Introduction
Periodontitis can cause progressive destruction of
dental support tissue, which is the primary risk factor for tooth
loss in adults (1-3). Traditional non-surgical treatments,
such as scaling, curettage and root surface leveling, can only
remove plaque and reduce inflammation (4), while surgical treatments such as
guided tissue regeneration and guided bone regeneration have
limited regenerative effects (5).
Technologies that can achieve periodontal tissue regeneration and
functional recovery currently remain under investigation. In recent
years, periodontal tissue engineering has developed rapidly,
providing a new direction for research. Periodontal tissue
engineering is a technique that uses tissue engineering to
construct a complex composed of one or more elements (such as
cells, scaffolds and signaling molecules) for implantation into the
defect area to achieve functional tissue regeneration (6,7).
However, ongoing challenges for this technique include the
identification of the best source of cells to construct such
tissue-engineered complexes, as well as the selection of biological
signaling molecules to stimulate the regenerative potential of
implanted stem cells in order to enhance their therapeutic
effects.
It has been reported that the ephrinB2/ephrin type-B
receptor 4 (EphB4) signaling pathway serves an important role in
bone remodeling processes. The reverse signaling pathway mediated
by ephrinB2 inhibits the function of osteoclasts and bone
resorption by decreasing c-Fos and NFATc1 activity (8). By contrast, the forward signaling
pathway mediated by EphB4 enhances osteoblast differentiation and
promotes bone formation by decreasing RhoA activity (9). In addition, ephrinB2-Fc can bind to
EphB4 receptors on the surface of endothelial cells and promote the
migration of vascular endothelial cells by activating the
phosphatidylinositol 3-kinase signaling pathway to accelerate
angiogenesis (10-12). Our previous experiments verified
that ephrinB2 gene-transfected dental stem cells enhanced the
osteogenic ability and promoted the angiogenesis of peripheral
vascular endothelial cells (13),
as well as promoted their angiogenesis in co-culture systems
(14). In addition, we observed
that the extracellular matrix of umbilical vein endothelial cells
was able to promote the differentiation of odontogenic stem cells
into endothelial cells (15).
Therefore, it can be speculated that the ephrinB2 signaling
molecule enhances cell migration, promotes odontogenic stem cell
osteogenesis and enhances peripheral angiogenesis. As such,
ephrinB2 is an ideal biological signaling molecule for osteogenic
regeneration.
Previous studies reported that a multitude of mature
stem cells can be derived from dental tissues, including
periodontal ligament stem cells (PDLSCs), dental pulp stem cells
(DPSCs), apical papilla and stem cells from human exfoliated
deciduous teeth (SHEDs), all of which exhibit a potent capacity for
differentiation into mesodermal lineages (16-18). Mesenchymal stem cells possessing
stem-like features can produce osteo/odontogenic, adipogenic and
neurogenic lineages. Notably, DPSCs and SHEDs have been
successfully induced into endothelial cells both in vitro
and in vivo (19-22), as have bone marrow mesenchymal
stem cells and human adipose-derived stem cells (23,24). Collectively, these results
indicate that dental stem cells are a suitable candidate for tissue
engineering.
While non-animal methods are widely used in
biomedical research, they cannot replace all use of animals.
Currently, the canine mandible defect model is widely used in
preclinical studies (25-27). Notably, beagle dogs are
recommended as potential animal models for medical testing due to
their genetic, biological and behavioral characteristics that
closely resemble those of humans; furthermore, numerous symptoms of
human conditions can be replicated in dogs (28). Previous studies comparing canine
and human dental mesenchymal stem cells have demonstrated that
canine DPSCs were able to differentiate into odontoblast-like cells
exhibiting osteogenic potential similar to human DPSCs (29). However, in vitro
transfection of canine PDLSCs (cPDLSCs) with ephrinB2 and the use
of transfected cells as seed cells for tissue engineering have
seldom been investigated. In addition, how ephrinB2/EphB4 signaling
may facilitate the osteogenic/odontogenic differentiation of
cPDLSCs has not been previously investigated. Therefore, the main
focus of the current study was to address these issues.
Materials and methods
Isolation, culture and identification of
cells
cPDLSCs were isolated from 6-month-old beagle dogs.
A total of 4 6-month old male beagle dogs (JC0853, 8.0 kg; JC0857,
7.5 kg; JC0889, 6.6 kg; JC0899, 7.4 kg) were obtained from the
Experimental Animal Center of Xuzhou Medical University (Xuzhou,
China). The dogs were bred carefully and exclusively by
professional breeders and housed in individual and clean cages at
room temperature with a humidity of 60%, and regular food and water
were provided twice daily. Ethical approval for the present study
was obtained from the Institutional Animal Care and Use Committee
of Xuzhou Medical University (Xuzhou, China; approval no.
20161108). Briefly, the dogs were induced to initial anesthesia by
intravenous injection of 6 mg/kg propofol and maintained by
inhalation of isoflurane at 2%, with the oxygen flow maintained at
3 l/min for 1 min and then adjusted to 0.4 l/min. The upper and
lower anterior teeth were extracted, using phosphate-buffered
saline containing penicillin and streptomycin to wash and separate
the periodontal ligament. Next, the periodontal ligament tissue was
scraped from the teeth and shattered on a super-clean table using a
mixture of 3 mg/ml collagenase I (Gibco; thermo Fisher Scientific,
Inc.) and 4 mg/ml neutral protease (Dispase; (Gibco; thermo Fisher
Scientific, Inc.) to digest and extract the cells. The cells were
cultured in α-minimum essential medium (α-MEM; Gibco; thermo Fisher
Scientific, Inc.) supplemented with 20% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc.) in a 37°C, 5% CO2
incubator. The limiting dilution method was adopted to separate
cPDLSCs from total canine periodontal ligament cells and their
morphology was monitored under an inverted microscope. Flow
cytometric analysis for STRO-1 (cat. no. 14-6688-82; thermo Fisher
Scientific, Inc.), CD45 (cat. no. MCA2035S; Bio-Rad Laboratories,
Inc.), CD73 (cat. no. 202122; Abcam), CD90 (cat. no. ab139364;
Abcam) and CD105 (cat. no. ab156756; Abcam) antibodies (all 1:500)
was employed to detect the phenotype of cells. Briefly, cells were
incubated with primary antibodies for 2 h in dark at room
temperature and then washed with PBS. This was followed by
incubation with the following appropriate secondary antibodies:
Goat anti-mouse IgG H&L (Alexa Fluor® 488)
pre-adsorbed (1:2,000; cat. no. ab150117; Abcam) and goat
anti-rabbit IgG H&L (Alexa Fluor® 488; 1:2,000; cat.
no. ab150077; Abcam) for 30 min in the dark at room temperature. A
BD FACSVerse™ flow cytometer (BD Biosciences) was used
to detect cells and BD FACSuite software V1.0 (BD Biosciences) were
used to detect and analyze flow cytometry data.
Furthermore, other appropriate methods were
utilized, as described below, to assess multidirectional
differentiation ability of the cells. For osteogenic
differentiation, cPDLSCs were seeded at a density of
1×104/well in 12-well plates and cultured to 70%
confluence. Subsequently, the osteogenic induction medium (α-MEM
containing 10% FBS, 50 µg/ml L-ascorbic acid phosphate, 10
mmol/l β-glycerophosphate and 10 nmol/l dexamethasone) was added.
Following 4 weeks of culture at 37°C, cells were fixed with 4%
paraformaldehyde for 30 min and stained with 0.2% Alizarin Red S
solution for 10 min at room temperature for mineralized nodules.
For adipogenic differentiation, cPDLSCs, seeded at a density of
1×104/well, were cultured to 70% confluence and cultured
with odipogenic induction medium (α-MEM containing 10% FBS, 1
µg/ml insulin, 1 µmol/l dexamethasone and 0.5 mmol/l
3-isobutyl-1-methylxanthine) for 4 weeks at 37°C. Finally, the
lipid droplets were stained with 0.5% Oil Red O for 10 min at room
temperature. For neurogenic differentiation, cPDLSCs seeded at a
density of 1×104/well in 12-well plates were cultured in
Neurobasal A medium (Gibco; thermo Fisher Scientific, Inc.)
supplemented with 20 ng/ml epidermal growth factor (Peprotech,
Inc.) and 40 ng/ml basic fibroblast growth factor (bFGF; Peprotech,
Inc.) for 4 weeks at 37°C. βIII-tubulin was detected by
immunofluorescence.
Immunofluorescence analysis
Following rinsing with PBS, cPDLSCs were grown on
glass cover slides, fixed with 4% paraformaldehyde for 10 min at
room temperature and blocked with 1% BSA for 30 min at room
temperature. Slides were incubated with primary mouse
anti-βIII-tubulin antibody (1:500; Abcam; cat. no. ab119100) at
room temperature in a humidified chamber overnight. Following
thorough rinsing with PBS, slides were then incubated with goat
anti-mouse IgG (H+L) secondary antibody overnight. Subsequently,
4,6-diamidino-2-phenylindole was used to label cell nuclei. The
cover slides were sealed with an aqueous-based mounting medium
(Vector Laboratories). For immunofluorescence microscopy, an
inverted Nikon fluorescence microscope (Nikon Corporation) with a
digital camera (magnification, ×20) and deconvolution software
(Slidebook 4.0; Intelligent Imaging) were used.
Transfection and identification of
ephrinB2 gene-transfected cPDLSCs
cPDLSCs were transfected with a null-control green
fluorescent protein (GFP)-Blasticidin Vector (Vector-cPDLSCs) or
NFNB2 GFP-Blasticidin Vector (EfnB2-cPDLSCs). Approximately 18-24 h
before transfection, adherent second-generation cPDLSCs were
transferred to a 24-well plate at a density of
1×105/well. The density of lentivirus-transfected cells
was ~2×105/well. The following day, the original medium
was replaced with 2 ml fresh medium containing 6 µg/ml
polybrene, and 10 µg of the virus suspension was added for
incubation at 37°C for 4 h. Next, 2 ml fresh medium was added to
dilute the polybrene, and the cells were cultured for a further 24
h. Subsequently, the medium containing the virus was replaced with
fresh medium, and cells were cultured for another 24 h. After 24 h
of transfection, GFP fluorescence was observed under a
fluorescent-inverted phase-contrast microscope (Olympus
Corporation). After 3-4 days of transfection, Blasticidin-specific
antibiotics (Merck KGaA) were added continuously with monitoring to
ensure that the cells were subcultured normally. Following
Blasticidin selection, 1×105 EfnB2-cPDLSCs and
vector-PDLSCs were cultured in six-well plates at 37°C for 48 or 72
h. Reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blotting were used to respectively detect
gene and protein expression levels of ephrinB2 in vector-cPDLSCs
and EfnB2-cPDLSCs following transfection. Wild-type cPDLSCs were
used as control. All experiments were performed in triplicate.
Cell proliferation assay
A Cell Counting Kit-8 (CCK-8; Sigma Aldrich; Merck
KGaA) assay was used to assess cell proliferation. Briefly,
cPDLSCs, EfnB2-cPDLSCs or Vector-cPDLSCs were seeded at a density
of 1×104 cells per well into a 96-well plate overnight
(37°C, 5% CO2). Next, 10 µl CCK-8 solution was
added to each well, followed by incubation for 4 h (37°C, 5%
CO2). The absorbance of each well was read at 450 nm
with a microplate reader on days 0, 2, 4, 6 and 8 to assess the
cell numbers.
Cell migration assay
A Transwell cell migration assay was used to detect
the cell migration as previously described (30). Briefly, transwell filter inserts
were placed in 24-well tissue culture plates (Boyden chambers; BD
LabWare), and after 12 h of serum starvation, 5×104
cPDLSCs, EfnB2-cPDLSCs or Vector-cPDLSCs were cultured in the upper
chamber with serum-free α-MEM. α-MEM supplemented with 15% fetal
bovine serum was added to the lower chamber. At 6, 8, 10 and 12 h,
viable cells that had migrated to the lower membrane surface were
fixed with 4% (w/v) paraformaldehyde for 30 min at room temperature
and stained with 0.1% (w/v) crystal violet for 10 min at room
temperature. Images were captured using an inverted microscope
(Olympus Corporation) and counted to quantify the migrated
cells.
Detection of osteogenic and angiogenic
abilities of cPDLSCs in vitro
Vector-cPDLSCs and EfnB2-cPDLSCs were separately
cultured in osteogenic induction medium (α-MEM containing 10% FBS,
50 µg/ml L-ascorbic acid phosphate, 10 mmol/l
β-glycerophosphate, and 10 nmol/l dexamethasone) for 7, 14 or 21
days. The expression levels of osteogenesis- and
angiogenesis-associated proteins were then detected by western
blotting, as described later in the text. Alkaline phosphatase
(ALP) assay and Alizarin Red S staining were also performed.
Briefly, for ALP assay, the BCIP/NBT ALP Color Development kit
(Beyotime Institute of Biotechnology) was used on day 14 of
induction, and the ALP activity was assessed by an ALP Detection
kit (Jiancheng Bioengineering Institute) on days 14, following the
manufacturer's protocol. For Alizarin Red S staining, the cultures
were fixed with 4% paraformaldehyde for 30 min at room temperature
after 3 weeks of incubation (37°C, 5% CO2) and then
stained with 0.2% Alizarin Red S (Sigma-Aldrich; Merck KGaA)
solution for 10 min at room temperature for mineralized nodules. To
quantify the calcium mineralization of EfnB2-cPDLSCs and
Vector-cPDLSCs within each well, sodium dodecyl sulfate (SDS; 10%
w/v) solution was added, and the cells were incubated overnight at
37°C. The absorbance of lysates at 405 nm was then read with a
SpectraMAX 340® microplate reader (Molecular Devices,
LLC).
In order to investigate the mechanism of ephrinB2 in
regulating osteogenic differentiation, ephrinB2 was blocked with
ephrinB2 small interfering RNA (EphrinB2-siRNA; thermo Fisher
Scientific, Inc.). Briefly, cPDLSCs were seeded on 6-well plates
until 70% confluence was reached, and then 30 µl Opti-MEM
along with 500 µl Lipofectamine 2000 and RNAiMAX reagent
(all from thermo Fisher Scientific, Inc.) were added to the cells.
Next, 3 µl siRNA was diluted in 150 µl Opti-MEM prior
to mixing it with the diluted RNAiMAX and Lipofectamine at a ratio
of 1:1. The siRNA-lipid complex was incubated for 5 min at room
temperature. Subsequently, cPDLSCs were incubated with the
siRNA-lipid complex for 24 h prior to the osteogenic induction
experiments, and then ALP and Alizarin Red S staining assays were
performed. FAM-labeled negative control (NC)-siRNA (CN2001; Biomics
Biotechnologies Co., Ltd.) was used as the control in this
experiment.
RT-qPCR
EfnB2-PDLSCs and Vector-cPDLSCs were cultured for 7,
14 or 21 days in normal α-MEM or osteogenic/odontogenic induction
medium, and then total RNA was extracted with an RNeasy Plus Mini
kit (Qiagen) and quantified using a NanoDrop2000 spectrophotometer
(thermo Fisher Scientific, Inc.). Subsequently, 1.0 µg total
RNA was reverse transcribed into cDNA in a total reaction mixture
of 10 µl using Super-Script VILO Master mix (Invitrogen;
Thermo Fisher Scientific, Inc.). qPCR was then conducted using an
ABI Prism 7000 Sequence Detection system with SYBR-Green reagent
(both from Applied Biosystems; thermo Fisher Scientific, Inc.). The
reaction program used for qPCR was as follows: 95°C for 10 min, 40
cycles of 95°C for 15 sec and 60°C for 1 min, followed by melt
curve analysis at 95°C for 15 sec, 60°C for 1 min, 95°C for 15 sec
and 60°C for 15 sec. The relative expression values of the target
genes were determined using the 2−ΔΔCq method (31) by normalization with the
housekeeping gene GAPDH as an internal control. The primer
sequences are listed in Table I.
Standards and samples were run in triplicate.
 | Table IPrimers used in polymerase chain
reaction. |
Table I
Primers used in polymerase chain
reaction.
| Canine gene | Primers
(5′-3′) |
|---|
| BMP2 | F:
CCCTACATGCTGGACCTGTA |
| R:
ATTTCTGGCAGTTCTTCCAA |
| Runx2 | F:
TACCACACCTACCTGCCACCAC |
| R:
GCGGAAGCATTCTGGAAGGAGAC |
| Col1 | F:
GGTTCAGCTAAGTTGGAGGTACT |
| R:
CCAATGTTGCCAGGGTAAC |
| OCN | F:
CTGGTCCAGCAGATGCAAAG |
| R:
CCGCTTGGACACGAAGGTT |
| BSP | F:
TTGCTCAGCATTTTGGGAATGG |
| R:
AACGTGGCCGATACTTAAAGACC |
| ALP | F:
TTCAAACCGAGACACAAGCACT |
| R:
GGGTCAGTCACGTTGTTCCTGT |
| DMP1 | F:
CAGGAGCACAGGAAAAGGAG |
| R:
CTGGTGGTATCTTGGGCACT |
| DSPP | F:
GTCCTAGTGGGAATGGAGCA |
| R:
TCTTCAGGGCCATCATCTTC |
| GAPDH | F:
TGTCCCCACCCCCAATGTATC |
| R:
CTCCGATGCCTGCTTCACTACCTT |
Western blot analysis
Vector-cPDLSCs and EfnB2-cPDLSCs were seeded at a
density of 1×105 cells per well in 6-well plates. When
the cells reached 70% confluence, 2 µg/ml
erythropoietin-producing human hepatocellular receptors B4 fragment
(EphB4-FC, R&D Systems) was added to activate ephrinB2 ligand.
The cells were stimulated with EphB4-FC for 0, 5, 10, 20, 30 and 60
min or induced with osteogenic medium for 0, 12, 24, 48 and 72 h,
and then M-PER™ Protein Extraction buffer containing 1X protease
inhibitor cocktail (thermo Fisher Scientific, Inc.) was used to
extract the total protein. A BCA kit (thermo Fisher Scientific,
Inc.) was used to quantify the protein concentrations. Next,
protein samples were separated by 7.5 or 12% SDS-polyacrylamide gel
electrophoresis and transferred onto an ImmunBlot PVDF membrane
(EMD Millipore). Membranes were blocked with 5% bovine serum
albumin in Tris-phosphate buffer containing 0.05% Tween-20 (TBS-T)
for 1 h at room temperature, and further incubated overnight at 4°C
with primary antibodies specific for ephrinB2 (1:1,000; ab131536;
Abcam), phospho-ephrinB2 (1:1,000; ab119323; Abcam), EphB4
(1:1,000; sc-5536; Santa Cruz Biotechnology), phospho-EphB4 (1:500;
12720; Signalway Antibody), collagen 1 (1:500; COL1; ab6308;
Abcam), Runt-related transcription factor 2 (1:1,000; Runx2;
ab76956; Abcam), osteocalcin (1:1,000; OCN; 33-5400; thermo Fisher
Scientific, Inc.) and β-actin (1:2,000; sc-47778; Santa Cruz
Biotechnology). Following three washes with TBS-T for 5 min each,
the membranes were incubated with horseradish peroxidase-conjugated
anti-rabbit (1:1,000; 7074; Cell Signaling Technology, Inc.) or
anti-mouse (1:1,000; 7076; Cell Signaling Technology, Inc.)
secondary antibodies for 1 h at room temperature, and then washed
three times with TBS-T. Blots were visualized and digitized using
enhanced chemiluminescence (thermo Fisher Scientific, Inc.) and the
results were quantified with ImageJ v1.8.0 software (National
Institutes of Health). The experiment was repeated three times.
Statistical analysis
All numerical data are presented as the mean ±
standard deviation. One-way analysis of variance with Bonferroni's
method was performed with SPSS 24.0 software (IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Isolated cPDLSCs exhibit stem cell
characteristics
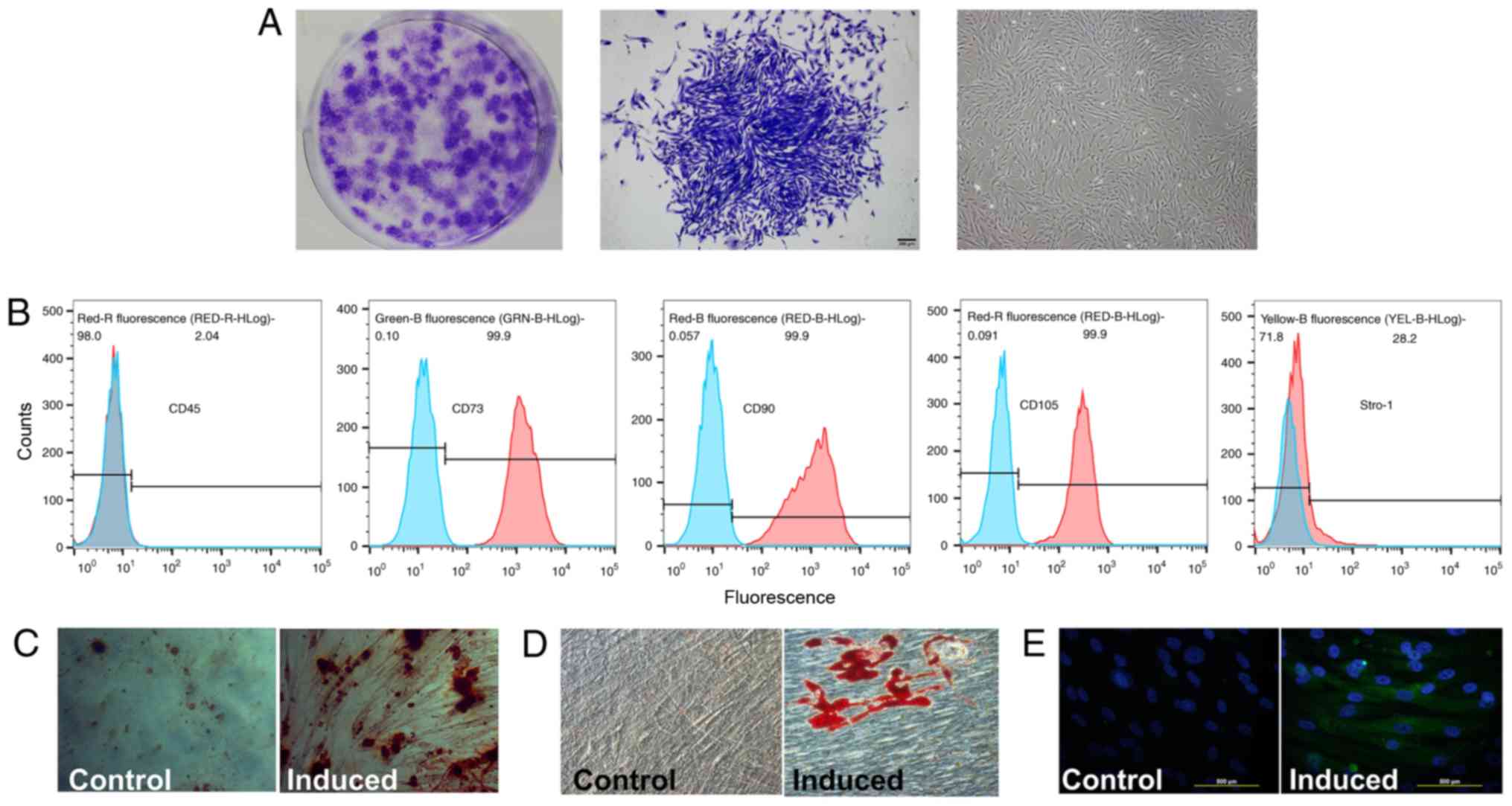
cPDLSCs displayed a fibroblast-like or
stellate-shaped morphology under an inverted microscope (Fig. 1A), while mesenchymal stem cell
markers were examined by flow cytometric analysis (Fig. 1B). STRO-1, CD90, CD105, CD73 and
CD45 were selected as the critical markers to compare the
similarities between cPDLSCs and mesenchymal stem cells (32,33). A relatively high number of cPDLSCs
expressed CD73 (99.9%), CD90 (99.9%) and CD105 (99.9%), while a low
number of cPDLSCs expressed CD45 (2.04%). STRO-1 was expressed at
more moderate levels in cPDLSCs (28.2%). Alizarin Red S and Oil Red
O staining demonstrated the presence of mineralization (Fig. 1C) and lipid droplets (Fig. 1D) following induction in
osteogenic and adipogenic media, respectively. In addition,
neurogenic induction caused morphological changes and increased the
expression of the neurogenic marker βIII-tubulin in cPDLSCs
(Fig. 1E).
EphrinB2 gene-modified cPDLSCs exhibit
increased GFP fluorescence, and mRNA and protein expression levels
of ephrinB2
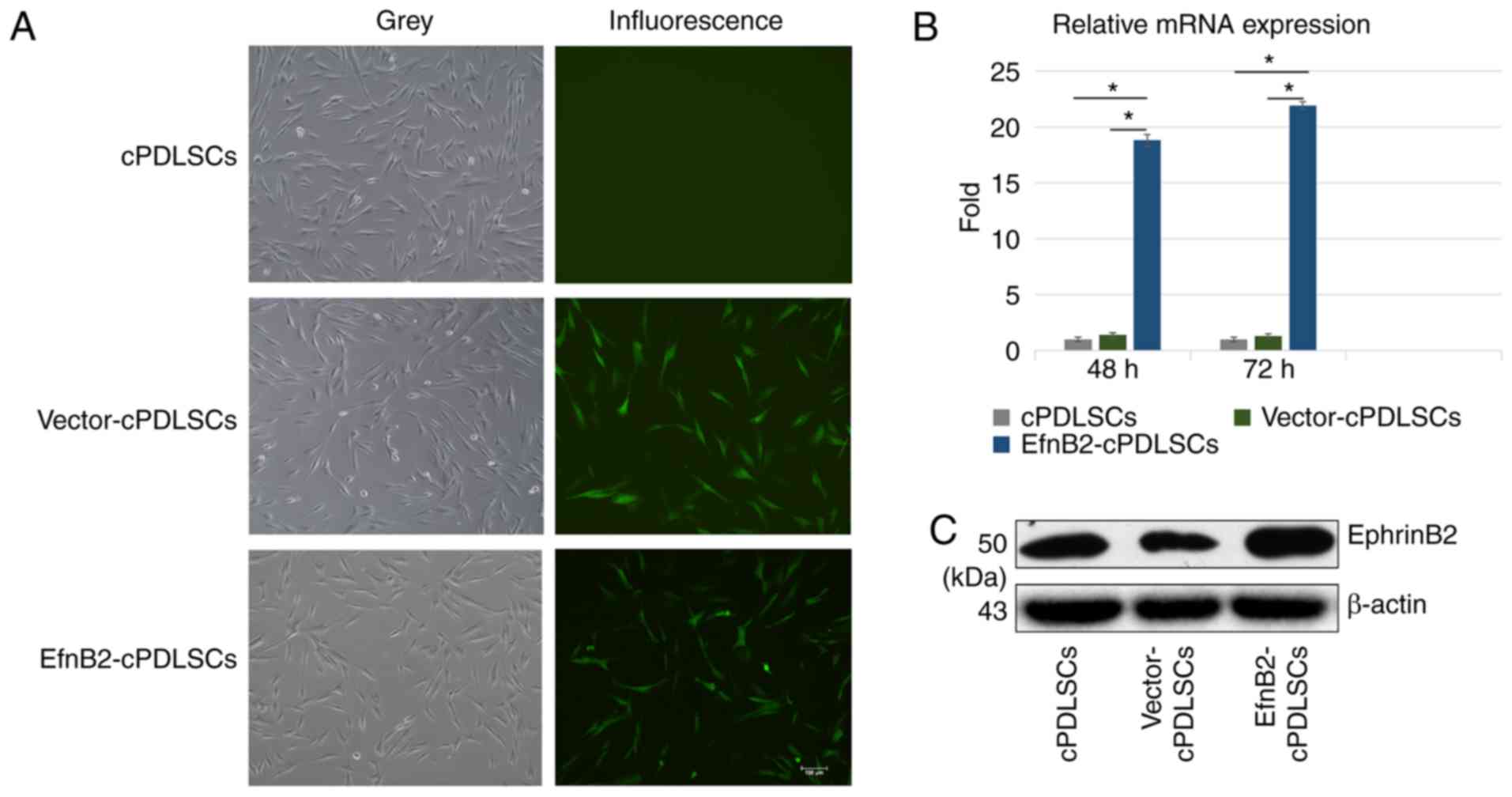
The fluorescent-inverted phase-contrast microscope
was used to detect the GFP expression in wild-type cPDLSCs,
EfnB2-cPDLSCs and Vector-cPDLSCs. Using microscopy, GFP was
detected in Vector-cPDLSCs and EfnB2-cPDLSCs, while no detectable
fluorescence signal was observed in wild-type cPDLSCs (Fig. 2A). In addition, RT-qPCR and
western blot analyses were performed to respectively detect the
mRNA and protein expression levels of ephrinB2 in wild-type
cPDLSCs, EfnB2-cPDLSCs and Vector-cPDLSCs. It was observed that the
expression of ephrinB2 mRNA was ~20-fold higher in EfnB2-cPDLSCs
compared with that in cPDLSCs and Vector-cPDLSCs. Western blot
analysis confirmed the significantly increased protein expression
levels of ephrinB2 in EfnB2-cPDLSCs compared with cPDLSCs and
Vector-cPDLSCs. These results indicated the successful upregulation
of EfnB2 gene in trans-fected cPDLSCs.
EphrinB2 gene-modified cPDLSCs exhibit
enhanced migration and proliferation
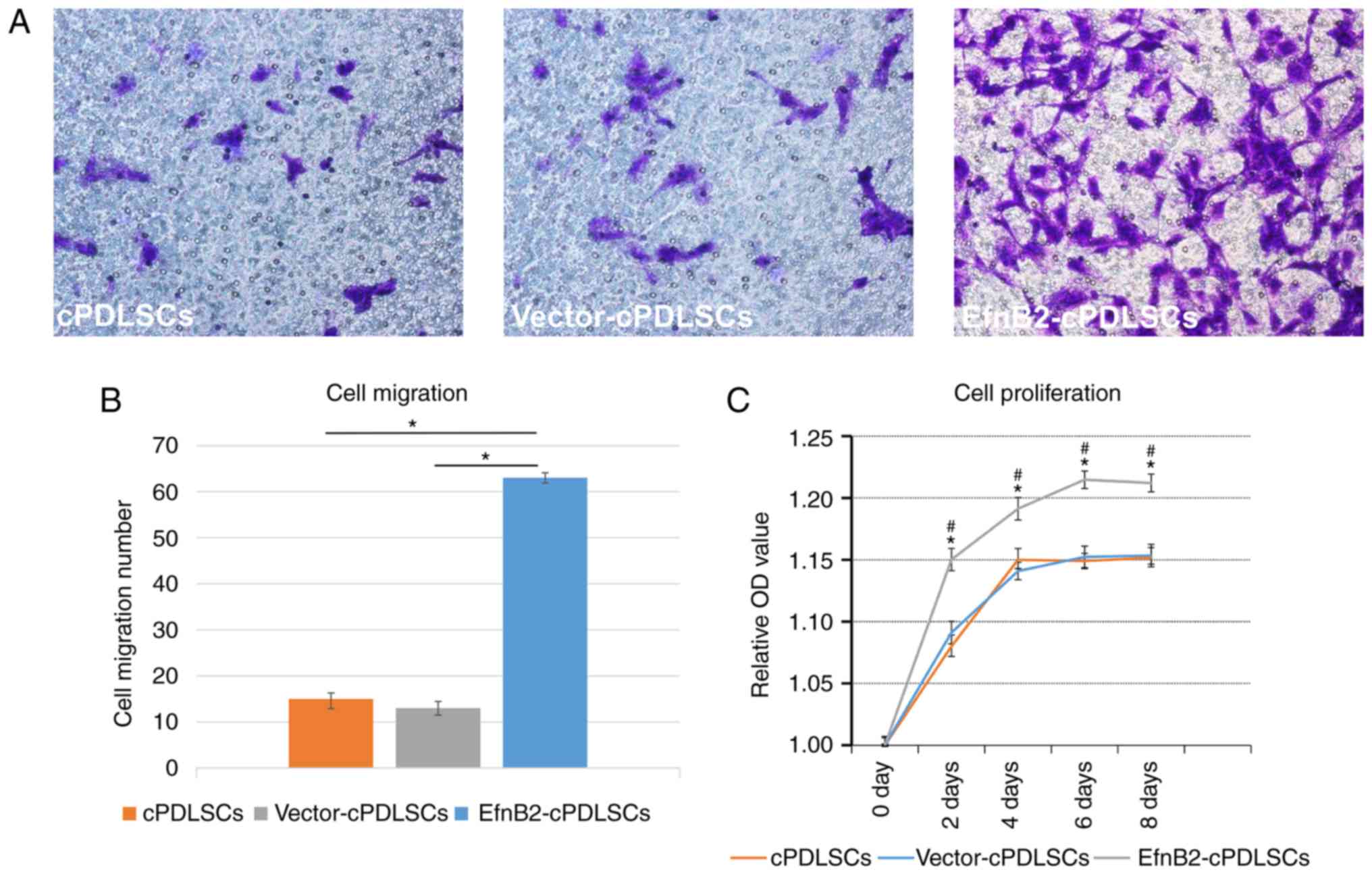
As observed in Fig.
3A, EfnB2-cPDLSCs exhibited a significantly increased migratory
ability compared with the cPDLSCs and Vector-cPDLSCs (P<0.05).
In addition, EfnB2-cPDLSCs exhibited significantly enhanced
proliferation on days 2, 4, 6 and 8 compared with the other two
groups (Fig. 3B).
EphrinB2 gene-modified cPDLSCs exhibit
enhanced osteogenic ability
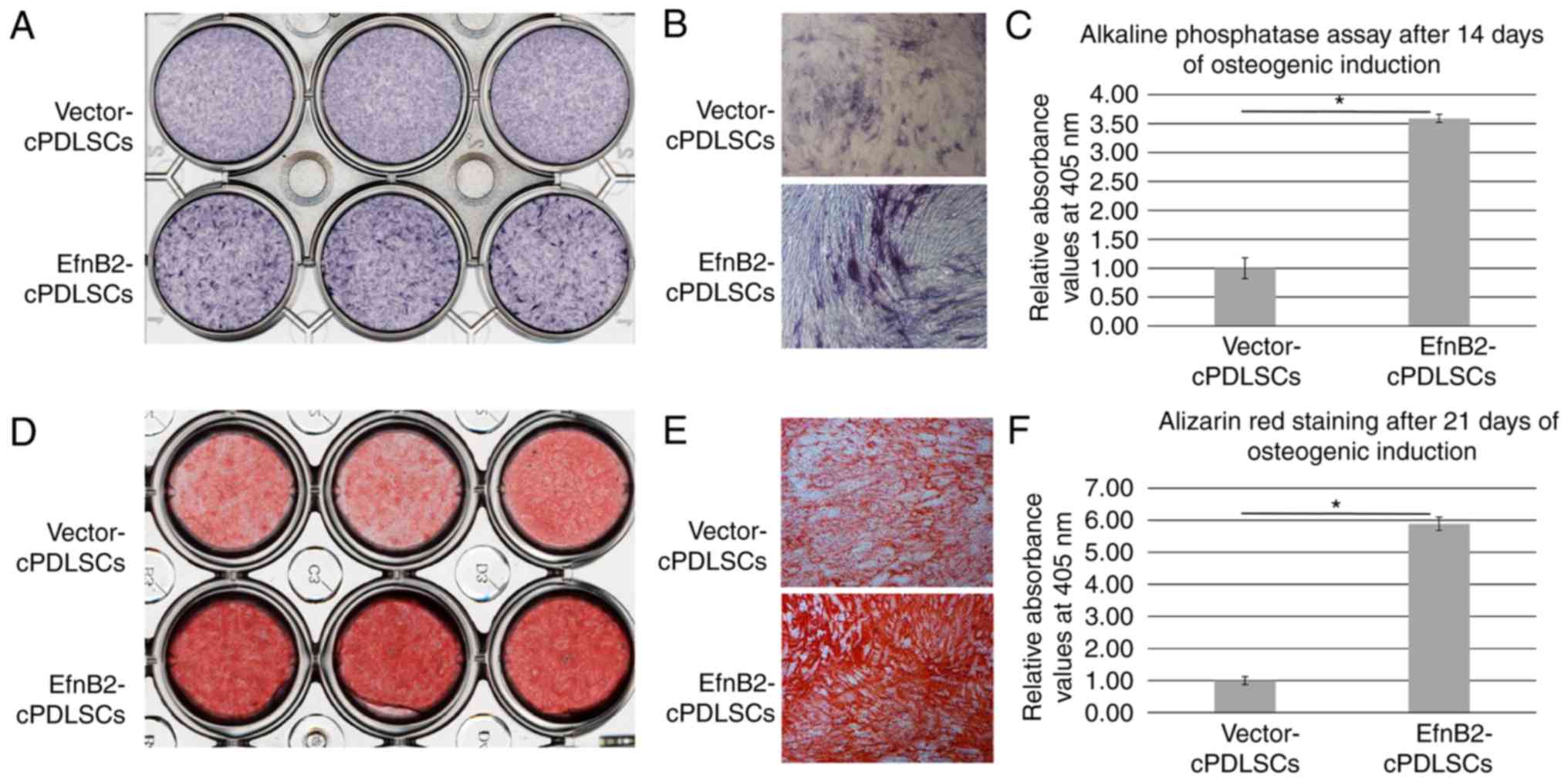
To examine the osteogenic ability of cells, an ALP
assay was performed following osteogenic induction for 14 days. As
shown in Fig. 4A-C, EfnB2-cPDLSCs
displayed an enhanced ALP activity compared with Vector-cPDLSCs. In
addition, Alizarin Red S staining performed after 21 days of
osteogenic induction revealed more prominent mineralized nodules in
EfnB2-cPDLSCs compared with those observed in Vector-cPDLSCs
(Fig. 4D-F).
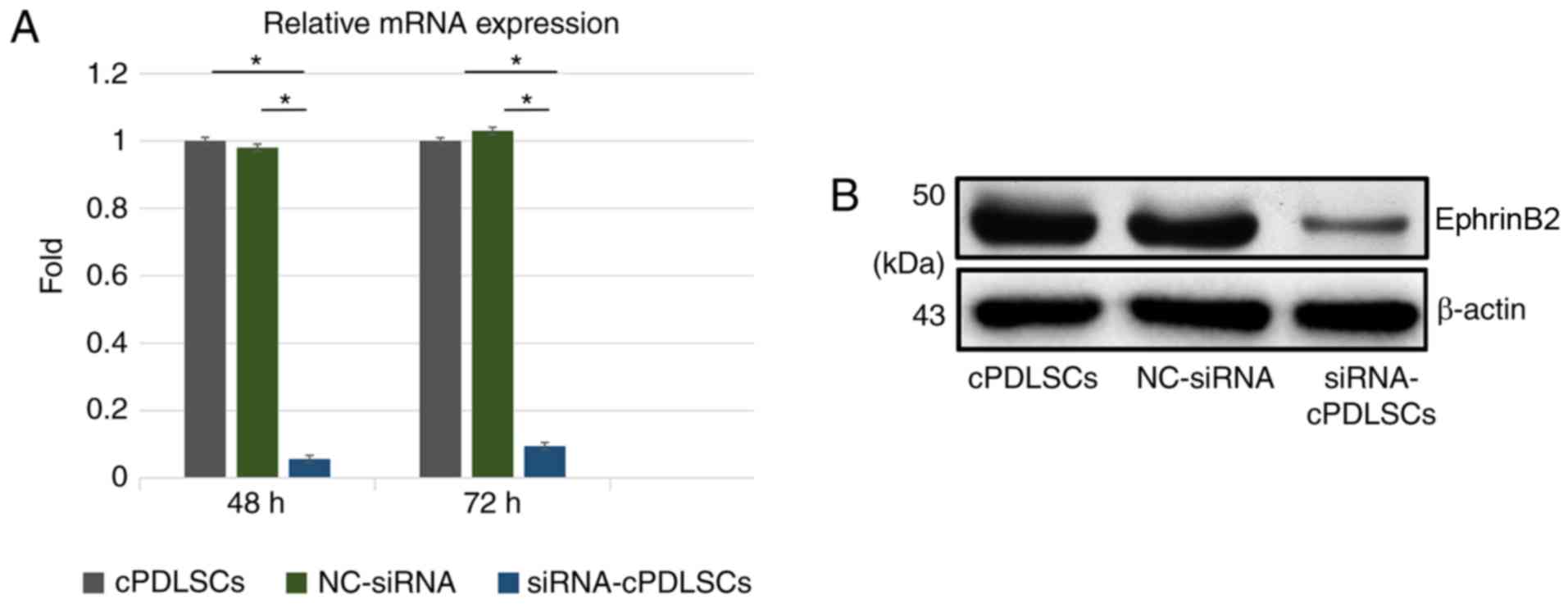
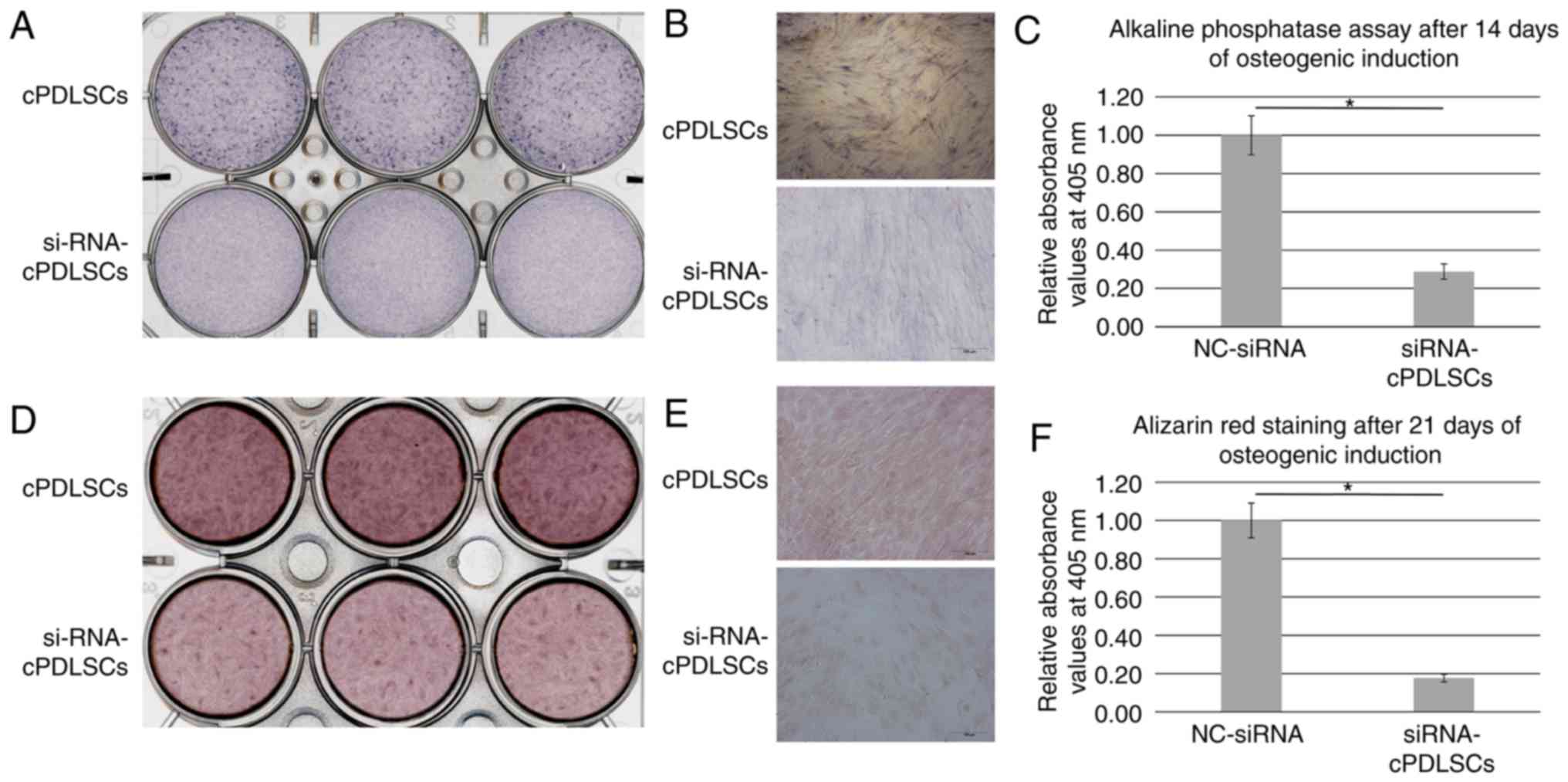
Inhibition of EphrinB2 expression
suppresses the osteogenic ability of cells
EphrinB2 expression was blocked with EphrinB2-siRNA
transfection, as indicated by the significant reduction in the mRNA
and protein levels of ephrinB2 in cPDLSCs (Fig. 5). Next, ALP staining was performed
subsequent to osteogenic induction for 14 days. As shown in
Fig. 6A-C,
EphrinB2-siRNA-transfected cPDLSCs (siRNA-cPDLSCs) displayed
decreased ALP activity as compared with the NC-siRNA-transfected
cPDLSCs. In addition, Alizarin Red S staining performed after 21
days of osteogenic induction revealed a reduction in the
mineralized nodules in siRNA-cPDLSCs as compared with those
observed in cPDLSCs transfected with NC-siRNA (Fig. 6D-F).
Effects of ephrinB2 transfection on the
mRNA expression levels of osteogenic markers in cPDLSCs following
osteogenic/odontogenic induction
As shown in Fig.
7, the mRNA expression levels of ALP, bone morphogenetic
protein 2 (BMP2), COL1 and Runx2 were markedly increased in
EfnB2-cPDLSCs compared with those in Vector-cPDLSCs on days 7 and
14 of osteogenic/odontogenic induction (Fig. 7A-D). However, no significant
differences were observed on day 21. Furthermore, the mRNA
expression levels of bone sialoprotein (BSP), dentin matrix
protein-1 (DMP1) and dentin sialophosphoprotein (DSPP) were
significantly higher in EfnB2-cPDLSCs after 14 days of induction,
whereas no significant differences were observed on days 7 or 21
(Fig. 7E-G). For OCN mRNA
expression, no significant differences were observed between
EfnB2-cPDLSCs and Vector-cPDLSCs on days 7 and 14, whereas a
significantly higher gene expression was observed on day 21 of
osteogenic induction (Fig.
7H).
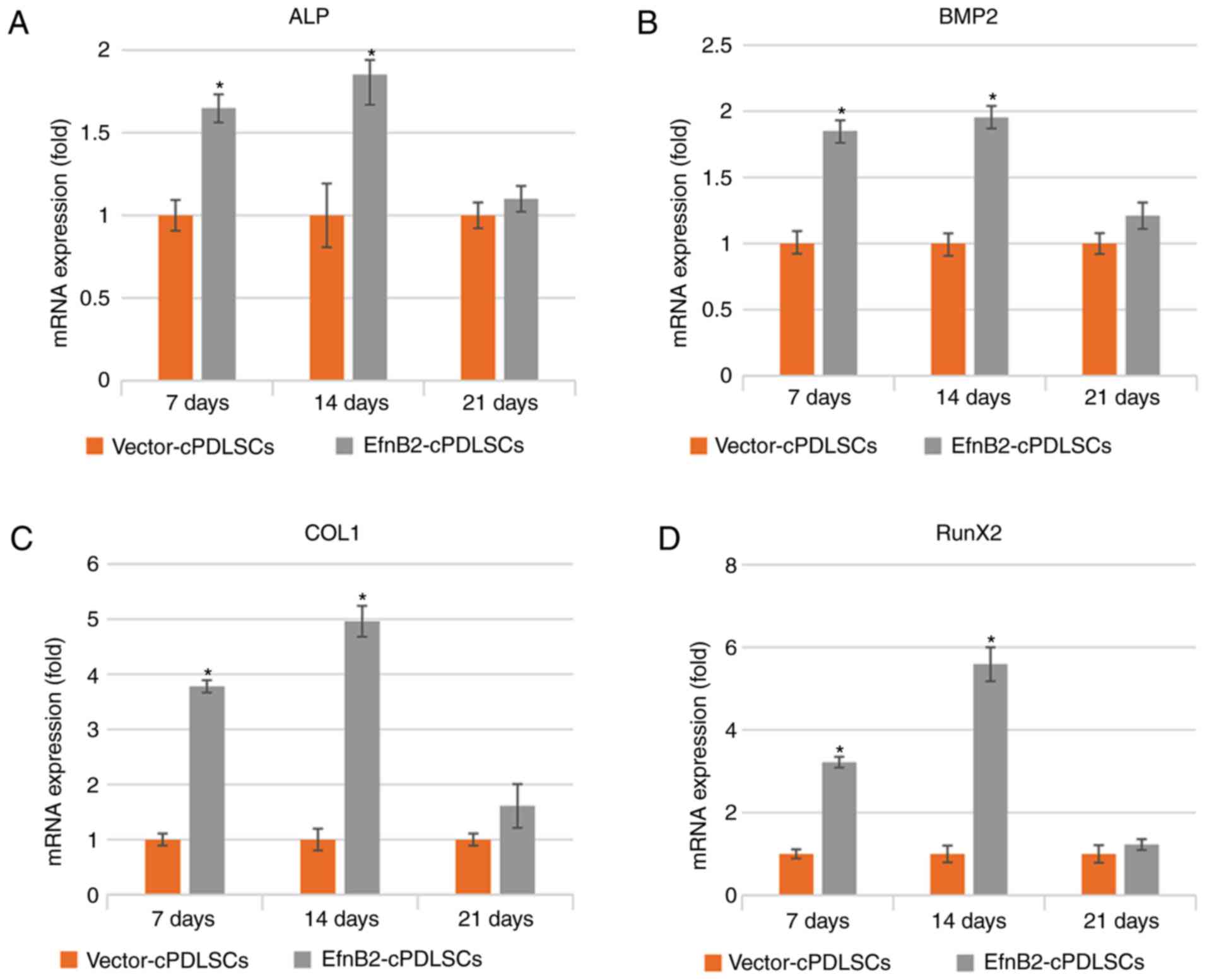 | Figure 7Reverse transcription-quantitative
polymerase chain reaction analysis of mRNA expression levels of (A)
ALP, (B) BMP2, (C) COL1, (D) Runx2, (E) BSP, (F) DMP1 (G) DSPP and
(H) OCN in Vector-cPDLSCs and EfnB2-cPDLSCs after 7, 14 and 21 days
of osteogenic/odontogenic induction. *P<0.05 vs.
Vector-cPDLSCs. cPDLSCs, canine periodontal ligament stem cells;
ALP, alkaline phosphatase; BMP2, bone morphogenetic protein 2;
COL1, collagen 1; Runx2, Runt-related transcription factor 2; BSP,
bone sialoprotein; DMP1, dentin matrix protein-1; DSPP, dentin
sialophosphoprotein; OCN, osteocalcin. |
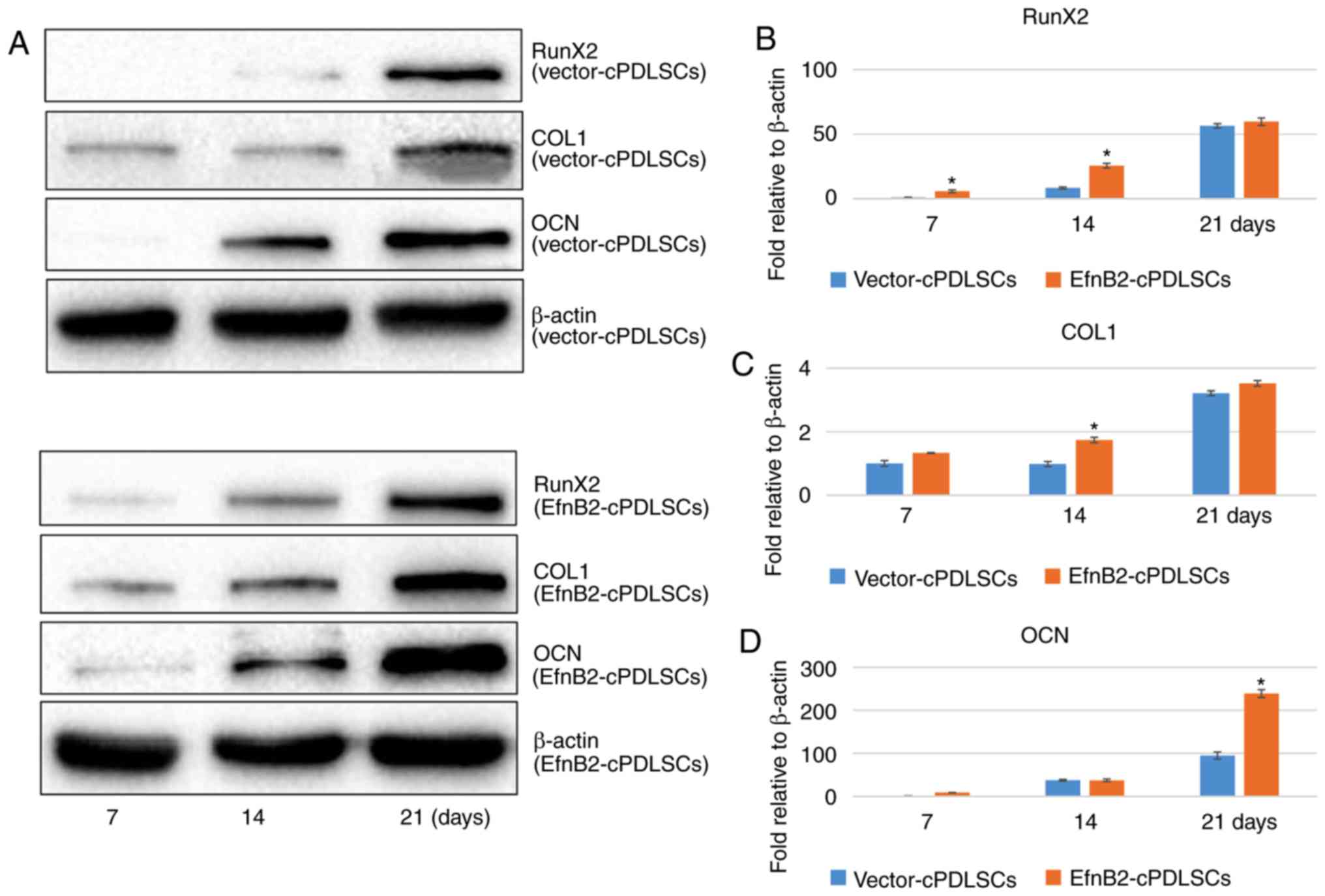
Protein expression levels of osteogenic
markers in cPDLSCs following osteogenic/odontogenic induction
As shown in Fig.
8, the protein expression levels of Runx2 were increased in
EfnB2-cPDLSCs compared with Vector-cPDLSCs on days 7 and 14 after
osteogenic/odontogenic induction; however, no significant
differences were observed on day 21. In addition, the protein
expression of OCN was significantly upregulated in EfnB2-cPDLSCs
compared with Vector-cPDLSCs on day 21, while no significant
differences were observed on days 7 and 14. Notably, the protein
expression levels of COL1 were increased in EfnB2-cPDLSCs compared
with Vector-cPDLSCs on day 14; however, no significant difference
were observed on days 7 and 21.
Endogenous levels of phosphorylated and
unphosphorylated ephrinB2 and EphB4 proteins during
osteogenic/odontogenic differentiation of EfnB2-cPDLSCs and
Vector-cPDLSCs
The western blot results revealed that maximal
levels of phospho-ephrinB2 occurred 30 min after stimulation of
EfnB2-cPDLSCs and Vector-cPDLSCs with EphB4-FC (Fig. 9A and B). The protein expression
levels of phospho-ephrinB2 remained increased in EfnB2-cPDLSCs
after 12 h of osteogenic/odontogenic induction and reached a
maximum at 24 h (Fig. 9C-E).
Although phospho-ephrinB2 was detected at a low expression level in
Vector-cPDLSCs at 12 h, and reached a maximum at 24 h, the
expression levels were markedly lower compared with EfnB2-cPDLSCs.
Similarly, there was a significant increase of phospho-EphB4 in
EfnB2-cPDLSCs compared with Vector-cPDLSCs at 48 h (Fig. 9C-E).
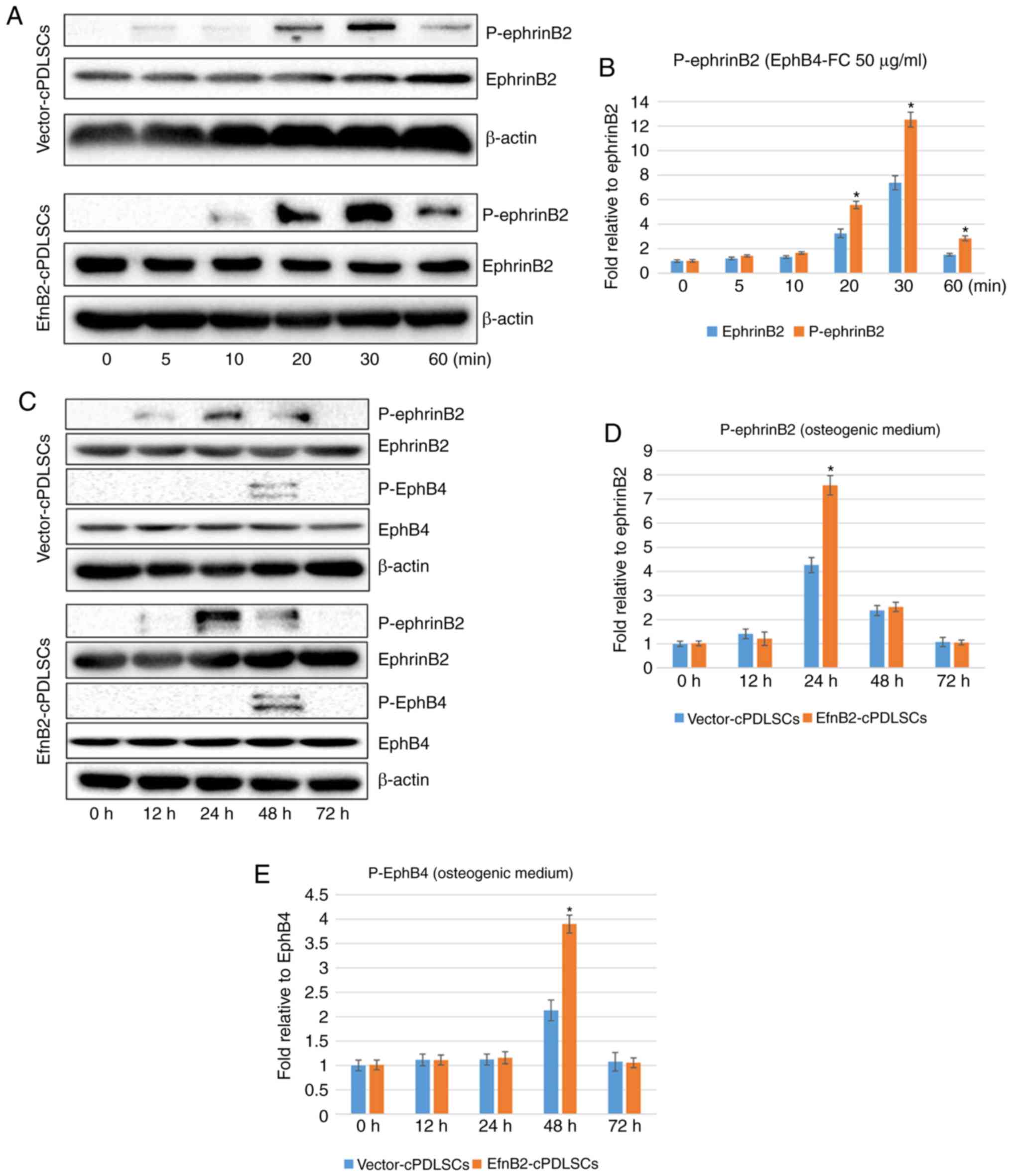 | Figure 9Endogenous expression of EphrinB2,
EphB4 and their phosphorylated proteins during osteogenic
differentiation of cPDLSCs. (A) Western blots of EphrinB2 and
P-ephrinB2 in Vector-cPDLSCs and EfnB2-cPDLSCs stimulated with
EphB4-FC for 0, 5, 10, 20, 30 and 60 min. Quantification of protein
expression bands of (B) P-ephrinB2 stimulated with EphB4-FC for 0,
5, 10, 20, 30 and 60 min. (C) Western blots of EphrinB2,
P-ephrinB2, EphB4 and P-EphB4 in Vector-cPDLSCs and EfnB2-cPDLSCs
following osteogenic induction for 0, 12, 24, 48 and 72 h.
Quantification of protein expression bands of (D) P-ephrinB2 and
(E) P-EphB4 following osteogenic induction for 0, 12, 24, 48 and 72
h. *P<0.05 vs. Vector-cPDLSCs. cPDLSCs, canine
periodontal ligament stem cells; P-, phosphorylated. |
Discussion
Currently, the selection of a carrier scaffold,
signaling molecules and seed cells is one of the key issues in
periodontal tissue engineering. A previous study has reported that
ephrinB2 ligand and its receptor EphB4 constitute a bidirectional
signaling pathway with a crucial role in bone remodeling (8). It has been indicated that the
ephrinB2-mediated reverse signaling pathway inhibits osteoclast
function and bone resorption by reducing the activity of c-Fos and
NFATc1, whereas the forward signaling pathway promotes bone
formation by reducing RhoA activity and enhancing osteoblast
differentiation (9).
Numerous studies have attempted to use ephrinB2 as a
molecular therapy for tissue engineering. Indeed, in animal
experiments, it has been demonstrated that the degree of bone
mineralization, bone hardness and osteoblast differentiation in
ephrinB2-deficient mice was significantly reduced compared with
those in wild-type mice (34).
Our previous study revealed that that ALP activity of PDLSCs was
significantly increased following transfection with ephrinB2 in
vitro, which resulted in more calcified nodules and increased
expression levels of osteogenesis-associated genes (BSP, COL1,
Runx2, DSPP and OCN), suggesting that upregulation of ephrinB2
promoted the osteogenic differentiation of PDLSCs (13). Collectively, these previous
results suggested that ephrinB2 is a potentially useful biomolecule
for tissue engineering. However, although in vitro and in
vivo studies have been performed, the effect of bidirectional
ephrinB2-EphB4 signaling has not been examined in cPDLSCs, and thus
this was the focus of the present study.
The present study first examined the stem cell-like
characteristics of isolated cPDLSCs, including self-renewal
capacity, multilineage differentiation ability and expression
levels of stem cell surface markers. It was observed that isolated
cPDLSCs displayed a fibroblast-like or stellate-shaped morphology,
and grew in colonies. A characteristic set of mesenchymal stem
cells markers, including STRO-1, CD90, CD105, CD73 and CD45, were
selected to perform a flow cytometric assay (32,33). A percentage of cPDLSCs as high as
28.2% expressed STRO-1, which is considered to be a mesenchymal
stem cell surface marker (35).
In addition, Alizarin Red S staining for mineralized nodules, Oil
Red O staining for lipid droplets and immunofluorescence staining
for βIII-tubulin were also performed following induction of
osteogenesis, adipogenesis or neurogenesis using the appropriate
media. The findings suggested that cPDLSCs may be a favorable
candidate for dental tissue engineering.
Next, transfection of cPDLSCs with ephrinB2 was
conducted in the present study, and the fluorescent protein
expression, as well as the mRNA and protein expression levels of
ephrinB2 were examined. The results demonstrated continuous and
stable expression of ephrinB2 mRNA and protein in EfnB2-cPDLSCs,
thus verifying the successful transfection of ephrinB2 into
cPDLSCs. In addition, to evaluate the proliferative and migratory
abilities of cells following ephrinB2 transfection, Transwell and
CCK-8 assays were performed, as previously described (13). EfnB2-cPDLSCs exhibited enhanced
migration and proliferation compared with cPDLSCs and
Vector-cPDLSCs. These results are consistent with previous research
demonstrating that ephrinB2 reverse signaling promotes cell
migration (36) and mediates cell
proliferation (37).
To further investigate the effect of ephrinB2 gene
transfection on osteogenic differentiation of cPDLSCs, ALP and
Alizarin Red S staining assays were also performed in the current
study. As expected, when cells were cultured under osteoinductive
conditions, EfnB2-cPDLSCs displayed enhanced ALP activity and
formed more mineralized nodules as compared with Vector-cPDLSCs. In
addition, the expression of middle and late osteogenic genes and
proteins was significantly increased, suggesting that ephrinB2
signaling was associated with late bone metabolism (38). Notably, in contrast to our
previous study (39), in which
EphB4-FC was used to stimulate human PDLSCs and no significant
differences were observed in the expression levels of early
osteogenic genes in the EphB4-FC-stimulated groups, there were
significant differences in the gene expression levels of early
osteogenic genes in EfnB2-cPDLSCs in the present study. Thus, it is
speculated that ephrinB2 reverse signaling is associated not only
with bone metabolism, but also with early bone formation (34). Furthermore, EphB4-FC stimulation
can only establish ephemeral and unsustainable signaling, while
gene transfection of ephrinB2 is more intense and sustainable
(40).
Previous research reported that activation of the
ephrinB2-EphB4 signaling pathway promoted early osteogenic
differentiation, leading to earlier osteogenic process and earlier
expression of Runx2 (41);
However, other research reported that high levels of ephrinB2
overexpression increases the osteogenic differentiation of human
mesenchymal stem cells, while no significant changes in Runx2
expression were found when they attempted to elucidate the
molecular mechanisms of ephrinB2 overexpression (42). This suggests that the anaphase
effect of cell-mediated mineralization is associated with other
signaling pathways.
To further evaluate the effect and mechanisms of
ephrinB2 and its receptor EphB4, western blot assays were conducted
in the present study. Following stimulation with EphB4-FC,
phospho-ephrinB2 expression was detectable. Notably, the
phospho-ephrinB2 protein expression levels in EfnB2-cPDLSCs were
significantly higher than that in Vector-cPDLSCs. Following
osteogenic/odontogenic induction, expression of phospho-ephrinB2
and phospho-EphB4 proteins was detected in EfnB2-cPDLSCs and
Vector-cPDLSCs, demonstrating that reverse and forward signaling
was induced. These results are consistent with the findings of
previous studies, suggesting that ephrinB2 signaling can be
activated, and that forward and reverse signaling can be induced in
cPDLSCs (43,44).
In conclusion, the findings of the present study
indicated that ephrinB2 gene-modified cPDLSCs exhibited enhanced
osteogenic differentiation, and that the ephrinB2 reverse signaling
and EphB4 forward signaling pathways served a key role in this
process. In addition, ephrinB2 gene modification promoted cell
migration and proliferation. Based on these results, it is
speculated that, since ephrinB2 is a transmembrane protein, it can
only transmit signals through direct contact between cells; thus,
when it is used for endogenous periodontal tissue regeneration,
ephrinB2-expressing cells can only selectively come into direct
contact with EphB4-expressing cells. Although this limits the
therapeutic effect to the defect area, as a result, there are no
side effects caused by diffusion of secretory cytokines following
transplantation. Furthermore, it is proposed that the migratory
ability of endogenous stem cells following transfection is
enhanced, and thus these cells become more conducive to migration
toward the tissue defect area, which is a key factor for tissue
repair.
Funding
This study was supported by the Seed Funding of
Xuzhou Medical University (grant no. 2018KJ22), the Xuzhou Science
and Technology Project (grant no. KC19137) and the National Natural
Science Foundation Youth Science Foundation Project (grant no.
81700954).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PW and MG conceived and designed the study. SZ, ZL
and CY performed the experiments. YL, YY and HW analyzed the data.
SZ and CZ were responsible for interpreting the data and writing
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Ethical approval for the present study was obtained
from the Institutional Animal Care and Use Committee of Xuzhou
Medical University (Xuzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Pihlstrom BL, Michalowicz BS and Johnson
NW: Periodontal diseases. Lancet. 366:1809–1820. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Taba M Jr, Kinney J, Kim AS and Giannobile
WV: Diagnostic biomarkers for oral and periodontal diseases. Dent
Clin North Am. 49:551–571. vi2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Darveau RP: Periodontitis: A polymicrobial
disruption of host homeostasis. Nat Rev Microbiol. 8:481–490. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heitz-Mayfield LJ and Lang NP: Surgical
and nonsurgical periodontal therapy. Learned and unlearned
concepts. Periodontol 2000. 62:218–231. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pellegrini G, Pagni G and Rasperini G:
Surgical approaches based on biological objectives: GTR versus GBR
techniques. Int J Dent. 2013:5215472013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu X, Li X, Wang J, He XT, Sun HH and Chen
FM: Concise review: Periodontal tissue regeneration using stem
cells: Strategies and translational considerations. Stem Cells
Transl Med. 8:392–403. 2019. View Article : Google Scholar :
|
|
7
|
Venkataiah VS, Handa K, Njuguna MM,
Hasegawa T, Maruyama K, Nemoto E, Yamada S, Sugawara S, Lu L,
Takedachi M, et al: Periodontal regeneration by allogeneic
transplantation of adipose tissue derived multi-lineage progenitor
stem cells in vivo. Sci Rep. 9:9212019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao C, Irie N, Takada Y, Shimoda K,
Miyamoto T, Nishiwaki T, Suda T and Matsuo K: Bidirectional
ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab.
4:111–121. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pasquale EB: Eph-ephrin bidirectional
signaling in physiology and disease. Cell. 133:38–52. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bochenek ML, Dickinson S, Astin JW, Adams
RH and Nobes CD: Ephrin-B2 regulates endothelial cell morphology
and motility independently of Eph-receptor binding. J Cell Sci.
123:1235–1246. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pennisi A, Ling W, Li X, Khan S,
Shaughnessy JD Jr, Barlogie B and Yaccoby S: The ephrinB2/EphB4
axis is dysregulated in osteoprogenitors from myeloma patients and
its activation affects myeloma bone disease and tumor growth.
Blood. 114:1803–1812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Füller T, Korff T, Kilian A, Dandekar G
and Augustin HG: Forward EphB4 signaling in endothelial cells
controls cellular repulsion and segregation from ephrinB2 positive
cells. J Cell Sci. 116:2461–2470. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu SY, Wang PL, Liao CS, Yang YQ, Yuan
CY, Wang S, Dissanayaka WL, Heng BC and Zhang CF: transgenic
expression of ephrinB2 in periodontal ligament stem cells (PDLSCs)
modulates osteogenic differentiation via signaling crosstalk
between ephrinB2 and EphB4 in PDLSCs and between PDLSCs and
pre-osteoblasts within co-culture. J Periodontal Res. 52:562–573.
2017. View Article : Google Scholar
|
|
14
|
Yuan C, Wang P, Zhu S, Zou T, Wang S, Xu
J, Heng BC, Diogenes A and Zhang C: EphrinB2 stabilizes
vascularlike structures generated by endothelial cells and stem
cells from apical papilla. J Endod. 42:1362–1370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gong T, Heng BC, Xu J, Zhu S, Yuan C, Lo
EC and Zhang C: Decellularized extracellular matrix of human
umbilical vein endothelial cells promotes endothelial
differentiation of stem cells from exfoliated deciduous teeth. J
Biomed Mater Res A. 105:1083–1093. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gronthos S, Mankani M, Brahim J, Robey PG
and Shi S: Postnatal human dental pulp stem cells (DPSCs) in vitro
and in vivo. Proc Natl Acad Sci USA. 97:13625–13630. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miura M, Gronthos S, Zhao M, Lu B, Fisher
LW, Robey PG and Shi S: SHED: Stem cells from human exfoliated
deciduous teeth. Proc Natl Acad Sci USA. 100:5807–5812. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang GT, Sonoyama W, Liu Y, Liu H, Wang S
and Shi S: The hidden treasure in apical papilla: The potential
role in pulp/dentin regeneration and bioroot engineering. J Endod.
34:645–651. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Nor F, Oh M, Cucco C, Shi S and
Nör JE: Wnt/β-catenin signaling determines the vasculogenic fate of
postnatal mesenchymal stem cells. Stem Cells. 34:1576–1587. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bento LW, Zhang Z, Imai A, Nör F, Dong Z,
Shi S, Araujo FB and Nör JE: Endothelial differentiation of SHED
requires MEK1/ERK signaling. J Dent Res. 92:51–57. 2013. View Article : Google Scholar :
|
|
21
|
Dissanayaka WL, Zhu L, Hargreaves KM, Jin
L and Zhang C: Scaffold-free prevascularized microtissue spheroids
for pulp regeneration. J Dent Res. 93:1296–1303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cordeiro MM, Dong Z, Kaneko T, Zhang Z,
Miyazawa M, Shi S, Smith AJ and Nör JE: Dental pulp tissue
engineering with stem cells from exfoliated deciduous teeth. J
Endod. 34:962–969. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang N, Chen B, Wang W, Chen C, Kang J,
Deng SQ, Zhang B, Liu S and Han F: Isolation, characterization and
multi-lineage differentiation of stem cells from human exfoliated
deciduous teeth. Mol Med Rep. 14:95–102. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qazi TH, Mooney DJ, Pumberger M, Geissler
S and Duda GN: Biomaterials based strategies for skeletal muscle
tissue engineering: Existing technologies and future trends.
Biomaterials. 53:502–521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akizuki T, Oda S, Komaki M, Tsuchioka H,
Kawakatsu N, Kikuchi A, Yamato M, Okano T and Ishikawa I:
Application of periodontal ligament cell sheet for periodontal
regeneration: A pilot study in beagle dogs. J Periodontal Res.
40:245–251. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iwata T, Yamato M, Tsuchioka H, Takagi R,
Mukobata S, Washio K, Okano T and Ishikawa I: Periodontal
regeneration with multi-layered periodontal ligament-derived cell
sheets in a canine model. Biomaterials. 30:2716–2723. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsumanuma Y, Iwata T, Washio K, Yoshida T,
Yamada A, Takagi R, Ohno T, Lin K, Yamato M, Ishikawa I, et al:
Comparison of different tissue-derived stem cell sheets for
periodontal regeneration in a canine 1-wall defect model.
Biomaterials. 32:5819–5825. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bugueño J, Li W, Salat P, Qin L and
Akintoye SO: The bone regenerative capacity of canine mesenchymal
stem cells is regulated by site-specific multilineage
differentiation. Oral Surg Oral Med Oral Pathol Oral Radiol.
123:163–172. 2017. View Article : Google Scholar
|
|
29
|
Dissanayaka WL, Zhu X, Zhang C and Jin L:
Characterization of dental pulp stem cells isolated from canine
premolars. J Endod. 37:1074–1080. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Albini A and Benelli R: The chemoinvasion
assay: A method to assess tumor and endothelial cell invasion and
its modulation. Nat Protoc. 2:504–511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
32
|
Hermida-Gómez T, Fuentes-Boquete I,
Gimeno-Longas MJ, Muiños-López E, Díaz-Prado S, de Toro FJ and
Blanco FJ: Quantification of cells expressing mesenchymal stem cell
markers in healthy and osteoarthritic synovial membranes. J
Rheumatol. 38:339–349. 2011. View Article : Google Scholar
|
|
33
|
Greco SJ, Liu K and Rameshwar P:
Functional similarities among genes regulated by OCT4 in human
mesenchymal and embryonic stem cells. Stem Cells. 25:3143–3154.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tonna S, Takyar FM, Vrahnas C,
Crimeen-Irwin B, Ho PW, Poulton IJ, Brennan HJ, McGregor NE, Allan
EH, Nguyen H, et al: EphrinB2 signaling in osteoblasts promotes
bone mineralization by preventing apoptosis. FASEB J. 28:4482–4496.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ning H, Lin G, Lue TF and Lin CS:
Mesenchymal stem cell marker Stro-1 is a 75 kd endothelial antigen.
Biochem Biophys Res Commun. 413:353–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakada M, Anderson EM, Demuth T, Nakada S,
Reavie LB, Drake KL, Hoelzinger DB and Berens ME: The
phosphorylation of ephrin-B2 ligand promotes glioma cell migration
and invasion. Int J Cancer. 126:1155–1165. 2010.
|
|
37
|
Steinle JJ, Meininger CJ, Chowdhury U, Wu
G and Granger HJ: Role of ephrin B2 in human retinal endothelial
cell proliferation and migration. Cell Signal. 15:1011–1017. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Edwards CM and Mundy GR: Eph receptors and
ephrin signaling pathways: A role in bone homeostasis. Int J Med
Sci. 5:263–272. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Heng BC, Wang S, Gong T, Xu J, Yuan C and
Zhang C: EphrinB2 signaling enhances osteogenic/odontogenic
differentiation of human dental pulp stem cells. Arch Oral Biol.
87:62–71. 2018. View Article : Google Scholar
|
|
40
|
Ikeda Y, Sun Z, Ru X, Vandenberghe LH and
Humphreys BD: Efficient gene transfer to kidney mesenchymal cells
using a synthetic adeno-associated viral Vector. J Am Soc Nephrol.
29:2287–2297. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang L, Zhang J, Wang C, Qi Y, Du M, Liu
W, Yang C and Yang P: Low concentrations of TNF-α promote
osteogenic differentiation via activation of the ephrinB2-EphB4
signalling pathway. Cell Prolif. 50:2017. View Article : Google Scholar
|
|
42
|
Tierney EG, Mcsorley K, Hastings CL, Cryan
SA, O'Brien T, Murphy MJ, Barry FP, O'Brien FJ and Duffy GP: High
levels of ephrinB2 over-expression increases the osteogenic
differentiation of human mesenchymal stem cells and promotes
enhanced cell mediated mineralisation in a
polyethyleneimine-ephrinB2 gene-activated matrix. J Control
Release. 165:173–182. 2013. View Article : Google Scholar
|
|
43
|
Li C, Shi C, Kim J, Chen Y, Ni S, Jiang L,
Zheng C, Li D, Hou J, Taichman RS and Sun H: Erythropoietin
promotes bone formation through EphrinB2/EphB4 signaling. J Dent
Res. 94:455–463. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Matsuo K and Otaki N: Bone cell
interactions through Eph/ephrin: Bone modeling, remodeling and
associated diseases. Cell Adh Migr. 6:148–156. 2012. View Article : Google Scholar : PubMed/NCBI
|