Introduction
In recent years, the incidence and mortality rates
of patients with coronary heart disease (CHD) have increased
annually, and it is one of the primary causes of mortality of
hospitalized patients (1,2). Coronary artery bypass graft (CABG)
is one of the primary means of treating patients with CHD (3,4).
At present, the great saphenous veins are the most frequently used
vessel for graft material for CABG. Although significant
improvements have been made in surgical techniques and surgical
instruments, vein graft failure (VGF) remains a frequent outcome in
patients who have undergone CABG, and seriously limits the
effectiveness of surgery and consequent prognosis (5,6).
Studies analyzing CABG operations over a 10-year period have
indicated that the incidence of VGF is >50%. Of these failures,
>90% are acute VGF that occurred within 1 month following
surgery and were the result of an acute thrombosis (7-9).
Regular anticoagulation therapy may decrease the incidence of acute
VGF. However, >60% of chronic VGF occurring in a 10 year period
following CABG was due to the proliferation and migration of smooth
muscle cells (SMCs) (10,11). Furthermore, the proliferation and
migration of SMCs may mediate platelet adhesion and atheromatous
plaque formation, which may aggravate VGF (12). However, the mechanism of SMCs
proliferation and migration in VGF remains unclear. There are no
effective therapeutic means of inhibiting the proliferation and
migration of SMCs. Therefore, determining the mechanism of the
proliferation and migration of SMCs in VGF has been the subject of
increased attention in cardiac surgery research.
MicroRNA (miRNA) are a class of non-coding RNAs, ~22
nucleotides in length, with regulatory functions that serve a
pivotal role in the regulation of developmental timing, cell
proliferation and migration. Continuous studies investigating the
functions of miRNAs have demonstrated the association between miRNA
and cardiovascular diseases (13,14). A number of studies have indicated
that miRNAs regulate the proliferation and migration of vascular
endothelial cells (ECs) and SMCs (15-18). Inhibition of vascular SMC is an
effective method of preventing VGF (19,20). The plasma expression level of
miR-423 was upregulated in patients with CHD following CABG
(21). Previous studies revealed
abnormal expression of miR-423 in heart disease (22-24). In addition, miR-423 has been
reported to be involved in regulating the development of various
vascular diseases, including atherosclerosis (25), myocardial infarction (26), diabetic vascular complications
(27) and lymphoma (28). However, the role of miR-423 in
regulating the proliferation and migration of ECs and SMCs to
mediate VGF remains unclear.
A disintegrin and metalloproteinase with
thrombospondin motifs-7 (ADAMTS-7), the 7th member of the ADAMTS
family to be identified, serves a vital role in multiple biological
processes, including embryogenesis, vasculogenesis and blood
coagulation. For example, increased expression of ADAMTS-7 enhanced
the proliferation and migration capacity of SMCs (29). Upregulation of ADAMTS-7 promoted
atherosclerosis by regulating degradation and remodeling of the
vascular matrix (30). Wang et
al (31) suggested that
ADAMTS-7 overexpression accelerated the progression of carotid
artery injury in rats by promoting the proliferation and migration
of SMCs. Furthermore, ADAMTS-7 is involved in intima hyperplasia
following vascular injury (32).
However, the underlying mechanism is not yet understood. Therefore,
the importance of ADAMTS-7 in vein graft restenosis requires
additional investigation.
In the present study, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
used to measure the expression levels of miR-423 in plasma, and in
human umbilical vein endothelial cells (HUVECs) and human umbilical
vein smooth muscle cells (HUVSMCs), and the association between
miR-423 and ADAMTS-7 was assessed. A potential interaction between
miR-423 and ADAMTS-7 was hypothesized to regulate the proliferation
and migration of HUVSMCs and HUVECs in vitro. Furthermore,
the role of miR-423 overexpression in a rat vein graft model was
examined in vivo.
Materials and methods
Ethics statement
The present study obtained approval from The Ethics
Committee of The First People's Hospital of Yunnan Province
(approval no. 2017YYLH018) and complied with the guidelines and
principles of the Declaration of Helsinki. All participants
provided written informed consent. Experiments involving animals
were conducted according to the guidelines of the Animal Care and
Use Committees at The First People's Hospital of Yunnan
Province.
Plasma specimens
For the clinical part of the present study, 15
patients (age, 62.279.04 years; 9 males and 6 females) who were
preparing to undergo CABG surgery and 10 healthy volunteers (age,
58.6±4.48 years; 5 males and 5 females) from urban and rural areas
were recruited. The patients were recruited at the Department of
Cardiovascular Surgery of The First People's Hospital of Yunnan
Province between December 2017 to March 2018. The healthy
volunteers were recruited at the medical examination center of The
First People's Hospital of Yunnan Province between December 2017 to
March 2018. Peripheral blood samples (2 ml) were collected from the
patients into EDTA tubes 1 day prior to and 1, 5, 10 and 20 days
following surgery in both patients and healthy volunteers. Samples
were frozen in liquid nitrogen immediately and stored at −80°C for
subsequent experiments.
Cell culture
HUVECs and HUVSMCs were obtained from the BeNa
Culture Collection and 293T cells were purchased from the American
Type Culture Collection. Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.) containing 1%
penicillin-streptomycin and 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C and 5% CO2.
Cell transfection
A total of 24 h prior to transfection, HUVECs and
HUVSMCs were seeded in 6-well plates at a density of
2×105 cells/well and incubated overnight. Subentry, 50
nM miR-423 mimic, 100 nM miR-423 inhibitor and 50 nM negative
control were transfected into HUVECs and HUVSMCs using
Lipofectamine™ 3000 reagent and Opti-MEM medium (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols,
and incubated for 4 h. The mixed solution was then replaced with
complete growth medium and cultured for 24 h. The miR-423 mimics,
inhibitor, pcDNA3.1 ADAMTS-7, and control (blank plasmid) were
purchased from Tolo Biotech Co., Ltd. The Sequences corresponding
to miR-423 mimic were: 5′-GCCTGAGGGGCAGAGAGC-3′, miR-423 inhibitor,
5′-ATCTTTGGTGGCCGTAGACCT-3′, and scrambled negative control,
5′-GCCTAACTGTGTCAGAAGGAA-3′.
RT-qPCR
Total RNA was isolated from cultured plasma and
cells using TRIzol® reagent (Qiagen GmbH) and
revere-transcribed into cDNA using the PrimeScript™ RT reagent kit
using a gDNA Eraser (Takara Bio, Inc.). RT-qPCR Master mix was
purchased from (Takara Bio, Inc.). The sequence of the primers for
qPCR are presented in Table I. U6
was used as an internal control. The 2−∆∆Cq method
(33) was used to calculate the
relative expression of miR-423. qPCR was performed in
triplicate.
 | Table IName and sequences of the
primers. |
Table I
Name and sequences of the
primers.
| Name | Primer
sequences |
|---|
| miR-423 | F:
5′-GCCTGAGGGGCAGAGAGC-3′ |
| R:
5′-CCACGTGTCGTGGAGTC-3′ |
| U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
| R:
5′-AACGCTTCACGAATTTGCGT-3′ |
| ADAMTS-7 | F:
5′-GTCATCGACTTCCCTTCCATAC-3′ |
| R:
5′-TGTCCATGTCATCGCAGAAG-3′ |
| GAPDH | F:
5′-ATGCCTCCTGCACCACCA-3′ |
| R:
5′-AGTCCCTCCACGATGCCAA-3′ |
CCK-8 assay
Cell Counting Kit-8 (Sigma-Aldrich; Merck KGaA) was
used to assess the proliferation rates of the HUVECs and HUVSMCs.
Cells were seeded in 96-well plates at a density of
1×105 cells per well and cultured with 5% CO2
at 37°C for 2 h to allow cells to adhere. A total of 10 µl
CCK-8 solution was added to each well and mixed, and cells were
incubated for an additional 2 h at 37°C. A dual-wavelength
microplate reader was used to measure proliferation at 450 nm
(Beckman Coulter, Inc.). Assays were performed in triplicate.
Transwell migration assay
A total of 2×105 cells/ml HUVECs or
HUVSMCs were plated in 200 µl serum-free medium in the upper
layer of the Transwell chambers (Corning, Inc.) and 800 µl
medium supplemented with 10% FBS was added to the bottom chamber.
After incubation for 24 h at 37°C, the cells that had migrated were
fixed with 4% paraformaldehyde at 37°C, rinsed three times with
PBS, stained with 0.1% crystal violet for 10 min at 37°C and rinsed
three times with PBS. The images of migrated cells were taken using
a light microscope (Olympus Corporation). For quantification, 5
randomly selected fields were analyzed at magnification, ×40.
miR-423 target prediction
TargetScan 7.2 (http://www.Targetscan.org) was performed to predict
the target genes of miR-423 according to the previous studies
(34-37).
Dual-luciferase reporter gene assay
The 3′-UTR fragments of ADAMTS-7 containing the
wild-type (WT) miR-423-binding site or a mutant (MUT)
miR-423-binding site were amplified by Shanghai GenePharma Co.,
Ltd. and cloned into the pmirGLO luciferase reporter gene vector
(Promega Corporation). The generated luciferase reporter plasmids
were designated as WT-ADAMTS-7-3′UTR and MUT-ADAMTS-7-3′UTR.
WT-ADAMTS-7-3′UTR or MUT-ADAMTS-7-3′UTR were co-transfected with
miR-423 mimics (50 nM) or mimics control (50 nM) into 293T cells
using Lipofectamine® 3000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). After 48 h of incubation at 37°C, the
activity of luciferase was determined using a dual-luciferase
reporter assay system (cat. no. PR-E1960; Promega Corporation). The
firefly lucif-erase activity was normalized to Renilla
luciferase activity.
Western blot analysis
Total protein was extracted for western blot
analysis. Briefly, total proteins were extracted from tissues
samples or cells using RIPA lysis buffer (Beijing Solarbio Science
& Technology Co., Ltd) and protein concentration was determined
using a BCA Protein assay kit (Thermo Fisher Scientific, Inc.).
Protein (40 µg/lane) from each sample was separated by 12%
SDS-PAGE and transferred onto PVDF membranes (Sigma-Aldrich; Merck
KGaA). The membranes were blocked at 37°C for 2 h with 5% fat-free
milk diluted with TBS containing 0.1% Tween-20. Subsequently, the
PVDF membranes were incubated overnight at 4°C with the primary
antibodies against ADAMTS-7 (cat. no. ab203027; 1:1,000; Abcam),
α-smooth muscle actin (α-SMA; cat. no. ab32575; 1:1,000; Abcam),
matrix metalloproteinase (MMP2; cat. no. ab215986; 1:1,000; Abcam),
MMP9 (cat. no. ab219372; 1:1,000; Abcam) and proliferating cell
nuclear antigen (PCNA; cat. no. ab92552; 1:1,000; Abcam). After
washing, the membranes were incubated for 1 h at 37°C with
horseradish peroxidase-conjugated secondary antibodies (cat. nos.
ab6721 and ab6728; 1:500; Abcam) at 37°C for 2 h. The signal was
visualized using enhanced chemiluminescence reagent (Bio-Rad
Laboratories, Inc.) according to the manufacturer's protocol.
Densitometric analysis was performed using ImageJ 1.8.0 software
(National Institute of Health). The expression levels of protein in
each sample were normalized to GAPDH.
Immunohistochemistry assay
Vessel graft wall tissues were collected and fixed
in 4% paraformaldehyde for 48 h at 37°C. Subsequently, tissues were
embedded in paraffin, and 4 µm-thick slices were dewaxed and
rehydrated in a graded series of ethanol solutions (50, 75, 85, 95
and 100%; 5 min per solution). Xylene and a graded series of
ethanol (100, 95, 85, and 75%) were used to dewax and hydrate the
samples, followed by 30 min of antigen retrieval in Tris-EDTA (pH
9.0) in a 720 W microwave. Subsequently, a DAB horseradish
peroxidase color development kit (Beyotime Institute of
Biotechnology) with a Ki-67 antibody (cat. no. ab15580; 1:1,000;
Abcam) was used to stain the samples at room temperature for 15
min. The, slides were then dyed with hematoxylin for 30 sec at
37°C, dehydrated and fixed, and sealed with neutral glue. Stained
images were observed and photographed under a fluorescence
microscope (Olympus Corporation) at magnification, ×400.
Hematoxylin and eosin (H&E) staining
and Masson staining
Whole vessel graft lumen tissues obtained from the
rat vein graft models were dissected and fixed with 4%
paraformaldehyde for 24 h at room temperature. Subsequently,
samples were dehydrated and embedded in paraffin blocks. Blocks of
6-µm thickness were cut and stained at room temperature
using hematoxylin for 20 min and eosin for 5 min to measure the
neointimal thickness. Meanwhile, sections of rat vein grafts were
also stained with Masson's trichrome for the evaluation of collagen
expression. Masson's trichrome kit (cat. no. Mst-8003/8004; Fuzhou
Maixin Biotech Co., Ltd.) was used at room temperature according to
the manufacturer's protocol, and observed under a light microscope
(magnification, ×200; Olympus Corporation).
Rat vein graft models
A total of 30 male Sprague-Dawley rats (8-9
week-old; weight, 250-300 g) were purchased from the Experimental
Animal Center of Kunming Medical University. All animals were
housed in an animal facility with a 12:12 h light: Dark cycle at
18-22°C and 40-60% humidity, and had ad libitum access to
rodent chow and water. Subsequently, all rats were anesthetized
with 300 mg/kg chloral hydrate by intraperitoneal injection and
systemically heparinized, and no rats exhibited signs of
peritonitis following the administration of chloral hydrate. Next,
all rats were randomly separated into three groups, with 10 rats
per group. A right sternocleidomastoid incision was performed in
all the groups. Subsequently, exposure of the right jugular vein of
a sufficient size was performed and a section ~2 cm was trimmed for
vessel graft. Then, the vessel graft to the right carotid artery
was replaced. During surgery, 8 nmol miR-423 agomir dissolved in
normal saline (0.9%) according to the manufacturer's instructions,
was perfused into the vein graft under a distending pressure of 20
mmHg for 10 min at room temperature prior to the arteriovenous
anastomosis. Rats in the sham group were injected with the same
volume of saline at the same location. Rats in all groups were
sacrificed immediately, and the jugular vein was harvested and
further analyzed.
Statistical analysis
All statistical analyses were performed using SPSS
v.22.0 software (IBM Corp.). Data are presented as the mean ±
standard deviation. Differences between multiple groups were
analyzed using a one-way analysis of variance followed by Tukey's
post hoc test. A Student's t-test was used to analyze differences
between two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of miR-423 in patients with
CHD following CABG surgery
To investigate the effects of miR-423 on autologous
vein graft restenosis, the clinicopathological characteristics were
compared. There were no significant differences in sex, age,
smoking habits, drinking habits, family history, diabetes mellitus
and hypertension between the patients who underwent CABG and
healthy controls (Table II).
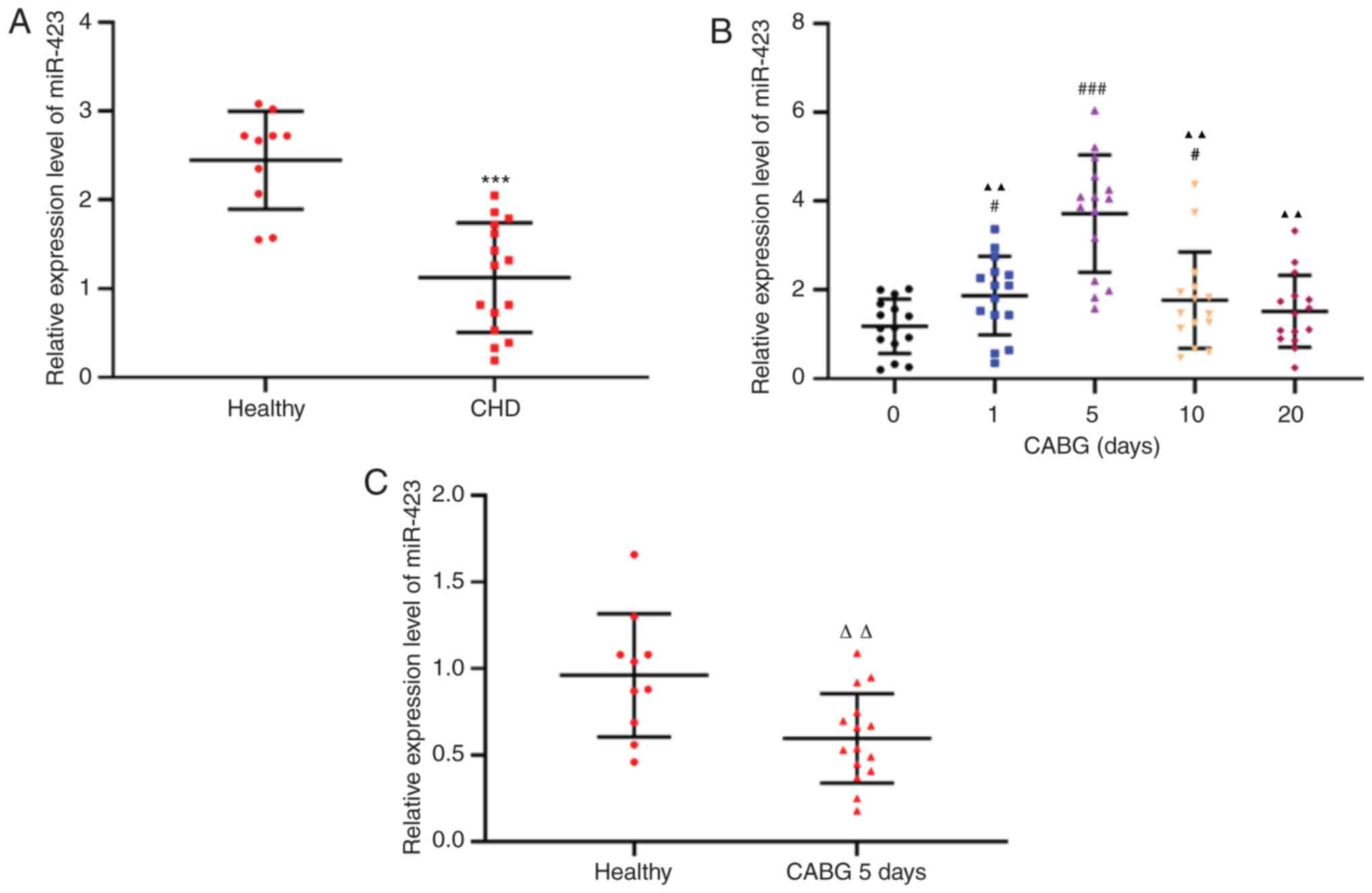
RT-qPCR was used to detect miR-423 expression levels in the plasma
of patients with CHD (n=15) and healthy controls (n=10). The
expression levels of miR-423 in the plasma of patients with CHD
were decreased compared with the healthy subjects (P<0.001;
Fig. 1A). The expression levels
of plasma miR-423 in all samples from patients with CHD 1 day prior
to CABG surgery and 1, 5, 10 and 20 days following CABG surgery
were decreased by the 10th day following CABG and returned to the
preoperative levels 20 days after CABG (Fig. 1B). In addition, even the maximum
expression levels of miR-423 in patients with CHD following CABG
were decreased compared with those in the healthy subjects
(P<0.01; Fig. 1C). Therefore,
an abnormal increase in miR-423 expression in patients with CHD
prior to and following CABG may be involved in the protection of
transplanted blood vessels in surgery, although the mechanism
remains unclear.
| Table IIClinicopathological
characteristics. |
Table II
Clinicopathological
characteristics.
|
Characteristics | Healthy people
(n=10) | CABG patients
(n=15) | P-value |
|---|
| Sex | | | |
| Male, n (%) | 5 (50) | 9 (60) | 0.5836 |
| Female, n (%) | 5 (50) | 6 (40) | 0.6124 |
| Age, years
(range) | 58.6±4.48
(50-65) | 62.27±9.04
(45-72) | 0.1293 |
| Drinking, n
(%) | 4 (40) | 7 (46.67) | 0.6502 |
| Smoking, n (%) | 4 (40) | 6 (40) | 0.7954 |
| Family history, n
(%) | 3 (30) | 5 (30) | 0.8135 |
| Diabetes mellitus,
n (%) | 2 (20) | 3 (20) | 0.7106 |
| Hypertension, n
(%) | 2 (20) | 4 (26.67%) | 0.6341 |
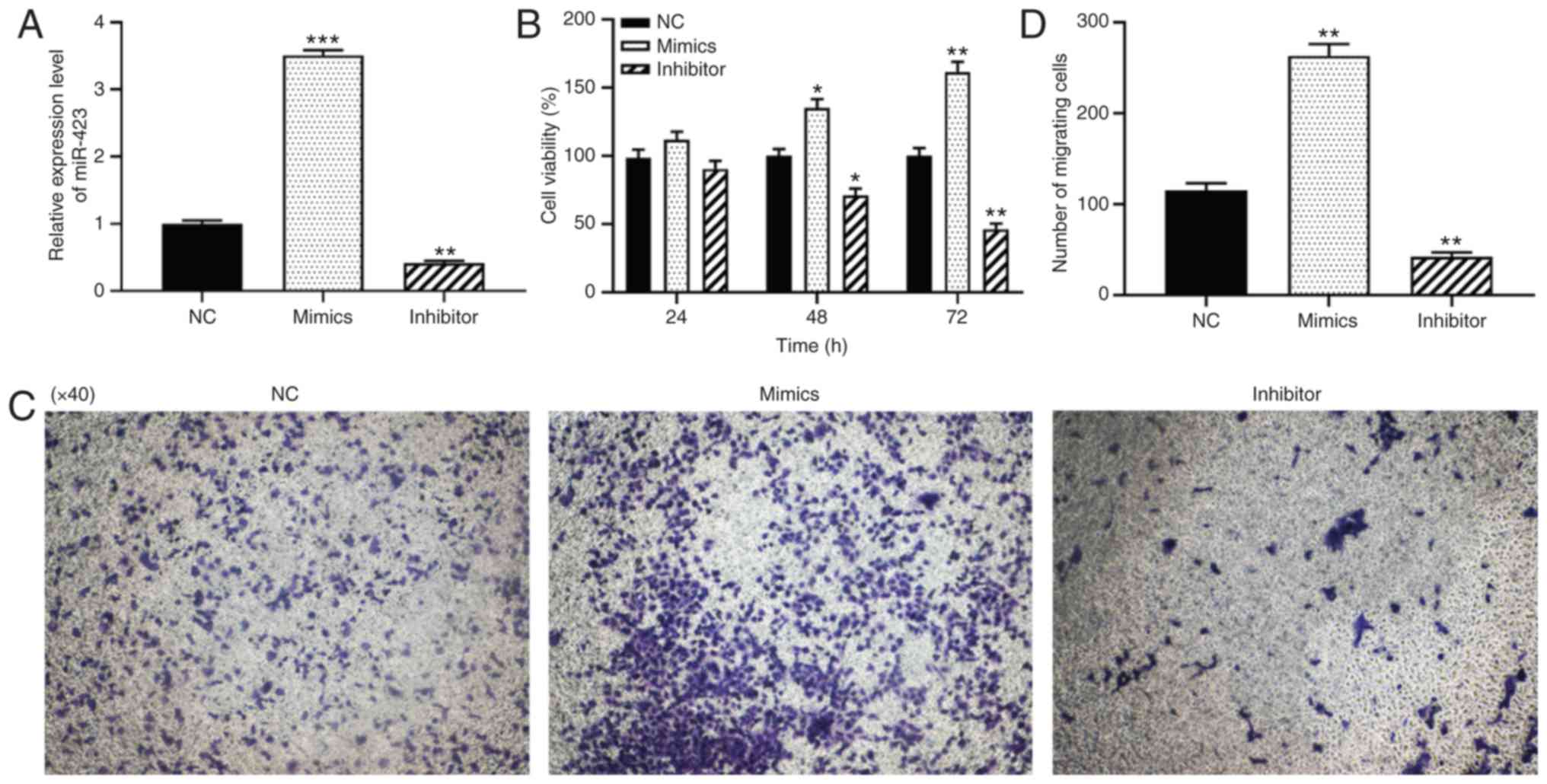
Effect of miR-423 on the proliferation
and migration of HUVECs
To further verify the function of miR-423 in
patients with CHD following CABG surgery, the effects of miR-423 on
the proliferation and migration levels of HUVECs were examined
using CCK-8 and Transwell assays. HUVECs were transfected with
miR-423 mimics or inhibitor to alter the expression of miR-423.
RT-qPCR analysis indicated that, compared with the control group,
the expression levels of miR-423 were significantly increased in
cells transfected with miR-423 mimics (P<0.001; Fig. 2A) and significantly decreased in
cells transfected with miR-423 inhibitor (P<0.01; Fig. 2A). The CCK-8 assay results
indicated that the over-expression of miR-423 notably promoted cell
viability in HUVECs at 48 (P<0.05) and 72 h (P<0.01) compared
with the NC group (Fig. 2B). Data
from the Transwell migration assays demonstrated that the
upregulation of miR-423 markedly increased the migration of HUVECs
compared with the NC group. (P<0.01; Fig. 2C and D), and knockdown of miR-423
resulted in the opposite effects on proliferation and migration
(P<0.01). Taken together, the overexpression of miR-423 was
demonstrated to increase the proliferation and migration levels of
HUVECs.
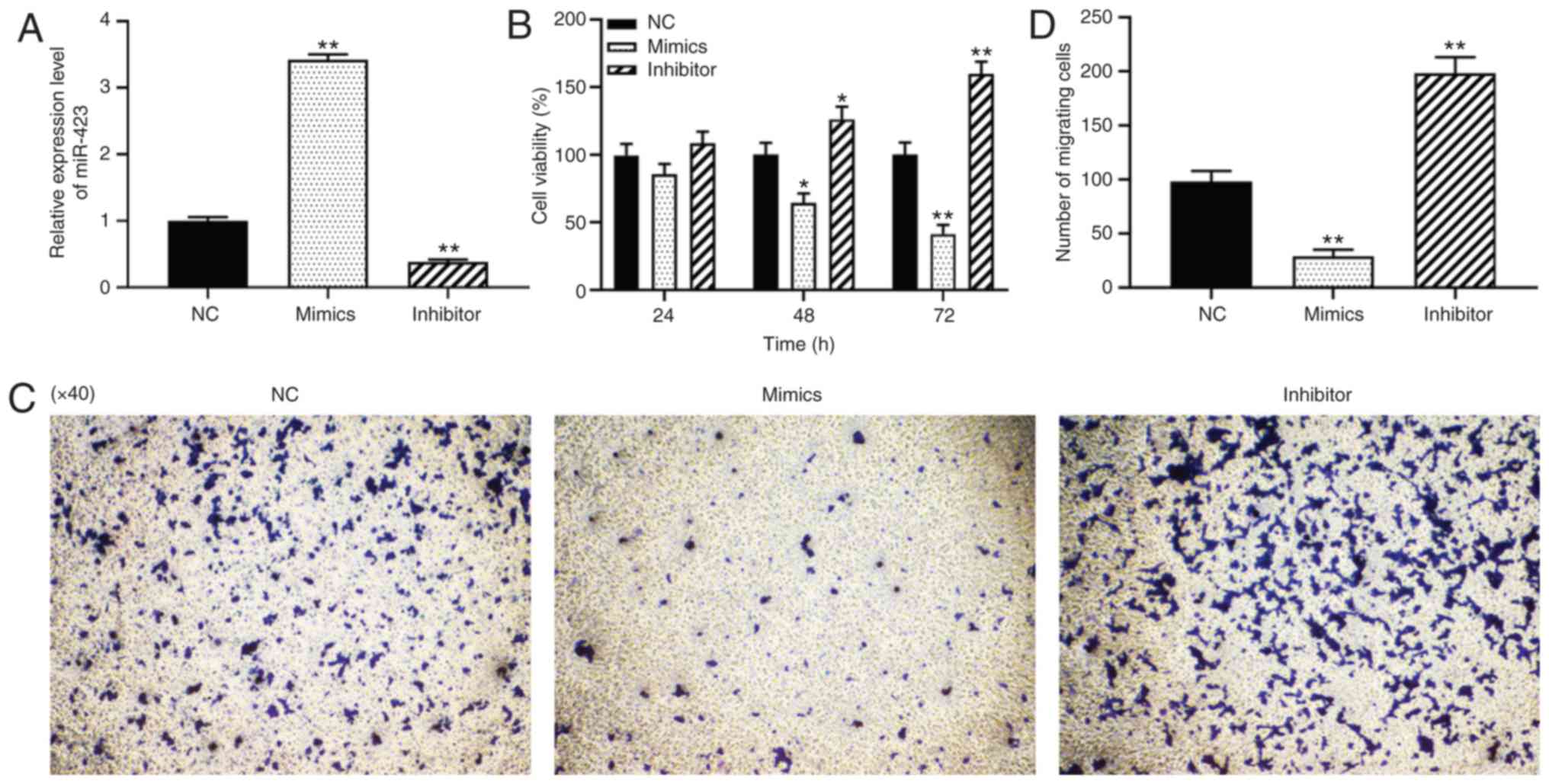
Effect of miR-423 on the proliferation
and migration of HUVSMCs
To investigate the loss- and gain-of-function of
miR-423 in vascular SMCs of vein grafts, miR-423 mimics or
inhibitors were transfected into HUVSMCs, and the results are
presented in Fig. 3A. CCK-8 and
Transwell migration assays were used to detect the effects of
miR-423 on the proliferation and migration of HUVSMCs. As indicated
in Fig. 3B-D, knockdown of
miR-423 significantly promoted the proliferation at 48 (P<0.05)
and 72 h (P<0.01; Fig. 3B) and
migration capacity of HUVSMCs (P<0.01; Fig. 3C and D), whereas overexpression of
miR-423 resulted in the opposite effect (P<0.01). These results
suggest that miR-423 overexpression significantly inhibited the
proliferation and migration of HUVSMCs in vitro.
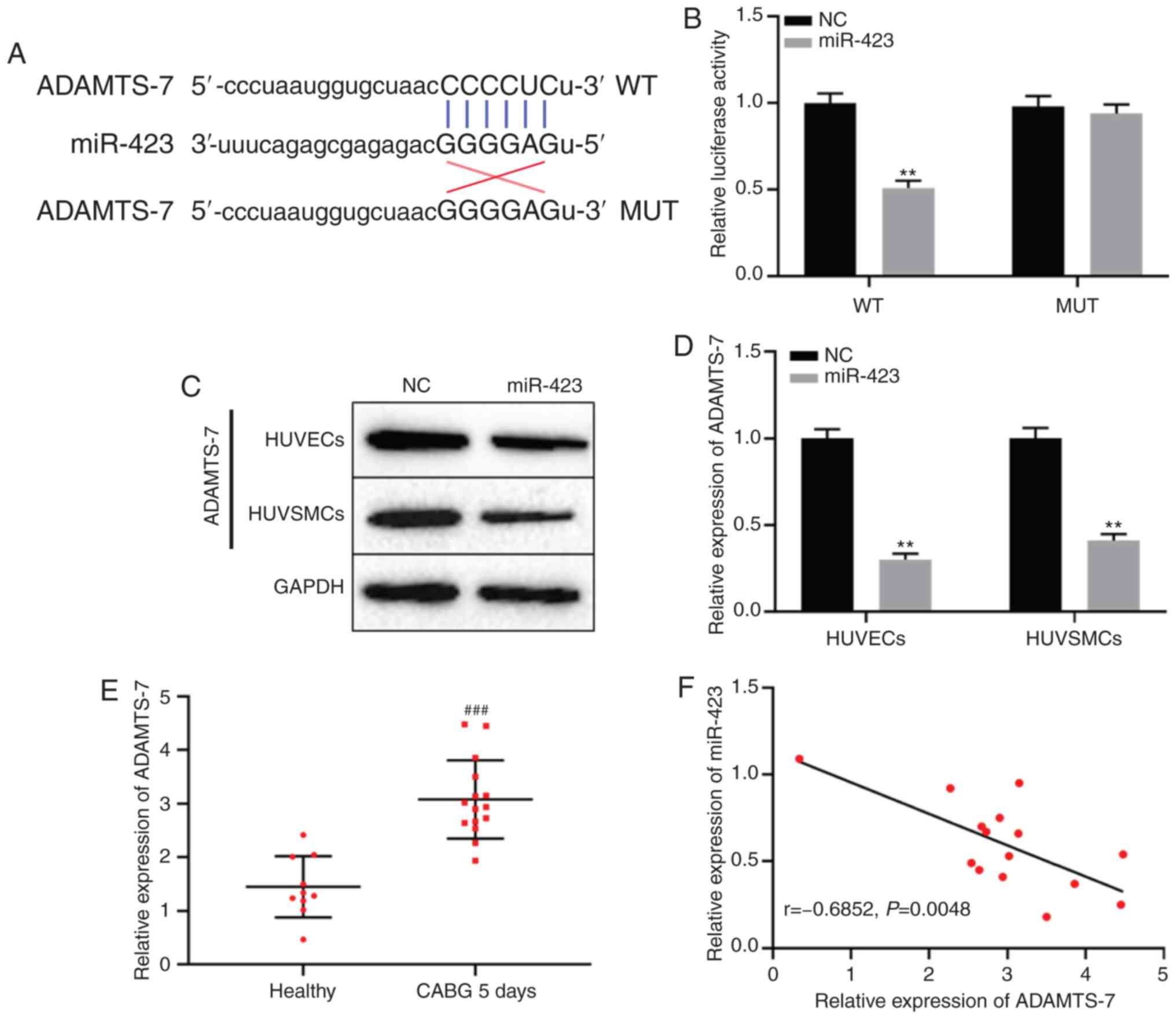
miR-423 directly targets ADAMTS-7
TargetScan database analysis indicated that miR-423
may target ADAMTS-7 directly (Fig.
4A). To determine whether miR-423 specifically bound to the
3′-untranslated region of ADAMTS-7 mRNA and regulated the
expression of ADAMTS-7, a dual-luciferase reporter gene assay was
used. The results demonstrated that luciferase activity in the
ADAMTS-7-WT + miR-423 mimics group was decreased compared with the
ADAMTS-7-WT + miR-NC group, but there was no significant difference
between miR-423 mimics or NC combined with ADAMTS-7-MUT group
(P<0.01; Fig. 4B). Protein
expression levels of ADAMTS-7 in cells transfected with miR-423
mimic compared with the NC group (P<0.01; Fig. 4C and D). Additionally, the
expression levels of ADAMTS-7 were significantly increased in
patients with CHD following CABG after 5 days compared with the
healthy subjects (P<0.001; Fig.
4E). Spearman's correlation analysis results revealed that
there was a remarkable negative correlation between the expression
of miR-423 and ADAMTS-7 in patients with CHD following CABG after 5
days (r=−0.685; P<0.01; Fig.
4F). These results suggest that ADAMTS-7 was directly targeted
by miR-423, and negatively regulated ADAMTS-7 expression.
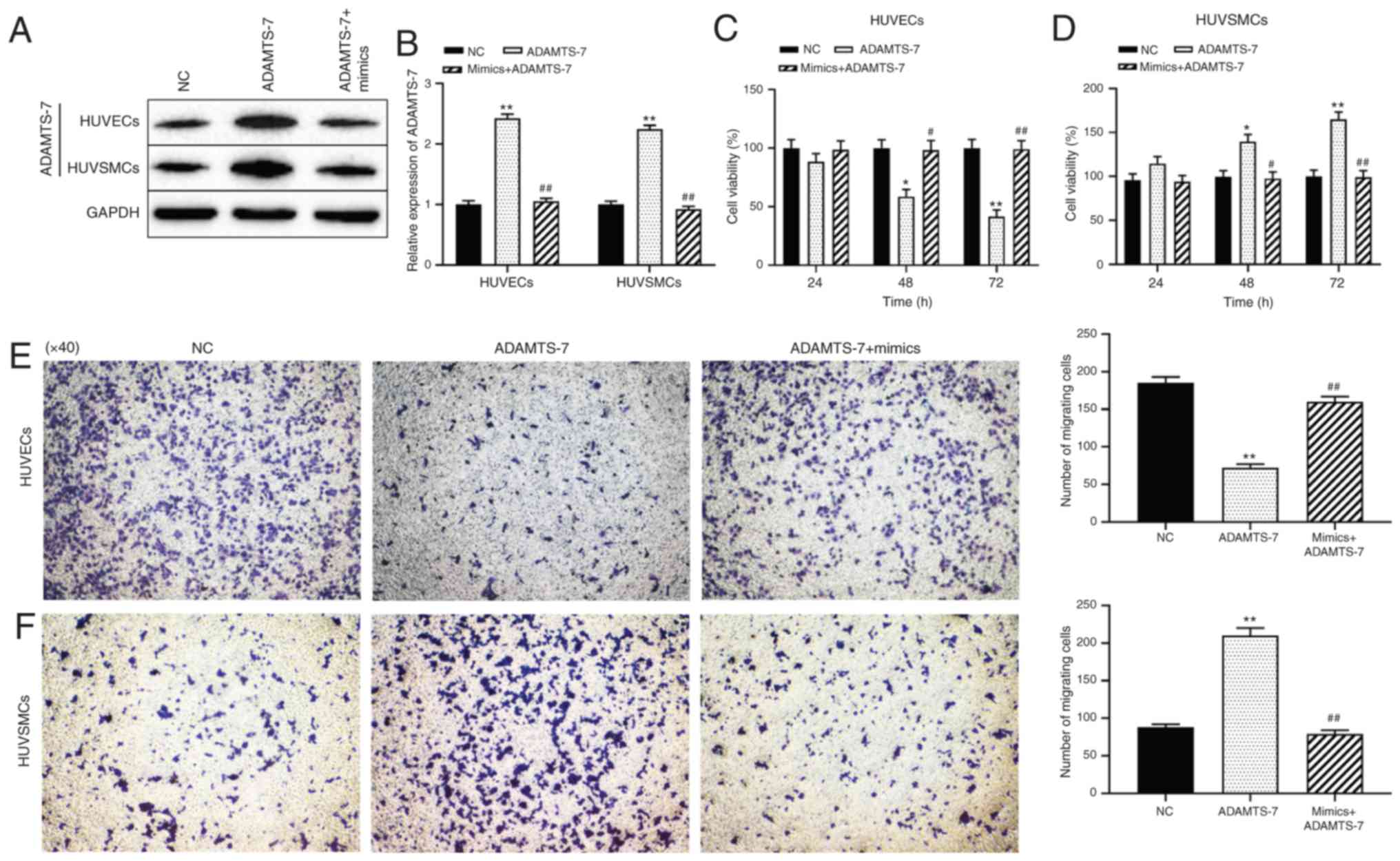
Effect miR-423/ADAMTS-7 axis on the
proliferation and migration of HUVECs and HUVSMCs
The mechanism by which miR-423 modulated cell
proliferation and migration of HUVECs and HUVSMCs was then
assessed. Western blot analysis demonstrated that transfection of
ADAMTS-7 was successful and protein expression levels were
increased, and that the increase in expression was reversed in
cells co-transfected with miR-423 mimics + pcDNA-ADAMTS-7
(P<0.01; Fig. 5A and B).
Compared with the NC group, overexpression of ADAMTS-7
significantly decreased the proliferation and migration ability of
HUVECs at 48 (P<0.05) and 72 h (P<0.01; Fig. 5C and E), but promoted the
proliferation and migration of HUVSMCs at 48 (P<0.05) and 72 h
(P<0.01; Fig. 5D and F). The
effect of increased ADAMTS-7 levels on proliferation and migration
of HUVSMCs or HUVECs was reversed by transfection with miR-423
mimics. These results suggest that upregulation of miR-423
decreases ADAMTS-7 expression, thereby promoting the proliferation
and migration of HUVECs and decreasing these behaviors in HUVSMCs
in vitro.
Effect of miR-423 upregulated on
autologous vein graft restenosis in vivo
Based on the effect of miR-423 on the proliferation
and migration in HUVSMCs or HUVECs, the function of miR-423 in the
distension on vein graft restenosis in vivo was determined.
Hematoxylin and eosin staining indicated that overexpression of
miR-423 markedly decreased neointimal thickness compared with the
vein graft group (Fig. 6A).
However, Masson staining results revealed that the percentage of
the neointima area occupied by blue-colored fibers (collagen) was
significantly different between the miR-423 agomir-treated group
and the Sham group or Vein graft group (Fig. 6B). Immunohistochemistry staining
demonstrated that increased levels of miR-423 decreased the
expression of Ki-67 in carotid arteries compared with the vein
graft group (Fig. 6C).
Furthermore, western blot analysis was applied to detect the effect
of miR-423 overexpression on the proliferation of vascular SMCs.
Upregulation of miR-423 decreased the expression vein graft-induced
ADAMTS-7, PCNA, MMP2, MMP9 and α-SMA (P<0.01; Fig. 6D). Expression of miR-423 was
upregulated in the miR-423 agomir group compared with the vein
graft group or the Sham group (P<0.001; Fig. 6E). These results suggest that
overexpression of miR-423 may attenuate autologous vein graft
restenosis via targeting ADAMTS-7 in vivo.
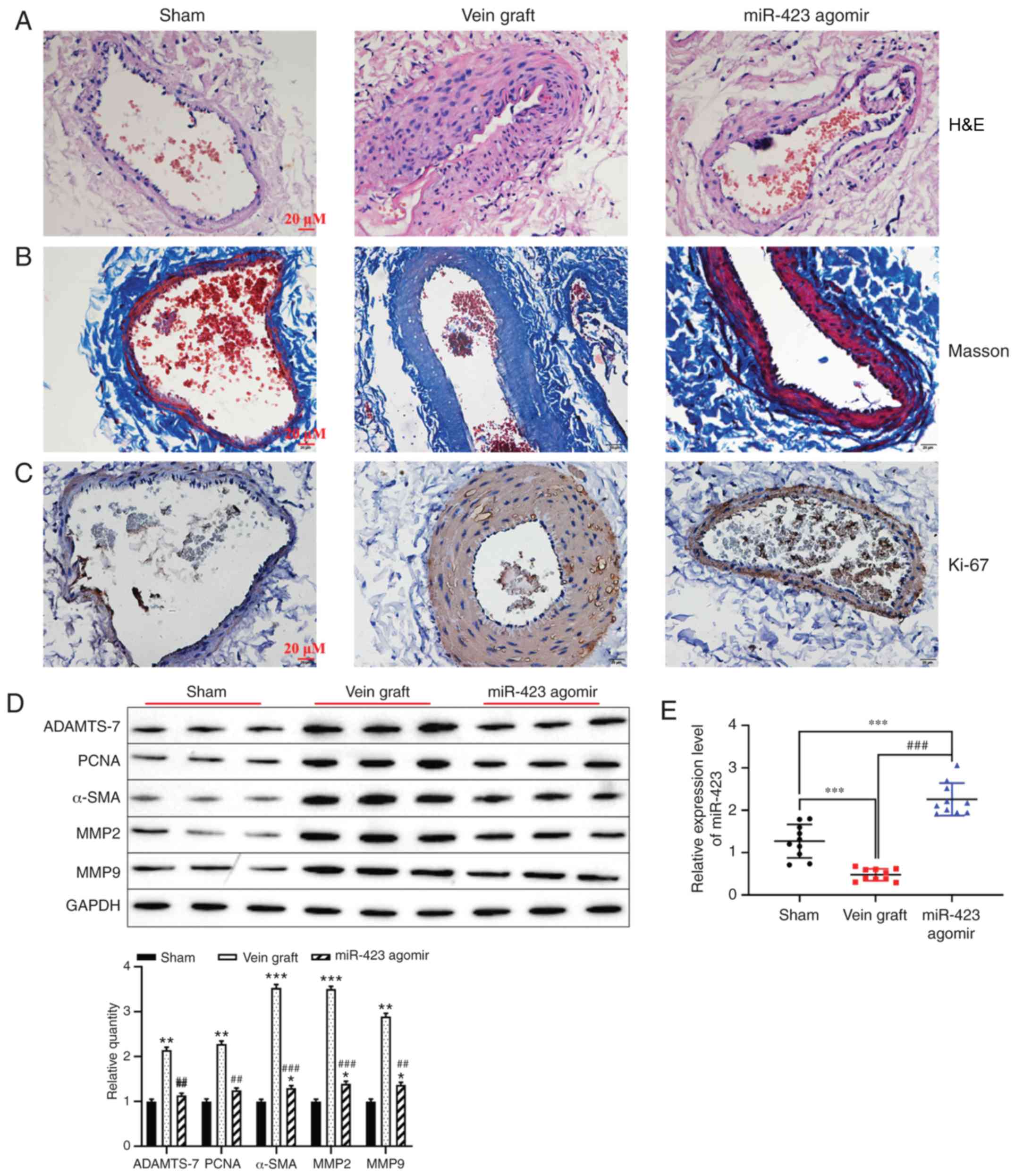 | Figure 6Effect of miR-423 on autologous vein
graft restenosis in vivo. (A) Effect of miR-423
overexpression on the thickening of vascular intima in vivo
was evaluated using H&E staining. Images are at magnification,
×20. (B) Masson-trichrome staining was applied to evaluate the
percentage of collagen in the neointima area. Images are at
magnification, ×20. (C) Proliferation of vascular cells, visualized
using immunostaining. Images are at magnification, ×20. (D)
Expression of ADAMTS-7, PCNA, α-SMA, MMP2, MMP9 were detected using
western blot analysis. (E) Expression of miR-423 was detected using
reverse transcription-quantitative polymerase chain reaction.
*P<0.05, **P<0.01 and
***P<0.001 vs. Sham group. ##P<0.01 and
###P<0.001 vs. vein graft group. ADAMTS-7, a
disintegrin and metalloproteinase with thrombospondin motifs-7;
PCNA, proliferating cell nuclear antigen; α-SMA, α-smooth muscle
actin; MMP, matrix metalloproteinase; miR, miRNA; H&E,
hematoxylin and eosin. |
Discussion
CHD is a common disease that affects the quality of
life patients and causes great economic burden to the patients and
the whole society (38). In the
present study, the role of the miR-423/ADAMTS-7 axis in the
proliferation and migration of HUVECs and HUVSMCs in vitro
and in vivo was examined. miR-423 was downregulated in the
plasma of patients with CHD compared with healthy volunteers.
Upregulation of miR-423 decreased the expression level of ADAMTS-7,
resulting in increases in the rates of proliferation and migration
of HUVECs, and accelerated the endothelialization of the artificial
vessel in vivo. In the past decade, numerous studies in the
field of miRNAs have provided evidence to support the role of
miRNAs in cardiovascular function and disease, and miRNAs have
exhibited significant clinical potential (39,40).
The biological behavior of HUVSMCs and HUVECs allows
them to serve an important role in restenosis and long-term
occlusion of vein bridges following coronary angioplasty (41-44). Increasing evidence has indicated
that the abnormal expression of miRNAs serves an important role in
transplanted vascular diseases and they have received much interest
in the research of various diseases (21,45-47). Proliferation, migration and
phenotypic transformation of SMCs or endothelial cells are
associated with various miRNAs (48) including miR-145 (49,50), miR-222 (51), miR-195 (52) and miR-21 (53). For example, Liu et al
(54) confirmed that the
upregulation of miR-378a targeted cyclin-dependent kinase 1 to
promote the proliferation and migration of vascular SMCs and
increase the incidence of in-stent restenosis following stenting.
Huang et al (55)
suggested that miR-22-3p overexpression inhibited proliferation and
migration of human artery vascular SMCs and prevented neointimal
hyperplasia by targeting high mobility group box-1. Endothelial
cells-derived miR-195 significantly suppressed the proliferation
and migration of vascular SMCs (52). Qu et al (56) identified that all patients with
severe CHD, who were preparing for CABG had a significantly
decreased level of miR-126-3p expression compared with the healthy
subjects. Taken together, inhibition of the biological function of
SMCs may be possible using specific drugs or biomolecules and this
may prevent graft vein failure. The results of the present study
revealed that the expression levels of miR-423 the plasma of
patients with CHD were decreased compared with those in the healthy
subjects, and that upregulation of miR-423 increased proliferation
and migration of HUVECs and had anti-proliferative and
anti-migratory effects on HUVSMCs. This result is in agreement with
previous studies indicating that the expression levels of miRNAs
related to the regulation of endothelial cell biological behavior
and vasculoprotective are significantly decreased in patients with
stable CHD (57).
Previous studies have demonstrated that ADAMTS-7 is
an important member of the depolymerized protein-like
metal-loproteinase family containing platelet binding protein
motifs, and its gene polymorphism is significantly associated with
the susceptibility to human coronary artery disease (30,58). Recently, ADAMTS-7 levels were
demonstrated to be associated with the severity of disease in
patients with CHD or saphenous vein grafts restenosis following
CABG (59,60). Furthermore, abnormally upregulated
expression levels of ADAMTS-7 promoted the proliferation and
migration of vascular SMCs and induced neointima formation in
vitro and in vivo (29,61). ADAMTS-7 promoted the proliferation
and migration of vascular SMCs by decomposing the extracellular
matrix, and thereby accelerating intimal hyperplasia, resulting in
restenosis (62,63). In addition, upregulation of
ADAMTS-7 decreased the proliferation and migration of endothelial
cells and restenosis following vascular injury by interrupting the
homeostasis between thrombospondin-1 and its natural inhibitor
cartilage oligomeric matrix protein (COMP) (31,32). In the present study,
overexpression of ADAMTS-7 significantly promoted the proliferation
and migration of HUVSMCs and induced neointima formation following
aorta injury. Knockdown of ADAMTS-7 significantly decreased the
expression of metalloproteinases such as MMP-2 and MMP-9. Previous
studies have indicated that MMPs contributed to the activation of
vascular SMCs proliferation (29,64). Overexpression of ADAMTS-7 degrades
matrix COMP in vessels (31), and
COMP decreases vascular SMCs migration by maintaining them in a
quiescent/contractile state (65). Based on the results of previous
studies, upregulation of ADAMTS-7 induces neointima formation
following injury to the aorta through promoting growth and
migration of vascular SMCs, perhaps as a result of degradation of
COMP. Furthermore, ADAMTS-7 was significantly highly expressed in
the neointima in vein grafts, and the expression level was markedly
decreased in vein grafts treated with miR-423 agomir.
In conclusion, the miR-423/ADAMTS-7 axis may be
involved in regulating the proliferation and migration of HUVSMCs
and HUVECs in vitro and in vivo, and may serve a
pivotal role in neointima formation in response to injury. The
present study may highlight a novel molecular target for treating
patients with CHD following CABG and new biomarkers for diagnosis
and prognosis.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Applied Basic
Research Project of Yunnan Provincial Science and Technology
Department, Kunming Medical University [Kunming, China; grant no.
2018FE001(−290)].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL conceived and designed the study. WR, LL, FW, and
NM performed the experiments and collected the data. WR, LZ, WH,
DX, and YC contributed to the analysis of the data and data
acquisition. HL contributed to the analysis of data and manuscript
preparation. WR and LL drafted the paper and revised manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study obtained approval from The Ethics
Committee of The First People's Hospital of Yunnan Province
(approval no. 2017YYLH018) and complied with the guidelines and
principles of the Declaration of Helsinki. All participants
provided written informed consent. Experiments involving animals
were conducted according to the guidelines of the Animal Care and
Use Committees at The First People's Hospital of Yunnan
Province.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang X, Li Y, Ren X, Xiong X, Wu L, Li J,
Wang J, Gao Y, Shang H and Xing Y: Effects of exercise-based
cardiac rehabilitation in patients after percutaneous coronary
intervention: A meta-analysis of randomized controlled trials. Sci
Rep. 7:447892017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anderson L, Oldridge N, Thompson DR,
Zwisler AD, Rees K, Martin N and Taylor RS: Exercise-based cardiac
rehabilitation for coronary heart disease: Cochrane systematic
review and meta-analysis. J Am Coll Cardiol. 67:1–12. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kaur K, Bedi G, Kaur M, Vij A and Kaur I:
Lipid peroxidation and the levels of antioxidant enzymes in
coronary artery disease. Indian J Clin Biochem. 23:33–37. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang X, He T, Han S, Zhang X, Sun Y, Xing
Y and Shang H: The role of traditional chinese medicine in the
regulation of oxidative stress in treating coronary heart disease.
Oxid Med Cell Longev. 2019:32314242019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hausenloy DJ, Boston-Griffiths E and
Yellon DM: Cardioprotection during cardiac surgery. Cardiovasc Res.
94:253–265. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hegewald J, Wegewitz UE, Euler U, van Dijk
JL, Adams J, Fishta A, Heinrich P and Seidler A: Interventions to
support return to work for people with coronary heart disease.
Cochrane Database Syst Rev. 3:CD0107482019.PubMed/NCBI
|
|
7
|
McKavanagh P, Yanagawa B, Zawadowski G and
Cheema A: Management and prevention of saphenous vein graft
failure: A review. Cardiol Ther. 6:203–223. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dianati Maleki N, Ehteshami Afshar A and
Parikh PB: Management of saphenous vein graft disease in patients
with prior coronary artery bypass surgery. Curr Treat Options
Cardiovasc Med. 21:122019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Virk HUH, Lakhter V, Ahmed M, O'Mucrchu B
and Chatterjee S: Radial artery versus saphenous vein grafts in
coronary artery bypass surgery: A literature review. Curr Cardiol
Rep. 21:362019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kenagy RD, Kikuchi S, Chen L, Wijelath ES,
Stergachis AB, Stamatoyannopoulos J, Tang GL, Clowes AW and Sobel
M: A single nucleotide polymorphism of cyclin-dependent kinase
inhibitor 1B (p27(Kip1)) associated with human vein graft failure
affects growth of human venous adventitial cells but not smooth
muscle cells. J Vasc Surg. 67:309–317. e72018. View Article : Google Scholar
|
|
11
|
Wadey K, Lopes J, Bendeck M and George S:
Role of smooth muscle cells in coronary artery bypass grafting
failure. Cardiovasc Res. 114:601–610. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Campos LCG, Ribeiro-Silva JC, Menegon AS,
Barauna VG, Miyakawa AA and Krieger JE: Cyclic stretch-induced Crp3
sensitizes vascular smooth muscle cells to apoptosis during vein
arterialization remodeling. Clin Sci (Lond). Feb 2–2018, Epub ahead
of print. View Article : Google Scholar
|
|
13
|
Gu H, Liu Z and Zhou L: Roles of miR-17-92
cluster in cardiovascular development and common diseases. Biomed
Res Int. 2017:91029092017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moghaddam AS, Afshari JT, Esmaeili SA,
Saburi E, Joneidi Z and Momtazi-Borojeni AA: Cardioprotective
microRNAs: Lessons from stem cell-derived exosomal microRNAs to
treat cardiovascular disease. Atherosclerosis. 285:1–9. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang W, Liu D, Han X, Ren J, Zhou P and
Ding P: MicroRNA-451 inhibits vascular smooth muscle cell migration
and intimal hyperplasia after vascular injury via Ywhaz/p38 MAPK
pathway. Exp Cell Res. 379:214–224. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zheng B, Yin WN, Suzuki T, Zhang XH, Zhang
Y, Song LL, Jin LS, Zhan H, Zhang H, Li JS and Wen JK:
Exosome-mediated miR-155 transfer from smooth muscle cells to
endothelial cells induces endothelial injury and promotes
atherosclerosis. Mol Ther. 25:1279–1294. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gabani M, Liu J, Ait-Aissa K, Koval O, Kim
YR, Castaneda D, Vikram A, Jacobs JS, Grumbach I, Trebak M, et al:
MiR-204 regulates type 1 IP3R to control vascular smooth muscle
cell contractility and blood pressure. Cell Calcium. 80:18–24.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Dong CQ, Peng GY, Huang HY, Yu YS,
Ji ZC and Shen ZY: MicroRNA-134-5p regulates media degeneration
through inhibiting VSMC phenotypic switch and migration in thoracic
aortic dissection. Mol Ther Nucleic Acids. 16:284–294. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cao BJ, Zhu L, Wang XW, Zou RJ and Lu ZQ:
MicroRNA-365 Promotes the contractile phenotype of venous smooth
muscle cells and inhibits neointimal formation in rat vein grafts.
IUBMB Life. 71:908–916. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Elsayed Y, Lekakou C and Tomlins P:
Modeling, simulations, and optimization of smooth muscle cell
tissue engineering for the production of vascular grafts.
Biotechnol Bioeng. 116:1509–1522. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Engler A, Dreja F, Koberle S, Thielmann M,
Peters J and Frey UH: Establishment of an easy and straight forward
heparinase protocol to analyse circulating and myocardial tissue
micro-RNA during coronary artery-bypass-graft surgery. Sci Rep.
8:13612018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van Boven N, Kardys I, van Vark LC,
Akkerhuis KM, de Ronde MWJ, Khan MAF, Merkus D, Liu Z, Voors AA,
Asselbergs FW, et al: Serially measured circulating microRNAs and
adverse clinical outcomes in patients with acute heart failure. Eur
J Heart Fail. 20:89–96. 2018. View Article : Google Scholar
|
|
23
|
Miyamoto S, Usami S, Kuwabara Y, Horie T,
Baba O, Hakuno D, Nakashima Y, Nishiga M, Izuhara M, Nakao T, et
al: Expression patterns of miRNA-423-5p in the serum and
pericardial fluid in patients undergoing cardiac surgery. PLoS One.
10:e01429042015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tutarel O, Dangwal S, Bretthauer J,
Westhoff-Bleck M, Roentgen P, Anker SD, Bauersachs J and Thum T:
Circulating miR-423-5p fails as a biomarker for systemic
ventricular function in adults after atrial repair for
transposition of the great arteries. Int J Cardiol. 167:63–66.
2013. View Article : Google Scholar
|
|
25
|
Zeng JF, Zeng ZL, Zhang K, Zhao Y, Liu YM,
Chen JJ, Tong H, Wei DH, Jiang ZS and Wang Z: miR-23b-3p and
miR-125b-5p downregulate apo(a) expression by targeting Ets1 in
HepG2 cells. Cell Biol Int. 42:313–323. 2018. View Article : Google Scholar
|
|
26
|
Bauters C, Kumarswamy R, Holzmann A,
Bretthauer J, Anker SD, Pinet F and Thum T: Circulating miR-133a
and miR-423-5p fail as biomarkers for left ventricular remodeling
after myocardial infarction. Int J Cardiol. 168:1837–1840. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hirota K, Keino H, Inoue M, Ishida H and
Hirakata A: Comparisons of microRNA expression profiles in vitreous
humor between eyes with macular hole and eyes with proliferative
diabetic retinopathy. Graefes Arch Clin Exp Ophthalmol.
253:335–342. 2015. View Article : Google Scholar
|
|
28
|
Ayoubian H, Ludwig N, Fehlmann T,
Menegatti J, Groger L, Anastasiadou E, Trivedi P, Keller A, Meese E
and Grasser FA: Epsteinbarr virus infection of cell lines derived
from diffuse large B-cell lymphomas alters microrna loading of the
Ago2 complex. J Virol. 93:e1297–e1318. 2019.
|
|
29
|
Zhang L, Yu F, Wang L, Zheng J, Du Y,
Huang Y, Liu B, Wang X and Kong W: ADAMTS-7 promotes vascular
smooth muscle cells proliferation in vitro and in vivo. Sci China
Life Sci. 58:674–681. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bengtsson E, Hultman K, Duner P, Asciutto
G, Almgren P, Orho-Melander M, Melander O, Nilsson J,
Hultgardh-Nilsson A and Goncalves I: ADAMTS-7 is associated with a
high-risk plaque phenotype in human atherosclerosis. Sci Rep.
7:37532017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang L, Zheng J, Bai X, Liu B, Liu CJ, Xu
Q, Zhu Y, Wang N, Kong W and Wang X: ADAMTS-7 mediates vascular
smooth muscle cell migration and neointima formation in
balloon-injured rat arteries. Circ Res. 104:688–698. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kessler T, Zhang L, Liu Z, Yin X, Huang Y,
Wang Y, Fu Y, Mayr M, Ge Q, Xu Q, et al: ADAMTS-7 inhibits
re-endothelialization of injured arteries and promotes vascular
remodeling through cleavage of thrombospondin-1. Circulation.
131:1191–1201. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Agarwal V, Bell GW, Nam J and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar :
|
|
35
|
García DM, Baek D, Shin C, Bell GW,
Grimson A and Bartel DP: Weak seed-pairing stability and high
target-site abundance decrease the proficiency of lsy-6 and other
miRNAs. Nat Struct Mol Biol. 18:1139–1146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of MicroRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
37
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shah T, Palaskas N and Ahmed A: An update
on gender disparities in coronary heart disease care. Curr
Atheroscler Rep. 18:282016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nouraee N and Mowla SJ: miRNA therapeutics
in cardiovascular diseases: Promises and problems. Front Genet.
6:2322015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
van Rooij E and Olson EN: MicroRNA
therapeutics for cardiovascular disease: Opportunities and
obstacles. Nat Rev Drug Discov. 11:860–872. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kostina D, Zverev D, Grebennik V, Gordeev
M, Ignatieva E, Voronkina I, Kostareva A and Malashicheva A: aortic
graft at coronary artery bypass surgery as a source of human aortic
smooth muscle cells. Cell Transplant. 26:1663–1668. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhu Y, Feng B, He S, Su Z and Zheng G:
Resveratrol combined with total flavones of hawthorn alleviate the
endothelial cells injury after coronary bypass graft surgery.
Phytomedicine. 40:20–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu X, Wang Z, Wu H, Jiang W and Hu R: Ras
ssDNA aptamer inhibits vascular smooth muscle cell proliferation
and migration through MAPK and PI3K pathways. Int J Mol Med.
35:1355–1361. 2015.PubMed/NCBI
|
|
44
|
Wang D, Wang Y, Ma J, Wang W, Sun B, Zheng
T, Wei M and Sun Y: MicroRNA-20a participates in the aerobic
exercise-based prevention of coronary artery disease by targeting
PTEN. Biomed Pharmacother. 95:756–763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Frey UH, Klaassen M, Ochsenfarth C, Murke
F, Thielmann M, Kottenberg E, Kleinbongard P, Klenke S, Engler A,
Heusch G, et al: Remote ischaemic preconditioning increases serum
extracellular vesicle concentrations with altered micro-RNA
signature in CABG patients. Acta Anaesthesiol Scand. 63:483–492.
2019.
|
|
46
|
Wang Z, Li X, Shen J, Tian D, Ji Q, Xia L
and Lv Q: Plasma microRNAs reflecting cardiac and inflammatory
injury in coronary artery bypass grafting surgery. J Surg Res.
224:58–63. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ram TP, Fomison-Nurse I, Gandhi S, Coffey
S, Saxena P, Galvin I, Bunton R, Williams MJA, Lamberts RR and
Katare R: The diagnostic sensitivity of circulating cardio-enriched
microRNAs is increased after normalization of high-density
lipoprotein levels. Int J Cardiol. 236:498–500. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Santulli G: microRNAs Distinctively
regulate vascular smooth muscle and endothelial cells: Functional
implications in angiogenesis, atherosclerosis, and in-stent
restenosis. Adv Exp Med Biol. 887:53–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yeh YT, Wei J, Thorossian S, Nguyen K,
Hoffman C, Del Alamo JC, Serrano R, Li YJ, Wang KC and Chien S:
MiR-145 mediates cell morphology-regulated mesenchymal stem cell
differentiation to smooth muscle cells. Biomaterials. 204:59–69.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hall IF, Climent M, Quintavalle M, Farina
FM, Schorn T, Zani S, Carullo P, Kunderfranco P, Civilini E,
Condorelli G and Elia L: Circ_Lrp6, a circular RNA enriched in
vascular smooth muscle cells, acts as a sponge regulating miRNA-145
function. Circ Res. 124:498–510. 2019. View Article : Google Scholar
|
|
51
|
Yasmeen S, Kaur S, Mirza AH, Brodin B,
Pociot F and Kruuse C: miRNA-27a-3p and miRNA-222-3p as novel
modulators of phosphodiesterase 3a (PDE3A) in cerebral
microvascular endothelial cells. Mol Neurobiol. 56:5304–5314. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gu J, Zhang H, Ji B, Jiang H, Zhao T,
Jiang R, Zhang Z, Tan S, Ahmed A and Gu Y: Vesicle miR-195 derived
from endothelial cells inhibits expression of serotonin transporter
in vessel smooth muscle cells. Sci Rep. 7:435462017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fang Q, Tian M, Wang F, Zhang Z, Du T,
Wang W, Yang Y, Li X, Chen G, Xiao L, et al: Amlodipine induces
vasodilation via Akt2/Sp1-activated miR-21 in smooth muscle cells.
Br J Pharmacol. 176:2306–2320. 2019.PubMed/NCBI
|
|
54
|
Liu S, Yang Y, Jiang S, Xu H, Tang N, Lobo
A, Zhang R, Liu S, Yu T and Xin H: MiR-378a-5p regulates
proliferation and migration in vascular smooth muscle cell by
targeting CDK1. Front Genet. 10:222019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Huang SC, Wang M, Wu WB, Wang R, Cui J, Li
W, Li ZL, Li W and Wang SM: Mir-22-3p inhibits arterial smooth
muscle cell proliferation and migration and neointimal hyperplasia
by targeting HMGB1 in arteriosclerosis obliterans. Cell Physiol
Biochem. 42:2492–2506. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Qu Q, Bing W, Meng X, Xi J, Bai X, Liu Q,
Guo Y, Zhao X and Bi Y: Upregulation of miR-126-3p promotes human
saphenous vein endothelial cell proliferation in vitro and prevents
vein graft neointimal formation ex vivo and in vivo. Oncotarget.
8:106790–106806. 2017. View Article : Google Scholar :
|
|
57
|
Fichtlscherer S, De Rosa S, Fox H,
Schwietz T, Fischer A, Liebetrau C, Weber M, Hamm CW, Röxe T,
Müller-Ardogan M, et al: Circulating microRNAs in patients with
coronary artery disease. Circ Res. 107:677–684. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pu X, Xiao Q, Kiechl S, Chan K, Ng FL, Gor
S, Poston RN, Fang C, Patel A, Senver EC, et al: ADAMTS7 cleavage
and vascular smooth muscle cell migration is affected by a
coronary-artery-disease-associated variant. Am J Hum Genet.
92:366–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wu W, Wang H, Yu C, Li J, Gao Y, Ke Y,
Wang Y, Zhou Y and Zheng J: Association of ADAMTS-7 levels with
cardiac function in a rat model of acute myocardial infarction.
Cell Physiol Biochem. 38:950–958. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu J, Zhou B, Yu H, Han J, Cui M, Zhang F,
Wang G, Guo L and Gao W: Association between plasma ADAMTS-7 levels
and severity of disease in patients with stable obstructive
coronary artery disease. Medicine (Baltimore). 95:e55232016.
View Article : Google Scholar
|
|
61
|
Jansen F, Stumpf T, Proebsting S, Franklin
BS, Wenzel D, Pfeifer P, Flender A, Schmitz T, Yang X, Fleischmann
BK, et al: Intercellular transfer of miR-126-3p by endothelial
microparticles reduces vascular smooth muscle cell proliferation
and limits neointima formation by inhibiting LRP6. J Mol Cell
Cardiol. 104:43–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wu W, Zhou Y, Li Y, Li J, Ke Y, Wang Y and
Zheng J: Association between plasma ADAMTS-7 levels and ventricular
remodeling in patients with acute myocardial infarction. Eur J Med
Res. 20:272015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Du Y, Gao C, Liu Z, Wang L, Liu B, He F,
Zhang T, Wang Y, Wang X, Xu M, et al: Upregulation of a disintegrin
and metallo-proteinase with thrombospondin motifs-7 by miR-29
repression mediates vascular smooth muscle calcification.
Arterioscler Thromb Vasc Biol. 32:2580–2588. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bendeck MP, Conte M, Zhang M, Nili N,
Strauss BH and Farwell SM: Doxycycline modulates smooth muscle cell
growth, migration, and matrix remodeling after arterial injury. Am
J Pathol. 160:1089–1095. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang L, Zheng J, Du Y, Huang Y, Li J, Liu
B, Liu CJ, Zhu Y, Gao Y, Xu Q, et al: Cartilage oligomeric matrix
protein maintains the contractile phenotype of vascular smooth
muscle cells by interacting with alpha(7)beta(1) integrin. Circ
Res. 106:514–525. 2010. View Article : Google Scholar
|