Introduction
The skin is the largest organ of the body and is the
interface between the environment and internal milieu. The
development of efficient sensory and effector capabilities to
differentially react to changes in the external environment is a
vital property of skin. Melanogenesis comprises a biosynthetic
pathway under complex regulatory control by multiple factors
(1). A key enzyme, tyrosinase,
catalyzes the first and only rate-limiting steps in melanogenesis.
The tyrosinase family genes TYR, tyrosinase related protein-1 and
dopachrome tautomerase, responsible for pigmentation, are
transcriptionally regulated by microphthalmia associated
transcription factor (2). Within
the context of the skin as a stress organ, the significance of
melanogenesis extends beyond the assignment of a color trait.
Melanin pigment serves a critical role in social communication and
protection against the harmful effects of solar radiation (2). The lack of melanin pigment such as
in vitiligo commonly affecting the face, can have significant
psychological effects and impair the quality of life of the
affected individuals. Due to their negative effect on physical
appearance, conditions in which pigment is lacking may act as a
potential barrier to social relationships and cause social anxiety.
The lack of melanin pigment also makes the skin more sensitive to
sunburn (3,4). Melanogenic activity serves as a
unique molecular sensor and transducer of noxious signals and as a
regulator of local homeostasis (5).
Vitiligo is a pigmentary disorder characterized by
the selective destruction of melanocytes (6,7).
To date, the cause of melanocyte death remains unknown. Evidence
indicates that oxidative stress is a pivotal etiological aspect for
the destruction of epidermal melanocytes in patients with vitiligo.
Increased levels of reactive oxygen species (ROS) have been found
in lesional and non-lesional skin in vitro and in
vivo (8,9). The generation of ROS in melanocytes
can disturb cell metabolism, proliferation and differentiation,
which further induces autoimmune responses towards melanocytes,
leading to their reversible cell damage (9,10).
The nuclear factor E2-related factor 2 (Nrf2)
signaling pathway is important in the antioxidative stress
mechanism of cells (11,12). Nrf2 is a transcription factor that
is constitutively degraded through the ubiquitin proteasome pathway
under quiescent conditions. Nrf2-kelch-like ECH-associated protein
1 (Keap1) is an adaptor of the ubiquitin ligase complex that
targets Nrf2 (13,14). Nrf2 is released from the
Nrf2-Keap1 complex under oxidative stress and is transferred to the
nucleus, where it binds to adenylate-uridylate-rich elements and
induces the release of downstream antioxidant genes. Such genes
include heme oxygenase-1 (HO-1), NADH quinine oxidoreductase 1
(Nqo1), glutamate cysteine ligase catalytic subunit (Gclc) and
glutamyl cysteine ligase modulator subunit (Gclm) (12,15). The Nrf2-keap1 signaling pathway
serves a key role in protecting melanocytes from hydrogen peroxide
(H2O2)-induced oxidative damage (16). Dysfunction of the Nrf2 signaling
pathway increases the sensitivity of vitiligo melanocytes to
H2O2-induced oxidative damage (17). The knockdown of Keap1 in
melanocytes promotes cell proliferation and survival. Keap1
silencing in melanocytes also induces melanogenesis through the
HO-1-associated activation of β-catenin (18).
Glycyrrhizin (GR) is a natural compound in the roots
and rhizomes of licorice. Previous studies have suggested that GR
has anti-inflammatory, antiviral, antimicrobial, anticancer,
immunomodulatory, hepatoprotective and cardioprotective effects
(19-22). Studies have shown that GR serves
an important role in the treatment of skin diseases. It ameliorates
imiquimod-induced psoriasis lesions in mice (23). GR also ameliorates the symptoms of
atopic dermatitis in the dinitrochlorobenzene-induced mouse model
(24). GR induces melanogenesis
by elevating the level of cAMP in b16 melanoma cells (25). Our previous study showed that
combined therapy of orally administered GR and UVB improved
active-stage generalized vitiligo (26). GR has also been reported to reduce
oxidative stress by activating the Nrf2 pathway (27-29).
The effect of GR on vitiligo and whether GR can
protect human melanocytes from oxidative stress remains unclear.
The present study investigated the potential protective effect of
GR against oxidative stress in normal human epidermal melanocytes
(NHEMs) and the mechanisms involved. It was found that GR protected
the NHEMs from H2O2-induced oxidative damage
via the Nrf2-dependent induction of HO-1, providing evidence for
the application of GR in the treatment of vitiligo.
Materials and methods
Skin specimens and ethics statement
Skin specimens were obtained from six healthy male
Chinese donors (mean age 6 years) who underwent circumcision in the
Department of Urology of The First Affiliated Hospital of Xi'an
Jiaotong University (Xi'an, China) between December 2017 and
January 2018. The present study followed the guidelines of the
Helsinki Declaration (1964) and subsequent amendments. The study
was approved by the Research Ethics Board of the First Affiliated
Hospital of Xi'an Jiaotong University. The guardians of all healthy
donors provided written informed consent prior to inclusion in the
study.
NHEM culture
Fresh tissues from children undergoing circumcision
in the Department of Urology were obtained and digested for 16 h at
4°C with 0.25% dispase II (Roche Diagnostics, Indianapolis, IN,
USA). The tissues were cut into pieces and 0.25% trypsin was added,
followed by incubation at 37°C for 30 min and subsequent
centrifugation for 5 min at 800 × g at room temperature. The cell
suspension was filtered through a 70-mm cell strainer and seeded in
a 10-cm cell culture dish at a density of 4-6×106
cells/ml. Melanocyte culture medium (Epilife Medium 254, Cascade
Biologics/Invitrogen, Portland, OR, USA) containing human
melanocyte growth supplement was added to the melanocyte culture,
which was incubated at 37°C with 5% CO2. Primary
melanocytes at passages three to eight were used in the present
study.
Antibodies and reagents
The rabbit anti-human Nrf2 antibody (Abcam,
Cambridge, MA, USA; cat. no. ab62352) and Alexa Fluor 488 goat
anti-rabbit IgG (H+L) antibody (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA; cat. no. A-11008) were used.
Analytical grade H2O2 was from Tianjin
Chemical Reagent Factory (Tianjin, China). GR, propidium iodide
(PI) and ZnPP were purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany).
Cell viability assay
Melanocyte viability was assessed using a CCK-8 cell
proliferation test kit (KeyGen Biotech Co., Ltd., Nanjing, China),
according to the manufacturer's instructions. Briefly, the primary
melanocytes were seeded in 96-well plates at a density of
2.5×104 cells/well. H2O2 (0.5 mM)
was added to the plates at room temperature. Morphological changes
in the melanocytes were observed after 24 h with a Nikon Eclipse
TS2 fluorescence microscope (Nikon Corporation, Tokyo, Japan). The
medium was replaced together with 10 µl/well cell proliferation and
toxicity test solution, and the cells were cultured for 1-2 h at
37°C. The color change was monitored and measured at an absorbance
at 450 nm using a microplate reader (Perkin-Elmer, Inc., Waltham,
MA, USA).
ROS detection
The NHEMs were treated with 1 mM GR for 24 h in
6-well plates at a density of 1×106 cells/well at room
temperature. The culture medium was discarded and replaced with
DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing 10 mM
H2-DCFDA (Invitrogen; Thermo Fisher Scientific, Inc.), followed by
incubation at 37°C for 30 min. The cells were then washed with PBS
and assayed by flow cytometry (FACSCanto II) with BD FACSDiva
v8.0.1 software (both from BD Biosciences, San Jose, CA, USA).
Small interfering RNA (siRNA)-targeted
gene silencing
Nrf2 siRNA and control (scrambled) siRNA
oligonucleotides were synthesized by Shanghai GenePharma Co. Ltd.
(Shanghai, China). The sequences are listed in Table I. For siRNA transfection, cells
were plated at a density of 1×106 cells/well in 24-well
plates. At 60% confluence, 30 nM Nrf2 siRNA and 250 µl
Lipofectamine® RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.) in Opti-MEM were mixed and added to cells in
24-well plates at room temperature. The knockdown efficiencies were
assessed by immunoblotting 48 h following siRNA transfection.
 | Table ISequences of siRNAs for Nrf2. |
Table I
Sequences of siRNAs for Nrf2.
| Name | Sequence
(5′-3′) |
|---|
|
Scrambled-siRNA | F:
UUCUCCGAACGUGUCACGUTT |
| R:
ACGUGACACGUUCGGAGAATT |
| Nrf2(1) | F:
GGGAGGAGCUAUUAUCCAUTT |
| R:
AUGGAUAAUAGCUCCUCCCTT |
| Nrf2(2) | F:
GCCCAUUGAUGUUUCUGAUTT |
| R:
AUCAGAAACAUCAAUGGGCTT |
| Nrf2(3) | F:
CCCGUUUGUAGAUGACAAUTT |
| R:
AUUGUCAUCUACAAACGGGTT |
Western blot analysis
The cells were lysed with RIPA lysis buffer and
mixed with 5X loading buffer. The protein sample was quantified by
Bradford assay and was boiled for 10 min and denatured. Each
protein sample (50 µg) was subjected to 10% SDS-PAGE. The separated
proteins were transferred onto a PVDF membrane. Following blocking
with 5% (w/v) dry non-fat milk in PBS for 2 h at room temperature,
the primary antibody was added to the PVDF membrane, followed by
overnight incubation at 4°C. Blotting was performed with primary
antibody against Nrf2 (cat. no. ab62352; Abcam; 1:1,000 dilution).
An HRP-labeled goat-anti-rabbit secondary antibody (cat. no. 7074;
Cell Signaling Technology, Inc., Danvers, MA, USA; 1:5,000
dilution) was then applied for 1 h at 37°C. β-actin (cat. no. 4967;
Cell Signaling Technology, Inc.; 1:1,000 dilution) and Histone H3
(cat. no. 9715; Cell Signaling Technology, Inc.; 1:1,000 dilution)
were used as controls. The membrane was washed three times with
TBST, and the blots were developed using an Enhanced
Chemiluminescence system (Pierce; Thermo Fisher Scientific, Inc.).
ImageJ 1.8.0 (National Institutes of Health, Bethesda, MD, USA) was
used to quantify the gray values of each blot.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using a Universal RNA
Extraction kit (Takara Bio, Inc., Otsu, Japan) and reverse
transcribed into cDNA with PrimeScript RT Master Mix kit (Takara
Bio, Inc., Otsu, Japan). The RT reaction was performed by
incubating the samples at 37°C for 15 min, followed by incubation
at 85°C for 5 sec and 4°C for 5 min. Equal quantities of cDNA were
used for RT-PCR analysis with a SYBR Premix Ex Taq II analysis kit
(Takara Bio, Inc.). The thermocycling conditions were as follows:
95°C for 5 min followed by 45 cycles of denaturation at 95°C for 5
sec, annealing at 60°C for 15 sec, and extension at 72°C for 30
sec. qPCR analyses were performed with the Bio-Rad CFX96™ Real-Time
PCR detection system and analyzed using CFXTM manager 3.0 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The specific primers used
are listed in Table II. Relative
expression levels were normalized to the expression of GAPDH and
were quantified using the comparative 2-ΔΔCq method
(30).
| Table IIPrimer sequences for polymerase chain
reaction. |
Table II
Primer sequences for polymerase chain
reaction.
| Gene | Sequences
(5′-3′) |
|---|
| Nrf2 | F:
CTTGGCCTCAGTGATTCTGAAGTG |
| R:
CCTGAGATGGTGACAAGGGTTGTA |
| HO-1 | F:
CAGGAGCTGCTGACCCATGA |
| R:
AGCAACTGTCGCCACCAGAA |
| NQO-1 | F:
GGATTGGACCGAGCTGGAA |
| R:
AATTGCAGTGAAGATGAAGGCAAC |
| GCLC | F:
GAAGTGGATGTGGACACCAGATG |
| R:
TTGTAGTCAGGATGGTTTGCGATAA |
| GCLM | F:
GGAGTTCCCAAATCAACCCAGA |
| R:
TGCATGAGATACAGTGCATTCCAA |
| GAPDH | F:
ATGACATCAAGAAGGTGGTG |
| R:
CATACCAGGAATGAGCTTG |
Flow cytometric analysis of
apoptosis
Following digestion with 0.25% trypsin, the primary
melanocytes were centrifuged at 800 × g for 5 min at room
temperature and resuspended with 100 µl binding buffer. Each sample
was incubated for 30 min in the dark with 2 μl Annexin
V-FITC and 2 μl PI staining buffer in binding buffer,
following which 300 μl binding buffer was added to each
sample. This was followed by analysis using the FACSCanto II flow
cytometer.
Immunofluorescence staining
The NHEMs were seeded on glass slides in a 12-well
plate. The glass slides were removed from the 12-well plate and
washed twice with PBS. The NHEMs were then fixed with 4%
paraformaldehyde for 30 min at room temperature. Following
permeabilization by incubation in 0.5% Triton X-100 for 15 min, the
NHEMs were blocked with 5% BSA (Beyotime Institute of
Biotechnology, Jiangsu, China) at 37°C for 1 h. The cells were then
incubated with the primary anti-Nrf2 antibody (1:200 dilution)
overnight at 4°C. The cells were washed with PBS five times (3 min
each wash) and then incubated with the Alexa Fluor 488-labeled
secondary antibody (1:200 dilution) at 37°C for 1 h. The nuclei
were counterstained with DAPI. The cells was visualized under a
confocal laser scanning microscope (TCS SP5 Leica GmbH, Wetzlar,
Germany).
Statistical analysis
Data are presented as the mean ± SD Student's t-test
was used to analyze differences between groups. Differences among
multiple groups were analyzed by one-way ANOVA with Dunnett's post
hoc or Tukey's post hoc test. P<0.05 was considered to indicate
a statistically significant difference. Statistical analyses were
performed using GraphPad Prism 6.0 (GraphPad Software Inc., San
Diego, C USA).
Results
GR protects human primary melanocytes
against H2O2-induced cell death
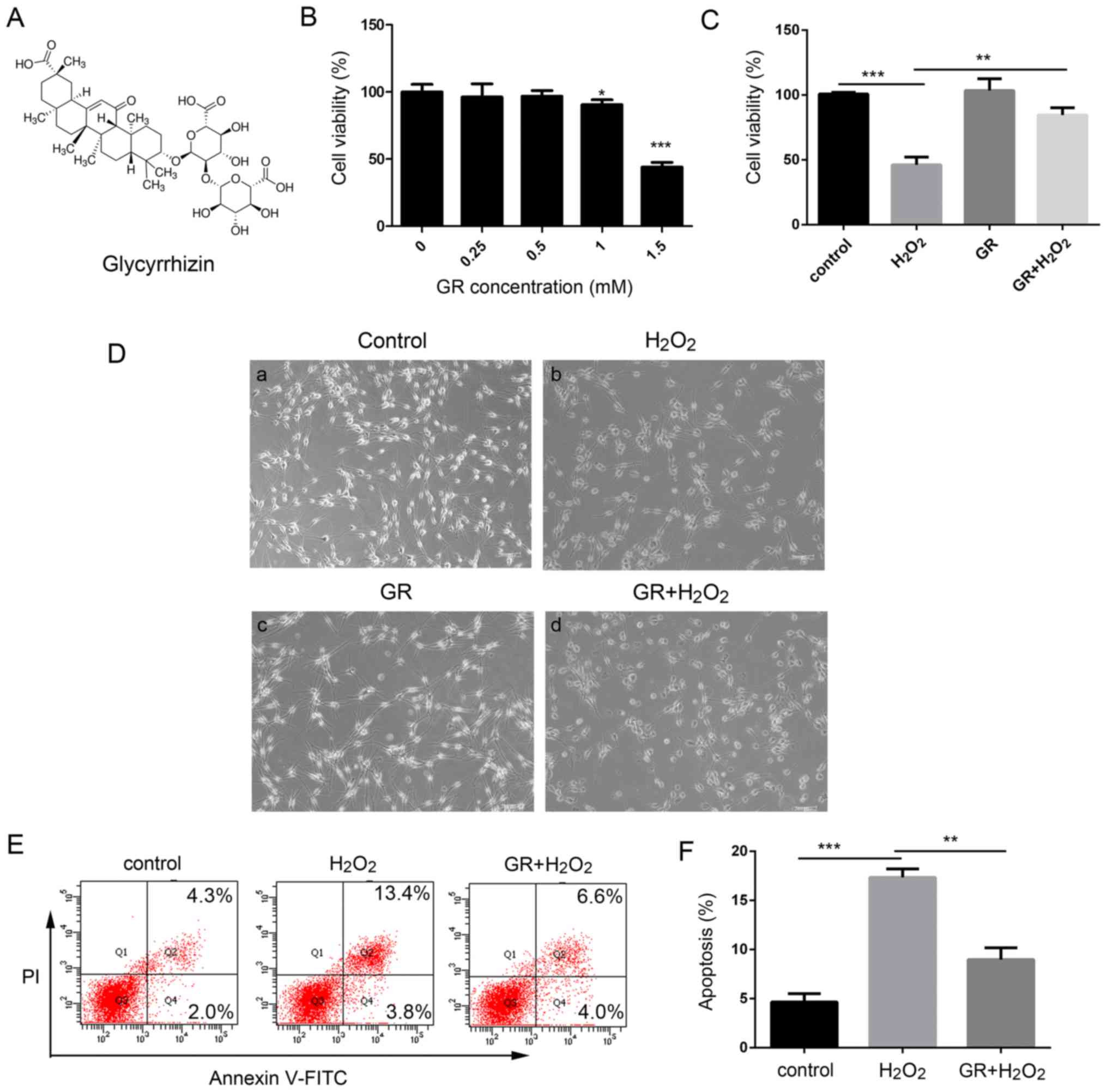
The present study investigated the effects of GR
(Fig. 1A) on the survival rate
and apoptosis of NHEMs treated with H2O2. Our
previous study determined that treatment of melanocytes with 0.5 mM
H2O2 for 24 optimal to induce oxidative
stress (31). As shown in
Fig. 1B there were no significant
changes in cell viability at low doses of GR (0.25, 0.5 and 1 mM),
whereas cell viability was significantly lower following treatment
with a high dose GR (1.5 mM) compared with that in untreated cells.
When the NHEMs were treated with 1.5 mM GR, the cell survival rate
decreased to 48.05±5.35% compared with that in untreated cells
(P<0.05). Therefore, 1 mM GR was used as the working
concentration in the following experiments. The primary human
melanocytes were treated with 0.5 mM H2O in the presence
or absence of 1 mM GR, and cell viability was then determined using
Cell Counting Kit-8 (CCK-8) assays. Following treatment with 0.5 mM
H2O2 for 24 h, morpho logical observation
(Fig. 1D-a-d) revealed the
melanocyte dendrites were shortened or had disappeared (Fig. 1D cell viability was decreased to
48.23±7.25% compared that in the control cells (Fig. 1C). However, pretreatment with 1 mM
GR significantly attenuated H2O2-induced
oxidative damage, as represented by an increase of cell viability
to 82.7±5.45% compared with that in control cells (Fig. 1C) In addition, the NHEMs were
pretreated with 1 mM GR for 24 h prior to 0.5 mM
H2O2 treatment and melanocyte apoptosis was
then examined using flow cytometry. As shown in Fig. 1E and F, following
H2O2 treatment, the percentage of apoptotic
cells increased to 17.5%, compared with 4.5% in the control group,
whereas pretreatment with 1 mM GR markedly inhibited
H2O2-induced apoptotic death, with the
percentage of Annexin V-stained cells at 8.8%.
GR inhibits
H2O2-induced ROS production
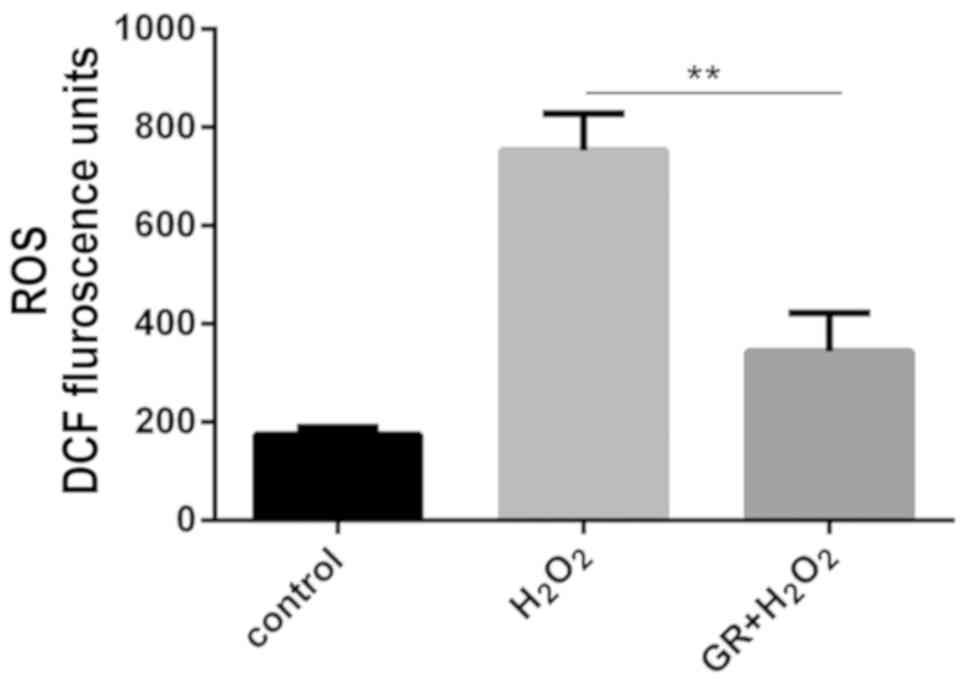
The present study subsequently examined whether
glycyrrhizin protected melanocytes against oxidative stress. The
effects of GR on ROS were measured in NHEMs treated with
H2O2. The ROS level was measured using the
probe H2-DCFH-DA. As shown in Fig.
2, following H2O2 treatment, there was an
increase in the level of DCF fluorescence, and the ROS level in
NHEMs was significantly reduced by GR pretreatment.
Activation of Nrf2 and induction of HO-1
by GR
To examine the molecular mechanism underlying the
effect of GR in reducing oxidative stress, the present study
determined whether GR treatment activated the classical antioxidant
pathway of Nrf2. As shown in Fig.
3A, Nrf2 was mainly localized in cytoplasm of the untreated
NHEMs. However, in cells treated with GR for 24 h, the expression
of Nrf2 in the nucleus was markedly increased. To confirm the
nuclear translocation of Nrf2, cytoplasmic and nuclear protein
fractions of NHEMs were extracted and immunoblotted with an
antibody against Nrf2 (Fig. 3B and
C). The expression of Nrf2 was increased significantly
following GR treatment, which was consistent with the results of
the immunofluorescence. Subsequently, the effects of GR on the
expression levels of four downstream genes of the Nrf2 antioxidant
pathway were examined. As shown in Fig. 3D, GR treatment increased the
expression levels of the four genes, of which the expression of
HO-1 was highly elevated.
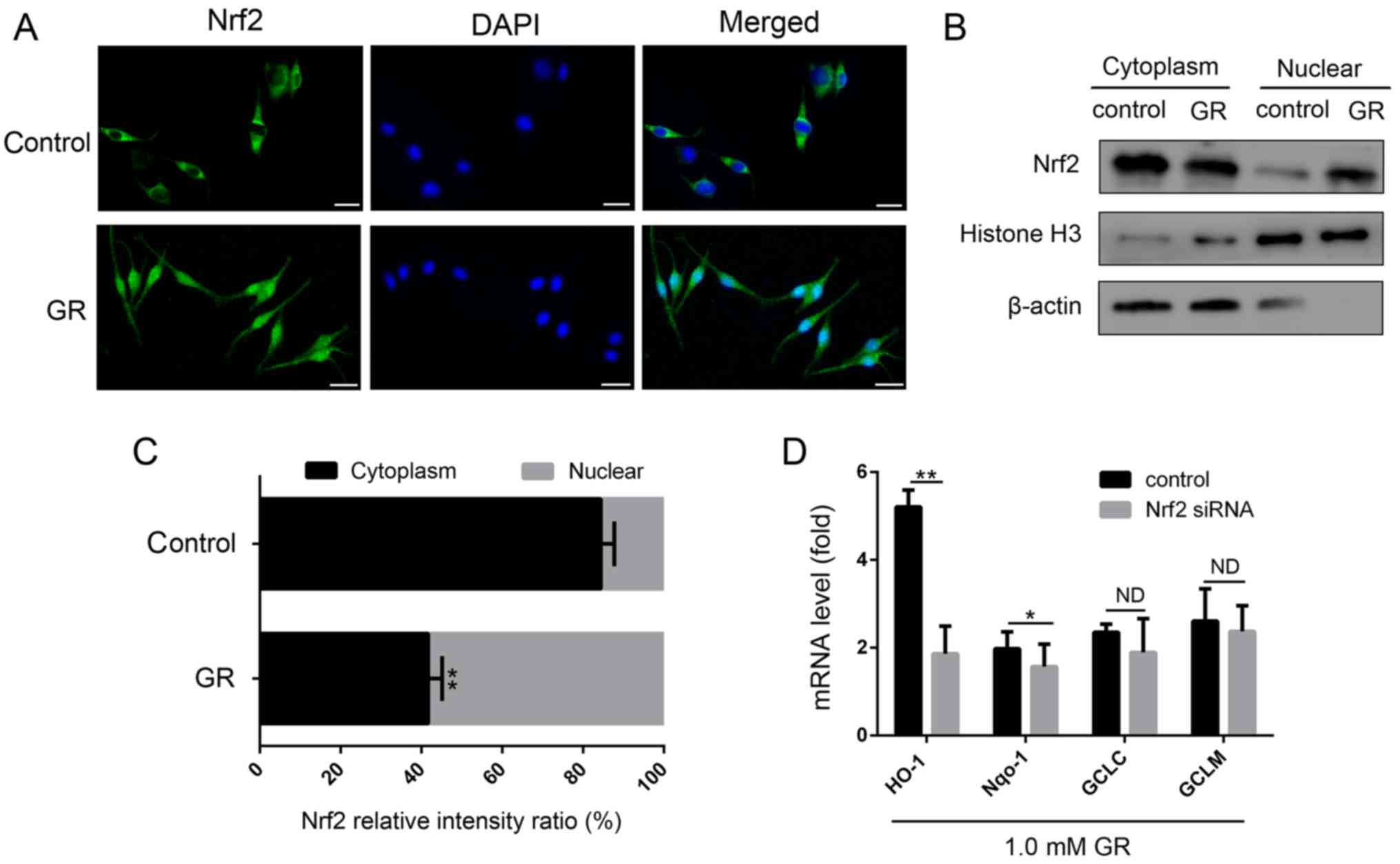 | Figure 3Influence of GR on the activation of
Nrf2 and expression of its target genes. (A) Nrf2
immunofluorescence was examined by confocal microscopy following
treatment of cells with 1.5 mM GR for 24 h (×400 magnification).
Nrf2 was immunostained with an Nrf2-specific antibody (green), and
DAPI was used to stain the nucleus (blue). Scale bar=25 μm.
(B) NHEMs were stimulated with 1.5 mM GR for 24 h, following which
Nrf2 in cytoplasmic and nuclear protein fractions were detected by
western blot analysis and (C) quantified. Histone H3 and β-actin
were used as internal references for nuclear and cytoplasmic
proteins, respectively. (D) NHEMs were transfected with Nrf2 siRNA
or scrambled siRNA for 48 h. mRNA levels of Nrf2 downstream
antioxidant genes (HO-1, NQO-1, GCLC and GCLM) were detected by
reverse transcription-quantitative polymerase chain reaction
analysis. Data are shown as gene expression following normalization
to the expression of GAPDH. *P<0.05 and
**P<0.01 compared with the control. NHEMs, normal
human epidermal melanocytes; Nrf2, nuclear factor E2-related factor
2; GR, glycyrrhizin; siRNA, small interfering RNA; HO-1, heme
oxygenase-1; NQO-1, NADH quinine oxidoreductase 1, GCLC, glutamate
cysteine ligase catalytic subunit; GCLM, glutamyl cysteine ligase
modulator subunit; ND, no difference. |
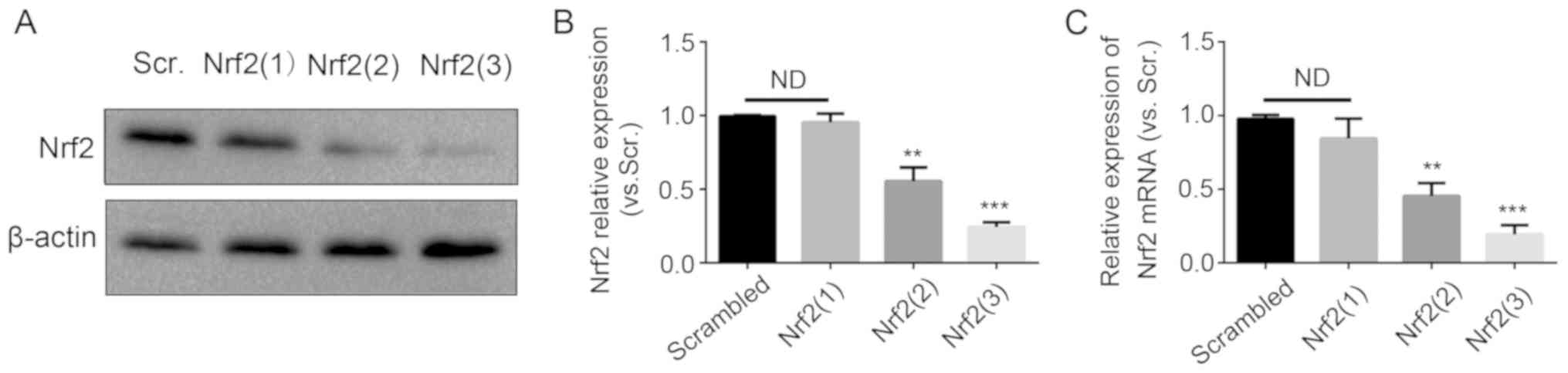
Subsequently, the primary melanocytes were
transfected with Nrf2 siRNAs. Three sequences of Nrf2 siRNAs were
used for knockdown. The Nrf2 knockdown efficiency was evaluated by
western blotting and RT-qPCR analysis (Fig. 4). The most efficient sequence was
the Nrf2 (3) siRNA. The
expression of HO-1 was significantly decreased following
transfection with Nrf2 (3) siRNA,
whereas the expression of the three other genes was not decreased
significantly. Therefore, GR exerted antioxidant effects on
melanocytes mainly through the Nrf2-mediated induction of HO-1.
Nrf2 knockdown reverses the protective
effect of GR on melanocytes against
H2O2-induced cytotoxicity
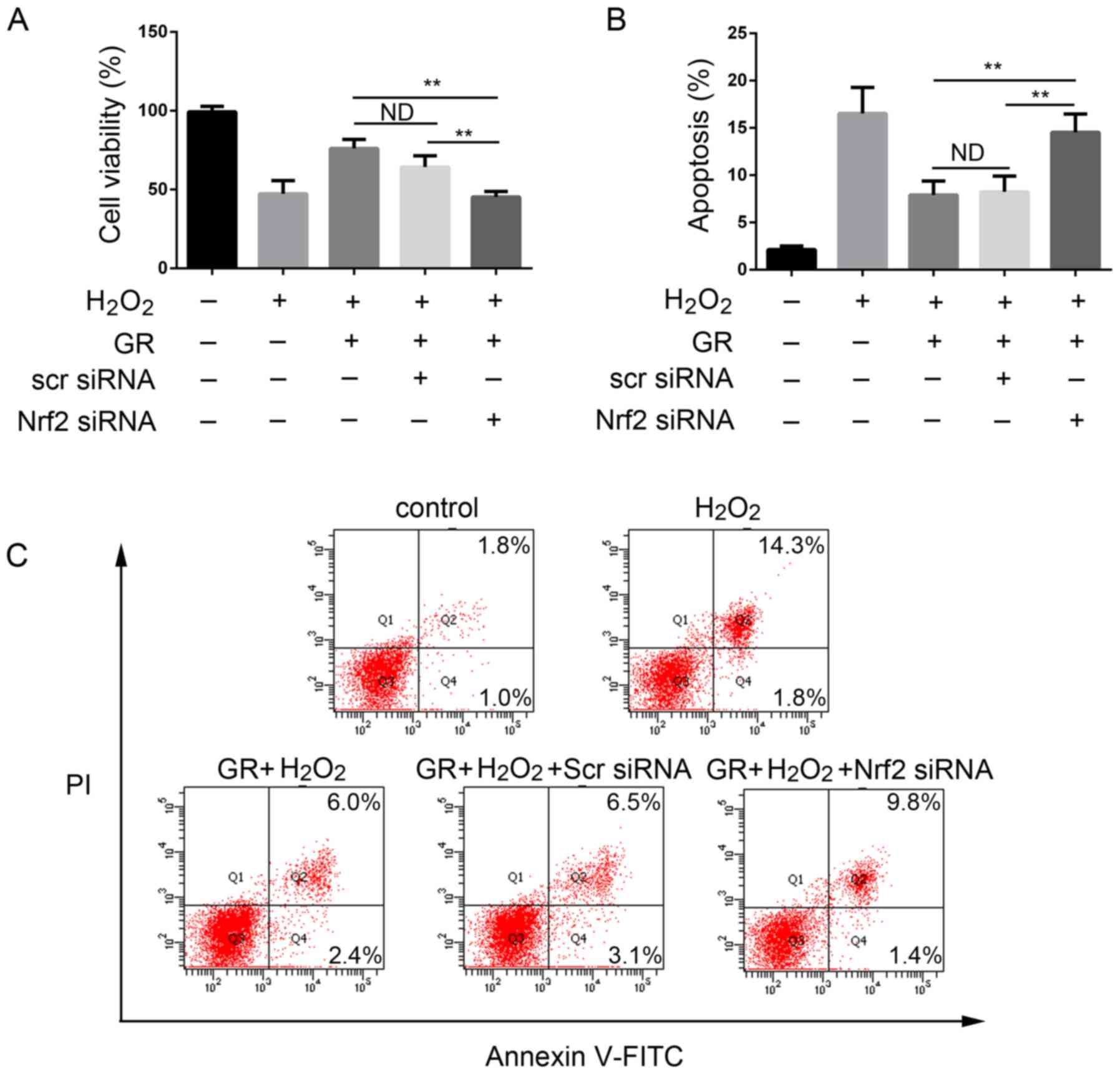
The present study then examined whether Nrf2 has a
direct role in the antioxidant effect of GR. The NHEMs were first
transfected with Nrf2 siRNA or scrambled siRNA for 48 h, followed
by 0.5 mM H2O2 treatment in the presence of 1
mM GR for 24 h. As shown in Fig.
5A, the viability of the NHEMs was significantly decreased
following silencing of Nrf2 compared with that of cells in the
control group. Transfection with scrambled siRNA had no
cytoprotective effect; cell viability was comparable with that of
cells treated with H2O2 and GR. GR
pretreatment had a significant protective effect on NHEMs, and the
apoptotic rate of the cells was decreased. When the Nrf2 gene was
knocked down, the protective effect of GR was decreased
significantly (Fig. 5B and C).
The cells treated with scrambled siRNA exhibited no changes in
GR-induced apoptotic rate under H2O2
treatment.
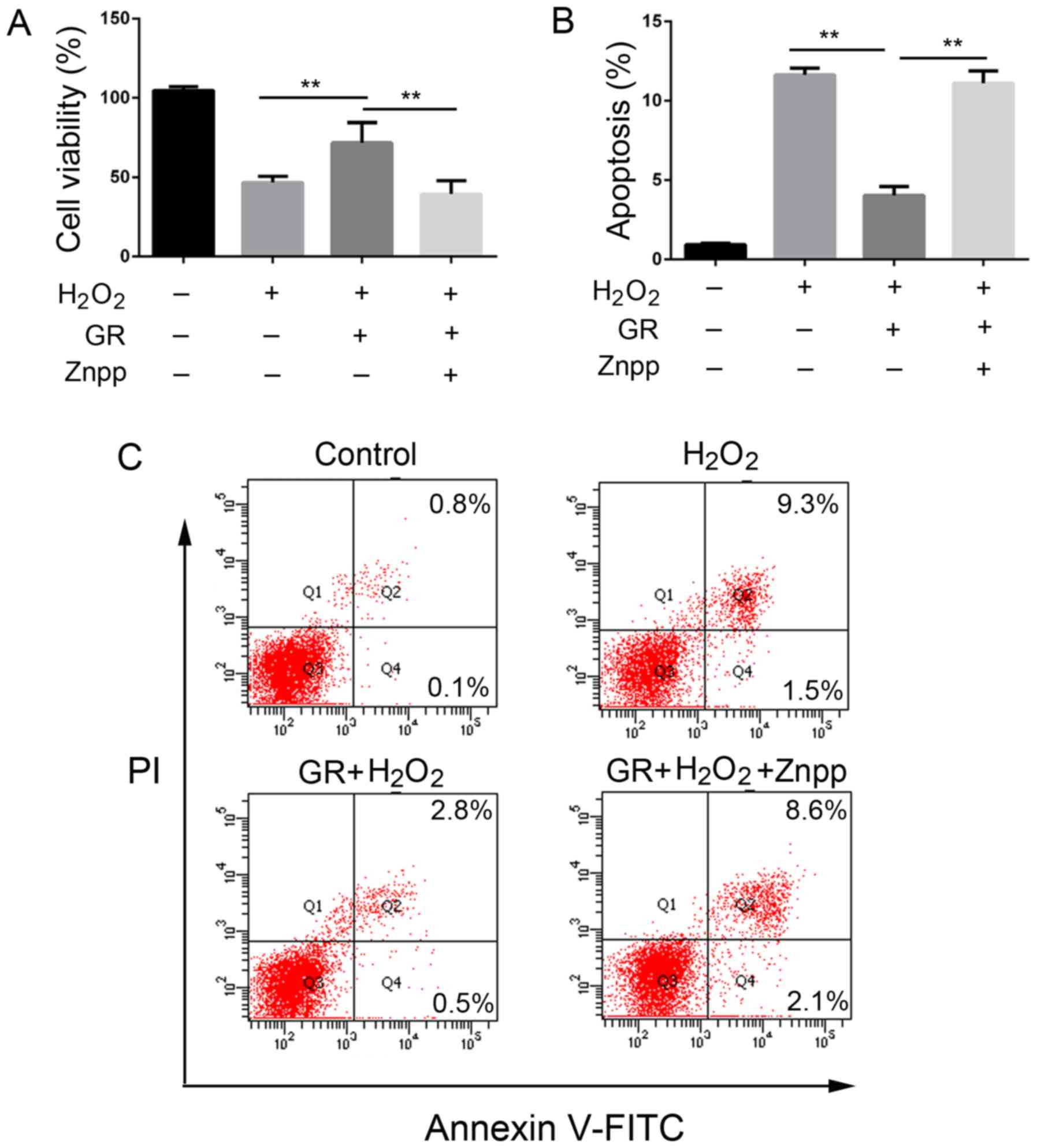
Protective effect of GR on the
cytotoxicity and oxidative stress induced by
H2O2 requires HO-1
To determine the role of the increased expression of
HO-1 in the antioxidative effect of GR on melanocytes, the NHEMs
were treated with ZnPP, an inhibitor of HO-1, for 24 h, followed by
treatment with H2O2 for 24 h. The cells were
then treated with GR for another 24 h. As shown in Fig. 6, pretreatment with GR increased
the viability of the NHEMs and resulted in a significant decrease
of apoptotic cells, whereas the addition of ZnPP reversed the
effect of GR by reducing the viability of the NHEMs and increasing
apoptotic cells (Fig. 6A-C).
Discussion
At present, the main clinical treatment methods for
active generalized vitiligo are narrow-band ultraviolet B light
combined with oral cortisone or another hormone therapy, topical
glucocorticoids, a calcineurin inhibitor (tacrolimus ointment), and
308 nm excimer laser exposure (32,33). Systemic hormones may be considered
for cases of rapid progression (34). However, hormonal side effects
should be considered, and reports indicate that the recurrence rate
following cortisone treatment is high (35). Therefore, the current treatments
for vitiligo are not always effective, and certain treatments are
limited to specific types of vitiligo.
Accumulating evidence shows that oxidative stress
serves a major role in the pathogenesis of vitiligo. Antioxidant
stress-based therapy is a promising strategy for vitiligo. In the
present study, it was found that GR protected human melanocytes
from H2O2-induced oxidative damage via the
Nrf2-dependent induction of HO-1. It has been reported that other
natural compounds, including melatonin and its metabolites, protect
melanocytes from UVB-induced DNA damage through activation of the
Nrf2-dependent pathway (36,37). GR also activates the Nrf2 pathway
in other cell types (28). In the
present study, it was first demonstrated that GR pretreatment
improved the survival rate and reduced the apoptotic rate of
melanocytes under oxidative stress. It was subsequently
demonstrated that GR also activated the Nrf2 antioxidant pathway in
melanocytes, and the gene knockdown of Nrf2 significantly reduced
the effect of GR.
GR activated downstream genes of the Nrf2 pathway,
and siRNA inhibited Nrf2, suggesting that HO-1 serves an important
role in antioxidation. It has been reported that HO-1 is an
important Nrf2 downstream gene in the protection of melanocytes
from oxidative stress (16). The
results obtained in the present study support this finding. Other
downstream genes of Nrf2 had no significant effect on the effects
of GR. The mechanism underlying the effect of GR through Nrf2/HO-1
remains unclear. GR binds to high-mobility group box 1 (HMGB1)
protein and inhibits its cytokine activities as an HMGB1 inhibitor
(38). Our previous study also
showed that oxidative stress stimulation triggers autocrine HMGB1
translocation and release from melanocytes, leading to the
suppressed expression of Nrf2 and downstream genes, which induces
melanocyte apoptosis (31). GR
may also upregulate the Nrf2 pathway by inhibiting HMGB1 and is
involved in antioxidant effects. The Nrf2-induced expression of
HO-1 also inhibits the expression of HMGB1, thereby reducing the
incidence of inflammation (39,40). Such Nrf2-HO-1-HMGB1 feedback loops
may be mechanisms involved in the effects of GR. The underlying
specific mechanisms require further investigation. GR promotes
melanogenesis in melanoma cells by increasing tyrosinase activity.
A limitation of the present study was that melanogenesis was not
assessed in human melanocytes, nor was the effect of GR on
tyrosinase activity in melanocytes examined.
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate that GR protects
melanocytes from oxidative stress by activation of the Nrf2
signaling pathway and inducing the expression of HO-1. Therefore,
GR may be a promising novel therapeutic drug for vitiligo.
Funding
This study was supported by grants from the National
Natural Sciences Foundation of China (grant nos. 81202352 and
81171504) and the Basic Research Program of Natural Science of
Shaanxi Province (grant no. 2017JM8140).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PL conceived and designed the study. KM, WP, DH, XW,
YM and FC performed the experiments, and acquired, analyzed and
interpreted the data; PL drafted and edited the manuscript. All
authors have read and revised the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Board of the First Affiliated Hospital of Xi'an Jiaotong
University. All participants provided written informed consent
prior to inclusion in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Slominski AT, Zmijewski MA, Skobowiat C,
Zbytek B, Slominski RM and Steketee JD: Sensing the environment:
Regulation of local and global homeostasis by the skin's
neuroendocrine system. Adv Anat Embryol Cell Biol. 212:vvii1–115.
2012.PubMed/NCBI
|
|
2
|
Slominski A, Tobin DJ, Shibahara S and
Wortsman J: Melanin pigmentation in mammalian skin and its hormonal
regulation. Physiol Rev. 84:1155–1228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salman A, Kurt E, Topcuoglu V and Demircay
Z: Social anxiety and quality of life in vitiligo and acne patients
with facial involvement: A cross-sectional controlled study. Am J
Clin Dermatol. 17:305–311. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Njoo MD and Westerhof W: Vitiligo.
Pathogenesis and treatment. Am J Clin Dermatol. 2:167–181. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slominski A, Zmijewski MA and Pawelek J:
L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators
of melanocyte functions. Pigment Cell Melanoma Res. 25:14–27. 2012.
View Article : Google Scholar
|
|
6
|
Krüger C and Schallreuter KU: A review of
the worldwide prevalence of vitiligo in children/adolescents and
adults. Int J Dermatol. 51:1206–1212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ezzedine K, Eleftheriadou V, Whitton M and
van Geel N: Vitiligo. Lancet. 386:74–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Picardo M, Dell'Anna ML, Ezzedine K,
Hamzavi I, Harris JE, Parsad D and Taieb A: Vitiligo. Nat Rev Dis
Primers. 1:150112015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schallreuter KU, Moore J, Wood JM, Beazley
WD, Gaze DC, Tobin DJ, Marshall HS, Panske A, Panzig E and Hibberts
NA: In vivo and in vitro evidence for hydrogen peroxide (H O
accumulation in the epidermis of patients with vitiligo and 2 2)
its successful removal by a UVB-activated pseudocatalase. J
Investig Dermatol Symp Proc. 4:91–96. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mohammed GF, Gomaa AH and Al-Dhubaibi MS:
Highlights in pathogenesis of vitiligo. World J Clin Cases.
3:221–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma Q: Role of nrf2 in oxidative stress and
toxicity. Annu Rev Pharmacol Toxicol. 53:401–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu H, Itoh K, Yamamoto M, Zweier JL and
Li Y: Role of Nrf2 signaling in regulation of antioxidants and
phase 2 enzymes in cardiac fibroblasts: Protection against reactive
oxygen and nitrogen species-induced cell injury. FEBS Lett.
579:3029–3036. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Villeneuve NF, Lau A and Zhang DD:
Regulation of the Nrf2-Keap1 antioxidant response by the ubiquitin
proteasome system: An insight into cullinring ubiquitin ligases.
Antioxid Redox Signal. 13:1699–1712. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taguchi K, Motohashi H and Yamamoto M:
Molecular mechanisms of the Keap1-Nrf2 pathway in stress response
and cancer evolution. Genes Cells. 16:123–140. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ichimura Y, Waguri S, Sou YS, Kageyama S,
Hasegawa J, Ishimura R, Saito T, Yang Y, Kouno T, Fukutomi T, et
al: Phosphorylation of p62 activates the Keap1-Nrf2 pathway during
selective autophagy. Mol Cell. 51:618–631. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jian Z, Li K, Liu L, Zhang Y, Zhou Z, Li C
and Gao T: Heme oxygenase-1 protects human melanocytes from
H2O2-induced oxidative stress via the
Nrf2-ARE pathway. J Invest Dermatol. 131:1420–1427. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jian Z, Li K, Song P, Zhu G, Zhu L, Cui T,
Liu B, Tang L, Wang X, Wang G, et al: Impaired activation of the
Nrf2-ARE signaling pathway undermines H response: A possible
mechanism for melanocyte degeneration in 2O2-induced oxidative
stress vitiligo. J Invest Dermatol. 134:2221–2230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JY, Lee H, Lee EJ, Kim M, Kim TG, Kim
HP and Oh SH: Keap1 knockdown in melanocytes induces cell
proliferation and survival via HO-1-associated β-catenin signaling.
J Dermatol Sci. 88:85–95. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee CH, Park SW, Kim YS, Kang SS, Kim JA,
Lee SH and Lee SM: Protective mechanism of glycyrrhizin on acute
liver injury induced by carbon tetrachloride in mice. Biol Pharm
Bull. 30:1898–1904. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Haleagrahara N, Varkkey J and Chakravarthi
S: Cardioprotective effects of glycyrrhizic acid against
isoproterenol-induced myocardial ischemia in rats. Int J Mol Sci.
12:7100–7113. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang ZH, Li T, Tong YG, Chen XJ, Chen XP,
Wang YT and Lu JJ: A systematic review of the anticancer properties
of compounds isolated from Licorice (Gancao). Planta Med.
81:1670–1687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ming LJ and Yin AC: Therapeutic effects of
glycyrrhizic acid. Nat Prod Commun. 8:415–418. 2013.PubMed/NCBI
|
|
23
|
Xiong H, Xu Y, Tan G, Han Y, Tang Z, Xu W,
Zeng F and Guo Q: Glycyrrhizin ameliorates imiquimod-induced
psoriasis-like skin lesions in BALB/c mice and inhibits
TNF-α-induced ICAM-1 expression via NF-κB/MAPK in HaCaT cells. Cell
Physiol Biochem. 35:1335–1346. 2015. View Article : Google Scholar
|
|
24
|
Wang Y, Zhang Y, Peng G and Han X:
Glycyrrhizin ameliorates atopic dermatitis-like symptoms through
inhibition of HMGB1. Int Immunopharmacol. 60:9–17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee J, Jung E, Park J, Jung K, Park E, Kim
J, Hong S, Park J, Park S, Lee S and Park D: Glycyrrhizin induces
melanogenesis by elevating a cAMP level in b16 melanoma cells. J
Invest Dermatol. 124:405–411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mou KH, Han D, Liu WL and Li P:
Combination therapy of orally administered glycyrrhizin and UVB
improved active-stage generalized vitiligo. Braz J Med Biol Res.
49:2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Abo El-Magd NF, El-Mesery M, El-Karef A
and El-Shishtawy MM: Glycyrrhizin ameliorates high fat diet-induced
obesity in rats by activating NrF2 pathway. Life Sci. 193:159–170.
2018. View Article : Google Scholar
|
|
28
|
Kim YM, Kim HJ and Chang KC: Glycyrrhizin
reduces HMGB1 secretion in lipopolysaccharide-activated RAW 264.7
cells and endotoxemic mice by p38/Nrf2-dependent induction of HO-1.
Int Immunopharmacol. 26:112–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu C, Liang C, Sun W, Chen J and Chen X:
Glycyrrhizic acid ameliorates myocardial ischemic injury by the
regulation of inflammation and oxidative state. Drug Des Devel
Ther. 12:1311–1319. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Mou K, Liu W, Miao Y, Cao F and Li P:
HMGB1 deficiency reduces H2 O2-induced
oxidative damage in human melanocytes via the Nrf2 pathway. J Cell
Mol Med. 22:6148–6156. 2018. View Article : Google Scholar :
|
|
32
|
Lotti T, Gori A, Zanieri F, Colucci R and
Moretti S: Vitiligo: New and emerging treatments. Dermatol Ther.
21:110–117. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lotti T, Buggiani G, Troiano M, Assad GB,
Delescluse J, De Giorgi V and Hercogova J: Targeted and combination
treatments for vitiligo. Comparative evaluation of different
current modalities in 458 subjects. Dermatol Ther. 21(Suppl 1):
S20–S26. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wassef C, Lombardi A, Khokher S and Rao
BK: Vitiligo surgical, laser, and alternative therapies: A review
and case series. J Drugs Dermatol. 12:685–691. 2013.PubMed/NCBI
|
|
35
|
Majid I and Imran S: Relapse after
methylprednisolone oral minipulse therapy in childhood vitiligo: A
12-month follow-up study. Indian J Dermatol. 58:113–116. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Skobowiat C, Brożyna AA, Janjetovic Z,
Jeayeng S, Oak ASW, Kim TK, Panich U, Reiter RJ and Slominski AT:
Melatonin and its derivatives counteract the ultraviolet B
radiation-induced damage in human and porcine skin ex vivo. J
Pineal Res. 65:e125012018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Janjetovic Z, Jarrett SG, Lee EF, Duprey
C, Reiter RJ and Slominski AT: Melatonin and its metabolites
protect human melanocytes against UVB-induced damage: Involvement
of NRF2-mediated pathways. Sci Rep. 7:12742017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mollica L, De Marchis F, Spitaleri A,
Dallacosta C, Pennacchini D, Zamai M, Agresti A, Trisciuoglio L,
Musco G and Bianchi ME: Glycyrrhizin binds to high-mobility group
box 1 protein and inhibits its cytokine activities. Chem Biol.
14:431–441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Faridvand Y, Nozari S, Vahedian V, Safaie
N, Pezeshkian M, Haddadi P, Mamipour M, Rezaie-Nezhad A, Jodati A
and Nouri M: Nrf2 activation and down-regulation of HMGB1 and MyD88
expression by amnion membrane extracts in response to the
hypoxia-induced injury in cardiac H9c2 cells. Biomed Pharmacother.
109:360–368. 2019. View Article : Google Scholar
|
|
40
|
Park EJ, Kim YM and Chang KC: Hemin
reduces HMGB1 release by UVB in an AMPK/HO-1-dependent pathway in
human keratinocytes HaCaT cells. Arch Med Res. 48:423–431. 2017.
View Article : Google Scholar : PubMed/NCBI
|