Introduction
A brief period of global brain ischemia causes
neuronal death/loss in vulnerable regions a few days following
reper-fusion. Pyramidal neurons in the Cornu Ammonis 1 (CA1)
subfield of the hippocampus proper are known to be the most
significantly affected neurons following 5 min of transient global
cerebral ischemia (tgCI); tgCI-induced death of pyramidal neurons
in the CA1 subfield occurs at 4-5 days following tgCI, therefore
this characteristic phenomenon is termed as delayed neuronal death
(DND) (1). Numerous factors
participate in the process of the tgCI-induced DND. For example,
glutamate-mediated excitotoxicity, oxidative stress and
neuroinflammatory process including activation of glial cells are
involved in DND (2-6).
Hyperpolarization-activated cation currents
(Ih), which are conducted by
hyperpolarization-activated cyclic nucleotide-gated (HCN) channels,
serve roles in the regulation of neuronal properties including
neuronal rhythmic activity (7).
In the hippocampus, Ih and HCN channels have been
established to participate in controlling neuronal excitability,
dendritic integration, synaptic transmission and plasticity,
long-term potentiation, and learning and memory formation (8-13).
Until now, 4 subunits of HCN channels (HCN1, HCN2, HCN3 and HCN4)
have been identified, and they are expressed in mammalian brains
(14). Among them, HCN1 conducts
Ih with fast kinetics and modest cAMP gating,
whereas HCN2, which is strictly regulated by cAMP, conducts
slow-kinetic Ih (15-17).
It has been demonstrated that alterations in
functions and/or expressions of HCN channels are associated with
abnormal and pathological hyperexcitability (18,19). A previous study has suggested that
the expression levels of HCN channels are increased in reactive
astrocytes in the rat brain following a permanent occlusion of the
middle cerebral artery, which causes permeant focal cerebral
ischemia (20). In the present
study, time-dependent changes of HCN1 and HCN2 protein expression
levels in the hippocampal CA1 subfield, which is the most
vulnerable to tgCI, were investigated following 5 min of tgCI in
gerbils.
Materials and methods
Experimental animals
The 6-month old male Mongolian gerbils (Meriones
unguiculatus) were obtained from the Experimental Animal
Center, Kangwon National University. All experimental procedures
were approved by the Institutional Animal Care and Use Committee at
Kangwon National University (approval no. KW-180124-1). The numbers
of animals used in this study and the suffering caused by the
procedures used in this experiment were minimized
Induction of tgCI
As described in the method of our previous studies
(2,21,22), the surgical procedure for tgCI was
performed as follows: Briefly, gerbils (total n=84) were
anesthetized with a mixture of 2.5% isoflurane in oxygen (34%) and
nitrous oxide (66%), with a modification of methods of previous
studies (23-26), and level of anesthesia was
confirmed by pedal reflex (firm toe pinch). Blood flow to the brain
was completely interrupted through the occlusion of bilateral
common carotid arteries for 5 min, and confirmed by the observation
of the inhibition of blood flow in the central artery in retinae
using an ophthalmoscope (HEINE Optotechnik). The body temperature
was monitored with a rectal temperature probe and controlled under
normothermic (37±0.5̊C) conditions using a thermometric blanket
prior and subsequent to surgery. Sham gerbils were subjected to the
same procedure without the bilateral common carotid artery
occlusion. No tgCI-induced mortality occurred in any of the sham-
or ischemia-operated groups.
Western blot analysis
HCN1 and HCN2 protein levels were analyzed in the
CA1 subfield at designated times (sham, 6, 12 h, 1, 2 and 4 days
after tgCI) using western blot analysis. The designated time
periods (from 6 h to 4 days after tgCI) were selected as
ischemia-induced neuronal death of CA1 pyramidal neurons occurred
~4 days after tgCI and because significant changes in various
factors, which were important in neuronal survival and death, were
significantly altered prior to the occurrence of ischemia-induced
neuronal death (21,22). As described in our previously
published method (22), 7 animals
at each time point were anaesthetized with intraperitoneal
injection of sodium pentobarbital (60 mg/kg) (JW Pharmaceutical
Co., Ltd.) (25-28), and their brains were removed. The
removed brains were serially and transversely cut into 400 µm
thickness on a vibratome (Leica Microsystems GmbH). Tissues of the
CA1 subfield were dissected from the hippocampi with a surgical
blade and homogenized in 50 mM PBS (pH 7.4) containing 0.1 mM EGTA
(pH 8.0), 0.2% Nonidet P-40, 10 mM EDTA (pH 8.0), 15 mM sodium
pyrophosphate, 100 mM β-glycerophosphate, 50 mM NaF, 150 mM NaCl, 2
mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride and 1
mM dithiothreitol (DTT). HCN1 and HCN2 protein concentrations were
determined using a Micro BCA protein assay kit (Pierce; Thermo
Fisher Scientific, Inc.) after the homogenized tissues were
centrifugated at 16,000 × g for 20 min at 4°C. Aliquot containing
total protein (20 mg) was boiled in loading buffer containing 150
mM Tris-HCI (pH 6.8), 6% SDS, 3 mM DTT, 0.3% bromophenol blue and
30% glycerol and then loaded onto 10% polyacrylamide gel. The gel
was transferred to nitrocellulose transfer membranes (Pall
Corporation) following electrophoresis. To reduce background
staining, the membranes were blocked with 5% non-fat dry milk in
PBS containing 0.1% Tween-20 (PBST) for 45 min at room temperature.
After washing three times with PBST (each for 5 min), the membrane
was incubated with rabbit anti-HCN1 (1:400; cat. no. MABN20; EMD
Millipore), rabbit anti-HCN2 (1:400; cat. no. MAB5596; EMD
Millipore), and mouse anti-β-actin (1:5,000; cat. no. A5316;
Sigma-Aldrich; Merck KGaA) antibodies overnight at 4°C. Finally,
the membrane was exposed to horseradish peroxidase-conjugated goat
anti-rabbit secondary antibody (1:5,000; cat. no. 12-348;
Sigma-Aldrich; Merck KGaA) or horseradish peroxidase-conjugated
goat anti-mouse secondary antibody (1:4,000; cat. no. sc-2031;
Santa Cruz Biotechnology, Inc.) for 2 h at room temperature and
visualized with an enhanced chemiluminescence kit (GE Healthcare
Life Sciences). Results of the western blot analysis were scanned,
and densitometric quantification of the bands was performed using
Image J 1.46 software (National Institutes of Health). The
quantification was represented by relative optical density (ROD). A
ratio of the ROD was calibrated as percentages: The sham group was
designated as 100%.
Tissue processing for histology
Brain sections containing the hippocampus of each
group (n=7 for each point in time) were prepared at designated
times (sham, 6, 12 h, 1, 2 and 4 days after tgCI). According to the
method described in our previous studies (2,21,22), the brain tissues were fixed by the
perfusion of 4% paraformaldehyde (in 0.1 M PBS) through the
ascending aorta. Then, the brains were removed and post-fixed with
the same fixative for 8 h at room temperature, then cryoprotected
by infiltration with 30% sucrose. Finally, the brain tissues were
serially sectioned into 25-µm thickness of coronal sections in a
cryostat (Leica Microsystems GmbH) for subsequent analysis.
Cresyl violet (CV) staining
To examine change in the distribution pattern and
morphology of cells or neurons in the hippocampus following tgCI,
the fixed sections were stained with CV, as we descried previously
(29). In brief, we made a
solution of 1.0% (w/v) CV acetate (Sigma) and added glacial acetic
acid (Sigma) to the solution. The sections were stained with the CV
solution for 2 min, washed and dehydrated by immersing in
ethanol.
Immunohistochemistry
According to the methods described in our previous
studies (2,21,22), immunohistochemical staining for
HCN1 and HCN2 was performed. Briefly, the fixed sections were
incubated with rabbit anti-HCN1 (1:100; EMD Millipore) or rabbit
anti-HCN2 (1:100; EMD Millipore) as primary antibodies. The
sections were then exposed to biotinylated goat anti-rabbit IgG
(1:200; Vector Laboratories, Inc.) and streptavidin peroxidase
complex (1:200; Vector Laboratories, Inc.). Finally, the sections
were visualized with 3,3′-diaminobenzidine (in 0.1 M Tris-HCl
buffer). In order to confirm the specificity of each
immunoreaction, a negative control test was performed using
pre-immune serum instead of each primary antibody. No
immunoreactivity was observed in the negative control tests.
For the quantitative analysis of HCN1 and HCN2
immunoreactivity, 6 sections with 120 µm intervals per animal were
selected. According to our published method (22), digital images of the HCN1 and HCN2
immunoreactive structures were captured in the hippocampus with
Axio Imager 2 microscope (Carl Zeiss AG) equipped with an Axiocam
digital camera (Carl Zeiss AG). The captured images were calibrated
into an array of 512×512 pixels. The density of HCN1- and
HCN2-immunoreactive structures was evaluated as an optical density
(OD). Each OD was obtained following the transformation of the mean
gray level using the formula: OD=log (256/mean gray level). The
background was captured from areas adjacent to the measured area
and subtracted. Finally, a ratio of the OD of image file was
calibrated as % (relative optical density, ROD) and analyzed using
NIH Image 1.59 software (National Institutes of Health). A ratio of
the ROD was calibrated as percentages, with the sham group
representing 100%.
Double immunofluorescence staining
To examine the cell type (neurons or glial cells)
exhibiting HCN1 or HCN2 immunoreactivity following tgCI, double
immunofluorescence staining was performed according to our
previously published protocol (21,22). In brief, rabbit anti-HCN1 (1:50;
cat. no. MABN20; EMD Millipore) or rabbit anti-HCN2 (1:50, cat. no.
MAB5596; Millipore)/mouse anti-ATP-sensitive inward rectifier
potassium channel 8 (Kir6.1; 1:100; cat. no. NBP2-59324; Novus
Biologicals, LLC) antibodies were selected to identify pericytes or
mouse anti-glial fibrillary acidic protein (GFAP; 1:200; cat. no.
AB5804; EMD Millipore) antibodies were used to identify astrocytes.
The fixed sections were then incubated in the mixture of antisera
and reacted in a mixture containing fluorescein
isothiocyanate-conjugated goat anti-rabbit IgG (1:200; cat. no.
111-095-003; Jackson ImmunoResearch, West Grove, PA, USA) and
Cy3-conjugated goat anti-mouse IgG (1:200; cat. no. 115-165-003;
Jackson ImmunoResearch Laboratories Inc.). Finally, the
immunoreactions were observed by using a LSM510 META NLO confocal
microscope (Carl Zeiss AG) at ×20 magnification.
Statistical analysis
The data are presented as the means ± standard error
of the means. Differences of the means among the groups were
analyzed by analysis of variance followed by Tukey's Honest
Significance Difference test, using SPSS 17.0 software (IBM Corp.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
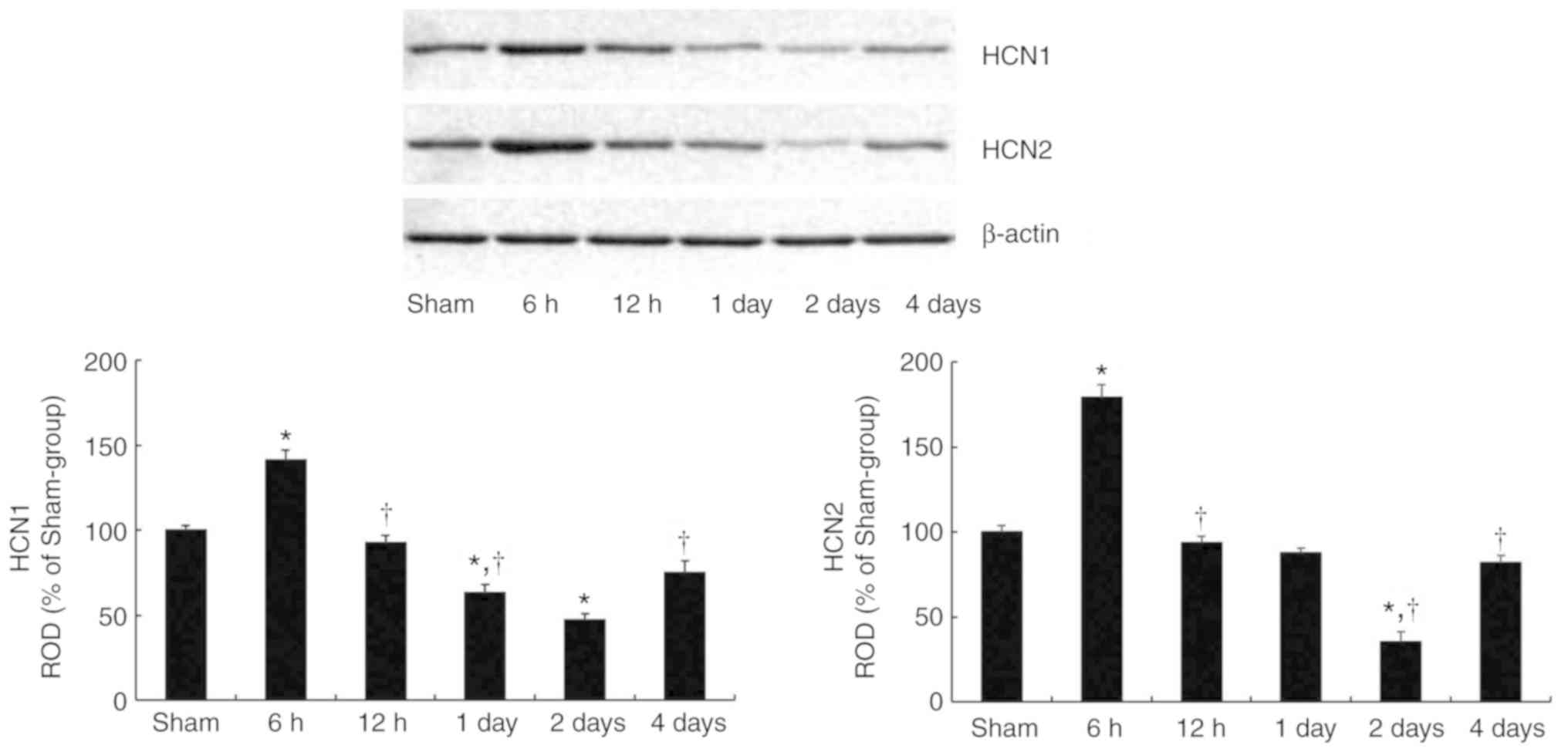
tgCI-induced changes in HCN1 and HCN2
protein levels
HCN1 and HCN2 protein levels in the hippocampal CA1
subfield became significantly altered with time following tgCI
(Fig. 1). HCN1 (P=0.0182) and
HCN2 (P= 0.0076) protein levels were significantly increased at 6 h
after tgCI compared with those in the sham-operated group.
Thereafter, the protein levels of HCN1 and HCN2 gradually decreased
in a time-dependent manner until 2 days after tgCI. And then, at 4
days after tgCI, the HCN1 (P=0.0324) and HCN2 (P=0.0194) protein
levels were significantly increased compared with those at 2 days
after tgCI.
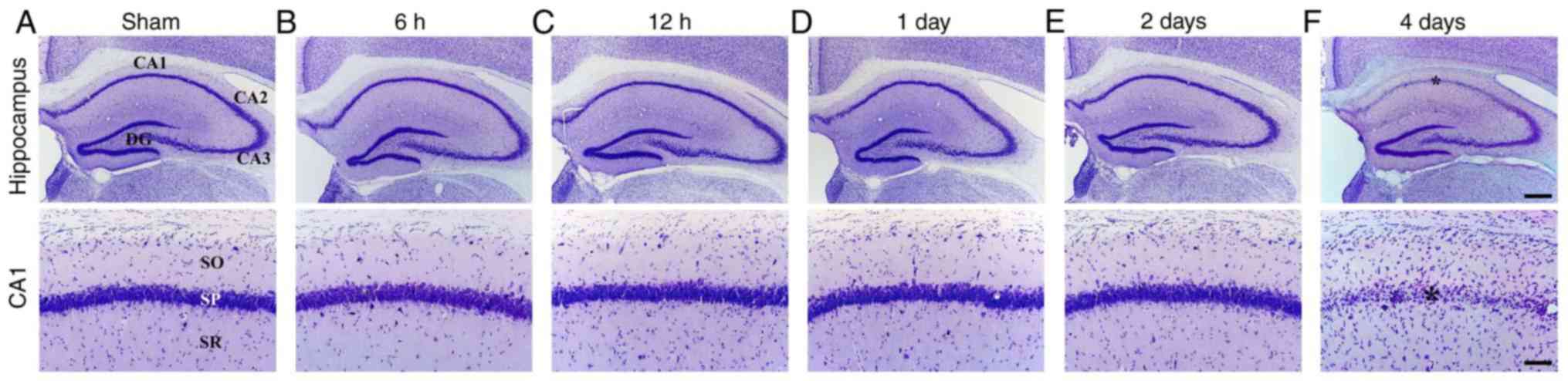
tgCI-induced delayed neuronal death
Ischemia-induced DND in the hippocampus was observed
in the CA1 subfield at 4 days after tgCI using CV staining
(Fig. 2). In the sham-operated
group, CV staining demonstrated the normal distribution of
hippocampal cells in all hippocampal subregions (Fig. 2A). In the ischemia-operated group,
the distribution pattern of CV-positive (CV+) cells was
not altered in the hippocampus until 2 days after tgCI (Fig. 2B-E). However, a significant loss
of CV+ cells was observed in the stratum pyramidale of
the CA1 subfield, not the other subregions, at 4 days after tgCI
(Fig. 2F).
tgCI-induced change in HCN1
immunoreactivity CA1 subfield
In the sham-operated group, HCN1 immuno-reactivity
was primarily detected in neurons of the stratum pyramidale in the
CA1 subfield, which are called CA1 pyramidal cells or neurons
(Fig. 3A). In the
ischemia-operated group, HCN1 immunoreactivity in the CA1 pyramidal
neurons was markedly increased (P=0.0022) at 6 h after tgCI
(Fig. 3B and G). Thereafter, HCN1
immunoreactivity in the CA1 pyramidal neurons was gradually
decreased in a time-dependent manner (Fig. 3C-G). HCN1 immunoreactivity in the
CA1 pyramidal neurons was barely detected in the CA1 pyramidal
neurons at 4 days after tgCI; however, at this time interval, HCN1
immunoreactivity began to be exhibited in cells in the striata
oriens and radiatum, which are non-pyramidal cells (Fig. 3F). Their cell type were examined
by double immunofluorescence staining and it was identified that
HCN1 immunoreactivity was colocalized with Kir6.1-immunoreactive
cells (pericytes) and GFAP-immunoreactive cells (astrocytes), which
are components of the blood brain barrier (BBB), in the striatum
radiatum at 4 days after tgCI (Fig.
3H-M).
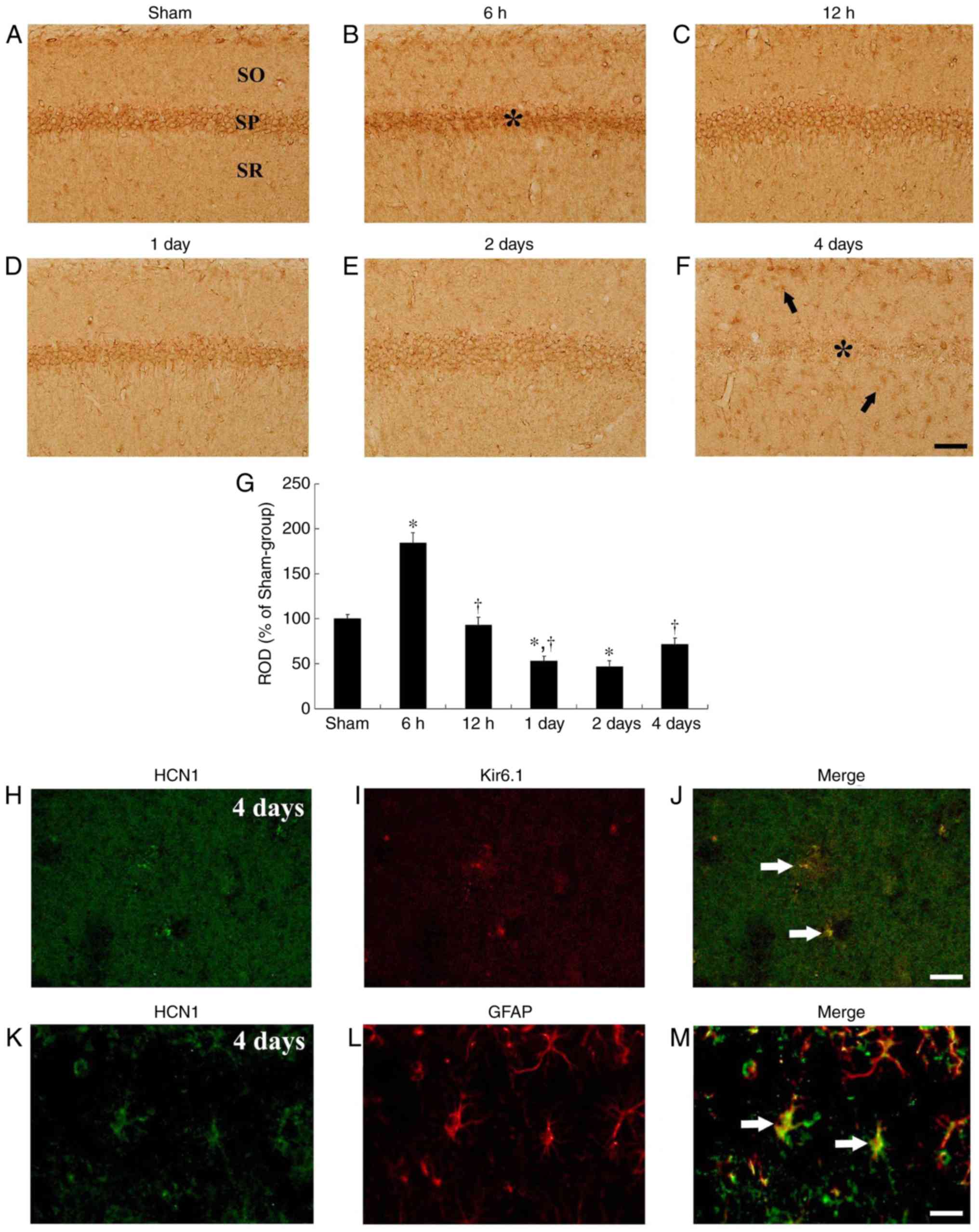 | Figure 3HCN1 immunohistochemistry in the
hippocampal CA1 region. (A-F) HCN1 immunohistochemistry in the CA1
subfield of the (A) sham and (B-F) ischemia-operated groups at (B)
6 h, (C) 12 h, (D) 1 day, (E) 2 days and (F) 4 days. HCN1
immunoreactivity was markedly increased in the CA1 pyramidal
neurons 6 h after tgCI. Thereafter, HCN1 immunoreactivity is
decreased in a time-dependent manner and was barely observed in the
CA1 pyramidal neurons 4 days after tgCI. Scale bar, 50 µm. The
asterisks and the arrows represent CA1 pyramidal neurons and
non-pyramidal cells, respectively. (G) ROD is presented as
percentages of HCN1 immunoreactive structures in the CA1 subfield
following tgCI (n=7 at each point in time. *P<0.05
vs. the sham-operated group, †P<0.05 vs. the previous
timepoint group. Bars indicate the means ± standard error of the
mean. (H-M) Double immunofluorescence staining for (H and K) HCN1
(green), (I) Kir6.1 (red), (L) GFAP (red) and (J and M) merged
images in the stratum pyramidale 4 days after tgCI. HCN1
immunoreactivity was localized in Kir6.1-immunoreactive pericytes
and GFAP-immunoreactive astrocytes. Arrows denote
Kir6.1-immunoreactive pericytes and GFAP-immunoreactive astrocytes
in the respective images. Scale bar, 20 µm. HCN1,
hyperpolarization-activated cyclic nucleotide-gated 1; CA1,
Cornu Ammonis 1; tgCI, transient global cerebral ischemia;
ROD, relative optical density; Kir6.1, ATP-sensitive inward
rectifier potassium channel 8; GFAP, anti-glial fibrillary acidic
protein. |
CA3 subfield
In the sham-operated group, HCN1 immunoreactivity
was clearly demonstrated in pyramidal neurons of the hippocampal
CA3 subfield (Fig. 4A). In the
ischemia-operated group, the level of HCN1 immunoreactivity was not
significantly altered in CA3 pyramidal neurons after tgCI (Fig. 4B-G).
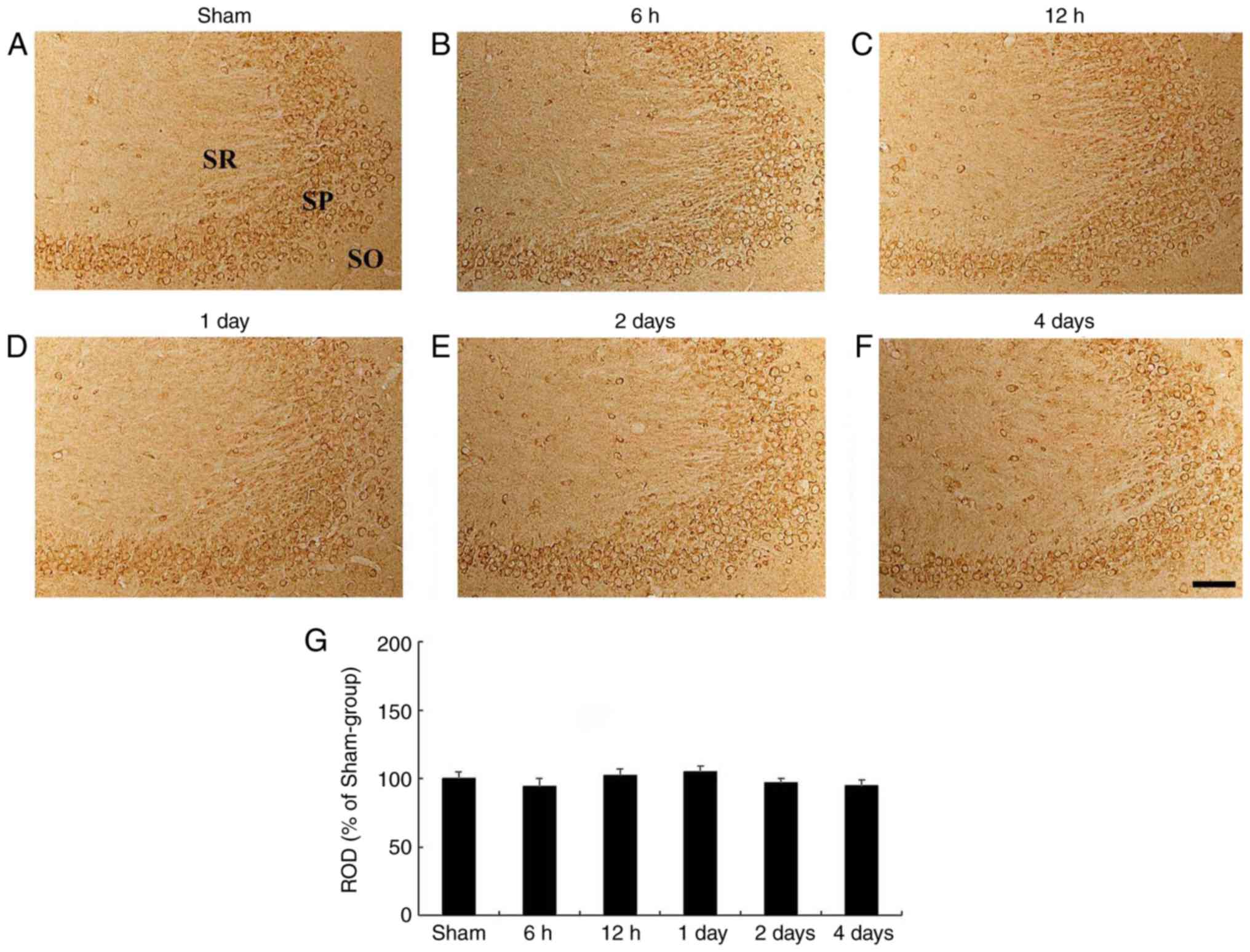 | Figure 4HCN1 immunohistochemistry in the
hippocampal CA3 region. (A-F) HCN1 immunohistochemistry in the CA3
subfield of the (A) sham- and ischemia-operated (B-F) groups at (B)
6 h, (C) 12 h, (D) 1 day, (E) 2 days and (F) 4 days. HCN1
immunoreactivity in the CA3 subfield was not signifi-cantly changed
in any of the experimental groups. Scale bar, 50 µm. (G) ROD is
presented as percentages of HCN1 immunoreactive structures in the
CA3 subfield following tgCI (n=7 at each point in time). HCN1,
hyperpolarization-activated cyclic nucleotide-gated 1; CA3,
Cornu Ammonis 3; SO, stratum oriens; SP, stratum pyramidale;
SR, stratum radiatum; ROD, relative optical density; tgCI,
transient global cerebral ischemia. |
tgCI-induced changes in HCN2
immunoreactivity CA1 subfield
In the sham-operated group, HCN2 immunoreactivity
was identified in the pyramidal neurons and non-pyramidal cells of
the hippocampal CA1 subfield (Fig.
5A). In the ischemia-operated group, HCN2 immunoreactivity in
pyramidal neurons and non-pyramidal cells of the CA1 subfield was
significantly (P<0.001) and transiently increased at 6 h after
tgCI and then gradually decreased until 2 days after tgCI (Fig. 5B-E and G). At 4 days after tgCI,
HCN2 immunoreactivity was significantly decreased in the CA1
pyramidal neurons, indicating that HCN2 immunoreactivity was
exhibited in the nuclei of certain CA1 pyramidal neurons and many
non-pyramidal cells (Fig. 5F). At
this time interval, the type of non-pyramidal cells was examined by
double immunofluorescence staining and it was identified that HCN2
immunoreactivity was colocalized in the Kir6.1-immunoreactive
pericytes and GFAP-immunoreactive astrocytes (Fig. 5H-M).
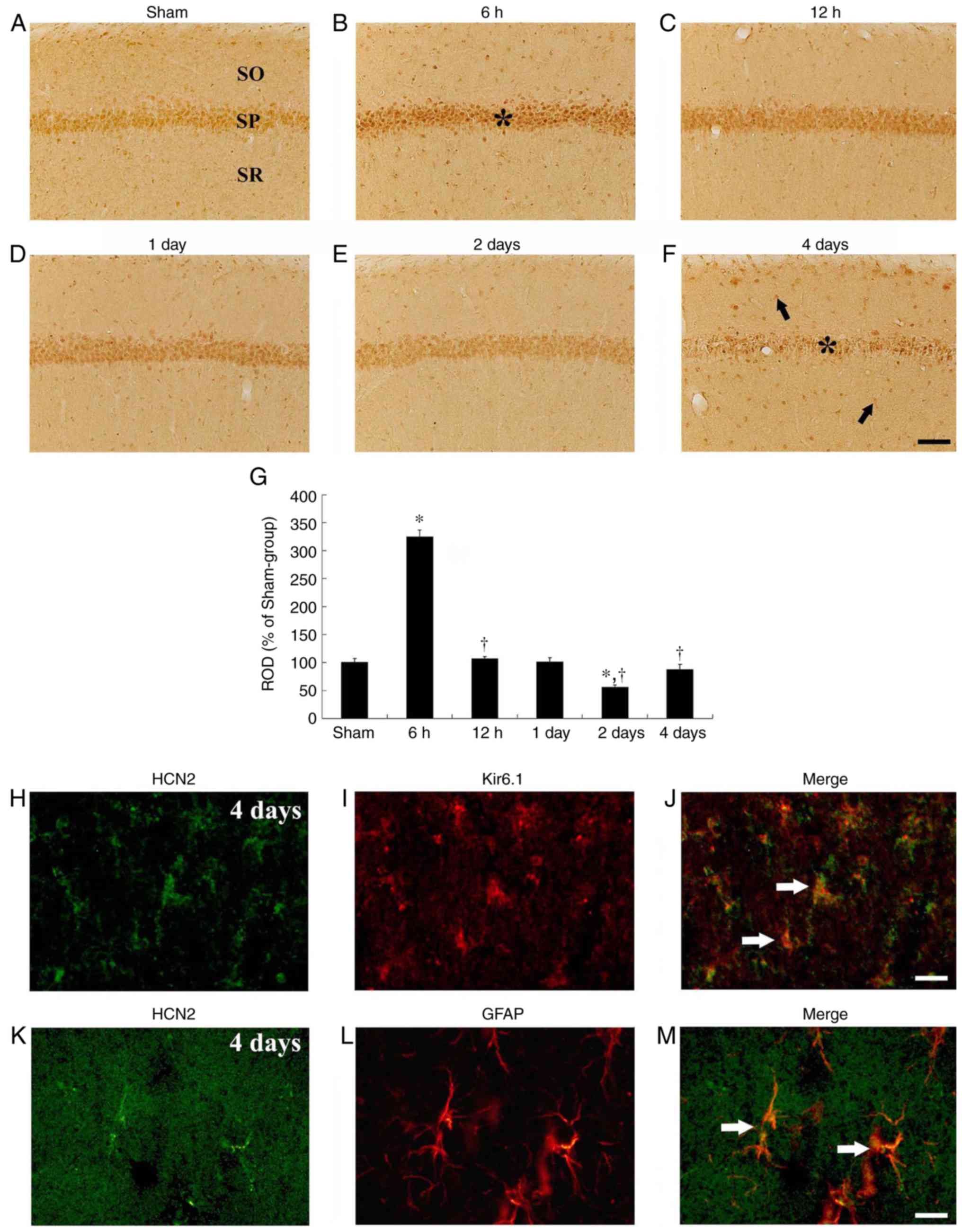 | Figure 5HCN2 immunohistochemistry in the
hippocampal CA1 region. HCN2 immunohistochemistry in the CA1
subfield of the (A) sham and (B-F) isch-emia-operated groups. HCN2
immunoreactivity is distinctly increased in CA1 pyramidal neurons 6
h after tgCI. Thereafter, HCN2 immunoreactivity is decreased with a
time-dependent manner and very weakly observed in CA1 pyramidal
neurons 4 days after tgCI. Scale bar, 50 µm. The asterisks and the
arrows represent the CA1 pyramidal neurons and non-pyramidal cells,
respectively. (G) ROD is presented as percentages of HCN2
immunoreactive structures in the CA1 subfield following tgCI (n=7
at each time point). *P<0.05 vs. the sham operated
group. †P<0.05 vs. the previous timepoint group. Bars
indicate the means ± standard error of the mean. (H-M) Double
immunofluorescence staining for (H and K) HCN1 (green), (I) Kir6.1
(red), (L) GFAP (red) and (J and M) merged images in the stratum
pyramidale 4 days after tgCI. HCN1 immunoreactivity was localized
in Kir6.1-immunoreactive pericytes and GFAP-immunoreactive
astrocytes. Arrows denote Kir6.1-immunoreactive pericytes and
GFAP-immunoreactive astrocytes in the respective images. Scale bar,
20 µm. HCN2, hyperpolarization-activated cyclic nucleotide-gated 2;
CA1, Cornu Ammonis 1; tgCI, transient global cerebral
ischemia; ROD, relative optical density; Kir6.1, ATP-sensitive
inward rectifier potassium channel 8; GFAP, anti-glial fibrillary
acidic protein. |
CA3 subfield
In the sham-operated group, HCN2 immuno-reactivity
was clearly observed in the pyramidal neurons of the hippocampal
CA3 subfield (Fig. 6A). In the
ischemia-operated group, no significant change in HCN2
immunoreactivity was observed in the CA3 pyramidal neurons after
tgCI (Fig. 6B-G).
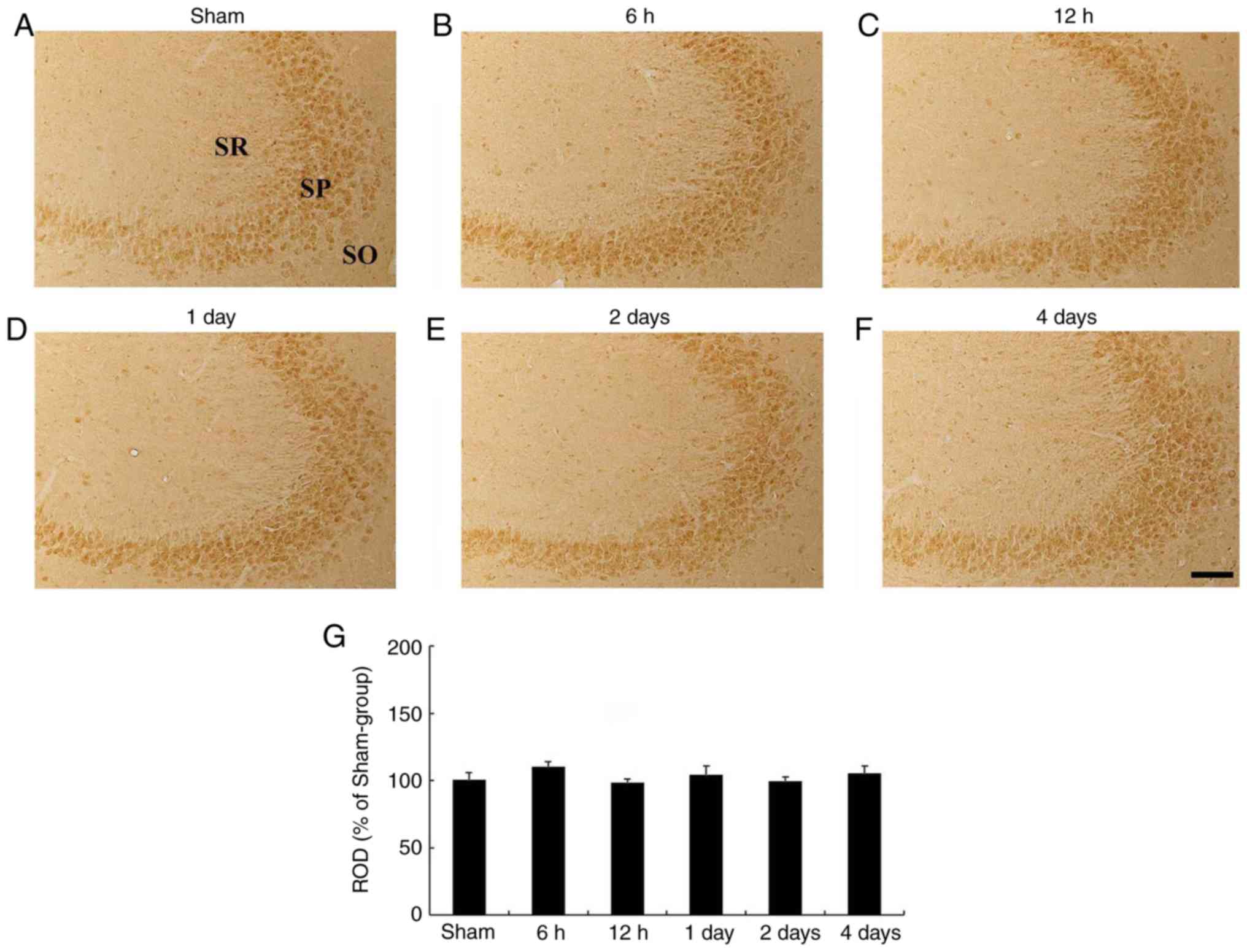 | Figure 6HCN2 immunohistochemistry in the
hippocampal CA3 region. (A-F) HCN2 immunohistochemistry in the CA3
subfield of the (A) sham- and (B-F) ischemia-operated groups at (B)
6 h, (C) 12 h, (D) 1 day, (E) 2 days and (F) 4 days. HCN2
immunoreactivity levels were not altered in the CA3 subfield
following tgCI. Scale bar, 50 µm. (G) ROD is presented as
percentages of HCN2 immunoreactive structures in the CA3 subfield
following tgCI (n=7 at each time point). HCN2,
hyperpolarization-activated cyclic nucleotide-gated 2; CA3,
Cornu Ammonis 3; SO, stratum oriens; SP, stratum pyramidale;
SR, stratum radiatum; ROD, relative optical density. |
Discussion
In our recently published study, it was demonstrated
that unilateral common carotid artery occlusion in gerbils produced
infarcts or selective neuronal death in a number of areas of the
brain, in which the pattern of neuronal death was significantly
different from that in transient global cerebral ischemia induced
by bilateral common carotid artery occlusion (30). Therefore, in the present study,
tgCI was induced by bilateral common carotid artery occlusion in
gerbils. In addition, all gerbils of the experimental groups
survived. This result was in accordance with a previous study that
demonstrated that the survival rate of gerbils following 5 min of
tgCI was 100% (31). Furthermore,
in the present study, ischemia-induced DND in the hippocampus was
observed in the CA1 subfield at 4 days after tgCI, which was
consistent with our previous studies (21,22).
HCN channels have been established to be
predominantly localized in neurons, not in glial cells, in the
mouse cortex and rat hippocampus, and Ih in hippocampal CA1
neurons is primarily mediated by HCN1 and HCN2 (16,17,20,32,33). In addition, it has been suggested
that HCN1 and HCN2 mRNAs are expressed in interneurons and in
principal neurons of the hippocampus in rodents (16,32). In particular, Notomi and Shiqemoto
(34) revealed that the pyramidal
cell layer of all hippocampal subfields in rats exhibited moderate
to intense HCN1 and HCN2 immunoreactivity levels. These results are
consistent with the results of the present study, which indicated
that HCN1 and HCN2 immunoreactivities were observed in pyramidal
neurons and interneurons, not in glial cells, of the hippocampal
CA1 subfield in the sham-operated-group.
It has been demonstrated that the expression levels
of HCN genes are significantly increased in mouse brain tissue 2
weeks after focal cerebral ischemia (20). Conversely, another study suggested
that HCN1 expression was significantly decreased in the hippocampal
CA1 subfield and neocortex of the rat following chronic cerebral
hypoperfusion; this study concluded that downregulated HCN1
expression may lead to impairment of learning and memory function
(35). In animal models of
epilepsy, Oh et al (36)
indicated that HCN1, not HCN2, immunoreactivity was transiently and
significantly increased in the rat hippocampus at 30 min and 12 h
following pilocarpine-induced status epilepticus and suggested that
enhanced HCN1 immuno-reactivity may result from elevated
hippocampal excitability for compensatory responses, as hippocampal
circuit activity may be important for a regulation of HCN
expression (36,37). In the present study, it was
identified that HCN1 and HCN2 immunoreactivity in the CA1 pyramidal
neurons were transiently and markedly increased at 6 h after tgCI
and decreased in a time-dependent manner thereafter. Therefore, it
may be hypothesized that transient elevations of HCN1 and HCN2
immunoreactivities in CA1 pyramidal neurons at 6 h after tgCI may
be one of compensatory responses to the tgCI-induced alteration in
hippocampal excitability and that time-dependent decreases of HCN1
and HCN2 immunoreactivities thereafter in the CA1 pyramidal neurons
may be associated with the process of tgCI-induced DND.
In the present study, it was identified that HCN1
and HCN2 immunoreactivities in the CA1 pyramidal neurons were
barely detected at 4 days after tgCI; however, at this time point,
HCN1 and HCN2 immunoreactivities were observed in
Kir6.1-immunoreactive pericytes and GFAP-immunoreactive astrocytes.
Unfortunately, why HCN1 and HCN2 immunoreac-tivity began to be
expressed in pericytes and astrocytes in the ischemic CA1 subfield
4 days after tgCI cannot be explained clearly. However, it has been
demonstrated that pericytes and astrocytes serve important roles in
the protection of nervous tissue against oxidative stress following
ischemia, by facilitation of BBB repair and attenuation of
neuroinflammatory response, respectively (38-41). In particular, Honsa et al
(20) demonstrated that HCN
channels were markedly expressed by reactive astrocytes in the CA1
subfield of the rat hippocampus following global cerebral ischemia
induced by transient bilateral common carotid artery occlusion
combined with hypoxia and that, in this model, electrophysiological
properties of reactive astrocytes were significantly altered
following ischemia; they concluded that HCN channels may
participate in the regeneration of ischemic tissue and ionic
homeostasis maintenance.
There are certain important limitations of the
present study. Firstly, data concerning behavioral and cognitive
changes following tgCI were not described, although it has been
established the tgCI-induced loss of hippocampal CA1 pyramidal
neurons leads to impairment of hippocampal-dependent learning and
memory and that significant changes in motor behavior are exhibited
at 1 day after tgCI in gerbils (29,42). Secondly, the present study did not
investigate how HCN1 and HCN2 expression affected the death of CA1
pyramidal neurons following tgCI at a molecular level. Therefore,
further studies are required to investigate the functional effects
of HCN1 and/or HCN2 on tgCI-induced neuronal death.
In brief, HCN1 and HCN2 immunoreactivities were
altered in pyramidal neurons of the hippocampal CA1 subfield in a
time-dependent manner following tgCI, and HCN1 and HCN2
immunoreactivity began to be observed in pericytes and astrocytes
in ischemic CA1 subfield at 4 days after tgCI. These results
indicate that tgCI-induced changes in HCN1 and HCN2 expression
levels may be closely associated with tgCI-induced DND of pyramidal
neurons in the hippocampal CA1 subfield.
Acknowledgements
Not applicable.
Funding
The present study was performed with the support of
'Cooperative Research Program for Agriculture Science and
Technology Development (project no. PJ01321101)' Rural Development
Administration, Republic of Korea, by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Science, ICT & Future Planning (grant
no. NRF-2017R1A2B4009079), and by the NRF funded by the Ministry of
Education (grant no. NRF-2017R1D1A1B03029311).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JHP, DWK, MHW and CHL were responsible for
experimental design, data collection, data analysis, and manuscript
writing. CWP, YEP and HAL performed the experiments, TKL and JHA
performed data analysis and provided critical comments throughout
the process of the present study. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All experimental procedures were approved by the
Institutional Animal Care and Use Committee at Kangwon National
University (approval no. KW-180124-1).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kirino T: Delayed neuronal death in the
gerbil hippocampus following ischemia. Brain Res. 239:57–69. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Park JH, Kim YH, Ahn JH, Choi SY, Hong S,
Kim SK, Kang IJ, Kim YM, Lee TK, Won MH and Lee CH: Atomoxetine
protects against NMDA receptor-mediated hippocampal neuronal death
following transient global cerebral ischemia. Curr Neurovasc Res.
14:158–168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Saito K, Suyama K, Nishida K, Sei Y and
Basile AS: Early increases in TNF-alpha, IL-6 and IL-1 beta levels
following transient cerebral ischemia in gerbil brain. Neurosci
Lett. 206:149–152. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sharma SS and Gupta S: Neuroprotective
effect of MnTMPyP, a superoxide dismutase/catalase mimetic in
global cerebral isch-emia is mediated through reduction of
oxidative stress and DNA fragmentation. Eur J Pharmacol. 561:72–79.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sugawara T, Lewén A, Noshita N, Gasche Y
and Chan PH: Effects of global ischemia duration on neuronal,
astroglial, oligodendroglial, and microglial reactions in the
vulnerable hippocampal CA1 subregion in rats. J Neurotrauma.
19:85–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Swanson RA, Ying W and Kauppinen TM:
Astrocyte influences on ischemic neuronal death. Curr Mol Med.
4:193–205. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Biel M, Wahl-Schott C, Michalakis S and
Zong X: Hyperpolarization-activated cation channels: From genes to
function. Physiol Rev. 89:847–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He W, Xu X, Lv Q and Guo L: Low dose
ZD7288 attenuates the ischemia/reperfusion-induced impairment of
long-term potentiation induction at hippocampal Schaffer
collateral-CA1 synapses. Cell Mol Neurobiol. 34:611–617. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang CC and Hsu KS: Reexamination of the
role of hyperpolarization-activated cation channels in short- and
long-term plasticity at hippocampal mossy fiber synapses.
Neuropharmacology. 44:968–981. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo P, Lu Y, Li C, Zhou M, Chen C, Lu Q,
Xu X, He Z and Guo L: Long-lasting spatial learning and memory
impairments caused by chronic cerebral hypoperfusion associate with
a dynamic change of HCN1/HCN2 expression in hippocampal CA1 region.
Neurobiol Learn Mem. 123:72–83. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lupica CR, Bell JA, Hoffman AF and Watson
PL: Contribution of the hyperpolarization-activated current (I(h))
to membrane potential and GABA release in hippocampal interneurons.
J Neurophysiol. 86:261–268. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mellor J, Nicoll RA and Schmitz D:
Mediation of hippocampal mossy fiber long-term potentiation by
presynaptic Ih channels. Science. 295:143–147. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nolan MF, Malleret G, Dudman JT, Buhl DL,
Santoro B, Gibbs E, Vronskaya S, Buzsáki G, Siegelbaum SA, Kandel
ER and Morozov A: A behavioral role for dendritic integration: HCN1
channels constrain spatial memory and plasticity at inputs to
distal dendrites of CA1 pyramidal neurons. Cell. 119:719–732.
2004.PubMed/NCBI
|
|
14
|
Monteggia LM, Eisch AJ, Tang MD, Kaczmarek
LK and Nestler EJ: Cloning and localization of the
hyperpolarization-activated cyclic nucleotide-gated channel family
in rat brain. Brain Res Mol Brain Res. 81:129–139. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Robinson RB and Siegelbaum SA:
Hyperpolarization-activated cation currents: From molecules to
physiological function. Annu Rev Physiol. 65:453–480. 2003.
View Article : Google Scholar
|
|
16
|
Santoro B, Chen S, Luthi A, Pavlidis P,
Shumyatsky GP, Tibbs GR and Siegelbaum SA: Molecular and functional
heterogeneity of hyperpolarization-activated pacemaker channels in
the mouse CNS. J Neurosci. 20:5264–5275. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wahl-Schott C and Biel M: HCN channels:
Structure, cellular regulation and physiological function. Cell Mol
Life Sci. 66:470–494. 2009. View Article : Google Scholar
|
|
18
|
Baruscotti M, Bottelli G, Milanesi R,
DiFrancesco JC and DiFrancesco D: HCN-related channelopathies.
Pflugers Arch. 460:405–415. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
DiFrancesco JC and DiFrancesco D:
Dysfunctional HCN ion channels in neurological diseases. Front Cell
Neurosci. 6:1742015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Honsa P, Pivonkova H, Harantova L, Butenko
O, Kriska J, Dzamba D, Rusnakova V, Valihrach L, Kubista M and
Anderova M: Increased expression of hyperpolarization-activated
cyclic nucleotide-gated (HCN) channels in reactive astrocytes
following ischemia. Glia. 62:2004–2021. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee CH, Park JH, Cho JH, Ahn JH, Yan BC,
Lee JC, Shin MC, Cheon SH, Cho YS, Cho JH, et al: Changes and
expressions of Redd1 in neurons and glial cells in the gerbil
hippocampus proper following transient global cerebral ischemia. J
Neurol Sci. 344:43–50. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park JH, Shin BN, Ahn JH, Cho JH, Kim IH,
Kim DW, Won MH, Hong S, Cho JH and Lee CH: Ischemia-induced changes
of PRAS40 and p-PRAS40 immunoreactivities in the gerbil
hippo-campal CA1 region after transient cerebral ischemia. Cell Mol
Neurobiol. 36:821–828. 2016. View Article : Google Scholar
|
|
23
|
Du X, Wang D, Li Y, Huo X, Li C, Lu J,
Wang Y, Guo M and Chen Z: Newly breeding an inbred strain of
ischemia-prone Mongolian gerbils and its reproduction and genetic
characteristics. Exp Anim. 67:83–90. 2018. View Article : Google Scholar :
|
|
24
|
Zhu XL, Yan BC, Tang C, Tang C, Qiu GW, Wu
Y, Wang Y and Bo P: Neuroprotective effect of Paeoniae Radix Rubra
on hippocampal CA1 region of mice induced by transient focal
cerebral ischemia via anti-gliosis and anti-oxidant activity. Chin
Herb Med. 11:86–91. 2019. View Article : Google Scholar
|
|
25
|
Davies LM, MacLellan CL, Corbett DR and
Colbourne F: Post-ischemic diazepam does not reduce hippocampal CA1
injury and does not improve hypothermic neuroprotection after
forebrain ischemia in gerbils. Brain Res. 1013:223–229. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shughrue P and Merchenthaler I: Estrogen
prevents the loss of CA1 hippocampal neurons in gerbils after
ischemic injury. Neuroscience. 116:851–861. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carpenter JW and Marion CJ: Exotic animal
formulary. Elsevier; St Louis, Missouri: 2018
|
|
28
|
Ahn JH, Shin BN, Song M, Kim H, Park JH,
Lee TK, Park CW, Park YE, Lee JC, Yong JH, et al: Intermittent
fasting increases the expressions of SODs and catalase in granule
and polymorphic cells and enhances neuroblast dendrite complexity
and maturation in the adult gerbil dentate gyrus. Mol Med Rep.
19:1721–1727. 2019.PubMed/NCBI
|
|
29
|
Lee JC, Park JH, Ahn JH, Kim IH, Cho JH,
Choi JH, Yoo KY, Lee CH, Hwang IK, Cho JH, et al: New GABAergic
neurogenesis in the hippocampal CA1 region of a gerbil model of
long-term survival after transient cerebral ischemic injury. Brain
Pathol. 26:581–592. 2016. View Article : Google Scholar
|
|
30
|
Ahn JH, Song M, Kim H, Lee TK, Park CW,
Park YE, Lee JC, Cho JH, Kim YM, Hwang IK, et al: Differential
regional infarction, neuronal loss and gliosis in the gerbil
cerebralhemisphere following 30 min of unilateral common carotid
artery occlusion. Metab Brain Dis. 34:223–233. 2019. View Article : Google Scholar
|
|
31
|
Kirino T, Tamura A and Sano K: A
reversible type of neuronal injury following ischemia in the gerbil
hippocampus. Stroke. 17:455–459. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bender RA, Brewster A, Santoro B, Ludwig
A, Hofmann F, Biel M and Baram TZ: Differential and age-dependent
expression of hyperpolarization-activated, cyclic nucleotide-gated
cation channel isoforms 1-4 suggests evolving roles in the
developing rat hippocampus. Neuroscience. 106:689–698. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Seo H, Seol MJ and Lee K: Differential
expression of hyper-polarization-activated cyclic nucleotide-gated
channel subunits during hippocampal development in the mouse. Mol
Brain. 8:132015. View Article : Google Scholar
|
|
34
|
Notomi T and Shigemoto R:
Immunohistochemical localization of Ih channel subunits, HCN1-4 in
the rat brain. J Comp Neurol. 471:241–276. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li S, He Z, Guo L, Huang L, Wang J and He
W: Behavioral alterations associated with a down regulation of HCN1
mRNA in hippocampal cornus ammon 1 region and neocortex after
chronic incomplete global cerebral ischemia in rats. Neuroscience.
165:654–661. 2010. View Article : Google Scholar
|
|
36
|
Oh YJ, Na J, Jeong JH, Park DK, Park KH,
Ko JS and Kim DS: Alterations in hyperpolarization-activated cyclic
nucleotidegated cation channel (HCN) expression in the hippocampus
following pilocarpine-induced status epilepticus. BMB Rep.
45:635–640. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brewster A, Bender RA, Chen Y, Dube C,
Eghbal-Ahmadi M and Baram TZ: Developmental febrile seizures
modulate hippocampal gene expression of hyperpolarization-activated
channels in an isoform- and cell-specific manner. J Neurosci.
22:4591–4599. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cai W, Liu H, Zhao J, Chen LY, Chen J, Lu
Z and Hu X: Pericytes in brain injury and repair after ischemic
stroke. Transl Stroke Res. 8:107–121. 2017. View Article : Google Scholar :
|
|
39
|
Sidoryk-Wegrzynowicz M, Wegrzynowicz M,
Lee E, Bowman AB and Aschner M: Role of astrocytes in brain
function and disease. Toxicol Pathol. 39:115–123. 2011. View Article : Google Scholar
|
|
40
|
Tian W, Sawyer A, Kocaoglu FB and
Kyriakides TR: Astrocyte-derived thrombospondin-2 is critical for
the repair of the blood-brain barrier. Am J Pathol. 179:860–868.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang S, Jin H, Zhu Y, Wan Y, Opoku EN, Zhu
L and Hu B: Diverse functions and mechanisms of pericytes in
ischemic stroke. Curr Neuropharmacol. 15:892–905. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Janać B, Radenović L, Selaković V and
Prolić Z: Time course of motor behavior changes in Mongolian
gerbils submitted to different durations of cerebral ischemia.
Behav Brain Res. 175:362–373. 2006. View Article : Google Scholar
|