Introduction
Parkinson's disease (PD), which was first documented
in 1817 by Dr James Parkinson, is the second most common
neurode-generative disorder with a prevalence of approximately 1-2%
among individuals >65 years of age (1) and approximately 4-5% among
individuals >85 years of age (2). The neuropathological hallmarks of PD
are the loss of dopaminergic neurons in the substantia nigra (SN)
pars compacta and the accumulation of Lewy bodies (LBs), which
mainly consist of α-synuclein (3), thus generating symptoms, including
bradykinesia, resting tremor, muscle rigidity and postural
instability (4). Previous studies
have demonstrated that various environmental factors, such as
nutrition, and exposure to metals and pesticides can affect the
progression of PD (5-7). Currently, however, the
identification of an efficient method with which to attenuate the
development of PD is warranted.
MicroRNAs (miRNAs or miRs) are a group of small and
non-coding RNAs comprising of 20-22 nucleotides in length, which
regulate genes associated with signaling pathways through either
mRNA cleavage or post-transcriptional silencing by targeting the
3′untranslated region (3′UTR) of their targets (8,9).
miRNAs are expressed throughout the developmental stages spatially
and temporally (10). miRNAs are
dysregulated in numerous diseases, including PD (11). In addition, numerous miRNAs have
been discovered to be associated with PD. For instance,
miRNA-155-5p expression has been shown to be increased, whereas the
expression of miRNA-146a-5p has been shown to be decreased in
patients with PD compared with healthy individuals (12). Furthermore, the decreased miR-433
and miR-133b expression levels are potential biomarkers for PD
(13). A previous study also
demonstrated that miR-410 exerts neuroprotective effects in an
in vitro model of 6-hydroxydopamine-induced PD (14). Recently, in another previous
study, in an in vitro model of rotenone-induced PD, an
increased miR-384-5p level was observed, and the downregulation of
this miRNA exerted a neuroprotective effect against rotenone by
targeting GRP78 (15). However,
the role of miR-384-5p in an in vivo model of PD remains
unknown. In addition, it remains unknown as to whether there are
other molecules that may be regulated by miR-384-5p. Therefore, the
present study aimed to investigate the role of miR-384-5p in an
in vitro and in vivo model of PD in order to
elucidate this matter. The findings of this study may prove to be
of paramount importance, as numerous miRNAs are being applied in
various clinical trial phases for the purpose of being used as
therapeutic drugs.
Materials and methods
Animals
Experiments were performed in 30 adult male C57B/6J
mice (weighing 20-30 g, 8 weeks old), which were obtained from the
Model Animal Research Center of Nanjing University. The mice were
kept in a controlled environment with a temperature of 22-25°C, a
12:12 h light/dark cycle, with free access to water and food.
Manipulations were conducted during the light phase of the day.
Efforts were made to minimize animal suffering and to reduce the
number of animals used. The present study was performed in
accordance with the guidelines of the Committee on Care and Use of
Experimental Animals Resources and with the approval of Ethical
Committee for The First Hospital of Yulin.
Experimental groups
The mice were randomly divided into 3 different
groups with 10 mice in each group, including the control group
(n=10), the dimethyl sulfoxide (DMSO) control group (n=10) and the
rotenone-induced PD group (n=10). The duration of the experiment
was 1 month.
Rotenone (Sigma Chemical Co.) was first dissolved in
DMSO, which was completed with sunflower oil. Rotenone was
administered orally once daily (0.1 ml/10 g, 30 mg/kg) for 30 days
to establish an in vivo model of PD, as previously described
(16). The mice in the DMSO
control group were orally administered once daily with DMSO for 30
days. The mice in the control group received no treatments.
Afterwards, the mice in each group were submitted to a swimming
test and traction test.
Swimming test
The swimming test was conducted to determine the
motor disability of the mice in each group using a round glass
swimming tank (length, 40 cm; width, 25 cm; height, 16 cm), which
was filled with water (at a temperature of 22-25°C) to a depth of
12 cm. The mice were scored using the following scale: 0, No
swimming with the head above the water; 1, occasional swim with
mice floating using the hind paws; 2, alternations between
swim-floating and passively floating; and 3, continuous swimming,
as previously described (17).
Traction test
The traction test was performed to determine the
muscle strength and equilibrium of the mice. The forepaws of the
mice were placed on a rope (diameter, 5 mm) which was approximately
horizontally 70 cm at a distance from the ground. The hind limb
placements of the mice were scored on a scale of 1 to 3, with the
lowest score indicating the most severe deficits. The score was
determined using the following criteria: 1 or 2, No or one hind
limb seizing the rope; and 3, both hind limbs seizing the rope.
Mice were allowed to hang upside down and stay on the rope for 30
sec, as previously described (18).
Tissue preparation
Animal health and behavior was monitored once every
2 days with the following principles: Soft and smooth hair, even
breath and no scabs, etc. During the process, no mouse had died.
After the swimming test and traction test, the animals were
euthanized by barbiturate overdose (pentobarbital sodium, 150
mg/kg; i.p.) and were immediately decapitated. Death was verified
by the absence of breathing, heartbeat or neurofeedback, etc. SN
tissues were then extracted from the mice and fresh-frozen at −80°C
which were then subjected to the following experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the SH-SY5Y cells
(please see below) and SN tissues using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). Reverse transcription and TaqMan
microRNA assay were carried out using the Hairpin-it™ miRNA qPCR
Quantitation kit (GenePharma Co.). Thereafter, qPCR was performed
using the SYBR-Green kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) on the ABI 7500 Real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: 95°C for 2 min, followed by 40 cycles
of 95°C for 15 sec, 58°C for 30 sec and 72°C for 30 sec. The primer
sequences were as follows: Sirtuin 1 (SIRT1) forward, 5′-CTG GGG
TGT CTG TTT CAT GTG G-3′ and reverse, 5′-GCT TGA GGA TCT GGA AGA
TCT GG-3′; GAPDH forward, 5′-CCA CTC CTC CAC CTT TGA CG and
reverse, 5′-CCA CCA CCC TGT TGC TGT AG-3′; forkhead box protein O1
(FOXO1) forward, 5′-TCC CAG TGA GCA GTA AAT C-3′ and reverse,
5′-CCA GCA GTT GAA CAA GTC-3′; p53 forward, 5′-GCT GGT TAG GTA GAG
GGA GTT G-3′ and reverse, 5′-GTG TGG GAT GGG GTG AGA TTT C-3′;
miR-384-5p, 5′-CGC GTA TGA ACA ATT TCT AGG AAT-3′; and U6, 5′-TTG
GTG AAG CGT TCC ATA TTT T-3′. The relative expression levels of
miRNA and mRNAs were determined using the 2−ΔΔCq method
(19). The expression of
miR-384-5p was normalized to U6, while the expression of SIRT1, p53
and FOXO1 was normalized to GAPDH.
Prediction of targeting mRNAs for
miR-384-5p
The binding sites between miR-384-5p and SIRT1 were
analyzed by TargetScan release 7.1 (http://www.targetscan.org/vert_71/).
Western blot analysis
Proteins were isolated from the SH-SY5Y cells
(please see below) or SN tissues by radioimmunoprecipitation assay
(RIPA) (Beyotime Institute of Biotechnology). The protein
concentrationin each sample was detected by BCA assay (Beyotime
Institute of Biotechnology). The same amount of proteins from each
sample (15 µg) were separated by 8-10% SDS-PAGE and
transferred onto PVDF membranes (Millipore). After blocking with 5%
non-fat milk at room temperature for 1 h, the PVDF membranes were
incubated with respective primary antibodies for α-synuclein rabbit
mAb (dilution, 1:1,000, cat. no. 4179), SIRT1 rabbit mAb (dilution,
1:1,000, cat. no. 9475), p53 rabbit mAb (dilution, 1:1,000, cat.
no. 2527), FOXO1 rabbit mAb (dilution, 1:1,000, cat. no. 2880), and
GAPDH rabbit mAb (dilution, 1:1,000, cat. no. 5174) overnight at
4°C and anti-rabbit IgG, HRP-linked secondary antibodies (dilution,
1:2,000, cat. no. 7074) at room temperature for 2 h, successively.
The above-mentioned antibodies were obtained from Cell Signaling
Technology. ECL system (Bio-Rad Laboratories Inc.) was used to
detect antibody bound proteins. Densitometric analysis was
performed using Image J 1.8.0 software (National Institutes of
Health).
Cells and cell culture
SH-SY5Y cells human dopaminergic neuroblastoma were
obtained from the American Type Culture Collection (cat. no.
ATCC® CRL-2266™; ATCC). STR profiling was used for the
authentication of the SH-SY5Y cells. The SH-SY5Y cells were
cultured in Dulbecco's modified Eagle's medium-Ham's Nutrient
Mixture F-12 (DMEM/F12; Invitrogen; Thermo Fisher Scientific) which
was supplemented with 10% fetal calf serum (FCS), and 1%
penicillin/streptomycin in a humidified atmosphere containing 5%
CO2 at 37°C. The cells were randomly divided into 3
groups, including the control group, DMSO group and
rotenone-induced PD group. The SH-SY5Y cells were exposed to
rotenone (20 µM) for 24 h, as previously described (20) before being harvested for use in
subsequent experiments.
Cell transfection
The SH-SY5Y cells (2×105 cells/well) were
seeded into 6-well plates and cultured till 90% confluence prior to
cell transfection. miR-NC inhibitor and miR-384-5p inhibitor were
obtained from GenePharma. Briefly, miR-384-5p inhibitor (2.5
µl) or miR-NC inhibitor (2.5 µl) and Lipofectamine
2000 (5 µl, Invitrogen; Thermo Fisher Scientific, Inc.) were
added into 250 µl DMEM. Subsequently, the mixture was mixed
thoroughly and added into the 6-well plates to a final work
concentration of 20 nmol/l. The above mixture was incubated for 48
h prior to use in the subsequent experiments.
Dual luciferase reporter assay
The 293 cells (ATCC) and SH-SY5Y cells transfected
with miR-384-5p inhibitor or miR-NC inhibitor were seeded into
96-well plates (1×103 cells/well) and cultured for 24 h.
The SIRT1 3′UTR was amplified from the cDNA of 293 cells and
inserted into pLight Switch Prom (Switchgear Genomics).
Subsequently, pLightSIRT1-WT-3′UTR reporter construct or
pLight-SIRT1-MUT-3′UTR was transfected into the 293 cells and
SH-SY5Y cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 24 h after cell transfection, Firefly
luciferase activity was determined by Light Switch Luciferase assay
(Switchgear Genomics) through normalization to Renilla
luciferase activity.
Cell apoptosis assay
The Annexin V-FITC/PI Staining Apoptosis Detection
kit was used for the detection of cell apoptosis. SH-SY5Y cells
were collected and washed with PBS. Thereafter, the SH-SY5Y cells
(3×105) were resuspended and incubated with Annexin
V-FIFC (5 µl) and PI (10 µl) at 37°C for 15 min. The
cell apoptotic rate was analyzed using a flow cytometer (BD
Biosciences).
Evaluation of α-synuclein
aggregation
Following incubation at 37°C for 48 h, the SH-SY5Y
cells were fixed with 4% formaldehyde for 15 min at 4°C, followed
by permeabilization with 0.2% Triton X-100 for 10 min at room
temperature. The primary antibody for α-synuclein rabbit mAb
(dilution, 1:200, cat. no. 4179) was incu bated at 4°C overnight.
Anti-rabbit IgG (Alexa Fluor 488 conjugate, cat. no. 4412;
dilution, 1:500) acted as a secondary antibody to detect
fluorescence and was incubated at room temperature for 2 h.
Subsequently, the cells were incubated with DAPI (cat. no. 4083;
0.5 µg/ml) for 10 min at room temperature for cell nuclei
staining. The above-mentioned antibodies were obtained from Cell
Signaling Technology. Images were captured under a confocal
microscope (TCS SP 5 II, Leica). Fluorescence intensity was
analyzed using MicroBrightField Stereo-Investigator (MBF
Bioscience).
Statistical analysis
GraphPad Prism 7 software (GraphPad Software, Inc.)
was applied for the analysis of the data. The results are expressed
as the means ± SEM. Comparisons between 2 groups were analyzed
using a Student's t-test, while comparisons among 3 groups were
analyzed by one-way analysis of variance followed by Newman-Keuls
post-hoc analysis. The correlation between miR-384-5p and SIRT1 was
analyzed by Pearson's correlation analysis. A value of P<0.05
was considered to indicate a statistically significant
difference.
Results
Increased a-synuclein and decreased
swimming/traction scores in mice with rotenone-induced PD
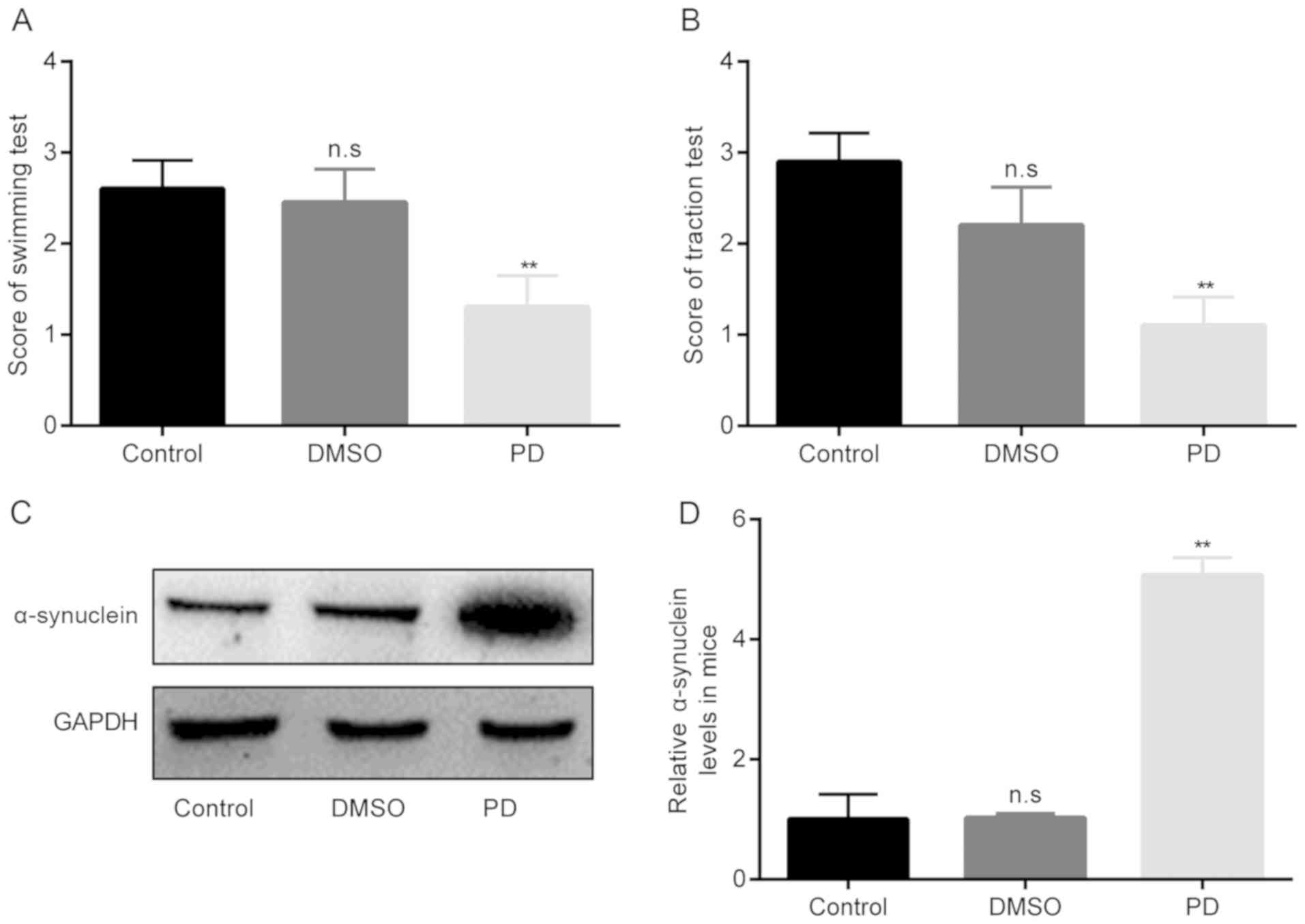
In the in vivo model of PD in this study, the
severity of parkinsonian signs was first evaluated through swimming
scores and traction scores at day 1 following rotenone induction,
as displayed in Fig. 1A and B.
The results demonstrated that there were no significant differences
in swimming scores between the control group and DMSO group,
whereas in the group of rotenone-exposed PD mice, the swimming
scores became significantly lower than those of the DMSO group
(Fig. 1A).
Furthermore, the data presented in Fig. 1B demonstrated that there were no
significant differences in traction scores between the control
group and DMSO group, whereas the induction of PD with rotenone led
to a significant decrease in the score of the traction test below
the DMSO values.
In addition, the protein expression levels of
α-synuclein were examined in each group. As shown in Fig. 1C and D, no significant were
observed between the control group and DMSO group; however, in the
group of rotenone-exposed PD mice, the protein expression levels of
α-synuclein became significantly higher than those of the DMSO
group. Taken together, these aforementioned results verify that the
constructed PD model was successful.
Increased miR-384-5p expression in mice
with rotenone- induced PD and targeting of SIRT1 by miR-384-5p in
293 cells
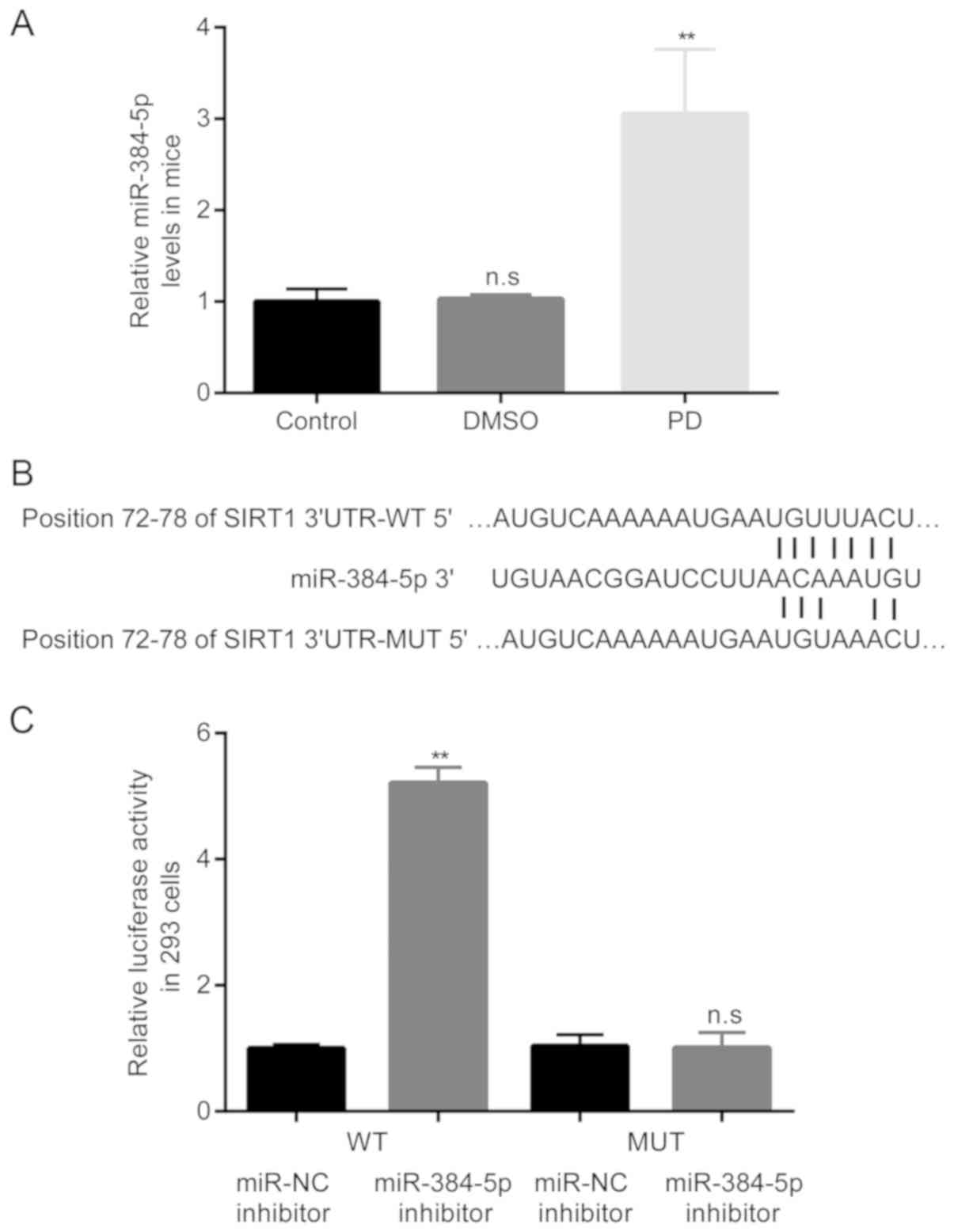
Subsequently, the differences in the expression
levels of miR-384-5p in the mice among the 3 groups were examined
by RT-qPCR. The results revealed that compared with the control
group, there were no obvious differences in miR-384-5p levels in
the DMSO group; however, in the group of mice with
rote-none-induced PD, the expression levels of miR-384-5p became
significantly higher than those of the mice in the DMSO group
(Fig. 2A).
The following experiment was performed in an aim to
predict the potential targets for miR-384-5p. As exhibited in
Fig. 2B, using the online
software TargetScan, it was discovered that the 3′UTR of SIRT1 had
the complementary site for miR-384-5p; moreover, the mutant
sequence of the 3′UTR of SIRT1 was obtained.
Subsequently, the interaction between miR-384-5p and
SIRT1 was confirmed by dual luciferase reporter assay. The results
demonstrated that transfection with miR-384-5p inhibitor
significantly induced the relative luciferase activity of 293 cells
transfected with luciferase activity plasmid containing SIRT
3′UTR-WT, but not SIRT 3′UTR-MUT (Fig. 2C).
Negative correlation between miR-384-5p
and SIRT1 expression in mice with rotenone-induced PD
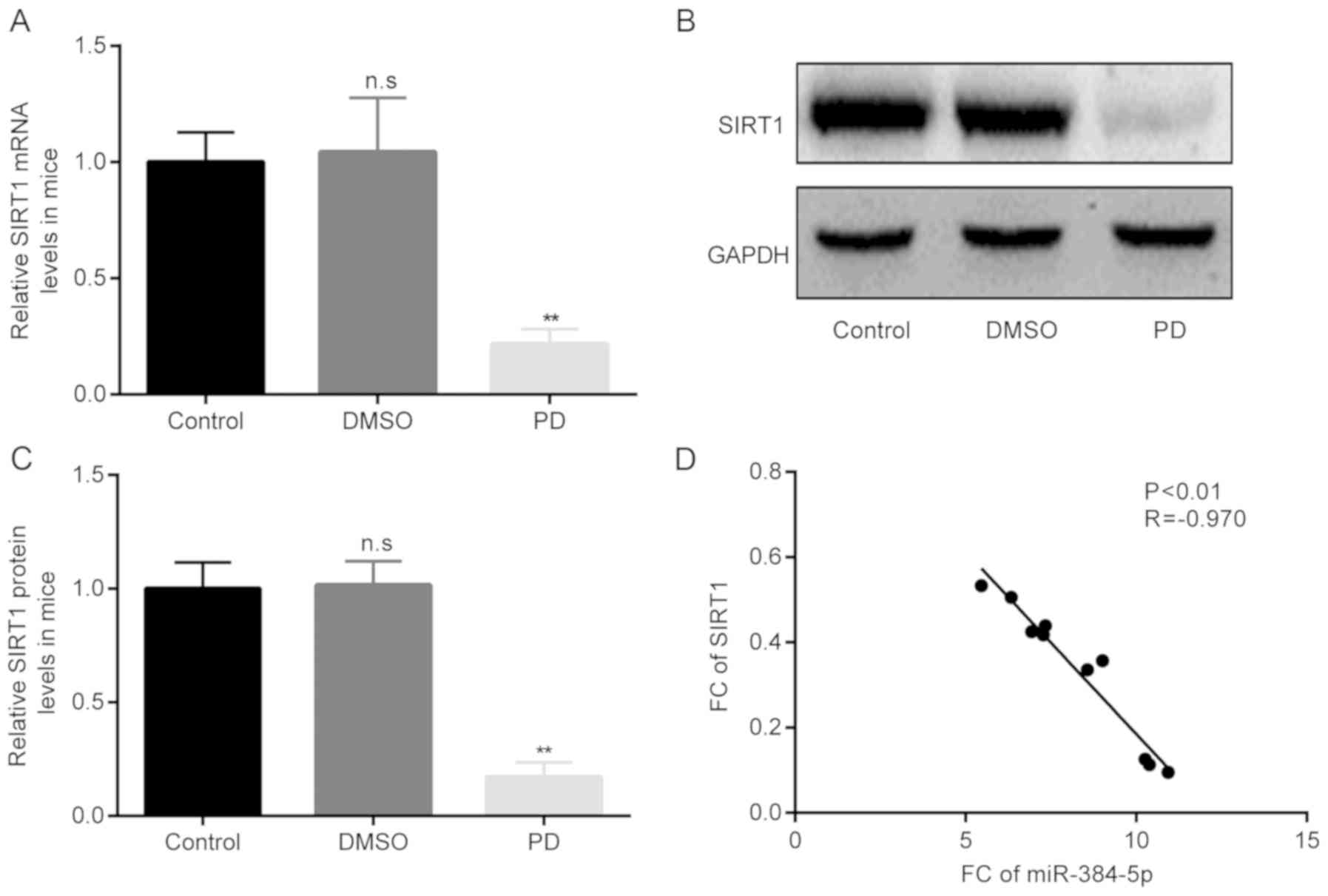
Thereafter, the differences in the mRNA and protein
expression levels of SIRT1 in mice with rotenone-induced PD mice
among the 3 groups were examined by RT-qPCR and western blot
analysis, respectively. The results of RT-qPCR revealed that,
compared with the control group, there were no obvious differences
in the SIRT1 mRNA levels between the control and the DMSO group;
however, in the group of mice with rotenone-induced PD, the mRNA
levels of SIRT1 became significantly lower than those of the mice
in the DMSO group (Fig. 3A).
Moreover, the results of western blot analysis revealed similar
results to those obtained with RT-qPCR (Fig. 3B and C).
Of note, the results of Pearson's correlation
analysis demonstrated that there was a significant negative
correlation between miR-384-5p and SIRT1 expression in the mice
with rotenone-induced PD (Fig.
3D).
Increased p53 and FOXO1 expression in
mice with rotenone- induced PD
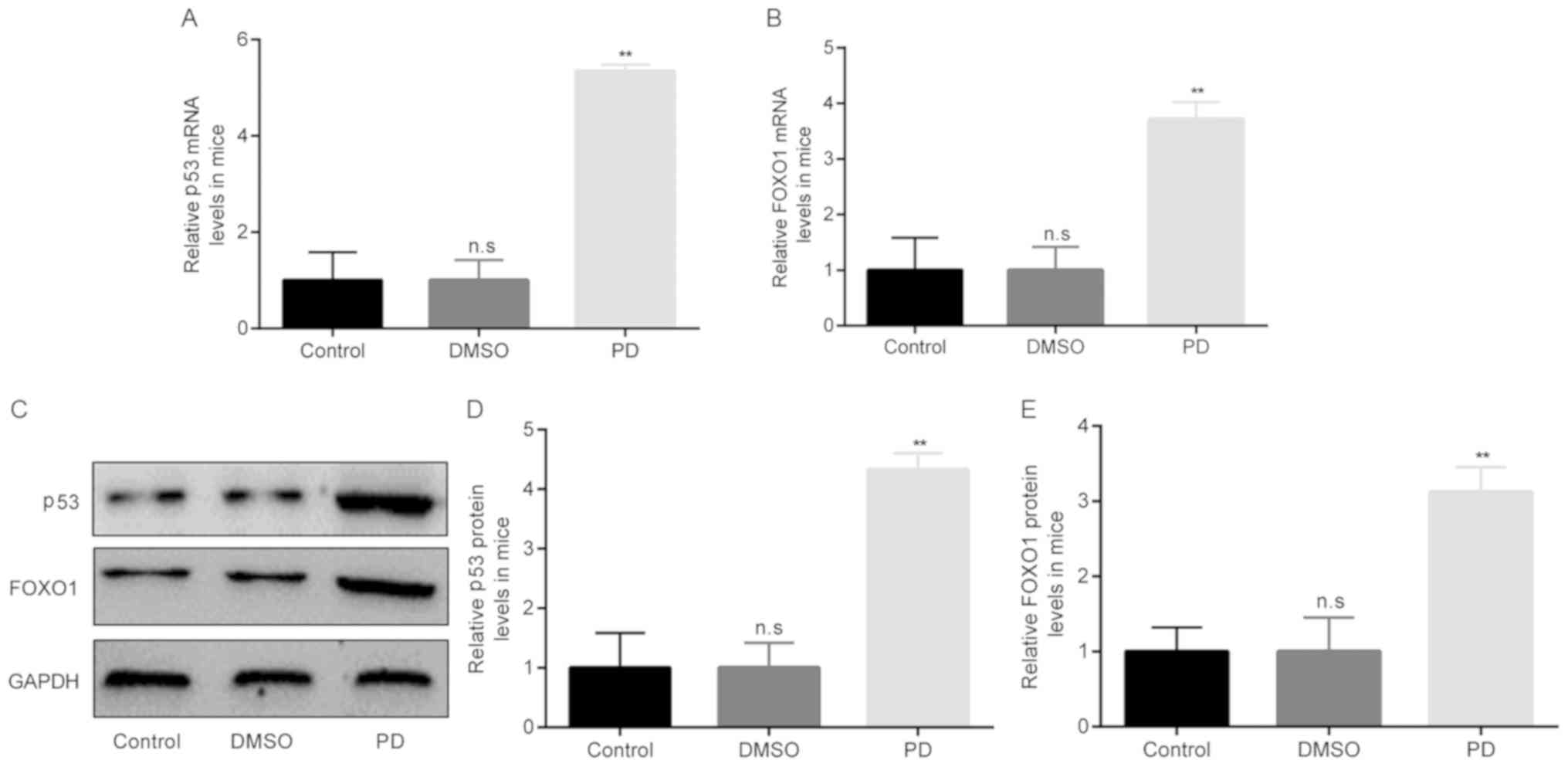
Based on the above-mentioned findings, the molecules
that are associated with SIRT1 in mice with rotenone-induced PD
were then investigated. A previous study found that p53 and FOXO1
were associated with SIRT1 in MPP+-induced SH-SY5Y cells
(21). Therefore, this study also
examined the association between SIRT1 and P53/FOXO1 in mice with
rotenone-induced PD.
The differences in the mRNA and protein expression
levels of p53/FOXO1 were determined among the 3 groups by RT-qPCR
and western blot analysis, respectively. The results of RT-qPCR
revealed that compared with the control group, there were no
obvious differences in the p53/FOXO1 mRNA levels in the DMSO group.
However, in the group of mice with rotenone-induced PD, the mRNA
levels of p53/FOXO1 became significantly higher than those of the
mice in the DMSO group (Fig. 4A and
B). Moreover, the results of western blot analysis revealed
similar results to those obtained with RT-qPCR (Fig. 4C-E).
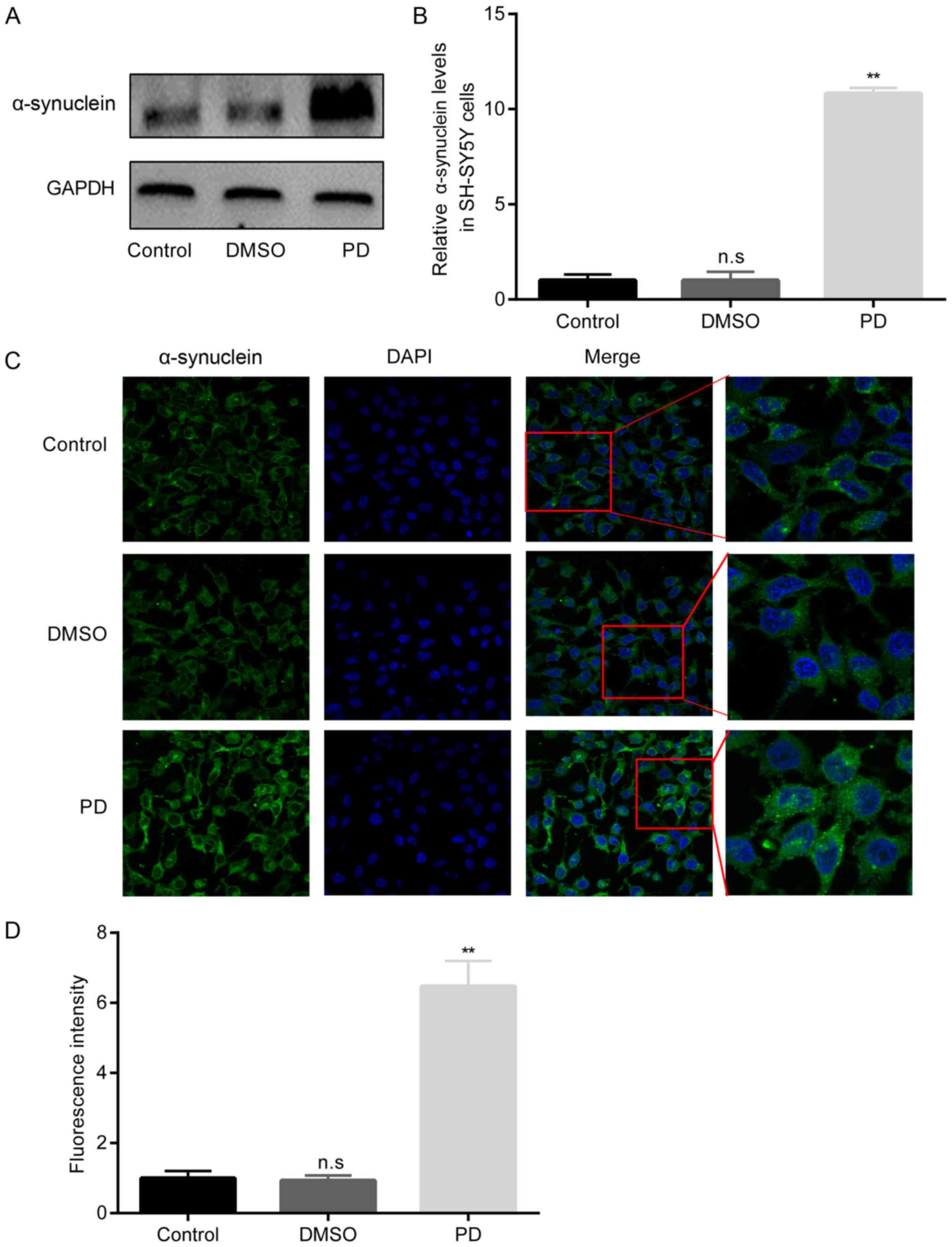
Increased α-synuclein in rotenone-treated
SH-SY5Y cells
In the in vitro model of PD in this study,
the protein expression level of α-synuclein was first evaluated in
the SH-SY5Y cells in the 3 groups. The results presented in
Fig. 5 demonstrated that there
were no significant differences in α-synuclein protein levels
between the control group and DMSO group. However, in the PD group
of rotenone-exposed SH-SY5Y cells, the α-synuclein protein levels
became significantly higher than those in the DMSO group.
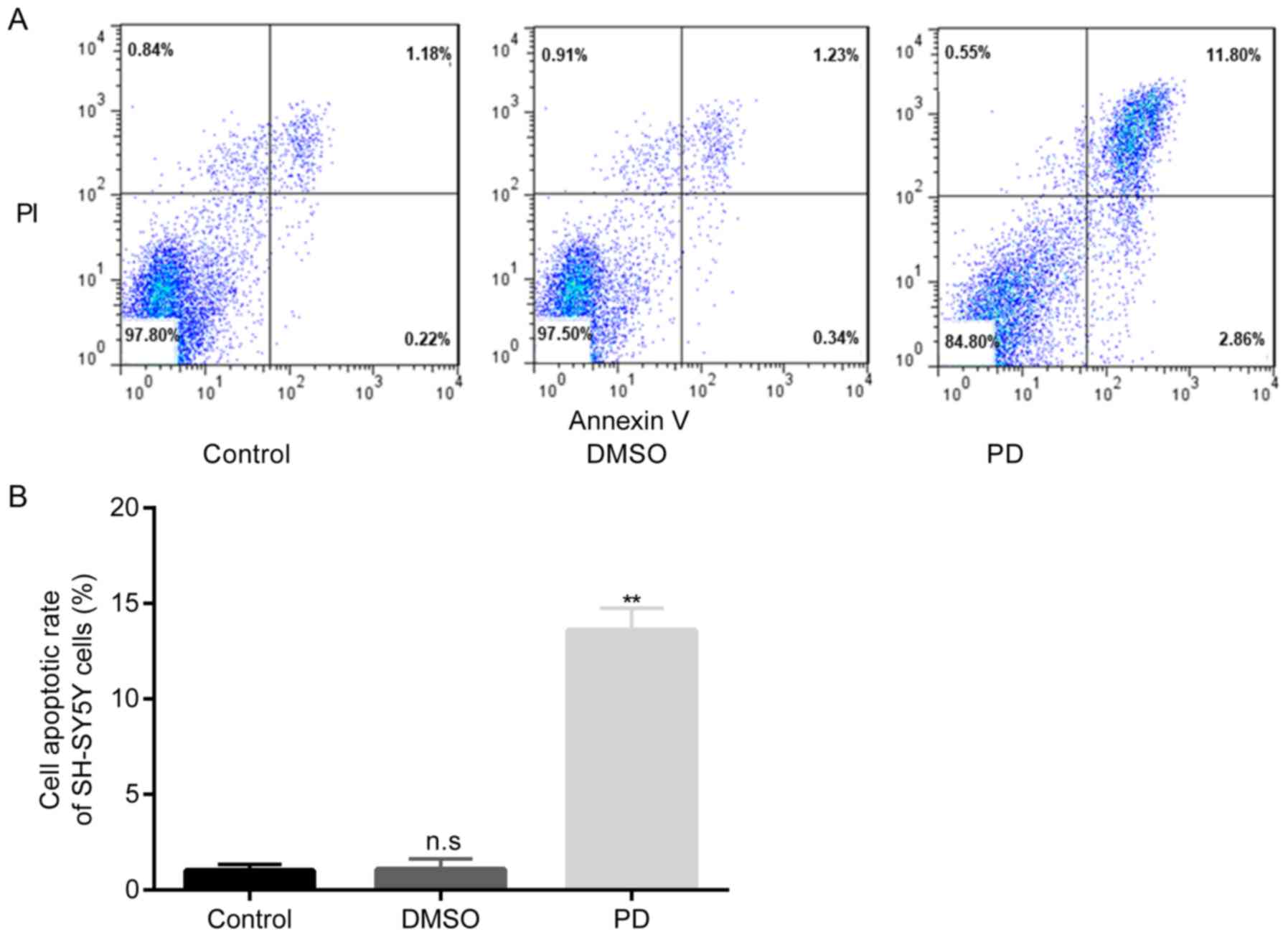
Increased apoptosis of rotenone-exposed
SH-SY5Y cells
The data presented in Fig. 6 demonstrated that there were no
significant differences in the apoptotic rate of SH-SY5Y cells
between the control group and DMSO group; however, the exposure of
the cells in the PD group to rotenone led to a significant increase
in the cell apoptotic rate compared with the DMSO group.
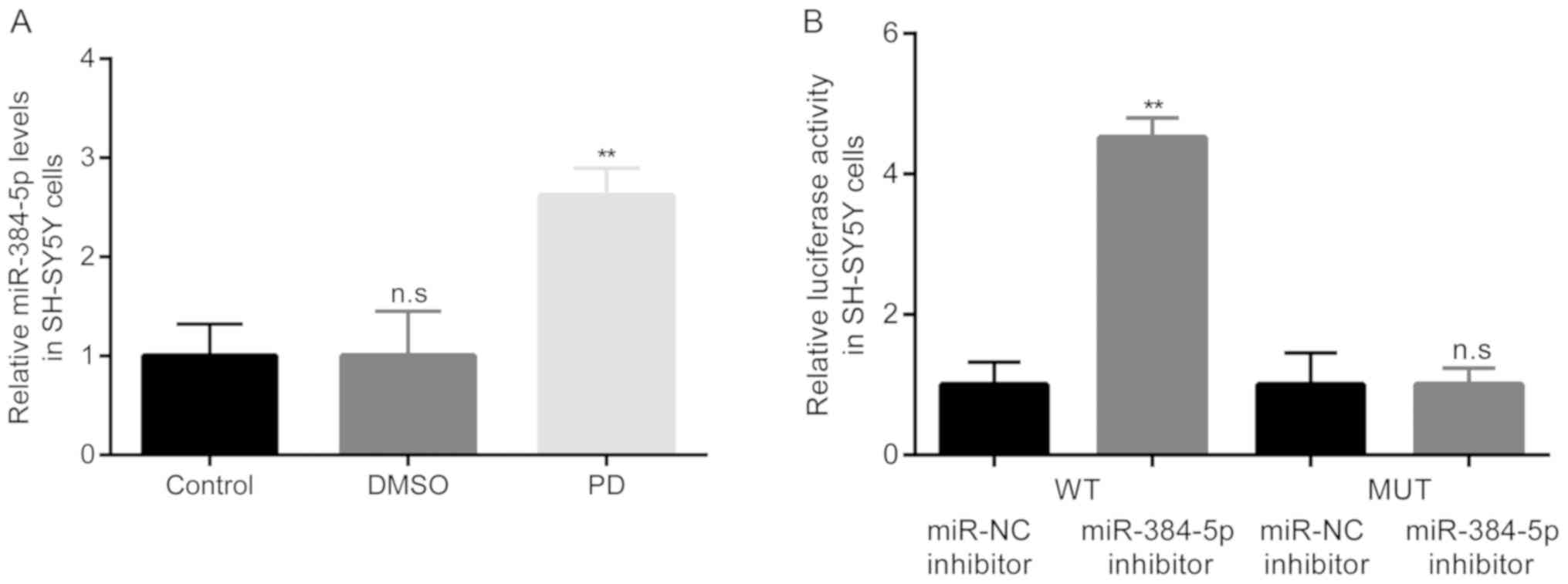
miR-384-5p targets SIRT1 in
rotenone-exposed SH-SY5Y cells
Subsequently, the differences in the expression
levels of miR-384-5p in the SH-SY5Y cells among the 3 groups were
examined by RT-qPCR. The results revealed that compared with the
control group, there were no obvious differences in miR-384-5p
levels in the DMSO group. Howev PD er, in the PD group of
rotenone-exposed SH-SY5Y cells, the expression levels of miR-384-5p
became significantly higher than those in the DMSO group (Fig. 7A).
Subsequently, the interaction between miR-384-5p and
SIRT1 was confirmed by dual luciferase reporter assay, and the
results demonstrated that transfection with miR-384-5p inhibitor
significantly induced the relative luciferase activity of SH-SY5Y
cells transfected with luciferase activity plasmid containing the
SIRT 3′UTR-WT, but not SIRT 3′UTR-MUT (Fig. 7B).
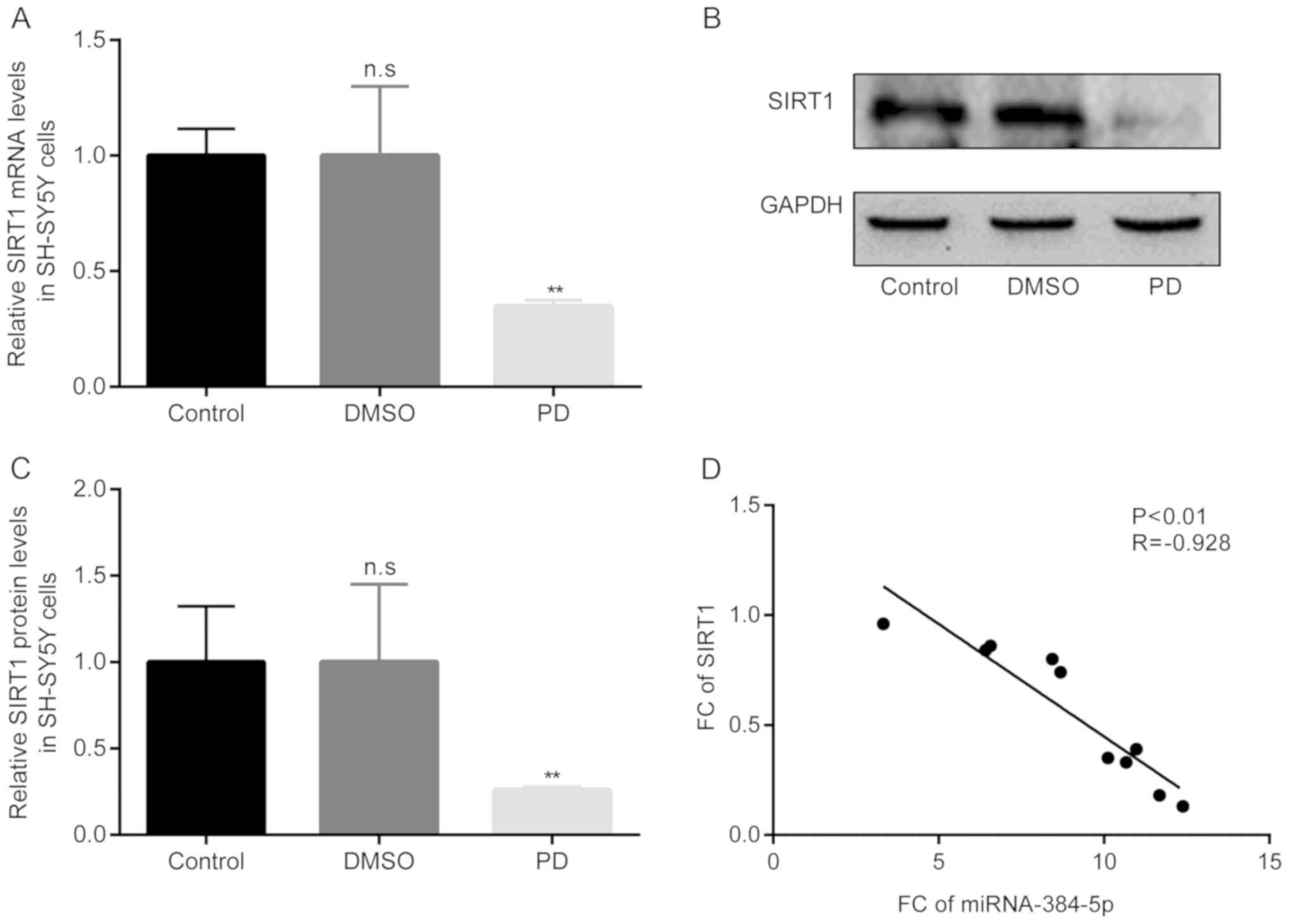
Negative correlation between miR-384-5p
and SIRT1 in rotenone-exposed SH-SY5Y cells
Thereafter, the differences in the mRNA and protein
expression levels of SIRT1 in the rotenone-exposed SH-SY5Y cells in
the PD group in the 3 groups were examined by RT-qPCR and western
blot analysis, respectively. The results of RT-qPCR revealed that
compared with the control group, there were no obvious differences
in SIRT1 mRNA levels in the DMSO group; however, in the PD group of
rotenone-exposed SH-SY5Y cells, the mRNA levels of SIRT1 became
significantly lower than those in the DMSO group (Fig. 8A). Moreover, the results of
western blot analysis revealed similar results to those obtained
with RT-qPCR (Fig. 8B and C).
Of note, the results of Pearson's correlation
analysis demonstrated that there was a significant negative
correlation between miR-384-5p and SIRT1 expression in the
rotenone-exposed SH-SY5Y cells in the PD group (Fig. 8D).
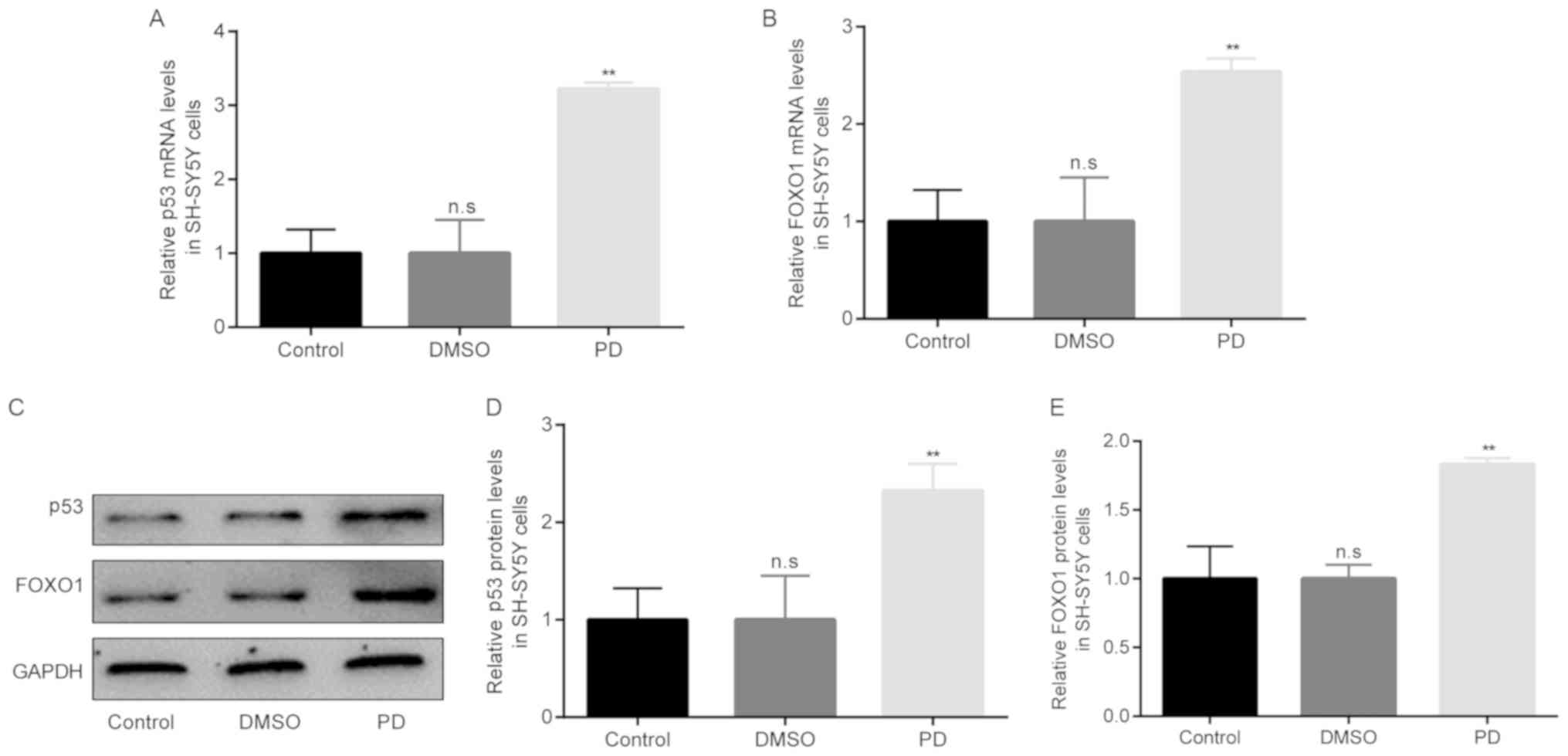
Increased p53 and FOXO1 expression in
rotenone-exposed SH-SY5Y cells
The association between SIRT1 and p53/FOXO1
expression in rotenone-exposed SH-SY5Y cells was then examined. The
differences in the mRNA and protein expression levels of p53/FOXO1
among the 3 groups were examined by RT-qPCR and western blot
analysis, respectively. The results of RT-qPCR revealed that
compared with the control group, there were no obvious differences
in p53/FOXO1 mRNA levels in the DMSO group. However, in the PD
group of rotenone-exposed SH-SY5Y cells, the mRNA levels of
p53/FOXO1 became significantly higher than those in the DMSO group
(Fig. 9A and B). Moreover, the
results of western blot analysis revealed similar results to those
of RT-qPCR (Fig. 9C-E).
Discussion
The prevalence of PD is 1-2% for patients >65
years of age (1) and 4-5% for
patients >85 years of age (2).
Numerous miRNAs have been discovered to be associated with PD. For
instance, miRNA-155-5p and miRNA-146a-5p (12), miR-433 and miR-133b (13) and miR-410 (14). In a recent study, in an in
vitro model of PD induced by rotenone, miR-384-5p expression
was increased, and the downregulation of this miRNA exerted a
neuroprotective effect against rotenone by targeting GRP78
(15). However, the role of
miR-384-5p in the in vivo model of PD and other molecules
that may be regulated by miR-384-5p have yet to be
investigated.
This study found that the swimming and traction
scores of the mice with rotenone-induced PD were reduced, while the
α-synuclein levels were increased by rotenone in the mice with PD
compared with the DMSO group. Moreover, the α-synuclein levels were
increased by rotenone in the SH-SY5Y cells in the PD group compared
with the DMSO group, indicating the successful establishment of the
in vivo and in vitro models of PD, respectively.
Thereafter, increased miR-384-5p levels were found
in the in vivo and in vitro PD groups compared with
the DMSO group, which was in line with the findings of a previous
study (15). miR-384-5p was found
to target SIRT1 in 293 cells and SH-SY5Y cells in the present
study.
SIRT1 has been reported to be downregulated in PD
(22). Consistently, this study
found that the SIRT1 levels were decreased in the in vivo
and in vitro PD groups compared with the DMSO group;
moreover, a negative correlation was found between miR-384-5p and
SIRT1 in mice with rote-none-induced PD and in SH-SY5Y cells
exposed to rotenone.
SIRT1 affects mitochondrial-related apoptotic
signaling pathways (23); and
reduces oxidative stress-induced neuronal cell death in PD
(22). SIRT1 also mediates
salidroside-elicited protective effects against the
MPP+-induced apoptosis of SH-SY5Y cells (24), regulates the development of PD and
is targeted by miR-9-5p (25).
This study found that rotenone induced a significant increase in
the apoptosis of SH-SY5Y cells compared with the DMSO group, and
these findings are in agreement with those of previous studies
showing the role of SIRT1 in apoptosis in PD (22,24,25).
Subsequently, this study aimed to explore the
molecules that are associated with SIRT1 in mice with
rotenone-induced PD and in SH-SY5Y cells exposed to rotenone. SIRT1
induces the function of p53 (26); SIRT1 has also been shown to
inhibit the H2O2-induced apoptosis of
osteoblasts via the FOXO1/β-catenin pathway (27). FOXOs and p53 are regulated by
SIRT1 under conditions of oxidative stress (28); p53 and FOXO1 have been shown to be
associated with SIRT1 in MPP+-induced SH-SY5Y cells
(21). Consistently, this study
found that in the in vivo and in vitro models of PD,
the mRNA and protein levels of p53/FOXO1 became significantly
higher than those in the DMSO group.
In conclusion, this study demonstrates that
miRNA-384-5p promotes the progression of PD by targeting SIRT1,
providing a potential therapeutic target for the treatment of PD.
However, there is a limitation to the present study: The detailed
mechanisms of SIRT1 as regards cell apoptosis were not elucidated.
Thus, further studies are required to investigate this in the
future.
Acknowledgments
Not applicable.
Funding
The current study was funded by the Science and
Technology Plan of Shaanxi Provincial Health Planning Commission
(2016D065).
Availability of materials and data
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
HT and YL carried out the experiments and analyzed
the data. YH designed the study, analyzed the data, and prepared
the manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was performed in accordance with
the guidelines of the Committee on Care and Use of Experimental
Animals Resources and with the approval of Ethical Committee for
The First Hospital of Yulin.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
de Rijk MC, Launer LJ, Berger K, Breteler
MM, Dartigues JF, Baldereschi M, Fratiglioni L, Lobo A,
Martinez-Lage J, Trenkwalder C and Hofman A: Prevalence of
Parkinson's disease in Europe: A collaborative study of
population-based cohorts. Neurologic Diseases in the Elderly
Research Group. Neurology. 54:S21–S23. 2000.PubMed/NCBI
|
|
2
|
de Lau LM and Breteler MM: Epidemiology of
Parkinson's disease. Lancet Neurol. 5:525–535. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dickson DW, Braak H, Duda JE, Duyckaerts
C, Gasser T, Halliday GM, Hardy J, Leverenz JB, Del Tredici K,
Wszolek ZK and Litvan I: Neuropathological assessment of
Parkinson's disease: Refining the diagnostic criteria. Lancet
Neurol. 8:1150–1157. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jankovic J: Parkinson's disease: Clinical
features and diagnosis. J Neurol Neurosurg Psychiatry. 79:368–376.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Seidl SE, Santiago JA, Bilyk H and
Potashkin JA: The emerging role of nutrition in Parkinson's
disease. Front Aging Neurosci. 6:362014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
White AR, Kanninen KM and Crouch PJ:
Metals and neurode-generation: Restoring the balance. Front Aging
Neurosci. 7:1272015. View Article : Google Scholar
|
|
7
|
Allen MT and Levy LS: Parkinson's disease
and pesticide exposure-a new assessment. Crit Rev Toxicol.
43:515–534. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hutvágner G and Zamore PD: A microRNA in a
multiple-turnover RNAi enzyme complex. Science. 297:2056–2060.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zeng Y and Cullen BR: Sequence
requirements for micro RNA processing and function in human cells.
RNA. 9:112–123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wienholds E and Plasterk RH: MicroRNA
function in animal development. FEBS Lett. 579:5911–5922. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thome AD, Harms AS, Volpicelli-Daley LA
and Standaert DG: MicroRNA-155 regulates alpha-synuclein-induced
infammatory responses in models of Parkinson disease. J Neurosci.
36:2383–2390. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caggiu E, Paulus K, Mameli G, Arru G,
Sechi GP and Sechi LA: Differential expression of miRNA 155 and
miRNA 146a in Parkinson's disease patients. eNeurologicalSci.
13:1–4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang X, Yang R, Hu BL, Lu P, Zhou LL, He
ZY, Wu HM and Zhu JH: Reduced circulating levels of miR-433 and
miR-133b are potential biomarkers for Parkinson's disease. Front
Cell Neurosci. 11:1702017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ge H, Yan Z, Zhu H and Zhao H: MiR-410
exerts neuroprotective effects in a cellular model of Parkinson's
disease induced by 6-hydroxydopamine via inhibiting the
PTEN/AKT/mTOR signaling pathway. Exp Mol Pathol. 109:16–24. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang M, Yun Q, Shi F, Niu G, Gao Y, Xie S
and Yu S: Downregulation of miR-384-5p attenuates rotenone-induced
neurotoxicity in dopaminergic SH-SY5Y cells through inhibiting
endoplasmic reticulum stress. Am J Physiol Cell Physiol.
310:C755–C763. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Inden M, Kitamura Y, Abe M, Tamaki A,
Takata K and Taniguchi T: Parkinsonian rotenone mouse model:
Reevaluation of long-term administration of rotenone in C57BL/6
mice. Biol Pharm Bull. 34:92–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Haobam R, Sindhu KM, Chandra G and
Mohanakumar KP: Swim-test as a function of motor impairment in MPTP
model of Parkinson's disease: A comparative study in two mouse
strains. Behav Brain Res. 163:159–167. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai SF, Han G, Li Y, Yu D, Zhang D, Feng
Y, Zhao JI and Sun Y: Effects of nicotine on the microglia of
Parkinson's disease mice. Asian J Pharmacodyn Pharmacokin.
8:319–323. 2008.
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Lin TK, Chen SD, Chuang YC, Lin HY, Huang
CR, Chuang JH, Wang PW, Huang ST, Tiao MM, Chen JB and Liou CW:
Resveratrol partially prevents rotenone-induced neurotoxicity in
dopaminergic SH-SY5Y cells through induction of heme oxygenase-1
dependent autophagy. Int J Mol Sci. 15:1625–1646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Salimian N, Peymani M, Ghaedi K and Nasr
Esfahani MH: Modulation in miR-200a/SIRT1axis is associated with
apoptosis in MPP+-induced SH-SY5Y cells. Gene.
674:25–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Singh P, Hanson PS and Morris CM: SIRT1
ameliorates oxida-tive stress induced neural cell death and is
down-regulated in Parkinson's disease. BMC Neurosci. 18:462017.
View Article : Google Scholar
|
|
23
|
Zhou Y, Wang S, Li Y, Yu S and Zhao Y:
SIRT1/PGC-1α signaling promotes mitochondrial functional recovery
and reduces apoptosis after intracerebral hemorrhage in rats. Front
Mol Neurosci. 10:4432018. View Article : Google Scholar
|
|
24
|
Wang CY, Sun ZN, Wang MX and Zhang C:
SIRT1 mediates salidroside-elicited protective effects against
MPP+ -induced apoptosis and oxidative stress in SH-SY5Y
cells: involvement in suppressing MAPK pathways. Cell Biol Int.
42:84–94. 2018. View Article : Google Scholar
|
|
25
|
Wang Z, Sun L, Jia K, Wang H and Wang X:
miR-9-5p modulates the progression of Parkinson's disease by
targeting SIRT1. Neurosci Lett. 701:226–233. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee JT and Gu W: SIRT1: Regulator of p53
deacetylation. Genes Cancer. 4:112–117. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yao H, Yao Z, Zhang S, Zhang W and Zhou W:
Upregulation of SIRT1 inhibits H2O2-induced osteoblast apoptosis
via FoxO1/β-catenin pathway. Mol Med Rep. 17:6681–6690.
2018.PubMed/NCBI
|
|
28
|
Hori YS, Kuno A, Hosoda R and Horio Y:
Regulation of FOXOs and p53 by SIRT1 modulators under oxidative
stress. PLoS One. 8:e738752013. View Article : Google Scholar : PubMed/NCBI
|