Introduction
Circular RNAs (circRNAs), which are formatted by
connecting the 3′ and 5′ ends together via exon circularization or
intron circularization, are non-coding RNAs that serve as competing
endogenous RNAs (1,2). circRNAs are resistant to RNase R
degradation and this stability underlies their abundant expression
and evolutionary conservation in human and mice (3). The vast majority of circRNAs are
expressed in the cytoplasm and contain microRNA (miRNA) response
elements to interact with miRNAs and regulate target gene
expression during and subsequent to transcription and (4). In addition, circRNAs exhibit
tissue-specific expression profiles.
circRNAs serve important roles in cardiovascular
diseases, including myocardial infarction, cardiac senescence,
atherosclerosis and pathological hypertrophy (5). For instance, heart-associated
circRNA (HACR) inhibits miR-223 activity to enhance the protective
role of ARC, thereby preventing cardiac hypertrophy and cardiac
failure (6). During myocardial
infarction, circRNA Cdr1as activates SP1 and PARP by functioning as
a sponge for miR-7 (7).
Conversely, circ-Foxo3 may inhibit the nuclear translocation of
transcription factors and promote cardiac senescence (8). Accordingly, recent studies have
suggested that circRNAs may be used as biomarkers and therapeutic
targets for cardiovascular diseases (9,10).
circRNAs exert their functions by binding to miRNAs,
indicating that circRNAs may serve as upstream regulators of miRNAs
(11). Conversely, miRNAs control
cardiac function by targeting a number of mRNA to regulate the
proliferation and apoptosis of cells during disease progression
(12). In addition, circRNAs are
more stable and abundant than miRNAs in serum and other body
fluids, and in extracellular vesicles (13). Therefore, circRNAs are ideal
candidates as potential disease biomarkers and therapeutic targets
(14). However, the mechanisms
underlying the roles of circRNAs in cardiomyocyte death induced by
anoxia/reoxygenation (A/R) or ischemia/reperfusion (I/R) injury are
not yet fully understood.
The present study investigated the roles of
circRNA-101237, encoded by the cyclin dependent kinase 8 (CDK8)
gene, and let-7a-5p in cardiomyocyte death induced by A/R injury
and the underlying mechanisms. It was demonstrated that
circRNA-101237 mediated apoptosis in cardiomyocytes by activating
autophagy that was dependent on let-7a-5p/insulin-like growth
factor 2 mRNA-binding protein 3 (IGF2BP3) axis. The present study,
which identified circRNA_101237 as a novel circRNA, also revealed
that the circRNA-101237/let-7a-5p/IGF2BP3 axis may serve as a
regulator of cardiomyocyte death and a potential therapeutic target
for the management of cardiovascular diseases.
Materials and methods
Primary cardiomyocyte culture and
treatment
Animal experiments were approved by the Ethics
Committee for Animal Research of The People's Hospital of Guangxi
Zhuang Autonomous Region. Cardiomyocytes were isolated from
2-day-old mice as previously described (15). The isolated cardiomyocytes were
cultured in DMEM/F-12 (Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 5% heat-inactivated horse serum and diluted to
1×106 cells/ml for 24 h. Subsequently, the
cardiomyocytes were passaged in culture dishes coated with laminin.
For the A/R treatment model, anoxia was induced by exposure to 5%
CO2 and 95% N2 for 2 h, followed by
reoxygenation with 95% O2 and 5% CO2 for 4, 8
or 12 h.
circRNA microarray analysis
Cardiomyocytes underwent A/R treatment for 12 h and
RNA was extracted using the RNeasy Mini kit (Qiagen China Co.,
Ltd.) according to the manufacturer's protocol. The total RNA was
used to analyze the circRNA expression profiles using the Mouse
Circular RNA Array Service (Aksomics Inc.). Differentially
expressed circRNAs were identified as fold change ≥2 and
P<0.001.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Thermo Fisher Scientific, Inc.). Following treatment with
DNAse I (Thermo Fisher Scientific, Inc.), RNA was reverse
transcribed with reverse transcriptase (Thermo Fisher Scientific,
Inc.). RT-qPCR for mature miRNAs was conducted using an All-in-One™
miRNA RT-qPCR (GeneCopoeia, Inc.) following the manufacturer's
recommended protocol on a CFX96 Real-Time PCR Detection System
(Bio-Rad Laboratories, Inc.). U6 was used as internal control for
miRNA expression analysis. The expression of circRNAs and mRNA was
measured by SsoFast™ EvaGreen® supermix (cat no.
1725201; Bio-Rad Laboratories, Inc.), according to the
manufacturer's protocols. Expression of GAPDH was used as an
internal control. qPCR was performed with the following
thermocycling conditions: 95°C for 3 min, and 39 cycles of 95°C for
10 sec and 60°C for 30 sec. The data were analyzed via the
comparative quantification cycle method (2−ΔΔCq;
−ΔCq=Cqtarget gene−Cqinternal control), as
previously described (16). The
sequences of the circRNAs divergent primers, let-7a-5p, IGF2BP3 and
GAPDH primers are listed in Table
SI.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labelling
(TUNEL) assay
Apoptotic cells were analyzed using the TUNEL
Chromogenic Apoptosis Detection kit (GeneCopoeia, Inc.) following
the manufacturer recommended protocol.
Viral constructions and infection
The circRNA_101237 vector was synthesized by Sangon
Biotech Co., Ltd. circRNA_101237 sequence (1 kb upstream) was
inserted into pcDNA3.1. circRNA_101237 without the downstream
reverse sequence was used as a negative control. All vectors,
including circRNA_101237 and IGF2BP3, were finally cloned into the
Adeno-X™ Tet-On Expression System (Seebio Biotech, (Shanghai) Co.,
Ltd.) according to the manufacturer's protocol. The target sequence
of IGF2BP3 small interfering RNA (siRNA) was 5′-AAA TGA TAT TGC TTC
CAT GAA TC TT-3′. The target sequence of circRNA_101237 siRNA was
5′-TCT GAT AGT AAG TCT TCG-3′. These siRNA adenoviruses were
constructed using the AAVPrime™ AAV System (GeneCopoeia, Inc.)
according to the manufacturer's protocol. All constructs were
amplified in GCI-AAV-293Ta cells (GeneCopoeia, Inc.). Primary
cardiomyocytes were infected with viral at multiplicity of
infection=50 for 48 h.
Let-7a-5p mimic and inhibitor
transfection
The let-7a-5p mimic and inhibitor, and their
negative control were obtained from GeneCopoeia (Guangzhou, China).
Let-7a-5p mimic and inhibitor transfection was performed by using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol.
Lactate dehydrogenase (LDH) activity
measurement
Primary cardiomyocytes were exposed to A/R treatment
for 0, 4, 8 or 12 h, and the medium was collected for LDH activity
measurement using an LDH Quantification kit (cat no. K726-500;
BioVision, Inc.) according to the manufacturer's protocol.
Caspase 3 activity measurement
Following transfection, primary cardiomyocytes were
exposed to A/R for 8 h, and the cells were collected for caspase 3
activity measurement using a Caspase-3 Fluorometric Assay lit (cat
no. K105-100; BioVision, Inc.) according to the manufacturer's
protocol.
Autophagic vacuoles staining
Following transfection with circRNA_101237 or
scramble control siRNA, primary cardio-myocytes were exposed to A/R
for 8 h, and the cells were collected for autophagic vacuoles
staining using a CYTO-ID autophagy detection kit (cat no.
ENZ-51031-0050; Enzo Life Sciences, Inc.) according to the
manufacturer's protocol.
Western blot analysis
Following the aforementioned treatments, the cells
were lysed by RIPA lysis buffer (Cell Signaling Technology, Inc.)
for 30 min on ice. The BCA Protein Concentration Assay kit (Thermo
Fisher Scientific, Inc.) was used to determine the protein
concentration of the sample, according to manufacturer's protocol.
The 50 g protein sample were subjected to 10% SDS-PAGE and
transferred to PVDF membranes. The membranes were blocked by 5%
non-fat milk for 1 h at room temperature. The membranes then were
incubated with primary antibodies [anti-microtubule-associated
proteins 1A/1B light chain 3B (LC3B), 1:1,000, cat. no. ab48394;
anti-IGF2BP3, 1:100, cat. no. ab225697; Beclin1, 1:1,000, cat. no.
ab210498; autophagy protein 5 (Atg5), 1:1,000, cat. no. ab228668;
and anti-GAPDH, 1:2,000, cat. no. ab8245] overnight at 4°C. All
antibodies were purchased from Abcam. The membranes were washed
three times with PBS. Following washing, the membranes were
incubated with appropriate secondary antibodies [goat anti-rabbit
IgG H&L (HRP), 1:3,000, cat. no. ab205718; and goat anti-mouse
IgG H&L (HRP), 1:3,000, cat. no. ab205719; both Abcam] for 1 h
at 37°C. The bands were measured using a chemiluminescence system
(Bio-Rad, USA) imaging system.
Luciferase activity assay
PCR was performed to generate the IGF2BP3
3′-untranslated region (UTR) as previously described (17). A QuikChange Lightning Multi
Site-Directed Mutagenesis kit (Agilent Technologies, Inc.) was used
to generate the mutated 3′-UTR of IGF2BP3. Sequencing was performed
to verify coding sequences. Wild-type and mutated 3′-UTRs of
IGF2BP3 were subcloned into the pGL3 vector immediately downstream
of the luciferase gene (Genomeditech). Primary cardiomyocytes were
transfected with wild-type or mutated 3′-UTRs of IGF2BP3, and
co-transfected with let-7a-5p mimics or mimics negative control
using Lipofectamine® 3000 (Thermo Fisher Scientific,
Inc.). The cells transfected with pGL3 vector were used as control.
At 48 h after transfection, the Dual-Luciferase Reporter Assay
System (Promega Corporation) was used to determine the luciferase
activity according to the manufacturer's protocol. Renilla
luciferase activity was used for normalization.
Biotinylated miRNA pull-down
The assay was performed as previously described
(1). The 5′-biotin labeled
let-7a-5p was synthesized by Sangon Biotech Co., Ltd.
Cardiomyocytes were transfected with biotinylated let-7a-5p for 24
h. Following transfection, cardiomyocytes were lysed in RIPA lysis
buffer (Cell Signaling Technology, Inc.) for 30 min on ice and
centrifuged at 12,000 × g at 4°C for 20 min to collect the
supernatant. A sample of 50 µl was used as an input control.
The supernatant was incubated with streptavidin agarose beads
(Invitrogen; Thermo Fisher Scientific, Inc.) at 4°C for 3 h. The
bound RNAs were purified by washing with low-salt buffer and
high-salt buffer (Wuhan Boster Biological Technology, Ltd.). The
purified RNA was used for RT-qPCR according to the aforementioned
protocol. GAPDH was used as a negative control.
Protein argonaute 2 (AGO2)
immunoprecipitation
Let-7a-5p and let-7a-5p mutants were transfected
into cardiomyocytes for 48 h. The cells were lysed in RIPA lysis
buffer (Cell Signaling Technology, Inc.) for 30 min on ice and
incubated with anti-AGO2 (1:100; cat. no. ab186733; Abcam) or
control IgG antibody-(1:100; cat. no. 3900s; Cell Signaling
Technology, Inc.) coupled Sepharose beads for 4 h at 4°C under
rotation. The bound RNAs were purified by washing with lysis buffer
(Wuhan Boster Biological Technology, Ltd.). The purified RNA was
used for RT-qPCR according to the aforementioned protocol. GAPDH
was used as a negative control.
Immunofluorescence assays
The cardiomyocytes were cultured on slides for 24 h.
Following blocking with non-fat 5% milk for 60 min at 37°C, the
slides were incubated with an anti-LC3B primary antibody (1:200
dilution; cat. no. ab48394; Abcam) at 4°C overnight. The slides
were incubated with appropriate secondary antibodies for 1 h at
37°C. Following washing with PBS, the slides were co-stained with
DAPI (1:2,000; cat. no. SC-3598; Santa Cruz Biotechnology, Inc.)
for 2 min at room temperature. Immunofluorescence images were
captured at ×200 magnification using a fluorescence microscope
(Nikon ECLIPSE 80i; Nikon Corporation).
Bioinformatics analysis
The bioinformatics software RNAhybrid (version 2.1;
https://bibiserv.cebitec.uni-bielefeld.
de/rnahybrid/)was used to identify potential miRNA binding sites
within circRNA_101237, as previously described (18). The bioinformatics tool miRwalk
(version 2; http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
was used to predict the potential targets of let-7a-5p as
previously described (19).
Statistical analysis
All statistical analyses were performed using
GraphPad Prism v.7.04 (GraphPad Software, Inc.). Continuity
variables are expressed as mean ± standard deviation of at least 3
independent experiments. A Student's t-test was used for comparison
between two groups; a one-way analysis of variance was used for
comparisons between three or more groups, followed by a Tukey's
post hoc test for comparison. All statistical analyses were
performed using a two-sided test. P<0.05 was considered to
indicate a statistically significant difference.
Results
circRNA_101237 is induced by A/R in
cardiomyocytes
To investigate the potential role of circRNAs
A/R-induced cardiomyocyte death, a circRNA microarray was performed
to screen the differentially expressed circRNAs in A/R-treated
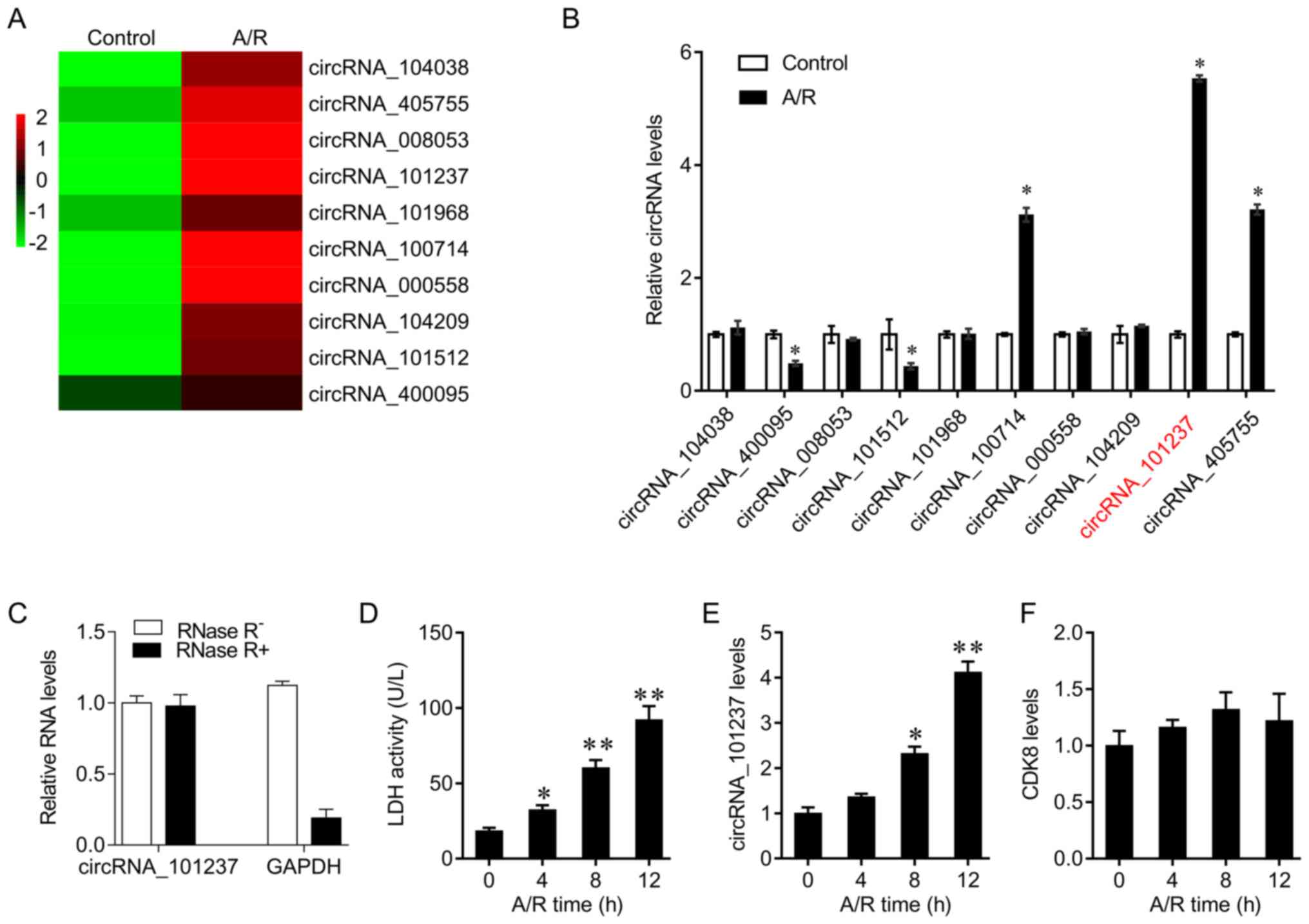
cardiomyocytes. The heatmap demonstrates the 10 circRNAs that were
differentially expressed following A/R treatment (Fig. 1A). Their expression levels were
confirmed by RT-qPCR. The results indicated that circRNA_400095 and
circRNA_101512 levels were decreased, and circRNA_100714,
circRNA_405755 and circRNA_101237 levels were increased upon A/R
treatment. circRNA_101237 levels, generated from the exon 10 to
exon 12 of the CDK8 gene, were significantly increased by A/R
stimulation (Fig. 1B). Due to
resistance to RNase R treatment, circRNAs cannot be degraded by
RNase R digestion. It was identified that GAPDH mRNA (linear RNA
transcripts) was degraded by RNase while circRNA_101237 was not
degraded (Fig. 1C). Primary
cardiomyocytes were then subjected to A/R for different times. LDH
is an enzyme located in the cytoplasm. When the cell membrane is
damaged by stimuli, such as A/R challenges, LDH is released. The
levels of LDH released may reflect the cellular response to A/R
(20). Therefore, LDH activity
was used as a positive control to evaluate whether the A/R was
successfully established. Primary cardiomyocytes were exposed to
A/R for 0, 4, 8, or 12 h, and the medium was collected for LDH
activity measurement using an LDH Quantification kit. LDH activity
was significantly upregulated by A/R treatment (Fig. 1D). In addition, circRNA_101237
levels increased in a time-dependent manner, while the expression
levels of host gene CDK8 were not significantly different among the
groups (Fig. 1E and F). These
data suggest that circRNA_101237 may contribute to oxygen
deprivation-induced cardiomyocyte death.
Knockdown of circRNA_101237 decreases
cardiomyocyte apoptosis
The role of circRNA_101237 in A/R-induced
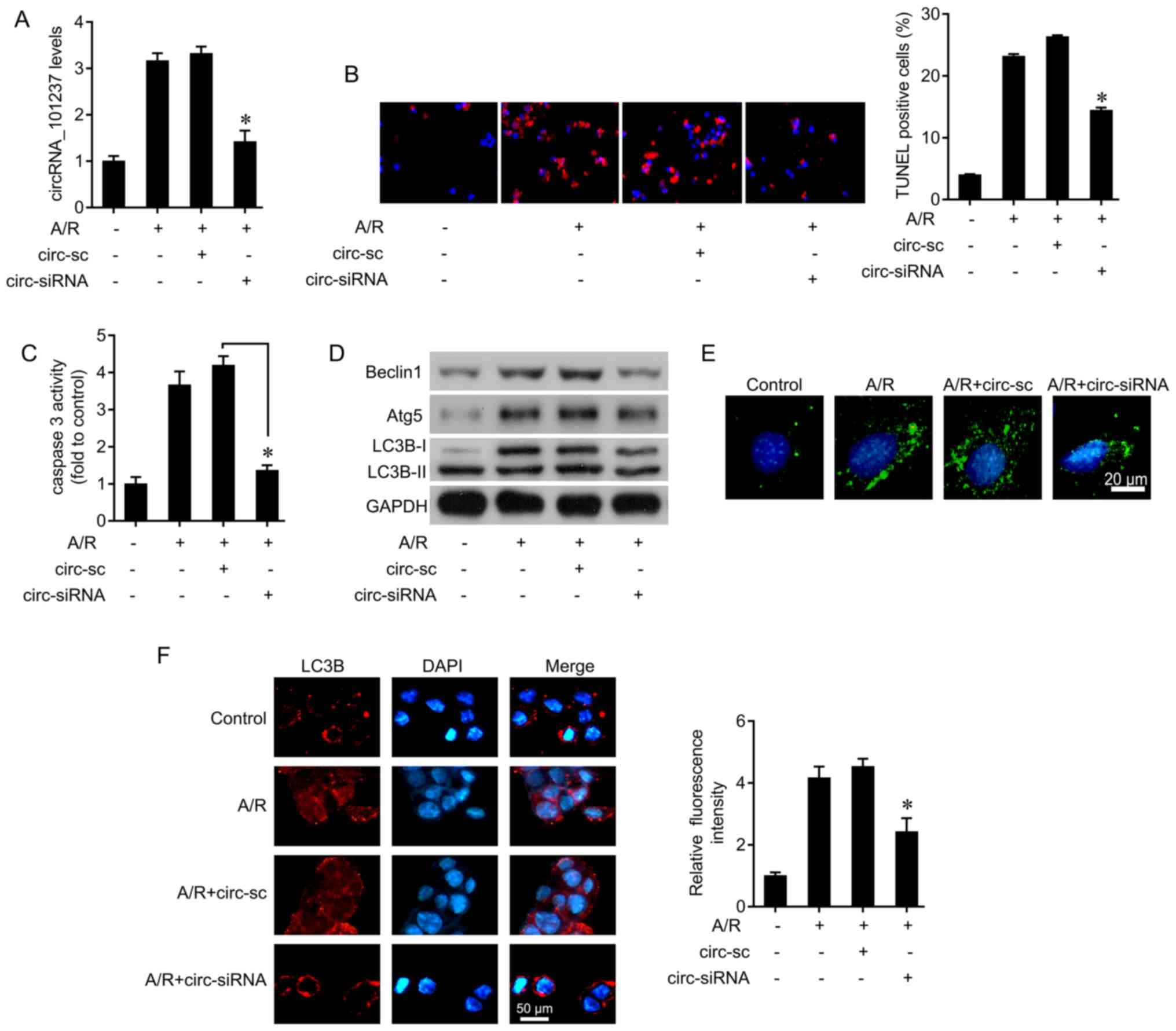
cardiomyocyte apoptosis was next examined by siRNA transfection.
circRNA_101237 knockdown efficiently inhibited the increase in
circRNA_101237 level induced by A/R in cardiomyocytes (Fig. 2A). A TUNEL assay was used to
determine the cell apoptosis induced by A/R. A/R treatment
significantly induced cell apoptosis. However, knockdown of
circRNA_101237 decreased the level of A/R-induced apoptosis,
identified by a decrease in the number of TUNEL-positive cells
(Fig. 2B). In addition, knockdown
of circRNA_101237 inhibited A/R-induced caspase 3 activity
(Fig. 2C).
Autophagic cell death serves important roles in
cardiac diseases, including ischemic heart disease and heart
failure (21). The autophagy
levels in cells were evaluated following A/R treatment and
circRNA_101237 siRNA transfection. It was observed that the
expression levels of Beclin1, Atg5, and LC3B were increased by A/R
treatment. circRNA_101237 siRNA transfection attenuated
A/R-mediated autophagy activation (Fig. 2D). Cyto-ID Green dye was used to
staining autophagic vacuoles, and it was identified that A/R
treatment significantly increased the number of autophagic
vacuoles, which was decreased by circRNA_101237 siRNA transfection
(Fig. 2E). In addition, the
immunofluorescence assay results also confirmed that circRNA_101237
knockdown reversed A/R-mediated LC3B upregulation (Fig. 2F). These data indicate that
circRNA_101237 knockdown inhibits cardio-myocytes apoptosis via the
autophagy pathway.
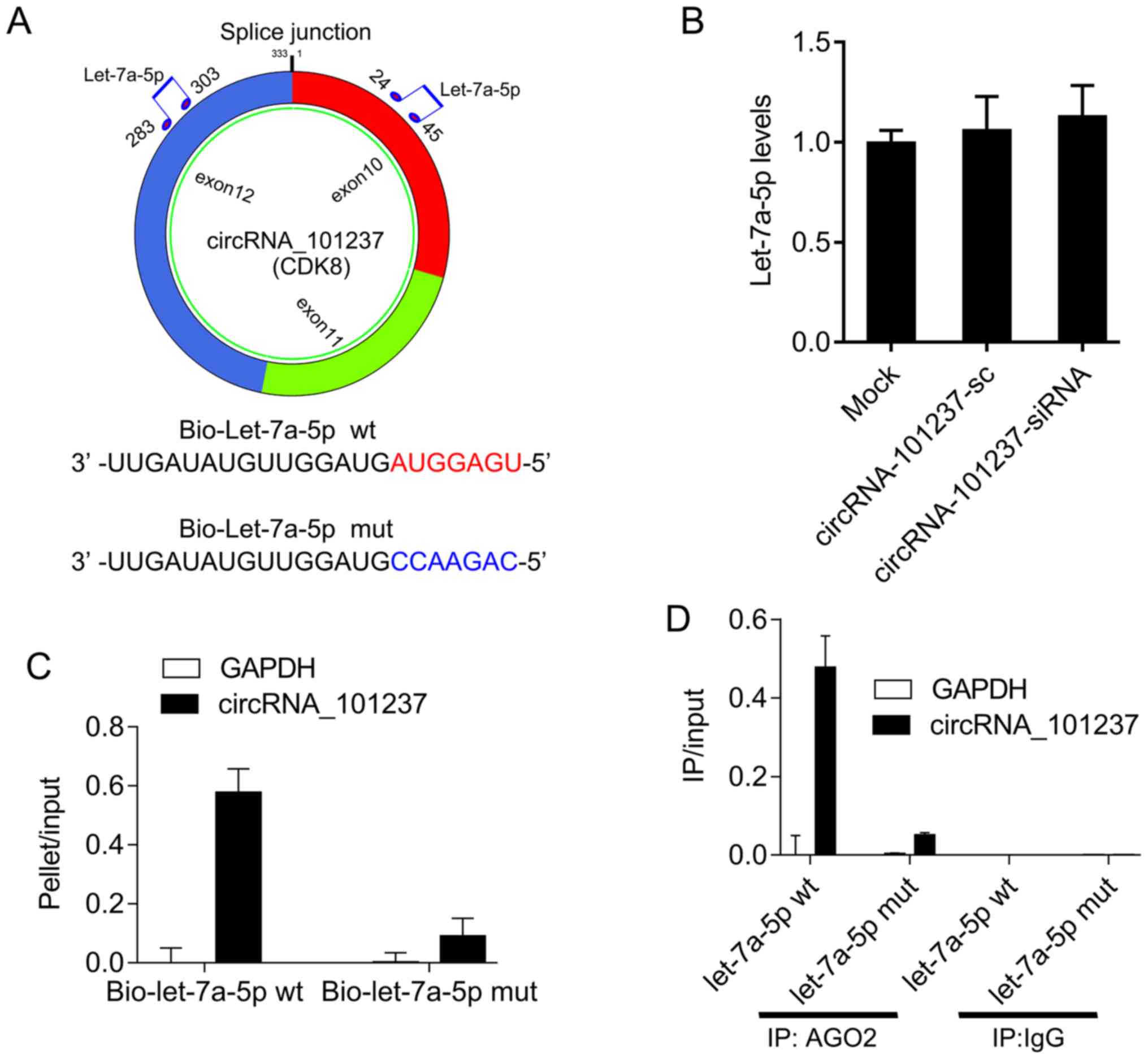
circRNA_101237 serves as a sponge for
let-7a-5p
circRNA_101237 is an exonic circRNA. The
bioinformatics software RNAhybrid was used to identify potential
miRNA binding sites within circRNA_101237. circRNA_101237
interacted with let-7a-5p at the two predicted binding sites
(Fig. 3A). However, it was
identified that circRNA_101237 knockdown had no effect on let-7a-5p
expression (Fig. 3B). The RNA
pull-down assays results demonstrated a higher enrichment of
circRNA_101237 in the bio-let-7a-5p-captured fraction compared with
the negative control (Fig. 3C).
Furthermore, AGO2 immunoprecipitation also suggested that
endogenous circRNA_101237 and let-7a-5p were co-located with AGO2,
but circRNA_101237 was not detectable in samples transfected with
mutant let-7a-5p mutant (Fig.
3D). These results suggested that let-7a-5p facilitates the
AGO2 association with circRNA_101237. Therefore, circRNA_101237
functions as a let-7a-5p sponge without regulating let-7a-5p
expression.
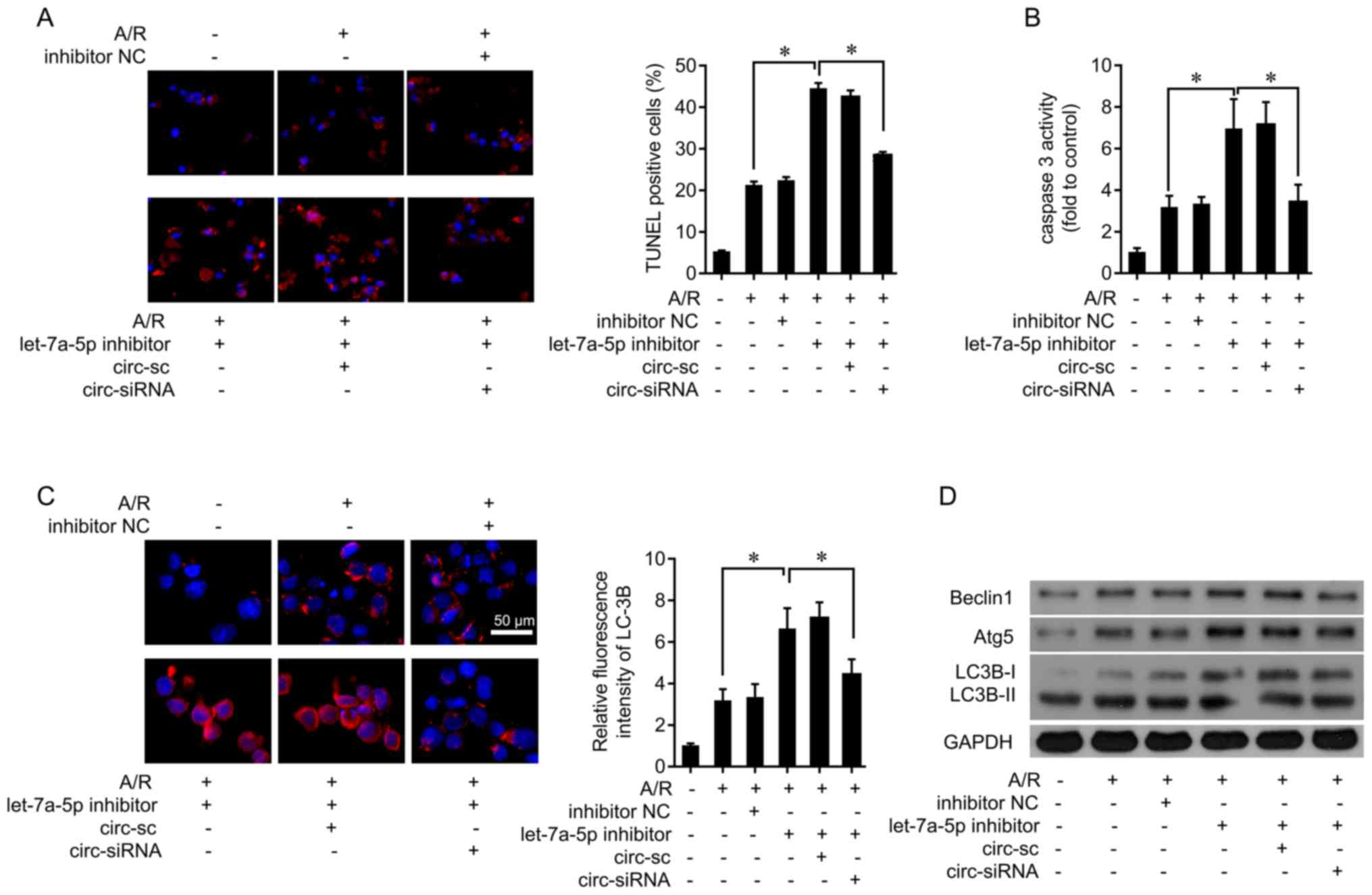
Let-7a-5p inhibitor attenuates the
effects of circRNA_101237 on cell apoptosis and autophagy in
cardiomyocytes
Next, whether endogenous let-7a-5p was implicated in
the cell death induced by A/R was explored. Let-7a-5p inhibitors
were transfected into primary cardiomyocytes (Fig. S1A). Let-7a-5p inhibition enhanced
A/R-induced apoptosis, while co-treatment of circRNA_101237 siRNA
attenuated apoptosis (Fig. 4A).
In addition, the let-7a-5p inhibitor upregulated A/R-induced
caspase 3 activity, which was then inhibited by circRNA_101237
siRNA transfection (Fig. 4B).
Furthermore, the let-7a-5p inhibitor significantly increased
A/R-induced LC3B levels, which was abolished by circRNA_101237
knockdown (Fig. 4C). The
expression levels of key molecules in the autophagy pathway were
also examined, and it was identified that treatment with the
let-7a-5p inhibitor increased Beclin1, Atg5, and LC3B levels
following A/R treatment (Fig.
4D). These data demonstrate that circRNA_101237 serves as a
let-7a-5p sponge to promote cardiomyocyte apop-tosis through
activating autophagy.
Let-7a-5p inhibits cardiomyocytes death
by targeting IGF2BP3
The bioinformatics tool miRwalk (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
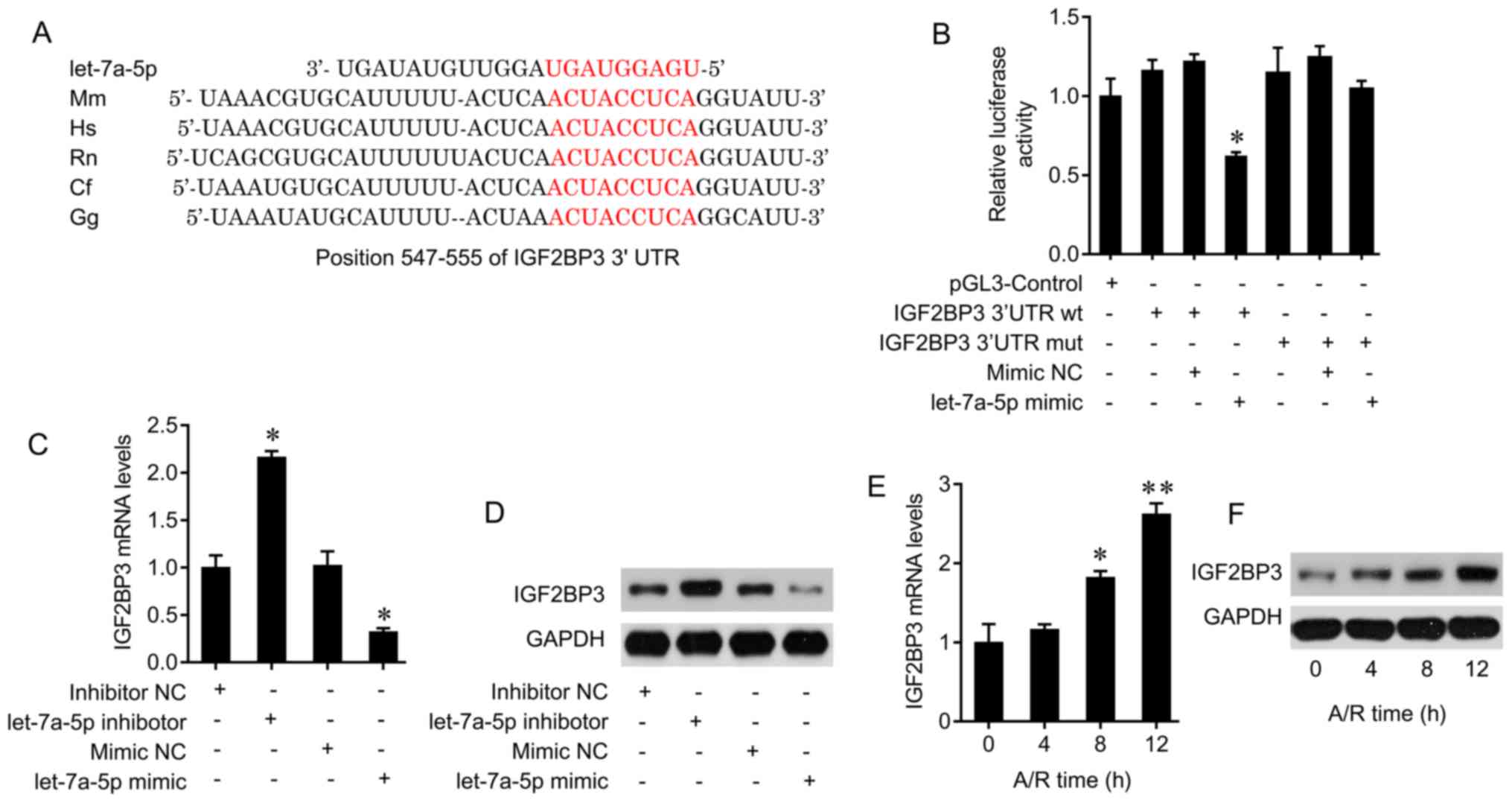
was used to predict the potential targets of let-7a-5p. The 3′ UTR
of the IGF2BP3 gene harbors the conserved binding sites of
let-7a-5p (Fig. 5A). The
luciferase assay system was used to confirm the interaction of
let-7a-5p and IGF2BP3. Let-7a-5p transfection led to a significant
decrease in luciferase activity in primary cardiomyocytes
transfected with the wild-type 3′UTR of IGF2BP3; this effect was
abolished by mutation of the IGF2BP3 gene (Fig. 5B). In addition, let-7a-5p
inhibitor treatment increased the mRNA and protein levels of
IGF2BP3, and let-7a-5p mimic transfection increased let-7a-5p
levels (Fig. S1A) and decreased
IGF2BP3 expression in primary cardiomyocytes (Fig. 5C and D). Therefore, let-7a-5p may
interact with IGF2BP3 and suppress IGF2BP3 expression.
It was also observed that A/R exposure increased the
mRNA and protein levels of IGF2BP3 in cardiomyocytes in a
time-dependent manner (Fig. 5E and
F). Furthermore, the effect of IGF2BP3 on the cell death
induced by A/R treatment was investigated. IGF2BP3 expression was
knocked down by siRNA transfection (Fig. S1B and C). IGF2BP3 silencing
decreased the number of TUNEL-positive cells induced by A/R
challenge, while co-treatment with let-7a-5p inhibitors
significantly increased the number of TUNEL-positive cells
(Fig. 6B). In addition, IGF2BP3
knockdown inhibited caspase 3 activity induced by A/R challenge,
which was increased by co-treatment with let-7a-5p inhibitors
(Fig. 6B).
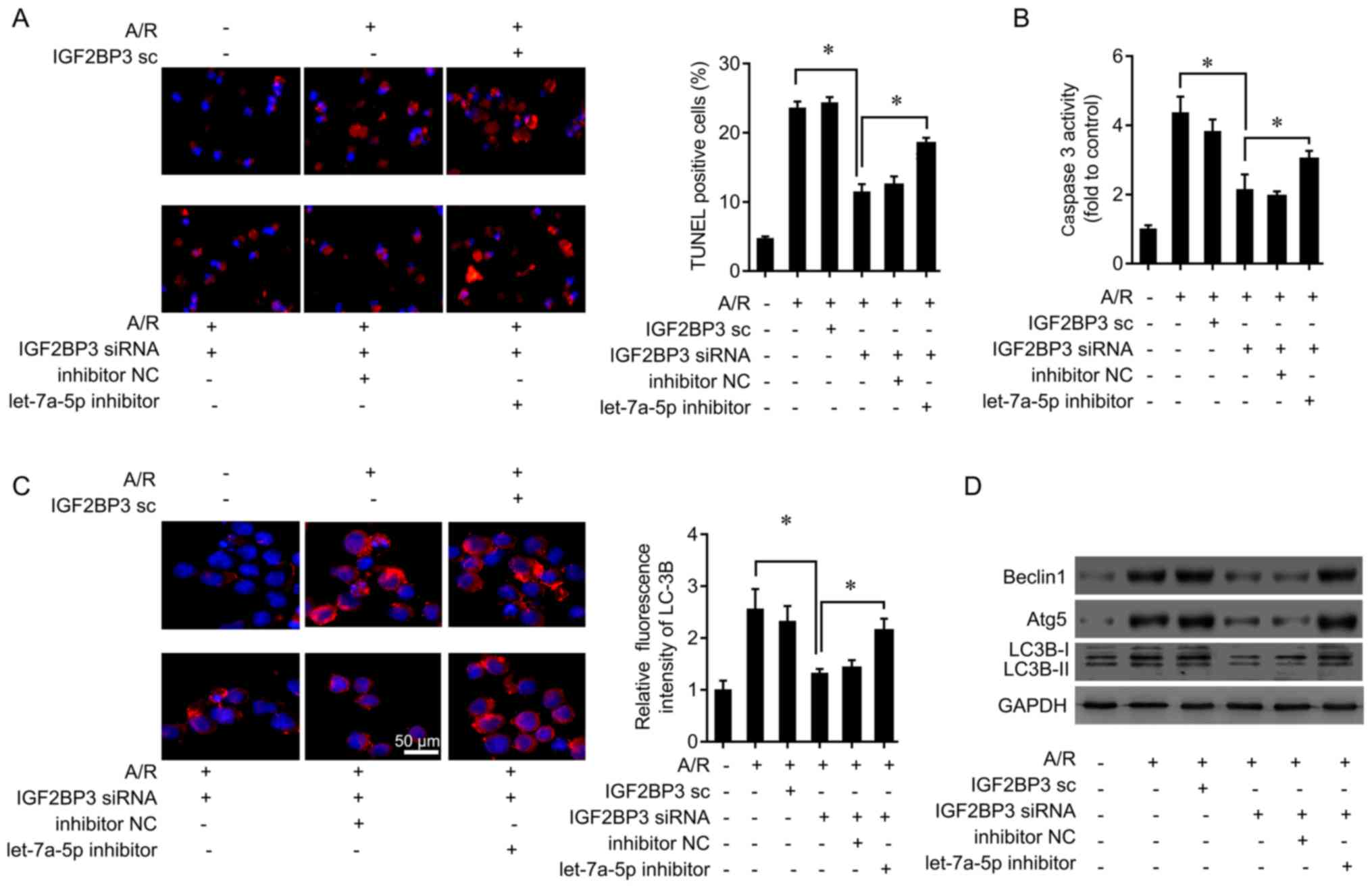 | Figure 6let-7a-5p inhibits cardiomyocytes
apoptosis by targeting IGF2BP3. (A) Cardiomyocytes were transfected
with IGF2BP3 siRNA or control oligo, or let-7a-5p inhibitor, and
then exposed to A/R for 12 h. A TUNEL assay was performed to
measure levels of cell apoptosis. *P<0.05. (B)
Caspase 3 activity was determined following IGF2BP3 knockdown and
let-7a-5p inhibitor treatment in cardiomyocytes. (C) Cardiomyocytes
were transfected with IGF2BP3 siRNA or control oligo, or let-7a-5p
inhibitor, and then exposed to A/R for 12 h. The autophagy marker,
LC3B, was analyzed by immunofluorescence. (D) The autophagy markers
Beclin1, Atg5 and LC3B was analyzed by western blot analysis
following IGF2BP3 knockdown and let-7a-5p inhibitor treatment in
cardiomyocytes. *P<0.05. IGF2BP3, insulin-like growth
factor 2 mRNA-binding protein 3; siRNA, small interfering RNA;
TUNEL, terminal deoxynu-cleotidyl-transferase-mediated dUTP nick
end labelling; A/R, anoxia/reoxygenation; LC3B,
microtubule-associated proteins 1A/1B light chain 3B; Atg5,
autophagy protein 5; NC, negative control. |
The results of the immunofluorescence assay revealed
that knockdown of IGF2BP3 inhibited A/R-mediated LC3B upregulation,
which was abolished by let-7a-5p inhibitor in primary
cardiomyocytes under A/R treatment (Fig. 6C). IGF2BP3 down-regulation
decreased Beclin1, Atg5, and LC3B levels following A/R treatment
(Fig. 6D). The effects of IGF2BP3
downregulation on Beclin1, Atg5, and LC3B levels was attenuated by
let-7a-5p inhibitor treatment (Fig.
6D). Therefore, let-7a-5p inhibited cardiomyocyte death by
targeting IGF2BP3 and inactivating autophagy.
Discussion
In cardiovascular diseases, several miRNAs have been
suggested as potential biomarkers with high diagnostic and
prognostic potential that were comparable to those of established
protein-based biomarkers, such as cardiac troponins and natriuretic
peptides (22,23). Additionally, miRNAs may also be
utilized as therapeutic agents in cardiovascular diseases (24). Recent studies have revealed that
circRNAs may function as endogenous miRNA sponges, ultimately
exerting a significant effect on downstream miRNA targets. For
example, the circRNA MFACR directly interacts with miR-652-3p and
inhibits its expression, subsequently inducing mitochondrial
protein, 18 kDa translation to regulate mitochondrial fission and
apoptosis (25), and circRNA HACR
serves as an endogenous miR-223 sponge to inhibit cardiac
hypertrophy and heart failure by targeting activity-regulated
cytoskeleton-associated protein (26). In the present study, a novel and
crucial circRNA, circRNA_101237, which regulated cardiomyocyte
apoptosis during A/R injury, was identified. It was also
demonstrated that circRNA_101237 targeted the let-7a-5p/IGF2BP3
signaling axis, which provided additional evidence for the sponge
activity of this circRNA (27).
The sponging of let-7a-5p leads to the upregulation
of IGF2BP3 protein and the activation of autophagy. The let-7
family contains 13 members, including let-7a-1 (let-7a-5p), let-7c,
let-7d, let-7e, let-7f, let-7g, let-7i, mir-98 and mir-202
(28). Let-7 has been identified
as a tumor suppressor in a number of types of human cancer
(29). Additionally, the members
of let-7 family have been identified to serve important roles in
cardiovascular biology and disease (30). Let-7 dysfunction has been observed
in various cardiovascular diseases. For example, a previous study
demonstrated the upregulation of let-7b and let-7c in cardiac
hypertrophy, and the downregulation of let-7a in angiotensin
II-induced cardiac hypertrophy (31). However, let-7 inhibition has also
been suggested to prevent deterioration of cardiac function via an
increase in the recruitment of epicardial cells and EMT (32). It is important to note that the
expression of the let-7 members is disease- and tissue-specific
(30). Let-7a is upregulated
during cardiac development and downregulated in human endothelial
cells during angiogenesis (33,34), indicating that the changes in the
expression of let-7 family members are specific to certain
pathological processes, and highlights the requirement for an
improved understanding of the let-7 family function. In the present
study, the results revealed that let-7a-5p expression was unchanged
by circRNA_101237 knockdown. However, it was also demonstrated that
let-7a-5p inhibition lead to the augmentation of A/R-mediated
cardiomyocyte death. Further examination identified IGF2BP3 as a
novel let-7a-5p target. IGF2BP3 is a oncofetal protein that refines
the post-transcriptional modulation of gene expression (35). IGF2BP3 can bind to the 5′ UTR of
IGF2 and regulate IGF2 expression and function (36). IGF2 activates the PI3K/AKT
pathway, which is involved in autophagy and apoptosis (37). Autophagy is an essential cellular
process in the heart and impaired autophagy serves a causative role
in cardiomyopathy. Stimuli such as A/R and I/R injury may lead to
excessive autophagic activity, which can damage a large fraction of
the cytoplasm and organelles, in particular the mitochondria,
subsequently leading to cell death (38). Therefore, maintaining cellular
autophagic homeostasis is important for cardiomyocyte survival.
In the present study, it was identified that IGF2BP3
was involved in A/R-induced cardiomyocyte apoptosis and, to the
best of our knowledge, demonstrated for the first time that IGF2BP3
was a target of let-7a-5p. It was also demonstrated that
circRNA_101237 served as a sponge for let-7a-5p and participated in
the regulation of autophagy and cardiomyocyte death in response to
A/R challenge. Cardiomyocyte death as a result of low oxygen levels
is regulated by complex signaling pathways (39). Although the present study
indicated that circRNA_101237 may regulate cardiomyocyte apoptosis
and autophagy induced by A/R injury, the mechanism underlying the
generation of circRNA_101237 and the molecular mechanisms
underlying the upregulation of circRNA_101237 in cardiomyocytes
remain unknown. As miRNAs can contain several circRNA binding
sites, it remains a possibility that other circRNAs in addition to
circRNA-101237 may also control the expression of let-7a-5p.
In conclusion, to the best of our knowledge, this
was the first study to demonstrate the regulatory mechanism and the
roles of circRNA_101237 in A/R injury. The results identified a
novel circRNA_101237/let-7a-5p/IGF2BP3 axis, which regulated
cardiomyocyte apoptosis via autophagy regulation, as a potential
therapeutic target for the management of cardiovascular
diseases.
Supplementary Data
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JG and HZ designed the study. JY and LS performed
the cell biological experiments. YL, ZL, and YX performed the
reverse transcription-quantitative polymerase chain reaction,
western blot, luciferase reporter assay and immunofluorescence
staining assays. All authors contributed to the writing of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The People's Hospital of Guangxi Zhuang Autonomous
Region.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang K, Long B, Liu F, Wang JX, Liu CY,
Zhao B, Zhou LY, Sun T, Wang M, Yu T, et al: A circular RNA
protects the heart from pathological hypertrophy and heart failure
by targeting miR-223. Eur Heart J. 37:2602–2611. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jakobi T, Czaja-Hasse LF, Reinhardt R and
Dieterich C: Profiling and validation of the circular RNA
repertoire in adult murine hearts. Genomics Proteomics
Bioinformatics. 14:216–223. 2016. View Article : Google Scholar :
|
|
3
|
Xu S, Zhou L, Ponnusamy M, Zhang L, Dong
Y, Zhang Y, Wang Q, Liu J and Wang K: A comprehensive review of
circRNA: From purification and identification to disease marker
potential. PeerJ. 6:e55032018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li JJ, Wang W, Wang XQ, He Y, Wang SS and
Yan YX: A novel strategy of identifying circRNA biomarkers in
cardiovascular disease by meta-analysis. J Cell Physiol.
234:21601–21612. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan X, Weng X, Zhao Y, Chen W, Gan T and
Xu D: Circular RNAs in cardiovascular Disease: An overview. Biomed
Res Int. 2017:51357812017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z
and Sharpless NE: Expression of linear and novel circular forms of
an INK4/ARF-associated non-coding RNA correlates with
atherosclerosis risk. PLoS Genet. 6:e10012332010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du WW, Yang W, Chen Y, Wu ZK, Foster FS,
Yang Z, Li X and Yang BB: Foxo3 circular RNA promotes cardiac
senescence by modulating multiple factors associated with stress
and senescence responses. Eur Heart J. 38:1402–1412. 2017.
|
|
9
|
Zhou Q, Zhang Z, Bei Y, Li G and Wang T:
Circular RNAs as novel biomarkers for cardiovascular diseases. Adv
Exp Med Biol. 1087:159–170. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang L, Meng X, Li G, Zhou Q and Xiao J:
Circular RNAs in cardiovascular diseases. Adv Exp Med Biol.
1087:191–204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tan WL, Lim BT, Anene-Nzelu CG,
Ackers-Johnson M, Dashi A, See K, Tiang Z, Lee DP, Chua WW, Luu TD,
et al: A landscape of circular RNA expression in the human heart.
Cardiovasc Res. 113:298–309. 2017.PubMed/NCBI
|
|
12
|
Altesha MA, Ni T, Khan A, Liu K and Zheng
X: Circular RNA in cardiovascular disease. J Cell Physiol.
234:5588–5600. 2019. View Article : Google Scholar
|
|
13
|
Kristensen LS, Andersen MS, Stagsted L,
Ebbesen KK, Hansen TB and Kjems J: The biogenesis, biology and
characterization of circular RNAs. Nat Rev Genet. 20:675–691. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bayoumi AS, Aonuma T, Teoh JP, Tang YL and
Kim IM: Circular noncoding RNAs as potential therapies and
circulating biomarkers for cardiovascular diseases. Acta Pharmacol
Sin. 39:1100–1109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
von Harsdorf R, Li PF and Dietz R:
Signaling pathways in reactive oxygen species-induced cardiomyocyte
apoptosis. Circulation. 99:2934–2941. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu Y, Liu X, Zhang F, Jiang S, Liu J and
Luo Y: Bortezomib-inducible long non-coding RNA myocardial
infarction associated transcript is an oncogene in multiple myeloma
that suppresses miR-29b. Cell Death Dis. 10:3192019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang K, Li W, Duan W, Jiang Y, Huang N, Li
Y, Ren B and Sun J: Resveratrol attenuates pulmonary embolism
associated cardiac injury by suppressing activation of the
inflammasome via the MALAT1miR223p signaling pathway. Int J Mol
Med. 44:2311–2320. 2019.PubMed/NCBI
|
|
18
|
Kruger J and Rehmsmeier M: RNAhybrid:
microRNA target prediction easy, fast and flexible. Nucleic Acids
Res. 34(Web Server Issue): W451–W454. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sticht C, De La Torre C, Parveen A and
Gretz N: miRWalk: An online resource for prediction of microRNA
binding sites. PLoS One. 13:e2062392018. View Article : Google Scholar
|
|
20
|
Wang S, Ye L and Wang L: Protective
mechanism of shenmai on myocardial ischemia-reperfusion through the
energy metabolism pathway. Am J Transl Res. 11:4046–4062.
2019.PubMed/NCBI
|
|
21
|
Pan JA, Tang Y, Yu JY, Zhang H, Zhang JF,
Wang CQ and Gu J: miR-146a attenuates apoptosis and modulates
autophagy by targeting TAF9b/P53 pathway in doxorubicin-induced
cardio-toxicity. Cell Death Dis. 10:6682019. View Article : Google Scholar
|
|
22
|
Barwari T, Joshi A and Mayr M: MicroRNAs
in cardiovascular disease. J Am Coll Cardiol. 68:2577–2584. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Romaine SP, Tomaszewski M, Condorelli G
and Samani NJ: MicroRNAs in cardiovascular disease: An introduction
for clinicians. Heart. 101:921–928. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schulte C, Karakas M and Zeller T:
microRNAs in cardiovascular disease-clinical application. Clin Chem
Lab Med. 55:687–704. 2017. View Article : Google Scholar
|
|
25
|
Wang K, Gan TY, Li N, Liu CY, Zhou LY, Gao
JN, Chen C, Yan KW, Ponnusamy M, Zhang YH and Li PF: Circular RNA
mediates cardiomyocyte death via miRNA-dependent upregulation of
MTP18 expression. Cell Death Differ. 24:1111–1120. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aufiero S, Reckman YJ, Pinto YM and
Creemers EE: Circular RNAs open a new chapter in cardiovascular
biology. Nat Rev Cardiol. 16:503–515. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
E S, Costa MC, Kurc S, Drożdż A,
Cortez-Dias N and Enguita FJ: The circulating non-coding RNA
landscape for biomarker research: Lessons and prospects from
cardiovascular diseases. Acta Pharmacol Sin. 39:1085–1099. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Roush S and Slack FJ: The let-7 family of
microRNAs. Trends Cell Biol. 18:505–516. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Balzeau J, Menezes MR, Cao S and Hagan JP:
The LIN28/let-7 pathway in cancer. Front Genet. 8:312017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bao MH, Feng X, Zhang YW, Lou XY, Cheng Y
and Zhou HH: Let-7 in cardiovascular diseases, heart development
and cardiovascular differentiation from stem cells. Int J Mol Sci.
14:23086–23102. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou X, Sun F, Luo S, Zhao W, Yang T,
Zhang G, Gao M, Lu R, Shu Y, Mu W, et al: Let-7a is an
antihypertrophic regulator in the heart via targeting calmodulin.
Int J Biol Sci. 13:22–31. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seeger T, Xu QF, Muhly-Reinholz M, Fischer
A, Kremp EM, Zeiher AM and Dimmeler S: Inhibition of let-7 augments
the recruitment of epicardial cells and improves cardiac function
after myocardial infarction. J Mol Cell Cardiol. 94:145–152. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Suarez Y, Fernandez-Hernando C, Pober JS
and Sessa WC: Dicer dependent microRNAs regulate gene expression
and functions in human endothelial cells. Circ Res. 100:1164–1173.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao L, Kong LP, Yu ZB, Han SP, Bai YF, Zhu
J, Hu X, Zhu C, Zhu S and Guo XR: microRNA expression profiling of
the developing mouse heart. Int J Mol Med. 30:1095–1104. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lederer M, Bley N, Schleifer C and
Huttelmaier S: The role of the oncofetal IGF2 mRNA-binding protein
3 (IGF2BP3) in cancer. Semin Cancer Biol. 29:3–12. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bhargava S, Patil V, Shah RA and
Somasundaram K: IGF2 mRNA binding protein 3 (IMP3) mediated
regulation of transcriptome and translatome in glioma cells. Cancer
Biol Ther. 19:42–52. 2018. View Article : Google Scholar :
|
|
37
|
Shimizu T, Sugihara E, Yamaguchi-Iwai S,
Tamaki S, Koyama Y, Kamel W, Ueki A, Ishikawa T, Chiyoda T, Osuka
S, et al: IGF2 preserves osteosarcoma cell survival by creating an
autophagic state of dormancy that protects cells against
chemotherapeutic stress. Cancer Res. 74:6531–6541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nishida K, Yamaguchi O and Otsu K:
Crosstalk between autophagy and apoptosis in heart disease. Circ
Res. 103:343–351. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Takemura G, Kanamori H, Okada H, Miyazaki
N, Watanabe T, Tsujimoto A, Goto K, Maruyama R, Fujiwara T and
Fujiwara H: Anti-apoptosis in nonmyocytes and pro-autophagy in
cardiomyocytes: Two strategies against postinfarction heart failure
through regulation of cell death/degeneration. Heart Fail Rev.
23:759–772. 2018. View Article : Google Scholar : PubMed/NCBI
|