Introduction
Osteosarcoma is the most common primary malignant
bone tumor in clinical practice; these tumors exhibit strong local
invasiveness, can easily metastasize at an early stage, and have a
poor prognosis (1-3). Osteosarcoma occurs mainly in
children and adolescents, and the 5-year survival rate is 65-75%
(1). Recent advances in surgery,
radiotherapy and neoadjuvant chemotherapy have greatly improved the
prognosis of osteosarcoma, but recurrence, metastasis, and drug
resistance still result in a poor prognosis. In addition, surgical
resection severely impairs the limb function of the patients, and
radiotherapy and chemotherapy are limited due to their severe
systemic toxicity (2). Therefore,
the development of new anti-osteosarcoma treatments with fewer
complications and side effects is urgently required (3).
Photodynamic therapy (PDT) is a new method for tumor
treatment that has the advantages of high selectivity, limited
damage, low toxicity, few side effects and reproducibility
(4,5). In addition, its curative effect is
improved when combined with surgery (6). PDT has been used for various
malignant diseases, including breast, gastric and skin cancer
(7-10). In PDT, a photosensitizer is
infused intravenously or injected locally into the body to ensure
that the photosensitizer is relatively enriched specifically in the
tumor tissue; then, the tumor tissue is locally irradiated with
light of a corresponding wavelength. When the photosensitizer is
excited by the light, photochemical and photobiological reactions
in the surrounding environment produce large amounts of reactive
oxygen species (ROS), which destroy tumor cells and tissues,
leading to tumor cell death (11-13). Early PDT methods used mainly
hematoporphyrin derivatives as photosensitizers. These
photosensitizers are slowly metabolized by the body, which leads to
long periods during which light must be avoided after treatment;
additionally, their composition is complex, the targeting is weak
and their skin phototoxicity is strong (14-16). Pyropheophorbide-α methyl ester
(MPPa) belongs to the second generation of photosensitizers; this
novel type of photosensitizer exhibits strong photosensitivity, a
clear chemical structure, a single component and a stable nature,
and it is metabolized rapidly by the body (17-19). MPPa-PDT has been studied in
multiple types of cancer such as breast, lung and prostate cancer
(20-22). de Miguel et al (23) have reported that photodynamic
therapy is a potential antitumoral treatment for surgically
inoperable osteosarcoma. However, further study of its mechanism
for treating osteosarcoma is required.
The present study aimed to explore the effects of
MPPa-PDT on the cell cycle, migration and invasion of human
osteosarcoma MG-63 cells. MPPa-PDT-induced apoptosis in
osteosarcoma MG-63 cells and the related mechanisms were examined
to provide an experimental basis for the clinical treatment of
osteosarcoma.
Materials and methods
Cell lines and reagents
The MG-63 cell line was purchased from. The human
fibroblast HFL-1 cell line was donated by Professor Zhou Jing,
Department of Respiratory, the First Affiliated Hospital of
Chongqing Medical University (Chongqing, China). MPPa and rapamycin
(RAPA) were purchased from Sigma-Aldrich; Merck KGaA. Dulbecco's
modified Eagle's medium (DMEM) and Matrigel were obtained from BD
Biosciences. Fetal bovine serum (FBS) and trypsin were purchased
from Gibco; Thermo Fisher Scientific, Inc. FLUO-3/AM was purchased
from Dojindo Molecular Technologies, Inc. An Annexin V-propidium
iodide (PI) double-staining test kit was purchased from Nanjing
Keygen Biotech Co., Ltd. Cyclin D1, Cyclin E, Cyclin A, Cyclin B1,
E-cadherin (E-cad), MMP-2, MMP-9, Akt, phosphorylated (p)-Akt,
mTOR, p-mTOR, 4EBP1, eukaryotic translation initiation factor
4E-binding protein 1 (4EBP1), p-4EBP1, P70S6K, p-P70S6K, Bip,
serine/threonine-protein kinase/endoribonuclease IRE1 (IRE1α),
eukaryotic translation initiation factor 2α kinase 3 (PERK),
protein disulfide isomerase (PDI), C/EBP-homologous protein 10
(CHOP), cleaved caspase-3, cleaved poly (ADP-ribose) polymerase 1
(PARP), microtubule-associated protein 1 light chain 3α (LC-3), P62
and β-actin antibodies were purchased from Cell Signaling
Technology, Inc. A cleaved caspase-12 antibody was purchased from
Wuhan Sanying Biotechnology, and a p-PERK antibody was supplied by
Santa Cruz Biotechnology. The PDT equipment was purchased from
Chongqing Jingyu Laser Technology Co. Ltd.
Experimental grouping and processing
The experiment comprised six groups as follows: i)
Control, untreated cells; ii) MPPa, cells treated with MPPa alone;
iii) LED, cells treated with a light-emitting diode (LED) alone;
iv) RAPA, cell treated with RAPA alone; v) MPPa-PDT, cell treated
with MPPa and light; and vi) MPPa-PDT-RAPA, cells treated with
MPPa, RAPA and light. MG-63 cells were placed in complete medium
containing 10% FBS, 90% DMEM and 100 µg/ml penicillin and
streptomycin and cultured at 37°C in a 5% CO2 incubator.
Upon reaching the logarithmic growth phase, the medium was changed.
RAPA (20 ng/ml) was added in the dark to the RAPA and MPPa-PDT-RAPA
groups, MPPa (0.75 µmol/l) was added in the dark to the
MPPa, MPPa-PDT and MPPa-PDT-RAPA groups, and equal volumes of
complete medium were added in the dark to the control and LED
groups. Each group was incubated in a 37°C and 5% CO2
incubator for 20 h in the dark; subsequently, the cultures were
washed twice with PBS, and complete medium was added. The LED,
MPPa-PDT and MPPa-PDT+RAPA groups were irradiated for 120 sec with
an integrated LED special light source (wavelength, 630 nm;
continuous output mode; optical power density, 40
mW/cm2) to provide the cells with a light dose of 4.8
J/cm2 based on a previous study, which demonstrated that
at a light dose of 4.8 J/cm2, the inhibition rate of the
group that received 0.75 µmol/l MPPa was 48.6±2.71%
(9). Following treatment, cells
were collected for subsequent experiments.
Cytotoxicity assay
The anti-proliferative effects of MPPa on human
fibroblast HFL-1 cells was assessed by Cell Counting Kit-8 (CCK-8)
assay (MedChemExpress LLC). Cells were treated with various
concentrations of MPPa (0, 0.5, 0.75 and 1.0 µmol/l) for 24
h. Subsequently, 10 µl CCK-8 was added to each well, and the
plates were incubated for 1-2 h at 37°C according to the
manufacturer's instructions. A microplate reader was used to detect
the absorbance at 450 nm. Data represented the mean of five
replicates. Cell viability was calculated using the following
equation: Cell viability (%)=Average OD in study group/average OD
in control group ×100%, where OD was the optical density. Each
assay was performed in triplicate. Based on the results of the cell
viability assay (Fig. S1), an
appropriate MPPa concentration was selected for subsequent
experiments.
Cell cycle assay
Human osteosarcoma MG-63 cells in the logarithmic
growth phase were inoculated into 6-well plates at a density of
2×106 cells/well, and the control and experimental
groups were established. Each group was placed in 3 replicate wells
that were treated with MPPa, LED or MPPa-PDT for 12 h, and the
supernatants were discarded. Following clearing and washing once
with PBS, the cells were collected, resuspended in 70% ice-cold
ethanol, fixed at 4°C overnight and centrifuged at 1,000 × g for 5
min at 4°C. Cells were washed once with 1 ml PBS, and 500 µl
PBS containing 50 µg/ml PI, 100 µg/ml RNase A and
0.2% Triton X-100 was added and incubated for 30 min at 4°C in the
dark. The fixation solution was discarded, and the cells were
resuspended in an appropriate amount of PBS and subjected to cell
cycle detection using CytoFLEX flow cytometer with CytExpert 2.1
software (Beckman Coulter, Inc.).
Wound-healing assay
Osteosarcoma MG-63 cells in the logarithmic growth
phase were seeded into 6-well plates (2×106 cells/well).
After 70% confluence was reached in each group, MPPa (0.75
µmol/l) was added to the MPPa and MPPa-PDT groups. Following
20-h MPPa treatment, a 10-µl pipette tip was used to make a
scratch in the cell monolayer. The wells were washed three times
with PBS to remove the floating cells, and 2 ml serum-free culture
medium was added to each group. The LED and MPPa-PDT groups were
treated with LED irradiation. Then, the plates were placed in a
37°C and 5% CO2 incubator. After 24 h, cell migration
was observed under an inverted light microscope, and the wells were
photographed to measure the scratch width using Image J 2.1.4
(National Institutes of Health).
Transwell invasion assay
Transwell chambers were placed in 24-well plates.
Matrigel diluted in serum-free medium (1:8) was added to the upper
chambers (100 µl per chamber), and the plates were placed in
37°C with 5% CO2 for 2 h. A total of 100 µl
(3×105 cells/ml) MG-63 cell suspension with or without
MPPa (0.75 µmol/l) was added to the upper chambers, and the
LED and MPPa-PDT groups were serum-starved and treated with MPPa
for 20 h. Following light irradiation, 1 ml medium containing 10%
FBS was added to the lower chambers. After incubation for 48 h at
37°C in a 5% CO2 incubator, the chambers were removed,
and the uninvaded cells in the upper chambers and the Matrigel were
carefully wiped away with a cotton swab. The invading cells that
had passed through the membrane were fixed with 100% methanol for 5
min at room temperature, stained with crystal violet for 15 min at
room temperature and counted using an Olympus upright light
microscope. The experiment was repeated three times.
Detection of intracellular calcium levels
by flow cytometry
MG-63 cells in the logarithmic growth phase were
inoculated at a density of 2×106 cells/well into 6-well
culture plates, incubated for 20 h at 37°C with 5% CO2,
treated according to their group and maintained for 2 h at 37°C in
the dark. Cells were collected and washed three times with PBS. The
calcium ion fluorescent probe FLUO-3/AM (5 µmol/l in DMSO)
was added to the cells and incubated at 37°C for 15 min in the
dark. Cells were washed three times with PBS and collected to make
single-cell suspensions. The intracellular fluorescence intensity
measured by flow cytometry (CytoFLEX flow cytometer; CytExpert 2.1)
represented the calcium ion level.
Apoptosis measurement by flow
cytometry
MG-63 cells were seeded into 6-well plates at a
density of 2×106 cells/well. Suspended and adherent
cells were collected after treatment and analyzed with an Annexin
V/PI apoptosis kit. Briefly, 5 µl PI and FITC-labeled
Annexin V was added to the cells, followed by mixing and incubation
for 15 min at room temperature in the dark. Subsequently, 400
µl 1X binding buffer was added, and the cells were analyzed
by flow cytometry (CytoFLEX flow cytometer; CytExpert 2.1).
Western blotting
After MG-63 cells in the logarithmic growth phase
were treated according to their group, the cells in were collected,
and total protein was extracted using RIPA cell lysate
(Sigma-Aldrich; Merck KGaA). The protein concentrations were
determined using the bicinchoninic acid assay method (5 g/l), and
30 µg total protein per sample was used after boiling and
denaturation. Following 10% SDS-PAGE, the PVDF membranes were
incubated with 5% milk [non-fat milk powder in TBS +0.05% Tween-20
(TBST)] at room temperature for 10 min, and the corresponding
primary antibodies were then added and incubated at 4°C overnight.
After rewarming in room temperature for 30 min, the membranes were
rinsed four times with TBST (10 min/rinse), and a horseradish
peroxidase-labeled secondary antibody was added to the blots and
incubated for 1.5 h at room temperature. Following washing with
TBST, the images were analyzed by ECL chemiluminescence using
Quantity One 4.6.2 software. The antibody information in Table I.
 | Table IAntibody details. |
Table I
Antibody details.
| Protein name | Source | Cat. no. | Dilution | Secondary
antibody |
|---|
| Cyclin D1 | CST | 2978 | 1:1,000 | Anti-rabbit |
| Cyclin E | CST | 20808 | 1:1,000 | Anti-rabbit |
| Cyclin A | CST | 4656 | 1:2,000 | Anti-mouse |
| Cyclin B1 | CST | 12231 | 1:1,000 | Anti-rabbit |
| E-cadherin | CST | 3195 | 1:1,000 | Anti-rabbit |
| MMP-2 | CST | 40994 | 1:1,000 | Anti-rabbit |
| MMP-9 | CST | 13667 | 1:1,000 | Anti-rabbit |
| Akt | CST | 4691 | 1:1,000 | Anti-rabbit |
| p-Akt | CST | 4060 | 1:2,000 | Anti-rabbit |
| mTOR | CST | 2983 | 1:1,000 | Anti-rabbit |
| p-mTOR | CST | 5536 | 1:1,000 | Anti-rabbit |
| 4EBP1 | CST | 9644 | 1:1,000 | Anti-rabbit |
| p-4EBP1 | CST | 2855 | 1:1,000 | Anti-rabbit |
| P70S6K | CST | 2708 | 1:1,000 | Anti-rabbit |
| p-P70S6K | CST | 9208 | 1:1,000 | Anti-rabbit |
| Bip | CST | 3177 | 1:1,000 | Anti-rabbit |
| IRE1α | CST | 3294 | 1:1,000 | Anti-rabbit |
| PERK | CST | 5683 | 1:1,000 | Anti-rabbit |
| Protein
disulfide | CST | 3501 | 1:1,000 | Anti-rabbit |
| CHOP | CST | 2895 | 1:1,000 | Anti-mouse |
| Cleaved
caspase-3 | CST | 9664 | 1:1,000 | Anti-rabbit |
| Cleaved PARP | CST | 5625 | 1:1,000 | Anti-rabbit |
| LC-3 | CST | 13118 | 1:1,000 | Anti-rabbit |
| P62 | CST | 23214 | 1:1,000 | Anti-rabbit |
| p-PERK | Santa Cruz | Sc-32577 | 1:1,000 | Anti-rabbit |
| β-actin | CST | 4970 | 1:1,000 | Anti-rabbit |
|
Goat-anti-rabbit | Invitrogen | G-21040 | 1:10,000 | |
|
Goat-anti-mouse | Invitrogen | 31430 | 1:10,000 | |
Statistical analysis
The data are presented as the mean ± SD, and
statistical comparisons were performed using one-way ANOVA with
Dunnett's test. All statistical analyses were performed using SPSS
software (version 22.0; IBM Corp.). Each experiment was repeated at
least three times. The statistical significance tests were
two-tailed. P<0.05 was considered to indicate a statistically
significant difference.
Results
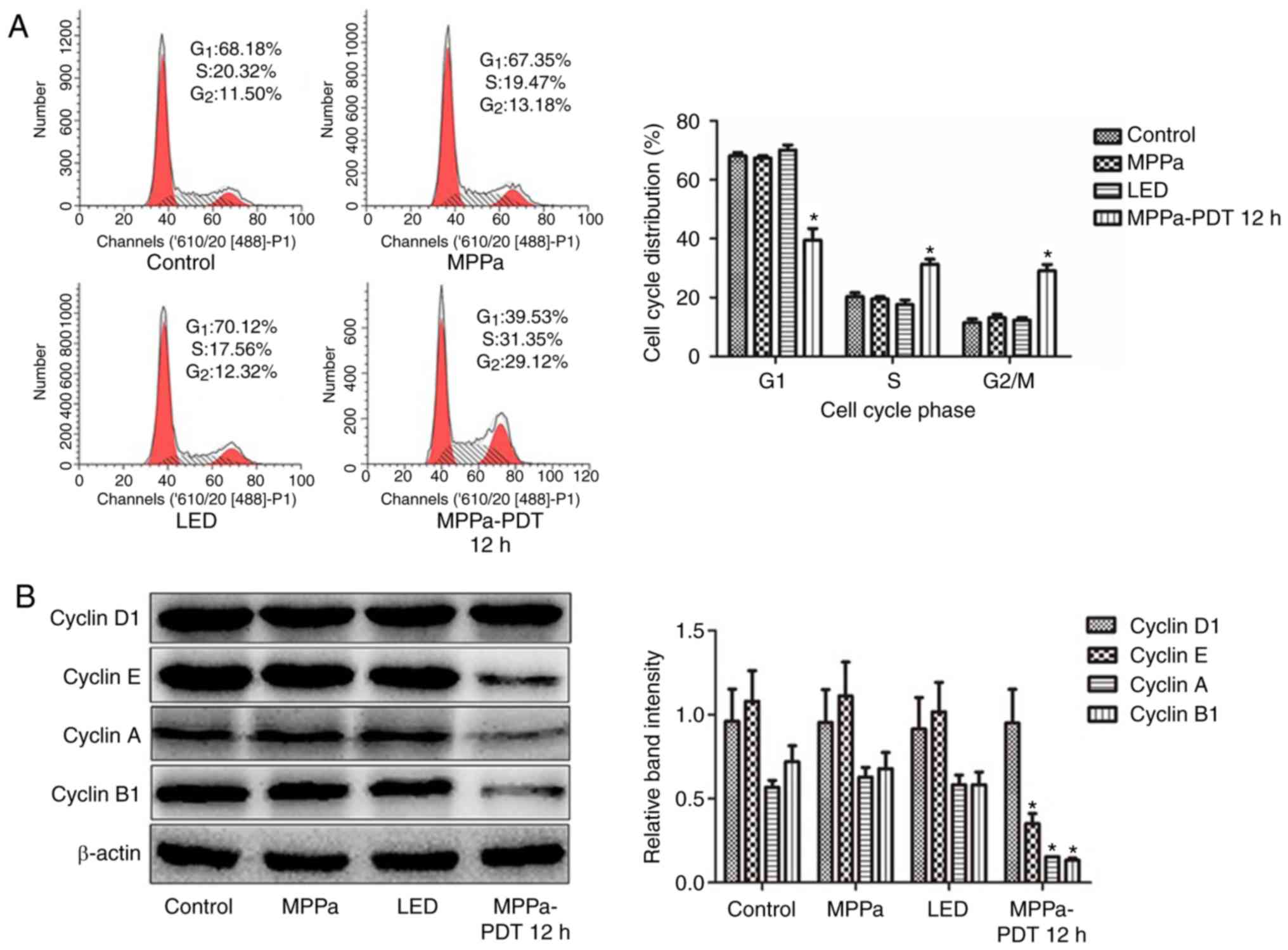
MPPa-PDT induces MG-63 cell cycle
arrest
Previous studies reported that MPPa-PDT inhibited
MG-63 cell proliferation (13).
Therefore, the present study used flow cytometry to evaluate
whether MPPa-PDT blocked cell proliferation by affecting the cell
cycle. Compared with the proportions in the control, MPPa and LED
groups, the proportion of cells in the G1 phase
significantly decreased, the proportion of cells in the
G2/M phase significantly increased and the proportion of
cells in the S phase significantly increased in the MPPa-PDT group
(P<0.05; Fig. 1A). No
significant changes in the cell cycle ratios were observed among
the control, MPPa and LED groups (P>0.05; Fig. 1A).
To investigate the mechanism of MPPa-PDT-induced
cell cycle arrest, western blotting analysis was performed. Cyclin
D1 and Cyclin E are essential proteins in the G1 phase
of cells, and Cyclin A and Cyclin B1 are key proteins in
G2/M phase. Cyclin E, Cyclin A and Cyclin B1 protein
expression levels were significantly lower in the MPPa-PDT group
compared with those in the control, MPPa and LED groups (P<0.05;
Fig. 1B), whereas Cyclin D1
protein expression was not significantly different (P>0.05;
Fig. 1B). No significant
differences were observed in protein expression among the control,
MPPa and LED groups (P>0.05; Fig.
1B). Following MPPa-PDT treatment, the expression of the
G1/S phase checkpoint-associated proteins was partially
downregulated, whereas the expression of G2/M
checkpoint-associated proteins was notably downregulated. These
results were consistent with those obtained for MG-63 cells by flow
cytometry, suggesting that MG-63 cell division was arrested at the
G2/M phase following MPPa-PDT treatment due to the
inhibition of G2/M cycle-related protein expression and
thus cell proliferation.
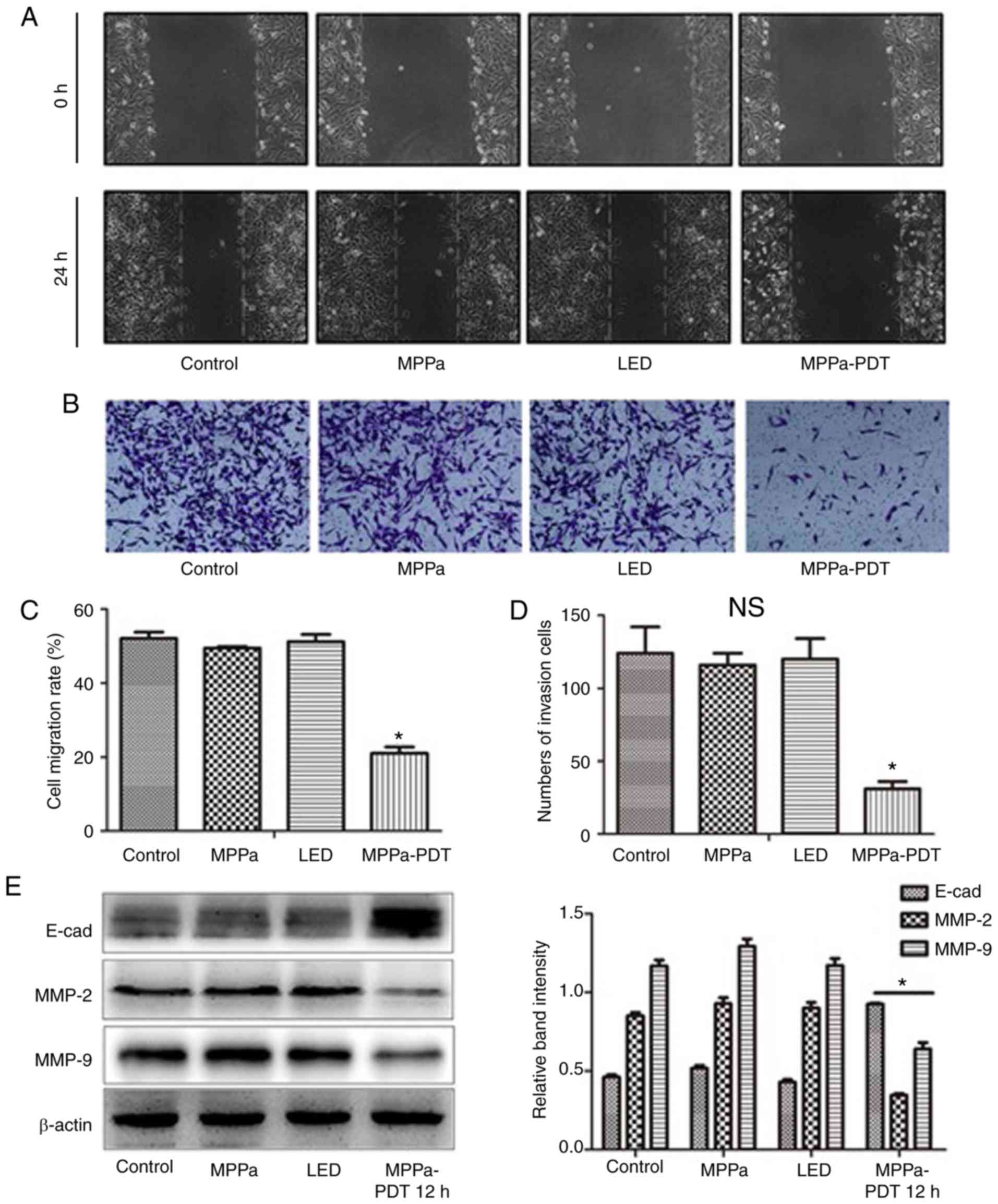
MPPa-PDT inhibits the migration and
invasion of MG-63 cells
To evaluate whether MPPa-PDT inhibited the migration
and invasion of MG-63 cells, wound-healing and invasion assays
using MG-63 cells were performed. The wound-healing assay results
demonstrated that following MPPa-PDT treatment, the cell migration
distance was significantly shorter in the MPPa-PDT group compared
with that in the control, MPPa and LED groups (P<0.05; Fig. 2A and C). No significant
differences were observed in the migration distances among the
control, MPPa and LED groups (P>0.05; Fig. 2A and C). Transwell invasion assay
results demonstrated that after MPPa-PDT treatment, the number of
transmembrane cells was significantly lower in the MPPa-PDT group
compared with that in the control, MPPa and LED groups (P<0.05;
Fig. 2B and D). No significant
differences were observed in the numbers of cells among the
control, MPPa and LED groups (P>0.05; Fig. 2B and D).
Further western blotting results demonstrated that
following MPPa-PDT treatment, the expression levels of the
migration-related protein E-cad in MG-63 cells were higher compared
with those in the control, MPPa and LED groups. By contrast,
expression of the invasion-related proteins MMP-2 and MMP-9 was
decreased in the MPPa-PDT group compared with all other groups
(P<0.05; Fig. 2E). No
significant differences were observed among the control, MPPa and
LED groups (P>0.05; Fig. 2E).
The results were consistent with those from the MG-63 cell
migration and invasion assays. These results suggested that MG-63
cell migration and invasion were inhibited by MPPa-PDT
treatment.
Photodynamic inhibition of the AKT/mTOR
pathway activation in MG-63 cells
Previous studies have demonstrated that the
phosphoinositide 3-kinase (PI3K)/AKT/mTOR pathway serves an
important role in tumor cell proliferation, angiogenesis and
metastasis, as well as in the antagonism of radiotherapy and
chemotherapy (24-26). To investigate whether MPPa-PDT
inhibited the AKT/mTOR pathway activation, western blotting was
performed. Following MPPa-PDT treatment, the levels of the p-AKT,
p-mTOR, 4-EBP-1 and p-P70S6K proteins were significantly lower and
the levels of the corresponding non-phosphorylated AKT, mTOR,
4-EBP-1 and P70S6K proteins were significantly higher in the
MPPa-PDT group compared with the control, MPPa, and LED groups
(P<0.05; Fig. 3A). However, no
significant changes in protein expression were observed among the
control, MPPa and LED groups (P>0.05; Fig. 4A). These results suggested that
the phosphorylation and activation of the AKT/mTOR pathway proteins
in MG-63 cells were inhibited by MPPa-PDT treatment.
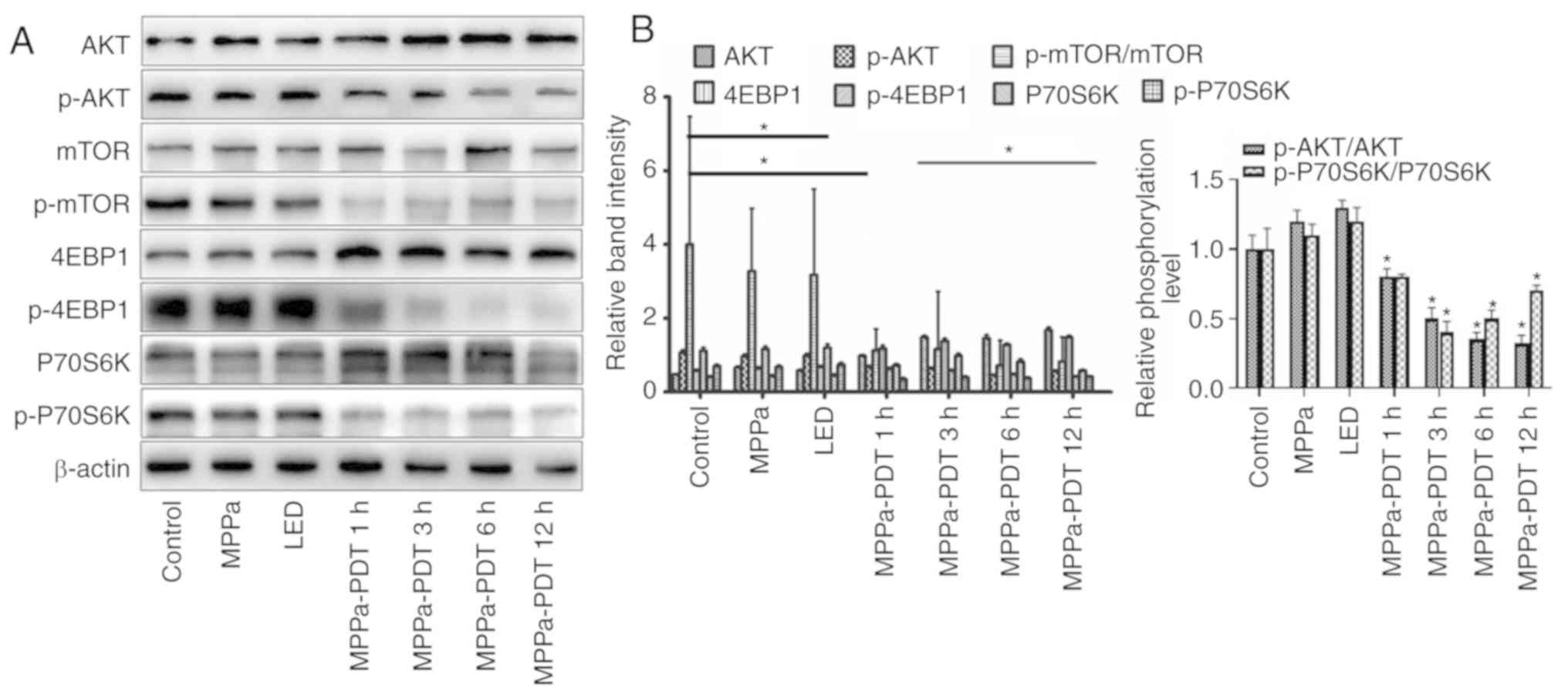 | Figure 3Effects of MPPa-PDT inhibition on the
PI3K/Akt/mTOR signaling pathway. (A) p-Akt, Akt, p-mTOR, mTOR,
p-4EBP1, 4EBP1, p-P70S6K and P70S6K protein expression levels were
analyzed by western blotting analysis. (B) Densitometric analysis
of protein expression. *P<0.05 vs. control. MPPa,
pyropheophorbide-α methyl ester; PDT, photodynamic therapy; LED,
cells treated with a light-emitting diode; p, phosphorylated;
4EBP1, eukaryotic translation initiation factor 4E-binding protein
1. |
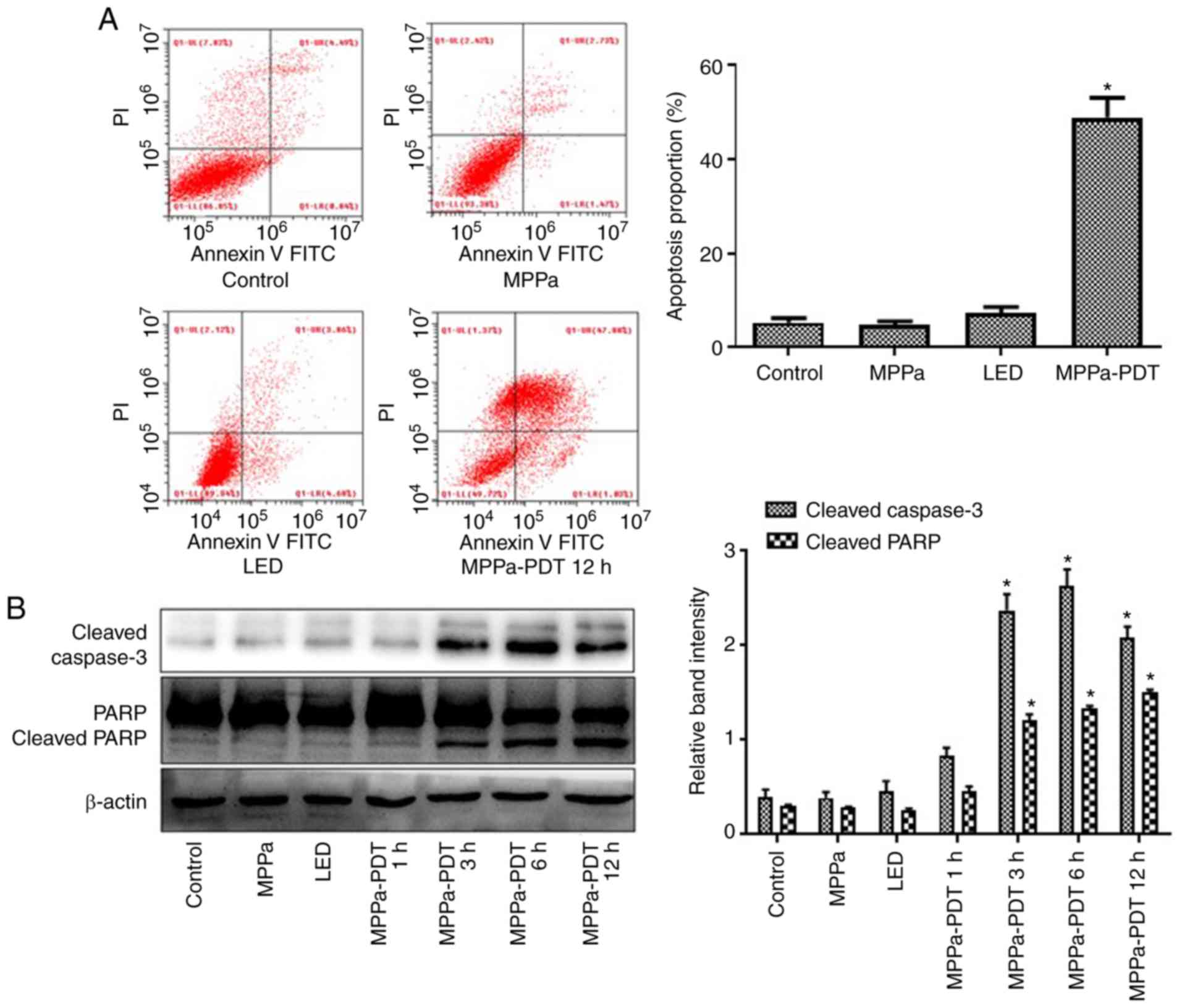
MPPa-PDT induces MG-63 cell
apoptosis
Following MPPa-PDT treatment, the apoptotic rate was
significantly higher in the MPPa-PDT group compared with the
control, MPPa and LED groups according to the flow cytometry assay
(P<0.05; Fig. 4A), whereas no
significant differences were observed among the control, MPPa and
LED groups (P>0.05; Fig. 4A).
Western blotting results indicated that cleaved caspase-3 and
cleaved PARP expression increased after 3 h of MPPa-PDT treatment
(P<0.05; Fig. 4B), but no
notable changes were observed among the control, MPPa and LED
groups (P>0.05; Fig. 4B).
These results suggested that MPPa-PDT induced apoptosis in
osteosarcoma MG-63 cells.
The ERS pathway is involved in
MPPa-PDT-induced apoptosis in MG-63 cells
Flow cytometry assays were performed to investigate
whether ERS contributed to the MPPa-PDT-induced apoptosis in
osteosarcoma MG-63 cells. The intracellular calcium level was
higher in the MPPa-PDT group compared with that in the control,
MPPa and LED groups (P<0.05; Fig.
5A). No significant changes in the calcium ion levels were
observed among the control, MPPa and LED groups (P>0.05;
Fig. 5A). These results suggested
that the MG-63 cells exhibited endoplasmic reticulum damage
following MPPa-PDT treatment.
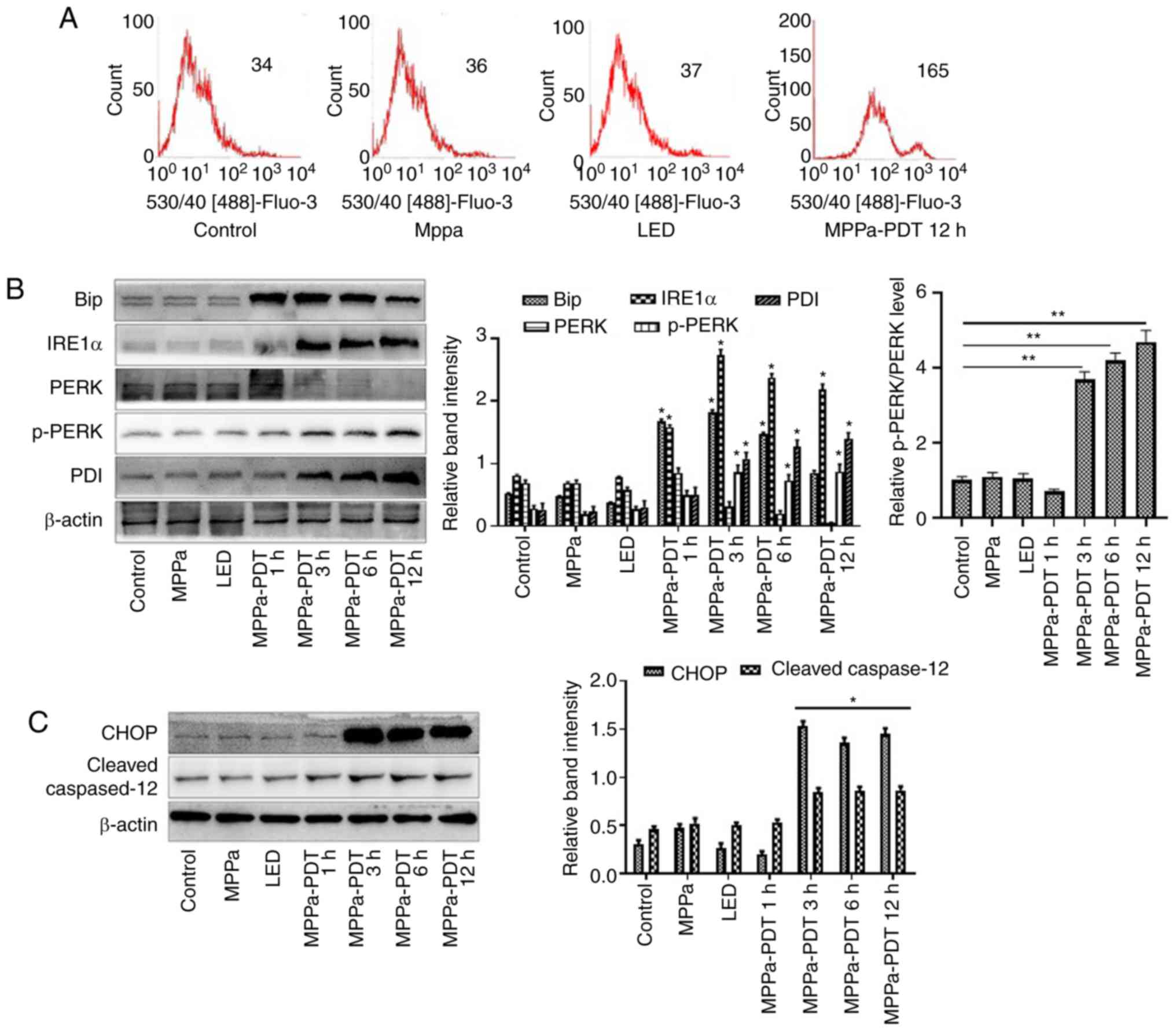 | Figure 5The ERS pathway is involved in
MPPa-PDT-induced MG-63 cell apoptosis. (A) The calcium levels in
the cells were measured by flow cytometry. (B and C) At 12 h,
whole-cell lysates were prepared to determine the expression of (B)
ERS-related proteins (Bip, IRE1α, PERK, p-PERK, and PDI) and (C)
ERS-related apoptotic proteins (CHOP and cleaved caspase-12) by
western blotting analysis. Data are presented as the mean ± SD from
three independent experiments. *P<0.05 vs. control.
MPPa, pyropheophorbide-α methyl ester; PDT, photodynamic therapy;
LED, cells treated with a light-emitting diode; ERS, endoplasmic
reticulum stress; IRE1α, serine/threonine-protein
kinase/endoribonuclease IRE1; PERK, eukaryotic translation
initiation factor 2α kinase 3; PDI, protein disulfide; CHOP,
C/EBP-homologous protein 10. |
In addition, western blotting revealed that at 1, 3,
6 and 12 h after MPPa-PDT treatment, the expression levels of
ERS-related proteins Bip, IRE1α, PERK, p-PERK and PDI and
ERS-related apoptotic proteins CHOP and cleaved caspase-12 were
higher, whereas the expression level of PERK was lower in the
MPPa-PDT-treated MG-63 cells compared with the control groups
(P<0.05; Fig. 5B and C). No
significant changes in these indicators were observed among the
control, MPPa and LED groups (P>0.05; Fig. 5B and C). These results indicated
that ERS was induced by MPPa-PDT in MG-63 cells and that
ERS-induced apoptosis was involved in this process.
mTOR targeting promotes apoptosis by
influencing photodynamic therapy-induced autophagy of osteosarcoma
MG-63
mTOR is not only an important factor regulating cell
growth and proliferation, but also a key promoter of autophagy that
inhibits autophagy after activation (26,27). A previous study has demonstrated
that MPPa-PDT can induce autophagy in MG-63 cells to promote
apoptosis (13). The present
study investigated whether the expression of autophagy-related
proteins of MG-63 cells was altered following MPPa-PDT induction
via mTOR phosphorylation inhibition to promote MG-63 cell apoptosis
and reduce MPPa-PDT tolerance.
The flow cytometry assay results revealed that
following pretreatment with RAPA, the numbers of apoptotic cells
were significantly higher in the MPPA-PDT + RAPA group compared
with the MPPa-PDT group (P<0.05; Fig. 6A), whereas no significant changes
in apoptosis were observed in the RAPA and control groups
(P>0.05; Fig. 6A). Western
blotting results indicated that mTOR expression was downregulated
and the p-mTOR phosphorylation level was further decreased in the
MG-63 cells following RAPA pretreatment; the expression of the
autophagy marker LC3-II was upregulated and P62 expression was
downregulated in MG-63 cells treated with RAPA and MPPa-PDT
compared with the control groups (P<0.05; Fig. 6B). The protein expression levels
of cleaved caspase-3 and cleaved PARP were significantly increased
(P<0.05; Fig. 6B), which was
consistent with the apoptosis results of the flow cytometry assay.
These results suggested that the addition of an mTOR
phosphorylation inhibitor suppressed the autophagy of osteosarcoma
MG-63 cells induced by photo-dynamic therapy and further promoted
apoptosis and reduced the tolerance of MG-63 cells to MPPa-PDT.
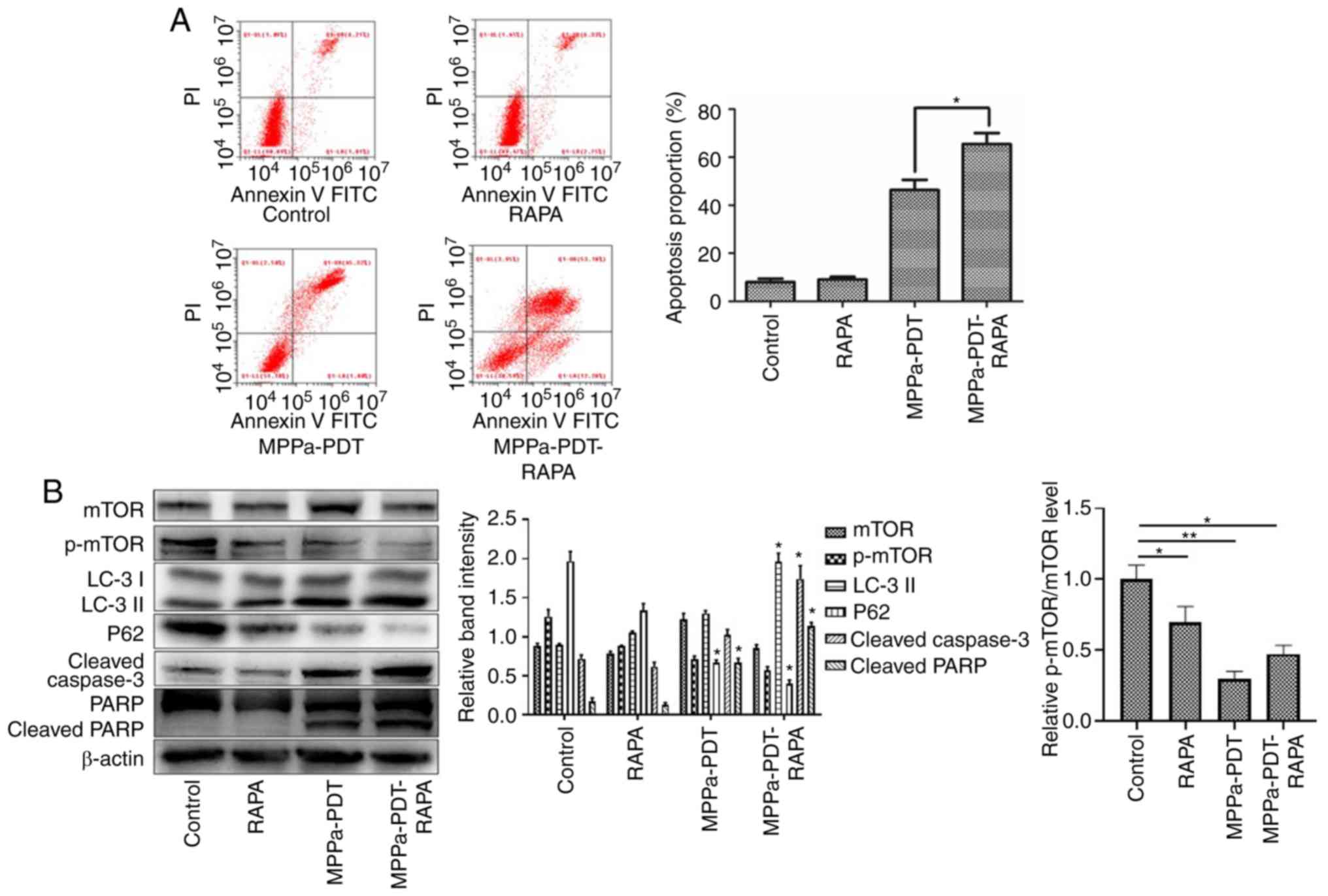 | Figure 6Apoptosis is promoted via mTOR to
enhance MPPa-PDT-induced autophagy in MG-63 cells. MG-63 cells were
pretreated with RAPA for 3 h in the presence or absence of
MPPa-PDT. Control cells received no pretreatment or only RAPA
pretreatment. (A) Following the corresponding treatment, apoptotic
rates were measured by flow cytometry. The apoptotic rate was
calculated as the percentage of early apoptotic (Annexin
V+/PI−) cells plus the percentage of late
apoptotic (Annexin V+/PI+) cells. (B) At 12 h
after treatment, whole-cell lysates were prepared to determine
p-mTOR, LC-3, P62, cleaved caspase-3 and cleaved PARP expression by
western blotting. Data are presented as the mean ± SD from three
independent experiments. *P<0.05 and
**P<0.01. vs. MPPa-PDT. MPPa, pyropheophorbide-α
methyl ester; PDT, photodynamic therapy; RAPA, rapamycin; PI,
propidium iodide; PARP, poly (ADP-ribose) polymerase 1; LC-3,
microtubule-associated protein 1 light chain 3α; p,
phosphorylated. |
Discussion
Currently, surgery combined with neoadjuvant
chemotherapy is the main treatment for osteosarcoma, but the
treatment remains problematic, including drug toxicity, side
effects, a high recurrence rate, chemotherapy resistance and
inoperability due to vascular nerve invasion (6). Therefore, new treatment methods are
urgently needed in clinical practice. PDT is a promising new
approach to cancer therapy that has been approved by the US Food
and Drug Administration for the treatment of tumors (7). Previous studies have reported that
PDT may inhibit the metastasis of osteosarcoma and restore the
immune function of osteosarcoma cells (28,29). MPPa belongs to the second
generation of photosensitizers and is a chlorophyll derivative
(30). To date, MPPa-PDT has been
used clinically for the treatment of a variety of tumors such as
osteosarcoma, lung and cervical cancer (17,19,31). The present study aimed to explore
the functional mechanism of MPPa-PDT in osteosarcoma.
The results of the present study suggested that
MPPa-PDT inhibited cell cycle activity by arresting osteosarcoma
MG-63 cells at the G2/M phase. Previous studies have
demonstrated that Cyclin D1 is an essential protein in the
G1 phase of cells; Cyclin E binds to CDK2 to form a
complex that promotes cells into the S phase, whereas Cyclin A and
Cyclin B1 are key proteins in the G2/M phase (32-34). In the present study, compared with
that in the control group, Cyclin D1 expression was not
significantly different in the MPPa-PDT group, and the decrease in
Cyclin E expression was consistent with the slight decrease in the
proportion of cells in the G1 phase observed in the flow
cytometry analysis of the cell cycle. These results suggested that
MPPa-PDT had a minimal effect on the G1 phase of the
cells. Cyclin A and Cyclin B1 protein expression was significantly
downregulated, and the flow cytometry results demonstrated that the
proportion of cells remaining in the G2/M phase
increased significantly following MPPa-PDT treatment compared with
the other groups, which induced cell cycle arrest and inhibited
cell proliferation.
Migration and invasion are the most prominent
malignant features of osteosarcoma; human osteosarcoma MG-63 cells
exhibit strong migratory and invasive abilities (35,36). The results of the preset study
demonstrated that the wound-healing ability of MG-63 cells and the
number of transmembrane MG-63 cells were significantly decreased
following MPPa-PDT treatment compared with the control groups,
suggesting that MPPa-PDT significantly reduced the migratory and
invasive abilities of osteosarcoma MG-63 cells in vitro. The
E-cad protein serves an important role in the metastasis of
malignant tumors, and the decrease in E-cad protein expression in
tumor tissues leads to intercellular adhesion weakening,
extracellular matrix degradation and basement membrane defects,
which promote the release of tumor cells from their original
positions and allow them to enter the blood and travel to distant
sites (37,38). Zhang et al (39) reported that the specific
inhibition of E-cad protein expression contributed to the distant
metastasis of osteosarcoma MG-63 cells. The results of the present
study demonstrated that E-cad protein expression was upregulated in
osteosarcoma MG-63 cells following MPPa-PDT treatment. In addition,
malignant tumors secrete MMPs and degrade the extracellular matrix
(40). In vivo, MMPs
interact with tissue inhibitors of metalloproteinases (TIMPs) to
maintain the balance of extracellular matrix synthesis and
degradation (41,42); however, MMP expression in
malignant tumor cells is greater than TIMP expression, resulting in
more extracellular matrix degradation than synthesis (43). Therefore, tumor cells are more
susceptible to invasion and metastasis (40-43). Guo et al (44) demonstrated that tectorigenin
significantly inhibited the invasion of osteosarcoma cells by
downregulating MMP-2 and MMP-9 expression. MMP-2 and MMP-9
expression in osteosarcoma MG-63 cells was significantly decreased
after MPPa-PDT treatment compared with the control cells in the
present study. These results suggested that the migratory and
invasive abilities of MG-63 cells were inhibited by MPPa-PDT
treatment.
The PI3K/AKT/mTOR pathway is the main signaling
pathway involved in protein synthesis. It is widely involved in
cell proliferation, differentiation and migration, promotes cell
cycle progression and regulates apoptosis and autophagy (45). Following PI3K activation, AKT
phosphorylation activates downstream signaling molecules, such as
mTOR, and exerts corresponding biological effects (24-26). Studies have demonstrated that mTOR
is an important factor regulating cell growth and proliferation
(24,27). Transient activation of the
AKT/mTOR signaling pathway serves an important role in the
development of a number of malignant tumors such as lung,
colorectal and breast cancer; the downstream serine/threonine
protein kinase P70S6K can increase the translation efficiency of
5′-mTOR mRNA after phosphorylation activation and promote protein
biosynthesis, whereas 4EBP1 phosphorylation promotes its activation
of the eIF4E protein following separation from eIF4E and thus
initiates protein translation (46,47). The results of the present study
demonstrated that phosphorylation of AKT, mTOR, 4EBP1 and P70S6K
was reduced by MPPa-PDT treatment, which inhibited the AKT/mTOR
pathway activity and decreased the quality and efficiency of
protein biosynthesis in MG-63 cells. Indirectly, MPPa-PDT may
inhibit the expression of cell cycle-associated proteins by
blocking the AKT/mTOR pathway activity and thus the MG-63 cell
cycle, thus inhibiting cell proliferation. Previous studies have
reported that MPPa-PDT induced apoptosis in lung, cervical and
breast cancer cells (17,18,30). In the present study, after
MPPa-PDT treatment, flow cytometry assay results demonstrated that
the apoptotic rate of the MPPa-PDT group was significantly
increased. In addition, the expression of the apoptosis-related
proteins cleaved caspase-3 and cleaved PARP was significantly
enhanced. These results indicated that MPPa-PDT induced apoptosis
in human osteosarcoma MG-63 cells.
The endoplasmic reticulum is a very complex
organelle with the ability to control protein synthesis,
transmembrane transport, integration, post-translational
modifications, glyco-sylation modifications, phospholipid and
cholesterol synthesis and intracellular calcium homeostasis. Under
pathological conditions, the disturbance of endoplasmic reticulum
function leads to ERS (48). ERS
triggers an unfolded protein response and intracellular calcium
homeostasis imbalance and increases the level of protein
phosphorylation or expression by activating the endoplasmic
reticulum transmembrane proteins PERK, IRE1α, and ATF6; this step
activates downstream signaling molecules or gene expression and
maintains cell homeostasis, which is essential for cell survival.
However, persistent or excessive ERS promotes the expression of
endoplasmic reticulum-associated death proteins and induces
apoptosis (49-52). In the present study, the level of
intracellular calcium in MG-63 cells was significantly increased
after 3-h MPPa-PDT treatment. Western blotting results also
indicated that Bip, IRE1α and p-PERK expression was significantly
increased following MPPa-PDT treatment. The expression of the
endoplasmic reticulum chaperone protein PDI was also upregulated to
promote the proper folding of unfolded or misfolded proteins and to
reduce cellular damage (48). The
above results indicated that MPPa-PDT may induce ERS in human
osteosarcoma MG-63 cells. CHOP, which is a member of the C/EBP
transcription factor family, is an ERS-specific transcription
factor, and increased expression of this protein mediates apoptosis
(53). Caspase-12, which is a
member of the caspase family, is located on the endoplasmic
reticulum membrane and is a key molecule mediating stress-induced
apoptosis in the endoplasmic reticulum; caspase-12 is activated and
cleaved only when ERS-related apoptosis occurs and mediates further
apoptosis (51). In the present
study, CHOP and cleaved caspase-12 expression in MG-63 cells was
significantly increased after MPPa-PDT treatment compared with the
control groups. These results indicated that MPPa-PDT induced ERS
in MG-63 cells and upregulated CHOP and cleaved caspase-12
expression, which activated the ERS apoptosis pathway and
ultimately induced apoptosis.
The sensitivity of MG-63 cells to MPPa-PDT was
enhanced using mTOR as an intervention target to influence
autophagy, promote osteosarcoma cell apoptosis and increase the
effect of MPPa-PDT. The mTOR molecule is a key promoter of the
autophagy process and can downregulate cellular autophagy activity
upon activation (54,55). A previous study reported that
autophagy induced by MPPa-PDT promoted MG-63 cell apoptosis
(13). Following the addition of
RAPA, which is a phosphorylation inhibitor of mTOR, the degree of
mTOR phosphorylation was further inhibited, the expression of
autophagy-related proteins was significantly altered, and the
apoptotic rate of the MG-63 cells was further enhanced compared
with MPPa-PDT treatment alone. Therefore, with mTOR as a target,
the effects of MPPa-PDT on MG-63 cells can be improved by
downregulating the degree of mTOR phosphorylation.
In conclusion, the results of the present study
demonstrated that MPPa-PDT blocked the MG-63 cell cycle, inhibited
cell migration, invasion and the AKT/mTOR pathway activation, and
induced ERS and ERS-induced apoptosis in MG-63 cells. At the same
time, mTOR may enhance the effects of MPPa-PDT on MG-63 cells as an
intervention target. These results enrich our understanding of
MPPa-PDT in the treatment of osteosarcoma cells; however, the
present results and targets of MPPa-PDT for the treatment of
osteosarcoma require further studies in multiple cell lines and
in vivo experiments prior to clinical application.
Supplementary Data
Funding
This study was supported by the National Natural
Science Foundation of China (81572634).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC and HY performed the experimental work, data
collection and interpretation. YT and YO participated in the design
and coordination of the experimental work and the acquisition of
data. SZ and HYY participated in the study design, data collection
and analysis and manuscript preparation. JL and ZB participated in
the study design, analyzed and interpreted the data and drafted the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy and integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The human fibroblast HFL-1 cell line was donated by
Professor Zhou Jing, Department of Respiratory, the First
Affiliated Hospital of Chongqing Medical University (Chongqing,
China).
References
|
1
|
Ando K, Heymann MF, Stresing V, Mori K,
Redini F and Heymann D: Current therapeutic strategies and novel
approaches in osteosarcoma. Cancers (Basel). 5:591–616. 2013.
View Article : Google Scholar
|
|
2
|
Yu W, Wang Y, Zhu J, Jin L, Liu B, Xia K,
Wang J, Gao J, Liang C and Tao H: Autophagy inhibitor enhance
ZnPc/BSA nanoparticle induced photodynamic therapy by suppressing
PD-L1 expression in osteosarcoma immunotherapy. Biomaterials.
192:128–39. 2019. View Article : Google Scholar
|
|
3
|
Arai K, Sakamoto R, Kubota D and Kondo T:
Proteomic approach toward molecular backgrounds of drug resistance
of osteosarcoma cells in spheroid culture system. Proteomics.
13:2351–2360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cao D, Zhu W, Kuang Y and Zhao S: A safe
and effective treatment: Surgery combined with photodynamic therapy
for multiple basal cell carcinomas. Photodiagnosis Photodyn Ther.
28:133–135. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Ramamurthy G, Shirke AA, Walker E,
Mangadlao J, Wang Z, Wang Y, Shan L, Schluchter MD, Dong Z, et al:
Photodynamic therapy is an effective adjuvant therapy for
image-guided surgery in prostate cancer. Cancer Res. 80:156–162.
2020.
|
|
6
|
Wu ZJ, Tan JC, Qin X, Liu B and Yuan ZC:
Significance of circulating tumor cells in osteosarcoma patients
treated by neoadjuvant chemotherapy and surgery. Cancer Manag Res.
10:3333–3339. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nishie H, Kataoka H, Yano S, Yamaguchi H,
Nomoto A, Tanaka M, Kato A, Shimura T, Mizoshita T, Kubota E, et
al: Excellent antitumor effects for gastrointestinal cancers using
photodynamic therapy with a novel glucose conjugated chlorin e6.
Biochem Biophys Res Commun. 496:1204–1209. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Savoia P, Deboli T, Previgliano A and
Broganelli P: Usefulness of photodynamic therapy as a possible
therapeutic alternative in the treatment of basal cell carcinoma.
Int J Mol Sci. 16:23300–23317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Phuong PTT, Lee S, Lee C, Seo B, Park S,
Oh KT, Lee ES, Choi HG, Shin BS and Youn YS: Beta-carotene-bound
albumin nanoparticles modified with chlorin e6 for breast tumor
ablation based on photodynamic therapy. Colloids Surf B
Biointerfaces. 171:123–133. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fujishiro T, Nonoguchi N, Pavliukov M,
Ohmura N, Kawabata S, Park Y, Kajimoto Y, Ishikawa T, Nakano I and
Kuroiwa T: 5-Aminolevulinic acid-mediated photodynamic therapy can
target human glioma stem-like cells refractory to antineoplastic
agents. Photodiagnosis Photodyn Ther. 24:55–68. 2018. View Article : Google Scholar
|
|
11
|
Kessel D and Oleinick NL: Cell death
pathways associated with photodynamic therapy: An update. Photochem
Photobiol. 94:213–218. 2018. View Article : Google Scholar
|
|
12
|
Zhang Q and Li L: Photodynamic
combinational therapy in cancer treatment. J BUON. 23:561–567.
2018.PubMed/NCBI
|
|
13
|
Huang Q, Ou YS, Tao Y, Yin H and Tu PH:
Apoptosis and autophagy induced by pyropheophorbide-α methyl
ester-mediated photodynamic therapy in human osteosarcoma MG-63
cells. Apoptosis. 21:749–760. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chang JE, Yoon IS, Sun PL, Yi E, Jheon S
and Shim CK: Anticancer efficacy of photodynamic therapy with
hematoporphyrin-modified, doxorubicin-loaded nanoparticles in liver
cancer. J Photochem Photobiol B. 140:49–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Silva AP, Kurachi C, Bagnato VS and Inada
NM: Fast elimination of onychomycosis by hematoporphyrin
derivative-photodynamic therapy. Photodiagnosis Photodyn Ther.
10:328–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zeng H, Sun M, Zhou C, Yin F, Wang Z, Hua
Y and Cai Z: Hematoporphyrin monomethyl ether-mediated photodynamic
therapy selectively kills sarcomas by inducing apoptosis. PLoS One.
8:e777272013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tao Y, Ou Y, Yin H, Chen Y, Zhong S, Gao
Y, Zhao Z, He B, Huang Q and Deng Q: Establishment and
characterization of human osteosarcoma cells resistant to
pyropheophorbide-α methyl ester-mediated photodynamic therapy. Int
J Oncol. 51:1427–1438. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qian G, Wang L, Zheng X and Yu T:
Deactivation of cisplatin-resistant human lung/ovary cancer cells
with pyropheophorbide-α methyl ester-photodynamic therapy. Cancer
Biol Ther. 18:984–989. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tu PH, Huang WJ, Wu ZL, Peng QZ, Xie ZB,
Bao J and Zhong MH: Induction of cell death by pyropheophorbide-α
methyl ester-mediated photodynamic therapy in lung cancer A549
cells. Cancer Med. 6:631–639. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu J, Tian S, Li KT, Chen Q, Jiang Y, Lin
HD, Yu LH and Bai DQ: Inhibition of breast cancer cell growth by
methyl pyropheophenylchlorin photodynamic therapy is mediated
though endoplasmic reticulum stress-induced autophagy in vitro and
vivo. Cancer Med. 7:1908–1920. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tu PH, Huang WJ, Wu ZL, Peng QZ, Xie ZB,
Bao J and Zhong MH: Induction of cell death by pyropheophorbide-α
methyl ester-mediated photodynamic therapy in lung cancer A549
cells. Cancer Med. 6:631–639. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tian Y, Leung W, Yue K and Mak N: Cell
death induced by MPPa-PDT in prostate carcinoma in vitro and in
vivo. Biochem Biophys Res Commun. 348:413–420. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
de Miguel GC, Abrantes AM, Laranjo M,
Grizotto AYK, Camporeze B, Pereira JA, Brites G, Serra A, Pineiro
M, Rocha-Gonsalves A, et al: A new therapeutic proposal for
inoperable osteosarcoma: Photodynamic therapy. Photodiagnosis
Photodyn Ther. 21:79–85. 2018. View Article : Google Scholar
|
|
24
|
Gao W, Wu XL, Li DZ and Liu HD: HOTTIP
participates in mammary cancer by promoting cell proliferation via
PI3K/AKT pathway. Eur Rev Med Pharmacol Sci. 22:4181–4187.
2018.PubMed/NCBI
|
|
25
|
Liu HW, Bi WT, Huang HT, Li RX, Xi Q, Feng
L, Bo W, Hu M and Wen WS: Satb1 promotes Schwann cell viability and
migration via activation of PI3K/AKT pathway. Eur Rev Med Pharmacol
Sci. 22:4268–4277. 2018.PubMed/NCBI
|
|
26
|
Wang J, Sun P, Chen Y, Yao H and Wang S:
Novel 2-pheny-loxypyrimidine derivative induces apoptosis and
autophagy via inhibiting PI3K pathway and activating MAPK/ERK
signaling in hepatocellular carcinoma cells. Sci Rep. 8:109232018.
View Article : Google Scholar
|
|
27
|
Lijun J, Shan H, Xiaoran Y, Zan Y, Guo Y
and Han L: Quercetin suppresses the mobility of breast cancer by
suppressing glycolysis through Akt-mTOR pathway mediated autophagy
induction. Life Sci. 208:123–130. 2018. View Article : Google Scholar
|
|
28
|
Meier D, Botter SM, Campanile C, Robl B,
Gräfe S, Pellegrini G, Born W and Fuchs B: Foscan and foslip based
photodynamic therapy in osteosarcoma, in vitro, and in intratibial
mouse models. Int J Cancer. 140:1680–1692. 2017. View Article : Google Scholar
|
|
29
|
Zhang F, Zhu Y, Fan G and Hu S:
Photodynamic therapy reduces the inhibitory effect of osteosarcoma
cells on dendritic cells by upregulating HSP70. Oncol Lett.
16:5034–5040. 2018.PubMed/NCBI
|
|
30
|
Huang L, Lin H, Chen Q, Yu L and Bai D:
MPPa-PDT suppresses breast tumor migration/invasion by inhibiting
Akt-NF-κB-dependent MMP-9 expression via ROS. BMC Cancer.
19:11592019. View Article : Google Scholar
|
|
31
|
Li W, Tan G, Cheng J, Zhao L, Wang Z and
Jin Y: A novel photosensitizer 3¹,13¹-phenylhydrazine-Mppa (BPHM)
and Its in vitro photodynamic therapy against HeLa cells.
Molecules. 21:pii: E558. 2016.
|
|
32
|
Santti K, Ihalainen H, Rönty M, Böhling T,
Karlsson C and Haglund C: High cyclin A expression, but not Ki67,
is associated with early recurrence in desmoid tumors. J Surg
Oncol. 118:192–198. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lal N, Nemaysh V and Luthra PM: Proteasome
mediated degradation of CDC25C and Cyclin B1 in Demethoxycurcumin
treated human glioma U87 MG cells to trigger G2/M cell cycle
arrest. Toxicol Appl Pharmacol. 356:76–89. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Marampon F, Gravina G, Ju X, Vetuschi A,
Sferra R, Casimiro M, Pompili S, Festuccia C, Colapietro A, Gaudio
E, et al: Cyclin D1 silencing suppresses tumorigenicity, impairs
DNA double strand break repair and thus radiosensitizes
androgen-independent prostate cancer cells to DNA damage.
Oncotarget. 7:5383–5400. 2016.
|
|
35
|
Cai H, Miao M and Wang Z: miR-214-3p
promotes the proliferation, migration and invasion of osteosarcoma
cells by targeting CADM1. Oncol Lett. 16:2620–2628. 2018.PubMed/NCBI
|
|
36
|
Qiu J, Zhang Y, Chen H and Guo Z:
MicroRNA-488 inhibits proliferation, invasion and EMT in
osteosarcoma cell lines by targeting aquaporin 3. Int J Oncol.
53:1493–1504. 2018.PubMed/NCBI
|
|
37
|
Campbell K and Casanova J: A role for
E-cadherin in ensuring cohesive migration of a heterogeneous
population of non-epithelial cells. Nat Commun. 6:79982015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shan Z, Wei Z and Shaikh ZA: Suppression
of ferroportin expression by cadmium stimulates proliferation, EMT,
and migration in triple-negative breast cancer cells. Toxicol Appl
Pharmacol. 356:36–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang LZ, Mei J, Qian ZK, Cai XS, Jiang Y
and Huang WD: The role of VE-cadherin in osteosarcoma cells. Pathol
Oncol Res. 16:111–117. 2010. View Article : Google Scholar
|
|
40
|
Li F, Zhang J, Guo J, Jia Y, Han Y and
Wang Z: RNA interference targeting CD147 inhibits metastasis and
invasion of human breast cancer MCF-7 cells by downregulating
MMP-9/VEGF expression. Acta Biochim Biophys Sin (Shanghai).
50:676–684. 2018. View Article : Google Scholar
|
|
41
|
Liao CL, Chu YL, Lin HY, Chen CY, Hsu MJ,
Liu KC, Lai KC, Huang AC and Chung JG: Bisdemethoxycurcumin
suppresses migration and invasion of human cervical cancer HeLa
cells via inhibition of NF-ĸB, MMP-2 and -9 pathways. Anticancer
Res. 38:3989–3997. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu M, Jiang H, Wang H, Liu J, Liu B and
Guo Z: SB225002 inhibits prostate cancer invasion and attenuates
the expression of BSP, OPN and MMP-2. Oncol Rep. 40:726–736.
2018.PubMed/NCBI
|
|
43
|
Shimoda M, Jackson HW and Khokha R: Tumor
suppression by stromal TIMPs. Mol Cell Oncol. 3:e9750822016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Guo Y, Chen YH, Cheng ZH, Ou-Yang HN, Luo
C and Guo ZL: Tectorigenin inhibits osteosarcoma cell migration
through downregulation of matrix metalloproteinases in vitro.
Anticancer Drugs. 27:540–546. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu JZ, Hu YL, Feng Y, Guo YB, Liu YF,
Yang JL, Mao QS and Xue WJ: Rafoxanide promotes apoptosis and
autophagy of gastric cancer cells by suppressing PI3K/Akt/mTOR
pathway. Exp Cell Res. 385:1116912019. View Article : Google Scholar
|
|
46
|
Zhou M, Shen S, Zhao X and Gong X:
Luteoloside induces G0/G1 arrest and pro-death autophagy through
the ROS-mediated AKT/mTOR/p70S6K signalling pathway in human
non-small cell lung cancer cell lines. Biochem Biophys Res Commun.
494:263–269. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu Y, Rao Q, Zhang X and Zhou X: Galangin
induced antitumor effects in human kidney tumor cells mediated via
mitochondrial mediated apoptosis, inhibition of cell migration and
invasion and targeting PI3K/AKT/mTOR signalling pathway. J BUON.
23:795–799. 2018.PubMed/NCBI
|
|
48
|
Santamaria PG, Mazón MJ, Eraso P and
Portillo F: UPR: An upstream signal to EMT induction in cancer. J
Clin Med. 8:pii: E624. 2019. View Article : Google Scholar
|
|
49
|
Santofimia-Castaño P, Izquierdo-Alvarez A,
Plaza-Davila M, Martinez-Ruiz A, Fernandez-Bermejo M,
Mateos-Rodriguez JM, Salido GM and Gonzalez A: Ebselen impairs
cellular oxidative state and induces endoplasmic reticulum stress
and activation of crucial mitogen-activated protein kinases in
pancreatic tumour AR42J cells. J Cell Biochem. 119:1122–1133. 2018.
View Article : Google Scholar
|
|
50
|
Hetz C and Saxena S: ER stress and the
unfolded protein response in neurodegeneration. Nat Rev Neurol.
13:477–491. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Badiola N, Penas C, Miñano-Molina A,
Barneda-Zahonero B, Fadó R, Sánchez-Opazo G, Comella JX, Sabriá J,
Zhu C, Blomgren K, et al: Induction of ER stress in response to
oxygen-glucose deprivation of cortical cultures involves the
activation of the PERK and IRE-1 pathways and of caspase-12. Cell
Death Dis. 2:e1492011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guha P, Kaptan E, Gade P, Kalvakolanu DV
and Ahmed H: Tunicamycin induced endoplasmic reticulum stress
promotes apoptosis of prostate cancer cells by activating mTORC1.
Oncotarget. 8:68191–68207. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sidhu A, Miller JR, Tripathi A, Garshott
DM, Brownell AL, Chiego DJ, Arevang C, Zeng Q, Jackson LC, Bechler
SA, et al: Borrelidin induces the unfolded protein response in oral
cancer cells and chop-dependent apoptosis. ACS Med Chem Lett.
6:1122–1127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou J, Tan SH, Nicolas V, Bauvy C, Yang
ND, Zhang J, Xue Y, Codogno P and Shen HM: Activation of lysosomal
function in the course of autophagy via mTORC1 suppression and
autophagosome-lysosome fusion. Cell Res. 23:508–523. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tao YF, Li ZH, Du WW Xu LX, Ren JL, Li XL,
Fang F, Xie Y, Li M, Qian GH, et al: Inhibiting PLK1 induces
autophagy of acute myeloid leukemia cells via mammalian target of
rapamycin pathway dephosphorylation. Oncol Rep. 37:1419–1429. 2017.
View Article : Google Scholar : PubMed/NCBI
|