Introduction
There is an increasing need for the production of
recombinant therapeutic proteins generated via a range of
transgenic techniques, with the optimal approach to such expression
being to produce these recombinant proteins in bioreactors such as
bacteria, yeast, plants, mammalian cells, or transgenic animals
(1-7). Among these bioreactors, mammary
gland bioreactors in transgenic animals offer the advantage of
being fully compatible with humans and being approved by the FDA
(8). Such mammary gland-derived
recombinant proteins have already been implemented for clinical use
(9) For example, recombinant
human antithrombin III (Atryn®) is produced from the
milk of transgenic goats (10).
Such mammary gland bioreactors are highly advantageous for the
production of those proteins that require post-translational
modifications in order to mediate their stability or activity
(11). Producing recombinant
proteins in mammary glands therefore represents a more profitable
approach to the production of recombinant human proteins. Pantano
et al suggested that relatively few mammary cells in
transgenic animals ultimately express recombinant proteins
(12), underscoring that there is
an urgent need to determine how to bolster the in vivo
expression of these recombinant proteins using optimized expression
vector systems.
Achieving high-level production of recombinant
proteins within the milk of transgenic animals depends upon
ensuring high-level transcription of the introduced cDNA. This
makes it essential to select appropriate cis-acting elements,
including promoters and enhancers, for the introduced genes. Large
quantities of β-casein proteins are produced in goats during
lactation in response to hormonal stimulation with β-casein
concentrations being 43% higher in goat milk relative to bovine
milk (13,14). It is thought to be a binding site
for STAT5a in the -300 bp region of the goat β-casein promoter, and
this binding site serves to mediate responses to lactogenic hormone
stimulation (15). This lays the
theoretical foundation for the selection of a goat β-casein
promoter, thus allowing for the efficient expression of proteins in
mammary glands.
Although the goat β-casein promoter has previously
been widely used to drive the transcription of many recombinant
proteins in transgenic goats, the expression of these proteins has
not been sufficiently high for commercial applications. A variety
of approaches have been employed in an effort to boost mammary
expression of these recombinant proteins, including the use of
distal regulatory elements/large genomic DNA fragments (16), insulators (17), matrix-attached regions (18), and targeted site integrations
(19). The cytomegalovirus (CMV)
promoter is a high-efficiency promoter/enhancer widely used for
transgene expression in cells. Zarrin et al found this
promoter to be more efficient than alternatives such as the SV40,
Rous sarcoma virus (RSV), and Vλl promoters for certain B-cell
lines (20). There are few
reports, however, regarding the use of a goat β-casein/CMV chimeric
promoter to facilitate protein production in the mammary glands of
medium and large transgenic animals.
The properties of a given protein determine its
protein purification strategy. LF is a cationic protein, thus
making it well suited to purification via cation-exchange
chromatography (21,22). This approach is widely used for
bovine LF purification by bLF-producing companies. Concanavalin A
affinity chromatography or metal ion affinity chromatography are
also viable strategies for purifying LF owing to its glycosylation
and Fe3+-binding activity (23,24).
In this study, we generated a transgenic goat
harboring the human lactoferrin transgene driven by a chimeric goat
β-casein/CMV promoter. This animal was generated using goat fetal
fibroblasts microinjected with the pCL25-rhLF-neo vector as SCNT
donor cells, allowing for mammary gland-specific transgene
expression (25,26), while retaining the biological
characteristics necessary for better efficacy as a drug or food
additive. We additionally conducted ELISAs, western blot, and
antibacterial activity assays to confirm that human lactoferrin was
efficiently expressed in transgenic goat milk while retaining its
normal biological activity.
Materials and methods
Ethics statement
Animal experiments and procedures were performed in
accordance with the guide for the Care and Use of Laboratory
Animals (Ministry of Science and Technology of the People's
Republic of China) and approved by the animal care and use
committee of Yangzhou University, Yangzhou, China [license no.
SYXK(Su)2017-0044]. A total of 50 female dairy goats (45-60 kg,
13-18 months old; Jiangsu Academy of Agricultural Sciences,
Nanjing, China) used in the current study were raised at room
temperature (25±2°C), with a 12 h day/night cycle, and allowed free
access to food and water. All animals were anesthetized using
xylazine hydrochloride injection (0.001-0.002 ml/kg) purchased from
huamu Animal health Products Co., Ltd. during surgery, with all
possible effort being made to reduce their pain, distress, and
suffering.
Lactoferrin expression vector
construction
Human lactoferrin (GenBank: KT006756.1) cDNA was
synthesized by Genscript (China), using cDNA containing 5′ and 3′
terminal XhoI sites. The sequence encoding the mature
lactoferrin peptide was fused to both the goat β-lactoglobulin
signal peptide as well as the Kozak translation initiation
sequence.
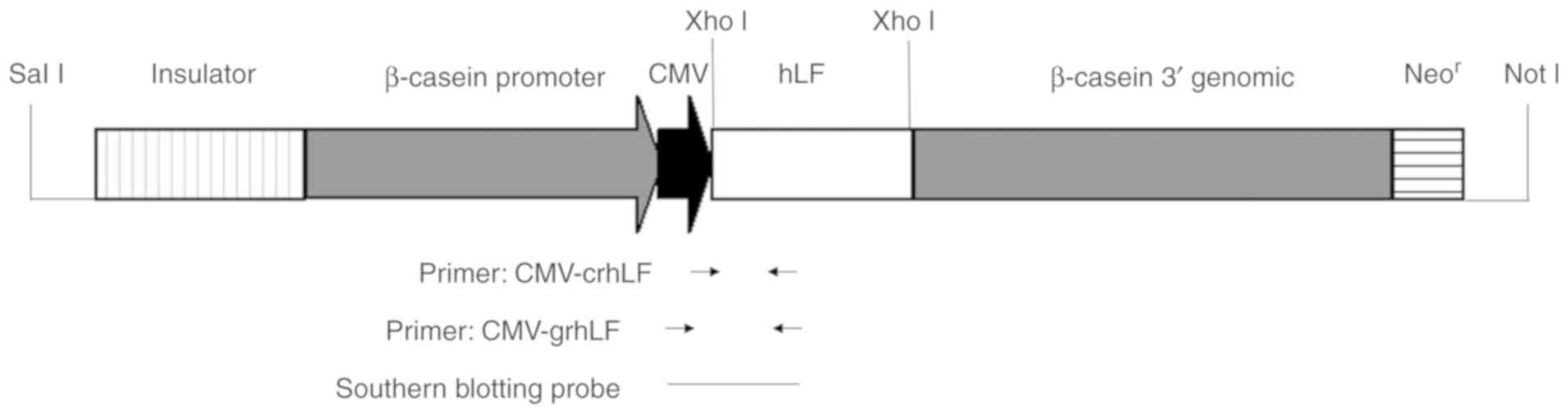
The synthesized lactoferrin gene was cloned into the
pCL25 vector (generated internally) containing the goat
β-casein/CMV chimeric promoter and a neo-selectable cassette in the
goat β-casein 3′ genomic region near the vector NotI site
(Fig. 1). NotI and
SalI were used for vector digestion, and a QIAquick Gel
Extraction kit (28704, Qiagen, Germany) was used to purify the
resultant fragments.
Cell culture and transgene
expression
A 30-day old fetus was surgically removed from a
Sannen dairy goat and used to generate fibroblasts. Briefly, fetal
tissue was cut into small fragments following the removal of the
internal organs, head, and limbs, and these fragments underwent
0.05% trypsin-EDTA-mediated digestion. Fibroblasts were then
isolated from the supernatant portion of this digestion and grown
using DMEM/F12 (Sh30023.01; hyclone) containing 10% FBS
(Sh30406.01; hyclone), and 1% penicillin-streptomycin (SV30010;
hyclone) at 37°C in a 5% Co2 humidified incubator. Cells
underwent passaging at 80% confluency, and after the second passage
cells were aliquoted and frozen in freezing media containing 10%
DMSo (D2650; Sigma) and 20% FBS.
Aliquots of cells were frozen, and once they grew to
80% confluency they were microinjected with 5 ng/µl of the
purified pCL25-rhLF-Neo DNA fragment using an Eppendorf InjectMan
(NI2; Eppendorf) and then cultured as above. After 24 h, the cells
were grown in selective medium containing 800 ng/µl G418
(SV3006801; hyclone) for approximately 10 days. A cloning ring was
used to isolate and expand healthy colonies following selection,
and these clones were subcultured as above. Some of these
subcultures were frozen for long-term storage, while the rest were
screened for expression of the transgene via polymerase chain
reaction (PCR).
Generation of a transgenic goat via
SCNT
Somatic cell nuclear transfer (SCNT) was conducted
after identifying transgene-positive clones. Enucleated oocytes
served as recipients for transgenic cell nuclei, with a super
electro cell fusion generator (EGFE21; nepa Gene) being used for
the SCnT procedure. next, 5 µmol/l ionomycin (I0634; Sigma)
and 7.5 µg/ml cytochalasin B (C6762; Sigma) in M16 medium
(M7292; Sigma) was used to activate these reconstructed embryos for
5 min, and then cells were treated with M16 containing 2 mmol/l
6-dimethylaminopurine (D2629; Sigma) and 7.5 µg/ml
cytochalasin B for 5 h. After activation, the embryos were
implanted into recipient goats, and after a 1-month period these
animals were assessed via ultrasound to confirm pregnancy.
Approximately 150 days later, kids were delivered naturally. For
all kids, a small portion of the ear was taken as a biopsy sample
from which DNA was isolated and used to assess transgene
incorporation by PCR and southern blotting. DL2000 DNA marker
(3427A; Takara) was purchased from Takara Biotech (Dalian) Co.,
Ltd.
Confirmation of transgene integration in
cloned goats
Genomic DNA from the transgenic donor cells and ear
tissues of cloned goats was prepared with an Easy Pure Genomic DNA
kit (EE101-1; TransGen). A pair of primers specific for human LF
was used to determine which donor cells had incorporated the
transgene. Primer sequences used were: CMV-crhLF-1: ATG GGC GTG GAT
AGC GGT TTG AC and CMV-crhLF-2: CCA CCA TCA AGG GTC ACA GCA TCG. To
identify transgenic goats, the following primers were instead used:
CMV-grhLF-1: ATA GTA ACG CCA ATA GGG A and CMV-grhLF-2: GGT CG CA
GTT TGT AGG G. The following conditions were used for all PCR
reactions: 94°C for 5 min, 33 cycles at 94°C for 1 min, 56°C for 1
min, then 72°C for 38 sec, and finally held at 72°C for 10 min.
Product sizes for the two primer pairs were 450 and 775 bp,
respectively. Sequencing analysis was performed by Sangon Biotech
(Shanghai) Co., Ltd.
Southern blotting was next employed to confirm
specific transgene DNA integration in goats. Ear biopsy-derived DNA
from transgenic and wild-type (WT) goats underwent overnight
BamHI digestion, with the PCL25-CMV-rhLF-neo plasmid serving
as a positive control. A digoxigenin-labeled probe then underwent
PCR amplification with the CMV-grhLF-1 and CMV-ghLF-2 primer pair.
Samples underwent 4-h agarose gel electrophoresis, after which DNA
was transferred to a nylon membrane (11417240001; Roche) for
blotting. This membrane next underwent probe hybridization for 18
h, followed by incubation with biotin-labeled mouse anti-Digoxin
for 30 min. A positive band was expected to be approximately 9.1 kb
in size. Southern blotting reagents were purchased from Boster Co.
(Wuhan).
ELISAs
Milk samples collected from lactating transgenic and
WT goats were centrifuged at 10,000 × g for 30 min at 4°C for whey
isolation. The samples were diluted at 1:10 with PBS, and were used
for ELISA reactions with a rabbit-anti-lactoferrin polyclonal
primary antibody (dilution 1:2,000, 4% FBS/PBS; Sangon;
D121815-0025). After incubation at 37°C for 1 h and being washed
three times with PBS-T (PBS containing 0.05% Tween-20), wells were
probed with an hRP-conjugated goat-anti-rabbit secondary antibody
(dilution 1:1,000, 4% FBS/PBS; Sangon) at 37°C for 1 h. The samples
then underwent a colorimetric reaction via adding TMB substrate to
each well, after which absorbance at 450 nm was measured via
microplate reader (Rayto). Protein standards (SRP6519; Sigma) were
used for standard curve generation, and sample rhLF concentrations
were determined based on this standard curve.
Purification of rhLF from the transgenic
cloned goat
Fat and other undissolved substances were removed
from milk via centrifugation at 12,000 × g for 30 min at 4°C, after
which the pH was reduced to 4.0 in order to facilitate casein
precipitation. Milk was then centrifuged at 4°C at 100,000 × g for
1 h. Supernatant pH was adjusted to 6.0 using acetic acid, after
which the samples were centrifuged again as in the previous step. A
protein purification system (ÄKTAprimeTM PLuS; GE healthcare) was
used for all purification reactions. First, after equilibration in
a column containing Buffer A (0.07 mol/l HAc, pH 3.1), samples were
loaded onto a HiTrap Capto S cation exchange column (1 ml; GE
healthcare) and the bound proteins were eluted via a step gradient
of 30 and 100% Buffer B (0.5 mol/l NaCl, 0.07 mol/l Tris-HAc, pH
7.5). The eluate containing 100% Buffer B was collected and
desalted via a Bestdex G-25 column (1.6×2.5 cm; BestChrom) for use
in downstream experiments. SDS-PAGE analysis was then used to
assess protein purity.
Western blotting
Whey was isolated as above and then boiled in SDS
loading buffer for 10 min, after which samples were
electrophoretically separated using 12% polyacrylamide Tris-glycine
gels. Afterwards, these gels were stained using Coomassie Brilliant
Blue G-250, and the sample purity and concentrations were
determined using Tanon Gis software (Bio-Tanon). For western
blotting, separated proteins were then transferred onto PVDF
membranes (F019531; Sangon). The membranes were blocked using 5%
BSA/TBST overnight at 4°C, and then probed using a polyclonal
rabbit-anti-LF antibody (1:2,000, 10% FBS/TBST; Sangon) at 37°C for
1.5 h. next, an hRP-conjugated secondary goat-anti-rabbit IgG
(1:1,000, 10% FBS/TBST; Sangon) was used to probe blots at 37°C for
1 h. The blots were then washed three times in TBST (20 mM
Tris-base, 137 mM naCl, 0.05% Tween-20), and protein bands were
detected with an ECL substrate solution (Millipore Corporation)
based on provided directions.
Bacteriostatic activity assessment
Lactoferrin has been shown to be able to inhibit the
growth of both Gram-positive and -negative bacteria, including
important pathogenic species such as Helicobacter pylori,
Staphylococcus aureus, Shigella flexneri,
enteropathogenic Escherichia coli (EPEC) and Salmonella
enterica serovar typhimurium (2,27-31). We therefore selected E.
coli K88 grown on LB plates as a model strain to test the
bacteriostatic activity of transgenic goat milk.
A single E. coli K88 colony was transferred
into 15 ml LB culture medium, shaken and kept overnight at 37°C.
The resultant bacteria were then streaked evenly across an LB agar
plate using a cotton swab.
To test bacteriostatic activity of the milk, sterile
filter papers (8 mm in diameter) were placed onto the plate
surface, and 100 µl sample aliquots were added on top of
this paper. After a 4-h incubation at 37°C, the size of the growth
inhibition area surrounding a given sample was used to assess
bacteriostatic activity.
Results
Lactoferrin expression vector
construction
We successfully inserted the LF cDNA fragment into
the pCL25 vector, thereby producing a pCL25-rhLF-Neo recombinant
vector that was found to be of appropriate size based on
restriction enzyme digestion and sequencing. Sequencing confirmed
that the rhLF coding region was fused in-frame upstream of
pCL25.
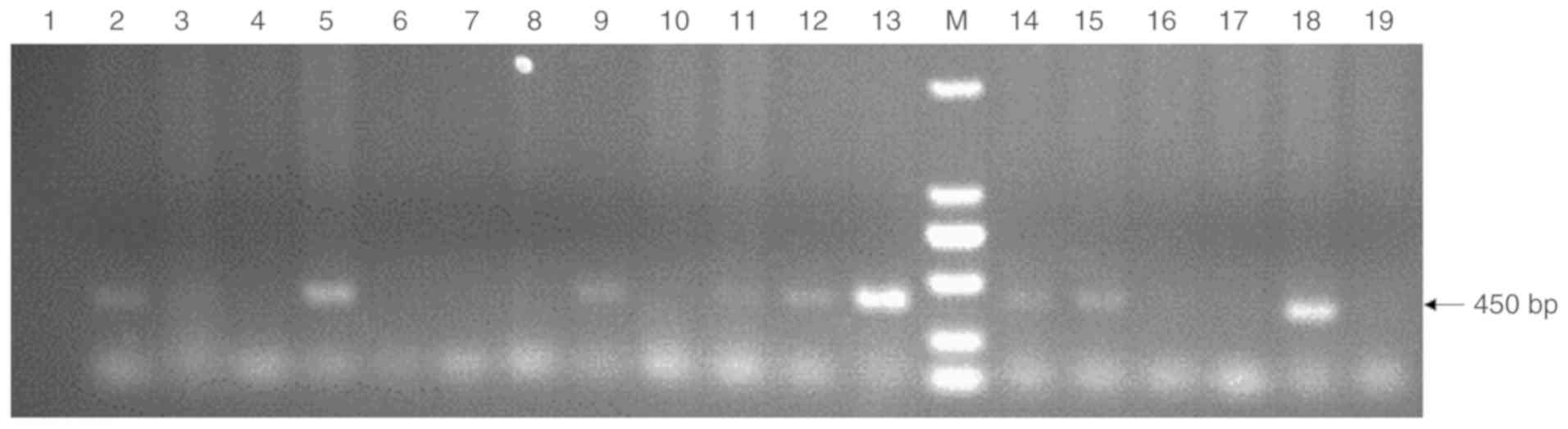
Fetal goat fibroblast transfection
Goat fetal fibroblasts were microinjected with 5
ng/µl of the purified pCL25-rhLF-neo DNA fragment, and were
selected using G418. A total of 16 G418-resistant transgenic cells
were obtained by single cell amplification. of these, 9 were
determined to express the hLF transgene via PCR using the
CMV-crhLF-1 and CMV-crhLF-2 primers (Fig. 2). In total, 56.25% (9/16) of the
cell clones had confirmed pCL25 integration. Clone no. 4 cells
served as SCNT donors as they were found to exhibit the best
viability and quality.
SCNT-mediated transgenic cloned goat
generation
SCNT was used to produce transgenic goats as
previously identified (27). We
transferred 65 reconstructed embryos into 5 recipient goats,
leading to the birth of a single female kid that was found by PCR
to harbor the pCL25-rhLF-neo transgene (Table I). The female kid was designated
as LF-1 (Fig. 3), and upon
reaching sexual maturity underwent mating with a WT ram.
 | Table INuclear transfer outcomes. |
Table I
Nuclear transfer outcomes.
| Cell lines | Oocytes | Reconstructed
embryos | Transferred
embryos | Recipients | Pregnancies at day
30 | No. of
newborns |
|---|
| No. 4 | 95 | 70 | 65 | 5 | 1 | 1 |
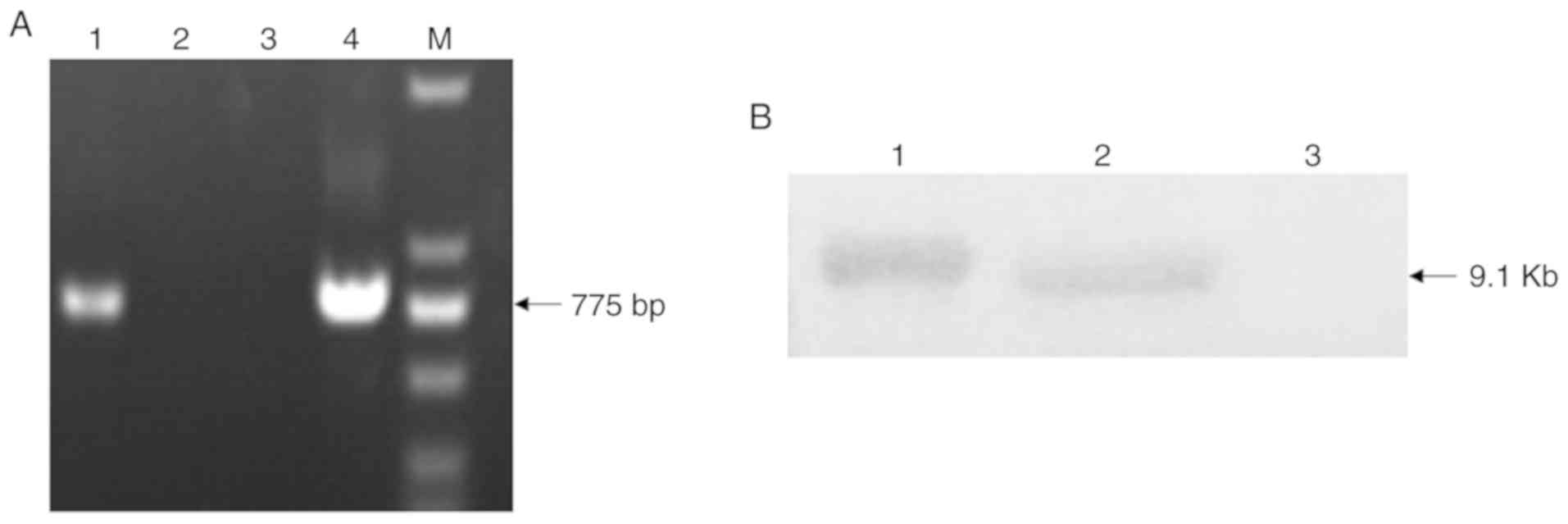
Confirmation of transgene integration in
cloned goats
PCR and Southern blotting were used to confirm that
the transgenic goat integrated the rhLF transgene. human
LF-specific primers (CMV-grhLF-1 and CMV-grhLF-2) were used to
identify the cloned goats by PCR, while digoxigenin-labeled
versions of these primers were used as probes for Southern
blotting.
Following PCR, we were able to amplify a 775 bp
product, confirming successful rhLF transgene integration into this
cloned goat (Fig. 4A). Southern
blotting further confirmed this finding (Fig. 4B).
Assessment of milk rhLF expression in
transgenic goats
Expression of rhLF in WT and transgenic goat milk
samples was next assessed via ELISA. Milk was collected during days
1-30 of lactation following delivery. We found that the rhLF
concentration reached a peak of 4.7 mg/ml on day 4, with an average
concentration of 3.89±0.82 mg/ml from days 1-30 of lactation.
To assess the possible ectopic expression of rhLF in
this transgenic goat, rhLF levels in the serum and saliva of
lactating goat were measured via ELISA. There was no indication of
rhLF expression in the serum or saliva of this transgenic goat
(data not shown).
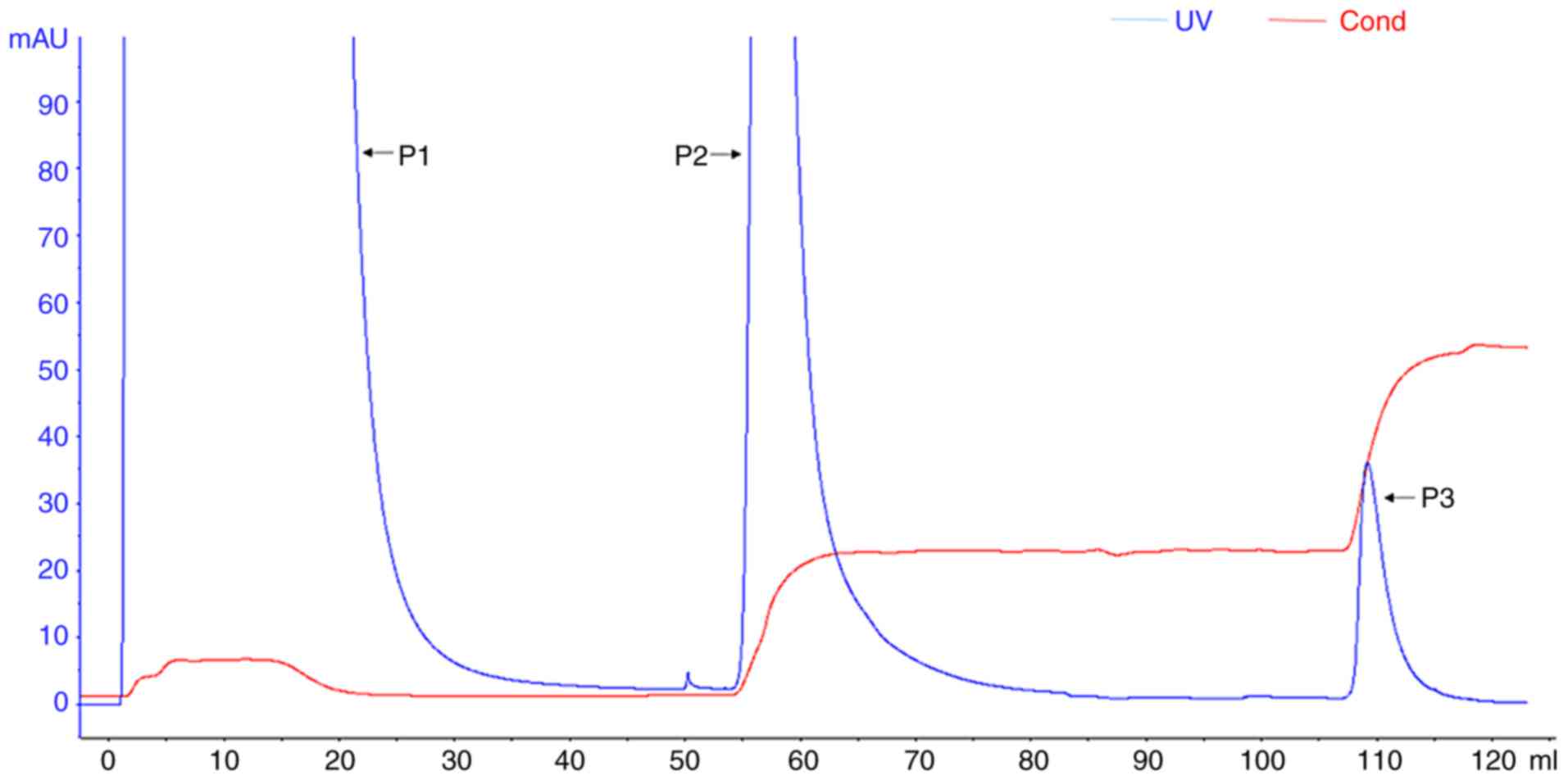
Purification of rhLF from the transgenic
cloned goat
Cation exchange chromatography can be used to
separate lactoferrin from milk, as lactoferrin has a net-positive
charge. In order to explore the optimal elution conditions for
separation and purification of rhLF via cation exchange
chromatography, we first assessed the optimal solution conductivity
for rhLF. Stepwise elution was used to achieve one-step elution and
separation of the target protein.
Two elution peaks were obtained from the HiTrap
Capto S cation exchange column eluted with a step gradient of 30
and 100% Buffer B. SDS-PAGE and western blotting revealed that
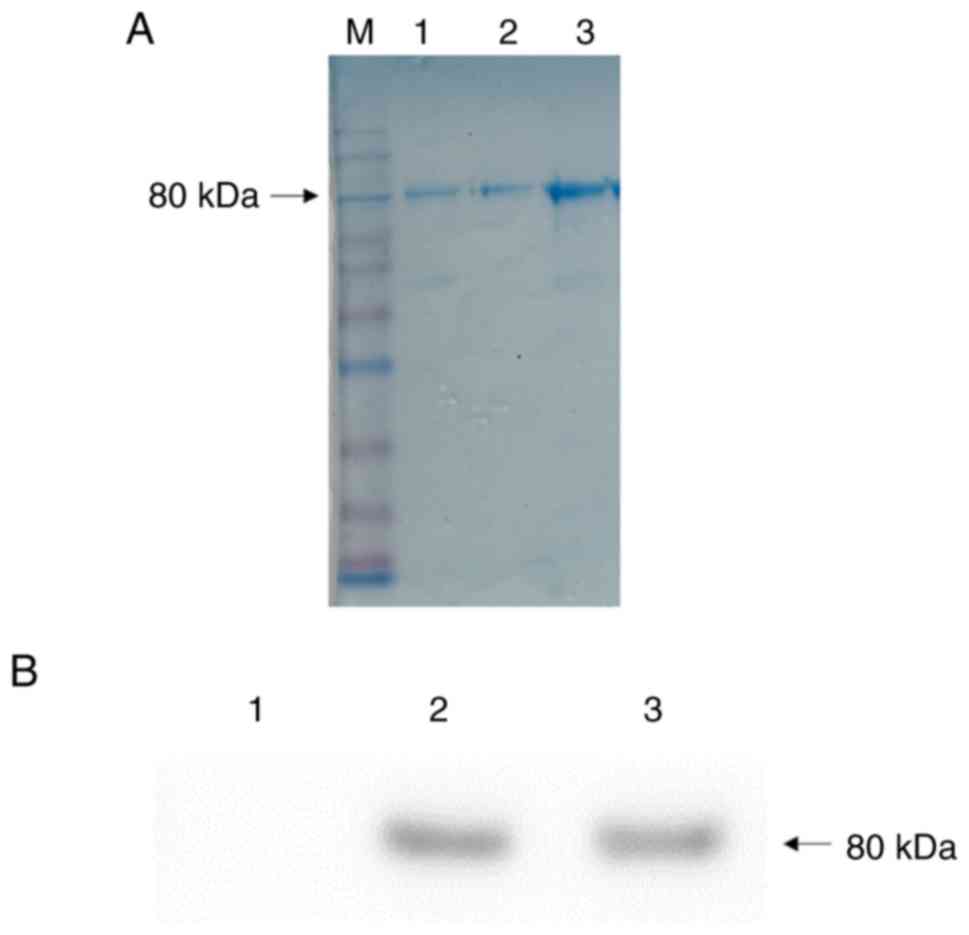
high-purity rhLF was successfully collected in peak P3 of the
eluent (Fig. 5), with a size of
80 kDa. The concentration of the purified rhLF was found to be 1.25
mg/ml by spectrophotometry (One Drop 1000+; OneDrop Technologies,
Inc.). The purity was determined to be 95.8% based on densitometric
scanning of the SDS-PAGE gel.
Western blotting confirmed that samples in the
transgenic goat were identical to native hLF control samples, with
a size of approximately 80 kDa (Fig.
6). Bands were absent in the WT control goat sample, as
expected.
Assessment of transgenic goat milk
bacteriostatic activity
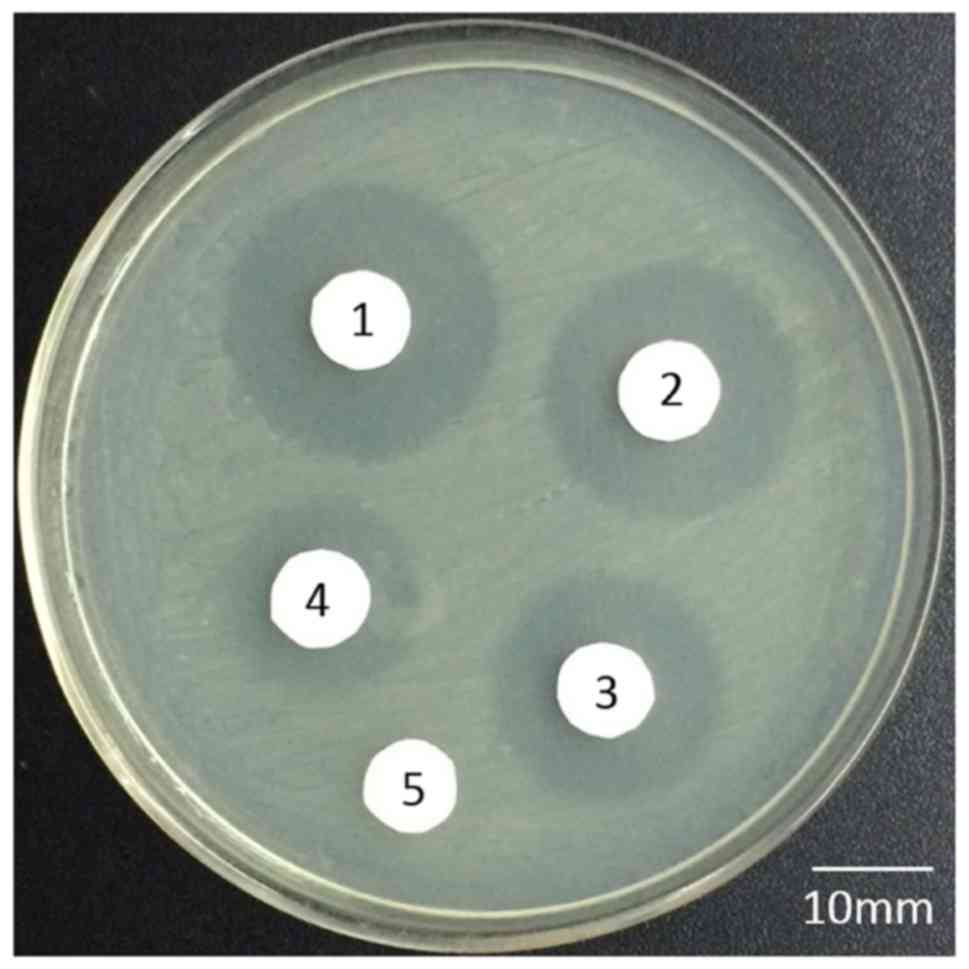
The bacteriostatic activity of the rhLF in the
transgenic goat milk was assessed via an agar disc diffusion method
in order to allow for observation of bacteriostatic activity in
vitro. Sterile filter paper was placed onto agar plates
containing E. coli K88, and bacteriostatic activity was
estimated based on inhibition zone sizes surrounding the sterile
filter papers following a 4 h incubation at 37°C. These results
revealed that rhLF from transgenic milk exhibited comparable
bacteriostatic activity to that of hLF (inhibition zone diameters
of 17 and 19 mm, respectively). WT goat milk served as a negative
control, with no inhibition zone being evident. We also found that
rhLF purified by cation-exchange chromatography exhibited similar
bacteriostatic activity (an inhibition zone diameter of 13 mm)
(Fig. 7).
Discussion
In this study, we successfully used SCNT as a means
of generating a transgenic goat producing rhLF in mammary cells,
using transgenic goat fetal fibroblast cells as donor cells. To
date, there have been no previous reports of using fetal
fibroblasts microinjected with rhLF gene as donor cells for SCNT.
We detected no abnormalities in the founder transgenic goat or its
offspring, indicating no effect of the vector on goat biology. To
determine whether the rhLF transgene could be stably transmitted to
offspring, the female founder transgenic goat was mated with a
wild-type ram and a single male lamb was birthed. A subsequent PCR
assay demonstrated that it was transgenic for rhLF (data not
shown), indicating that the rhLF transgene can be inherited by
offspring.
There are many reports regarding high-level rhLF
expression in transgenic mice, rabbits, and cows (32-34). Mice and rabbits, however, are not
suitable for large-scale commercial rhLF production due to their
limited milk production and short life span (2-3 years for mice,
8-10 years for rabbits). Cows also are not appropriate models for
producing rhLF because bovine milk has more allergenic protein than
does goat milk (35,36). Therefore, goats are more suitable
as a biologic mammary reactor for the large-scale production of
rhLF given that goat milk has been reported to contain smaller fat
globules and a distinct casein composition relative to bovine milk,
making it less allergenic (37,38). At present, many transgenic animals
are produced via SCNT or pronucleus microinjection, including sheep
(39), goats (40,41), cows (42), mice (43) and rabbits (44). The success rate of SCNT remains
low and varies based upon factors such as the vector used, the
source of recipient and donor cells, the exact SCnT protocol
employed, and the influence of exogenous genes on embryonic
development (45-47). The quality of donor cells is
critical for producing transgenic animals via SCnT. Preparation of
transgenic animals using electroporation-mediated transfection
requires optimization of transfection conditions and is often
associated with a high rate of cell death. however, cell
microinjection avoids these challenges, instead offering a high
integration rate while remaining suitable for genetic engineering
and the establishment of transgenic animals. In this study, we
improved upon the process of preparing transgenic goats using goat
fetal fibroblasts microinjected with the rhLF gene as donor cells
for SCNT.
In previous reports, we constructed various mammary
gland-specific vectors containing a CMV enhancer and a chimeric
promoter [goat β-casein, bovine αs1-casein, and goat
β-lactoglobulin (BLG)] based on milk protein promoter sequences.
These vectors allowed for 1.17-8.10 mg/ml hLF levels in transgenic
murine milk-roughly 100,000-fold higher than the levels produced
from control promoters (7-40 ng/ml). We also found that the
inclusion of the CMV enhancer significantly increased hLF
expression in these mice. Use of hLF cDNA did not achieve
expression levels as high as those from hLF genomic DNA in these
mice (25). Many factors can
influence recombinant milk protein expression levels, including
copy number, site of chromosomal insertion, and species-specific
differences in expression patterns (48,49). rhLF expression levels in
transgenic goats can be as high as 4.7 mg/ml-levels which are
markedly higher than the levels observed in transgenic goats
without a CMV enhancer (50).
There were no indications of rhLF expression in the serum or saliva
of the transgenic goat, as the goat β-casein promoter is
specifically expressed only in lactating mammary tissue and not at
other ectopic sites. This means that there is no potential risk of
transgenic animals expressing heterologous proteins in the mammary
gland when using this goat β-casein/CMV chimeric promoter. using
western blotting we further confirmed that the size of the rhLF
expressed in transgenic goats was roughly 80 kDa, which is
comparable to the size of hLF.
The secretion of lactoferrin in milk is directly
related to the nutritional status and environmental conditions of
the mother, and as such the secretion of lactoferrin in milk can be
improved by improving maternal housing conditions and other
factors. however, for transgenic animals, in addition to these
growth conditions and environmental factors, improving the
inheritance and stability of foreign genes remains a major
challenge. In this study, our aim was to produce transgenic-cloned
goats as living mammary bioreactors that exhibited a high-level of
rhLF expression in their milk. An optimized construct is essential
in order to achieve a high level expression of recombinant
proteins. We used ELISA to confirm the expression of rhLF in
transgenic goat milk on days 1-30 of lactation following delivery,
revealing that rhLF was continuously expressed in goat milk during
this 30-day period. There were no clear decreases in rhLF
expression during the lactation period. These results thus clearly
show that a transgenic goat carrying the pCL25-rhLF-Neo mammary
gland-specific expression vector encoding goat β-casein/CMV
chimeric promoter can express rhLF stably in the mammary gland.
At present, phosphoric buffer (PB) is widely used as
an eluent when extracting lactoferrin via cation-exchange
chromatography (23). however, PB
easily associates with common Ca2+ ions, Mg2+
ions, and heavy metal ions to form precipitates, and it can also
inhibit certain biochemical processes as well as the activity of
most enzymes. PB is thus not an ideal eluent choice when purifying
lactoferrin by cation-exchange chromatography. In order to achieve
superior purified rhLF activity, we therefore used a commercially
available HiTrap Capto S cation exchange column for its effective
purification from the milk of a transgenic goat, using the Tris-HAc
buffer as an eluent. Similarly to the rhLF purified in other
previous reports (24,51), rhLF purification efficiency in
transgenic goats was high (≥95.8%). When we assessed the
bacteriostatic activity of this rhLF, we found it to be comparable
to that of natural hLF, penicillin, and streptomycin, which
suggests that rhLF may be an effective antibiotic for future
use.
In conclusion, we have successfully used SCNT to
produce a transgenic goat, with goat fetal fibroblast cells serving
as donor cells microinjected with the expression vector pCL25. Our
results conclusively demonstrate that the pCL25 vector, which
contains goat β-casein/CMV chimeric promoter, can drive transgenic
goats to stably express a biologically active form of rhLF. This
study offers an initial strategy for rhLF production for
incorporation into drugs or food products, thereby facilitating
future studies of this protein.
Acknowledgments
We thank all lab members for their help in this
study.
Funding
This study was supported by the National Key
Research and Development Program of China, China (no.
2016YFE0126000), the Priority Academic Program Development of
Jiangsu higher Education Institutions, China (PAPD) and the
Yangzhou university-enterprise Cooperation Project, China
(YZ2017283).
Availability of data and materials
All data and materials are available from the
authors.
Authors' contributions
TZ, YY, RL, SX, MZ, TY, YL, and KY performed the
studies and analyzed the data. TZ and YC wrote and revised the
manuscript. YC conceived and guided the study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The Guide for the Care and use of Laboratory Animals
(Ministry of Science and Technology of the People's Republic of
China) was followed for all surgical studies described herein,
which received prior approval from the Yangzhou University Animal
Care and Use Committee (license no. SYXK(Su)2017-0044). All animals
were anesthetized during surgery, with all possible effort being
made to reduce the pain, distress, and suffering of the study
animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
García-Montoya IA, Cendón TS,
Arévalo-Gallegos S and Rascón-Cruz Q: Lactoferrin a multiple
bioactive protein: An overview. Biochim Biophys Acta. 1820:226–236.
2012. View Article : Google Scholar
|
|
2
|
Wu J, Hu Y, Du C, Piao J, Yang L and Yang
X: The effect of recombinant human lactoferrin from the milk of
transgenic cows on Salmonella enterica serovar typhimurium
infection in mice. Food Funct. 7:308–314. 2016. View Article : Google Scholar
|
|
3
|
Fliedl L, Grillari J and Grillari-Voglauer
R: human cell lines for the production of recombinant proteins: On
the horizon. New Biotechnol. 32:673–679. 2015. View Article : Google Scholar
|
|
4
|
Fu Z, Baker D, Cheng A, Leighton J,
Appelbaum E and Aon J: Characterization of a saccharomyces
cerevisiae fermentation process for production of a therapeutic
recombinant protein using a multivariate bayesian approach.
Biotechnol Prog. 32:799–812. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gupta SK and Shukla P: Advanced
technologies for improved expression of recombinant proteins in
bacteria: Perspectives and applications. Crit Rev Biotechnol.
36:1089–1098. 2016. View Article : Google Scholar
|
|
6
|
Kim H, Yoo SJ and Kang HA: Yeast synthetic
biology for the production of recombinant therapeutic proteins.
FEMS Yeast Res. 15:1–16. 2015.
|
|
7
|
Song S, Ge X and Cheng Y, Lu R, Zhang T,
Yu B, Ji X, Qi Z, Rong Y, Yuan Y and Cheng Y: high-level expression
of a novel recombinant human plasminogen activator (rhPA) in the
milk of transgenic rabbits and its thrombolytic bioactivity in
vitro. Mol Biol Rep. 43:775–783. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lavine G: FDA approves first biological
product derived from transgenic animal. Am J Health Syst Pharm.
66:5182009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Davis B and Bernstein JA: Conestat alfa
for the treatment of angioedema attacks. Ther Clin Risk Manag.
7:265–273. 2011.PubMed/NCBI
|
|
10
|
The farmyard drug store: Nature.
443:16–17. 2006. View
Article : Google Scholar
|
|
11
|
Bösze Z, Baranyi M and Whitelaw CB:
Producing recombinant human milk proteins in the milk of livestock
species. Adv Exp Med Biol. 606:357–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pantano T, Rival-Gervier S, Prince S,
Menck-Le Bourhis C, Maeder C, Viglietta C, Houdebine LM and Jolivet
G: In vitro and in vivo effects of a multimerized alphas 1-casein
enhancer on whey acidic protein gene promoter activity. Mol Reprod
Dev. 65:262–268. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huma N, Ghaffar F, Rafiq S, Pasha I,
Sameen A, Hayat I and Hussain I: Characterization of milk proteins
from different animal species through Gel electrophoresis. Pak J
Zool. 50:1983–1986. 2018. View Article : Google Scholar
|
|
14
|
Dayal R, Hurlimann J, Suard YM and
Kraehenbuhl JP: Chemical and immunochemical characterization of
caseins and the major whey proteins of rabbit milk. Biochem J.
201:71–79. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kung MH, Lee YJ, Hsu JT, Huang MC and JU
YT: A functional study of proximal goat β-casein promoter and
intron 1 in immortalized goat mammary epithelial cells. J Dairy
Sci. 98:3859–3875. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Giraldo P and Montoliu L: Size matters:
Use of YACs, BACs and PACs in transgenic animals. Transgenic Res.
10:83–103. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujiwara Y, Miwa M, Takahashi R, Kodaira
K, Hirabayashi M, Suzuki T and Ueda M: high-level expressing YAC
vector for transgenic animal bioreactors. Mol Reprod Dev.
52:414–420. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sharp JA, Cane Kn, Mailer SL, Oosthuizen
WH, Arnould JP and Nicholas KR: Species-specific cell-matrix
interactions are essential for differentiation of alveoli like
structures and milk gene expression in primary mammary cells of the
Cape fur seal (Arctocephalus pusillus pusillus). Matrix Biol.
25:430–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
McKnight RA, Shamay A, Sankaran L, Wall RJ
and Hennighausen L: Matrix-attachment regions can impart position-
independent regulation of a tissue-specific gene in transgenic
mice. Proc Natl Acad Sci USA. 89:6943–6947. 1992. View Article : Google Scholar
|
|
20
|
Zarrin AA, Malkin L, Fong I, Luk KD, Ghose
A and Berinstein NL: Comparison of CMV, RSV, SV40 viral and
Vlambda1 cellular promoters in B and T lymphoid and non-lymphoid
cell lines. Biochim Biophys Acta. 1446:135–139. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Law BA and Reiter B: The isolation and
bacteriostatic properties of lactoferrin from bovine milk whey. J
Dairy Res. 44:595–599. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yoshida S, Wei Z, Shinmura Y and Fukunaga
N: Separation of lactoferrin-a and -b from bovine colostrum. J
Dairy Sci. 83:2211–2215. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nuijens JH, van Berkel PH and Schanbacher
FL: Structure and biological actions of lactoferrin. J Mammary
Gland Biol neoplasia. 1:285–295. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Aguila A, Herrera AG, Toledo W, Rodríguez
AA, Garcia HM, Hernández J, Cádiz A, Sierra G and Brock JH:
Isolation and structure-functional characterization of human
colostral lacto-ferrin. Biotecnol Appl. 17:177–182. 2000.
|
|
25
|
Cheng Y, An LY, Yuan YG, Wang Y, Du FL, Yu
BL, Zhang ZH, Huang YZ and Yang TJ: hybrid expression cassettes
consisting of a milk protein promoter and a cytomegalovirus
enhancer significantly increase mammary-specific expression of
human lactoferrin in transgenic mice. Mol Reprod Dev. 79:573–585.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan YG, An L, Yu B, Song S, Zhou F, Zhang
L, Gu Y, Yu M and Cheng Y: Expression of recombinant human
alpha-lactalbumin in the milk of transgenic goats using a hybrid
pomoter/enhancer. J Anal Methods Chem. 2014:2810312014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gomez HF, Ochoa TJ, Herrera-Insua I,
Carlin LG and Cleary TG: Lactoferrin protects rabbits from Shigella
flexneri-induced inflammatory enteritis. Infect Immun.
70:7050–7053. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ochoa TJ, Brown EL, Guion CE, Chen JZ,
McMahon RJ and Cleary TG: Effect of lactoferrin on
enteroaggregative E. coli (EAEC). Biochem Cell Biol. 84:369–376.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ochoa TJ, Noguera-Obenza M, Ebel F, Guzman
CA, Gomez HF and Cleary TG: Lactoferrin impairs type III secretory
system function in enteropathogenic Escherichia coli. Infect Immun.
71:5149–5155. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Farnaud S and Evans RW: Lactoferrin-a
multifunctional protein with antimicrobial properties. Mol Immunol.
40:395–405. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mosquito S, Ochoa TJ, Cok J and Cleary TG:
Effect of bovine lactoferrin in Salmonella ser. Typhimurium
infection in mice. Biometals. 23:515–521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nuijens JH, van Berkel PH, Geerts ME,
Hartevelt PP, de Boer HA, van Veen HA and Pieper FR:
Characterization of recombinant human lactoferrin secreted in milk
of transgenic mice. J Biol Chem. 272:8802–8807. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li L, Shen W, Min L, Dong H, Sun Y and Pan
Q: human lactoferrin transgenic rabbits produced efficiently using
dimethylsulfoxide- sperm-mediated gene transfer. Reprod Fertil Dev.
18:689–695. 2006. View Article : Google Scholar
|
|
34
|
van Berkel PH, Welling MM, Geerts M, van
Veen HA, Ravensbergen B, Salaheddine M, Pauwels EK, Pieper F,
Nuijens JH and Nibbering PH: Large scale production of recombinant
human lactoferrin in the milk of transgenic cows. Nat Biotechnol.
20:484–487. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pastuszka R, Barłowska J and Litwińczuk Z:
Allergenicity of milk of different animal species in relation to
human milk. Postepy Hig Med Dosw (online). 70:1451–1459. 2016.
View Article : Google Scholar
|
|
36
|
Høst A, Husby S, Gjesing B, Larsen JN and
Løwenstein H: Prospective estimation of IgG, IgG subclass and IgE
antibodies to dietary proteins in infants with cow milk allergy.
Levels of antibodies to whole milk protein, BLG and ovalbumin in
relation to repeated milk challenge and clinical course of cow milk
allergy. Allergy. 47:218–229. 1992. View Article : Google Scholar
|
|
37
|
Pisanu S, Marogna G, Pagnozzi D, Piccinini
M, Leo G, Tanca A, Roggio AM, Roggio T, Uzzau S and Addis MF:
Characterization of size and composition of milk fat globules from
Sarda and Saanen dairy goats. Small Ruminant Res. 109:141–151.
2013. View Article : Google Scholar
|
|
38
|
Juvarajah T, Wan-Ibrahim WI, Ashrafzadeh
A, Othman S, Hashim OH, Fung SY and Abdul-Rahman PS: Human milk fat
globule membrane contains hundreds of abundantly expressed and
nutritionally beneficial proteins that are generally lacking in
caprine milk. Breastfeed Med. 13:631–637. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schramm FR: The Dolly case, the Polly
drug, and the morality of human cloning. Cad Saude Publica.
15(Suppl 1): 51–64. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
An LY, Yuan YG, Yu BL, Yang TJ and Cheng
Y: Generation of human lactoferrin transgenic cloned goats using
donor cells with dual markers and a modified selection procedure.
Theriogenology. 78:1303–1311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ohkoshi K, Takahashi S, Koyama S, Akagi S,
Adachi N, Furusawa T, Fujimoto J, Takeda K, Kubo M, Izaike Y and
Tokunaga T: In vitro oocyte culture and somatic cell nuclear
transfer used to produce a live-born cloned goat. Cloning Stem
Cells. 5:109–115. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang M, Sun Z, Yu T, Ding F, Li L, Wang X,
Fu M, Wang H, Huang J, Li N and Dai Y: Large-scale production of
recombinant human lactoferrin from high-expression, marker-free
transgenic cloned cows. Sci Rep. 7:107332017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gordon JW: Studies of foreign genes
transmitted through the germ lines of transgenic mice. J Exp Zool.
228:313–324. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hu J, Peng X, Schell TD, Budgeon LR,
Cladel NM and Christensen ND: An hLA-A2.1-transgenic rabbit model
to study immunity to papillomavirus infection. J Immunol.
177:8037–8045. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yanagimachi R: Cloning: Experience from
the mouse and other animals. Mol Cell Endocrinol. 187:241–248.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Oback B and Wells D: Donor cells for
nuclear cloning: Many are called, but few are chosen. Cloning Stem
Cells. 4:147–168. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Westhusin ME, Long CR, Shin T, Hill JR,
Looney CR, Pryor JH and Piedrahita JA: Cloning to reproduce desired
genotypes. Theriogenology. 55:35–49. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Robertson G, Garrick D, Wu W, Kearns M,
Martin D and Whitelaw E: Position-dependent variegation of globin
transgene expression in mice. Proc Natl Acad Sci USA. 92:5371–5375.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dobie KW, Lee M, Fantes JA, Graham E,
Clark AJ, Springbett A, Lathe R and McClenaghan M: Variegated
transgene expression in mouse mammary gland is determined by the
transgene integration locus. Proc Natl Acad Sci USA. 93:6659–6664.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang J, Li L, Cai Y, Xu X, Chen J, Wu Y,
Yu H, Yu G, Liu S, Zhang A, et al: Expression of active recombinant
human lacto-ferrin in the milk of transgenic goats. Protein Expr
Purif. 57:127–135. 2008. View Article : Google Scholar
|
|
51
|
Tomita M, Wakabayashi H, Yamauchi K,
Teraguchi S and Hayasawa H: Bovine lactoferrin and lactoferricin
derived from milk: Production and applications. Biochem Cell Biol.
80:109–112. 2002. View Article : Google Scholar : PubMed/NCBI
|