Introduction
Sepsis is a major cause of admission to the
intensive care unit and mortality in the clinic (1), and it is usually characterized by
dysregulation of the host response followed by infection, which can
lead to organ failure and death (2). Currently, sepsis and its associated
shock are major healthcare problems affecting millions of people
worldwide each year. In Germany, the percentage of patients with
severe sepsis was reported to rise from 27% in 2007 to >40% in
2013, and the in-hospital morality of sepsis rose from 2.7% to ~24%
over the same period (3). Among
these patients, at least one in four succumb to sepsis each year
(4). Although there has been much
focus on awareness campaigns and clinician-friendly care bundles,
compliance with all six necessary steps within the first hour of
admission remains unsatisfactory (5). Therefore, it is important to further
investigate the exact molecular mechanism of sepsis. Interactions
among the epithelium, local immune system and microbiome are
essential for the maintenance of host health, and sepsis alters the
intestinal environment to promote epithelial cell dysfunction
(6). Apoptosis is considered to
serve an important role in sepsis, which leads directly to
metabolism dysfunction and organ failure (7,8),
and results in severe immunosuppression (9,10).
Therefore, it is essential to uncover the regulatory network of
epithelial cell apoptosis in sepsis as it may provide an effective
strategy for sepsis treatment.
MicroRNAs (miRNAs) are small non-coding RNAs that
are ~22 nucleotides in length (11). A recent study has revealed that
miRNAs serve critical roles in the progression of sepsis (12). Gao et al (13) reported that miR-146 attenuates
cardiac dysfunction in polymicrobial sepsis by targeting
interleukin-1 receptor-associated kinase 1 and TNF
receptor-associated factor 6. Roderburg et al (14) demonstrated that circulating
miR-150 serum levels predict the survival of patients with sepsis.
Another study also confirmed that levels of circulating miR-133a
serve as a biomarker to predict mortality in patients with severe
sepsis (15). These findings
suggest that miRNAs serve critical roles in the development of
sepsis. miR-195 is regarded as a tumour suppressor in cancer
development and progression (16). Almeida et al (17) reported that miR-195 regulates
proliferation, osteogenesis and paracrine activity during
angiogenesis in primary mesenchymal stromal/stem cells. In
addition, Wu et al (18)
revealed that the expression level of miR-195 is significantly
increased in whole blood samples from a sepsis model compared with
samples from a sham-operated group. However, to the best of our
knowledge, the biological role of miR-195 in the development of
sepsis remains largely unknown.
Sirtuin 1 (SIRT1) is a member of the mammalian
sirtuin family, which is a conserved family of
NAD+-dependent deacetylases and ADP-ribosyltransferases
that serve critical roles in several cellular processes, including
DNA repair, gene expression and metabolic regulation (19). A previous study revealed that
SIRT1 restrains lung inflammasome activation in a sepsis animal
model (20). Li et al
(21) demonstrated that
resveratrol relieves acute lung injury in a lipopolysaccharide
(LPS)-induced sepsis animal model by activating SIRT1. Furthermore,
enhancing the activation of SIRT1 markedly alters the transcription
profiles in experimental sepsis models (22). Considering these findings, SIRT1
may serve a critical role in the process of sepsis; however, to the
best of our knowledge, no studies on the role of miR-195 in the
regulation of SIRT1 in sepsis have been conducted.
The present study investigated the function of
miR-195 in an LPS-induced cell model of sepsis and the mechanism
through which miR-195 modulates SIRT1 expression. The current study
demonstrated that miR-195 promoted cell injury in sepsis via
targeting SIRT1/eukaryotic translation initiation factor 2A (eIF2a)
signalling, which suggests that miR-195 may be a potentially
effective therapeutic target for patients with sepsis.
Materials and methods
Cell culture
NCM460 cells were obtained from American Type
Culture Collection. Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Sigma-Aldrich; Merck KGaA), 100
U/ml streptomycin (Thermo Fisher Scientific, Inc.) and 100 U/ml
penicillin (Thermo Fisher Scientific, Inc.) at 37°C with a 5%
CO2 atmosphere in a humidified incubator. Sodium
butyrate (purity >98.5) was purchased from Sigma-Aldrich; Merck
KGaA. When cells reached >80% confluence, they were trypsinized
with 0.25% trypsin-EDTA (Sigma-Aldrich; Merck KGaA).
Cell viability detection
Cell viability was determined using a Cell Counting
kit-8 (CCK-8) assay. Briefly, NCM460 cells were seeded in a 96-well
plate at a density of 3×103/well and maintained in DMEM
for 24 h at 37°C. Subsequently, the cells were exposed to LPS (from
Escherichia coli 0111:B4; Sigma-Aldrich; Merck KGaA) at
concentrations of 0, 1, 5, 10, 25, 50 and 100 µg/ml.
Following incubation for 24 h at 37°C, 10 µl/well CCK-8
solution (Dojindo Molecular Technologies, Inc.) was added to the
medium and cultured for 3 h at 37°C. Subsequently, the plate was
agitated for 5 min, and the absorbance of each well at 450 nm was
determined using an automatic microplate reader (Multiskan MK3;
Thermo Fisher Scientific, Inc.). Each sample was analysed in
triplicate, and the mean absorbance was calculated as the final
result.
Evaluation of apoptosis by flow
cytometry
Firstly, 2×106 cells/well were seeded
into a six-well plate for 24 h and then exposed to LPS at
concentrations of 0, 1, 5 or 10 µg/ml for 24 h at 37°C.
Next, the cells were washed with PBS three times and immediately
fixed with 4% paraformaldehyde at 4°C for 15 min. Then, the cells
were stained with an Annexin V-FITC/propidium iodide double
staining apoptosis detection kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Subsequently, the
apoptosis of cells was determined by an Accuri C6 flow cytometer
(BD Biosciences) using FlowJo 7.2 software (FlowJo, LLC). Each
experiment was performed in triplicate, and the mean value was
calculated as the final result.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
After 24 h of transfection of miR-195 inhibitor, the
cell medium was removed and total RNA was isolated using
TRIzol® reagent (Takara Bio, Inc.) according to the
manufacturer's protocol, and quantified using an SSP-3000 Nanodrop
Spectrophotometer (Infinigen Biotechnology, Inc.). Subsequently, 1
µg RNA was reverse transcribed into complementary DNA using
a Reverse Transcription kit (Takara Bio, Inc.), according to the
manufacturer's protocol. Subsequently, the samples were amplified
and analysed using a SYBR Premix Ex Taq™ kit (cat. no. RR820A;
Takara Bio, Inc.) and an Applied Biosystems 7500 system (Thermo
Fisher Scientific, Inc.). For mRNA, GAPDH was used as the internal
control, and for miRNA, U6 was used as the internal control. The
fold-change in gene expression was calculated using the
2−∆∆Cq method (23).
The thermocycling conditions were as follows: 95°C for 3 min,
followed by 35 cycles of 95°C for 30 sec, 59°C for 45 sec and 72°C
for 40 sec. All PCR assays were replicated at least three times.
The primers used in the RT-qPCR assays were as follows: miR-195
forward, 5′-GGC TAG CAG CAC AGA AAT-3′ and reverse, 5′-GTG CAG GGT
CCG AGG T-3′; SIRT1 forward, 5′-CCA GAT CCT CAA GCC ATG T-3′ and
reverse, 5′-TTG GAT TCC TGC AAC CTG-3′; U6 forward, 5′-CTC GCT TCG
GCA GCA CAT ATA CT-3′ and reverse, 5′-ACG CTT CAC GAA TTT GCG TGT
C-3′; and GAPDH forward, 5′-AAT GGG CAG CCG TTA GGA AA-3′ and
reverse, 5′-TGA AGG GGT CAT TGA TGG CA-3′.
Plasmid construction and cell
transfection
miR-195 mimic (5′-UAG CAG CAC AGA AAU GGC-3′),
antagomir (5′-GCC AAU AUU UCU GUG CUG CUA-3′) and scrambled
negative control (NC) miRNAs (NC mimic, 5′-UUC UCC GAA CGU GUC ACG
UTT-3′ and NC antagomir, 5′-CAG UAC UUU UGU GUA GUA CAA-3′) were
purchased from GenePharma Co., Ltd. Cells were seeded into 96-well
plates and cultured to 70-80% confluence. The cells were then
transfected with miRNA mimic (100 nM) or antagomir (200 nM) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. A
total of 48 h after transfection, the cells were harvested.
Luciferase activity reporter assay
Bioinformatics analysis was performed to search for
the potential targets of miR-195. TargetScan 5.0 software
(http://www.targetscan.org/) validated
SIRT1 as a putative target of miR-195. A luciferase vector
including the 3′-untranslated region (3′-UTR) of human SIRT1 (WT
luc-SIRT1), which contains the SIRT1-miR-195 response elements, was
purchased from Addgene, Inc. A mutant containing two SIRT1-miR-195
response elements in the 3′-UTR of SIRT1 (mutant luc-SIRT1) was
generated using site-directed gene mutagenesis. The mutant
sequences were designed as follows: 5′-UAA UAU UUU GGA Cug cugUU-3′
(the five lowercase nucleotides were deleted) and 5′-TAA AGT ATT
CCT CTG TAC GAT-3′ (the four italic nucleotides were
substituted for TGCT). A reporter vector consisting of a luciferase
gene followed by the miR-195 binding consensus sequence was
purchased from Signosis, Inc. NCM460 cells (2×105 /well)
were seeded in a 12-well plate for 24 h. Subsequently, 200 ng WT
luc-SIRT1 or mutant luc-SIRT1 and miR-195-5p mimic, antagomir or
scrambled oligonucleotide were co-transfected into NCM460 cells
using Lipofectamine® 2000, according to the
manufacturer's protocol. A pRL-CMV vector containing the CMV
enhancer and early promoter elements, which express Renilla
luciferase, served as the internal control. Luciferase activity was
measured using a dual luciferase reporter assay system (Progema
Corporation) 24 h after transfection. Renilla luciferase
activity was used as the internal control.
Western blot analysis
Following treatment, the medium was removed and the
cells were lysed using RIPA lysis buffer (Sigma-Aldrich; Merck
KGaA) at 4°C for 15 min. The cell lysates were then harvested and
centrifuged for 10 min at 11,000 × g at 4°C. The supernatants were
collected and quantified using the BCA method. Subsequently, 30
µg total protein from each sample was separated by 10%
SDS-PAGE and transferred to PVDF membranes. Next, the PVDF
membranes were blocked with 5% non-fat milk dissolved in TBS and
20% Tween-20 (TBST) at room temperature for 1 h. After rinsing with
TBST three times for 5 min each time, the membranes were incubated
at 4°C overnight with specific antibodies against the following:
(1:100; cat. no. sc-74465, Santa Cruz Biotechnology, Inc.), Bcl-2
(1:1,000; cat. no. 15071, Cell Signaling Technology, Inc.), Bax
(1:1,000; cat. no. 14796, Cell Signaling Technology, Inc.),
phosphorylated-eIF2a (1:1,000; cat. no. 3398, Cell Signaling
Technology, Inc.), eIF2a (1:1,000; cat. no. 5324, Cell Signaling
Technology, Inc.), activating transcription factor 4 (ATF4;
1:1,000; cat. no. 11815, Cell Signaling Technology, Inc.), C/EBP
homologous protein (CHOP; 1:1,000; cat. no. 2895, Cell Signaling
Technology, Inc.) GRP78 (1:1,000; cat. no. ab28615, Abcam), growth
arrest and DNA damage-inducible protein (GADD34; cat. no. ab9869,
1:1,000; Abcam), activating transcription factor 6 (ATF6; 1:1,000;
cat. no. ab37149, Abcam), IRE1 (1:1,000; cat. no. ab37073, Abcam)
and GAPDH (1:5,000; cat. no. 5174, Cell Signaling Technology,
Inc.). After rinsing with TBST three times for 5 min each time, the
PVDF membranes were incubated with horseradish
peroxidase-conjugated goat-anti-rabbit secondary antibody (1:2,000;
cat. no. 32460; Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 1 h. Subsequently, the membranes were rinsed with
TBST three times (10 min each time) and visualized using an ECL
plus kit (GE Healthcare). Data were analyzed by ImageJ software
(version 1.6; National Institutes of Health).
Statistical analyses
In the current study, each experiment was repeated
at least three times with consistent results. All data are
presented as the mean ± standard deviation, and were analyzed using
GraphPad Prism (version 7.0; GraphPad Software, Inc.). One-way
ANOVA followed by the Newman-Keuls test was conducted for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
LPS treatment upregulates the expression
of miR-195 in NCM460 cells
To investigate the effects of LPS on NCM460 cells,
cell viability after treatment with LPS was detected using a CCK-8
assay. LPS significantly decreased the viability of NCM460 cells in
a dose-dependent manner (Fig.
1A). According to the data, three LPS concentrations (1, 5 and
10 µg/ml) were selected for further investigation.
Subsequently, the flow cytometry assay also demonstrated that LPS
could promote the apoptosis of NCM460 cells with increasing LPS
concentrations (Fig. 1B and C).
In addition, the expression pattern of miR-195 was determined. As
presented in Fig. 1D, LPS
treatment significantly induced the upregulation of miR-195 in
NCM460 cells. Thus, these data indicated that LPS treatment
inhibited NCM460 cells and increased miR-195 expression, which may
indicate that miR-195 serves an important role in LPS-induced
apoptosis of NCM460 cells.
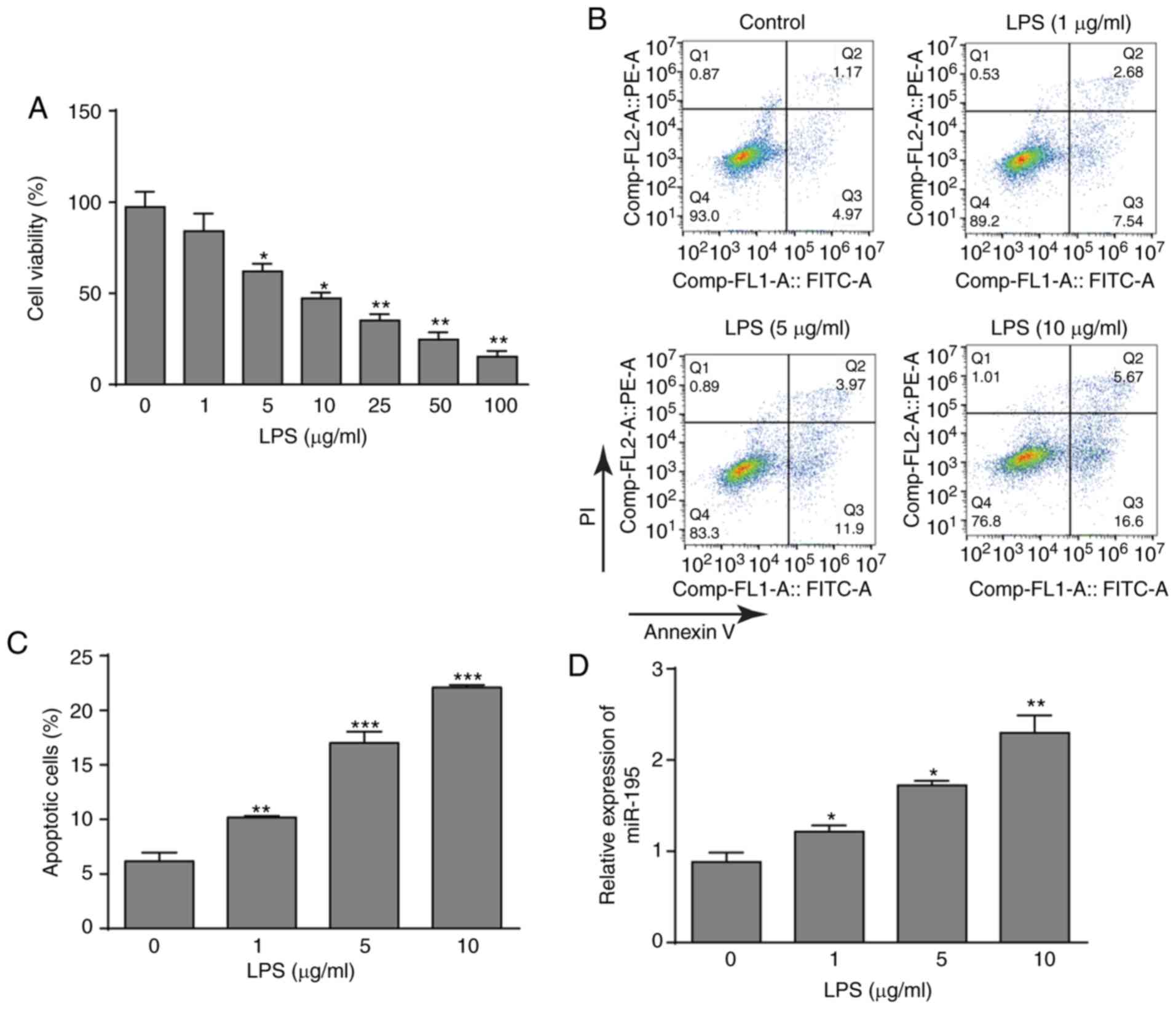 | Figure 1Effects of LPS on cell viability,
apoptosis and miR-195 expression. (A) Cultured NCM460 cells were
incubated with LPS (0, 1, 5, 10, 25, 50 or 100 µg/ml) for 24
h, and cell viability was then examined by CCK-8 assay. (B)
Apoptosis of NCM460 cells following exposure to LPS (0, 1, 5 or 10
µg/ml) was detected by flow cytometry. (C) Quantification of
the LPS-induced apoptosis rate of NCM460 cells. (D) miR-195
expression level in NCM460 cells following exposure to LPS (0, 1, 5
or 10 µg/ml) was examined by reverse
transcription-quantitative PCR and compared with that in the
control group. The miRNA expression level is expressed as a ratio
of U6. *P<0.05, **P<0.01 and
***P<0.001 vs. control (0 µg/ml). LPS,
lipopolysaccharide; miR-195, microRNA-195; PI, propidium
iodide. |
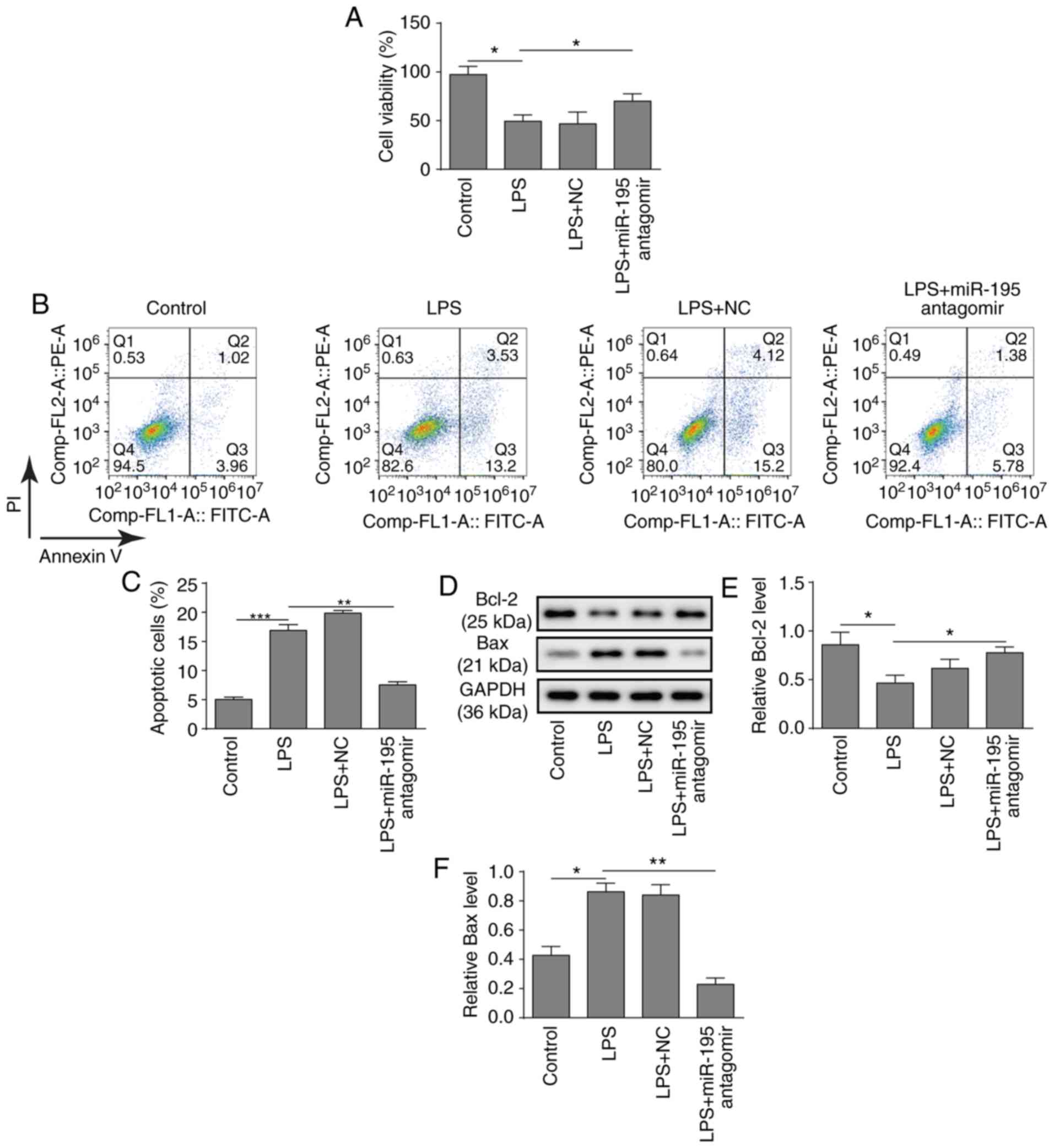
Silencing miR-195 inhibits the apoptosis
of NCM460 cells
To further investigate the potential role of
miR-195, a miR-195 antagomir was applied to silence the expression
of miR-195 in NCM460 cells. Subsequently, the biological behaviours
of NCM460 cells were investigated. First, a CCK-8 assay
demonstrated that silencing miR-195 significantly relieved the
inhibition of cell viability induced by LPS in NCM460 cells
(Fig. 2A). Consistently,
silencing miR-195 also significantly decreased the apoptosis of
NCM460 cells induced by LPS (Fig. 2B
and C). Furthermore, a western blot analysis of
apoptosis-related proteins revealed that LPS treatment
significantly inhibited the expression of Bcl-2 and promoted the
expression of Bax, while silencing miR-195 significantly increased
the expression of Bcl-2 and suppressed the expression of Bax
(Fig. 2D-F). In summary, these
findings suggest that silencing miR-195 could attenuate the
apoptosis of NCM460 cells induced by LPS.
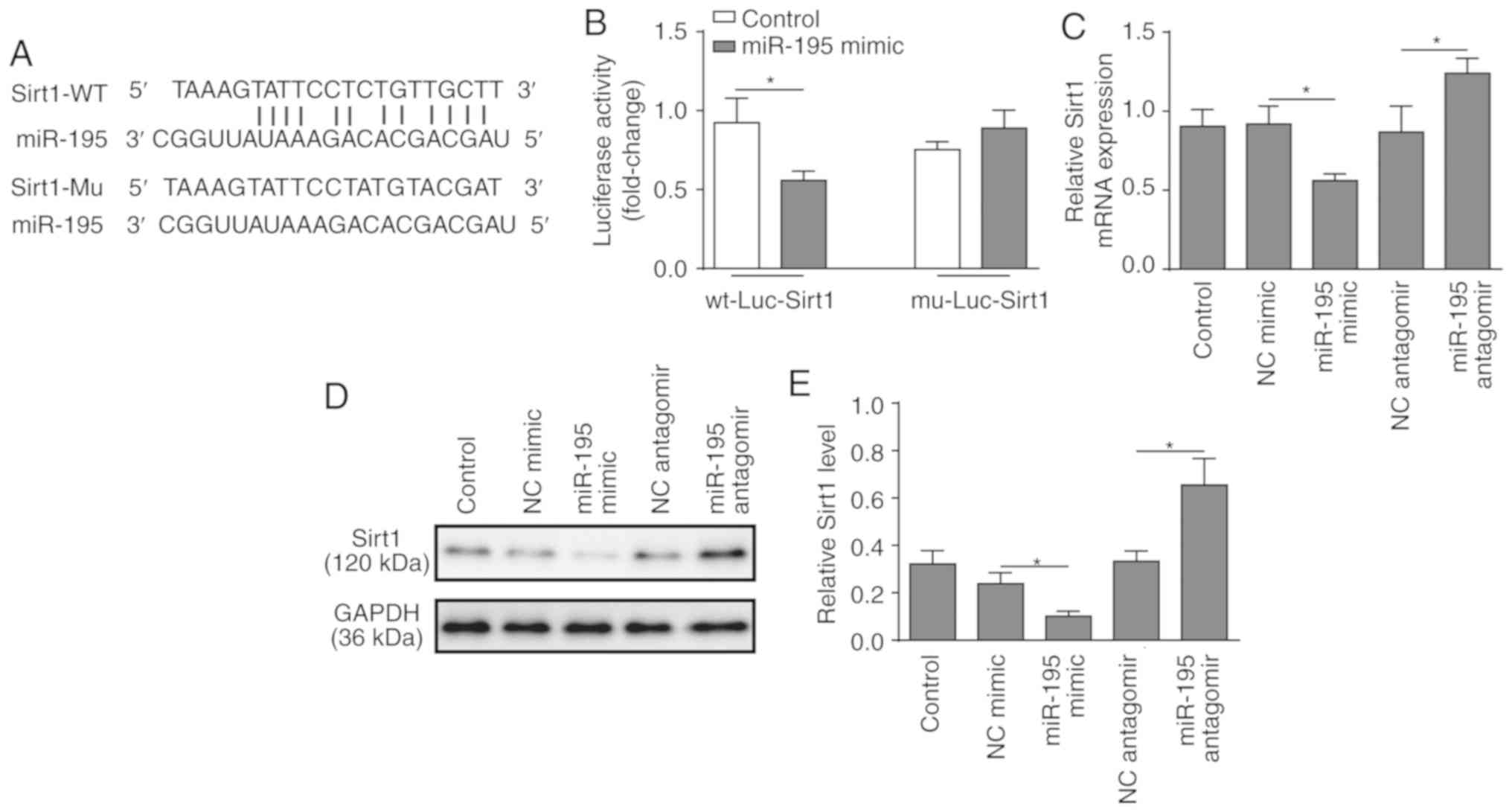
miR-195 directly regulates the expression
of SIRT1
SIRT1 was identified as a predicted target of
miR-195 using bioinformatics analysis (Fig. 3A). To verify the association
between miR-195 and SIRT1, a dual-luciferase reporter assay was
performed. miR-195 mimic and luciferase reporter plasmids with
cloned miR-195 binding sites of WT SIRT1-3′UTR or MUT SIRT1-3′UTR
were co-transfected. The results revealed that miR-195 mimic
significantly reduced the lucif-erase activity of the WT
SIRT1-3′UTR, but had no effect on the MUT SIRT1-3′UTR (Fig. 3B). Following that, loss or
gain-of-function of miR-195 was employed using miR-195 mimic and
antagomir (Fig. S1). RT-qPCR and
western blot analyses demonstrated that silencing miR-195
significantly increased the expression of SIRT1, while
overexpression of miR-195 significantly suppressed the expression
of SIRT1 (Fig. 3C-E). These
findings support the hypothesis that SIRT1 is a direct target of
miR-195.
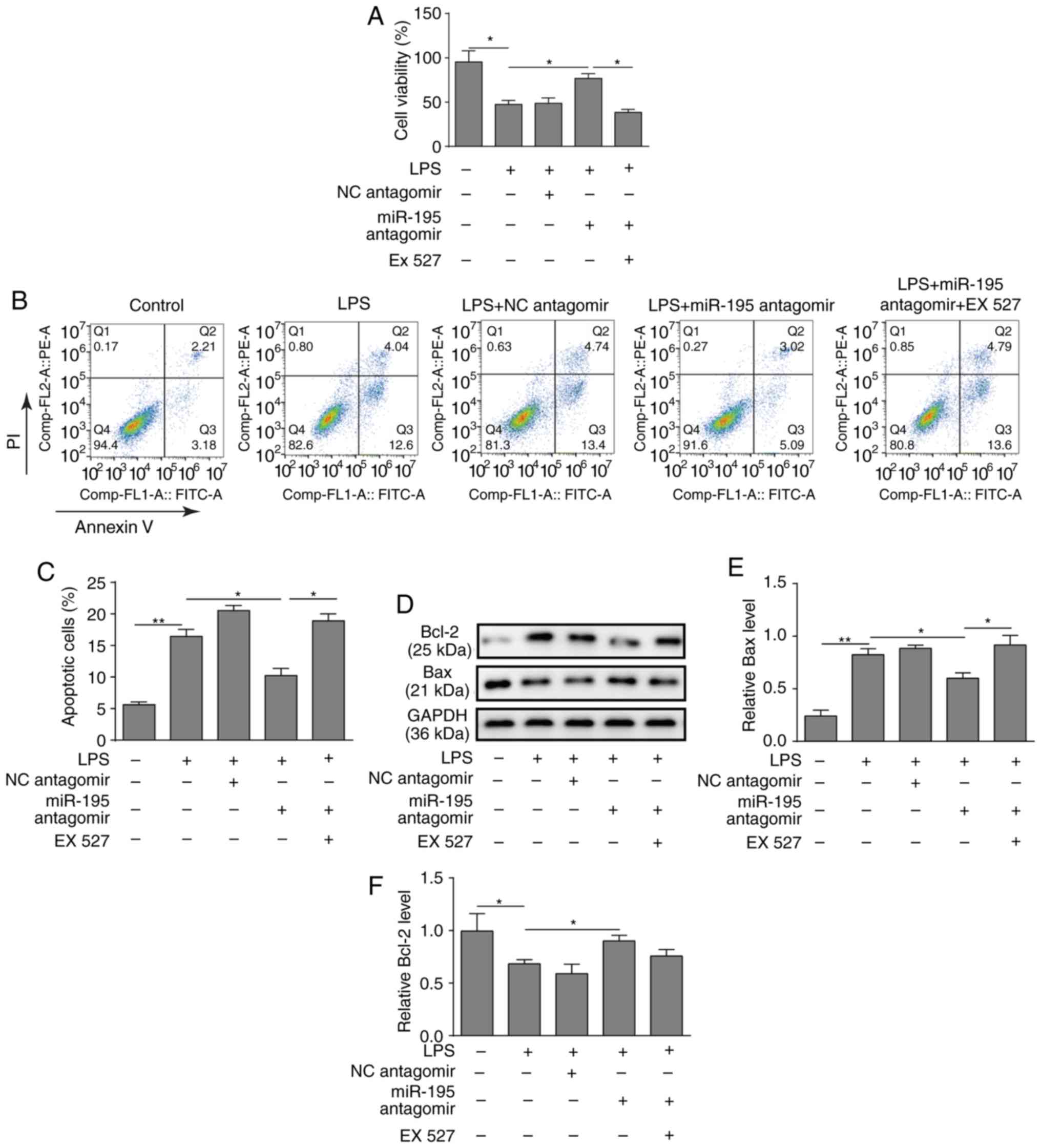
Inhibition of SIRT1 reverses the effects
of miR-195 antagomir on LPS-induced NCM460 cells
To investigate the function of the miR-195/SIRT1
regulatory network in LPS-induced apoptosis of intestinal
epithelial cells, NCM460 cells were pre-treated with the SIRT1
inhibitor EX527. As presented in Fig.
4A, the results of the CCK-8 assay demonstrated that silencing
miR-195 increased the viability of NCM460 cells treated with LPS,
while the SIRT1 inhibitor EX527 inhibited this effect (Fig. 4A). Similarly, a flow cytometry
assay revealed that the SIRT1 inhibitor EX527 reversed the
protective effects of miR-195-silencing on NCM460 cell survival
(Fig. 4B and C). Furthermore,
western blotting demonstrated that silencing miR-195 could promote
the expression of Bcl-2 but reduce the expression of Bax, while
application of EX527 significantly reversed the effects on Bax
level (Fig. 4D-F).
miR-195 promotes apoptosis of NCM460
cells via targeting SIRT1/eIF2a
A previous study indicated that SIRT1 serves a
protective role in myocardial cells by regulating eIF2a and its
downstream effectors (24).
Therefore, the signalling pathway of SIRT1/eIF2a was investigated
in the endoplasmic reticulum (ER) stress response of NCM460 cells
in the present study. Western blot analysis revealed that silencing
miR-195 could inhibit apoptosis derived from ER stress via
upregulating the expression of SIRT1 and downregulating the
expression of ATF4, CHOP, GRP78 and GADD34 and reducing the
phosphorylation of eIF2a. However, the SIRT1 inhibitor EX527
significantly increased the expression of CHOP, GRP78 and GADD34,
as well as the phosphorylation of eIF2a, which reversed the
protective effects of miR-195-silencing (Fig. 5A and B). In addition, the other
two pathways (ATF6 and IRE1) of ER stress were further
investigated, and the results were similar to the PERK pathway
markers (Fig. S2A and B). Taken
together, it can be concluded that silencing miR-195 serves a
positive role in inhibiting NCM460 cell apoptosis via regulating
SIRT1/eIF2a. A schematic figure presenting the association of
miR-195, SIRT1 and ER stress is presented in Fig. 5C.
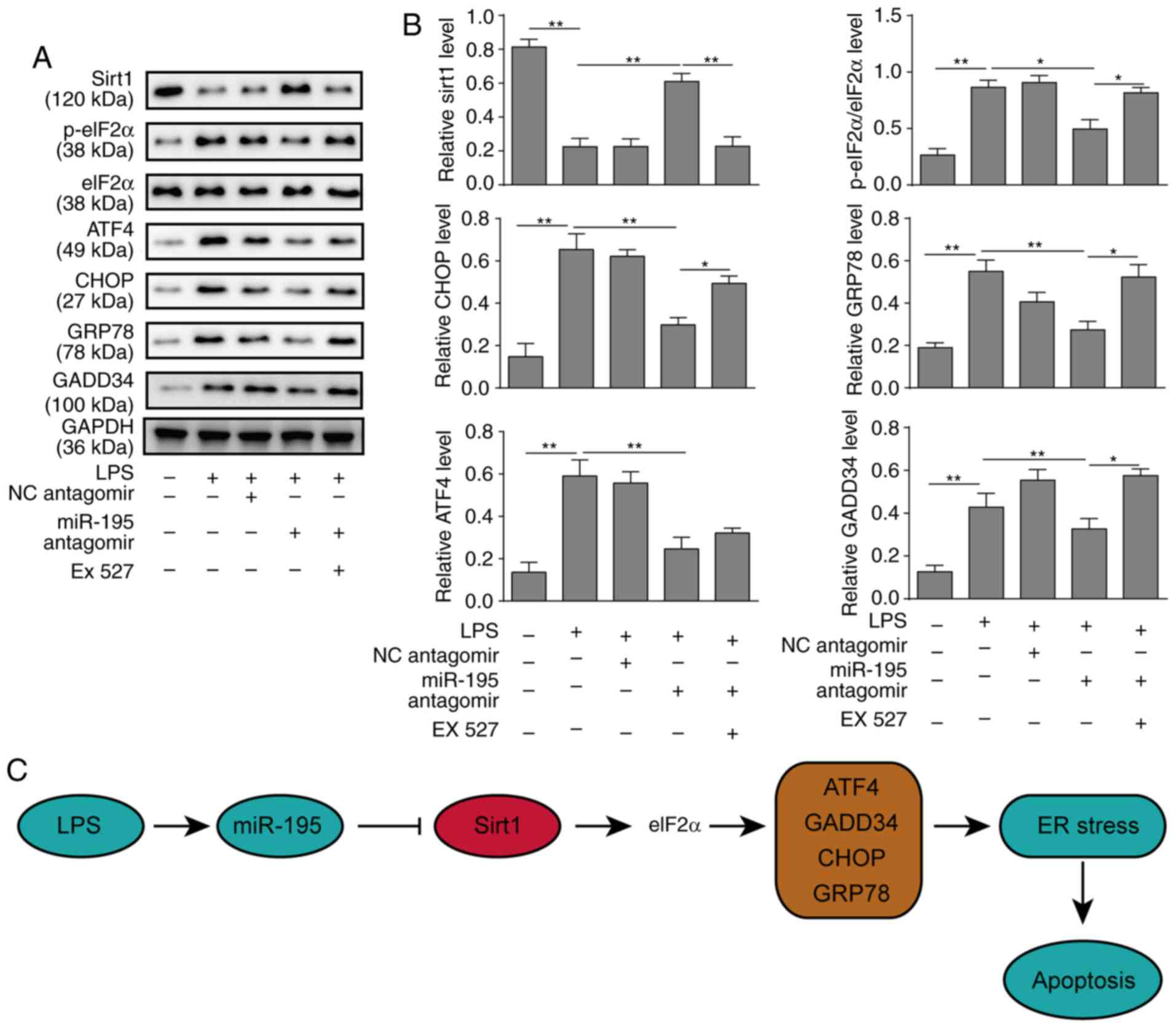 | Figure 5miR-195 promotes apoptosis of NCM460
cells via targeting SIRT1/eIF2a. NCM460 cells were transfected with
miR-195 antagomir or a scrambled oligonucleotide as a control. A
total of 24 h after transfection, the cells were incubated with
LPS. To further investigate the underlying mechanisms of miR-195,
the cells were incubated with the SIRT1 inhibitor EX527 after
transfection. (A) The expression levels of markers of the
SIRT1/eIF2a signalling pathway were measured by western blot
analysis. (B) Quantification of markers in the SIRT1/eIF2a
signalling pathway. (C) A schematic figure demonstrating the roles
of miR-195, Sirt and ER stress response. The data are presented as
the mean ± standard deviation from three independent experiments.
*P<0.05, **P<0.01. LPS,
lipopolysaccharide; NC, negative control; SIRT1, sirtuin 1;
miR-195, microRNA-195; p-, phosphorylated; eIF2a, eukaryotic
translation initiation factor 2A; ATF4, activating transcription
factor 4; CHOP, C/EBP homologous protein; GADD34, growth arrest and
DNA damage-inducible protein; ER, endoplasmic reticulum. |
Discussion
In the present study, miR-195 was identified to be
significantly upregulated in an LPS-induced cell sepsis model.
Furthermore, transfection with a miR-195 antagomir could
significantly decrease apoptosis but increase the viability of
intestinal epithelial NCM460 cells. Further analyses revealed that
miR-195 could bind to the 3′UTR of SIRT1 and regulate the
expression of SIRT1. In addition, silencing miR-195 protected the
viability of NCM460 cells treated with LPS, but the SIRT1 inhibitor
EX527 reversed this effect by silencing miR-195. These findings
indicate that miR-195 regulates the apoptosis of intestinal
epithelial cells in LPS-induced sepsis via SIRT1.
Sepsis is a major public health concern and is
usually characterized by intravascular or extravascular microbial
infection, systemic inflammation and microcirculatory dysfunction,
which results in tissue damage, organ failure or even death
(25). Notably, dysregulated
apoptosis and increased neutrophil function lead to immune and
organ dysfunction in sepsis and its associated multiple organ
failure (26). LPS, a membrane
component of gram-negative bacteria, is typically used to induce
sepsis in cell and animal models, in which pro-inflammatory
cytokines in the serum are increased with septic clinic
manifestations (27). A previous
review has documented that miRNAs, such as miR-195, serve critical
roles in the process of sepsis (28). Wu et al (18) revealed that miR-195 is
significantly upregulated in a caecal ligation and puncture-induced
experimental sepsis model. Another study revealed that serum
miR-195 serves as a biomarker for Chinese patients with sepsis. The
present study also detected upregulation of miR-195 in an
LPS-induced sepsis cell model, which was in line with other work.
Furthermore, Zheng et al (29) demonstrated that inhibition of
miR-195 suppresses apoptosis and multiple organ injury in a sepsis
mouse model. The present study demonstrated that silencing miR-195
could protect NCM460 cells from LPS-induced apoptosis. This
suggests that miR-195 may serve a critical role in the pathogenesis
of sepsis.
SIRT1 is a NAD+-dependent histone
deacetylase that serves critical roles in various biological
processes, including the ER stress response and cell survival.
During sepsis adaptation, SIRT1 switches monocyte energy sources
from glycolysis to fatty acid oxidation (30). Gao et al (20) revealed that SIRT1 could restrain
inflammasome activation in the lungs of a murine sepsis model. Li
et al (21) reported that
resveratrol attenuates acute lung injury via activating SIRT1 in
LPS-induced sepsis. Vachharajani et al (31) revealed that inhibition of SIRT1
improves immunity and outcomes of the hypo-inflammatory phenotype
of sepsis. Thus, accumulating evidence supports the protective role
of SIRT1 in sepsis. In addition, miR-195 has been reported to
regulate the initiation and development of tumours by targeting
multiple genes. For example, Zhou et al (32) demonstrated that miR-195 inhibits
cervical cancer migration and invasion via binding Smad3. miR-195
has also been identified to suppress colon cancer proliferation by
targeting Wnt family member 3A (33). Furthermore, Zhu et al
(34) have reported that miR-195
promotes palmitate-induced apoptosis in cardiomyocytes by reducing
the expression of SIRT1. Considering these findings, we speculated
whether there was such a similar connection between miR-195 and
SIRT1 in sepsis. The present study verified that SIRT1 is a target
of miR-195 in NCM460 cells, and inhibition of miR-195 could
significantly upregulate the expression of SIRT1. To further
confirm the underlying mechanism, the SIRT1 inhibitor EX527 was
used to investigate the biofunctional changes in LPS-treated NCM460
cells. The results demonstrated that silencing miR-195 could reduce
LPS-induced apoptosis in NCM460 cells, while pre-treatment with
EX527 could promote apoptosis. These findings demonstrated that
SIRT1 may be a critical downstream target of miR-195-mediated
apoptosis in intestinal epithelial cells.
Although the underlying mechanisms by which
silencing miR-195 affects SIRT1 and reduces apoptosis in sepsis
remain to be investigated, other studies may provide some
suggestions. Liu et al (35) identified that downregulating
Smad3/ATF4 is essential for the SIRT1 suppression of ER
stress-induced apoptosis. Furthermore, Koga et al (36) reported that ER stress promotes
hepatocellular injury via increasing the expression of SIRT1 in the
PI3K/Akt-GSK3β signalling pathway. Prola et al (37) demonstrated that SIRT1 attenuates
ER stress-induced cell apoptosis in cardiomyocytes via eIF2a
deacetylation. These findings suggest that SIRT1 may serve critical
roles in ER stress-induced apoptosis. In the present study, the
activation of the ER stress response was also revealed. The results
demonstrated that silencing miR-195 could significantly increase
the expression level of SIRT1, while decreasing the expression
levels of ATF4, CHOP, GRO78 and GADD34 and inhibiting the
phosphorylation of eIF2a. By contrast, inhibiting SIRT1 increased
the expression levels of ATF4, CHOP, GRO78 and GADD34 and the
phosphorylation of eIF2a. The present results confirmed that
miR-195 could promote cell apoptosis by downregulating the
SIRT1/eIF2a signalling pathway in the ER.
In summary, the present study revealed that
upregulation of miR-195 in a sepsis cell model could serve a
pivotal role in promoting intestinal epithelial cell apoptosis and
result in sepsis aggravation. Furthermore, molecular mechanism
experiments indicated that miR-195 directly targets SIRT1 and
promotes apoptosis in intestinal epithelial cells via the
SIRT1/eIF2a signalling pathway.
Supplementary Data
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China. (grant no. 81400031).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JGL is responsible for the integrity of the entire
study, designed the study, analyzed the data, performed statistical
analysis, and edited and reviewed the manuscript. YT designed the
study, performed the literature research and experimental studies,
acquired the data, performed data and statistical analyses, and
prepared the manuscript. SY performed the experimental studies and
statistical analysis, and acquired the data. SYD performed the
experimental studies. LZ performed the experimental studies and
data analysis, and prepared the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kaukonen KM, Bailey M, Pilcher D, Cooper
DJ and Bellomo R: Systemic inflammatory response syndrome criteria
in defining severe sepsis. New Engl J Med. 372:1629–1638. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thimmulappa RK, Lee H, Rangasamy T, Reddy
SP, Yamamoto M, Kensler TW and Biswal S: Nrf2 is a critical
regulator of the innate immune response and survival during
experimental sepsis. J Clin Invest. 116:984–995. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fleischmann C, Thomas-Rueddel DO, Hartmann
M, Hartog CS, Welte T, Heublein S, Dennler U and Reinhart K:
Hospital incidence and mortality rates of sepsis. Dtsch Arztebl
Int. 113:159–166. 2016.PubMed/NCBI
|
|
4
|
Rhodes A, Evans LE, Alhazzani W, Levy MM,
Antonelli M, Ferrer R, Kumar A, Sevransky JE, Sprung CL, Nunnally
ME, et al: Surviving sepsis campaign: International guidelines for
management of sepsis and septic shock: 2016. Crit Care Med.
45:486–552. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hunter JG, Pritchett C, Pandya D, Cripps A
and Langford R: Sim-sepsis: Improving sepsis treatment in the
emergency department? BMJ Simul Technol Enhanc Learn.
2018:bmjstel2018.
|
|
6
|
Fay KT, Ford ML and Coopersmith CM: The
intestinal micro-environment in sepsis. Biochim Biophysica Acta Mol
Basis Dis. 1863:2574–2583. 2017. View Article : Google Scholar
|
|
7
|
Yoseph BP, Klingensmith NJ, Liang Z, Breed
ER, Burd EM, Mittal R, Dominguez JA, Petrie B, Ford ML and
Coopersmith CM: Mechanisms of intestinal barrier dysfunction in
sepsis. Shock. 46:52–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gill SE, Rohan M and Mehta S: Role of
pulmonary microvascular endothelial cell apoptosis in murine
sepsis-induced lung injury in vivo. Respir Res. 16:1–13. 2015.
View Article : Google Scholar
|
|
9
|
Luan Y, Yao Y, Xiao X and Sheng Z:
Insights into the apoptotic death of immune cells in sepsis. J
Interferon Cytokine Res. 35:17–22. 2015. View Article : Google Scholar :
|
|
10
|
Delano MJ and Ward PA: Sepsis-induced
immune dysfunction: Can immune therapies reduce mortality? J Clin
Invest. 126:23–21. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fang W and Bartel DP: The menu of features
that define primary microRNAs and enable de novo design of microRNA
genes. Mol Cell. 60:131–145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Giza DE, Fuentesmattei E, Bullock MD,
Tudor S, Goblirsch MJ, Fabbri M, Lupu F, Yeung SJ, Vasilescu C and
Calin GA: Cellular and viral microRNAs in sepsis: Mechanisms of
action and clinical applications. Cell Death Differ. 23:1906–1918.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao M, Wang X, Zhang X, Ha T, Ma H, Liu L,
Kalbfleisch JH, Gao X, Kao RL and Williams DL: Attenuation of
cardiac dysfunction in polymicrobial sepsis by MicroRNA-146a is
mediated via targeting of IRAK1 and TRAF6 expression. J Immunol.
195:672–682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roderburg C, Luedde M, Vargas Cardenas DV,
Vucur M, Scholten D, Frey N, Koch A, Trautwein C, Tacke F and
Luedde T: Circulating MicroRNA-150 serum levels predict survival in
patients with critical illness and sepsis. PLoS One. 8:e546122013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tacke F, Roderburg C, Benz F, Cardenas DV,
Luedde M, Hippe HJ, Frey N, Vucur M, Gautheron J, Koch A, et al:
Levels of circulating miR-133a are elevated in sepsis and predict
mortality in critically ill patients. Crit Care Med. 42:1096–1104.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu J, Ji A, Wang X, Zhu Y, Yu Y, Lin Y,
Liu Y, Li S, Liang Z, Xu X, et al: MicroRNA-195-5p a new regulator
of Fra-1, suppresses the migration and invasion of prostate cancer
cells. J Transl Med. 13:2892015. View Article : Google Scholar
|
|
17
|
Almeida MI, Silva AM, Vasconcelos DM,
Almeida CR, Caires H, Pinto MT, Calin GA, Santos SG and Barbosa MA:
MiR-195 in human primary mesenchymal stromal/stem cells regulates
proliferation, osteogenesis and paracrine effect on angiogenesis.
Oncotarget. 7:7–22. 2016. View Article : Google Scholar :
|
|
18
|
Wu SC, Yang JC, Rau CS, Chen YC, Lu TH,
Lin MW, Tzeng SL, Wu YC, Wu CJ and Hsieh CH: Profiling circulating
MicroRNA expression in experimental sepsis using cecal ligation and
puncture. PLoS One. 8:e779362013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Price NL, Gomes AP, Ling AJ, Duarte FV,
Martin-Montalvo A, North BJ, Agarwal B, Ye L, Ramadori G, Teodoro
JS, et al: SIRT1 is required for AMPK activation and the beneficial
effects of resve-ratrol on mitochondrial function. Cell Metab.
15:675–690. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao R, Ma Z, Hu Y, Chen J, Shetty S and Fu
J: Sirt1 restrains lung inflammasome activation in a murine model
of sepsis. Am J Physiol Lung Cell Mol Physiol. 308:L847–L853. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li T, Zhang J, Feng J, Li Q, Wu L, Ye Q,
Sun J, Lin Y, Zhang M, Huang R, et al: Resveratrol reduces acute
lung injury in a LPS-induced sepsis mouse model via activation of
Sirt1. Mol Med Rep. 7:1889–1895. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Opal S, Ellis JL, Suri V, Freudenberg JM,
Vlasuk GP, Li Y, Chahin AB, Palardy JE, Parejo N, Yamamoto M, et
al: Sirt1 activation markedly alters transcription profiles and
improves outcome in experimental sepsis. Shock. 28:559–567.
2015.
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Prola A, Pires Da Silva J, Guilbert A,
Lecru L, Piquereau J, Ribeiro M, Mateo P, Gressette M, Fortin D,
Boursier C, et al: SIRT1 protects the heart from ER stress-induced
cell death through eIF2α deacetylation. Cell Death Differ.
24:343–356. 2017. View Article : Google Scholar
|
|
25
|
Djoumerska-Alexieva I, Pashova S, Vassilev
T and Pashov A: The protective effect of modified intravenous
immunoglobulin in LPS sepsis model is associated with an increased
IRA B cells response. Autoimmun Rev. 12:653–656. 2013. View Article : Google Scholar
|
|
26
|
Paunel-Görgülü A, Flohé S, Scholz M,
Windolf J and Lögters T: Increased serum soluble Fas after major
trauma is associated with delayed neutrophil apoptosis and
development of sepsis. Critical Care. 15:R202011. View Article : Google Scholar :
|
|
27
|
Cai B, Deitch EA and Ulloa L: Novel
insights for systemic inflammation in sepsis and hemorrhage. Med
Inflamm. 2010:6424622010. View Article : Google Scholar
|
|
28
|
Kingsley SMK and Bhat BV: Role of
microRNAs in sepsis. Inflamm Res. 66:1–17. 2017. View Article : Google Scholar
|
|
29
|
Zheng D, Yu Y, Li M, Wang G, Chen R, Fan
GC, Martin C, Xiong S and Peng T: Inhibition of MicroRNA 195
prevents apoptosis and multiple-organ injury in mouse models of
sepsis. J Infect. 213:1661–1670. 2016. View Article : Google Scholar
|
|
30
|
Liu TF, Vachharajani V, Millet P,
Bharadwaj MS, Molina AJ and Mccall CE: Sequential actions of
SIRT1-RELB-SIRT3 coordinate nuclear-mitochondrial communication
during immunometabolic adaptation to acute inflammation and sepsis.
J Biol Chem. 290:396–408. 2015. View Article : Google Scholar :
|
|
31
|
Vachharajani VT, Liu T, Brown CM, Wang X,
Buechler NL, Wells JD, Yoza BK and Mccall CE: SIRT1 inhibition
during the hypoinflammatory phenotype of sepsis enhances immunity
and improves outcome. J Leuk Biol. 96:785–796. 2014. View Article : Google Scholar
|
|
32
|
Zhou Q, Han LR, Zhou YX and Li Y: MiR-195
suppresses cervical cancer migration and invasion through targeting
smad3. Int J Gynecol Cancer. 26:817–824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li B and Wang S and Wang S: MiR-195
suppresses colon cancer proliferation and metastasis by targeting
WNT3A. Mol Genet Genomics. 293:1245–1253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu H, Yang Y, Wang Y, Li J, Schiller PW
and Peng T: MicroRNA-195 promotes palmitate-induced apoptosis in
cardiomyocytes by down-regulating Sirt1. Cardiovasc Res. 92:75–84.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu Z, Gu H, Lu G, Xu Y, Fei F, Saeed M
and Chao S: Reducing Smad3/ATF4 was essential for Sirt1 inhibiting
ER stress-induced apoptosis in mice brown adipose tissue.
Oncotarget. 8:9267–9279. 2016.PubMed/NCBI
|
|
36
|
Koga T, Suico MA, Shimasaki S, Watanabe E,
Kai Y, Koyama K, Omachi K, Morino-Koga S, Sato T, Shuto T, et al:
Endoplasmic reticulum (ER) stress induces sirtuin 1 (SIRT1)
expression via the PI3K-Akt-GSK3β signaling pathway and promotes
hepatocellular injury. J Biol Chem. 290:30366–30374. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Prola A, Silva JP, Guilbert A, Lecru L,
Piquereau J, Ribeiro M, Mateo P, Gressette M, Fortin D, Boursier C,
et al: SIRT1 protects the heart from ER stress-induced cell death
through eIF2α deacetylation. Cell Death. 24:343–356. 2016.
View Article : Google Scholar
|