Introduction
Breast cancer is a serious threat to human health
(1) and a major cause of
morbidity and mortality among women, accounting for ~23% of female
malignancies (2,3). With the development of medical
technology and the prevalence of precancerous screening technology,
breast cancer morbidity and mortality rates have declined in
developed countries, but incidence and mortality are rising
worldwide (4-6). Currently, the main treatment methods
for breast cancer include surgical resection, radiotherapy and
chemotherapy, as well as immunotherapy (7). However, due to its high metastatic
rate, strong invasiveness and recurrence, the prognosis of patients
with breast cancer remains poor (7,8).
Accumulating evidence suggested that breast cancer
metastasis can occur early and has significant organ specificity
(9,10), which may explain the high
expression levels of adhesion molecules, such as vascular cell
adhesion protein 1 (11), in
addition to a series of genes associated with breast cancer
metastasis. Breast cancer cells expressing the epidermal growth
factor receptor oncogene possess neurotropic properties, and
commonly metastasize to the central nervous system (12). A recent study demonstrated a
significant correlation between the low expression level of breast
cancer type 1 susceptibility protein in the tumor tissues of
patients with breast cancer and brain metastasis, but the specific
mechanism remains to be fully elucidated (13). In a previous study, investigation
of circular RNA (circRNA) expression by high-throughput sequencing
identified numerous circRNAs as potential molecular markers for
brain metastasis in breast cancer (14). Among those, the interaction
between hsa_circ_0001944 and microRNA-509 was found to be the most
promising (14). In addition to
brain metastasis, a subset of breast cancer cells expressing C-X-C
chemokine receptor (CXCR2) tend to metastasize to lung tissues rich
in stromal cell-derived factor 1 (SDF1) (15). Although the role of the SDF1/CXCR4
axis in breast cancer metastasis has not been fully elucidated, the
results from a previous meta-analysis suggested that the SDF1/CXCR4
protein expression level is a reliable prognostic biomarker for
breast cancer (16). Bone and
liver metastases (17,18) also play a critical role in breast
cancer. In vivo, it was confirmed that interleukin-1 was a
mediator of bone metastasis in breast cancer cells (19). Although certain biomarkers of
breast cancer can predict organ specificity, the mechanism of
cancer metastasis in complex living systems is not fully
understood. Epithelial-mesenchymal transition (EMT) has been
considered to be responsible for tumor metastasis (20,21). Several studies identified that
tumor cells display decreased epithelial properties and enhanced
mesenchymal properties, associated with reduced expression of
epithelial-related markers, including cytokeratin and E-cadherin,
and increased expression of mesenchymal-related markers, such as
vimentin, N-cadherin and matrix metalloproteinases (MMPs). This is
accompanied by activation of the mitogen-activated protein kinase
(MAPK) signaling pathway (22-24). Amyloid precursor protein (APP) has
a long extracellular domain, a transmembrane domain and a shorter
intracellular domain, and is widely expressed in most cells
(25). Early studies have
reported that β-APP is formed via the hydrolysis of extracellular
fragments accumulated in the brain tissue, affecting the structure
and function of the nervous system, and is one of the factors
associated with the pathogenesis of Alzheimer's disease (26-29). In addition, tumor cells can
promote hematogenous metastasis by increasing the expression of APP
(30). In addition, the
intracellular domain of APP can interact with adaptor proteins,
activating downstream intracellular signaling molecules, thereby
affecting the physiological functions of nerves and immune cells,
influencing cell migration and invasion (31).
APP is upregulated in a variety of cancer types
(32,33). A previous study found that APP is
an independent predictor of poor prognosis in non-intraluminal
breast cancer and an important risk factor for breast cancer distal
lymph node metastasis (34). In
the present study, immunohistochemistry and in vitro
experiments were performed to examine the association between APP
expression in breast cancer and clinical symptoms in patients with
breast cancer. The present results suggested that APP was
positively correlated with the expression of androgen receptor (AR)
and Ki-67. In vitro experiments from the present study
demonstrated that the bioactive androgen dihydrotestosterone
induced APP mRNA transcription in a dose- and time-dependent
manner, while hydroxyflutamide, an AR blocking agent, effectively
inhibited this process. Moreover, the proliferative activity of
breast cancer cells is associated with the expression levels of APP
(35). However, little is known
on the role of APP in breast cancer progression. In the present
study, the effects of APP on the migration and invasion of breast
cancer cells were investigated using APP overexpression and
knockdown cell lines. The present results provides theoretical
support for the development of APP as a novel therapeutic targets
for the management of breast cancer.
Materials and methods
Cell lines
MDA-MB-231, MCF-7, MCF-10, BT549 and BT474 breast
cancer cell lines were obtained from the Shanghai Institute of Life
Sciences Cell Bank and cultured according to the manufacturer's
instructions.
Related reagents
DMEM and FBS were purchased from Gibco (Thermo
Fisher Scientific, Inc.). The empty plasmid pEGFP-n1-APP (cat. no.
69924) and pENTR APP short hairpin (sh)RNA (cat. no. 30135)
plasmids were supplied by Addgene Inc. The transfection reagent
polyetherimide (PEI; cat. no. 03880) was supplied by Sigma-Aldrich
(Merck KGaA). PrimeScript RT reagent kit (Takara Bio, Inc.) and One
Step SYBR-Green PrimeScript RT-PCR kit II (Takara Bio, Inc.) kits
were used for reverse transcription (RT) and quantitative-PCR
(q-PCR), respectively. Transwell chambers and Matrigel were
purchased from BD Biosciences. Rabbit anti-human APP (1:2,000 for
western blot analysis; 1:300 for immunohistochemistry; cat. no.
2452S), mouse anti-human E-cadherin (1:2,000; cat. no. 14472),
mouse anti-human N-cadherin (1:2,000; cat. no. 14215), mouse
anti-human cytokeratin (1:2,000; cat. no. 4545), mouse anti-human
vimentin (1:2,000; cat. no. 49636), mouse anti-human MMP-9
(1:2,000; cat. no. 3852), rabbit anti-human MMP-2 (1:2,000; cat.
no. 4022), rabbit anti-human MMP-3 (1:2,000; cat. no. 14351) and
rabbit anti-human mitogen-activated protein kinase kinase kinase 11
(MLK3) primary antibodies (1:2,000; cat. no. 2817) were purchased
from Cell Signaling Technology, Inc. Rabbit anti-human MEK4
(1:2,000; cat. no. ab33912), rabbit anti-human phosphorylated
(p)-MEK4 (1:2,000; cat. no. ab131353), rabbit anti-human p-MLK3
(1:2,000; cat. no. ab191530), rabbit anti-human JNK3 (1:2,000; cat.
no. ab126591), rabbit anti-human p-JNK3 (1:2,000; cat. no.
ab124956) and rabbit anti-human β-actin primary antibodies
(1:4,000; cat. no. ab179467), as well as horseradish peroxidase
(HRP)-conjugated goat anti-rabbit (1:5,000; cat. no. ab6721) and
goat anti-mouse (1:3,500; cat. no. ab6789) secondary antibodies
were purchased from Abcam. TRIzol® reagent was obtained
from Thermo Fisher Scientific, Inc. qPCR primers were synthesized
by Shanghai Biotech.
Cell culture
MDA-MB-231, MCF-7 and BT474 cells were cultured in
DMEM containing 10% FBS and 1% streptomycin mixture, and then
placed in a humidified atmosphere with 5% CO2 at 37°C.
Cell passaging was conducted using 0.25% trypsin + EDTA.
Human breast carcinoma tissues and
immunohistochemistry
A total of eight female patients with breast cancer
(age, 37-62 years) underwent clinical and histopathological
diagnosis at the First Affiliated Hospital of Xiamen University
between January and December 2018. All patients included in the
study had clinical TNM stage III or IV breast cancer, and had not
been treated with radiotherapy or chemotherapy prior to surgery.
Written informed consent was obtained from each patient. The study
protocol was approved by the Ethics Committee of the First
Affiliated Hospital of Xiamen University. All immunohistochemical
samples were retrieved from the Department of Pathology of the
First Affiliated Hospital of Xiamen University. Immunohistochemical
staining was performed on tissues. Samples were soaked in 10%
neutral buffered formalin at 4°C and embedded in paraffin.
Subsequently, the paraffined slices (thickness, 5 µm) were
dewaxed and rehydrated, then placed on glass slides. Antigen
retrieval was performed in a pressure cooker for 3 min at 120°C.
Slices were then incubated with primary antibody (1:500) at 4°C
overnight, followed by incubation with corresponding secondary
antibodies for 2 h at room temperature. After washing in PBS, the
slices were added with diaminobenzidine, stained with 10%
hematoxylin for 10 sec at room temperature and sealed with neutral
gum. Sections were visualized and photographed using an optical
microscope (Leica Microsystems GmbH; magnification, ×200). Results
were quantified using Image Pro Plus software (version 6.0; Media
Cybernetics).
Plasmid transfection
MDA-MB-231, MCF-7 and BT474 cells in the logarithmic
growth phase were trypsinized and counted, and then seeded into a
6-well plate at a density of 5×105 cells/well. To
determine the effects of APP overexpression, five groups were
defined as follows: i) Untransfected cells (PEI but no plasmid);
ii) control plasmid (PEI with 2 µg control pEGFP plasmid);
iii) control shRNA (PEI with 2 µg control pENTR plasmid);
iv) APP overexpression (PEI with 2 µg pEGFP-n1-APP plasmid);
and v) APP shRNA (PEI with 2 µg pENTR APP shRNA plasmid).
For the subsequent experiments, cells were transfected and
categorized into three groups as follows: i) Control (PEI but no
plasmid); ii) overexpression (PEI with 2 µg pEGFP-n1-APP
plasmid); and iii) silencing (PEI with 2 µg pENTR APP shRNA
plasmid) groups. The transfected cells were used in subsequent
experiments after 48 h.
Drug treatment
MDA-MB-231 cells were cultured in six-well plates at
a density of 5×105 cells/well, and then treated with the
MEK inhibitor PD0325901 (Sigma-Aldrich; Merck KGaA; 1 nM) for 48 h
at 37°C. When drug treatment and transfection were combined,
MDA-MB-231 cells were cultured in six-well plates at a density of
5×105 cells/well, and then transfected with APP
overexpressing plasmid for 24 h, followed by treatment with the MEK
inhibitor PD0325901 (1 nM) for 48 h.
Migration assay
MDA-MB-231, MCF-7 and BT474 cells in the logarithmic
growth phase were trypsinized and counted, then seeded in a 6-well
plate at a density of 5×105 cells per well. After
reaching complete adherence and ~60% confluence, the cells were
transfected with the corresponding plasmids. After 48 h of
incubation, the cells were digested and counted, resuspended in
serum-free medium, and inoculated into the upper Transwell chamber
at a density of 1×104 cells/well. The lower chamber was
filled with 500 µl complete medium containing 10% FBS in a
24-well plate. After the upper and lower chambers were assembled,
they were placed in a cell culture incubator for 4 h, and the upper
chamber was removed. The Transwell inserts were scraped with cotton
swabs to remove residual cells. Cells on the lower surface of the
well were stained with 0.1% crystal violet solution for 30 min at
room temperature and visualized using an optical microscope (Leica
Microsystems GmbH; magnification, ×100).
Invasion assay
The matrix collagen solution (8 mg/ml) was diluted
8X with serum-free medium. The diluted Matrigel (50 µl) was
then added to the upper chamber of the Transwell insert in advance,
and placed in a cell culture incubator for a minimum of 1 h at
37°C. After treating the cells as described in the migration assay,
cells resuspended in the serum-free medium were seeded at a density
of 1×104 cells/well into the upper chamber of the
Transwell insert, where the Matrigel solution had been previously
added at room temperature and then put into culture incubator at
37°C for 30 min. The lower chamber was filled with 500 µl
complete medium containing 10% FBS. After the upper and lower
chambers were assembled, they were placed in a cell culture
incubator for 12 h at 37°C. Next, the cells were treated as
described in the migration assay section. Stained cells were
photographed using an optical microscope (Leica Microsystems GmbH;
magnification, ×100) and counted in five randomly selected
fields.
Gap closure assay
A total of 1×106 MDA-MB-231 cells were
seeded into a 6-well plate, on which the UV-sterilized
polydimethylsiloxane blocks (1 mm ×2 cm) were placed before. The
culture medium was supplemented with 10% FBS. When the cells
reached a confluence of 90%, the blocks were removed and the width
was photographed using an optical microscope (Leica Microsystems
GmbH; magnification, ×40) and measured at 24, 48 and 72 h. The gap
closure values were calculated as follows: Percentage of gap
closure= 1 -(widtht/width0) ×100% (36).
RT-qPCR
Total RNA was extracted from MDA-MB-231, MCF-7 and
BT474 cells using TRIzol reagent (Thermo Fisher Scientific, Inc.)
at 4°C, according to the manufacturer's instructions. A ratio of
A260/A280 was considered to be acceptable within the range of
1.8-2.0. The RT reaction was performed as follows: Initial
incubation at 37°C for 5 min, followed by sequential steps at 42°C
for 15 min, at 85°C for 10 sec and at 4°C for 60 min. β-actin was
selected as the reference gene. For the q-PCR, the thermocycling
conditions were as follows: Pre-denaturation at 95°C for 5 min,
followed by 40 cycles of denaturation at 95°C for 15 sec and of
annealing at 60°C for 15 sec. The 2−ΔΔCq method
(37) was used to quantify gene
expression. The primer sequences used were as follows: β-actin
forward, 5′-CCT CGC CTT TGC CGA TCC-3′ and reverse, 5′-GGA TCT TCA
TGA GGT AGT CAG TC-3′; MMP-9 forward, 5′-AAT CTC ACC GAC AGG CAG
CT-3′ and reverse, 5′-CCA AAC TGG ATG ACG ATG TC-3′; APP forward,
5′-TCT CGT TCC TGA CAA GTG CAA-3′ and reverse, 5′-GCA AGT TGG TAC
TCT TCT CAC TG-3′; cytokeratin forward, 5′-ACC AAG TTT GAG ACG GAA
CAG-3′ and reverse, 5′-CCC TCA GCG TAC TGA TTT CCT-3′; vimentin
forward, 5′-GCC CTA GAC GAA CTG GGT C-3′ and reverse, 5′-GGC TGC
AAC TGC CTA ATG AG-3′; MMP-2 forward, 5′-TAC AGG ATC ATT GGC TAC
ACA CC-3′ and reverse, 5′-GGT CAC ATC GCT CCA GAC T-3′; MMP-3
forward, 5′-CTG GAC TCC GAC ACT CT G GA-3′ and reverse, 5′-CAG GAA
AGG TTC TGA AGT G AC C-3′; E-cadherin forward, 5′-CGA GAG CTA CAC
GTT CAC GG-3′ and reverse, 5′-GGG TGT CGA GGG AAA AAT AGG-3′;
N-cadherin forward, 5′-TCA GGC GTC TGT AGA GGC TT-3′ and reverse,
5′-ATG CAC ATC CTT CGA TAA GA C TG-3′.
Western blot analysis
The cells were lysed with RIPA buffer (Beyotime
Institute of Biotechnology) to extract the total protein from each
group. Protein lysates were centrifuged at 12,000 × g for 15 min at
4°C and the protein concentration was determined using a
bicinchoninic acid assay. Samples (30 µg per lane) were
isolated using 8-10% SDS-PAGE and transferred to PVDF membranes,
followed by blocking with 5% skimmed milk at room temperature for 2
h. Target proteins were detected with specific antibodies overnight
at 4°C and observed on Bio-Rad Gel Imager using HRP-conjugated
secondary antibodies (1:5,000; incubated for 3 h at room
temperature) and enhanced chemiluminescence reagents (Western
Lightning, Inc.). The bands were quantified using Quantity One 4.6
software (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data analysis were performed using SPSS 21.0 (IBM
Corp.). Graphs were drawn using GraphPad Prism 6.0 (GraphPad
Software, Inc.). Multivariate mean comparisons were performed using
one-way ANOVA with Dunnett's test. Student's t-test was used to
analyze the difference between two groups. All experimental data
were obtained from ≥3 independent experiments. Data are the
presented as mean ± SD. P<0.05 was considered to indicate a
statistically significant difference.
Results
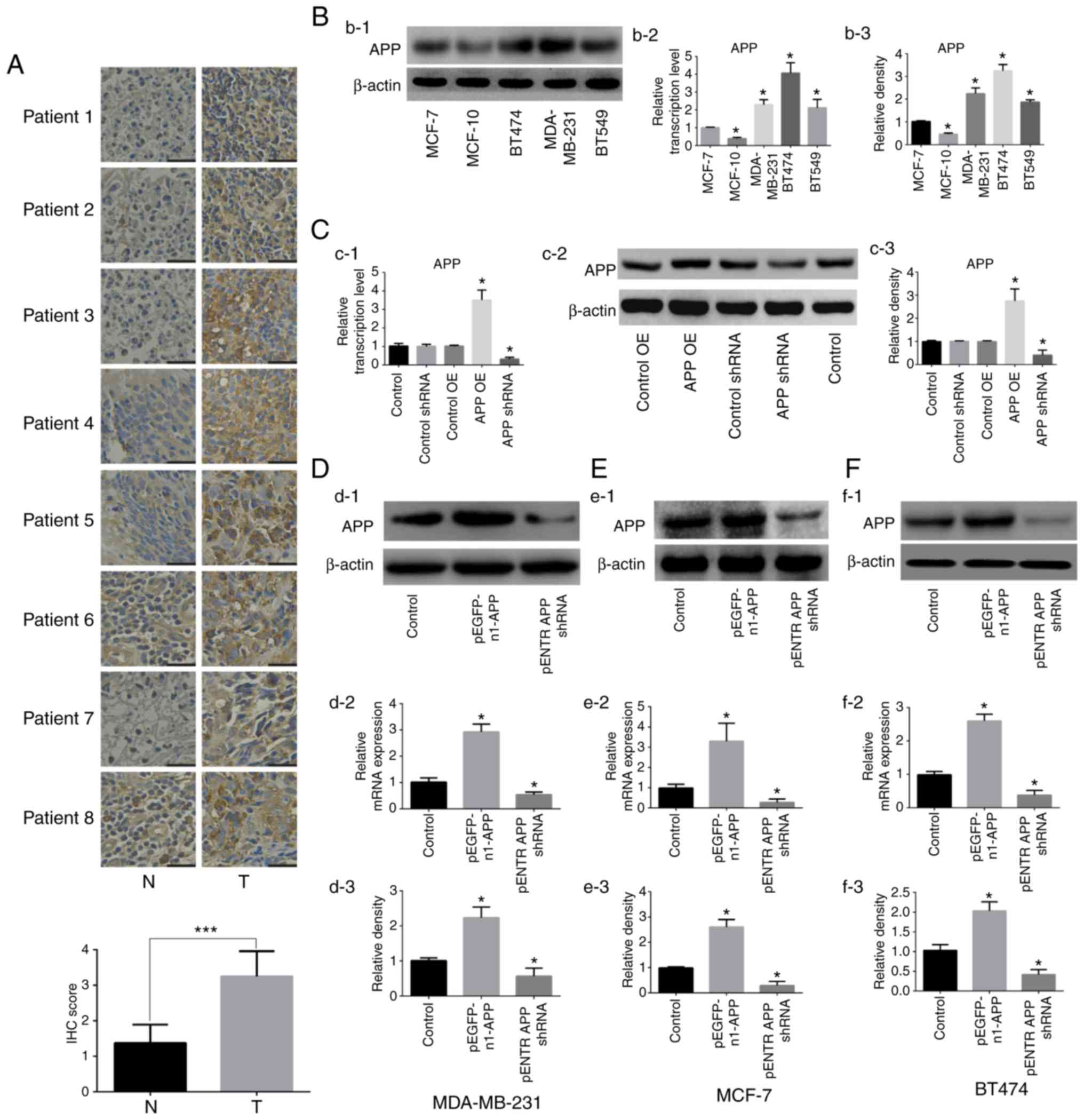
APP expression in human breast cancer
tissue samples and verification of transfection efficiency
following APP overexpression and knockdown in breast cancer
cells
The expression of APP was detected by
immunohistochemistry in human breast cancer tissues. The results
shown in Fig. 1A indicated a
significant upregulation of APP in breast cancer tissues compared
with adjacent tissues (P<0.001). Subsequently, the expression
levels of APP in five breast cancer cell lines were examined
(Fig. 1B), and three cell lines
(MDA-MB-231, MCF-7 and BT474) showing a moderate expression of APP
were selected for further experiments. APP overexpression and
knockdown were then performed on these selected cell lines.
Following transfection with the corresponding
plasmids, it was found that the empty plasmids had no effects on
the expression of APP in MDA-MB-231 cell line, therefore cells
treated with PEI alone were used as the control in the subsequent
experiments (Fig. 1C). The
expression levels of APP in MDA-MB-231, MCF-7 and BT474 cells were
detected by RT-qPCR and western blot analysis. The mRNA and protein
expression levels of APP in cells transfected with the pEGFP-n1-APP
plasmid were significantly increased compared with control cells
(Fig. 1D-F; P<0.05). APP mRNA
and protein expression levels in pENTR APP shRNA cells were
significantly decreased compared with control cells (P<0.05).
The present results suggested that the transfection experiments
were successful.
APP promotes the migratory and invasive
ability of breast cancer cells
The results of the Transwell assay showed that the
migratory (Fig. 2A-C) and
invasive (Fig. 2E-G) abilities of
pEGFP-n1-APP-transfected cells were significantly enhanced compared
with the control group (P<0.05), whereas pENTR APP
shRNA-transfected cells exhibited significantly impaired migratory
and invasive abilities compared with the control group (P<0.05).
The results of the gap closure assay revealed that APP
overexpression significantly promoted gap closure in MDA-MB-231
cells, while APP knockdown inhibited gap closure (Fig. 2D; P<0.05).
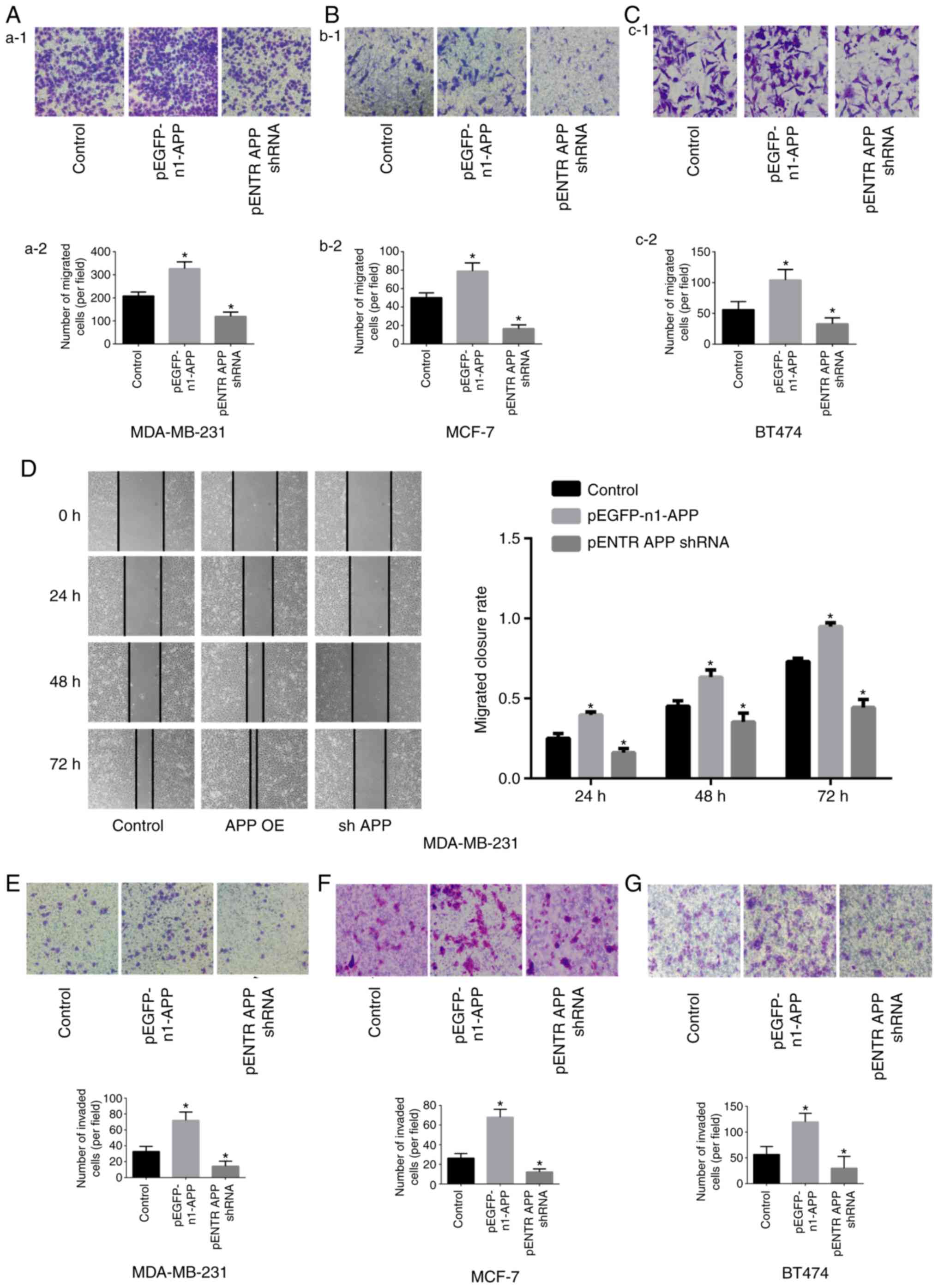 | Figure 2Effects of APP overexpression and
silencing on the migration of MDA-MB-231, MCF-7 and BT474 breast
cancer cells. Staining images from the Transwell assay. The effects
of APP overexpression and silencing were investigated in (Aa-1)
MDA-MB-231 cells and (Aa-2) quantified, in (Bb-1) MCF-7 cells and
(Bb-2) quantified, and in (Cc-1) BT474 breast cancer cells and
(Cc-2) quantified. Magnification in Aa-1 and Bb-1, ×100. Scale bar,
200 µm. Magnification in Cc-1, ×400. Scale bar, 50
µm. (D) Representative gap closure assay images and
statistical analysis, showing the effects of APP overexpression and
silencing on the cell migration of MDA-MB-231 cells. Magnification,
×40. Scale bar, 250 µm. Representative images of the effects
of APP overexpression and silencing on the invasive ability of
breast cancer (E) MDA-MB-231, (F) MCF-7 and (G) BT474 cells
(magnification, ×100; Scale bar, 200 µm) and the statistical
results of the number of invading cells. Five representative fields
were randomly imaged and quantified for each well in cell migration
and invasion assays. Data are presented as the mean ± SD from three
independent experiments. *P<0.05 vs. control. APP,
amyloid precursor protein; shRNA, short hairpin RNA; OE,
overexpression. |
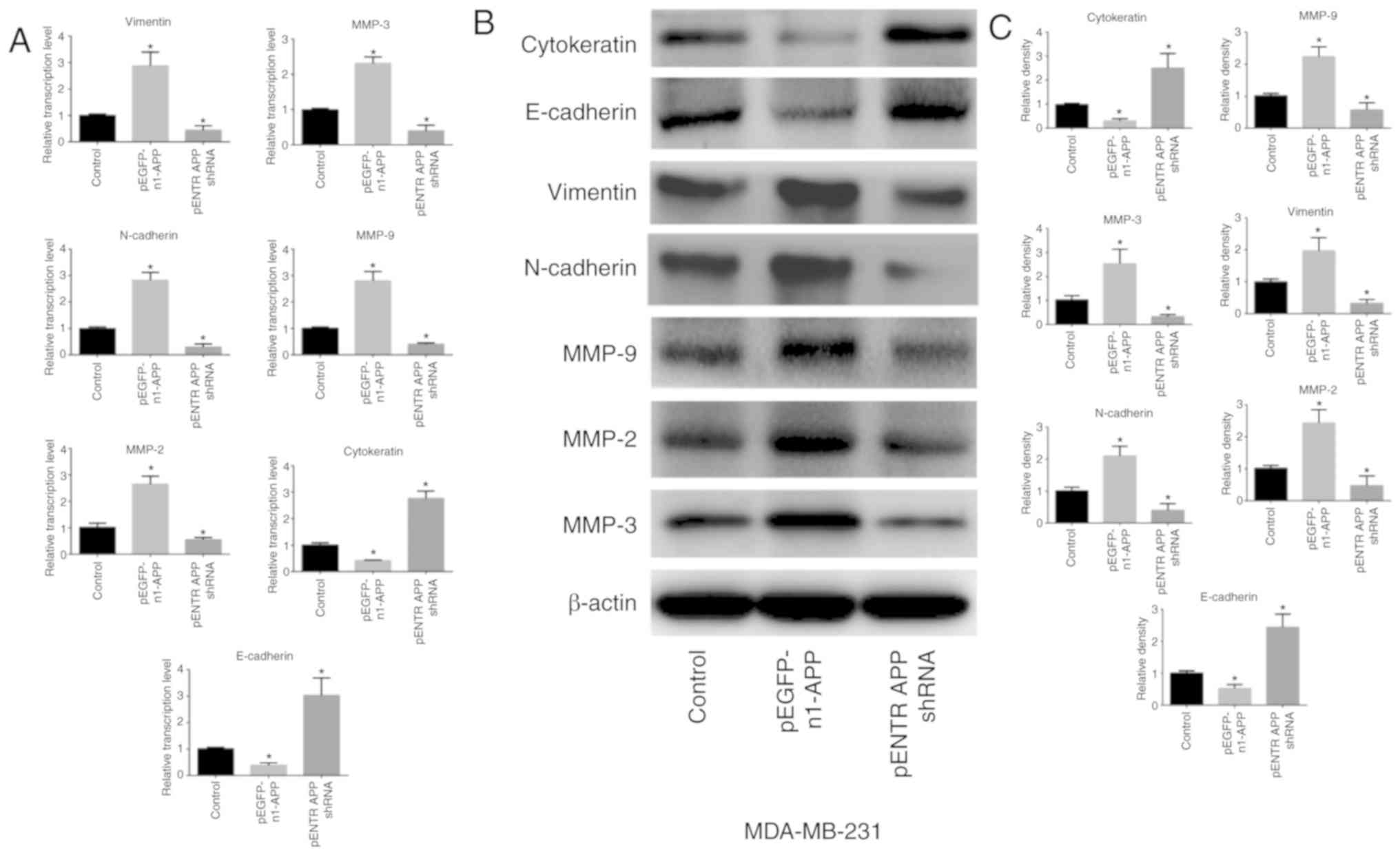
APP regulates the expression of
EMT-related genes in breast cancer
APP overexpression significantly upregulated the
expression levels of the mesenchymal-related genes MMP-9, MMP-2,
MMP-3, N-cadherin and vimentin in MDA-MB-231 cells (P<0.05),
whereas the expression levels of the epithelial-related genes
E-cadherin and cytokeratin were significantly attenuated (Fig. 3; P<0.05). APP knockdown
exhibited the opposite effects (P<0.05).
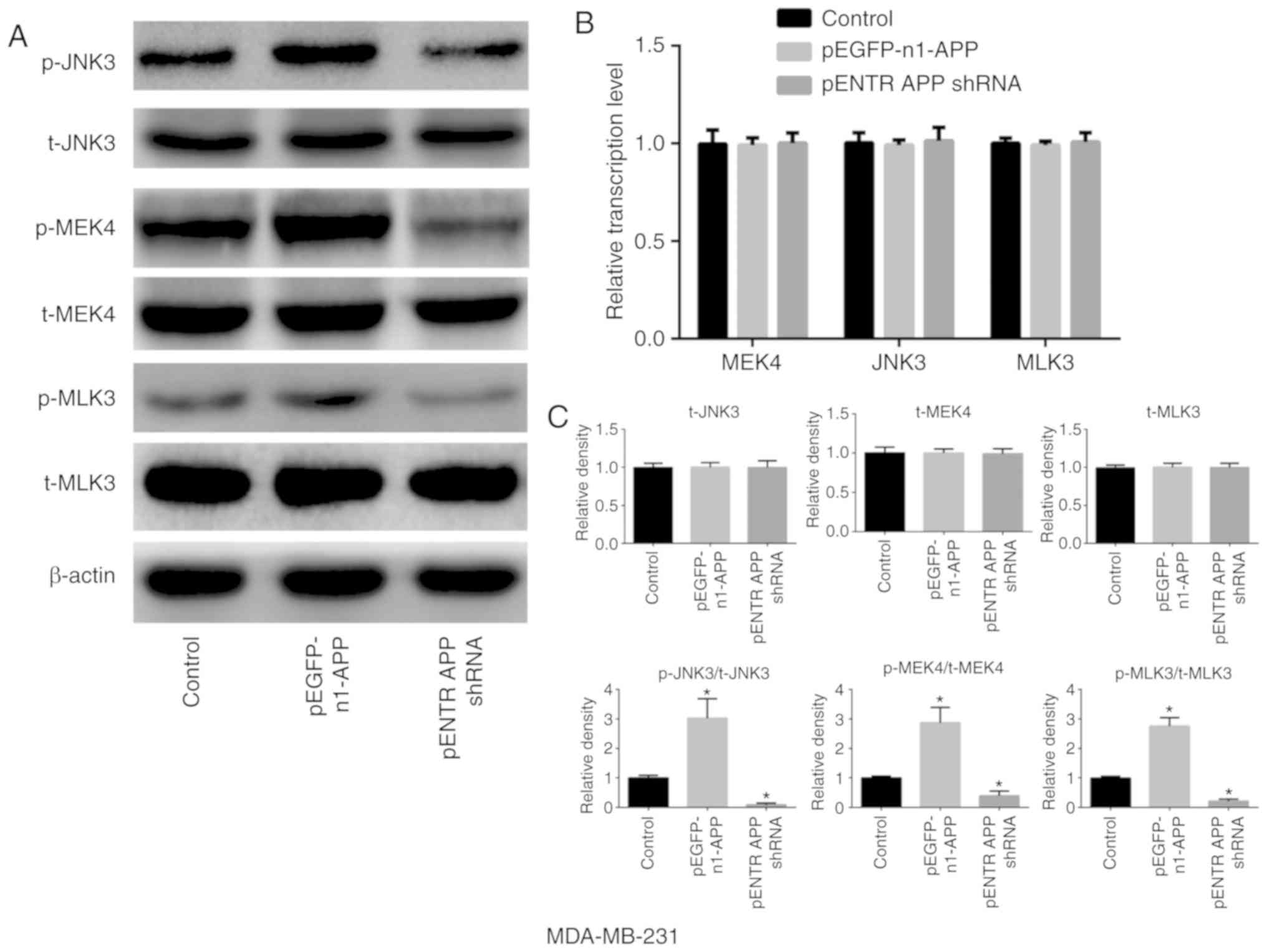
APP is involved in the expression and
phosphorylation of MAPK signaling pathway-related molecules in
breast cancer
APP overexpression notably increased the
phosphorylation of MLK3, MEK4 and JNK3 in breast cancer MDA-MB-231
cells compared with the control group (Fig. 4; P<0.05). However, no
significant differences were observed in the expression levels of
total MLK3, MEK4 and JNK3 in any of the groups compared with the
control. However, the phosphorylation levels of MLK3, MEK4 and JNK3
were significantly decreased following APP silencing
(P<0.05).
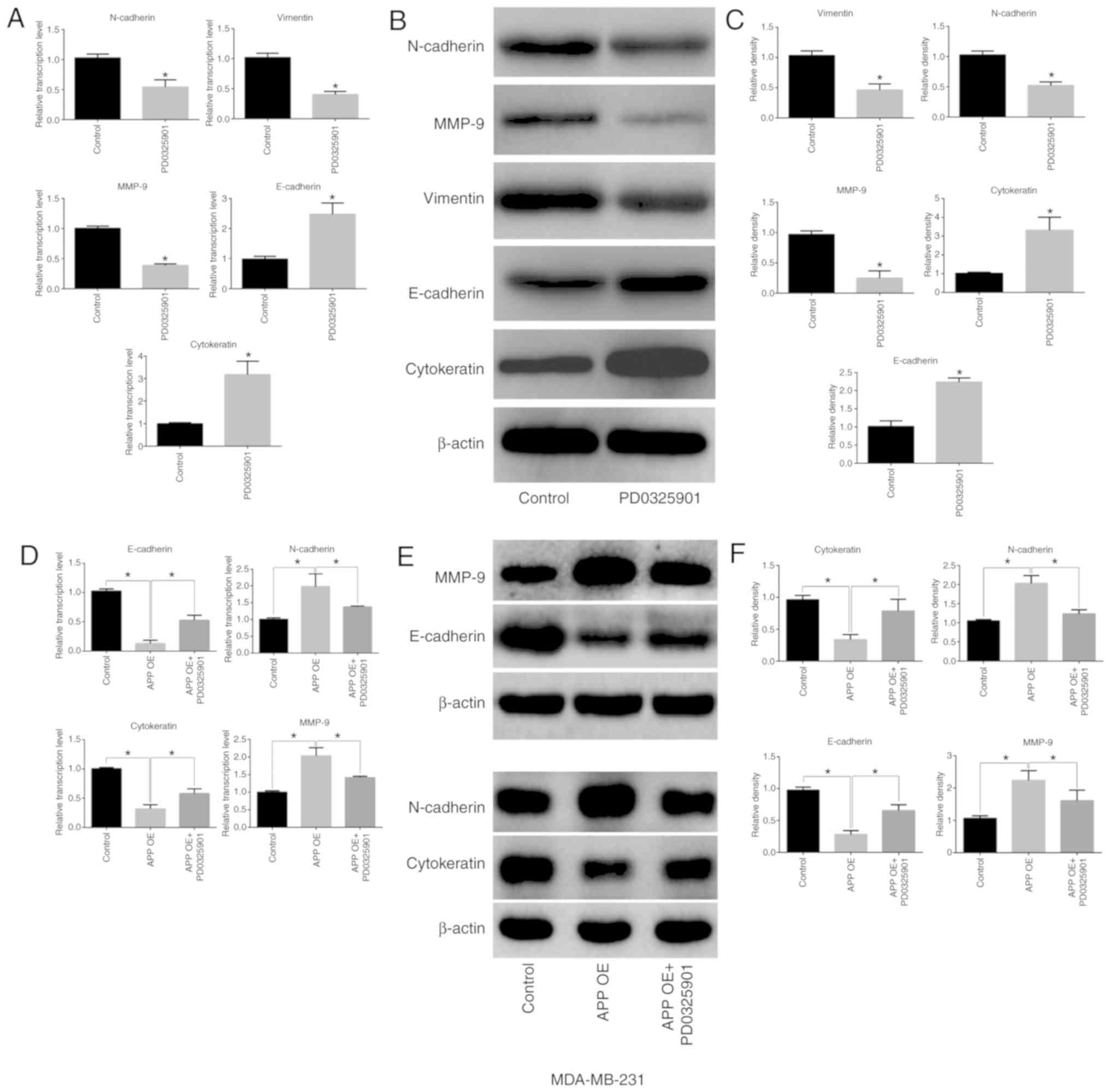
MEK inhibitor PD0325901 does not affect
the expression of APP but impairs the migratory and invasive
ability of breast cancer cells
MDA-MB-231 cells were cultured and seeded into
6-well plates at a density of 5×105 cells/well. They
were then treated with the MEK inhibitor PD0325901 (1 nM) for 48 h.
Cells not treated with the inhibitor were used as the control
group. After 48 h, total protein was collected to detect APP
protein expression. In addition, Transwell and Matrigel assays were
performed. PD0325901 treatment alone did not affect the expression
levels of APP (Fig. 5A), but
significantly inhibited the migratory and invasive abilities of
breast cancer cells compared with the control group (Fig. 5B and C; P<0.05). Moreover,
following APP overexpression, MDA-MB-231 cells (Fig. 5D) were treated with MEK inhibitor
PD0325901, and it was found that PD0325901 could significantly
reduce the migratory and invasive ability increased following APP
overexpression, as shown by migration, invasion and gap closure
assays (Fig. 5E-H;
P<0.05).
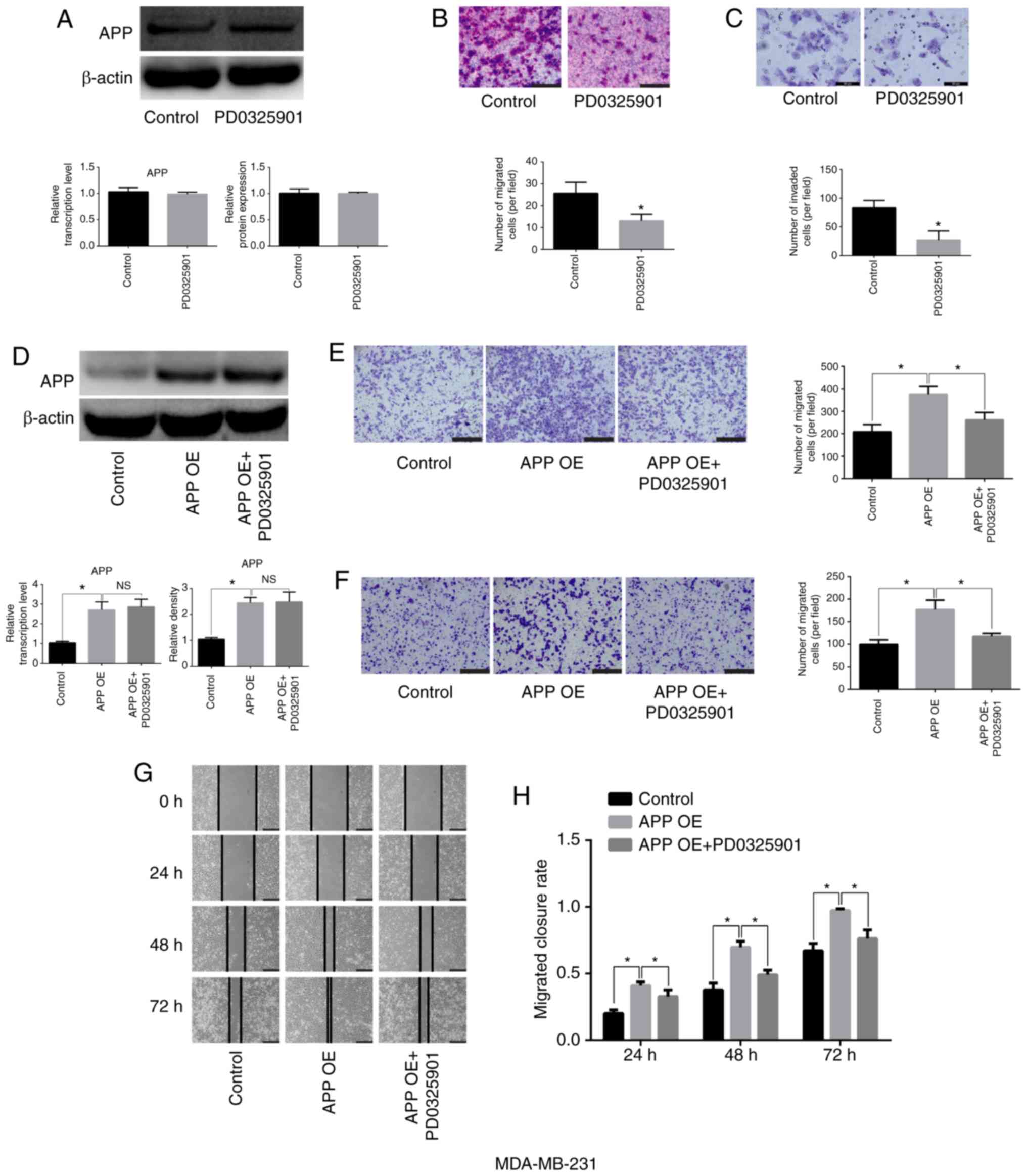 | Figure 5Effects of the MEK inhibitor
PD0325901 on the expression level of APP and the migratory and
invasive abilities of MDA-MB-231 breast cancer cells. (A) MEK
inhibitor exerted no effect on the mRNA and protein expression
levels of APP, as indicated by RT-qPCR and western blot analysis.
(B) Representative staining images showing the effect of PD0325901
on the migratory ability of breast cancer cells and statistical
analysis of the number of migrated cells. Magnification, ×200.
Scale bar, 100 µm. (C) Representative staining images
showing the effect of PD0325901 on the invasive ability of breast
cancer cells and statistical analysis of the number of invaded
cells. Magnification, ×400. Scale bar, 50 µm. (D) PD0325901
exerted no effect on APP-overexpressing MDA-MB-231 breast cancer
cells. Following APP overexpression and treatment with the MEK
inhibitor PD0325901, PD0325901 significantly reduced the migratory
and invasive ability of breast cancer cells, as indicated by the
(E) migration and (F) invasion assays. Scale bar, 200 µm.
Magnification, ×100. (G) Representative gap closure assay images
and (H) statistical analysis. Magnification, ×40. Scale bar, 250
µm. In total, five representative fields were randomly
imaged and quantified for each well in the cell migration and
invasion assays. All the experiments were performed in triplicate.
Data are presented as the mean ± SD. *P<0.05. APP,
amyloid precursor protein; MEK, mitogen-activated protein kinase
kinase; OE, overexpression; RT-qPCR, reverse
transcription-quantitative PCR; NS, not significant. |
MEK inhibitor PD0325901 regulates the
expression of EMT-related factors in breast cancer cells
PD0325901 treatment alone significantly inhibited
the expression levels of MMP-9, N-cadherin and vimentin in
MDA-MB-231 breast cancer cells (P<0.05), but significantly
increased the expression levels of E-cadherin and cytokeratin
(Fig. 6A-C; P<0.05). A
decreased expression of epithelial-associated markers (E-cadherin
and Cytokeratin) and increased expression of mesenchymal markers
(MMP-9 and N-cadherin) was identified in APP overexpressing breast
cancer cells compared with the control group (Fig. 6D-F; P<0.05). Conversely,
PD0325901 treatment significantly reversed the expression levels of
EMT-related genes caused by APP overexpression (Fig. 6D-F; P<0.05).
MEK inhibitor PD0325901 is involved in
the expression and phosphorylation of MAPK signaling
pathway-related factors in breast cancer cells
The present results suggested that PD0325901
treatment significantly inhibited the activation of JNK3, which is
downstream of MEK (Fig. 7A-C;
P<0.05), but did not affect the expression of total JNK3
compared with untreated cells. In addition, no significant
differences were observed in the expression and activation levels
of MEK4 and its upstream factor, MLK3. The overexpression of APP
increased the phosphorylation level of MEK, MLK3 and JNK3. In
addition, in MDA-MB-231 cells transfected with APP overexpressing
plasmid and treated with the MEK inhibitor PD0325901 (1 nM), the
phosphorylation levels of MEK4 and MLK3 were not significantly
changed compared with the overexpression group (Fig. 7F). By contrast, PD0325901
treatment markedly reversed the phosphorylation of JNK3 caused by
APP overexpression (P<0.05). Moreover, cell transfection and
drug treatment had no effect on the protein expression levels of
the unphosphorylated forms of MEK4, JNK3 and MLK3 in MDA-MB-231
cells (Fig. 7D-F).
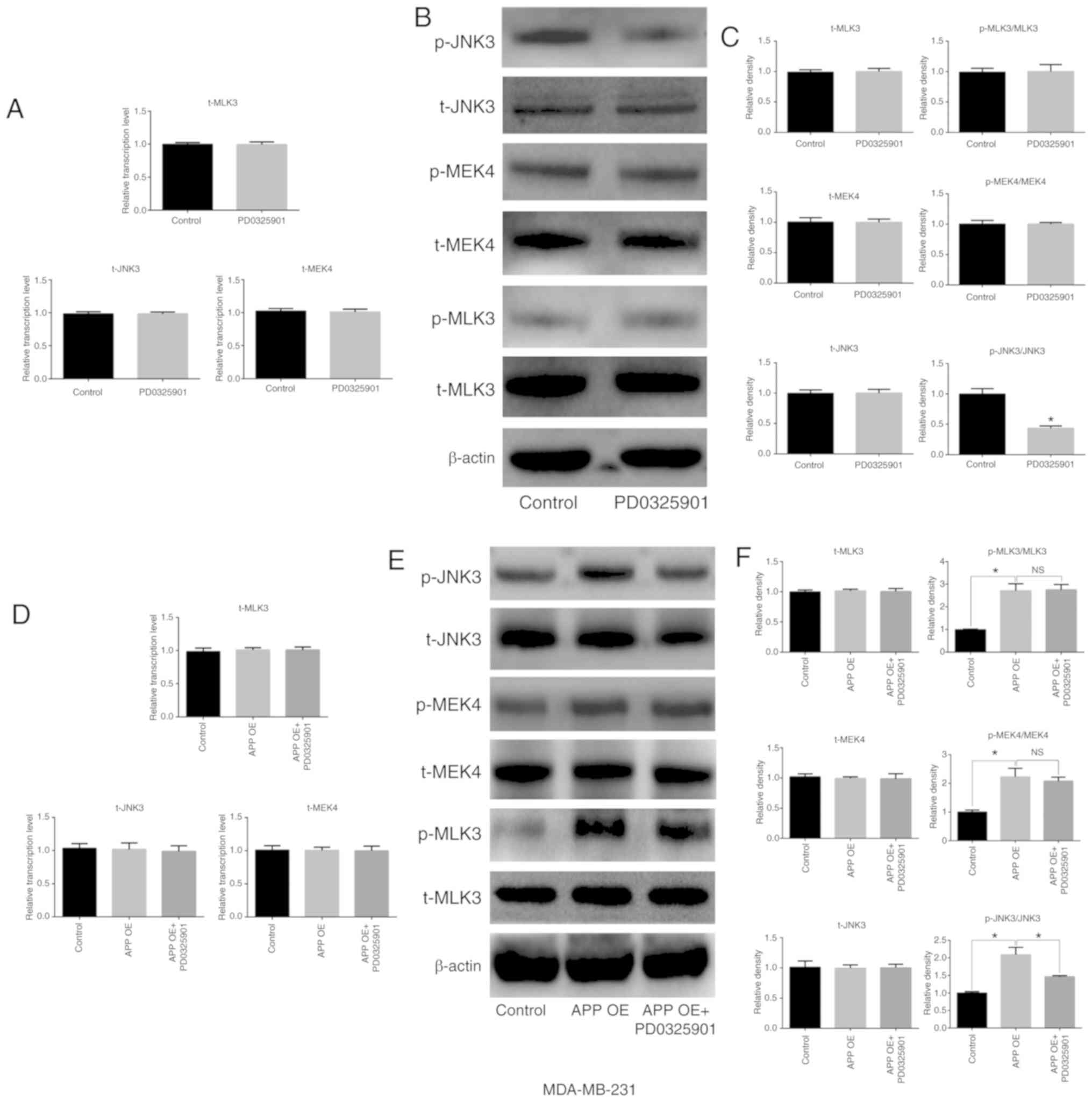 | Figure 7Effects of the MEK inhibitor
PD0325901 on the expression and phosphorylation of MAPK signaling
pathway-related molecules in MDA-MB-231 breast cancer cells. (A)
Effect of PD0325901 on the mRNA expression levels of MAPK signaling
pathway-related genes in breast cancer cells by RT-qPCR. (B) Effect
of PD0325901 on the protein expression levels of MAPK signaling
pathway-related molecules in breast cancer cells by western blot
analysis, and (C) statistical analysis of the western blot bands.
APP overexpression in breast cancer cells treated with PD0325901.
(D) mRNA expression level of MAPK signaling pathway-related genes
in breast cancer cells was detected by RT-qPCR. (E) Protein
expression levels of MAPK signaling pathway-related molecules in
breast cancer cells were determined by western blot analysis. (F)
Statistical analysis of the relevant western blot bands. All the
experiments were performed in triplicate. Data are presented as the
mean ± SD. *P<0.05. MEK, mitogen-activated protein
kinase kinase; MAPK, mitogen-activated protein kinase; APP, amyloid
precursor protein; RT-qPCR, reverse transcription-quantitative PCR;
MEK4, mitogen-activated protein kinase kinase 4; OE,
overexpression; p-, phosphorylated; t-, total; NS, not
significant. |
Discussion
Breast cancer metastasis is associated with a
variety of factors (38,39). Previous studies have suggested
that cancer metastasis mainly occurs at the later stages, when the
primary tumor reaches a certain size and tumor cells escape from
its periphery (9). However, it
has been reported that tumor metastasis also occurs at the early
stages, as explained by the parallel metastasis theory (9). It was previously demonstrated that
overexpression of APP can promote hematogenous metastasis in
melanoma and lung cancer cells (30). When tumor cells penetrate the
vascular endothelial barrier, they bind to death receptor 6
expressed on the endothelial cell through the APP extracellular
domain fragment on the cell surface, inducing programmed necrosis
of endothelial cells and allowing tumor cells to enter the
bloodstream and metastasize to distant sites (30). However, whether APP affects the
migratory and invasive abilities of tumor cells remains
unclear.
APP has been shown to be upregulated in a variety of
cancer types, including breast, lung and cervical cancer (40-44), but the effect of APP
overexpression on the biological features of tumor cells has not
been clearly determined. In the present study, following APP
overexpression, the migration and invasion of breast cancer cells
were significantly increased. Conversely, APP silencing reduced the
migratory and invasive ability of breast cancer cells. It has been
previously reported that APP promotes cancer cell migration and
invasion, and a similar phenotype has been observed in prostate
cancer (44). By silencing APP in
LNCaP and DU145 prostate cancer cells, cell proliferation was
impaired, and cell migration and invasion were notably attenuated
(44). Moreover, APP silencing in
prostate cancer cells significantly inhibited the expression of
EMT-related genes, in particular of MMP family members (44). However, it is unclear how the
expression of EMT-related genes is regulated by APP silencing.
EMT is a necessary step in the process of tumor
metastasis and affects the prognosis of patients with tumors
(23). When tumor cells undergo
EMT, their migratory and invasive abilities are significantly
increased, promoting cancer metastasis (22). The present study investigated the
effect of APP on EMT-related gene expression in breast cancer
cells. Both RT-qPCR and western blot analysis results indicated
that the overexpression of APP in MDA-MB-231 cells increased the
expression levels of the mesenchymal markers MMP-9, MMP-2, MMP-3,
N-cadherin and vimentin, whereas the expression levels of the
epidermal-associated markers N-cadherin and cytokeratin were
significantly reduced. Therefore, the present results suggested
that APP affected the migration and invasion of breast cancer cells
by regulating the expression of EMT-related proteins. Various
studies have reported that MCF-7 cells, as a metastatic cell line,
have migratory and invasive properties (45,46). The present results suggested that
MCF-7, MDA-MB-231 and BT474 cells presented similar responses to
altered APP expression.
The present results suggested that APP affected the
expression of EMT-related genes in breast cancer cells; however,
the specific mechanism of action remains unclear. A previous study
demonstrated that the intracellular domain fragment of the APP
protein activates the MAPK signaling pathway (47), which is associated with EMT
activation (48). When MAPK is
activated, its downstream transcription factors enter the nucleus
and promote the expression of mesenchymal-related genes, thereby
promoting the progression of EMT (22,49). The present study indicated that
the overexpression of APP activated MAPK signaling pathway
components, including MLK3, MEK4 and JNK3, whose phosphorylation
levels were found to be significantly upregulated following APP
overexpression. However, APP overexpression exerted no effect on
the total expression levels of MLK3, MEK4 and JNK3. Additionally,
PD0325901 (MEK inhibitor) treatment significantly reversed the
expression levels of EMT-related genes and the phosphorylation of
the MAPK signaling pathway-related factor JNK3 induced by APP
overexpression. Collectively, the regulation of the MAPK signaling
pathway may be a possible mechanism through which APP regulates EMT
in breast cancer cells, affecting their migratory and invasive
abilities. In conclusion, the present results suggested that APP
could regulate the expression of EMT-related genes in breast cancer
cells by activating MAPK signaling pathway-related proteins. The
activation of these factors could promote the EMT of breast cancer
cells, thus increasing the migration and invasion of breast cancer
cells. However, the present study presents certain limitations,
including the limited number of human tissues and the lack of
animal models, which are required to further investigate whether
APP may be used as a therapeutic target for metastatic breast
cancer.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CL conceived and designed the study. XW and SC
performed the experiments. CL provided experimental technical
guidance. All authors performed the data analysis and drafted the
manuscript. All authors have read and approved the final version of
the manuscript for publication.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of the First Affiliated Hospital of Xiamen University
(approval no. 2016-318S). Written informed consent was obtained
from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Antoni MH and Dhabhar FS: The impact of
psychosocial stress and stress management on immune responses in
patients with cancer. Cancer. 125:1417–1431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cedolini C, Bertozzi S, Londero AP,
Bernardi S, Seriau L, Concina S, Cattin F and Risaliti A: Type of
breast cancer diagnosis, screening, and survival. Clin Breast
Cancer. 14:235–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Criscitiello C, Viale G and Curigliano G:
Peptide vaccines in early breast cancer. Breast. 44:128–134. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Feng C, Zhang H, Chen J, Wang S, Xin Y, Qu
Y, Zhang Q, Ji W, Yamashita F, Rui M and Xu X: Ratiometric
co-encapsulation and co-delivery of doxorubicin and paclitaxel by
tumor-targeted lipo-disks for combination therapy of breast cancer.
Int J Pharm. 560:191–204. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Balazy KE, Benitez CM, Gutkin PM, Jacobson
CE, von Eyben R and Horst KC: Association between primary language,
a lack of mammographic screening, and later stage breast cancer
presentation. Cancer. 125:2057–2065. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peart O: Breast intervention and breast
cancer treatment options. Radiol Technol. 86:535M–558M; quiz
559-562. 2015.PubMed/NCBI
|
|
8
|
Castaneda SA and Strasser J: Updates in
the treatment of breast cancer with radiotherapy. Surg Oncol Clin N
Am. 26:371–382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Klein CA: Parallel progression of primary
tumours and metastases. Nat Rev Cancer. 9:302–312. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yousefi M, Nosrati R, Salmaninejad A,
Dehghani S, Shahryari A and Saberi A: Organ-specific metastasis of
breast cancer: Molecular and cellular mechanisms underlying lung
metastasis. Cell Oncol (Dordr). 41:123–140. 2018. View Article : Google Scholar
|
|
11
|
Sharma R, Sharma R, Khaket TP, Dutta C,
Chakraborty B and Mukherjee TK: Breast cancer metastasis: Putative
therapeutic role of vascular cell adhesion molecule-1. Cell Oncol
(Dordr). 40:199–208. 2017. View Article : Google Scholar
|
|
12
|
Uemura MI, French JT, Hess KR, Liu D,
Raghav K, Hortobagyi GN, Arun BK, Valero V, Ueno NT, Alvarez RH, et
al: Development of CNS metastases and survival in patients with
inflammatory breast cancer. Cancer. 124:2299–2305. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Szarszewska M, Markowska A, Jach R,
Marszałek A, Filas V, Bednarek W, Olejek A, Tomczak P, Sajdak S,
Nowak-Markwitz E, et al: Significance of BRCA1 expression in breast
and ovarian cancer patients with brain metastasis-A multicentre
study. Adv Med Sci. 64:235–240. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fu B, Zhang A, Li M, Pan L, Tang W, An M,
Liu W and Zhang J: Circular RNA profile of breast cancer brain
metastasis: Identification of potential biomarkers and therapeutic
targets. Epigenomics. 10:1619–1630. 2018. View Article : Google Scholar
|
|
15
|
Rice SL and Friedman KP: Clinical PET-MR
imaging in breast cancer and lung cancer. PET Clin. 11:387–402.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu H, Li Z, Deng M, Liu Q, Zhang T, Guo
W, Li P and Qiao W: Prognostic and clinicopathological value of
CXCL12/SDF1 expression in breast cancer: A meta-analysis. Clin Chim
Acta. 484:72–80. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
BacalbaȘa N, Balescu I, Dima S and Popescu
I: Long-term survivors after liver resection for breast cancer
liver metastases. Anticancer Res. 35:6913–6917. 2015.
|
|
18
|
Ma R, Feng Y, Lin S, Chen J, Lin H, Liang
X, Zheng H and Cai X: Mechanisms involved in breast cancer liver
metastasis. J Transl Med. 13:642015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holen I, Lefley DV, Francis SE, Rennicks
S, Bradbury S, Coleman RE and Ottewell P: IL-1 drives breast cancer
growth and bone metastasis in vivo. Oncotarget. 7:75571–75584.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Neelakantan D, Zhou H, Oliphant MUJ, Zhang
X, Simon LM, Henke DM, Shaw CA, Wu MF, Hilsenbeck SG, White LD, et
al: EMT cells increase breast cancer metastasis via paracrine GLI
activation in neighbouring tumour cells. Nat Commun. 8:157732017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chaffer CL, San Juan BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu M, Wang S, Wang Y, Wu H, Frank JA,
Zhang Z and Luo J: Role of p38γ MAPK in regulation of EMT and
cancer stem cells. Biochim Biophys Acta Mol Basis Dis.
1864:3605–3617. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tian M and Schiemann WP: TGF-β stimulation
of EMT programs elicits non-genomic ER-α activity and anti-estrogen
resistance in breast cancer cells. J Cancer Metastasis Treat.
3:150–160. 2017. View Article : Google Scholar :
|
|
24
|
Mutlu M, Saatci Ö, Ansari SA, Yurdusev E,
Shehwana H, Konu Ö, Raza U and Şahin Ö: miR-564 acts as a dual
inhibitor of PI3K and MAPK signaling networks and inhibits
proliferation and invasion in breast cancer. Sci Rep. 6:325412016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang X, Zhou X, Li G, Zhang Y, Wu Y and
Song W: Modifications and trafficking of APP in the pathogenesis of
Alzheimer's disease. Front Mol Neurosci. 10:2942017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hefter D and Draguhn A: APP as a
protective factor in acute neuronal insults. Front Mol Neurosci.
10:222017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hatami A, Monjazeb S, Milton S and Glabe
CG: Familial Alzheimer's disease mutations within the amyloid
precursor protein alter the aggregation and conformation of the
amyloid-β peptide. J Biol Chem. 292:3172–3185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wilkins HM and Swerdlow RH: Amyloid
precursor protein processing and bioenergetics. Brain Res Bull.
133:71–79. 2017. View Article : Google Scholar :
|
|
29
|
Habib A, Sawmiller D and Tan J: Restoring
soluble amyloid precursor protein α functions as a potential
treatment for Alzheimer's disease. J Neurosci Res. 95:973–991.
2017. View Article : Google Scholar
|
|
30
|
Strilic B, Yang L, Albarrán-Juárez J,
Wachsmuth L, Han K, Müller UC, Pasparakis M and Offermanns S:
Tumour-cell-induced endothelial cell necroptosis via death receptor
6 promotes metastasis. Nature. 536:215–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lei H, Zhang Y, Huang L, Xu S, Li J, Yang
L, Wang L, Xing C, Wang X and Peng Y: L-3-n-Butylphthalide
regulates proliferation, migration, and differentiation of neural
stem cell in vitro and promotes neurogenesis in APP/PS1 mouse model
by regulating BDNF/TrkB/CREB/Akt pathway. Neurotox Res. 34:477–488.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Harder H, Holroyd P, Burkinshaw L, Watten
P, Zammit C, Harris PR, Good A and Jenkins V: A user-centred
approach to developing bWell, a mobile app for arm and shoulder
exercises after breast cancer treatment. J Cancer Surviv.
11:732–742. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Di Stadio CS, Altieri F, Minopoli G,
Miselli G, Rippa E and Arcari P: Role of human GKN1 on APP
processing in gastric cancer. Biochimie. 135:149–153. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsang JYS, Lee MA, Ni YB, Chan SK, Cheung
SY, Chan WW, Lau KF and Tse GMK: Amyloid precursor protein is
associated with aggressive behavior in nonluminal breast cancers.
Oncologist. 23:1273–1281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Takagi K, Ito S, Miyazaki T, Miki Y,
Shibahara Y, Ishida T, Watanabe M, Inoue S, Sasano H and Suzuki T:
Amyloid precursor protein in human breast cancer: An
androgen-induced gene associated with cell proliferation. Cancer
Sci. 104:1532–1538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mao Y, Zhang Y, Fan S, Chen L, Tang L,
Chen X and Lyu J: GALNT6 promotes tumorigenicity and metastasis of
breast cancer cell via β-catenin/MUC1-C signaling pathway. Int J
Biol Sci. 15:169–182. 2019. View Article : Google Scholar :
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
38
|
Schwartz RS and Erban JK: Timing of
metastasis in breast cancer. N Engl J Med. 376:2486–2488. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peart O: Metastatic breast cancer. Radiol
Technol. 88:519M–539M. 2017.PubMed/NCBI
|
|
40
|
Pandey P, Sliker B, Peters HL, Tuli A,
Herskovitz J, Smits K, Purohit A, Singh RK, Dong J, Batra SK, et
al: Amyloid precursor protein and amyloid precursor-like protein 2
in cancer. Oncotarget. 7:19430–19444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Golde TE, Cai XD, Shoji M and Younkin SG:
Production of amyloid beta protein from normal amyloid beta-protein
precursor (beta APP) and the mutated beta APPS linked to familial
Alzheimer's disease. Ann N Y Acad Sci. 695:103–108. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao L, He D, Jiao M, Kong L, Shao C, Chen
J, Fang Z, Ma X, Chen H, Li L, et al: Overexpression of histone
deacetylase and amyloid precursor protein in hepatocellular
carcinoma. Technol Cancer Res Treat. 16:586–594. 2017. View Article : Google Scholar :
|
|
43
|
Sobol A, Galluzzo P, Liang S, Rambo B,
Skucha S, Weber MJ, Alani S and Bocchetta M: Amyloid precursor
protein (APP) affects global protein synthesis in dividing human
cells. J Cell Physiol. 230:1064–1074. 2015. View Article : Google Scholar :
|
|
44
|
Miyazaki T, Ikeda K, Horie-Inoue K and
Inoue S: Amyloid precursor protein regulates migration and
metalloproteinase gene expression in prostate cancer cells. Biochem
Biophys Res Commun. 452:828–833. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu Y, Lv F, Liang D, Yang Q, Zhang B, Lin
H, Wang X, Qian G, Xu J and You W: HOTAIR may regulate
proliferation, apoptosis, migration and invasion of MCF-7 cells
through regulating the P53/Akt/JNK signaling pathway. Biomed
Pharmacother. 90:555–561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou J, Zhang WW, Peng F, Sun JY, He ZY
and Wu SG: Downregulation of hsa_circ_0011946 suppresses the
migration and invasion of the breast cancer cell line MCF-7 by
targeting RFC3. Cancer Manag Res. 10:535–544. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shi C, Zhu X, Wang J and Long D: Estrogen
receptor α promotes non-amyloidogenic processing of platelet
amyloid precursor protein via the MAPK/ERK pathway. J Steroid
Biochem Mol Biol. 144(Pt B): 280–285. 2014. View Article : Google Scholar
|
|
48
|
Zhao XW, Zhou JP, Bi YL, Wang JY, Yu R,
Deng C, Wang WK, Li XZ, Huang R, Zhang J and Tao DT: The role of
MAPK signaling pathway in formation of EMT in oral squamous
carcinoma cells induced by TNF-α. Mol Biol Rep. 46:3149–3156. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kumar K, Chow CR, Ebine K, Arslan AD, Kwok
B, Bentrem DJ, Eckerdt FD, Platanias LC and Munshi HG: Differential
regulation of ZEB1 and EMT by MAPK-interacting protein kinases
(MNK) and eIF4E in pancreatic cancer. Mol Cancer Res. 14:216–227.
2016. View Article : Google Scholar
|