Introduction
Influenza viruses cause worldwide outbreaks and
seasonal pandemics and pose serious risks to human and animal
health. Due to the lack of proofreading mechanisms in the negative
sense RNA genomes during replication, the virus has a high rate of
mutation (1). Thus, considerable
time is required to update vaccines when novel mutated viruses
appear. Thus, it is necessary to continue the development of novel
and effective antiviral drugs against the influenza viruses,
particularly drugs that do not cause resistance.
Current anti-influenza virus drugs include
amantadine, rimantadine, oseltamivir, zanamivir and paramivir,
which target the life cycle of a virus (2,3).
However, worldwide resistance to these drugs due to prolonged usage
and the resulting immuno-compromised status already exists
(4,5). For example, the influenza strains
H3N2 and pdmH1N1 have been reported to be adamantine-resistant
(6) and the H7N9 virus is also
resistant to oseltamivir (7).
Herbal/plant-based medicine, already a global trend in the
pharmaceutical industry, is a potential resource for novel
therapeutic agents (8,9). Traditional Chinese Medicine (TCM)
has been used in the treatment of illnesses for centuries,
including influenza and influenza-like illnesses (10). TCM tends to regulate the host
response and to treat disease by using multi-host targets,
rendering TCM unlikely prone to drug resistance. Following these
insights, there has been an increasing focus on the development of
anti-influenza drugs using traditional medicines as novel prospects
for influenza management (11,12).
Eleutheroside B1, a coumarin compound extracted from
herba sarcandrae, demonstrated a wide spectrum of anti-human
influenza virus efficacy in a previous study (13). However, the mechanisms of action
of eleutheroside B1 remain elusive. More than 85% of the human
genome is transcribed into RNA; however, only <3% of the genome
encodes proteins. Thus, most transcripts of the human genome are
non-coding RNA (ncRNAs) without protein-coding capacity. The
pervasive transcription of the human genome produces thousands of
previously unidentified long intergenic ncRNAs (14). Recent studies have proven that
ncRNAs, including microRNAs (miRNAs or miRs) and long non-coding
RNAs (lncRNAs) play an important role in a number of biological
phenomena and human diseases (15-18). Some studies have demonstrated that
the segment-specific non-coding sequences of influenza A virus and
host non-coding RNA are crucial for influenza A virus replication
(19,20). It has also been recognized that
some ncRNAs are critically involved in the virus-host interaction
as key regulators of transcription or post-transcription during
viral infection. There is increasing evidence to indicate the
functional involvement of these regulatory miRNAs, vault
complex-associated RNAs (vtRNAs) and lncRNAs in influenza virus
replication (21,22). For example, some studies have
demonstrated that ncRNAs can regulate the activation of pattern
recognition receptor (PRR)-associated signaling and transcription
factors, as well as the production of interferons (IFNs) and the
expression of critical IFN-stimulated genes (ISGs) (23,24). In addition to these ncRNAs,
circular RNAs (circRNAs) are a novel type of RNA that, unlike
linear RNAs, form a covalently closed continuous loop, and are
highly represented in the eukaryotic transcriptome (25,26). Some studies have suggested that
circRNAs, derived from both the host and viruses, interact with
these double-stranded RNAs (dsRNAs) by binding antiviral proteins
and interface with the host-virus interaction (27,28). The present study investigated
whether eleutheroside B1 regulates host ncRNAs and cellular
pathways for its anti-influenza virus and anti-inflammation
activities.
High-throughput methods for obtaining biological
data, such as RNA-sequencing and isobaric tags for relative and
absolute quantification (iTRAQ) assay have undergone rapid
technological advances that have led to the identification of novel
molecules for the effective treatment of diseases, and the
investigation of the underlying mechanisms of actions and the
specific targets of interactions, such as DNA, RNA, protein and
enzymes (29,30). Previous studies have investigated
the mRNA expression profiles of A549 cells, following infection by
the influenza virus and treatment with potential drug targets
(31,32). However, these studies were limited
to mRNA, and did not provide an overview of the transcriptome-wide
responses to changes in host cells, nor explored their association
with the differentially expressed ncRNAs and proteins. In the
present study, human lung cancer cells (A549) were treated with
eleutheroside B1 following influenza A virus infection, in order to
assess the pathway profiles of ncRNAs and proteins. RNA-sequencing
and iTRAQ assay were used to comprehensively investigate the
mechanisms responsible for the eleutheroside B1 activity against
influenza A viral infection. Moreover, molecular insights from
target gene enrichment of altered ncRNAs and differentially
expressed proteins provided critical knowledge to enhance the
understanding of the anti-influenza virus mechanisms of
eleutheroside B1.
Materials and methods
Compound, cells and viruses
Eleutheroside B1, extracted from Sarcandra
glabra (also known as herba sarcandrae) was characterized by
proton and carbon nuclear magnetic resonance (1H and
13C NMR) spectroscopy as previously described (13,32). The results of ultra-performance
liquid chromatography in time-of-flight mass spectrometry indicated
>89% purity for eleutheroside B1. Eleutheroside B1 was dissolved
in dimethyl sulfoxide (DMSO) at a concentration of 50 mg/ml and
stored at −20°C.
Human alveolar basal epithelial adenocarcinoma cells
(A549 cells) were purchased from the American Tissue Culture
Collection (ATCC) and cultured in Dulbecco's modified Eagle's
medium (DMEM), supplemented with 10% fetal bovine serum (FBS) under
standard conditions (37°C, 5% CO2). Influenza virus
A/PR/8/34 (H1N1) was also purchased from ATCC and propagated in the
allantoic cavities of chicken eggs (9 days). After 48 h, these
chicken eggs were broken for the collection of chicken embryo
allantoic fluid with the influenza virus.
Cell culture, viral infection and sample
preparation
A549 cells were trypsinized with 0.25% trypsin,
containing 10 mM EDTA (pH 7.4), and seeded in 6-well culture plates
(BD Bioscciences) at up to 80% confluence. Following 24 h, the A549
cells were infected with A/PR/8/34 (H1N1) (MOI=0.1), and incubated
with serum-free medium at 37°C. After removing the inoculums, the
cells were treated with or without eleutheroside B1 (100
µg/ml, TC50=250 µg/ml) (13). The cells were lysed in TRIzol
reagent (Thermo Fisher Scientific, Inc.), following 24 h of
infection, for RNA sequencing and iTRAQ assay. All samples were
stored at -80°C.
RNA sequencing
The total RNA extracts from each sample were
obtained following the manufacturer's instructions (Thermo Fisher
Scientific, Inc.). The quality of the RNA was evaluated by
electrophoresis on a 1% agarose gel. The A260/A280 ratio was
determined using a NanoDrop spectrophotometer (NanoDrop
Technologies, Thermo Fisher Scientific, Inc.) and was accepted
between 1.8 and 2.0. RNA integrity was assessed by Agilent 2100
Tape Station analysis (Agilent Technologies, Inc.) and RIN >7
were found to be acceptable (33). RNA sequencing was performed on an
Illumina X-ten RNA-seq sequence production system (Illumina,
Inc.).
Identification of novel ncRNAs
The raw data were first filtered to eliminate
low-quality reads, based on the reading mapping to the reference
genome, and StringTie was used to assemble clean data. The combined
transcripts were annotated using the GFFCompare program. The
filtering of the RNA coding for the putative protein was performed
using the minimum length and the threshold of the number of exons.
ncRNAs included: i) Small ncRNAs (<200 nt), such as miRNAs,
small interfering RNA (siRNA) and PIWI-interacting RNAs (piRNAs);
ii) ncRNAs, such as lncRNAs (transcript lengths of >200 nt); and
iii) long antisense RNAs (34).
CPC/CNCI/PFAM was used to further screen the coding and non-coding
genes. The coding potential score of <1 was considered as a
novel ncRNA by CPC software (35).
Prediction of the target and functional
analysis
Many ncRNAs, particularly intergenic ncRNAs can
regulate gene transcription via different mechanisms, including
cis-regulatory mechanisms. A previous study demonstrated that
intergenic ncRNAs were more likely to be similar to regulatory
genes, and intergenic ncRNAs located within 5 kb gene-flanking
regions as 'gene-proximate intergenic ncRNAs' (36). These intergenic ncRNAs are
potentially cis-regulatory and often regulate regulatory
genes (36). Therefore, ncRNA
analyses were performed on protein-coding genes that were located
in the 5 kb flanking region of the individual cDNAs. Whilst
predicting the genes encoding the target protein of a
trans-acting ncRNA, the sequence of each ncRNA was obtained,
based on the browser of the human genome, and Gene Ontology (GO;
http://www.geneontology.org/) was used
to functionally classify these neighboring genes in humans
(37). For GO analysis, the R
package was used to separately prepare biological processes,
molecular functions and cellular components. Kyoto Encyclopedia of
Genes and Genomes (KEGG, http://www.genome.jp/kegg/) pathway analysis was
performed to obtain enriched pathways using the predicted target
genes. In order to analyze the regulatory network of differentially
expressed ncRNAs and target mRNAs, the DIANA-LncBase database
(http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex)
was used for prediction. In addition, Circnet (38) was used to predict the association
between differentially expressed ncRNA and circRNAs. The default
parameter in the website was used in this study.
iTRAQ assays
A total of 1×107 cells were mixed with
1,000 µl RIPA buffer (with protease inhibitor cocktail) in
1.5 ml tubes, then sonicated on ice. The cells were treated with or
without eleutheroside B1 (100 µg/ml) 24 h following
infection and were used for both iTRAQ-based proteomic analysis and
RNA sequencing. Following centrifugation for 15 min at 4°C at
12,000 × g the supernatant was transferred to new tubes and kept on
ice. The protein in the supernatant was quantified using the BCA
assay (Pierce BCA Protein Assay kit; Thermo Fisher Scientific,
Inc.) and detected by SDS electrophoresis. Peptides were obtained
following reduction, alkylation, acetone precipitation and trypsin
digestion. Equal amounts of peptides from each sample were
individually labeled with specific TMT reagents (126C,
127C, 128C, 129C,
130C). After cleaning with sodium deoxycholate, the
peptides were desalted using C18 solid phase extraction. A total of
100 µg peptides were fractionated to 120 fractions with high
pH RPRP-HPLC and combined to eight fractions. For each fraction, 2
µg peptide were separated and analyzed with a
Nano-HPLC/EASY-nLC1200 (Thermo Fisher Scientific, Inc.) coupled to
Q-Exactive mass spectrometry (Thermo Finnigan). Separation was
performed using a reversed-phase column (100 µm, ID × 15 cm,
Reprosil-Pur 120 C18-AQ, 1.9 µm, Dr. Math). Mobile phases
contained H2O with 0.1% FA, 2% ACN (phase A) and 80%
ACN, 0.1% FA (phase B). Samples were separated with a 120 min
gradient at 300 nl/min flow rate. Gradient B: 5% for 3 min, 8-35%
for 92 min, 35-45% for 20 min, 45-100% for 2 min, 100% for 2 min,
100-2% for 2 min and 2% for 2 min.
Data acquisition and iTRAQ analysis
Data-dependent acquisition was performed in profile
and positive mode with a Orbitrap analyzer at a resolution of
70,000 (200 m/z) and m/z range of 350-1,600 for MS1. For MS2, the
resolution was set to 17,500 (200 m/z) with a fixed first mass of
120 m/z. The automatic gain control (AGC) target for MS1 was set to
3.0e+06 and 1.0e+05 for MS2. The top 20 most intense ions were
fragmented by HCD with normalized collision energy (NCE) of 32%,
and isolation window of 2 m/z. The dynamic exclusion time window
was 30 sec. Raw MS files were processed with MaxQuant (version
1.5.6.0). The Human protein sequence database
(Uniprot_HUMAN_2016_09) was downloaded from UNIPROT. This database
and its reverse decoy were used by MaxQuant software. The
quantification type was reporter ion MS2 with 6-plex TMT specific
to Lys (K) and unmodified N-term; Filter by PIF (0.75). Trypsin was
set as specific enzyme with up to 2 miss cleavage; oxidation [M]
and acetyl [protein N-term] were considered as variable
modification, Carbamidomethyl [C] was set as fixed modification;
min peptide length was 7 and max peptide mass was 4600. Both
peptide and protein FDR should be <0.01. Only unmodified unique
peptides were used for quantification. The iBAQ label-free
quantification was also measured with log fit checked. All the
other parameters were reserved as default.
All the identified proteins were annotated and
classified by GO and KEGG. The differentially expressed proteins
were then processed by DAVID Functional Annotation Tool 6.8
(http://david.abcc.ncifcrf.gov/) for the
analysis of term enrichment. The results were filtered on the basis
of a Fisher Exact statistical methodology, as previously described.
The GO biological network was evaluated using the ClueGO of
Cytoscape software (http://www.cytoscape.org/, Cytoscape 3.7.2). The
analysis of the protein-protein interaction was performed by STRING
v10.0 (http://www.string-db.org/), and a high
coefficient value of 0.7 was used as a factor reduction. A cluster
analysis was performed to identify the protein expression profiles
differentially expressed (fold change ≥1.2 or ≤0.8 and P<0.05)
using hcluster (https://pypi.python.org/pypi/hcluster/0.2.0).
Integrative analysis of proteome and
transcriptome data
Transcriptome data obtained from a previous study
(39) was used for integrative
analysis with proteome data. In order to identify proteins that
have been consistently expressed at RNA and protein levels, the
differentially expressed proteins were compared with differentially
expressed ncRNAs. Through previous studies and the starBase
database (40), potential target
genes of the differently expressed ncRNAs were predicted. The
predicted potential target genes of the differently expressed
ncRNAs were also compared with the differentially expressed
proteins. The interaction network of the corresponding proteins and
ncRNAs related genes was constructed by the STRING v10.0 software
and Cytoscape software.
Reverse transcription-quantitative PCR
(RT-qPCR)
The A549 cells were seeded in a 6-well plate at 37°C
with 5% CO2, and then infected with influenza virus
(PR8, 0.1 MOI). Following incubation for 2 h, the cells were
treated with eleutheroside B1 (100 µg/ml). At 24 h
post-infection, the cells were collected for the mRNA expression
testing of selected genes [nuclear paraspeckle assembly transcript
1 (NEAT1) and L antigen family member 3 (LAGE3)] by RT-qPCR. The
primer sequences of NEAT1 and LAGE3 are as presented in Table I. RNA was extracted with RnaExTM
Total RNA Isolation Solution (GENEray, Inc.). The production of
cDNA was then achieved using the Rayscript cDNA Synthesis kit
(GENEray, Inc.) with 60 min at 37°C, and 5 min at 85°C.
Subsequently, cDNA was used for qPCR using SYBR-Green Power qPCR
PreMix (GENEray, Inc.). Primers of NEAT1 and LAGE3 were designed
with Entrez Gene: 283131 and Entrez Gene: 8270. The thermocycling
conditions were 1 cycle conditions including 10 min of initial
denaturation at 95°C and 40 cycles of 10 sec denaturation at 95°C,
34 sec annealing at 60°C, 15 sec denaturation at 95°C, and 1
solubility curve cycle of 60 sec of annealing at 60°C, 30 sec
annealing at 95°C, 15 sec annealing at 60°C. The method of
quantification used was that of Livak and Schmittgen
(2−ΔΔCq) (39).
 | Table IThe primers of NEAT1, LAGE3 and GAPDH
mRNA. |
Table I
The primers of NEAT1, LAGE3 and GAPDH
mRNA.
| Gene | Primers | Sequence
(5′→3′) |
|---|
| NEAT1 | Forward |
GTTCCGTGCTTCCTCTTCTG |
| Reverse |
GTGTCCTCCGACTTTACCAG |
| LAGE3 | Forward |
AAACCGCAGCCTCTCAAC |
| Reverse |
TCTCTGTGGCTCCTTCCC |
| GAPDH | Forward |
GCTGAGTATGTTGTGGAGTC |
| Reverse |
GCAGAAGGAGCAGAGATGA |
Western blot analysis
Eleutheroside B1 (100 µg/ml) was added to the
A549 cells following 2 h of incubation with influenza virus (PR8)
at 37°C. The samples, including A549 cells, A549 cells infected
with PR8, PR8-infected A549 cells plus eleutheroside B1 (100
µg/ml) and A549 cells treated with eleutheroside B1 (100
µg/ml) were collected for protein extraction. Whole cell
extracts were obtained by using cell lysis buffer (50 mM Tris-HCl
pH 8.0, 150 mM NaCl, 1% NP-40, protease inhibitors, 1 mM PMSF). As
determined by BCA assay, 1 µg proteins in the cell extract
was loaded per lane and run on 10% SDS-PAGE gel, separated,
transferred to PVDF membranes through a Trans-Blot Semi-Dry
transfer machine, then blocked with PBST with milk (5% w/v milk,
0.05% v/v Tween in PBS), followed by incubation in PBST containing
milk and incubation with a 1:1,000 dilution of the primary antibody
to LAGE3 (PA5-46520, Thermo Fisher Scientific, Inc.) overnight at
4°C and a 1:2,000 dilution goat anti-rabbit IgG cross-adsorbed
secondary antibody (G-21234, Thermo Fisher Scientific, Inc.) for 1
h at room temperature. The membrane was covered with ECL detection
reagent and detected by the Bio-Rad ChemiDoc™ MP system. The gray
value of the result was analyzed using ImageJ software (https://imagej.en.softonic.com/mac) and GraphPad
Prism 7 (GraphPad Software Inc).
Statistical analysis
Differences between 2 groups were analyzed using a
Student's t-test. The differences between multiple groups were
analyzed using one-way ANOVA with Fishers' Least Significant
Difference test. A value of P<0.05 was considered to indicate a
statistically significant difference. Genes in 2 groups, whose
|logFC|>2 and q value were <0.05, were defined as
differential expression genes in the present study. For RNA-seq and
iTRAQ, 3 repeated samples in each group were combined for testing.
For RT-qPCR and western blot analysis, 3 independent experiments
were performed.
Results
Sequencing and alignment
To elucidate the molecular mechanisms of action of
eleutheroside B1 against influenza A virus infection, RNA-seq of
influenza A virus-infected human lung epithelial (A549) cells were
performed in the presence or absence of eleutheroside B1 treatment
using an Illumina X-ten RNA-seq sequence production system. There
were 3 experimental conditions: A549 cells without infection
(A549), A549 cells infected with A/PR8/34/(H1N1) (PR8) and A549
cells infected with A/PR8/34/(H1N1) and treated with Eleutheroside
B1 (100 µg/ml) (PR8+eleu). More than 13 million raw reads
for each sample were generated from constructed RNA-seq libraries.
After filtering, 34.75 M (97.11%, A549), 33.97 M (95.93%, PR8),
34.79 M (95.45%, PR8+eleu) sequencing reads were unambiguously
mapped against the human reference genome. Furthermore, CPAT v.1.2.
2 was used to predict whether these transcripts are coding or
non-coding. The results demonstrated that most of these were
protein-coding RNAs, and some were novel transcripts which were not
found in the database (Fig. 1).
For non-coding RNAs, predicted RNAs indicate mRNAs which are near
to ncRNAs and exhibit similar functions. The expression value
(genes) for each sample is summarized in Table II.
| Table IISummary of expression value for
samples. |
Table II
Summary of expression value for
samples.
| #SN | >0 | >1 | >2 | >3 |
|---|
| A549 | 37,994 | 35,031 | 30,627 | 24,075 |
| PR8 | 38,477 | 35,335 | 30,157 | 23,529 |
| PR8+eleu | 38,217 | 34,855 | 29,385 | 23,100 |
Non-coding RNA expression profiles in
A549 cells infected with influenza A virus
In order to determine specific ncRNA expression
induced by the influenza A virus, the differentially expressed
non-coding RNAs between the uninfected A549 cells and the
virus-infected cells were analyzed. Compared with the control cell
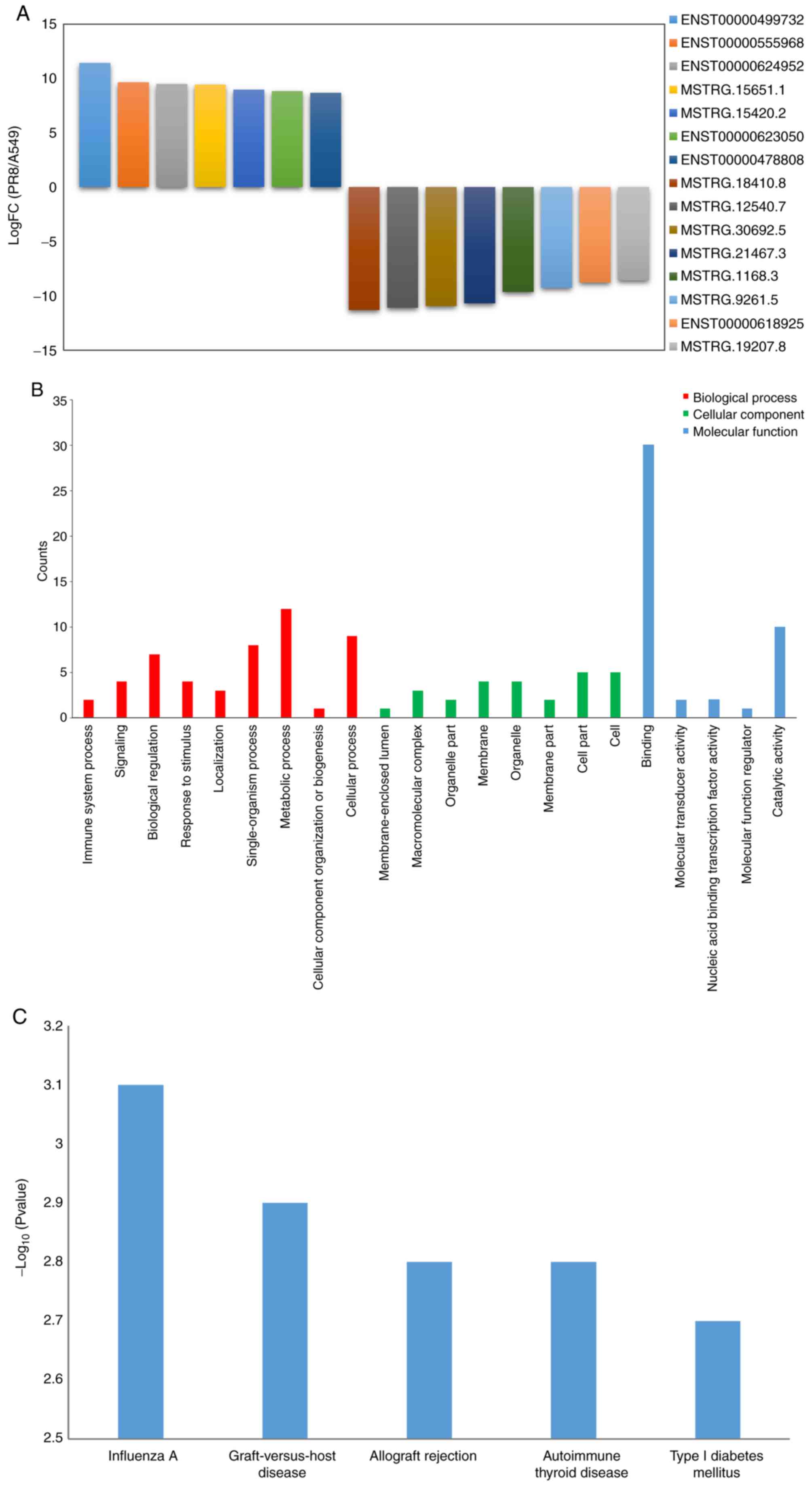
group, 7 ncRNAs that were upregulated and 8 ncRNAs that were
downregulated were identified in the A549 cells following influenza
virus infection (logFC=log2|PR8/A549|); q≤0.05). The differentially
expressed ncRNAs included NEAT1, RP11-66N24.3, R P11- 609D21.3, MST
RG.15651, MST RG.15420, RP11-717F1.2, USP30-AS1, MSTRG.18410,
MSTRG.12540, MST RG. 3 0 69 2, MST RG. 214 67, MST RG.116 8,
MSTRG.9261, MALAT1 and MSTRG19207 (alternative transcript names:
ENST00000499732, ENST00000555968, ENST00000624952, MSTRG.15651.1,
MSTRG.15420.2, ENST00000623050, ENST00000478808, STRG.18410.8,
MSTRG.12540.7, MSTRG.30692.5, MSTRG.21467.3, MSTRG.1168.3,
MSTRG.9261.5, ENST00000618925 and MSTRG.19207.8) (Fig. 2A). Of these ncRNAs, NEAT1 is a
lncRNA that is upregulated in both influenza A virus and HSV-1
infections, causing larger paraspeckles. NEAT1 also modulates HIV-1
post-transcriptional expression. To predict the function of
differentially expressed ncRNAs, the genes closely associated with
ncRNAs were analyzed by the GO and KEGG pathways. In the present
study, 4,091 of these intergenic ncRNAs were identified, and 14,969
genes were predicted to interact with these ncRNAs. Following
statistical analysis (logFC=log2 (PR8/A549), q≤0.05), 7 upregulated
ncRNAs and 8 downregulated ncRNAs were selected for GO
classification and KEGG pathway analysis. According to the GO
classification, these target genes were enriched (P<0.05) in the
biological process (localization, metabolic process, cellular
process and single-organism process); cellular component (cell
part, membrane part and macromolecular matrix); and molecular
function (binding and catalytic activity) (Fig. 2B). The KEGG pathway analysis
revealed that these genes were involved in influenza A,
graft-versus-host disease, allograft rejection, autoimmune thyroid
disease, type I diabetes mellitus and RIG-I-like Receptor (Fig. 2C).
Expression profile of ncRNAs in pulmonary
epithelial cells infected with influenza A virus (A549) and treated
with eleutheroside B1
In order to identify non-coding RNA expression in
influenza virus-infected cells treated with eleutheroside B1, the
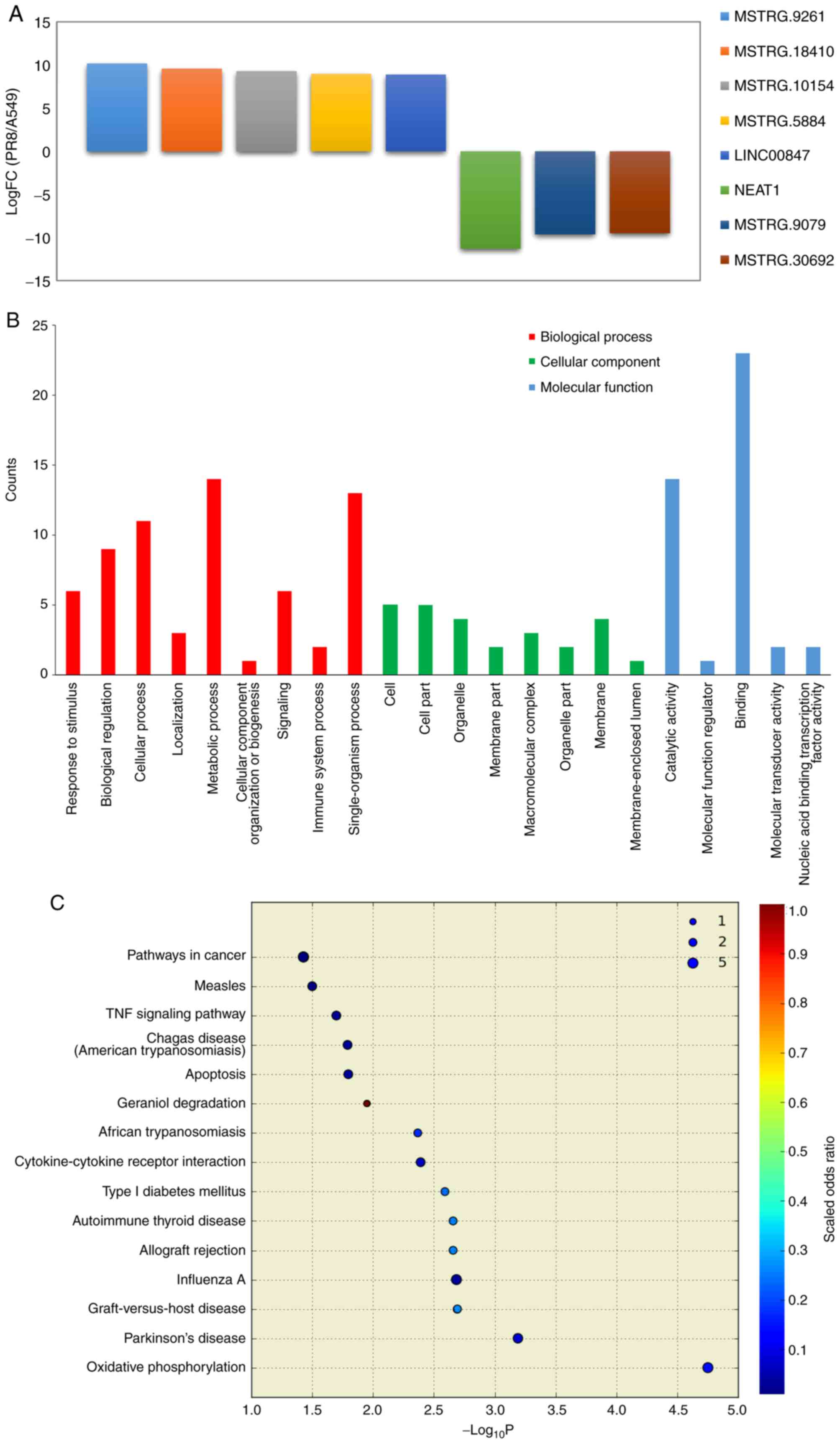
PR8+eleu and PR8 groups were compared and 5 upregulated and 3
downregulated ncRNAs were identified (Fig. 3A), namely NEAT1, MSTRG.9261,
MSTRG.9079, MSTRG.18410, MSTRG.30692, MSTRG.10154, MSTRG.5884,
LINC00847 (alternative transcript names: ENST00000499732,
MSTRG.9261.5, MSTRG.9079.2, MSTRG.18410.8, MSTRG.30692.23,
MSTRG.10154.6, MSTRG.5884.8, ENST00000502162). NEAT1 was
downregulated in the influenza virus-infected cells treated with
eleutheroside B1, although it was upregulated in the influenza
virus-infected cells without drug treatment. In order to determine
differences in ncRNA expression between the 2 groups, genes
associated with ncRNAs were also analyzed through GO and KEGG
analysis to predict their function. The GO enrichment analysis
demonstrated that the majority of these ncRNAs were enriched
(P<0.05) for biological process (single-organism process,
biological regulation, metabolic process, cellular process);
cellular component (macromolecular complex, organelle, membrane,
cell part, cell); and molecular function (catalytic activity,
binding) with some variations from those predicted by ncRNA
(Fig. 3B). According to the KEGG
pathway analysis, the genes associated with the differentially
expressed ncRNAs were involved in oxidative phosphorylation,
Parkinson's disease, graft-versus-host disease, influenza A,
allograft rejection, autoimmune thyroid disease, type I diabetes
mellitus and cytokine-cytokine receptor interaction (Fig. 3C). GO and KEGG analysis of the
differentially expressed ncRNAs between the PR8 and A549 cells
suggested similar pathways, such as catalytic activity, binding (GO
enrichment) and influenza A (KEGG pathway analysis), which were
also enriched by differentially expressed ncRNAs in the
PR8+eleu/PR8 cells.
Regulatory network of ncRNAs and
mRNAs
To date, previous results have demonstrated that
lncRNAs have at least 6 regulatory functions, such as directly
regulating the structure of DNA, transcription and translation of
RNAs (41). lncRNAs also inhibit
the target gene regulation of miRNAs to indirectly regulate gene
expression. Conversely, ncRNAs may target different parts of an
mRNA for its function. In this study, to examine the molecular
mechanisms of ncRNA involvement in influenza virus infection, cells
were treated with eleutheroside B1. The regulatory network analyses
of differentially expressed ncRNAs and mRNAs were performed through
predication on the DIANA-LncBase database (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex).
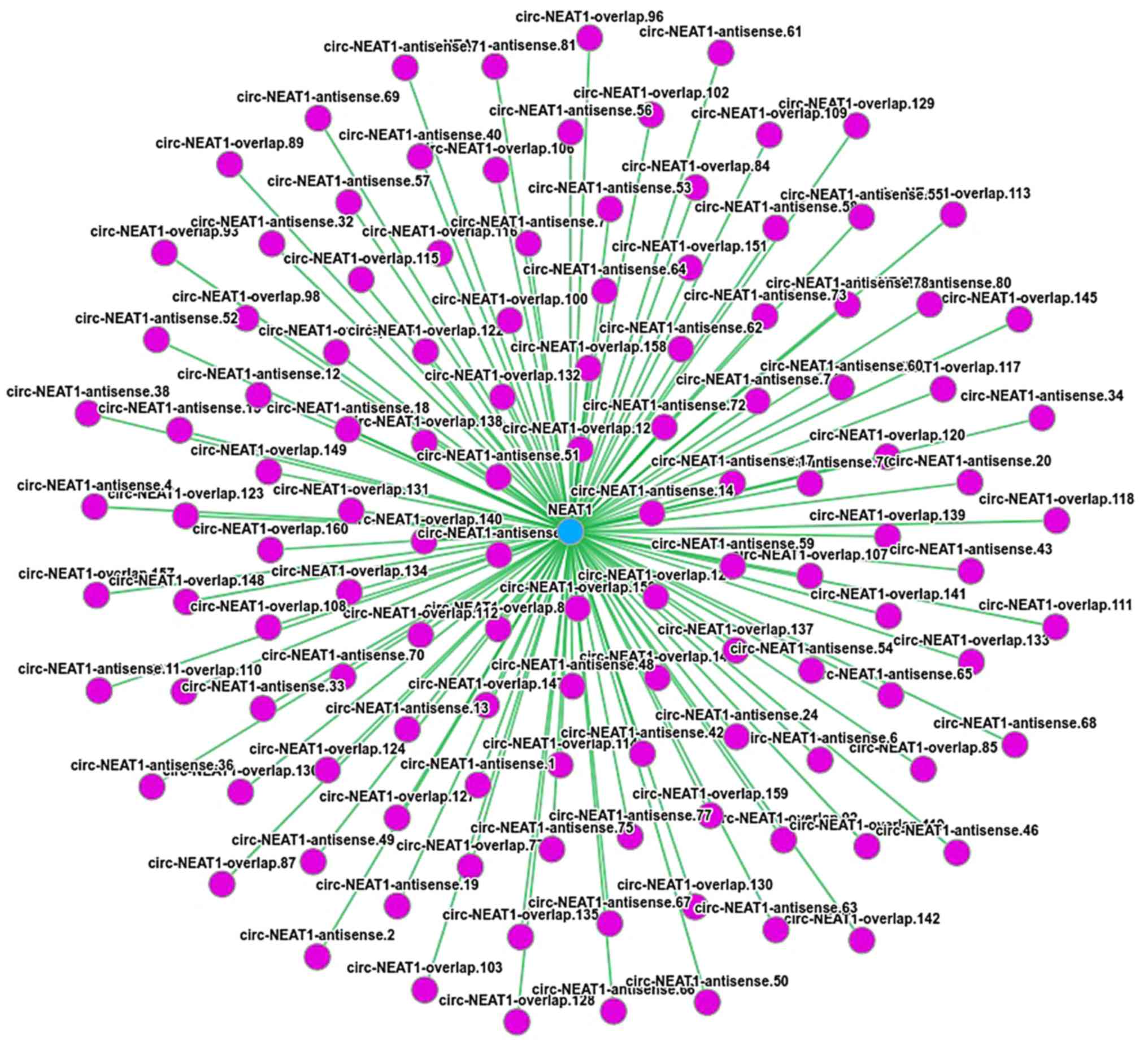
In the predication analysis, NEAT1 targeted 144 mRNAs through
miRNAs (Table III). According
to the predication results in Circnet (38), NEAT1 exhibited an association with
120 circRNAs (Fig. 4). These
results illustrated the regulatory association between ncRNAs and
mRNAs in the mechanisms of eleutheroside B1 in inhibiting the
influenza virus.
| Table IIIPredicted miRNAs and mRNA interacting
with NEAT1. |
Table III
Predicted miRNAs and mRNA interacting
with NEAT1.
| lncRNA | miRNA | mRNA |
|---|
| NEAT1 | hsa-miR-23b-3p | ATG12, PRAP1,
HIF1A, MET, BECN1, ATG12, HMGA2, TSC1, NOTCH1, RGS5, PLAU, SRC,
MAP3K14, TSC1, NKX3-2, CCNG1, MAP3K14, CA2VEGFA, HIP1R, TRAF5,
ATG12, TRAF5, CDH1, IL6R, PPARGC1A, MAP3K1, VEGFA, ETS1, HAS2, VHL,
ZEB1, TAB2, NOTCH2, TAB3, TGIF1, KLF3, MYC, ZNF71, HMGB2, PRDX3,
TAB3, MAP3K1, VEGFA, CCND1, FOXA1, ST7LFZD5, HOXB4, LPAR1 |
| NEAT1 | hsa-miR-7-5p | SNCA, RGS5, FANCG,
IRS2, SMARCD1, BCL2, RAF1, TET2, EGFR, PIK3CD, REL, HOXB5, BCL2,
IGF1R, PSME3, PIK3CD, FOS, GDF5, UBE2A, HOXB3, RNF183, SKP2, XIAP,
XRCC2, PIK3CG, FOS, MSH3, KLF4, TET2, PAK1, BAX, CUL5, RNF183,
PAX6, KMT5A, RNF183, VDAC1, IRS2,HELLS, RAF1, HOXB5, HOXB3, SRSF1,
RELA, RAF1, EGFR, XIAP, TET2, PTK2, IGF1R, MSH3, HELLS, SERPINB5,
IRS2, RGS5, RELA, RNF183 |
| NEAT1 | hsa-miR-30a-3p | THBS1, TMEM2,
SLC7A6, BECN1, RUNX2 |
| NEAT1 |
hsa-miR-146a-5p | RAD54L, TRAF6,
BCLAF1, CLIP1, NUMB, IER5L, SOX2, CXCL12, LFNG, ERBB4, LFNG, HDAC7,
ZDHHC13, CD86, WASF2, FANCM, NFKB1, TLR2, TGFB1, CCND1 |
| NEAT1 | hsa-miR-224-3p | RB1CC1 |
| NEAT1 | hsa-miR-3928-3p
D | ICER1 |
| NEAT1 | hsa-miR-153-5p | RICTOR, TGFB2 |
| NEAT1 | hsa-miR-214-5p | RASSF5, CDK3,
IGF1R, CDK3 |
| NEAT1 |
hsa-miR-216a-5p | CEMIP, SMAD7, CD44,
CDC42 |
| NEAT1 | hsa-miR-328-3p | PLCE1, CD44, H2AFX,
CD44, PTPRJ |
| NEAT1 | hsa-miR-23a-3p | STAT3, FOXA1,
HMGB2 |
| NEAT1 | hsa-miR-339-5p | BCL6 |
| NEAT1 |
hsa-miR-148a-5p | CENPF, CDKN1B |
Proteomic expression in A549 cells
infected with the influenza virus
The proteomic expression levels in the A549 cells at
24 h following influenza virus infection were analyzed in order to
identify the specific proteomic expression of cells treated with
eleutheroside B1. The analysis of the LC-MS/MS data generated a
total of 70,249 peptides. A total of 5,809 proteins were identified
with at least one unique peptide with a confidence level of
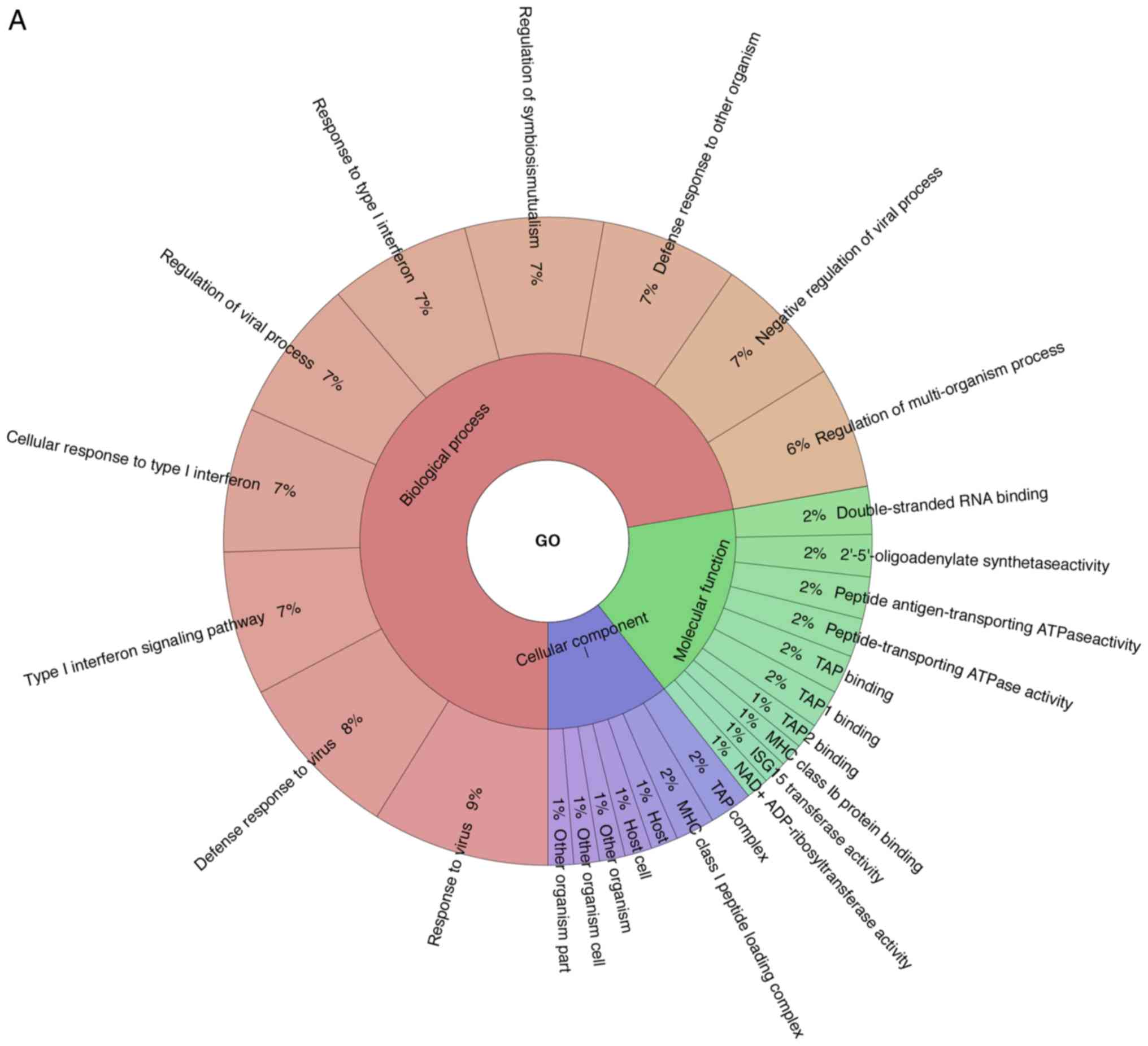
>95%. Through GO analysis, 71 differentially expressed proteins
were enriched for the biological process, cellular component and
the molecular function (Fig. 5A).
The GO results demonstrated that the majority of the proteins
expressed were enriched in these categories of biological
processes, such as virus response, type I interferon signaling
pathway, cellular response to type I interferon, regulation of
viral process and response to type I interferon. The TAP complex
and MHC class I peptide loading complex were the two most abundant
categories in cellular component. For molecular function,
double-stranded RNA binding, 2′-5′-oligoadenylate synthetase
activity, peptide antigen-transporting ATPase activity,
peptide-transporting ATPase activity were primary categories for
proteins. As indicated by KEGG analysis, these proteins were
primarily involved in Herpes simplex virus infection, influenza A
virus, measles, hepatitis C virus, NOD-like receptor signaling
pathway, antigen processing and presentation, RIG-I-like receptor
signaling pathway, phagosome, chemokine signaling pathway,
Staphylococcus aureus infection and osteoclast
differentiation (Fig. 5B).
Differentially expressed proteins in
influenza virus infection and eleutheroside B1 treatment
Proteins seldom function alone, but rather interact
with other proteins to perform various functions. In this study, to
explore protein interaction patterns in influenza virus infection
and eleutheroside B1 treatment, differently expressed proteins
identified were analyzed using STRING software and were enriched
through GO and KEGG pathway. A total of 90 proteins were detected
from all the test groups. Differentially expressed proteins (fold
change of ≥1.5 or ≤0.666 and P<0.05) were identified between the
influenza virus-infected cells treated with eleutheroside B1 and
the untreated cells. Only one protein (LAGE3) was downregulated in
the PR8+eleu vs. PR8 group, and 70 upregulated and 5 downregulated
proteins in the PR8+eleu vs. cells group, in which no significance
difference was found for the expression of LAGE3 (data not
shown).
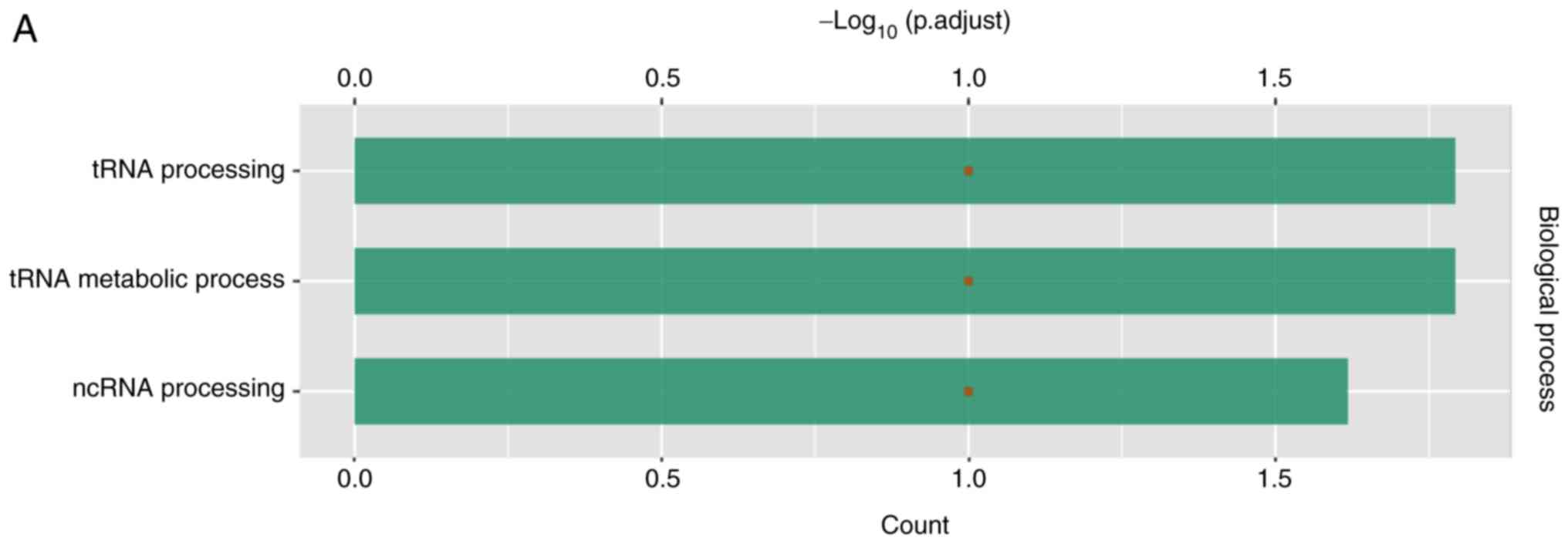
According to GO analysis, LAGE3 was involved in tRNA
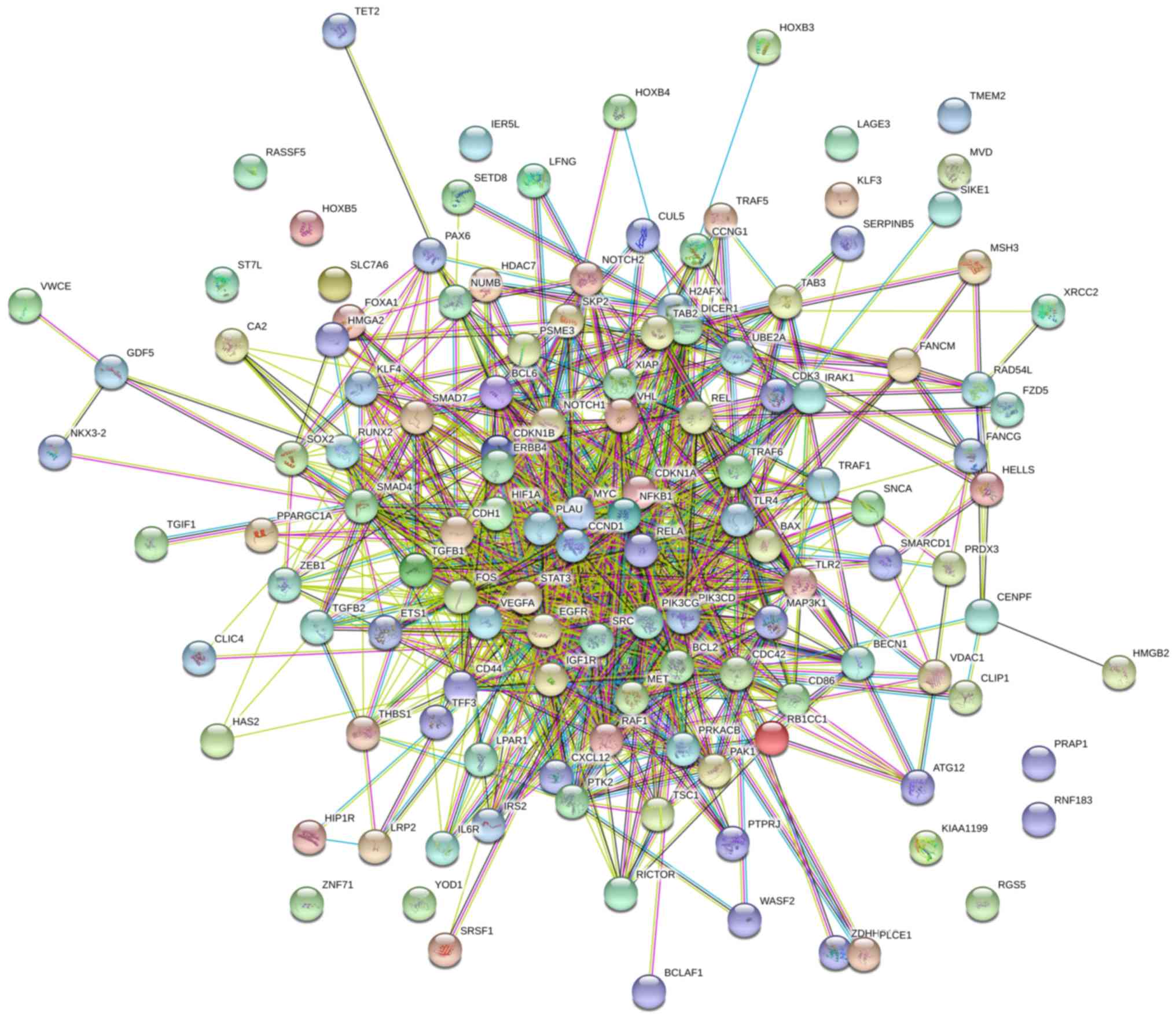
processing, tRNA metabolic process and lncRNA processing (Fig. 6A). Through analysis with STRING,
LAGE3 was shown to interact with a series of other proteins
(Fig. 6B).
Interaction network of integrative
proteome and transcriptome analysis
Integrative analysis of the proteome and
transcriptomes were performed to determine the mechanisms through
which proteins are regulated by eleutheroside B1 at the
translational and transcriptional level. LncRNAs can regulate gene
expression by acting on miRNAs. Based on prediction, the
differentially expressed ncRNA NEAT1 in the PR8+eleu vs. PR8 groups
exhibited an interaction with 144 proteins through miRNAs. The
differential expression of LAGE3 in the 2 groups was regulated in
the same manner as its mRNAs in the RNA sequences (data not shown).
The potential ncRNA targets were used with proteomics data to
construct an interaction network, which indicated that the
potential ncRNA targets, such as ATG12, EGFR, CD44, CXCL12 and
CDC42 were interacting, whereas there was no direct interaction
between these targets and LAGE3 in the database (Fig. 7). The interaction between these
proteins was also intricated in the GO terms and KEGG pathways,
such as the positive regulation of cellular metabolic process
(GO:0031325), regulation of cell proliferation (GO:0042127),
protein binding (GO:0005515), enzyme binding (GO:0019899), nuclear
part (GO:0044428), microRNAs in cancer (05206), FoxO signaling
pathway (04068), indicating that eleutheroside B1 can affect cell
function by regulating both ncRNA and protein. Combining the
RNA-seq and iTRAQ results, it was demonstrated that eleutheroside
B1 regulated some pathways, which were associated with cell
proliferation and gene expression at different levels to inhibit
the replication of influenza virus.
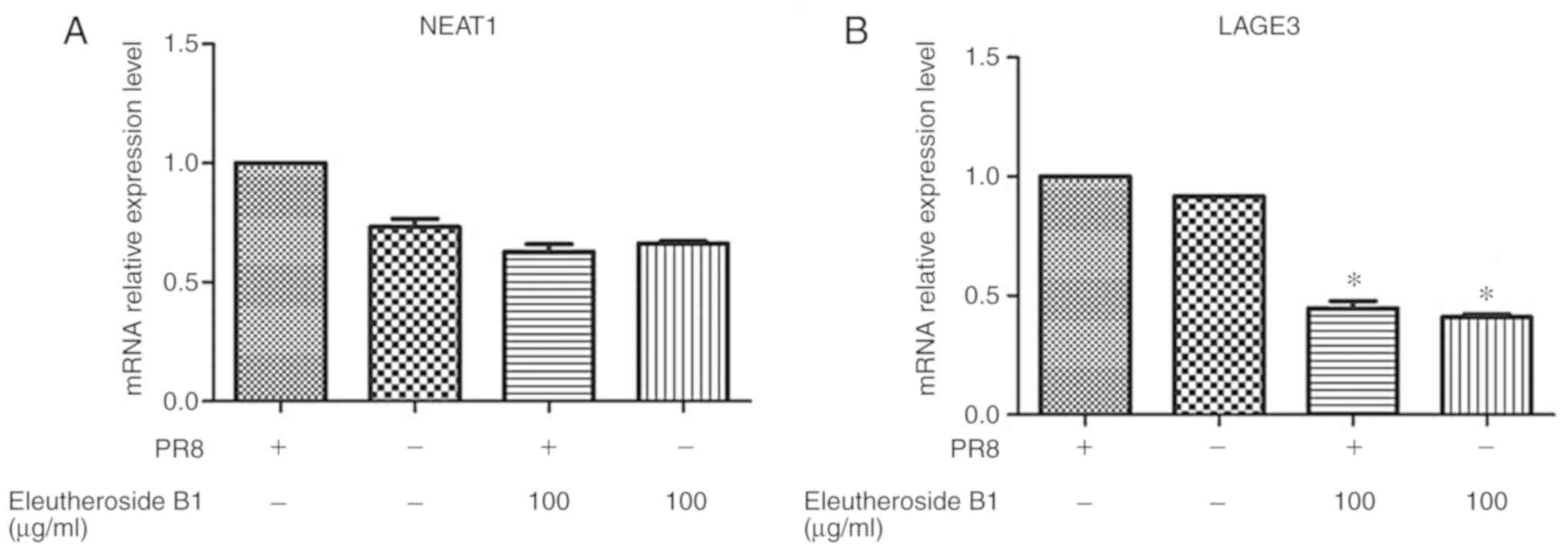
Validation of NEAT1 and LAGE3
The mRNA expression levels of ncRNA NEAT1 and LAGE3
were decreased by eleutheroside B1 treatment at concentrations of
100 µg/ml (Fig. 8).
However, the mRNA expression level of LAGE3 was significantly
downregulated by eleutheroside B1 (P<0.05). The effect of
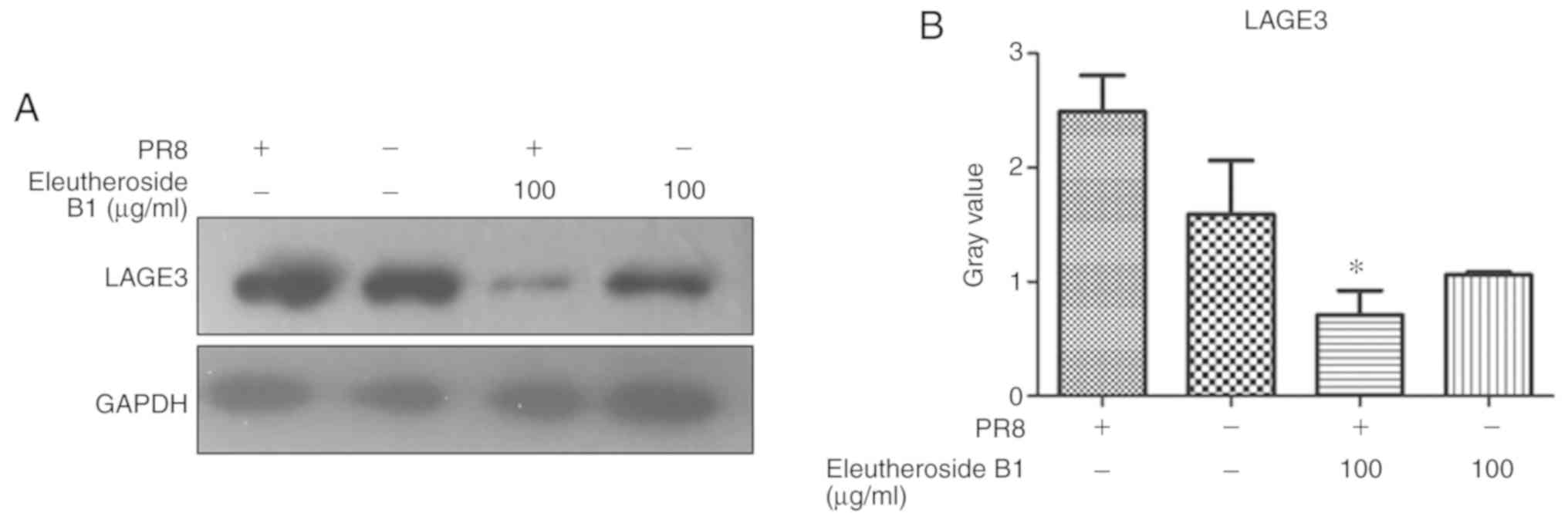
eleutheroside B1 on the expression of LAGE3 protein was also
confirmed by western blot analysis, and the result revealed a
decreased protein expression of LAGE3 in the eleutheroside B1
treatment group (Fig. 9).
Discussion
The underlying mechanisms of TCM against the
influenza virus at the molecular level are complex. High-throughput
RNA sequencing and iTRAQ assays are good methods which provide
comprehensive analysis of the mRNA, non-coding RNA and proteins
induced by disease pathogenesis and drugs (31,42,43). Utilizing these methods, multiple
aspects of mRNA, ncRNA and protein expression profiles were
profiled and characterized in A549 cells in response to
eleutheroside B1, following influenza virus infection. The result
of mRNA expression profiles have been previously published
(39); thus, only ncRNA and
protein expression profiles are shown in this study. The
transcriptome analysis revealed 8 differentially expressed ncRNAs
in the PR8+eleu vs. PR8 groups (5 upregulated ncRNAs and 3
downregulated RNAs) and 15 differentially expressed ncRNAs in the
PR8 vs. A549 cell groups (7 upregulated ncRNAs and 8 downregulated
RNAs). In these group comparisons, the same differentially
expressed ncRNA (NEAT1) was found to regulate the expression of
IL-8 through sequestering splicing factor proline-glutamine rich
(SFPQ/PSF) in paraspeckles, and is deregulated by stress, IAV and
HSV infection, or poly I:C treatment (22). NEAT1 was also essential in the
expression of various genes associated with innate immunity
(44). The formation of
subnuclear paraspeckle structures depend on transcription and RNA
polymerase II in response to stress or cellular metabolic changes
(45). The mRNA data analysis
also indicated that the effect of eleutheroside B1 on RNA
polymerase and RNA synthesis was required for the anti-influenza
activity (46). Previous studies
have proven that NEAT1 is a crucial innate immune molecule with
dual functions (44). On the one
hand, it is a critical part of the antiviral response to increase
the expression of immune cytokines, which on the other hand, could
be 'hijacked' by viruses to facilitate viral gene expression and
viral replication (44,47,48). The results of the present study
demonstrated that NEAT1 was downregulated by eleutheroside B1 in
A549 cells infected with the influenza virus, while it is
upregulated in the PR8 vs. the control A549 cells. The induction of
NEAT1 by the influenza virus is necessary for the successful
replication of the virus. The downregulation of NEAT1 by
eleutheroside B1 suggested that eleutheroside B1 may inhibit viral
replication and may decrease the expression of immune cytokines
through NEAT1; this finding is consistent the findings of a
previous study (13). In
addition, NEAT1 also regulates some pathways, such as the
Wnt/β-catenin signaling pathway (44). In the future, the authors of this
study aim to construct NEAT1 overexpression and knockout cells in
order to investigate the effects on the anti-influenza virus
activity of eleutheroside B1.
The GO and pathway analyses for predicted target
genes of altered ncRNAs in the PR8+eleu/PR8 groups revealed that
the majority of the genes that were enriched (P<0.05) belong to
biological process (single-organism process, biological regulation,
metabolic process, cellular process), cellular component
(macromolecular complex, organelle, membrane, cell part, cell, and
molecular function (catalytic activity, binding). According to the
KEGG pathway analysis, these genes associated with the
differentially expressed ncRNAs were involved in oxidative
phosphorylation, Parkinson's disease, graft-versus-host disease,
influenza A, allograft rejection, autoimmune thyroid disease, type
I diabetes mellitus, cytokine-cytokine receptor interaction
(49). The influenza virus
recruits variable and complementary host cellular pathways during
its replication cycle (50-52). Current anti-influenza virus
medication often targets one part of the virus life cycle, which
potentially influences the development of drug-resistance (2). The data suggest that the involvement
of eleutheroside B1 in altering cellular pathways is necessary for
the virus life cycle, such as binding and cytokine-cytokine
receptor interaction, consistent with previous studies (13,53). Cytokine-receptor complexes are
ideal drug targets. However, the protein-protein interactions are
difficult to block using small molecules (53,54).
Further proteomics analysis of the function of
eleuthero-side B1 was performed through iTRAQ assays. Only one
protein (LAGE3) was downregulated in the PR8+eleu vs. PR8 groups,
whereas 70 were upregulated and 5 proteins were downregulated in
the PR8+eleu vs. cell groups. LAGE3 belongs to the NY-ESO gene
family, a component of the EKC/KEOPS complex. This complex is
required for the formation of a threonylcarbamoyl group on
adenosine at position 37 (t6A37) in tRNAs that read codons
beginning with adenine (55,56). Some researchers have reported that
the EKC/KEOPS complex is linked with the protein synthesis
machinery and cell growth (57).
According to GO analysis, LAGE3 was involved in tRNA processing,
tRNA metabolic processes and lncRNA processing. These
differentially expressed proteins could be potential targets of
eleutheroside B1 in regulating the ncRNAs of host cells. Transfer
RNAs (tRNAs) are substrates for protein synthesis and a central
part of the translation machinery in all living organisms, which
impact translational speed and fidelity by their abundance. A
'channeled tRNA cycle' includes the following: tRNAs are shuttled
directly from the ribosome to their cognate tRNA synthetase and
back to the ribosome for another cycle of translation without
reentering the cytosolic pool (58,59). The polysome-associated tRNA
population changes dramatically, following influenza virus
infection; however, the total cellular tRNA population remains
unaltered (60). This proves that
the changes in polysome-associated tRNA levels reflect the codon
usage of viral genes, suggesting the existence of local tRNA pools
optimized for viral translation and enhanced translational
efficiency of viral genes. Over the past decade, ncRNAs (microRNAs,
lncRNAs and vtRNAs) have been identified as an important class of
regulators involved in virus-host interaction, particularly in
anti-viral immune response (20).
Focus on ncRNAs has recently been increased. Advances in the
high-throughput sequencing techniques are generating increasing
numbers of newly discovered ncRNAs that are involved in infections
and immunological processes (61). In some reports, ncRNAs have been
shown to regulate histone modifications in influenza virus-infected
cells, interact with transcription factors during virus infection
and induce specific protein-coding gene expression, affect virus
replication in an interferon-independent manner and control protein
synthesis during influenza virus infection (62-66). The LAGE3 gene involved in lncRNA
processing warrants further investigation to determine the role of
eleuthero-side B1 as a lncRNA regulator for its anti-influenza
virus.
Overall, the present study demonstrates that ncRNAs
are involved in the interaction between host and influenza virus.
Although research on non-coding RNA is currently unpopular in
Chinese medicine, it is worth noting that ncRNAs, such as NEAT1,
the protein LAGE3, and their pathways identified in the present
study indicate the need to investigate further the details of the
ability of eleutheroside B1 to inhibit influenza virus. One of the
limitations of the present study was that it was only performed
using A549 cells. It would be ideal to include animal or human
samples to profile the RNA and protein expression, particularly in
influenza virus-infected patients treated with this drug to compare
whether the observed changes are comparable in a clinical setting.
Unfortunately, the existing method for extracting eleutheroside B1
does not yield sufficient compounds to allow animal experiments and
clinical tests. However, in the future the authors aim to study
clinical samples treated with herba sarcandrae. In addition,
another limitation is that ncRNA expression was not examined at
different time points. Previous research (46) has often selected 24 h after
infection for RNA-seq; therefore, this time point was used for
detection in this study. In the future, the authors aim to perform
further studies using different time points. Another limitation of
the present study is that only the iTRAQ assay was used to
investigate protein expression, which yields incomplete data due to
the use of data-dependent acquisition that only captures one
differentially expressed protein. Thus, it will be interesting to
examine protein expression profiles induced by eleutheroside B1 by
other proteomics methods, being cautious that ncRNAs and proteins
may function in a coordinated fashion, and therefore, the overall
anti-influenza virus response induced by eleutheroside B1 may be
due to the effect of a cumulative response to overall changes in
ncRNAs or proteins rather than individual ncRNAs or proteins.
Furthermore, certain studies open novel avenues to further explore
the molecular pathways based on ncRNA or protein functions.
Therefore, whether these networks were induced by eleutheroside B1
in the future should be investigated.
In the validation assays, the RT-qPCR results
revealed that the expression levels of ncRNA NEAT1 and mRNA LAGE3
were decreased by eleutheroside B1 (100 µg/ml) treatment.
The mRNA expression level of LAGE3 was significantly downregulated
by eleutheroside B1 (P<0.05). Western blot analysis also
demonstrated that the protein expression of LAGE3 was decreased
with eleutheroside B1 treatment. For NEAT1, the ncRNA and not the
mRNA, which cannot be investigated in protein level using recent
techniques. In this study, only RT-qPCR and western blot analysis
were used to examine these genes of interest. However, further
molecular assays, such as immunofluorescence assay, should be
adopted to examine these genes in the future. In addition, the
authors aim to perform further studies in which NEAT1- and
LAGE3-overexpressing cells and knockout cells will be constructed
to investigate their roles in the anti-influenza virus activity of
eleutheroside B1.
Funding
The present study was supported by the National
Natural Science Foundation of China (U1502226), the Engineering
technology research center (development) of Guangdong general
universities (GCZX-A1408), Natural Science Basic Research Program
of Shaanxi (Program No. 2019JM-513) and Shijiazhuang Yiling
Pharmaceutical Co.,Ltd (Shijiazhuang, China).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article or are available from
the corresponding author on reasonable request.
Authors' contributions
WY wrote the manuscript and performed experiments.
JC drafted the manuscript and performed experiments. ZW revised the
manuscript and performed experiments. XiaohuW designed, edited and
modified the manuscript. ZZ performed experiments and modified the
manuscript. DT designed this study and reviewed the manuscript.
YutaoW and XinhuaW obtained funds and designed this study. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Yoon SW, Webby RJ and Webster RG:
Evolution and ecology of influenza A viruses. Curr Top Microbiol
Immunol. 385:359–375. 2014.PubMed/NCBI
|
|
2
|
Amarelle L, Lecuona E and Sznajder JI:
Anti-influenza treatment: Drugs currently used and under
development. Arch Bronconeumol. 53:19–26. 2017.In English, Spanish.
View Article : Google Scholar
|
|
3
|
Takamatsu K, Marumo S, Fukui M and Hata A:
Safety and efficacy of anti-influenza drugs, intravenous peramivir
against influenza virus infection in elderly patients with
underlying disease. J Microbiol Immunol Infect. 50:541–544. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moss RB, Davey RT, Steigbigel RT and Fang
F: Targeting pandemic influenza: A primer on influenza antivirals
and drug resistance. J Antimicrob Chemother. 65:1086–1093. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Oh DY, Panozzo J, Vitesnik S, Farrukee R,
Piedrafita D, Mosse J and Hurt AC: Selection of multi-drug
resistant influenza A and B viruses under zanamivir pressure and
their replication fitness in ferrets. Antivir Ther. 23:295–306.
2018. View
Article : Google Scholar
|
|
6
|
Deyde VM, Xu X, Bright RA, Shaw M, Smith
CB, Zhang Y, Shu Y, Gubareva LV, Cox NJ and Klimov AI: Surveillance
of resistance to adamantanes among influenza A(H3N2) and A(H1N1)
viruses isolated worldwide. J Infect Dis. 196:249–257. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yen HL, McKimm-Breschkin JL, Choy KT, Wong
DD, Cheung PP, Zhou J, Ng IH, Zhu H, Webby RJ, Guan Y, et al:
Resistance to neuraminidase inhibitors conferred by an R292K
mutation in a human influenza virus H7N9 isolate can be masked by a
mixed R/K viral population. mBio. 4:pii: e00396-13. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gurib-Fakim A: Medicinal plants:
Traditions of yesterday and drugs of tomorrow. Mol Aspects Med.
27:1–93. 2006. View Article : Google Scholar
|
|
9
|
Pan SY, Zhou SF, Gao SH, Yu ZL, Zhang SF,
Tang MK, Sun JN, Ma DL, Han YF, Fong WF and Ko KM: New perspectives
on how to discover drugs from herbal medicines: CAM's outstanding
contribution to modern therapeutics. Evid Based Complement Alternat
Med. 2013:6273752013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang L, Zhang RM, Liu GY, Wei BL, Wang Y,
Cai HY, Li FS, Xu YL, Zheng SP and Wang G: Chinese herbs in
treatment of influenza: A randomized, double-blind,
placebo-controlled trial. Respir Med. 104:1362–1369. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin TJ, Lin CF, Chiu CH, Lee MC and Horng
JT: Inhibition of endosomal fusion activity of influenza virus by
Rheum tanguticum (da-huang). Sci Rep. 6:277682016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu W, Li R, Li X, He J, Jiang S, Liu S and
Yang J: Quercetin as an antiviral agent inhibits influenza a virus
(IAV) entry. Viruses. 8:pii: E6. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Yan W, Chen Q, Huang W, Yang Z, Li
X and Wang X: Inhibition viral RNP and anti-inflammatory activity
of coumarins against influenza virus. Biomed Pharmacother.
87:583–588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Harrow J, Frankish A, Gonzalez JM,
Tapanari E, Diekhans M, Kokocinski F, Aken BL, Barrell D, Zadissa
A, Searle S, et al: GENCODE: The reference human genome annotation
for the ENCODE project. Genome Res. 22:1760–1774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He JH, Han ZP and Li YG: Association
between long non-coding RNA and human rare diseases (Review).
Biomed Rep. 2:19–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li W, Li N, Shi K and Chen Q: Systematic
review and meta-analysis of the utility of long non-coding RNA GAS5
as a diagnostic and prognostic cancer biomarker. Oncotarget.
8:66414–66425. 2017.PubMed/NCBI
|
|
17
|
Shin SY, Jeong JS, Lim JY, Kim T, Park JH,
Kim JK and Shin C: Transcriptomic analyses of rice (Oryza sativa)
genes and non-coding RNAs under nitrogen starvation using multiple
omics technologies. BMC Genomics. 19:5322018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie H, Ma B, Gao Q, Zhan H, Liu Y, Chen Z,
Ye S, Li J, Yao L and Huang W: Long non-coding RNA CRNDE in cancer
prognosis: Review and meta-analysis. Clin Chim Acta. 485:262–271.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ng SS, Li OT, Cheung TK, Malik Peiris JS
and Poon LL: Heterologous influenza vRNA segments with identical
non-coding sequences stimulate viral RNA replication in trans.
Virol J. 5:22008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Winterling C, Koch M, Koeppel M,
Garcia-Alcalde F, Karlas A and Meyer TF: Evidence for a crucial
role of a host non-coding RNA in influenza A virus replication. RNA
Biol. 11:66–75. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li F, Chen Y, Zhang Z, Ouyang J, Wang Y,
Yan R, Huang S, Gao GF, Guo G and Chen JL: Robust expression of
vault RNAs induced by influenza A virus plays a critical role in
suppression of PKR-mediated innate immunity. Nucleic Acids Res.
43:10321–10337. 2015.PubMed/NCBI
|
|
22
|
Ma Y, Ouyang J, Wei J, Maarouf M and Chen
JL: Involvement of host non-coding RNAs in the pathogenesis of the
influenza virus. Int J Mol Sci. 18:pii: E39. 2016. View Article : Google Scholar
|
|
23
|
Valadkhan S and Plasek LM: Long non-coding
RNA-mediated regulation of the interferon response: A new
perspective on a familiar theme. Pathog Immun. 3:126–148. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Y, Zhang J, Huo C, Ding N, Li J, Xiao
J, Lin X, Cai B, Zhang Y and Xu J: Dynamic organization of lncRNA
and circular RNA regulators collectively controlled cardiac
differentiation in humans. EBioMedicine. 24:137–146. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Okholm TLH, Nielsen MM, Hamilton MP,
Christensen LL, Vang S, Hedegaard J, Hansen TB, Kjems J, Dyrskjøt L
and Pedersen JS: Circular RNA expression is abundant and correlated
to aggressiveness in early-stage bladder cancer. NPJ Genom Med.
2:362017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen YG, Kim MV, Chen X, Batista PJ,
Aoyama S, Wilusz JE, Iwasaki A and Chang HY: Sensing self and
foreign circular RNAs by intron identity. Mol Cell. 67:228–238.e5.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Liu CX, Xue W, Zhang Y, Jiang S, Yin
QF, Wei J, Yao RW, Yang L and Chen LL: Coordinated circRNA
biogenesis and function with NF90/NF110 in viral infection. Mol
Cell. 67:214–227.e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brahmachari G: Discovery and development
of antidiabetic agents from natural products. 1st edition. Waltham
MA: Elsevier Inc; 2016
|
|
29
|
Luo R and Zhao H: Protein quantitation
using iTRAQ: Review on the sources of variations and analysis of
nonrandom missingness. Stat Interface. 5:99–107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Smith SB, Dampier W, Tozeren A, Brown JR
and Magid-Slav M: Identification of common biological pathways and
drug targets across multiple respiratory viruses based on human
host gene expression analysis. PLoS One. 7:e331742012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou B, Li J, Liang X, Yang Z and Jiang Z:
Transcriptome profiling of influenza A virus-infected lung
epithelial (A549) cells with lariciresinol-4-β-D-glucopyranoside
treatment. PLoS One. 12:e01730582017. View Article : Google Scholar
|
|
32
|
Li X, Zhang Y, Zeng X, Yang L and Deng Y:
Chemical profiling of bioactive constituents in Sarcandra glabra
and its preparations using ultra-high-pressure liquid
chromatography coupled with LTQ Orbitrap mass spectrometry. Rapid
Commun Mass Spectrom. 25:2439–2447. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schroeder A, Mueller O, Stocker S,
Salowsky R, Leiber M, Gassmann M, Lightfoot S, Menzel W, Granzow M
and Ragg T: The RIN: An RNA integrity number for assigning
integrity values to RNA measurements. BMC Mol Biol. 7:32006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han Li C and Chen Y: Small and long
non-coding RNAs: Novel targets in perspective cancer therapy. Curr
Genomics. 16:319–326. 2015. View Article : Google Scholar
|
|
35
|
Kang YJ, Yang DC, Kong L, Hou M, Meng YQ,
Wei L and Gao G: CPC2: A fast and accurate coding potential
calculator based on sequence intrinsic features. Nucleic Acids Res.
45(W1): W12–W16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qu Z and Adelson DL: Bovine ncRNAs are
abundant, primarily intergenic, conserved and associated with
regulatory genes. PLoS One. 7:e426382012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Qu Z and Adelson DL: Identification and
comparative analysis of ncRNAs in human, mouse and zebrafish
indicate a conserved role in regulation of genes expressed in
brain. PLoS One. 7:e522752012. View Article : Google Scholar
|
|
38
|
Liu YC, Li JR, Sun CH, Andrews E, Chao RF,
Lin FM, Weng SL, Hsu SD, Huang CC, Cheng C, et al: CircNet: A
database of circular RNAs derived from transcriptome sequencing
data. Nucleic Acids Res. 44(D1): D209–D215. 2016. View Article : Google Scholar :
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
40
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42(Database Issue): D92–D97. 2014. View Article : Google Scholar
|
|
41
|
Rinn JL: lncRNAs: Linking RNA to
chromatin. Cold Spring Harb Perspect Biol. 6:pii: a018614. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hu J, Gao Z, Wang X, Gu M, Liang Y, Liu X,
Hu S, Liu H, Liu W, Chen S, et al: iTRAQ-based quantitative
proteomics reveals important host factors involved in the high
pathogenicity of the H5N1 avian influenza virus in mice. Med
Microbiol Immunol. 206:125–147. 2017. View Article : Google Scholar
|
|
43
|
Xiao YL, Kash JC, Beres SB, Sheng ZM,
Musser JM and Taubenberger JK: High-throughput RNA sequencing of a
formalin-fixed, paraffin-embedded autopsy lung tissue sample from
the 1918 influenza pandemic. J Pathol. 229:535–545. 2013.
View Article : Google Scholar :
|
|
44
|
Imamura K, Imamachi N, Akizuki G, Kumakura
M, Kawaguchi A, Nagata K, Kato A, Kawaguchi Y, Sato H, Yoneda M, et
al: Long noncoding RNA NEAT1-dependent SFPQ relocation from
promoter region to paraspeckle mediates IL8 expression upon immune
stimuli. Mol Cell. 53:393–406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mao YS, Zhang B and Spector DL: Biogenesis
and function of nuclear bodies. Trends Genet. 27:295–306. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yan W, Zheng C, He J, Zhang W, Huang XA,
Li X, Wang Y and Wang X: Eleutheroside B1 mediates its
anti-influenza activity through POLR2A and N-glycosylation. Int J
Mol Med. 42:2776–2792. 2018.PubMed/NCBI
|
|
47
|
Hirose T, Virnicchi G, Tanigawa A,
Naganuma T, Li R, Kimura H, Yokoi T, Nakagawa S, Bénard M, Fox AH
and Pierron G: NEAT1 long noncoding RNA regulates transcription via
protein sequestration within subnuclear bodies. Mol Biol Cell.
25:169–183. 2014. View Article : Google Scholar :
|
|
48
|
Landeras-Bueno S, Jorba N, Pérez-Cidoncha
M and Ortin J: The splicing factor proline-glutamine rich
(SFPQ/PSF) is involved in influenza virus transcription. PLoS
Pathog. 7:e10023972011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Samji T: Influenza A: Understanding the
viral life cycle. Yale J Biol Med. 82:153–159. 2009.PubMed/NCBI
|
|
50
|
Li C, Bankhead A III, Eisfeld AJ, Hatta Y,
Jeng S, Chang JH, Aicher LD, Proll S, Ellis AL, Law GL, et al: Host
regulatory network response to infection with highly pathogenic
H5N1 avian influenza virus. J Virol. 85:10955–10967. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Luo M: Influenza virus entry. Adv Exp Med
Biol. 726:201–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rudnicka A and Yamauchi Y: Ubiquitin in
influenza virus entry and innate immunity. Viruses. 8:pii: E293.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Schreiber G and Walter MR:
Cytokine-receptor interactions as drug targets. Curr Opin Chem
Biol. 14:511–519. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Spangler JB, Moraga I, Mendoza JL and
Garcia KC: Insights into cytokine-receptor interactions from
cytokine engineering. Annu Rev Immunol. 33:139–167. 2015.
View Article : Google Scholar
|
|
55
|
Long M, Wang Y, Chen D, Wang Y, Wang R,
Gong D, He H, Rock DL, Hao W and Luo S: Identification of host
cellular proteins LAGE3 and IGFBP6 that interact with orf virus
protein ORFV024. Gene. 661:60–67. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wan LC, Maisonneuve P, Szilard RK, Lambert
JP, Ng TF, Manczyk N, Huang H, Laister R, Caudy AA, Gingras AC, et
al: Proteomic analysis of the human KEOPS complex identifies
C14ORF142 as a core subunit homologous to yeast Gon7. Nucleic Acids
Res. 45:805–817. 2017. View Article : Google Scholar :
|
|
57
|
Rojas-Benitez D, Ibar C and Glavic Á: The
Drosophila EKC/KEOPS complex: Roles in protein synthesis
homeostasis and animal growth. Fly (Austin). 7:168–172. 2013.
View Article : Google Scholar
|
|
58
|
Petrushenko ZM, Budkevich TV, Shalak VF,
Negrutskii BS and El'skaya AV: Novel complexes of mammalian
translation elongation factor eEF1A.GDP with uncharged tRNA and
aminoacyl-tRNA synthetase. Implications for tRNA channeling. Eur J
Biochem. 269:4811–4818. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Stapulionis R and Deutscher MP: A
channeled tRNA cycle during mammalian protein synthesis. Proc Natl
Acad Sci USA. 92:7158–7161. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pavon-Eternod M, David A, Dittmar K,
Berglund P, Pan T and Bennink JR: Vaccinia and influenza A viruses
select rather than adjust tRNAs to optimize translation. Nucleic
Acids Res. 41:1914–1921. 2013. View Article : Google Scholar :
|
|
61
|
Landeras-Bueno S and Ortin J: Regulation
of influenza virus infection by long non-coding RNAs. Virus Res.
212:78–84. 2016. View Article : Google Scholar
|
|
62
|
Barriocanal M, Carnero E, Segura V and
Fortes P: Long non-coding RNA BST2/BISPR is induced by IFN and
regulates the expression of the antiviral factor tetherin. Front
Immunol. 5:6552015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
D'Orso I and Frankel AD: RNA-mediated
displacement of an inhibitory snRNP complex activates transcription
elongation. Nat Struct Mol Biol. 17:815–821. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kambara H, Niazi F, Kostadinova L, Moonka
DK, Siegel CT, Post AB, Carnero E, Barriocanal M, Fortes P, Anthony
DD and Valadkhan S: Negative regulation of the interferon response
by an interferon-induced long non-coding RNA. Nucleic Acids Res.
42:10668–10680. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mariner PD, Walters RD, Espinoza CA,
Drullinger LF, Wagner SD, Kugel JF and Goodrich JA: Human Alu RNA
is a modular transacting repressor of mRNA transcription during
heat shock. Mol Cell. 29:499–509. 2008. View Article : Google Scholar : PubMed/NCBI
|