar D: Ameliorative
effect of ursolic acid on renal fibrosis in adenine-induced chronic
kidney disease in rats. Biomed Pharmacother. 101:972–980. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bhat RA, Lingaraju MC, Pathak NN, Kalra J,
Kumar D, Kumar D and Tandan SK: Effect of ursolic acid in
attenuating chronic constriction injury-induced neuropathic pain in
rats. Fundam Clin Pharmacol. 30:517–528. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lewinska A, Adamczyk-Grochala J,
Kwasniewicz E, Deregowska A and Wnuk M: Ursolic acid-mediated
changes in glycolytic pathway promote cytotoxic autophagy and
apoptosis in phenotypically different breast cancer cells.
Apoptosis. 22:800–815. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gan D, Zhang W, Huang C, Chen J, He W,
Wang A, Li B and Zhu X: Ursolic acid ameliorates CCl4-induced liver
fibrosis through the NOXs/ROS pathway. J Cell Physiol.
233:6799–6813. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ben Trivedi A, Kitabatake N and Doi E:
Toxicity of dimethyl sulfoxide as a solvent in bioassay system with
HeLa cells evaluated colorimetrically with
3-(4,5-dimethylthiazol-2-yl)-2,5-di-phenyl-tetrazolium bromide.
Agric Biol Chem. 54:2961–2966. 1990.PubMed/NCBI
|
|
24
|
Han NR, Moon PD, Ryu KJ, Kim NR, Kim HM
and Jeong HJ: Inhibitory effect of naringenin via IL-13 level
regulation on thymic stromal lymphopoietin-induced inflammatory
reactions. Clin Exp Pharmacol Physiol. 45:362–369. 2018. View Article : Google Scholar
|
|
25
|
Han NR, Moon PD, Yoo MS, Ryu KJ, Kim HM
and Jeong HJ: Regulatory effects of chrysophanol, a bioactive
compound of AST201701 in a mouse model of
2,4-dinitrofluorobenzene-induced atopic dermatitis. Int
Immunopharmacol. 62:220–226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han NR, Moon PD, Kim HM and Jeong HJ:
Cordycepin ameliorates skin inflammation in a DNFB-challenged
murine model of atopic dermatitis. Immunopharmacol Immunotoxicol.
40:401–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moon PD and Kim HM: Anti-inflammatory
effect of phenethyl isothiocyanate, an active ingredient of
Raphanus sativus Linne. Food Chem. 131:1332–1339. 2012. View Article : Google Scholar
|
|
28
|
Moon PD, Han NR, Lee JS, Kim HY, Hong S,
Kim HJ, Yoo MS, Kim HM and Jeong HJ: β-eudesmol inhibits thymic
stromal lymphopoietin through blockade of caspase-1/NF-κB signal
cascade in allergic rhinitis murine model. Chem Biol Interact.
294:101–106. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han NR, Moon PD, Ryu KJ, Jang JB, Kim HM
and Jeong HJ: β-eudesmol suppresses allergic reactions via
inhibiting mast cell degranulation. Clin Exp Pharmacol Physiol.
44:257–265. 2017. View Article : Google Scholar
|
|
30
|
Wu Z, Wang Y, Meng X, Wang X, Li Z, Qian
S, Wei Y, Shu L, Ding Y, Wang P and Peng Y: Total C-21 steroidal
glycosides, isolated from the root tuber of Cynanchum auriculatum
Royle ex Wight, attenuate hydrogen peroxide-induced oxidative
injury and inflammation in L02 cells. Int J Mol Med. 42:3157–3170.
2018.PubMed/NCBI
|
|
31
|
Geng Q, Wei Q, Wang S, Qi H, Zhu Q, Liu X,
Shi X and Wen S: Physcion 8-O-β-glucopyranoside extracted from
Polygonum cuspidatum exhibits anti-proliferative and
anti-inflammatory effects on MH7A rheumatoid arthritis-derived
fibroblast-like synoviocytes through the TGF-β/MAPK pathway. Int J
Mol Med. 42:745–754. 2018.PubMed/NCBI
|
|
32
|
Perkins ND: Integrating cell-signalling
pathways with NF-kappaB and IKK function. Nat Rev Mol Cell Biol.
8:49–62. 2007. View Article : Google Scholar
|
|
33
|
Rossol M, Pierer M, Raulien N, Quandt D,
Meusch U, Rothe K, Schubert K, Schöneberg T, Schaefer M, Krügel U,
et al: Extracellular Ca2+ is a danger signal activating the NLRP3
inflammasome through G protein-coupled calcium sensing receptors.
Nat Commun. 3:13292012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Numerof RP and Asadullah K: Cytokine and
anti-cytokine therapies for psoriasis and atopic dermatitis.
BioDrugs. 20:93–103. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Moon PD, Han NR, Lee JS, Kim HM and Jeong
HJ: Effects of Linalyl acetate on thymic stromal lymphopoietin
production in mast cells. Molecules. 23:E17112018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ziegler SF: The role of thymic stromal
lymphopoietin (TSLP) in allergic disorders. Curr Opin Immunol.
22:795–799. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Oyoshi MK, Venturelli N and Geha RS:
Thymic stromal lymphopoietin and IL-33 promote skin inflammation
and vaccinia virus replication in a mouse model of atopic
dermatitis. J Allergy Clin Immunol. 138:283–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mizuno K, Morizane S, Takiguchi T and
Iwatsuki K: Dexamethasone but not tacrolimus suppresses
TNF-α-induced thymic stromal lymphopoietin expression in lesional
keratinocytes of atopic dermatitis model. J Dermatol Sci. 80:45–53.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee HC and Ziegler SF: Inducible
expression of the proallergic cytokine thymic stromal lymphopoietin
in airway epithelial cells is controlled by NFkappaB. Proc Natl
Acad Sci USA. 104:914–919. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shen D, Xie X, Zhu Z, Yu X, Liu H, Wang H,
Fan H, Wang D, Jiang G and Hong M: Screening active components from
Yu-ping-feng-san for regulating initiative key factors in allergic
sensitization. PLoS One. 9:e1072792014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Humke EW, Shriver SK, Starovasnik MA,
Fairbrother WJ and Dixit VM: ICEBERG: A novel inhibitor of
interleukin-1beta generation. Cell. 103:99–111. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Moon PD, Choi IH and Kim HM: Naringenin
suppresses the production of thymic stromal lymphopoietin through
the blockade of RIP2 and caspase-1 signal cascade in mast cells.
Eur J Pharmacol. 671:128–132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Han NR, Moon PD, Kim HM and Jeong HJ:
Tryptanthrin ameliorates atopic dermatitis through downregulation
of TSLP. Arch Biochem Biophys. 542:14–20. 2014. View Article : Google Scholar
|
|
44
|
Tasaka K: Recent advances in the research
on histamine release. Nihon Yakurigaku Zasshi. 98:197–207. 1991.In
Japanese. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tang X, Gao J, Chen J, Fang F, Wang Y, Dou
H, Xu Q and Qian Z: Inhibition by [corrected] ursolic acid of
[corrected] calcium-induced mitochondrial permeability transition
and release of two proapoptotic proteins. Biochem Biophys Res
Commun. 337:320–324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Han NR, Kim HM and Jeong HJ: Thymic
stromal lymphopoietin is regulated by the intracellular calcium.
Cytokine. 59:215–217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Moon PD, Han NR, Kim HM and Jeong HJ:
High-fat diet exacerbates dermatitis through up-regulation of TSLP.
J Invest Dermatol. 20S0022–202X. (18): 32824–0. 2018.
|