Introduction
Colon cancer is one of the most common malignant
alimentary canal tumor types, and mainly occurs on the joint of the
rectum and colon sigmoideum (1).
A total of 1.2 million patients are diagnosed annually and ~0.6
million mortalities occur each year globally (1). Nearly 60% patients with colon cancer
will develop metastasis, which results in a high mortality of
patients with colon cancer (2,3).
Colon cancer is the fifth highest cause of cancer-associated
mortalities in China, and the morbidity continues to rapidly rise,
with a 5-year survival rate of 40-60% (4). Therefore, it is important to further
reveal the underlying mechanisms of colon cancer.
An increasing number of herbs have been used in
medicines and cosmetics for centuries. Among them, wild
chrysanthemum (Chrysanthemum indicum L.) has a range of
biological activities. In China, wild chrysanthemum is used to cure
numerous types of inflammatory diseases (5). Studies have documented that
flavonoids are the active ingredients in wild chrysanthemum,
including hispidulin, isorhoifolin and scutellarein (6-8).
Scutellarein, a flavone present in the perennial herb
Scutellaria baicalensis, is the aglycone of scutellarin with
a free hydroxyl in 7 position, and exerts a higher bioavailability
compared with scutellarin (9). A
variety of studies have demonstrated that scutellarein may inhibit
the viability of human lung cancer cells and fibrosarcoma cells
(9-11), in addition to serving an
anti-tumor function in human colon cancer (12,13). However, the underlying mechanisms
of scutellarin, derived from the wild chrysanthemum, in the
repression of colon cancer progression remain largely unknown.
Cluster of differentiation (CD) family proteins,
including CD44, CD50 and CD74, are all members of the
immuno-globulin superfamily, and are associated with cancer cell
migration and immune response (14,15). Melanoma cell adhesion molecule
(MCAM), also known as CD146, is a cell adhesion molecule and is
involved in several cellular processes, including cell invasion,
migration, immune response and signal transduction (16). The increased expression of MCAM in
the primary tumor types is associated with metastasis and prognosis
in several cancer types (16-18). Mucosal addressin cell adhesion
molecule 1 (Madcam1) is known as a type of adhesion molecule, which
is involved in inflammatory responses with no expression in the
majority of normal tissues (19,20). Receptor for advanced glycation end
products (RAGE) is a signal transduction receptor that is involved
in multiple pathologic conditions, including carcinogenesis and
inflammation relying on its diverse ligands (21,22). It is well acknowledged that RAGE
expression is associated with multifarious cancer types, including
colon cancer (23). These results
suggest that all of the aforementioned proteins serve a function in
carcinogenesis. However, whether scutellarein inhibits the
development of colon cancer through modulating these oncoproteins
(CD44, CD50, CD74, CD138, MCAM, CD151, CD166, CD206, RAGE and
Madcam1) is not yet known.
Ubiquitylation is considered to be an important
modification of nuclear and cytoplasmic proteins at the
post-translational level, as it is known that ubiquitylation serves
an important function in cell apoptosis, cell cycle and DNA damage
repair (24,25). A number of studies have
demonstrated that ubiquitylation is strongly associated with the
initiation and progression of carcinogenesis (26,27). Notably, numerous
ubiquitin-associated proteins have been revealed as tumor
suppressors/oncogenes and therapeutic targets in different tumor
types (28,29). However, whether ubiquitylation is
involved in scutellarein-mediated colon cancer repression remains
unknown.
The present study aimed to investigate the function
of scutellarein in ubiquitylation, and to determine the expression
of a number of ubiquitin-associated proteins in the presence of
scutellarein. In order to elucidate the functions of scutellarein
for the potential treatment of colon cancer, the anticancer
function of scutellarein from wild chrysanthemum in the development
of colon cancer cells and its associated mechanisms were
examined.
Materials and methods
Extraction of scutellarein
A whole plant of wild chrysanthemum was gathered
from the wild, dried in the air and then ground, followed by
extraction using 99.5% ethanol three times. Subsequently, the
ethanol was removed and the extract was suspended in water and
partitioned with petroleum ether and acetyl acetate. Under reduced
pressure, the acetyl acetate was evaporated and obtained. This was
handled with column chromatography on silica gel eluted with
gradient dichloromethane and purified by Sephadex LH-20 to obtain a
yellow powder. The structure of the compound was determined using
1H and 13C nuclear magnetic resonance (NMR) spectra and
electrospray ionization mass spectrometry analysis. The results
revealed the peaks to be at m/z 287[M+H]+ and
285[M−H]−, demonstrating that the molecular
weight was 286 g/mol. On the other hand, two single peaks at δ 6.77
(1H, s) and 6.57 (1H, s) were observed on the 1H NMR spectrum.
Furthermore, on the 13C NMR spectrum, 13 carbon signals were
detected. The purity of the scutellarein was identified by using
high performance liquid chromatography on an Agilent 1,100 high
performance liquid chromatography system (Agilent Technologies,
Inc.), and was determined to be 96.5%. The chromatographic column
was Waters Xterra Rp-C18 (3.0×100 mm; 3.5 µm),
the mobile phase A was acetonitrile solution, and the mobile phase
B was 0.1% formic acid water. The gradient elution conditions were
10-20% A (0-10 min), 20-25% A (10-30 min) and 25-45% A (30-60 min),
and the volume flow rate was 0.4 ml/min with a column temperature
of 20°C and an injection volume of 5 µl. The UV detection
wavelength was 275 nm, and the running time was 60 min.
Cell culture
Colon cell lines CL-40, SW480 and T84 were all
obtained from the American Type Culture Collection. T84 cells were
maintained in high glucose-DMEM medium (Gibco; Thermo Fisher
Scientific, Inc.) with the presence of 10% fetal bovine serum (FBS;
Biological Industries) and 1% (v/v) penicillin-streptomycin, while
SW480 cells were cultured in DMEM/F-12 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS and 1%
penicillin-streptomycin, and CL-40 cells were incubated with
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS and 1% penicillin-streptomycin. All cells
were maintained in a humidified condition of 5% CO2 at
37°C.
MTT analysis
MTT analysis was performed to investigate the half
maximal inhibitory concentration (IC50) of scutellarein
in colon cancer cells and cell viability. SW480 and T84 cells
(3,000 cells/well) were cultured in 96-well plates for 24 h
following adherence, and were then exposed to scutellarein at a
gradient concentration for 36 h. Subsequently, 10 µl MTT
solution (Beyotime Institute of Biotechnology) were added per well
and the cells were incubated for another 4 h at 37°C. DMSO was used
to dissolve the purple formazan. Then, the optical density (OD)
values were measured at 570 nm, and the IC50 value was
calculated. In addition, cell viability was also detected using
MTT. Cells were cultured in 96-well plates, and underwent different
treatments, including scutellarein, scutellarein+RAGE
overexpression (OE-RAGE), scutellarein+RAGE short hairpin RNA
(sh-RAGE), OE-CDC4, OE-CDC4+OE-RAGE, following adherence. Following
incubation at 37°C for 1, 2, 3, 4 and 5 days, the cells were
treated with MTT solution for 4 h, and the OD values were measured
at 570 nm.
Western blot analysis
The colon cancer SW480 and T84 cells were seeded
into 6-well plates and cultured for 12 h until adherence. Then,
cells underwent different treatments (scutellarein,
scutellarein+OE-RAGE, scutellarein+sh-RAGE, OE-CDC4 and
OE-CDC4+OE-RAGE) for 48 h, and then the cells were gathered. The
total protein was isolated using RIPAIII (Beijing Solarbio Science
& Technology Co., Ltd.) and centrifuged at 12,000 × g at 4°C
for 20 min. Proteins from supernatants were quantified using
bicinchoninic acid analysis (EMD Millipore) and boiled with 1x
loading buffer. A total of 20 mg protein was separated using 10%
SDS-PAGE, and was subsequently transferred onto polyvinylidene
fluoride membranes (EMD Millipore). The membranes were immersed
into 5% non-fat milk for 1 h at room temperature to block,
following by being washed with TBS with 0.1% (v/v) Tween-20 a total
of 3 times. Then, the membranes were incubated with primary
antibodies at 4°C overnight. Next day, the membranes were incubated
with the corresponding horseradish peroxidase (HRP)-conjugated
anti-mouse (cat. no. 7076) or anti-rabbit (cat. no. 7074) secondary
antibody (1:10,000 dilution; Cell Signaling Technology, Inc.) at
room temperature for 2 h. Subsequently, the complexes were tested
using the electrochemical luminescence of horseradish peroxidase
substrate (EMD Millipore) and analyzed using ImageJ software
(version 1.48; National Institutes of Health). The primary
antibodies were as follows: Anti-CD44 (1:1,000; cat. no. #3578),
anti-ring finger and CHY zinc finger domain containing 1 (RCHY1;
1:1,000; cat. no. #5754), anti-cleaved caspase3 (1:1,000; cat. no.
#9661), anti-caspase3 (1:1,000; cat. no. #9662), anti-cleaved
caspase7 (1:1,000; cat. no. #9492), anti-caspase7 (1:1,000, no.
#9491), anti-phosphorylated (p-) p65 (1:1,000; cat. no. #3033),
anti-p65 (1:1,000; cat no. #8242) and anti-protein kinase C (PKC;
1:1,000; cat. no. #2056) antibodies were all obtained from Cell
Signaling Technology, Inc. Anti-RAGE (1:1,000; cat. no. sc365154),
anti-cell division control protein 4 (CDC4; 1:1,000; cat. no.
sc293423), anti-CD50 (1:1,000; cat. no. sc71307), anti-CD74
(1:1,000; cat. no. sc166047), anti-CD138 (1:1,000; cat. no.
sc390791), anti-CD151 (1:1,000; cat. no. sc271216), anti-CD166
(1:1,000; cat. no. sc74558), anti-CD206 (1:1,000; cat. no.
sc70586), anti-MCAM (1:1,000; cat. no. sc376762), anti-Madcam1
(1:1,000; cat. no. sc374398), anti-Cbl proto-oncogene (CBL;
1:1,000; cat. no. sc1651), anti-Ubiquitin (1:1,000; cat. no.
sc166553), anti-SMAD specific ubiquitin protein ligase 1 (Smurf1;
1:1,000; cat. no. sc100616), anti-MDM2 proto-oncogene (MDM2;
1:1,000; cat. no. sc5304) and anti-GAPDH (1:1,000; cat. no.
sc47724) were all purchased from Santa Cruz Biotechnology, Inc.,
and anti-vascular endothelial growth factor (VEGF; 1:1,000; cat.
no. ab46154) was purchased from Abcam. The corresponding secondary
antibodies were obtained from OriGene Technologies, Inc.
Cycloheximide (CHX) analysis
The colon cancer SW480 and T84 cells were routinely
cultured and underwent different treatments. Then, the cells were
treated with CHX (100 µg/ml) for 1, 2, 4, 8 and 24 h at
37°C, following which the total protein was extracted.
Colony formation assay
Colon cancer SW480 and T84 cells (1,000 cells/dish)
were seeded in 3.5 cm dishes and cultured in a humidified
environment with 5% CO2 at 37°C for 24 h. Then,
different concentrations of scutellarein (10, 20, 40, 60 and 80
µM) were added into each dish and the cells were incubated
with scutellarein at 37°C for 15 days. Cells treated with DMSO were
used as the negative control. Subsequent to the incubation at 37°C
for 15 days, the cells were washed with phosphate buffered saline
(PBS) 3 times, following by staining with 1 ml crystal violet
solution for 10 min at room temperature. The cells were then washed
with PBS prior to the colonies being counted.
Reverse transcription-quantitative PCR
(RT-qPCR)
Colon cancer SW480 and T84 cells with different
treatments were obtained and mRNA was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. Subsequently,
reverse transcription (42°C for 30 min followed by 85°C for 5 min)
was performed using EasyScript Reverse Transcriptase (Beijing
Transgen Biotech Co., Ltd.) followed by qPCR on a DA7600 Real-time
Nucleic Acid Amplification Fluorescence Detection System (Bio-Rad
Laboratories, Inc.) in a 25 µl volume system using the
TransStart Green qPCR SuperMix (Beijing Transgen Biotech Co.,
Ltd.). The thermocycling conditions were as follows: Initial
denaturation for 3 min at 95°C, followed by 35 cycles of
denaturation at 95°C for 10 sec and annealing/extension at 55°C for
30 sec.
The primers were obtained from Sangon Biotech Co.,
Ltd. Melting-curve analysis was conducted to recognize the reaction
specificity. All experiments were performed in triplicate and the
data were analyzed using the 2−ΔΔCq method (30). Primers of RAGE, CDC4 and β-catenin
were as follows: RAGE forward, 5′-GGG GTA CCA AGG AAG CAG GTA GGC
AGC C-3′ and reverse, 5′-CCG CTC GAA TCC ATT CCT GTT CAT CTG C-3′;
CDC4 forward, 5′-GTG GGA CAT ACA GGT GGA-3′ and reverse, 5′-CAA CGC
ACA GTG GAA CTA-3′; β-actin forward, 5′-CAT GTA CGT TGCT ATC CAG
GC-3′ and reverse, 5′-CTC CTT AAT GTC ACG CAC GAT-3′.
Immunoprecipitation (IP)
An IP assay was performed using protein G plus
A-agarose beads as established, with CDC4 or RAGE antibodies
obtained from Santa Cruz Biotechnology, Inc. In brief, SW480 and
T84 cells were first rinsed with cold PBS and lysed in IP lysis
buffer (Thermo Fisher Scientific, Inc.); the total proteins were
obtained after being centrifuged at 20,000 × g for 30 min at 4°C
and served as the 'Input' sample. Then, the cell lysate containing
200 µg protein was incubated with Dynabeads®
protein G (Thermo Fisher Scientific, Inc.) for 1 h at room
temperature, and incubated with 2 µg of antibody against
RAGE (cat. no. sc365154; Santa Cruz Biotechnology, Inc.) or IgG
(cat. no. 5946, Cell Signaling Technology, Inc.; negative control)
overnight at 4°C, followed by incubation with Dynabeads®
protein G for another 1 h at room temperature to form the immune
complex. Then, the protein samples were loaded onto gels for
western blot analysis.
Cell transfection
T84 and SW480 cells (3×105 cells/well)
were seeded into 6-well plates and cultured with antibiotic-free
standard growth medium for 24 h until the confluence reached
60-80%. Subsequently, the cells were infected with the following
lentivirus vectors using 7 µg/ml polybrene (Hanbio
Biotechnology Co., Ltd.), including RAGE shRNA lentivirus vector
(cat. no. sc-36374-SH; Santa Cruz Biotechnology, Inc.), CDC4 shRNA
lentivirus vector (sh-CDC4; cat. no. sc-37547-SH; Santa Cruz
Biotechnology, Inc.) and the control shRNA plasmid-A (sh-NC; cat.
no. sc-108060; Santa Cruz Biotechnology, Inc.). In addition, the
RAGE clustered regularly interspaced short palindromic repeats
(CRISPR) Activation Plasmid (cat. no. sc-400284-ACT; Santa Cruz
Biotechnology, Inc.), and CDC4 CRISPR Activation Plasmid (cat. no.
sc-401257-ACT; Santa Cruz Biotechnology, Inc.) were used to
upregulate the expression of RAGE (OE-RAGE) and CDC4 (OE-CDC4) in
colon cancer, and were transfected into cells using Lipo3000 (cat.
no. L3000008; Thermo Fisher Scientific, Inc.) at 37°C for 48 h, in
addition to the Control CRISPR/dCas9 Activation Plasmid (OE-NC;
cat. no. sc-437275; Santa Cruz Biotechnology, Inc.). Subsequent to
48 h cell infection/transfection, the mRNA and protein samples were
extracted from the cells and submitted to RT-qPCR and western blot
analysis.
Flow cytometry
T84 and SW480 cells were seeded into 6-well plates
and cultured for 24 h. Following different treatments, the cells
were gathered into individual centrifuge tubes and suspended in
Annexin-binding buffer. Subsequently, the cells were incubated with
Annexin V-fluorescein isothiocyanate and propidium iodide solution
(BD Biosciences) at room temperature in the shade for 15 min,
according to the manufacturer's instructions. The apoptotic
percentages of all samples were analyzed using flow cytometry (BD
Biosciences). Cell apoptotic rates were analyzed using FlowJo v.10
(FlowJo, LLC).
Animal experiments
The function of scutellarein in inhibiting tumor
progression was assessed using a mouse model. A total of 30 male
BALB/c nude mice aged 5 weeks old (weight 18-20 g) were obtained
from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China)
and maintained under specific pathogen-free conditions at 20-26°C
with 55±5% humidity, a 12 h light/dark cycle and ad libitum
access to food and water. The experiment protocols were ethically
approved by the Institutional Animal Care and Use Committee of the
Affiliated Hospital of Southwest Medical University (Sichuan,
China). The T84 cells (5×106 in 200 µl PBS) were
tail-vein injected into the mice to generate a colon cancer-bearing
model. Once the tumors had grown, the mice were randomly separated
into 3 groups and intraperitoneally injected with scutellarein (0.5
µg/g body weight) and shRNA-CDC4 (0.1 µg/g body
weight). Mice treated with PBS alone were considered to be the
control group. The animal health and behavior, in addition to the
tumor sizes, were monitored every 3 days. A total of 20 days later,
the mice were euthanized by cervical dislocation and the heads were
broken and the tumor weight of each mouse was evaluated. Then, the
tumors were removed from the mice and subjected to
immunohistochemical staining with an anti-Ki-67 antibody (1:100
dilution; cat. no. 12202; Cell Signaling Technology, Inc.) and
hematoxylin (100%, 3 min) and eosin (0.5%, 30 sec) staining at room
temperature to assess the effect of scutellarein/CDC4 on the
expression of Ki-67 and pathological alteration. The diameter for
the maximum tumor was 1.8 cm and all mice were euthanized at the
end of the experiment.
For the immunohistochemical staining, tissue
sections were deparaffinized and rehydrated with xylene (100%) and
ethanol (100, 95, 80 and 70%). Subsequently, the antigens were
retrieved using citrate antigen retrieval solution at 95°C
(Beyotime Institute of Biotechnology) and blocked with 5% goat
serum (AmyJet Scientific, Inc.) diluted in TBS + 0.5% Tween-20 at
room temperature. The sections were probed with the primary
antibody against Ki-67 (1:100 dilution; cat. no. 12202; Cell
Signaling Technology, Inc.) at 4°C overnight. Following the primary
antibody incubation, sections were incubated with a HRP-conjugated
secondary antibody (1:500; cat. no. 8114; Cell Signaling
Technology, Inc.) at room temperature for 30 min, followed by
incubation with the chromogen 3,30-diaminobenzidine tetrachloride
(R&D Systems, Inc.) for 2-3 sec at room temperature. Cell
nuclei were stained with 1% Harris hematoxylin solution for 30 sec
at room temperature. The protein expression of Ki-67 was evaluated
by two pathologists on the base of the positive staining proportion
and the staining intensity, as previously described (31). The positive staining percentage
was scored as 0 for ≤5%, 1 for 6-25%, 2 for 26-50%, 3 for 51-75%
and 4 for >75%. Intensity was marked as follows: 0 represented
no staining, 1 represented weak staining, 2 represented moderate
staining and 3 represented strong staining. The final score was
obtained by multiplying the percentage score and intensity
score.
Statistical analysis
The results are representative of independent
experiments, and the data were presented as the mean ± standard
error of the mean. SPSS22.0 statistical software (IBM Corp.) was
used to analyze the data between groups with different treatments
using paired Student's t-tests for 2 groups and one way analysis of
variance followed by a Tukey's test for multiple groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Scutellarein treatment induces cell
apoptosis and inhibits cell viability in CL-40, T84 and SW480
cells
First, the present study investigated the effect of
scutellarein treatment on cell apoptosis and viability in colon
cancer. The CL-40, T84 and SW480 cells were exposed to gradient
concentrations of scutellarein, and the cytotoxic activity of
scutellarein was evaluated using an MTT assay. The IC50
values of scutellarein in CL-40, T84 and SW480 cell lines were
determined to be 40.9, 40 and 39.6 µM, respectively
(Fig. 1A, C and E). In addition,
the ratios of cleaved caspase3/total caspase3 and cleaved/total
caspase7 were significantly increased when the cells were treated
with scutellarein in a dose-dependent manner (Fig. S1A-C), in addition to the
apoptosis rates detected using a flow cytometry assay in CL-40, T84
and SW480 cell lines (Fig. 1B, D and
F) compared with the control group (P<0.05). Furthermore,
the function of scutellarein treatment in the clone formation
ability of CL-40, T84 and SW480 cells was examined using a colony
formation assay. As presented in Fig. S1D, compared with the control
group, the plates with scutellarein-treated cells demonstrated
fewer cell clones, and ≥40 µM scutellarein induced a
significant inhibition in the clone formation ability in a
dose-dependent manner compared with the 0 µM group
(P<0.05). These results illustrated that scutellarein treatment
was able to efficiently inhibit colon cancer cell viability and
induce cell apoptosis.
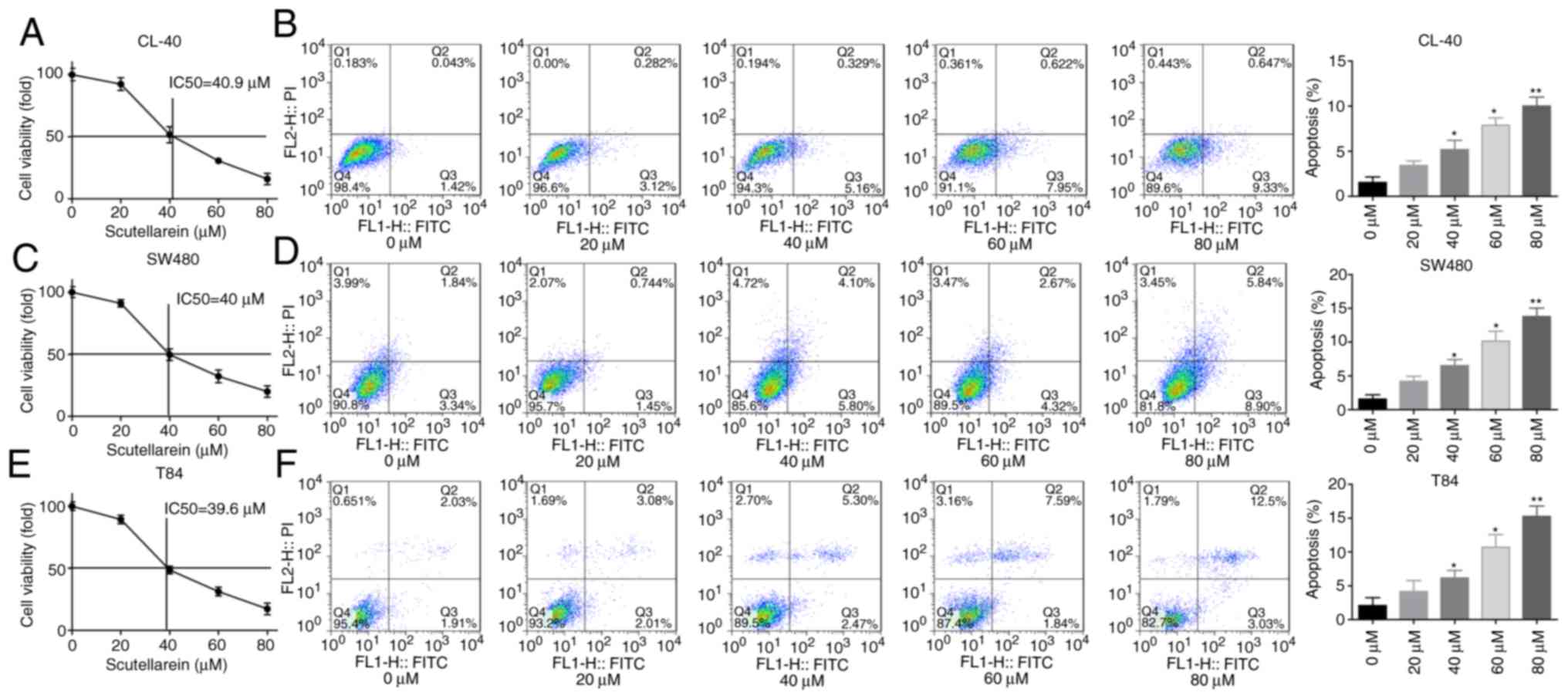 | Figure 1Scutellarein treatment inhibited cell
viability and promoted cell apoptosis in colon cancer cells (CL-40,
SW480 and T84). The cells were exposed to scutellarein (0, 20, 40,
60 and 80 µM) for 24 h, and then assays were performed. (A)
Cytotoxic effect of scutellarein on CL-40 cells were determined
using an MTT assay, and the IC50 value was determined to
be 40.9 µM. (B) Effect of scutellarein treatment on the
apoptosis rate of CL-40 cells was evaluated by using a flow
cytometry assay. (C) Cell viability of SW480 cells was determined
using an MTT assay, and the IC50 value was determined to
be 40 µM. (D) SW480 cell apoptosis was detected by using
flow cytometry technology. (E) Cytotoxic effect of scutellarein on
T84 cells was assessed using an MTT assay, and the IC50
value was determined to be 39.6 µM. (F) Cell apoptosis in
T84 cells was measured using a flow cytometry assay. The data were
represented as the mean ± standard error of the mean (n=3).
*P<0.05, **P<0.01 vs. the control
group. IC50, half maximal inhibitory concentration. |
Scutellarein inhibits cell proliferation
and promotes cell apoptosis via reducing RAGE expression in T84 and
SW480 cells
To reveal the mechanisms underlying scutellarein in
colon cancer inhibition, the present study then investigated the
effects of scutellarein on the expression of numerous oncoproteins,
including CD44, CD50, CD74, CD138, MCAM, CD151, CD166, CD206, RAGE
and Madcam1 (14,15). The results revealed that
scutellarein treatment (40 µM) significantly inhibited the
protein expression levels of RAGE compared with the control group
(P<0.05) in T84 (Fig. 2A) and
SW480 cells (Fig. 2B), with no
significant influence in the protein expression levels of CD44,
CD50, CD74, CD138, MCAM, CD151, CD166, CD206 and Madcam1
(P>0.05). However, scutellarein treatment exerted no significant
effect on the mRNA levels of RAGE compared with the control group
(P>0.05; Fig. 2C).
Furthermore, the present study revealed that the significantly
increased expression of RAGE induced by OE-RAGE transfection was
significantly reduced when T84 and SW480 cells were treated with
scutellarein (40 µM; P<0.05; Fig. 2D). These results demonstrated that
scutellarein treatment negatively regulated RAGE expression at the
protein level.
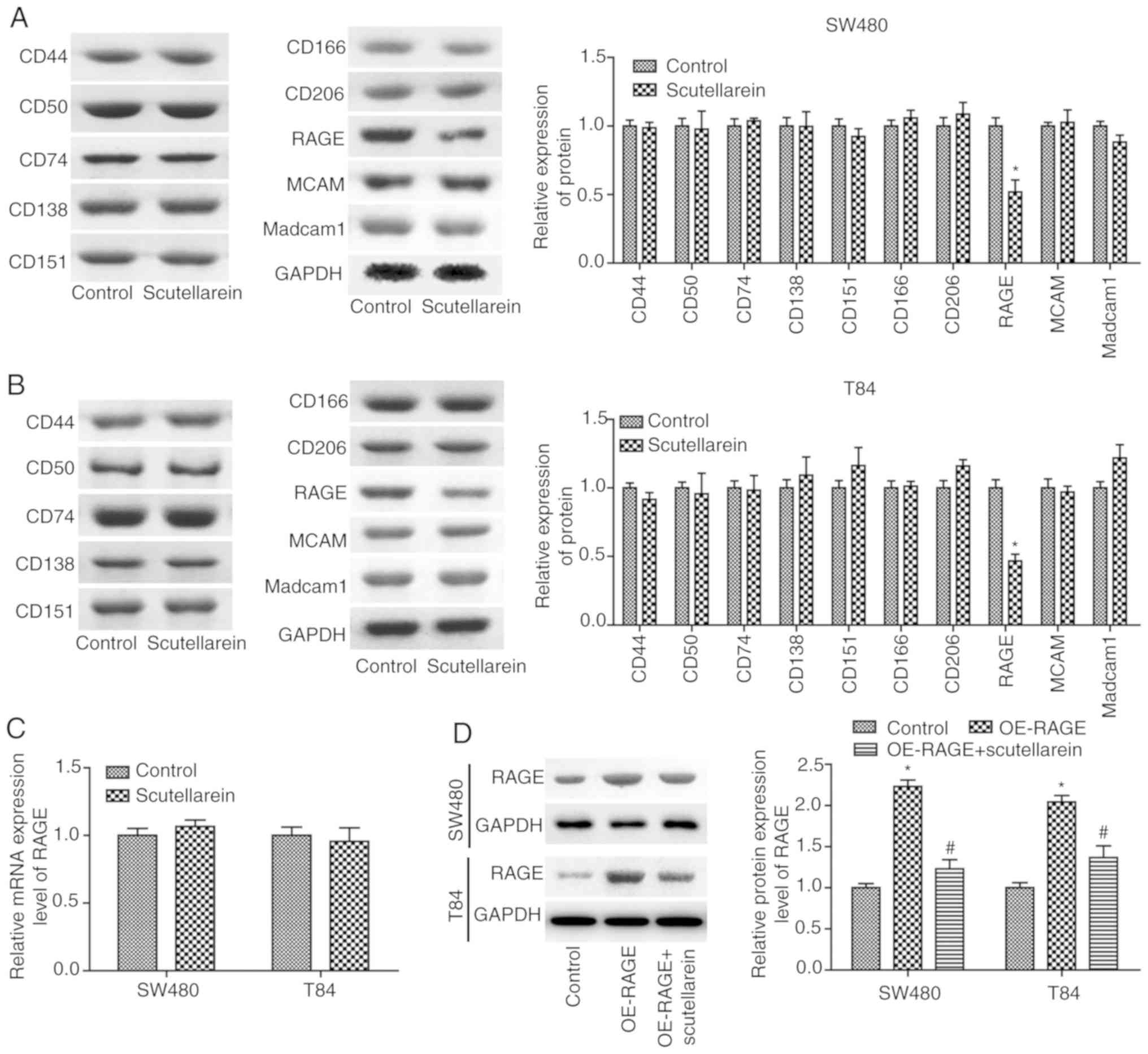 | Figure 2Scutellarein treatment decreased RAGE
protein levels in SW480 and T84 cells. Western blot analysis was
performed to determine the effects of scutella-rein on the
expression of CD44, CD50, CD74, CD138, MCAM, CD151, CD166, CD206,
RAGE and Madcam1 once the cells were treated with scutellarein (40
µM) in (A) SW480 cells and (B) T84 cells, and GAPDH was used
as a loading control. (C) mRNA expression levels of RAGE were
determined by using a reverse transcription-quantitative PCR assay
once the cells were treated with scutellarein (40 µM). (D)
Western blot analysis was performed to investigate the expression
of RAGE protein once cells were transfected with the OE-RAGE
plasmid together with scutellarein (40 µM) or not. The data
demonstrated are representative of three independent experiments.
n=3. *P<0.05 vs. the control group;
#P<0.05 vs. the OE-RAGE group. CD, cluster of
differentiation; MCAM, melanoma cell adhesion molecule; RAGE,
receptor for advanced glycation end products; Madcam1, mucosal
addressin cell adhesion molecule 1; OE, overexpression. |
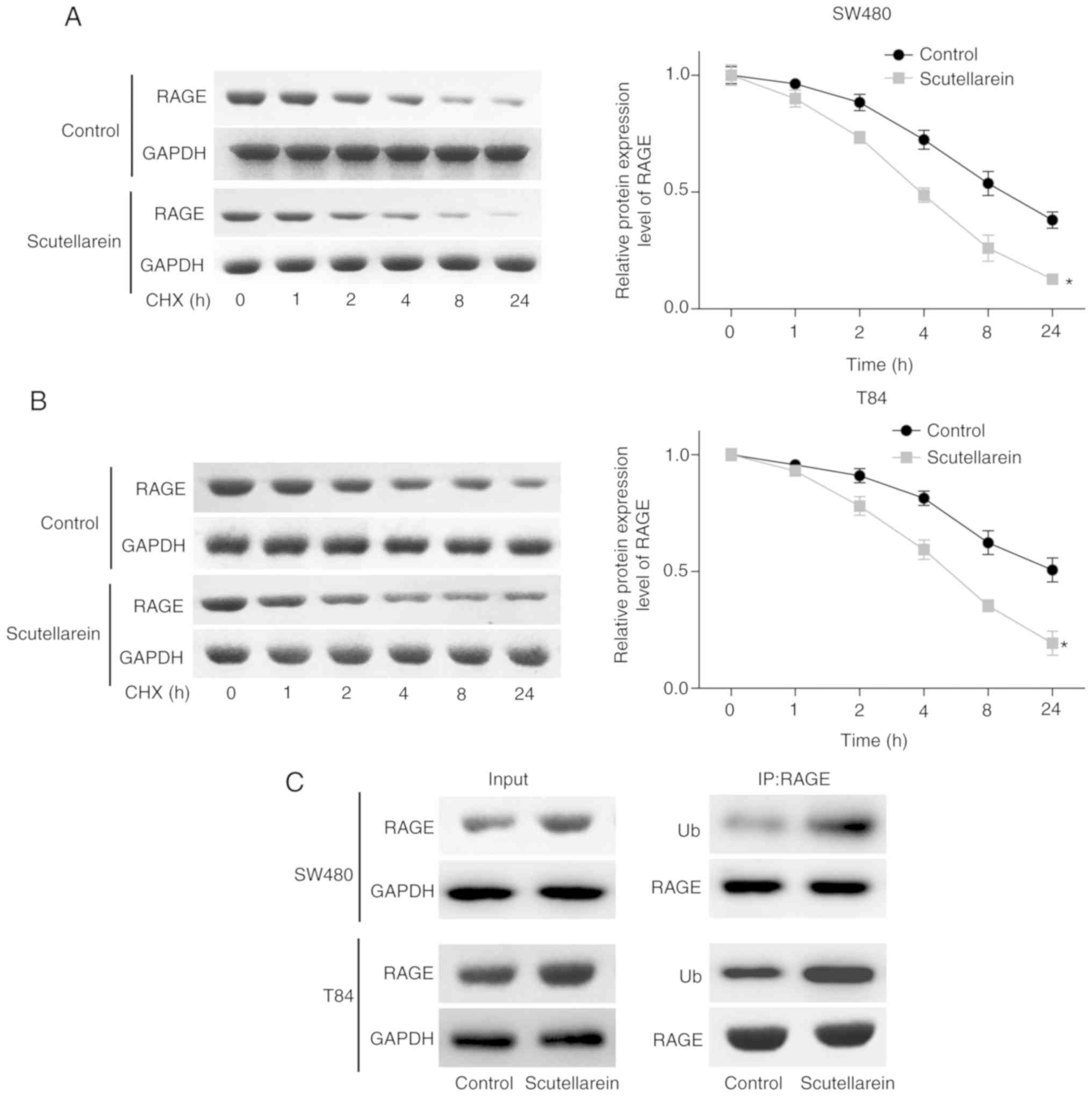
To reveal the underlying mechanism of the
scutellarein treatment-induced reduction in the RAGE protein
expression level, the present study then investigated the effect of
scutellarein treatment on the protein stability and ubiquitination
of RAGE. The CHX analysis demonstrated that scutellarein treatment
significantly reduced the stability of RAGE protein in SW480
(P<0.05; Fig. 3A) and T84 cell
lines compared with the control (P<0.05; Fig. 3B). In addition, scutellarein
treatment significantly enhanced the ubiquitination of the RAGE
protein (Fig. 3C). These results
revealed that scutellarein treatment may reduce RAGE protein
expression via enhancing its ubiquitination and impairing its
stability.
The present study then investigated the effects of
RAGE in scutellarein-mediated cell proliferation repression and
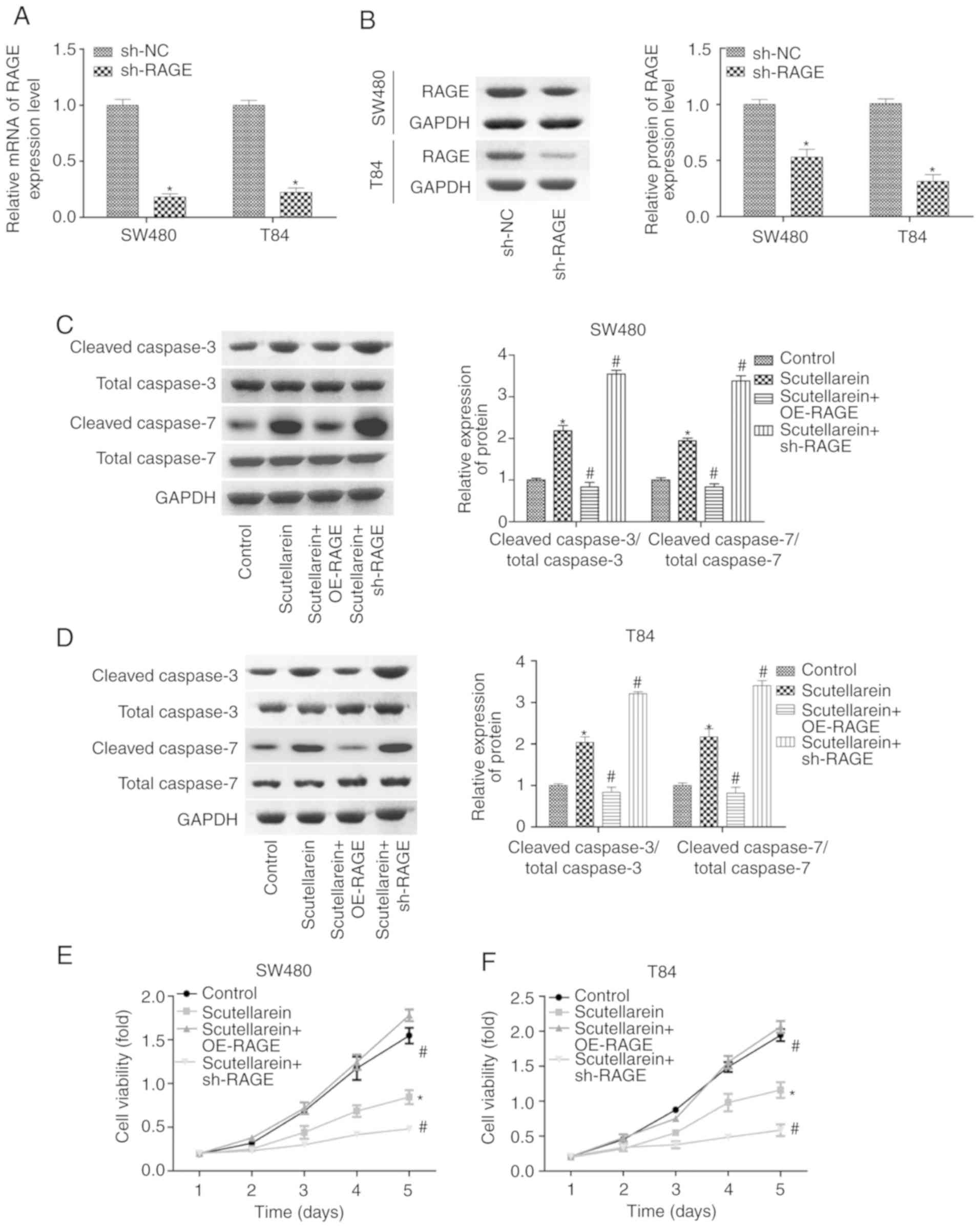
apoptosis promotion in T84 and SW480 cells. The expression levels
of RAGE were significantly reduced when the cells were infected
with shRNA targeting the RAGE gene at the mRNA and protein levels
when compared with the sh-NC group (P<0.05; Fig. 4A and B). As demonstrated in
Fig. 4C and D, the overexpression
of RAGE significantly blocked the function of scutellarein in
promoting the expression levels of cleaved caspase3/7, while the
knockdown of RAGE enhanced the function of scutellarein, with a
statistically significant difference compared with the
scutellarein-treated group (P<0.05; Fig. 4C and D). Additionally, the
proliferation of T84 and SW480 cells in the presence of
scutellarein with or without RAGE overexpression or knockdown was
also assessed. Compared with the control group, cell proliferation
was significantly repressed following scutellarein treatment, and
RAGE overexpression significantly weakened this function while RAGE
downregulation enhanced this function in SW480 and T84 cell lines
(P<0.05; Fig. 4E and F). In
addition, the present study detected the effect of the
scutellarein/RAGE axis on the activation of signals which have been
reported to be under the regulation of scutellarein, including
nuclear factor-κβ (32), PKC
(33) and VEGF (34). The results revealed that
scutellarein treatment significantly decreased the expression
levels of p-p65, PKC and VEGF compared with the control group
(P<0.05), whereas the expression levels of p-p65 and VEGF were
neutralized when RAGE was overexpressed, while RAGE downregulation
further decreased p-p65 and VEGF expression levels compared with
the scutellarein group (P<0.05; Fig. S2). However, the deregulation of
RAGE exhibited no significant influence in PKC expression in SW480
and T84 cell lines (P>0.05; Fig.
S2). These results demonstrated that scutellarein promoted
colon cancer cell apoptosis and repressed cell viability via
downregulating the expression of RAGE.
Scutellarein increases the combination of
CDC4 and RAGE
To investigate the mechanism of the suppressant
effects of scutellarein on RAGE levels, the present study detected
the expression of a series of ubiquitination-associated proteins,
including CDC4, RCHY1, MDM2, CBL and Smurf1 in T84 and SW480 cells
using western blot analysis. It was revealed that only the
expression levels of CDC4 were significantly increased in presence
of scutellarein compared with the control (P<0.05), with no
notable change in the expression levels of RCHY1, MDM2, CBL and
Smurf1 (Fig. 5A and C). As CDC4
was considered as one of the ubiquitin ligases, the increase of
CDC4 may have induced the degradation of downstream molecules. IP
was performed to investigate the combination of CDC4 and RAGE and
the results revealed that scutellarein enhanced the direct or
indirect combination of CDC4 and RAGE protein expression in SW480
cells (Fig. 5B) and T84 cells
(Fig. 5D). These results
indicated that CDC4 may serve a function in the
scutellarein-mediated reduction of RAGE expression.
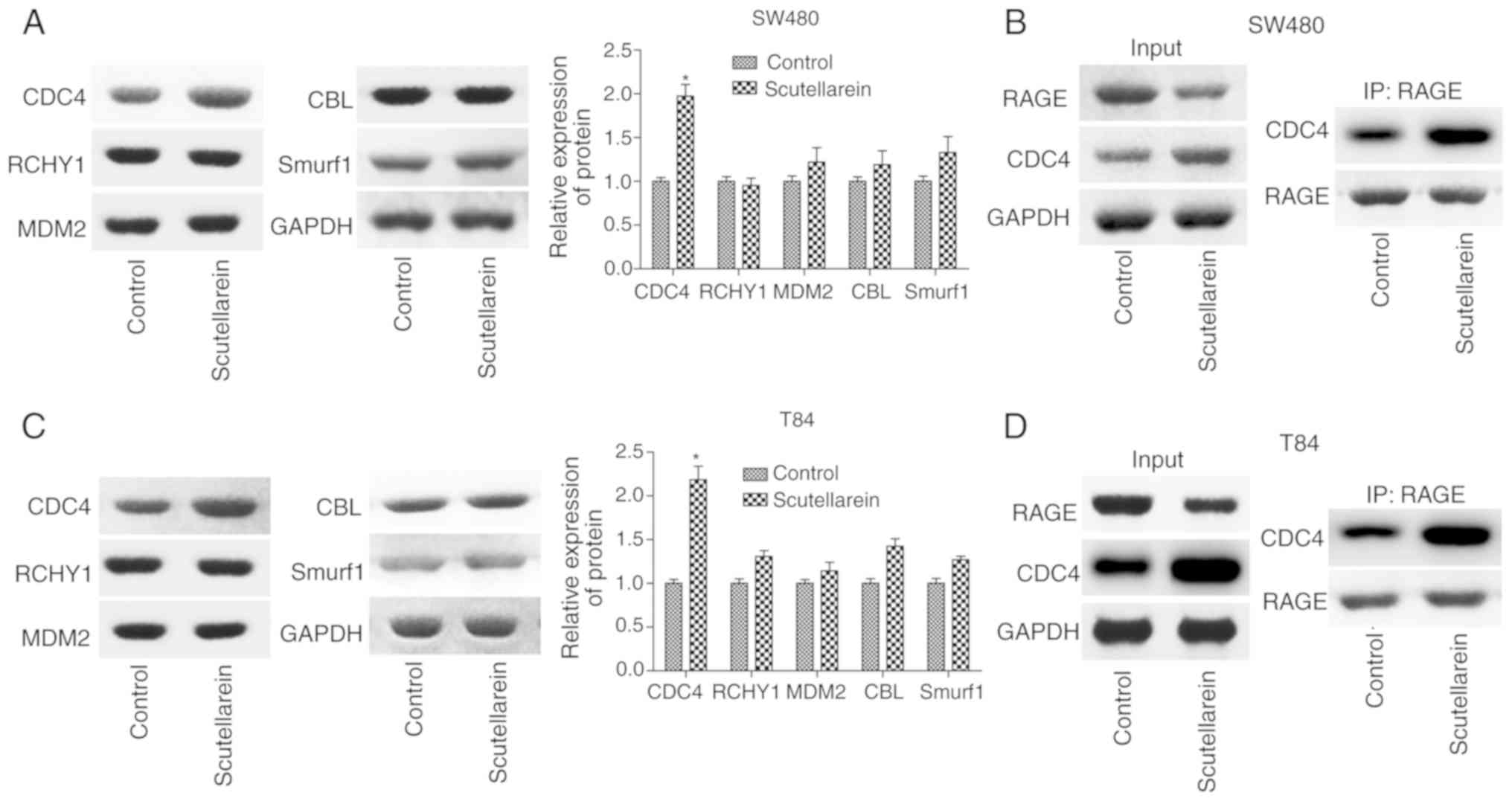 | Figure 5Effects of scutellarein on the
combination of CDC4 and RAGE proteins. (A) Western blot analysis
was performed to investigate the effects of scutellarein (40
µM) on the expression of ubiquitin-associated proteins,
including CDC4, RCHY1, MDM2, CBL and Smurf1 in SW480 cells. (B) An
IP assay was performed to detect the effects of scutellarein on the
combination of CDC4 and RAGE in SW480 cells. (C) Representative
data presenting the levels of ubiquitin-associated proteins (CDC4,
RCHY1, MDM2, CBL and Smurf1) in T84 cells treated with
scutellarein. (D) An IP assay was used to assess the effects of
scutellarein on the combination of CDC4 and RAGE proteins in T84
cells. n=3. *P<0.05 vs. the control group. RAGE,
receptor for advanced glycation end products; CDC4, cell division
control protein 4; IP, immunoprecipitation; RCHY1, ring finger and
CHY zinc finger domain containing 1; MDM2, MDM2 proto-oncogene;
CBL, Cbl proto-oncogene; Smurf1, SMAD specific ubiquitin protein
ligase 1. |
Scutellarein treatment inhibits cell
proliferation and promotes cell apoptosis via CDC4-mediated RAGE
downregulation
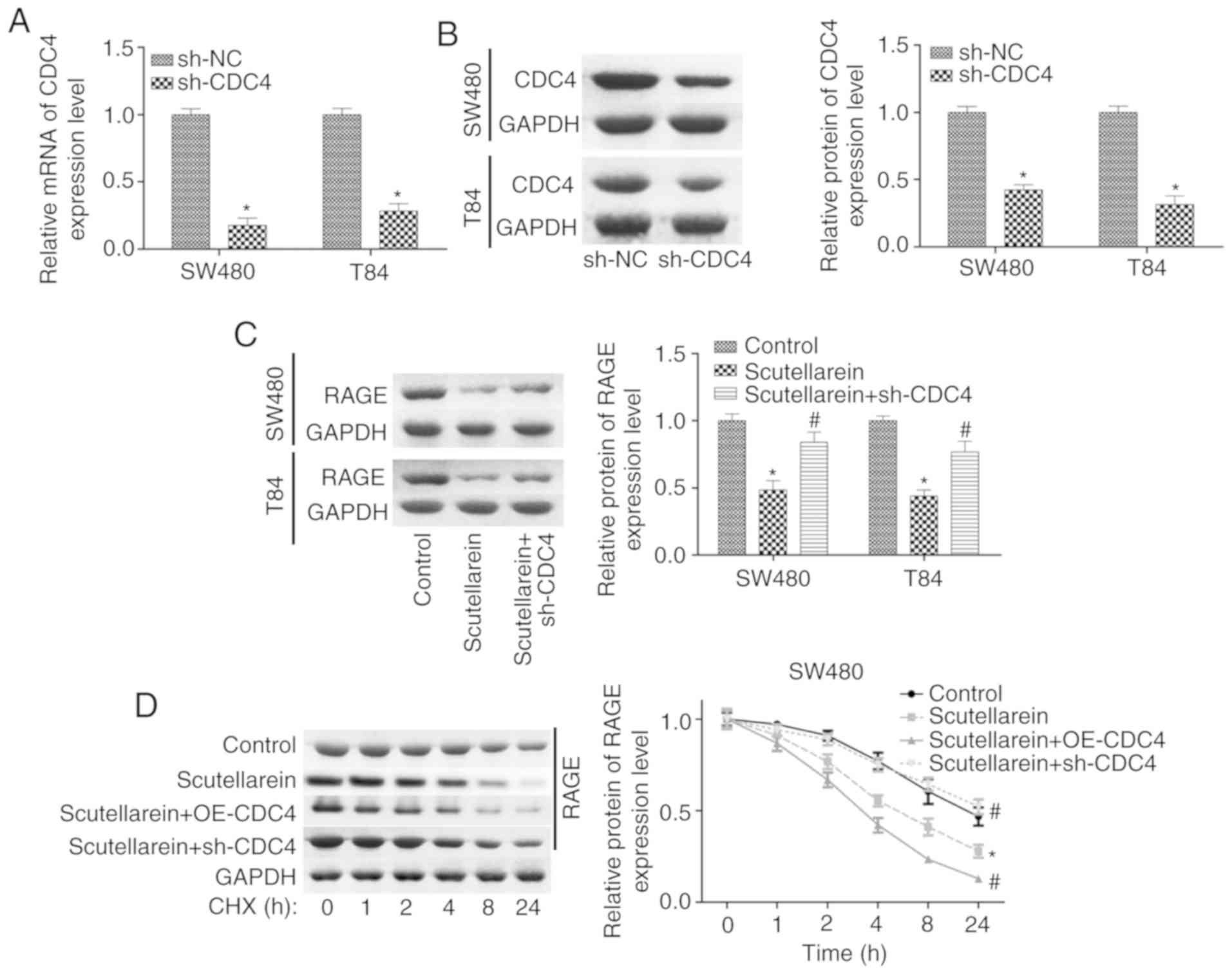
To further study the function CDC4 served in the
scutellarein-mediated reduction of RAGE expression and colon cancer
inhibition, the present study then performed CHX analysis to
investigate the effect of CDC4 on the stability of RAGE. The
results demonstrated that the overexpression of CDC4 significantly
accelerated the degradation of the RAGE protein in SW480 cells
(Fig. 6A) and T84 cells (Fig. 6B) compared with the control
(P<0.05). In addition, it was revealed that the over-expression
of CDC4 increased the expression of ubiquitinated RAGE compared
with the control (Fig. 6C). Cell
proliferation was significantly inhibited (P<0.05; Fig. 6D and E) while cell apoptosis
(P<0.05; Fig. 6F) was
significantly increased when CDC4 was overexpressed in SW480 and
T84 cells compared with the control, whereas these effects were
significantly abolished when RAGE was overexpressed at the same
time (P<0.05). These results indicated that CDC4 promoted colon
cancer cell apoptosis and inhibited cell proliferation via
increasing the RAGE ubiquitination levels.
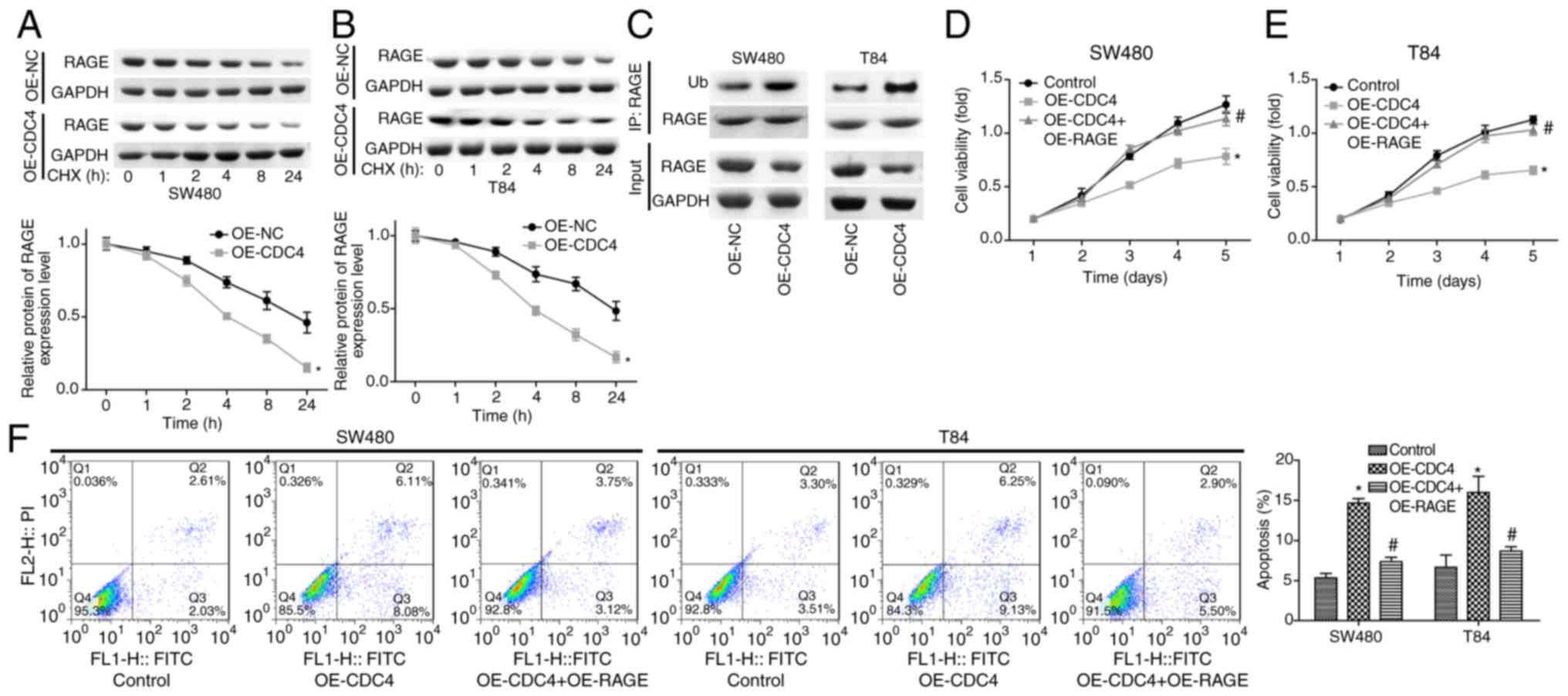 | Figure 6Effects of CDC4/RAGE on the
proliferation and apoptosis of SW480 and T84 cells. Following 24 h
of cell transfection with OE-NC or OE-CDC4, (A) SW480 and (B) T84
cells were treated with CHX (100 µg/ml) for 1, 2, 4, 8 or 24
h, and then a western blot assay was performed to detect the
protein levels of RAGE. (C) A western blot assay was performed to
investigate the ubiquitination of RAGE subsequent to SW480 and T84
cells being transfected with OE-CDC4 or OE-NC. Cell proliferation
of (D) SW480 and (E) T84 cells was detected using MTT analysis once
the cells were transfected with OE-CDC4, OE-CDC4+OE-PAGE or their
negative control vector. (F) Effects of CDC4 and RAGE
overexpression on the apoptosis of SW480 and T84 cells were
analyzed using flow cytometry. n=3. *P<0.05 vs.
control group; #P<0.05 vs. the OE-CDC4 group. RAGE,
receptor for advanced glycation end products; CDC4, cell division
control protein 4; OE, overexpression; NC, negative control; CHX,
cycloheximide. |
Furthermore, the present study investigated the
function of CDC4 in scutellarein-mediated proliferation inhibition
and apoptosis promotion in colon cancer. The expression of CDC4 was
significantly reduced when the cells were infected with sh-CDC4 at
the mRNA and protein levels compared with the negative control
(P<0.05; Fig. 7A and B).
Knockdown of CDC4 significantly rescued the reduction of RAGE in
SW480 and T84 cells mediated by scutellarein treatment (P<0.05;
Fig. 7C). CHX analysis revealed
that the overexpression of CDC4 significantly enhanced the effect
of scutellarein in reducing RAGE stability, while the knockdown of
CDC4 significantly weakened the effect of scutellarein (P<0.05;
Fig. 7D and E). Furthermore, it
was revealed that the increased expression levels of cleaved
caspase3/7 caused by scutellarein treatment were significantly
reduced when CDC4 was downregulated (P<0.05; Fig. 7F and G). In addition, the
knockdown of CDC4 significantly rescued scutellarein-mediated
reductions in the expression levels of p-p65 and VEGF (P<0.05),
with no notable change in the expression of PKC (P>0.05;
Fig. S3). These results
demonstrated that the increased level of CDC4 served a vital
function in scutellarein-mediated proliferation inhibition and
apoptosis promotion in colon cancer.
Scutellarein suppresses the progression
of colon cancer via increasing CDC4 expression in vivo
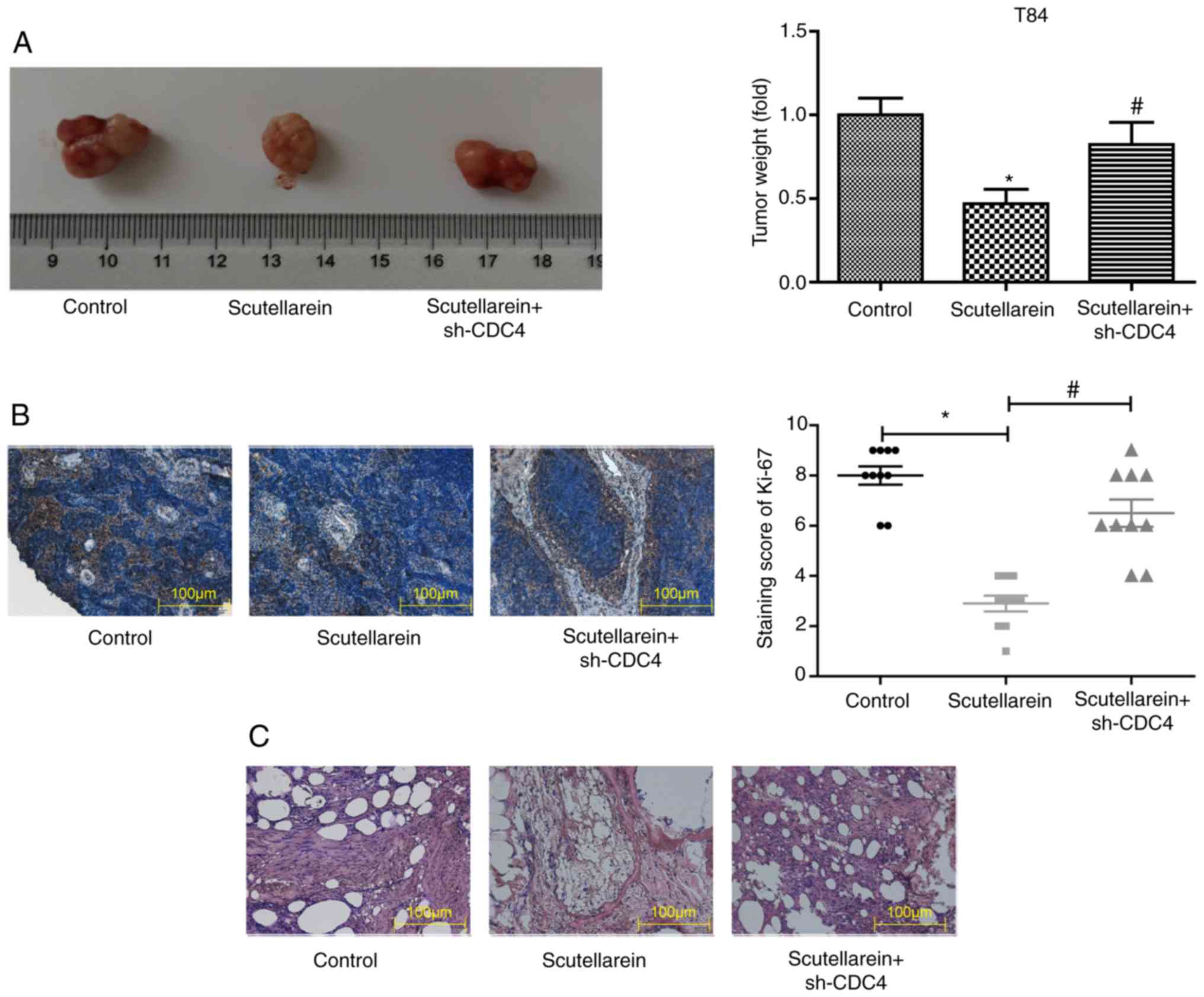
Next, the present study further investigated whether
scutellarein inhibited the progression of colon cancer in
vivo via upregulating CDC4 expression. A colon cancer-bearing
mice model was established by the injection of the colon cancer
cell line T84. The solid tumor types were removed and weighed once
the mice were sacrificed by cervical dislocation. In comparison
with the control group, scutellarein treatment significantly
reduced the tumor volume by nearly 50%, while the knockdown of CDC4
significantly increased the tumor volume compared with the
scutellarein alone treated group (P<0.05; Fig. 8A). In addition, the knockdown of
CDC4 also significantly neutralized the functions of scutellarein
treatment in reducing the expression levels of Ki-67 compared with
the control (P<0.05; Fig. 8B)
and the alleviation in the histological change of tumor tissues
(Fig. 8C). These results revealed
that scutellarein suppressed the in vivo tumor formation of
colon cancer via increasing CDC4 expression.
Discussion
As is well known, colon cancer is one of the most
malignant carcinoma types (35).
Wild chrysanthemum is the flower head of Chrysanthemum indicum
L., which may be obtained wildly in the majority of natural
habitats in China (5,36). The present study was designed to
investigate the effects and mechanism of scutellarein in the
development of colon cancer. The present study demonstrated that
scutellarin, derived from wild chrysanthemum, notably inhibited the
development of colon cancer in vitro and in vivo via
CDC4-mediated RAGE ubiquitination enhancement.
As early as 2005, Goh et al (13) reported that the chemically
standardized extract from Scutellaria barbata induced a
significant increase in the sub G1 phase and promoted cell
apoptosis in colon cancer LoVo cells. Subsequently, a study by Wang
et al (37) also
demonstrated the anti-tumor activity of scutellarein in colon
cancer. Recently, Guo et al (12) demonstrated that scutellarein
identified from Scutellaria barbata significantly increased
the apoptosis of colon cancer HCT116 cells via increasing the
production of intracellular reactive oxygen species. As CL-40, T84
and SW480 cell lines are from different types of colon cancer, in
that CL-40 cells are derived from the colon adenocarcinoma, T84
cells are derived from the lung metastatic site and SW480 cells are
derived from the primary site of colon cancer of Dukes' type B,
they were selected for the present study to investigate the effects
of scutellarein in the viability and apoptosis of colon cancer
cells. Consistent with previous results (12,13,37), the present study also observed
that scutellarein treatment significantly inhibited colony
formation and cell proliferation and accelerated cell apoptosis in
colon cancer CL-20, SW480 and T84 cells (P<0.05). In addition,
it was demonstrated that scutellarein significantly inhibited the
in vivo tumor formation ability of T84 cells using colon
cancer-bearing mice (P<0.05). The present results further
illustrate the inhibitory function of scutellarein in colon cancer
progression.
To investigate the underlying mechanism of
scutellarein-mediated repression in colon cancer, the present study
investigated the effects of scutellarein treatment on the
expression of numerous oncoproteins, including CD44, CD50, CD74,
CD138, MCAM, CD151, CD166, CD206, RAGE and Madcam1. As CL-40 and
SW480 are primary cell lines of colon cancer, one was selected
(SW480) together with T84 (a metastasis cell line) for the
subsequent experiments. The present results demonstrate that
scutellarein treatment significantly inhibited the expression of
RAGE compared with the control (P<0.05), with no substantial
influence in the expression levels of CD44, CD50, CD74, CD138,
MCAM, CD151, CD166, CD206 and Madcam1. An increasing number of
studies have demonstrated the importance of RAGE in the
pathogenesis of multiple human disease types, including cancer
(38-41). The deregulation of RAGE is
considered to be an important factor in tumorigenesis, and the
increase of RAGE is associated with a diverse range of malignancies
(22,42). In the present study, the results
demonstrated that scutellarein treatment significantly decreased
the protein levels of RAGE, but not the mRNA levels, compared with
the control (P<0.05). Thus, the present study investigated the
effect of scutellarein on RAGE protein stability and
ubiquitination, and the results revealed that scutellarein
treatment decreased the protein stability of RAGE and enhanced its
ubiquitination. Similarly, RAGE was also reported to be regulated
by the ubiquitination-degradation pathway by the ubiquitin E3
ligase subunit F-box protein O10 (43). Furthermore, the present study
observed that the overexpression of RAGE weakened while the
knockdown of RAGE enhanced scutellarein functions in promoting cell
apoptosis and inhibiting cell proliferation in colon cancer,
suggesting that scutellarein inhibited colon cancer progression via
downregulating RAGE.
Ubiquitination is a process in which ubiquitin
alters and specifically modifies the target proteins with the help
of a series of particular enzymes, including ubiquitin activating
enzyme, ubiquitin conjugating enzyme and ubiquitin ligase (44). An increasing number of studies
have demonstrated that ubiquitination is strongly implicated in the
pathogenesis of carcinogenesis (45,46). In the present study, the
underlying mechanism of the scutellarein-mediated ubiquitination of
RAGE protein was examined. An increase in CDC4 expression was
observed in the presence of scutellarein. CDC4/F-box and WD repeat
domain containing 7, located on the 4q, is considered to be a vital
anti-oncogene in various cancer types, in addition to a potential
therapeutic target, with mutations observed in pancreatic,
colorectal and ovarian tumor types (47). The results of the present study
confirmed that CDC4 may combine with the RAGE protein and then
induce its ubiquitination and the subsequent degradation.
Furthermore, a significant decrease in cell proliferation and an
increase in cell apoptosis rates were observed in SW480 and T84
cells with CDC4 overexpression compared with the control
(P<0.05), while this effect was impaired when RAGE was
downregulated, suggesting that CDC4 inhibited colon cancer
progression via decreasing the expression of RAGE. Furthermore, the
present results also revealed the important function of CDC4 in
scutellarein-mediated increases in cell apoptosis and inhibition in
cell proliferation and tumorigenesis in colon cancer.
However, there are a number of limitations to the
present study. In the present study it was mainly revealed that
scutellarein inhibited the expression of the oncoprotein RAGE via
increasing CDC4-mediated ubiquitination enhancement, which then
inhibited cell proliferation and induced cell apoptosis. However,
the present study did not investigate the molecular mechanism
underlying the scutellarein-mediated CDC4 upregulation. In
addition, the results revealed that scutellarein inhibited cell
proliferation and induced cell apoptosis in T84 and SW480 cells in
a similar manner. However, T84 cells are derived from a lung
metastatic site, while SW480 cells are derived from a primary site
of colon cancer of Dukes' type B, indicating that these are two
different types of cells with different malignant behaviors. It was
hypothesized that this may be induced by the same protein which is
located at the upstream of CDC4, but this was not further
investigated. These will be investigated in future studies through
performing sequencing to determine the gene which is modulated by
scutellarein in T84 and SW480 cell lines. Furthermore, the present
study did not examine the effect of scutellarein on the function of
CL-40 cells due to the limitations in funding and time. The present
study only examined the effects and underlying mechanisms of
scutellarein, an active ingredient of wild chrysanthemum, on the
progression of colon cancer. However, it is not clear whether
chamomile and other chrysanthemums have similar effects. In future
studies, the effects of chamomile and other chrysanthemums on colon
cancer progression will be examined.
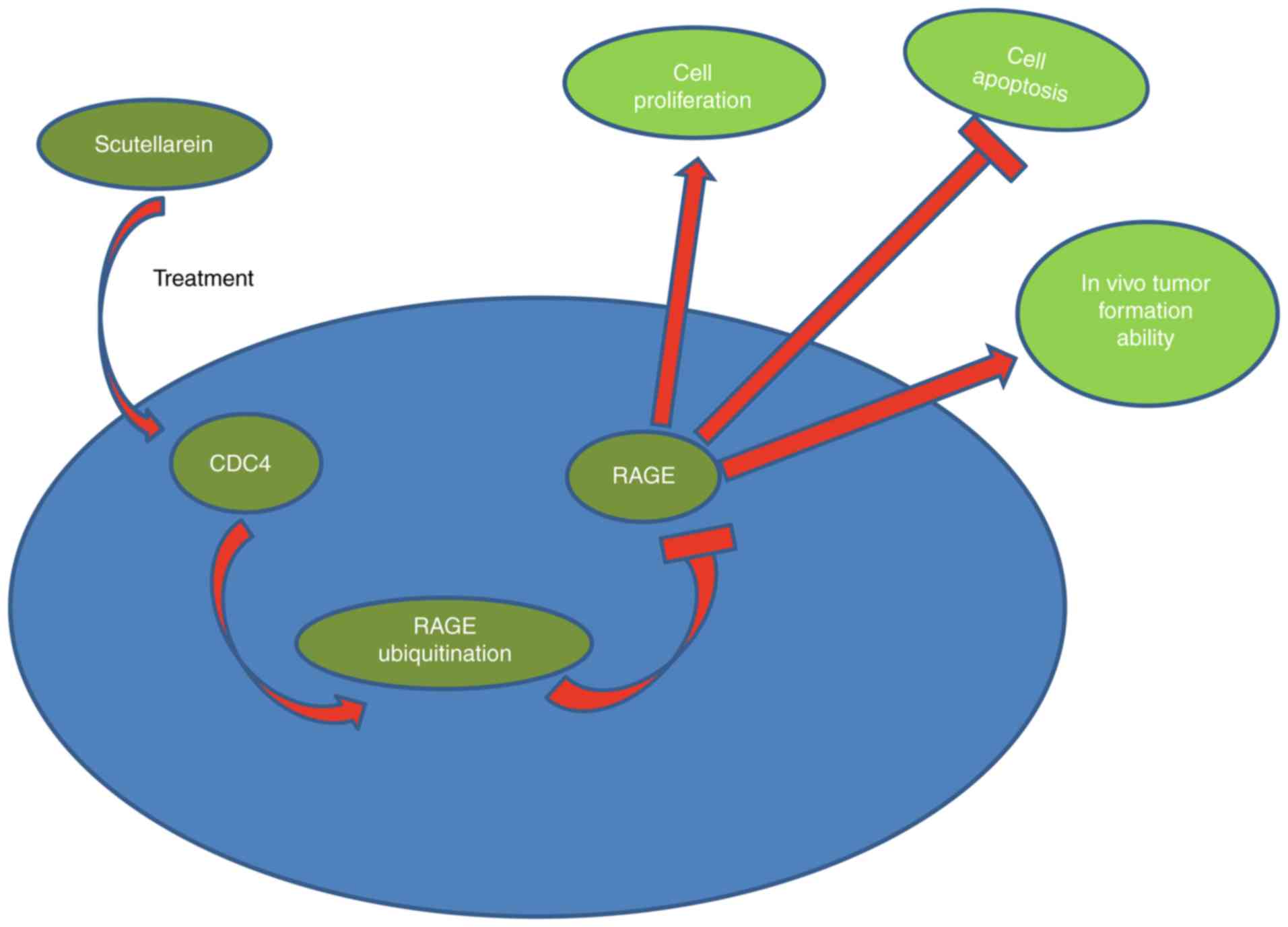
In conclusion, the present investigation revealed
that scutellarein derived from wild chrysanthemum may effectively
suppress the proliferation and tumorigenesis and induce the
apoptosis of colon cancer cells through downregulating RAGE
expression mediated by CDC4 upregulation (Fig. 9). The present results may provide
more useful information for the application of scutellarein in the
treatment of colon cancer.
Supplementary Data
Funding
The present study was supported by a research grant
for Doctors of the Affiliated Hospital of Southwest Medical
University and Luzhou People's Government-Southwest Medical
University Cooperative Scientific Research Project (grant no.
2019LZXNDJ26).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL and JW provided the idea and performed the
experiments, in addition to writing the manuscript. SZ and JL
performed the data analyses and parts of the experiments. WD
performed parts of the data analyses. YL revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Athymic nude mouse assays were performed in
accordance with the institutional principles for the concern and
use of animals and the protocol was ethically approved by the
ethical committee of the Affiliated Hospital of Southwest Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Labianca R, Beretta GD, Kildani B, Milesi
L, Merlin F, Mosconi S, Pessi MA, Prochilo T, Quadri A, Gatta G, et
al: Colon cancer. Crit Rev Oncol Hematol. 74:106–133. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Freeman HJ: Early stage colon cancer.
World J Gastroenterol. 19:8468–8473. 2013. View Article : Google Scholar :
|
|
3
|
Orangio GR: The economics of colon cancer.
Surg Oncol Clin N Am. 27:327–347. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Y, Shi J, Huang H, Ren J, Li N and
Dai M: Burden of colorectal cancer in China. Zhonghua Liu Xing Bing
Xue Za Zhi. 36:709–714. 2015.In Chinese. PubMed/NCBI
|
|
5
|
Sun S, Jiang P, Su W, Xiang Y, Li J, Zeng
L and Yang S: Wild chrysanthemum extract prevents UVB
radiation-induced acute cell death and photoaging. Cytotechnology.
68:229–240. 2016. View Article : Google Scholar :
|
|
6
|
Yang WS, Kim D, Yi YS, Kim JH, Jeong HY,
Hwang K, Kim JH, Park J and Cho JY: AKT-targeted anti-inflammatory
activity of the methanol extract of Chrysanthemum indicum var.
albescens J Ethnopharmacol. 201:82–90. 2017. View Article : Google Scholar
|
|
7
|
Deng Y, Jiang J, Chen S, Teng N, Song A,
Guan Z, Fang W and Chen F: Combination of multiple resistance
traits from wild relative species in Chrysanthemum via trigeneric
hybridization. PLoS One. 7:302012. View Article : Google Scholar
|
|
8
|
Hassanpouraghdam MB: Flowerhead volatile
oil composition of soilless culture-grown Chrysanthemum balsamita
L. Nat Prod Res. 23:672–677. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi X, Chen G and Liu X, Qiu Y, Yang S,
Zhang Y, Fang X, Zhang C and Liu X: Scutellarein inhibits cancer
cell metastasis in vitro and attenuates the development of
fibrosarcom in vivo. Int J Mol Med. 35:31–38. 2015. View Article : Google Scholar
|
|
10
|
Cheng CY, Hu CC, Yang HJ, Lee MC and Kao
ES: Inhibitory effects of scutellarein on proliferation of human
lung cancer A549 cells through ERK and NFκB mediated by the EGFR
pathway. Chin J Physiol. 57:182–187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parajuli P, Joshee N, Rimando AM, Mittal S
and Yadav AK: In vitro antitumor mechanisms of various Scutellaria
extracts and constituent flavonoids. Planta Med. 75:41–48. 2009.
View Article : Google Scholar
|
|
12
|
Guo F, Yang F and Zhu YH: Scutellarein
from Scutellaria barbata induces apoptosis of human colon cancer
HCT116 cells through the ROS-mediated mitochondria-dependent
pathway. Nat Prod Res. 33:2372–2375. 2019. View Article : Google Scholar
|
|
13
|
Goh D, Lee YH and Ong ES: Inhibitory
effects of a chemically standardized extract from Scutellaria
barbata in human colon cancer cell lines, LoVo. J Agric Food Chem.
53:8197–8204. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Behzad MM, Asnafi AA, Jaseb K, Jalali Far
MA and Saki N: Expression of CD markers' in immune thrombocytopenic
purpura: Prognostic approaches. APMIS. 125:1042–1055. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bacher P and Scheffold A: Flow-cytometric
analysis of rare antigen-specific T cells. Cytometry A. 83:692–701.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tripathi SC, Fahrmann JF, Celiktas M,
Aguilar M, Marini KD, Jolly MK, Katayama H, Wang H, Murage EN,
Dennison JB, et al: MCAM mediates chemoresistance in small-cell
lung cancer via the PI3K/AKT/SOX2 signaling pathway. Cancer Res.
77:4414–4425. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z and Yan X: CD146, a
multi-functional molecule beyond adhesion. Cancer Lett.
330:150–162. 2013. View Article : Google Scholar
|
|
18
|
Zhang H, Zhang J, Wang Z, Lu D, Feng J,
Yang D, Chen X and Yan X: CD146 is a potential marker for the
diagnosis of malignancy in cervical and endometrial cancer. Oncol
Lett. 5:1189–1194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tanida S, Mizoshita T, Mizushima T, Sasaki
M, Shimura T, Kamiya T, Kataoka H and Joh T: Involvement of
oxidative stress and mucosal addressin cell adhesion molecule-1
(MAdCAM-1) in inflammatory bowel disease. J Clin Biochem Nutr.
48:112–116. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Truffi M, Colombo M, Peñaranda-Avila J,
Sorrentino L, Colombo F, Monieri M, Collico V, Zerbi P, Longhi E,
Allevi R, et al: Nano-targeting of mucosal addressin cell adhesion
molecule-1 identifies bowel inflammation foci in murine model.
Nanomedicine (Lond). 12:1547–1560. 2017. View Article : Google Scholar
|
|
21
|
Logsdon CD, Fuentes MK, Huang EH and
Arumugam T: RAGE and RAGE ligands in cancer. Curr Mol Med.
7:777–789. 2007. View Article : Google Scholar
|
|
22
|
Palanissami G and Paul SFD: RAGE and Its
ligands: Molecular interplay between glycation, inflammation, and
hallmarks of cancer-a review. Horm Cancer. 9:295–325. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fuentes MK, Nigavekar SS, Arumugam T,
Logsdon CD, Schmidt AM, Park JC and Huang EH: RAGE activation by
S100P in colon cancer stimulates growth, migration, and cell
signaling pathways. Dis Colon Rectum. 50:1230–1240. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Foot N, Henshall T and Kumar S:
Ubiquitination and the regulation of membrane proteins. Physiol
Rev. 97:253–281. 2017. View Article : Google Scholar
|
|
25
|
Swatek KN and Komander D: Ubiquitin
modifications. Cell Res. 26:399–422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou MJ, Chen FZ and Chen HC:
Ubiquitination involved enzymes and cancer. Med Oncol. 31:932014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin X, Yang C, Fan P, Xiao J, Zhang W,
Zhan S, Liu T, Wang D and Wu H: CDK5/FBW7-dependent ubiquitination
and degradation of EZH2 inhibits pancreatic cancer cell migration
and invasion. J Biol Chem. 292:6269–6280. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gallo LH, Ko J and Donoghue DJ: The
importance of regulatory ubiquitination in cancer and metastasis.
Cell Cycle. 16:634–648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan C, Su H, Song X, Cao H, Kong L and Cui
W: Smad ubiquitination regulatory factor 1 (Smurf1) promotes
thyroid cancer cell proliferation and migration via
ubiquitin-dependent degradation of kisspeptin-1. Cell Physiol
Biochem. 49:2047–2059. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Xin B, He X, Wang J, Cai J, Wei W, Zhang T
and Shen X: Nerve growth factor regulates CD133 function to promote
tumor cell migration and invasion via activating ERK1/2 signaling
in pancreatic cancer. Pancreatology. 16:1005–1014. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang XW, Xu Y, Sui X, Lin H, Xu JM, Han
D, Ye DD, Lv GF, Liu YX, Qu XB and Duan MH: Scutellarein suppresses
Aβ-induced memory impairment via inhibition of the NF-κB pathway in
vivo an in vitro. Oncol Lett. 17:5581–5589. 2019.PubMed/NCBI
|
|
33
|
Tian X, Chang L, Ma G, Wang T, Lv M, Wang
Z, Chen L, Wang Y, Gao X and Zhu Y: Delineation of platelet
activation pathway of scutellarein revealed its intracellular
target as protein kinase C. Biol Pharm Bull. 39:181–191. 2016.
View Article : Google Scholar
|
|
34
|
Thirusangu P, Vigneshwaran V, Vijay Avin
BR, Rakesh H, Vikas HM and Prabhakar BT: Scutellarein antagonizes
the tumorigenesis by modulating cytokine VEGF mediated
neoangiogenesis and DFF-40 actuated nucleosomal degradation.
Biochem Biophys Res Commun. 484:85–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Labianca R, Nordlinger B, Beretta GD,
Mosconi S, Mandalà M, Cervantes A and Arnold D; ESMO Guidelines
Working Group: Early colon cancer: ESMO clinical practice
guidelines for diagnosis, treatment and follow-up. Ann Oncol.
24(Suppl 6): vi64–vi72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bosman FT and Yan P: Molecular pathology
of colon cancer. Pol J Pathol. 65(4 Suppl 1): S1–S11. 2014.In
Polish.
|
|
37
|
Wang F, Yang B, Zhao Y, Liao X, Gao C,
Jiang R, Han B, Yang J, Liu M and Zhou R: Host-guest inclusion
system of scutellarein with 2-hydroxypropyl-beta-cyclodextrin:
Preparation, characterization, and anticancer activity. J Biomater
Sci Polym Ed. 25:594–607. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van Zoelen MA, Achouiti A and van der Poll
T: RAGE during infectious diseases. Front Biosci (Schol Ed).
3:1119–1132. 2011. View
Article : Google Scholar
|
|
39
|
Lee TW, Kao YH, Chen YJ, Chao TF and Lee
TI: Therapeutic potential of vitamin D in AGE/RAGE-related
cardiovascular diseases. Cell Mol Life Sci. 76:4103–4115. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Azizan N, Suter MA, Liu Y and Logsdon CD:
RAGE maintains high levels of NFκB and oncogenic Kras activity in
pancreatic cancer. Biochem Biophys Res Commun. 493:592–597. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rahimi F, Karimi J, Goodarzi MT, Saidijam
M, Khodadadi I, Razavi AN and Nankali M: Overexpression of receptor
for advanced glycation end products (RAGE) in ovarian cancer.
Cancer Biomark. 18:61–68. 2017. View Article : Google Scholar
|
|
42
|
Tesarova P, Cabinakova M, Mikulova V, Zima
T and Kalousova M: RAGE and its ligands in cancer-culprits,
biomarkers, or therapeutic targets? Neoplasma. 62:353–364. 2015.
View Article : Google Scholar
|
|
43
|
Evankovich J, Lear T, McKelvey A, Dunn S,
Londino J, Liu Y, Chen BB and Mallampalli RK: Receptor for advanced
glycation end products is targeted by FBXO10 for ubiquitination and
degradation. FASEB J. 31:3894–3903. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rittinger K and Ikeda F: Linear ubiquitin
chains: Enzymes, mechanisms and biology. Open Biol. 7:pii: 170026.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ao N, Chen Q and Liu G: The small
molecules targeting ubiquitin-proteasome system for cancer therapy.
Comb Chem High Throughput Screen. 20:403–413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ding F, Xiao H, Wang M, Xie X and Hu F:
The role of the ubiquitin-proteasome pathway in cancer development
and treatment. Front Biosci (Landmark Ed). 19:886–895. 2014.
View Article : Google Scholar
|
|
47
|
Davis H and Tomlinson I: CDC4/FBXW7 and
the 'just enough' model of tumourigenesis. J Pathol. 227:131–135.
2012. View Article : Google Scholar : PubMed/NCBI
|