The extracellular matrix (ECM) serves a significant
role in modulating tissue genesis and remodeling not only by
connecting cells and providing support for them, but also by
regulating connections between the cells, and between the cell and
the matrix, inducing cell adhesion, motility and differentiation.
In the cardiovascular system, the ECM participates in maintaining
the structural continuity of the heart and vessels, providing
physical support for cell adhesion, controlling cell growth and
death, and regulating diastolic stiffness, as well as tissue repair
or remodeling to the cardiovascular damage (1). A number of these functions are
performed by a group of non-structural ECM proteins called
matricellular proteins, which includes thrombospondins (TSPs),
tenascins, periostin, osteopontin, CCN proteins and osteonectin
(2).
As a family of matricellular proteins, TSPs may be
secreted by various types of cells. A total of 5 members of the TSP
family (TSP1-5) have been identified so far, and are divided into
two subgroups, subgroup A and subgroup B, according to their
structural differences. Subgroup A contains TSP-1 and TSP-2, which
are trimeric and similar in structure, and subgroup B consists of
TSP-3, TSP-4 and TSP-5, which are pentameric and smaller compared
with those in subgroup A (3).
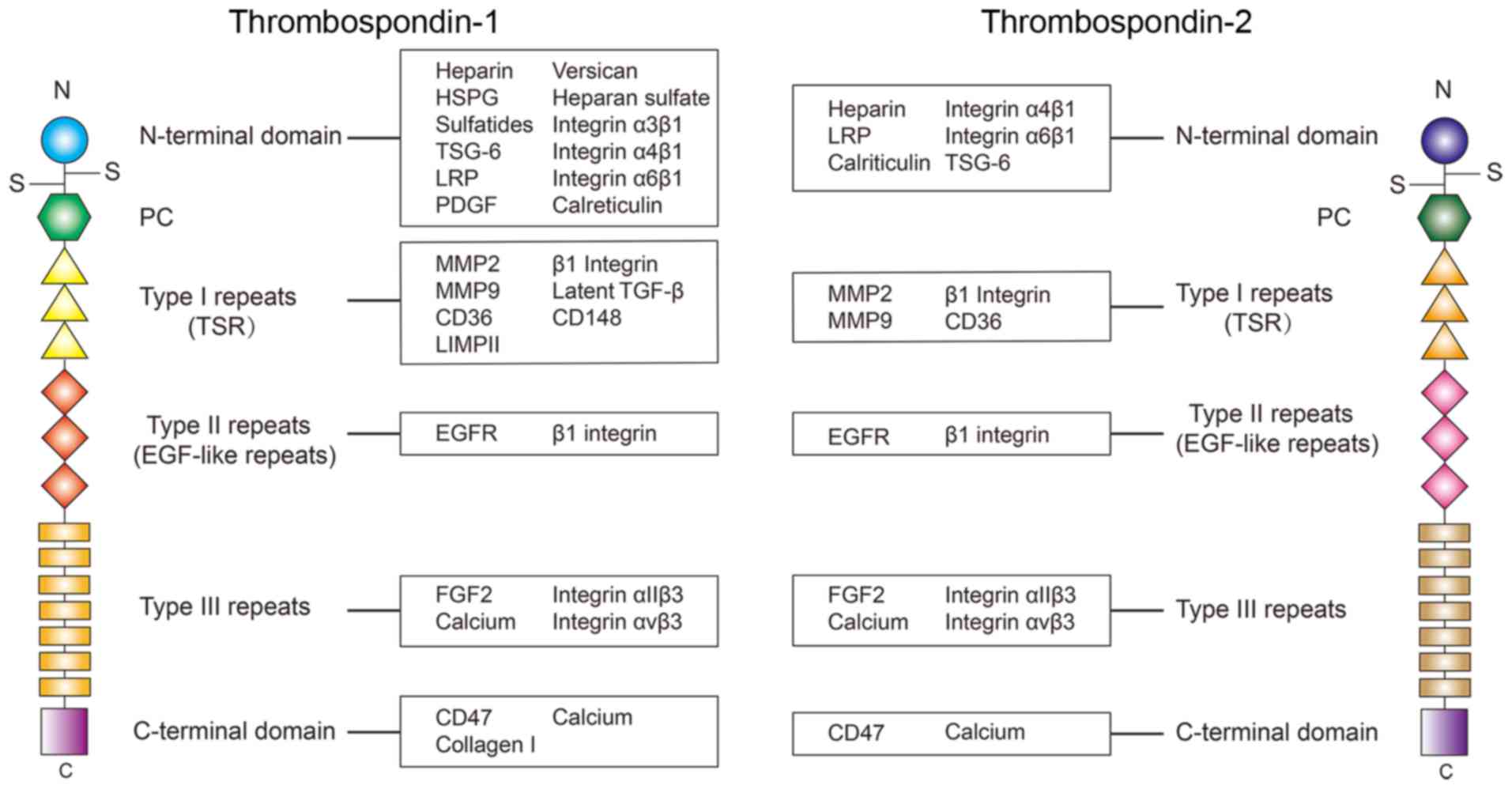
TSP-1 and TSP-2 are the most studied thrombospondins. In the
present review, the structure and the role of TSP-1 and TSP-2 in
cardiovascular diseases (CVDs; Table
I), and the potential pathways associated with these TSPs will
be discussed.
TSP-2 shares the same structure with TSP-1, however
the amino acid sequences are slightly different. The primary
differences in the amino acid sequence are located in the
N-terminal domain. Evidence reveals there is only a 32% amino acid
sequence similarity in the N-terminal heparin binding domain
between TSP-1 and TSP-2 (3).
Therefore, the ligands binding TSP-1 and TSP-2 at the N-terminal
domain are different in certain circumstances. For example, a
previous study identified that recombinant human TSP1 may activate
the latent TGF-β, but this phenomenon is not observed in
recombinant mouse TSP-2 (45).
Therefore, although TSP-1 and TSP-2 share the same structure, their
different amino acid sequences, especially in the N-terminal
domain, is the cause of their differences in binding ligands.
The interaction between TSP-1 and TSP-2 and their
ligands may also affect each other. For example, TSP-1 and TSP-2
are known to compete with each other for degradation through LRP,
which contributes to maintaining the levels of TSP-1 and TSP-2 in
certain situations, such as wound healing (46).
Due to their multidomain structure and the ability
to interact with multiple ligands, TSP-1 and TSP-2 are active in
various types of physiological and pathological processes. At
present, there have been multiple studies concerning the role of
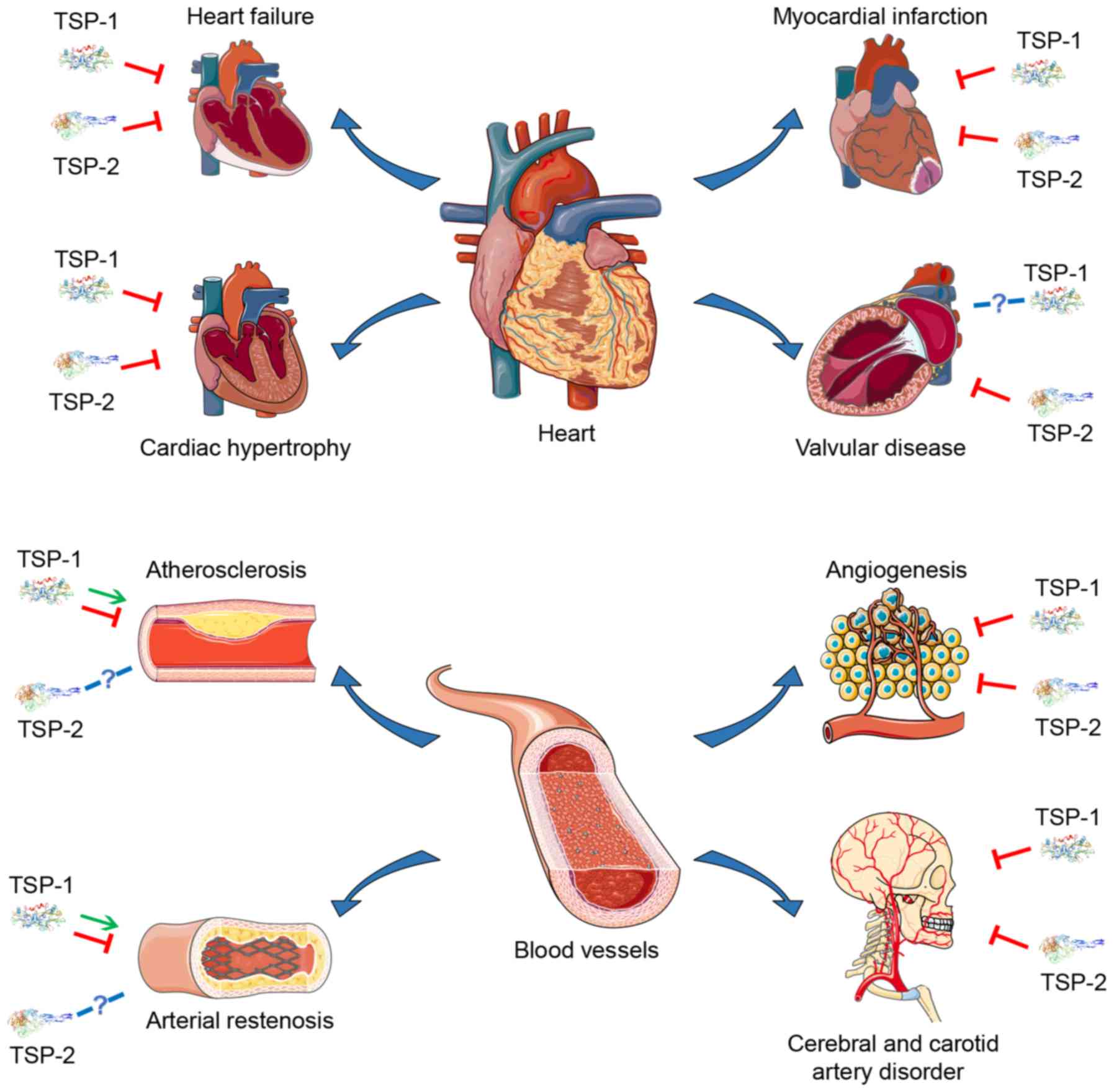
TSP-1 and TSP-2 in various CVDs (Table I and Fig. 2), suggesting that they may become
potential therapeutic targets.
MI is myocardial ischemic necrosis caused by a rapid
decrease or interruption of coronary blood supply, which leads to
high mortality rates worldwide. In patients with acute ST-segment
elevation MI, the expression of TSP-1 is significantly increased
(47), and the decrease of TSP-1
following percutaneous coronary intervention (PCI) is associated
with major adverse cardiac events (48). In rats with infarcted hearts,
TSP-1 was transiently induced and located in cells at the border
area of the infarction, which can be increased by
ischemia/reperfusion (49). In
canine and murine models of reperfusion injury, TSP-1 also
exhibited a selective distribution in the ECM, the microvascular
endothelium, and the mononuclear cells of the infarct border zone.
TSP-1-deficient mice have a severe post-infarction remodeling
compared with that in wild-type mice, although their infarct size
was almost equal, indicating that selective endogenous expression
of TSP-1 in the infarct border zone may limit the expansion of the
granulation tissue and protect the non-infarcted myocardium from
fibrotic remodeling (50).
Following MI, left ventricular (LV) remodeling
significantly contributes to LV dilation and dysfunction, which
leads to functional damage and poor prognosis. The pathological
process of LV remodeling includes scar formation in the infarcted
area, as well as hypertrophy and fibrosis in the non-infarcted
surrounding area. This is primarily due to the degradation of ECM,
the deposition of myofibroblasts, and the rapid increase in the
amount of collagen following infarction. Previous studies have
identified that extracellular collagen matrix (ECCM) remodeling
largely contributes to LV remodeling, and the decrease of ECCM is
associated with the increase of LV dilation and rupture, indicating
that TSP-1 may participate in the pathology of MI (51). Exposure to hypoxia markedly
induced the expression of TSP-2 in cardiomyocyte progenitor cells
(hCMPCs), and knockdown of TSP-2 resulted in increased
proliferation, migration and MMP activity of hCMPCs, indicating
that TSP-2 may participate in controlling the migratory and
invasive capacities of hCMPCs under hypoxic conditions (52). However, little is known regarding
the specific role of TSP-2 in MI.
Conversely, certain functional variants of the TSP-1
and TSP-2 genes may be associated with MI (53–55). The N700S polymorphism of TSP-1 is
a potential genetic risk factor for MI (56–58), as it disrupts calcium-binding
sites (59). Those who are
homozygous for the minor allele or are heterozygous (GG and TG
genotypes, respectively) have a significantly increased risk of MI
compared with those who are homozygous (TT genotype) for the major
allele (54). These data provide
a novel perspective on the TSP-associated therapeutic approach of
MI.
Cardiac hypertrophy is primarily induced by chronic
pressure overload, such as essential hypertension. Pathological
features of cardiac hypertrophy include increased growth of the
cardiomyocytes, proliferation of the cardiac fibroblasts and
increased ECM deposition. There have been some reports on the role
of TSP-1 and TSP-2 in cardiac hypertrophy. Compared with wild type
mice, TSP-1-deficient mice exhibited enhanced early hypertrophy and
late dilation when exposed to pressure overload (60). Despite this, TSP-1
(−/−) mice exhibited increased myocardial
MMP-3 and -9 activation following pressure overload (60). In obese diabetic DB/DB
mice, myocardial TSP-1 levels are significantly upregulated in the
perivascular and interstitial space. In comparison with normal
DB/DB mice, DB/DB TSP-1 (−/−)
mice exhibited an enhanced LV dilation, which was associated with
mild non-progressive systolic dysfunction, and TSP-1 could
incorporate into the matrix and inhibit leptin-induced MMP-2
activation (61). These previous
studies suggest that TSP-1 is upregulated in the diabetic heart and
prevents chamber dilation by exerting matrix-preserving actions on
the cardiac fibroblasts.
TSP-2 is also closely associated with cardiac
hypertrophy. Data suggests that older TSP-2
(−/−) mice are associated with an enhanced
dilated cardiomyopathy characteristic as impaired systolic function
as well as increased cardiac dilatation and myocardial fibrosis,
indicating that TSP-2 deficiency leads to an age-associated dilated
cardiomyopathy (62). Compared to
wild-type mice, TSP-2-knockout mice display increased mortality
accompanied by decreasing cardiac function, increased cardiomyocyte
apoptosis and ECM damage in a doxorubicin-induced cardiomyopathy
mouse model (63). The absence of
TSP-2 also results in decreased systolic function and enhanced
cardiac dilatation in human Coxsackie virus B3 (CVB3)-induced
myocarditis (64). Previous data
also identified that TSP-2 expression is activated uniquely in
hypertrophic hearts that may develop heart failure, which may be an
early-stage molecular program of heart failure (65).
Abnormal myocardium remodeling leads to myocardial
overload. If not treated promptly, long-term myocardial overload
may progress into heart failure. From the perspective of pathology,
heart failure is associated with abnormal inflammation, coagulation
activation and endothelial dysfunction. TSP-1 and TSP-2 also
participate in some of these changes. Previous studies have
revealed that TSP-1 expression is decreased in failing hearts,
which may be associated with ventricular dilatation (66,67). Treatment of cardiomyocytes with a
TSP1-derived peptide that activates CD47 leads to increased
cardiomyocyte hypertrophy in a Ca2+ and calmodulin
protein kinase II dependent manner, indicating that TSP-1 may
contribute to LV hypertrophy and heart failure (68). Using aged mouse models with
failure-resistant and failure-prone characteristics, a previous
study identified that micro(mi) RNA-18 and miRNA-19 may modulate
TSP-1 expression and cardiac ECM protein levels in age-associated
heart failure; therefore, decreased miRNA-18/19 and increased TSP-1
levels may contribute to the identification of failure-prone hearts
(69). TSP-1 levels in patients
with heart failure may also be decreased due to oral
anticoagulation therapy, which is used to prevent thromboembolic
events (70).
Elevated TSP-2 is primarily associated with poor
prognosis in patients with heart failure. Among patients with
coronary heart disease with symptomatic congestive heart failure
(CHF), circulating TSP-2 is increased, which is associated with
increased 3-year CHF-associated death, all-cause mortality and
recurrent hospitalization risk (71). In patients with preserved ejection
fraction heart failure, high serum levels of TSP-2 are associated
with poor prognosis (72,73). TSP-2 overexpression in wild-type
mouse hearts led to decreased cardiac inflammation and improved
cardiac function after CVB3 infection, suggesting that TSP-2 may
mitigate against cardiac injury, inflammation, and dysfunction
during acute viral myocarditis (64).
Calcific aortic valve disease (CAVD) is a
progressive disorder manifesting as sclerotic stiffening and
valvular thickening, eventually leading to aortic stenosis. The
pathological process of CAVD is accompanied by inflammatory cell
infiltration, lipid accumulation, fibrosclerosis, ECM disorder,
angiogenesis and nodular calcification (74). In fibrotic and stenotic aortic
valves, the mRNA levels of TSP-2 are increased 4.9-fold (P=0.037)
and 4.8-fold (P=0.001), respectively (75). TSP-1 can also be detected in the
fibrotic and stenotic valves, but the expression of TSP-1 is not
significantly different, indicating that CAVD was associated with
TSP-2 upregulation in aortic cusps (75). However, evidence suggesting an
association between TSP-1 and valvular diseases is limited, and the
specific role of TSP-2 in the pathological process of valvular
disease requires further study.
Cerebral and carotid artery disease are important
subgroups of peripheral vascular diseases, which have high
mortality rates worldwide. TSP-1 and TSP-2 may also serve a role in
cerebral and carotid artery disease. In symptomatic patients with
carotid artery diseases, TSP-1 expression on the surface of
circulating platelets is significantly increased (76). Compared with wild-type mice, TSP-1
(−/−) mice exhibit a decreased response to
fluvastatin in inhibiting intimal hyperplasia following carotid
artery ligation, indicating that the statin effect on intimal
hyperplasia may be dependent on TSP-1 (77). A previous study identified that
the expression of TSP-1 is increased following a stroke, and TSP-1
deficiency leads to impaired recovery (78). High expression levels of TSP-1 and
TSP-2 were identified in the ischemic rat brain following cerebral
ischemia/reperfusion, which may contribute to spontaneous
resolution of postischemic angiogenesis (79). In a spontaneous intracerebral
hemorrhage (ICH) rat model, thrombin treatment induced high
expression of TSP-1 or TSP-2 in the blood vessels around the
damaged brain region. These data provide support the hypothesis
that thrombin positively regulates the expression of TSP-1 and
TSP-2 following ICH, which may be involved in modulating
angiogenesis in injured brains (79–81).
In a rat carotid balloon angioplasty model,
intraluminal delivery of TSP-2 small inhibiting (si)RNA inhibited
the vascular response to the injury (82). TSP-2 is increased and colocalized
to the astrocytes following a stroke, and TSP-2 deficiency leads to
an impaired recovery following stroke (78,83). TSP-2 expression was increased in
the ischemic brain, which may contribute to the spontaneous
resolution of post-ischemic angiogenesis (79). These data suggest that TSP-2 may
promote angiogenesis and recovery following cerebral and carotid
artery injury.
Atherosclerosis is characterized by thickening,
hardening and decreased elasticity of the arterial wall. Lipid
levels, endothelial cell injury, inflammation and the migration of
vascular smooth muscle cells (VSMC) are considered as several
fundamental pathological processes of atherosclerosis. Previous
evidence suggested that TSP-1 can interact with some of the
aforementioned factors and further regulate the pathological
process of atherosclerosis through various mechanisms, while the
association between TSP-2 and atherosclerosis requires further
investigation. Following partial carotid ligation, disturbed blood
flow induced arterial stiffening through collagen deposition.
Compared with wild type carotid arteries, TSP-1 knockout animals
have significantly decreased arterial stiffening, indicating that
disturbed flow may promote arterial stiffening through TSP-1
(84). Conversely, proatherogenic
flow conditions may induce endothelial apoptosis via TSP-1
(40). The absence of TSP-1
accelerates the maturation of the atherosclerotic plaque in
apolipoprotein E (ApoE−/−) mice, indicating
that TSP-1 may function as an inhibitor of atherosclerosis
(85,86). TSP-1 may also interact with
lipoproteins. In hypercholesterolemic atherosclerotic rabbits, the
overexpressed TSP-1 secreted by injured arteries may bind to
very-low-density lipoprotein (VLDL), which may promote its
incorporation into nascent atherosclerotic plaques, simultaneously
delivering VLDL cholesterol into the lesions (87,88). These results indicate that TSP-1
may serve different roles in different pathological stages of
atherosclerosis. Therefore, it is necessary to further investigate
the specific role of TSP-1 in atherosclerosis.
An important pathological process of atherosclerosis
is the migration of media smooth muscle cells (SMCs) into the
intima and hyperplasia. The expression of TSP-1 has been
demonstrated to increase in VSMC in human atherosclerotic lesions
(89), which may contribute to
inflammation and atherogenesis. Hypoxia induces the migration of
the coronary artery SMCs, which is elicited by TSP-1 (90,91). An additional study identified that
TSP modulates SMCs migration, which may accelerate atherosclerotic
lesion development during vascular injury or inflammation (92).
TSP-1 may also modulate the interaction between
diabetes and atherosclerosis. Evidence reveals that TSP-1
expression is increased in large arteries of diabetic animals
however, the protein levels of TSP-1 in microvascular endothelial
cells are decreased when exposed to high glucose levels (89,93,94). In a hyperglycemic ApoE
(−/−) mouse model, lack of TSP-1 prevented
atherogenic lesion formation (95). The expression of TSP-1 is
increased in hypoxic pulmonary hypertension rats, which may
contribute to the pathogenesis of hypoxic pulmonary vascular
remodeling (96).
Compared with TSP-1, there is limited research on
the association between TSP-2 and atherosclerosis. In
atherosclerotic specimens, TSP-2 mRNA was absent from intraplaque
microvessels and endothelial cells lining the atheromatous plaque
(97). Therefore, the specific
mechanism of TSP-2 in the pathological process of atherosclerosis
requires further investigation.
Angiogenesis is a fundamental physiological process
associated with tissue repair following injury, which also promotes
tumor progression. This process is tightly modulated by various
growth factors and the interaction between cells and the ECM. TSP-1
and TSP-2 have been revealed to regulate angiogenesis by
interacting with specific growth factors, cells and ECM. Previous
evidence indicates that downregulation of endothelial cell TSP-1
causes an enhancement of in vitro angiogenesis (98). In vitro and in vivo
models indicated that factor XIII, a clotting factor, may also
promote angiogenesis by downregulating TSP-1 and stimulate
endothelial cell proliferation and migration (99). In TSP-1-deficient animals, tumor
burden and vasculature increase markedly, and TSP-1 overexpression
resulted in decreased tumor diameter and fewer tumor capillaries,
indicating that TSP-1 may inhibit tumor angiogenesis (22,100). The inhibitory effect of TSP-1 on
tumors may be accomplished via cross-talk with endothelial cells
(101). The overexpression of
TSP1-CD47 signaling in diabetes is associated with endothelial cell
dysfunction, which leads to impaired angiogenesis (102). In the ischemic retina,
glia-derived TSP-1 may inhibit angiogenic responses (103), and deficiency of TSP-1
contributes to enhanced neovascularization in the eye (104–108).
Similar to TSP-1, TSP-2 can also inhibit
angiogenesis and tumor growth, even with greater potency compared
with that of TSP-1 (109). In
vitro experiments indicated that TSP-2 inhibits proliferation
of microvascular endothelial cells (110,111), and the absence of TSP-2 is
associated with enhanced angiogenesis, partly due to the altered
endothelial cell and ECM interactions (112,113). Decreasing gelatinolytic activity
in situ leads to TSP-2-limited angiogenesis (114). In rheumatoid arthritis, TSP-2
overexpression also inhibits vascularization (115).
In older mice, the delay of TSP-2 and MMP2
expression in wounds may promote the impaired rate of wound healing
(116). TSP-2 gene knockout mice
exhibited increased blood vessel density, but no such alteration
was observed in TSP-1-deficient animals (117). This evidence indicates the role
of TSP-2 in anti-angiogenesis.
Restenosis of the arteries following cardiovascular
surgery, such as PCI, is a major problem, which leads to a poor
prognosis. The pathological process of arterial restenosis is
similar to atherosclerosis to a certain extent, including
endothelial injury, migration and proliferating of VSMCs into the
intima. Similar to atherosclerosis, the precise role of TSP-1 in
the pathological process of arterial restenosis is difficult to
define. In the balloon catheter injury rat model, TSP was markedly
increased in the thickening arterial wall, and the TSP antigen in
thickening arterial wall is primary secreted by VSMCs (118). In rat resistance arteries, TSP-1
was able to reverse the pathological inward remodeling caused by
spontaneous hypertension, indicating that TSP-1 may act as an
inhibitor of arterial restenosis (119). A previous study identified that
the interaction of TSP-1 and β1 integrin is associated with
platelet-stimulated SMC proliferation (120). However, there is also evidence
revealing that TSP-1 is not a major component of ECM in human
restenotic tissues, even in the presence of hypercellularity or
ongoing cellular proliferation (121).
In human aortic SMCs, TSP-2 silencing caused by
siRNA improves cell attachment but does not affect cell
proliferation and migration, suggesting that TSP-2 also
participates in the pathological process of arterial restenosis
(122), which represents a novel
hypothesis.
In addition to the aforementioned major CVDs, TSP-1
and TSP-2 also serve important roles in a number of other CVDs.
Evidence indicated that TSP-1 may contribute to the pathogenesis of
pulmonary hypertension associated with hypoxia (123). TSP-1 deficiency contributes
markedly to maladaptive remodeling of the ECM, causing an
acceleration of aortic aneurysm progression (124). During the abdominal aortic
aneurysm development, TSP-1 regulates the adhesion and migration of
mononuclear cells and promotes vascular inflammation (125). During autologous proangiogenic
cell therapy, TSP-1-derived peptide RFYVVMWK may interact with
priming CD34+ cells and enhance the vascular engraftment (126). TSP-2 (−/−)
mice exhibit a bleeding diathesis even if they have normal blood
coagulation and no thrombocytopenia (127), and an altered foreign body
reaction characterized by an enhanced vascularity (128,129). The plasma TSP-2 level is
elevated in acute Kawasaki disease, which may be a novel predictor
for intravenous immunoglobulin resistance (130). In a TSP-2-knockout mouse model,
significantly increased endothelial cell density and reduced
fibrosis were observed in the peri-graft region during the cardiac
cell transplantation (131).
These studies suggest that TSP-1 and TSP-2 also function in other
CVDs, such as pulmonary hypertension, aortic aneurysm progression
and acute Kawasaki disease.
Due to their multidomain structure, TSP-1 and TSP-2
can specifically bind to numerous types of different ligands.
Therefore, they are involved in various signal pathways regulating
cellular activities and ECM components in CVDs (Tables II and III). A comprehensive description of
these pathways may facilitate the understanding of the role TSP-1
and TSP-2 serve in the pathological processes of multiple CVDs at
the molecular level, which may provide certain potential
therapeutic strategies.
Interactions between various cells and the ECM cause
direct or indirect modulation of numerous cellular activities, such
as proliferation, adhesion, migration, differentiation and
apoptosis, which contributes markedly to numerous CVDs. TSP-1 binds
to HSPG with high affinity, which promotes human melanoma cell
migration (132). At the sites
of inflammation, TSP1 binding to tumor-specific glycoprotein 6 may
regulate hyaluronan metabolism, indicating a critical role of TSP-1
in mediating cellular interactions with hyaluronan (9). During the vascular smooth muscle
inflammatory response, TSP-1 and TSP-2 may bind to versican and
negatively modulate the ECM component (11). In certain circumstances, TSP-1 can
bind to LIMPII and promote cell adhesion (27). Previous data indicated that TSP-1
may bind to calreticulin (CRT) on the cell surface and induce focal
adhesion disassembly, as well as cell migration through the
association of CRT with lipoprotein LRP (10,13,133).
Integrins are a family of glycosylated,
heterodimeric transmembrane receptors that consist of α and β
subunits, which provide a physical link between the ECM and the
cytoskeleton. Previous studies identified that TSP-1 and TSP-2 may
also interact with various types of integrins. Binding of integrin
α3β1 to TSP-1 mediates efficient migration of ECs, indicating that
the binding of TSP-1 and integrin α3β1 stimulates cell adhesion and
migration (15,16). Despite this, integrin α3β1 binding
to TSP-1 can also mediate cell motility and inhibit angiogenesis
(15,17). Studies have demonstrated that α4β1
integrin mediates CD47-stimulated sickle red blood cells adhesion
to immobilized TSP-1 and modulate T cell behavior (18,134). In addition, the N-terminal
domain of TSP-1 is also a ligand for α6β1 integrin, which modulates
the adhesion of human microvascular endothelial to immobilized
TSP-1 and TSP-2 (19). TSP-1 and
TSP-2 may also interact with β1 integrin, contributing to the
adhesion of cells that express β1 integrin (28,135). The type III repeats of TSP-1 and
TSP-2 may interact with integrin αIIβ3 and αvβ3, promoting their
binding with platelets (136). A
previous study has demonstrated that TSP-2 may also contribute to
anti-angiogenesis in diabetes myocardium (137). These results indicate the
important role of the interaction between integrins and TSPs in the
ECM-receptor interaction.
The PI3K-Akt pathway can be activated by various
cellular stimuli, such as growth factors, and regulates numerous
fundamental cellular functions, such as cell proliferation,
migration and apoptosis. These cellular activities are critical for
the pathological process of CVDs. TSP-1 and TSP-2 also participate
in a number of these activities through the PI3K-Akt pathway. The
N-terminal domain of TSP-1 can interact with platelet-derived
growth factor, leading to mediation of VSMC proliferation and
migration (138). In addition to
the ability to degrade collagen, studies suggest that MMP-9 may
also release vascular endothelial growth factor to participate in
modulating the invasion and the morphogenesis of endothelial cells,
which can also be modulated by TSP-1 (139). The type II repeats of TSP-1
interacts with EGFR and increases cell migration (140). The type III repeat domain of
TSP-1 and TSP-2 may interact with integrin αIIβ3 and αvβ3,
promoting SMC migration (141).
Binding of TSP-1 and TSP-2 to FGF2 inhibits apoptosis (43) and triggers caspase-independent
cell death (142).
CD36 is a multi-ligand receptor that participates in
various pathological processes of CVDs, such as the formation of
atherosclerosis. CD47 is a glycoprotein on numerous types of cell
surfaces, which serves an important regulatory role in immune
response and inflammation. The binding of TSP-1 and CD36 inhibits
angiogenesis through promoting endothelial cell apoptosis and
inhibiting nitric oxide (NO) signal transduction (15,25,143). The C-terminal domain of TSP-1
and TSP-2 can interact with CD47, which may promote cell migration
and adhesion (18,42,144), inhibit cyclic guanosine
monophosphate synthesis, nitric oxide (NO) signaling (143,145) and cell cycle progression in ECs
(146). Previous data revealed
that the binding of TSP-1 and CD47 may also inhibit angiogenesis,
blood flow, and adhesion of monocytes and macrophages (147–150), which may promote foam cell
formation (151), pulmonary
arterial vasculopathy (152) and
LV heart failure (68). Through
these mechanisms, TSP-1 and TSP-2 serve a significant role in
numerous CVDs.
A wide range of different cellular functions, such
as cell proliferation, differentiation, migration and apoptosis,
can also be modulated by TGF-β, a member of the transforming growth
factor superfamily, which is a group of secreted cytokines. Studies
also revealed that TSP-1 regulates the above cellular activities
through the TGF-β pathway. Previous data suggested that the type I
repeats of TSP-1 may bind and activate latent TGF-β. The activated
TGF-β can further stimulate new matrix deposition and angiogenesis
(45,153–155), promote inflammatory response via
recruitment of inflammatory cells and increase myofibroblast
differentiation (156) through
the TGF-β pathway.
TSP-2 may also bind to latent TGF-β. However, TSP-2
cannot activate latent TGF-β. In addition, due to this reason,
TSP-1 and TSP-2 can regulate the activity of TGF-β and modulate the
downstream pathways by competitively binding to it (45).
Numerous pathological processes of CVDs are
accompanied by the destruction of ECM homeostasis. For example,
excessive accumulation of type I and type III collagen is a
significant feature of cardiac hypertrophy, which is due to the
higher collagen synthesis capacity compared with the degradation
ability. MMP2 and MMP9 serve crucial roles in maintaining ECM
homeostasis. Evidence revealed that TSP-1 and TSP-2 may interact
with MMP2 and MMP9, which can inhibit their activity and regulate
collagen homeostasis (20,157,158).
In addition, there is also evidence revealing that
collagens can interact with TSP-1 directly. The C-terminal domain
of TSP-1 may bind to collagen I, contributing to fibroblast
homeostasis (156). These
results suggest that TSP-1 and TSP-2 contribute markedly to ECM
homeostasis.
Phagocytosis of TSP-1 serves a critical role in
tissue remodeling and inflammation in CVDs, which is mediated by
various ligands. In vascular endothelial cells, the heparan sulfate
proteoglycans expressed on the cell surface are associated with the
process of binding and endocytosis of TSP-1, which leads to its
lysosomal degradation (8).
Evidence revealed that the HSPG on the endothelial cells may
mediate the binding and degradation of TSP-1 (159). Studies suggest that LRP may also
function in mediating phagocytosis of TSP-1 in certain types of
cells (160,161), indicating that LRP may serve a
significant role in the catabolism of TSP-1 in vivo. The
binding of TSP-1 and CD36 has been demonstrated to promote the
internalization of oxidized LDL, fatty acids and phospholipids,
leading to inhibition of atherosclerosis (162). However, little is known on the
specific role of TSP-2 in the phagosome pathway, and requires
further study.
Calcium is an indispensable ion involved in numerous
physiological processes in the human body. It participates in
maintaining the biopotentials on both sides of the cell membrane,
maintaining normal muscle expansion and relaxation, nerve
conduction and vasoconstriction. TSP-1 and TSP-2 can bind to
calcium and affect the function of modulating physiological
activities. Using a simulated model, previous studies have
identified that the change between fully calcium-loaded and
calcium-depleted TSP1-Sig1 may modulate its interactions, which may
become a novel therapeutic target (38,163). Binding of TSP-2 and FGF2 can be
inhibited by calcium, indicating that calcium can affect cell
function via intervening in interactions between other molecules
(36).
In addition to the aforementioned pathways, TSP-1
and TSP-2 also interact with numerous other ligands. During the
coagulation reaction, TSP-1 can interact with the vitamin D-binding
protein, contributing to the chemotaxis of coagulation factor C5a
(164). TSP-2 can interact with
cytochrome p450 1B1, promoting angiogenesis through the regulation
of oxidative stress (165). In
addition, as an important gas signal in the cardiovascular system,
NO can negatively regulate TSP-2 transcription and induce
angiogenesis (166).
A disintegrin and metalloproteinase with
thrombospondin motifs (ADAMTS) is a type of metalloproteinase which
has been demonstrated to be associated with numerous CVDs. Studies
have identified that ADAMTS1 contributes to wound closure and
inhibits the angiogenesis via interaction with TSP-1 and TSP-2
(167). Evidence has revealed
that there is a close association between ADAMTS7 and CVDs. TSP-1
and TSP-2 interaction with ADAMTS7 promotes the pathological
processes of atherosclerosis, coronary artery disease (168–172), aortic aneurysm (173) and vascular remodeling (174–176) through interacting with TSP-1 and
TSP-2. Conversely, there is also evidence revealing that ADAMTS7
may inhibit LV reverse remodeling following MI (177–179), suggesting ADAMTS7 may be a
critical regulator in CVDs.
The present review suggests that TSP-1 and TSP-2
serve significant roles in the pathological process of numerous
CVDs, and their multi-domain structural features and ability to
bind to different ligands may also provide novel targets for the
treatment of different CVDs at the molecular level.
However, there are two limitations of the present
study. Firstly, although both TSP-1 and TSP-2 have a similar
multidomain structure, both bind to different ligands and serve
different roles. There is limited research into the specific role
of TSP-2 in the pathogenesis of numerous CVDs, indicating that more
research is required. Secondly, numerous novel ligands remain to be
identified. Fortunately, with the development of new large-scale
techniques, including array-based surface plasmon resonance,
new-generation yeast two-hybrid and numerous novel computational
methods, novel TSP-1 and TSP-2 ligands may be identified (4). Identification of these ligands may
contribute to determination of the interaction networks of TSP-1
and TSP-2, which may provide an improved understanding of their
role in CVDs.
The authors would like to thank Dr Yu Han (Zhejiang
University, Hangzhou, China), Dr Xinyi Teng (Zhejiang University,
Hangzhou, China) and Dr Xin Guo (School of Medicine, Zhejiang
University, Hangzhou, China) for their valuable advice. In
addition, the authors acknowledge Dr Yasaman Iran Manesh (Zhejiang
University, Hangzhou, China) for her revising the article.
The present study was supported by the National
Natural Science Foundation of China (grant. nos. 81300236, 81670433
and 81970398), the Project of Zhejiang Medical Young Talents 2017
(grant. no. 20190301), the Zhejiang Medical and Health Science and
Technology Project (grant. no. 2020RC014) and the Natural Science
Foundation of Zhejiang Province (grant. no. LQ20H020008).
Not applicable.
KZ, ML and LY carried out literature search and
acquisition of references. KZ, ZL and GF were involved in the
conception and design of the manuscript. ZL performed manuscript
review and gave final approval of the version to be published. All
authors have read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Chistiakov DA, Melnichenko AA, Myasoedova
VA, Grechko AV and Orekhov AN: Thrombospondins: A role in
cardiovascular disease. Int J Mol Sci. 18:E15402017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bornstein P: Thrombospondins as
matricellular modulators of cell function. J Clin Invest.
107:929–934. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bornstein P: Thrombospondins: Structure
and regulation of expression. FASEB J. 6:3290–3299. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Resovi A, Pinessi D, Chiorino G and
Taraboletti G: Current understanding of the thrombospondin-1
interactome. Matrix Biol. 37:83–91. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anilkumar N, Annis DS, Mosher DF and Adams
JC: Trimeric assembly of the C-terminal region of thrombospondin-1
or thrombospondin-2 is necessary for cell spreading and fascin
spike organisation. J Cell Sci. 115:2357–2366. 2002.PubMed/NCBI
|
|
6
|
Yu H, Tyrrell D, Cashel J, Guo NH, Vogel
T, Sipes JM, Lam L, Fillit HM, Hartman J, Mendelovitz S, et al:
Specificities of heparin-binding sites from the amino-terminus and
type 1 repeats of thrombospondin-1. Arch Biochem Biophys.
374:13–23. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Elzie CA and Murphy-Ullrich JE: The
N-terminus of thrombospondin: The domain stands apart. Int J
Biochem Cell Biol. 36:1090–1101. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Feitsma K, Hausser H, Robenek H, Kresse H
and Vischer P: Interaction of thrombospondin-1 and heparan sulfate
from endothelial cells. Structural requirements of heparan sulfate.
J Biol Chem. 275:9396–9402. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kuznetsova SA, Day AJ, Mahoney DJ, Rugg
MS, Mosher DF and Roberts DD: The N-terminal module of
thrombospondin-1 interacts with the link domain of TSG-6 and
enhances its covalent association with the heavy chains of
inter-alpha-trypsin inhibitor. J Biol Chem. 280:30899–30908. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan Q, Murphy-Ullrich JE and Song Y:
Structural insight into the role of thrombospondin-1 binding to
calreticulin in calreticulin-induced focal adhesion disassembly.
Biochemistry. 49:3685–3694. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuznetsova SA, Issa P, Perruccio EM, Zeng
B, Sipes JM, Ward Y, Seyfried NT, Fielder HL, Day AJ, Wight TN and
Roberts DD: Versican-thrombospondin-1 binding in vitro and
colocalization in microfibrils induced by inflammation on vascular
smooth muscle cells. J Cell Sci. 119:4499–4509. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sweetwyne MT, Pallero MA, Lu A, Van Duyn
Graham L and Murphy-Ullrich JE: The calreticulin-binding sequence
of thrombospondin 1 regulates collagen expression and organization
during tissue remodeling. Am J Pathol. 177:1710–1724. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang L, Murphy-Ullrich JE and Song Y:
Molecular insight into the effect of lipid bilayer environments on
thrombospondin-1 and calreticulin interactions. Biochemistry.
53:6309–6322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Orr AW, Pallero MA, Xiong WC and
Murphy-Ullrich JE: Thrombospondin induces RhoA inactivation through
FAK-dependent signaling to stimulate focal adhesion disassembly. J
Biol Chem. 279:48983–48992. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ndishabandi D, Duquette C, Billah GE,
Reyes M, Duquette M, Lawler J and Kazerounian S: Thrombospondin-1
modulates actin filament remodeling and cell motility in mouse
mammary tumor cells in vitro. Discoveries (Craiova). 2:e312014.
View Article : Google Scholar
|
|
16
|
Chandrasekaran L, He CZ, Al-Barazi H,
Krutzsch HC, Iruela-Arispe ML and Roberts DD: Cell
contact-dependent activation of alpha3beta1 integrin modulates
endothelial cell responses to thrombospondin-1. Mol Biol Cell.
11:2885–2900. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Furrer J, Luy B, Basrur V, Roberts DD and
Barchi JJ Jr: Conformational analysis of an alpha3beta1
integrin-binding peptide from thrombospondin-1: Implications for
antiangiogenic drug design. J Med Chem. 49:6324–6333. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brittain JE, Han J, Ataga KI, Orringer EP
and Parise LV: Mechanism of CD47-induced alpha4beta1 integrin
activation and adhesion in sickle reticulocytes. J Biol Chem.
279:42393–42402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Calzada MJ, Sipes JM, Krutzsch HC,
Yurchenco PD, Annis DS, Mosher DF and Roberts DD: Recognition of
the N-terminal modules of thrombospondin-1 and thrombospondin-2 by
alpha-6beta1 integrin. J Biol Chem. 278:40679–40687. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bein K and Simons M: Thrombospondin type 1
repeats interact with matrix metalloproteinase 2. Regulation of
metalloproteinase activity. J Biol Chem. 275:32167–32173. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee T, Esemuede N, Sumpio BE and Gahtan V:
Thrombospondin-1 induces matrix metalloproteinase-2 activation in
vascular smooth muscle cells. J Vasc Surg. 38:147–154. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rodriguez-Manzaneque JC, Lane TF, Ortega
MA, Hynes RO, Lawler J and Iruela-Arispe ML: Thrombospondin-1
suppresses spontaneous tumor growth and inhibits activation of
matrix metalloproteinase-9 and mobilization of vascular endothelial
growth factor. Proc Natl Acad Sci USA. 98:12485–12490. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zeng T, Yuan J, Gan J, Liu Y, Shi L, Lu Z,
Xue Y, Xiong R, Huang M, Yang Z, et al: Thrombospondin 1 is
increased in the aorta and plasma of patients with acute aortic
dissection. Can J Cardiol. 35:42–50. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dawson DW, Pearce SF, Zhong R, Silverstein
RL, Frazier WA and Bouck NP: CD36 mediates the In vitro inhibitory
effects of thrombospondin-1 on endothelial cells. J Cell Biol.
138:707–717. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Silverstein RL and Febbraio M:
CD36-TSP-HRGP interactions in the regulation of angiogenesis. Curr
Pharm Des. 13:3559–3567. 2007. View Article : Google Scholar
|
|
26
|
Simantov R, Febbraio M and Silverstein RL:
The antiangiogenic effect of thrombospondin-2 is mediated by CD36
and modulated by histidine-rich glycoprotein. Matrix Biol.
24:27–34. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Crombie R and Silverstein R: Lysosomal
integral membrane protein II binds thrombospondin-1.
Structure-function homology with the cell adhesion molecule CD36
defines a conserved recognition motif. J Biol Chem. 273:4855–4863.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Calzada MJ, Annis DS, Zeng B,
Marcinkiewicz C, Banas B, Lawler J, Mosher DF and Roberts DD:
Identification of novel beta1 integrin binding sites in the type 1
and type 2 repeats of thrombospondin-1. J Biol Chem.
279:41734–41743. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Goel HL, Moro L, Murphy-Ullrich JE, Hsieh
CC, Wu CL, Jiang Z and Languino LR: Beta1 integrin cytoplasmic
variants differentially regulate expression of the antiangiogenic
extracellular matrix protein thrombospondin 1. Cancer Res.
69:5374–5382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahamed J, Janczak CA, Wittkowski KM and
Coller BS: In vitro and in vivo evidence that thrombospondin-1
(TSP-1) contributes to stirring- and shear-dependent activation of
platelet-derived TGF-beta1. PLoS One. 4:e66082009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kumar R, Mickael C, Kassa B, Gebreab L,
Robinson JC, Koyanagi DE, Sanders L, Barthel L, Meadows C, Fox D,
et al: TGF-β activation by bone marrow-derived thrombospondin-1
causes Schistosoma- and hypoxia-induced pulmonary hypertension. Nat
Commun. 8:154942017. View Article : Google Scholar
|
|
32
|
McGillicuddy FC, O’Toole D, Hickey JA,
Gallagher WM, Dawson KA and Keenan AK: TGF-beta1-induced
thrombospondin-1 expression through the p38 MAPK pathway is
abolished by fluvastatin in human coronary artery smooth muscle
cells. Vascul Pharmacol. 44:469–475. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Takahashi K, Mernaugh RL, Friedman DB,
Weller R, Tsuboi N, Yamashita H, Quaranta V and Takahashi T:
Thrombospondin-1 acts as a ligand for CD148 tyrosine phosphatase.
Proc Natl Acad Sci USA. 109:1985–1990. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Takahashi K, Sumarriva K, Kim R, Jiang R,
Brantley-Sieders DM, Chen J, Mernaugh RL and Takahashi T:
Determination of the CD148-interacting region in thrombospondin-1.
PLoS One. 11:e01549162016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Garg P, Yang S, Liu A, Pallero MA,
Buchsbaum DJ, Mosher DF, Murphy-Ullrich JE and Goldblum SE:
Thrombospondin-1 opens the paracellular pathway in pulmonary
microvascular endothelia through EGFR/ErbB2 activation. Am J
Physiol Lung Cell Mol Physiol. 301:L79–L90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rusnati M, Borsotti P, Moroni E, Foglieni
C, Chiodelli P, Carminati L, Pinessi D, Annis DS, Paiardi G,
Bugatti A, et al: The calcium-binding type III repeats domain of
thrombospondin-2 binds to fibroblast growth factor 2 (FGF2).
Angiogenesis. 22:133–144. 2019. View Article : Google Scholar
|
|
37
|
Kvansakul M, Adams JC and Hohenester E:
Structure of a thrombospondin C-terminal fragment reveals a novel
calcium core in the type 3 repeats. EMBO J. 23:1223–1233. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gupta A, Agarwal R, Singh A and Bhatnagar
S: Calcium-induced conformational changes of Thrombospondin-1
signature domain: Implications for vascular disease. J Recept
Signal Transduct Res. 37:239–251. 2017. View Article : Google Scholar
|
|
39
|
Joo SJ: Mechanisms of platelet activation
and integrin αIIβ3. Korean Circ J. 42:295–301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Freyberg MA, Kaiser D, Graf R,
Buttenbender J and Friedl P: Proatherogenic flow conditions
initiate endothelial apoptosis via thrombospondin-1 and the
integrin-associated protein. Biochem Biophys Res Commun.
286:141–149. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Freyberg MA, Kaiser D, Graf R, Vischer P
and Friedl P: Integrin-associated protein and thrombospondin-1 as
endothelial mechanosensitive death mediators. Biochem Biophys Res
Commun. 271:584–588. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
McDonald JF, Dimitry JM and Frazier WA: An
amyloid-like C-terminal domain of thrombospondin-1 displays CD47
agonist activity requiring both VVM motifs. Biochemistry.
42:10001–10011. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rath GM, Schneider C, Dedieu S, Rothhut B,
Soula-Rothhut M, Ghoneim C, Sid B, Morjani H, El Btaouri H and
Martiny L: The C-terminal CD47/IAP-binding domain of
thrombospondin-1 prevents camptothecin- and doxorubicin-induced
apoptosis in human thyroid carcinoma cells. Biochim Biophys Acta.
1763:1125–1134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pimanda JE, Annis DS, Raftery M, Mosher
DF, Chesterman CN and Hogg PJ: The von Willebrand factor-reducing
activity of thrombospondin-1 is located in the
calcium-binding/C-terminal sequence and requires a free thiol at
position 974. Blood. 100:2832–2838. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Schultz-Cherry S, Chen H, Mosher DF,
Misenheimer TM, Krutzsch HC, Roberts DD and Murphy-Ullrich JE:
Regulation of transforming growth factor-beta activation by
discrete sequences of thrombospondin 1. J Biol Chem. 270:7304–7310.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen H, Sottile J, Strickland DK and
Mosher DF: Binding and degradation of thrombospondin-1 mediated
through heparan sulphate proteoglycans and low-density-lipoprotein
receptor-related protein: Localization of the functional activity
to the trimeric N-terminal heparin-binding region of
thrombospondin-1. Biochem J. 318:959–963. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Befekadu R, Christiansen K, Larsson A and
Grenegard M: Increased plasma cathepsin S and trombospondin-1 in
patients with acute ST segment elevation myocardial infarction.
Cardiol J. 26:385–393. 2019. View Article : Google Scholar
|
|
48
|
Kaiser R, Grotemeyer K, Kalsch T, Graber
S, Wilkens H and Elmas E: Decreased TSP-1 following percutaneous
coronary intervention is associated with major adverse cardiac
events in ST-elevation myocardial infarction. Clin Hemorheol
Microcirc. 54:59–73. 2013. View Article : Google Scholar
|
|
49
|
Sezaki S, Hirohata S, Iwabu A, Nakamura K,
Toeda K, Miyoshi T, Yamawaki H, Demircan K, Kusachi S, Shiratori Y,
et al: Thrombospondin-1 is induced in rat myocardial infarction and
its induction is accelerated by ischemia/reperfusion. Exp Biol Med
(Maywood). 230:621–630. 2005. View Article : Google Scholar
|
|
50
|
Frangogiannis NG, Ren G, Dewald O, Zymek
P, Haudek S, Koerting A, Winkelmann K, Michael LH, Lawler J and
Entman ML: Critical role of endogenous thrombospondin-1 in
preventing expansion of healing myocardial infarcts. Circulation.
111:2935–2942. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jugdutt BI: Ventricular remodeling after
infarction and the extracellular collagen matrix: When is enough
enough? Circulation. 108:1395–1403. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
van Oorschot AA, Smits AM, Pardali E,
Doevendans PA and Goumans MJ: Low oxygen tension positively
influences cardiomyocyte progenitor cell function. J Cell Mol Med.
15:2723–2734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ashokkumar M, Anbarasan C, Saibabu R,
Kuram S, Raman SC and Cherian KM: An association study of
thrombospondin 1 and 2 SNPs with coronary artery disease and
myocardial infarction among South Indians. Thromb Res. 128:e49–e53.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Koch W, Hoppmann P, de Waha A, Schomig A
and Kastrati A: Polymorphisms in thrombospondin genes and
myocardial infarction: A case-control study and a meta-analysis of
available evidence. Hum Mol Genet. 17:1120–1126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhang XJ, Wei CY, Li WB, Zhang LL, Zhou Y,
Wang ZH, Tang MX, Zhang W, Zhang Y and Zhong M: Association between
single nucleotide polymorphisms in thrombospondins genes and
coronary artery disease: A meta-analysis. Thromb Res. 136:45–51.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zwicker JI, Peyvandi F, Palla R, Lombardi
R, Canciani MT, Cairo A, Ardissino D, Bernardinelli L, Bauer KA,
Lawler J and Mannucci P: The thrombospondin-1 N700S polymorphism is
associated with early myocardial infarction without altering von
Willebrand factor multimer size. Blood. 108:1280–1283. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhou X, Huang J, Chen J, Zhao J, Ge D,
Yang W and Gu D: Genetic association analysis of myocardial
infarction with thrombospondin-1 N700S variant in a Chinese
population. Thromb Res. 113:181–186. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Abdelmonem NA, Turky NO, Hashad IM, Abdel
Rahman MF, El-Etriby A and Gad MZ: Association of thrombospondin-1
(N700S) and thrombospondin-4 (A387P) gene polymorphisms with the
incidence of acute myocardial infarction in egyptians. Curr Pharm
Biotechnol. 18:1078–1087. 2017. View Article : Google Scholar
|
|
59
|
Stenina OI, Ustinov V, Krukovets I,
Marinic T, Topol EJ and Plow EF: Polymorphisms A387P in
thrombospondin-4 and N700S in thrombospondin-1 perturb calcium
binding sites. FASEB J. 19:1893–1895. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Xia Y, Dobaczewski M, Gonzalez-Quesada C,
Chen W, Biernacka A, Li N, Lee DW and Frangogiannis NG: Endogenous
thrombospondin 1 protects the pressure-overloaded myocardium by
modulating fibroblast phenotype and matrix metabolism.
Hypertension. 58:902–911. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gonzalez-Quesada C, Cavalera M, Biernacka
A, Kong P, Lee DW, Saxena A, Frunza O, Dobaczewski M, Shinde A and
Frangogiannis NG: Thrombospondin-1 induction in the diabetic
myocardium stabilizes the cardiac matrix in addition to promoting
vascular rarefaction through angiopoietin-2 upregulation. Circ Res.
113:1331–1344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Swinnen M, Vanhoutte D, Van Almen GC,
Hamdani N, Schellings MW, D’hooge J, Van der Velden J, Weaver MS,
Sage EH, Bornstein P, et al: Absence of thrombospondin-2 causes
age-related dilated cardiomyopathy. Circulation. 120:1585–1597.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
van Almen GC, Swinnen M, Carai P, Verhesen
W, Cleutjens JP, D’hooge J, Verheyen FK, Pinto YM, Schroen B,
Carmeliet P and Heymans S: Absence of thrombospondin-2 increases
cardiomyocyte damage and matrix disruption in doxorubicin-induced
cardiomyopathy. J Mol Cell Cardiol. 51:318–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Papageorgiou AP, Swinnen M, Vanhoutte D,
VandenDriessche T, Chuah M, Lindner D, Verhesen W, de Vries B,
D’hooge J, Lutgens E, et al: Thrombospondin-2 prevents cardiac
injury and dysfunction in viral myocarditis through the activation
of regulatory T-cells. Cardiovasc Res. 94:115–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Schroen B, Heymans S, Sharma U,
Blankesteijn WM, Pokharel S, Cleutjens JP, Porter JG, Evelo CT,
Duisters R, van Leeuwen RE, et al: Thrombospondin-2 is essential
for myocardial matrix integrity: Increased expression identifies
failure-prone cardiac hypertrophy. Circ Res. 95:515–522. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Batlle M, Perez-Villa F, Lazaro A,
García-Pras E, Vallejos I, Sionis A, Castel MA and Roig E:
Decreased expression of thrombospondin-1 in failing hearts may
favor ventricular remodeling. Transplant Proc. 41:2231–2233. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Vila V, Martinez-Sales V, Almenar L,
Lazaro IS, Villa P and Reganon E: Inflammation, endothelial
dysfunction and angiogenesis markers in chronic heart failure
patients. Int J Cardiol. 130:276–277. 2008. View Article : Google Scholar
|
|
68
|
Sharifi-Sanjani M, Shoushtari AH, Quiroz
M, Baust J, Sestito SF, Mosher M, Ross M, McTiernan CF, St Croix
CM, Bilonick RA, et al: Cardiac CD47 drives left ventricular heart
failure through Ca2+-CaMKII-regulated induction of
HDAC3. J Am Heart Assoc. 3:e0006702014. View Article : Google Scholar
|
|
69
|
van Almen GC, Verhesen W, van Leeuwen RE,
van de Vrie M, Eurlings C, Schellings MW, Swinnen M, Cleutjens JP,
van Zandvoort MA, Heymans S and Schroen B: MicroRNA-18 and
microRNA-19 regulate CTGF and TSP-1 expression in age-related heart
failure. Aging Cell. 10:769–779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Vila V, Sales VM, Almenar L, Lazaro IS,
Villa P and Reganon E: Effect of oral anticoagulant therapy on
thrombospondin-1 and von Willebrand factor in patients with stable
heart failure. Thromb Res. 121:611–615. 2008. View Article : Google Scholar
|
|
71
|
Berezin AE, Kremzer AA and Samura TA:
Circulating thrombospondine-2 in patients with moderate-to-severe
chronic heart failure due to coronary artery disease. J Biomed Res.
Mar 2–2015.[Epub ahead of print]. PubMed/NCBI
|
|
72
|
Kimura Y, Izumiya Y, Hanatani S, Yamamoto
E, Kusaka H, Tokitsu T, Takashio S, Sakamoto K, Tsujita K, Tanaka
T, et al: High serum levels of thrombospondin-2 correlate with poor
prognosis of patients with heart failure with preserved ejection
fraction. Heart Vessels. 31:52–59. 2016. View Article : Google Scholar
|
|
73
|
Hanatani S, Izumiya Y, Takashio S, Kimura
Y, Araki S, Rokutanda T, Tsujita K, Yamamoto E, Tanaka T, Yamamuro
M, et al: Circulating thrombospondin-2 reflects disease severity
and predicts outcome of heart failure with reduced ejection
fraction. Circ J. 78:903–910. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Freeman RV and Otto CM: Spectrum of
calcific aortic valve disease: Pathogenesis, disease progression,
and treatment strategies. Circulation. 111:3316–3326. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Pohjolainen V, Mustonen E, Taskinen P,
Näpänkangas J, Leskinen H, Ohukainen P, Peltonen T, Aro J, Juvonen
T, Satta J, et al: Increased thrombospondin-2 in human
fibrosclerotic and stenotic aortic valves. Atherosclerosis.
220:66–71. 2012. View Article : Google Scholar
|
|
76
|
Jurk K, Ritter MA, Schriek C, Van Aken H,
Droste DW, Ringelstein EB and Kehrel BE: Activated monocytes
capture platelets for heterotypic association in patients with
severe carotid artery stenosis. Thromb Haemost. 103:1193–1202.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Desai P, Helkin A, Odugbesi A, Stein J,
Bruch D, Lawler J, Maier KG and Gahtan V: Fluvastatin inhibits
intimal hyperplasia in wild-type but not Thbs1-null mice. J Surg
Res. 210:1–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liauw J, Hoang S, Choi M, Eroglu C, Choi
M, Sun GH, Percy M, Wildman-Tobriner B, Bliss T, Guzman RG, et al:
Thrombospondins 1 and 2 are necessary for synaptic plasticity and
functional recovery after stroke. J Cereb Blood Flow Metab.
28:1722–1732. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Lin TN, Kim GM, Chen JJ, Cheung WM, He YY
and Hsu CY: Differential regulation of thrombospondin-1 and
thrombospondin-2 after focal cerebral ischemia/reperfusion. Stroke.
34:177–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yang AL, Zhou HJ, Lin Y, Luo JK, Cui HJ,
Tang T and Yang QD: Thrombin promotes the expression of
thrombospondin-1 and -2 in a rat model of intracerebral hemorrhage.
J Neurol Sci. 323:141–146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhou HJ, Zhang HN, Tang T, Zhong JH, Qi Y,
Luo JK, Lin Y, Yang QD and Li XQ: Alteration of thrombospondin-1
and -2 in rat brains following experimental intracerebral
hemorrhage. Laboratory investigation. J Neurosurg. 113:820–825.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Bodewes TC, Johnson JM, Auster M, Huynh C,
Muralidharan S, Contreras M, LoGerfo FW and Pradhan-Nabzdyk L:
Intraluminal delivery of thrombospondin-2 small interfering RNA
inhibits the vascular response to injury in a rat carotid balloon
angioplasty model. FASEB J. 31:109–119. 2017. View Article : Google Scholar
|
|
83
|
Woo MS, Yang J, Beltran C and Cho S: Cell
surface CD36 protein in monocyte/macrophage contributes to
phagocytosis during the resolution phase of ischemic stroke in
mice. J Biol Chem. 291:23654–23661. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Kim CW, Pokutta-Paskaleva A, Kumar S,
Timmins LH, Morris AD, Kang DW, Dalal S, Chadid T, Kuo KM, Raykin
J, et al: Disturbed flow promotes arterial stiffening through
thrombospondin-1. Circulation. 136:1217–1232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Moura R, Tjwa M, Vandervoort P, Van
Kerckhoven S, Holvoet P and Hoylaerts MF: Thrombospondin-1
deficiency accelerates atherosclerotic plaque maturation in
ApoE−/− mice. Circ Res. 103:1181–1189. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Narizhneva NV, Razorenova OV, Podrez EA,
Chen J, Chandrasekharan UM, DiCorleto PE, Plow EF, Topol EJ and
Byzova TV: Thrombospondin-1 up-regulates expression of cell
adhesion molecules and promotes monocyte binding to endothelium.
FASEB J. 19:1158–1160. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Roth JJ, Gahtan V, Brown JL, Gerhard C,
Swami VK, Rothman VL, Tulenko TN and Tuszynski GP: Thrombospondin-1
is elevated with both intimal hyperplasia and hypercholesterolemia.
J Surg Res. 74:11–16. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Muraishi A, Capuzzi DM and Tuszynski GP:
Binding of thrombospondin to human plasma lipoproteins. Biochem
Biophys Res Commun. 193:1145–1151. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Barillari G, Iovane A, Bonuglia M,
Albonici L, Garofano P, Di Campli E, Falchi M, Condò I, Manzari V
and Ensoli B: Fibroblast growth factor-2 transiently activates the
p53 oncosuppressor protein in human primary vascular smooth muscle
cells: Implications for atherogenesis. Atherosclerosis.
210:400–406. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Osada-Oka M, Ikeda T, Akiba S and Sato T:
Hypoxia stimulates the autocrine regulation of migration of
vascular smooth muscle cells via HIF-1alpha-dependent expression of
thrombospondin-1. J Cell Biochem. 104:1918–1926. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Takahashi M, Oka M, Ikeda T, Akiba S and
Sato T: Role of thrombospondin-1 in hypoxia-induced migration of
human vascular smooth muscle cells. Yakugaku Zasshi. 128:377–383.
2008.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Yabkowitz R, Mansfield PJ, Ryan US and
Suchard SJ: Thrombospondin mediates migration and potentiates
platelet-derived growth factor-dependent migration of calf
pulmonary artery smooth muscle cells. J Cell Physiol. 157:24–32.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Raman P, Krukovets I, Marinic TE,
Bornstein P and Stenina OI: Glycosylation mediates up-regulation of
a potent antiangiogenic and proatherogenic protein,
thrombospondin-1, by glucose in vascular smooth muscle cells. J
Biol Chem. 282:5704–5714. 2007. View Article : Google Scholar
|
|
94
|
Stenina OI, Krukovets I, Wang K, Zhou Z,
Forudi F, Penn MS, Topol EJ and Plow EF: Increased expression of
thrombospondin-1 in vessel wall of diabetic Zucker rat.
Circulation. 107:3209–3215. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Ganguly R, Sahu S, Ohanyan V, Haney R,
Chavez RJ, Shah S, Yalamanchili S and Raman P: Oral chromium
picolinate impedes hyperglycemia-induced atherosclerosis and
inhibits proatherogenic protein TSP-1 expression in STZ-induced
type 1 diabetic ApoE−/− mice. Sci Rep. 7:452792017.
View Article : Google Scholar
|
|
96
|
Yang YJ, Cheng DY, Zheng XW, Li F and Yang
GL: Expression of thrombospondin-1 in the lung of hypoxic pulmonary
hypertension rats. Sichuan Da Xue Xue Bao Yi Xue Ban. 43:19–23.
2012.(In Chinese). PubMed/NCBI
|
|
97
|
Reed MJ, Iruela-Arispe L, O’Brien ER,
Truong T, LaBell T, Bornstein P and Sage EH: Expression of
thrombospondins by endothelial cells. Injury is correlated with
TSP-1. Am J Pathol. 147:1068–1080. 1995.PubMed/NCBI
|
|
98
|
DiPietro LA, Nebgen DR and Polverini PJ:
Downregulation of endothelial cell thrombospondin 1 enhances in
vitro angiogenesis. J Vasc Res. 31:178–185. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Dardik R, Solomon A, Loscalzo J, Eskaraev
R, Bialik A, Goldberg I, Schiby G and Inbal A: Novel proangiogenic
effect of factor XIII associated with suppression of thrombospondin
1 expression. Arterioscler Thromb Vasc Biol. 23:1472–1477. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Isenberg JS, Hyodo F, Ridnour LA, Shannon
CS, Wink DA, Krishna MC and Roberts DD: Thrombospondin 1 and
vasoactive agents indirectly alter tumor blood flow. Neoplasia.
10:886–896. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Tzeng HT, Tsai CH, Yen YT, Cheng HC, Chen
YC, Pu SW, Wang YS, Shan YS, Tseng YL, Su WC, et al: Dysregulation
of Rab37-mediated cross-talk between cancer cells and endothelial
cells via thrombospondin-1 promotes tumor neovasculature and
metastasis. Clin Cancer Res. 23:2335–2345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Bitar MS: Diabetes impairs angiogenesis
and induces endothelial cell senescence by up-regulating
thrombospondin-CD47-dependent signaling. Int J Mol Sci.
20:E6732019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yafai Y, Eichler W, Iandiev I, Unterlauft
JD, Jochmann C, Wiedemann P and Bringmann A: Thrombospondin-1 is
produced by retinal glial cells and inhibits the growth of vascular
endothelial cells. Ophthalmic Res. 52:81–88. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang S, Sorenson CM and Sheibani N: Lack
of thrombospondin 1 and exacerbation of choroidal
neovascularization. Arch Ophthalmol. 130:615–620. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Wu Z, Wang S, Sorenson CM and Sheibani N:
Attenuation of retinal vascular development and neovascularization
in transgenic mice over-expressing thrombospondin-1 in the lens.
Dev Dyn. 235:1908–1920. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wang Y, Wang S and Sheibani N: Enhanced
proangiogenic signaling in thrombospondin-1-deficient retinal
endothelial cells. Microvasc Res. 71:143–151. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Wang S, Wu Z, Sorenson CM, Lawler J and
Sheibani N: Thrombospondin-1-deficient mice exhibit increased
vascular density during retinal vascular development and are less
sensitive to hyperoxia-mediated vessel obliteration. Dev Dyn.
228:630–642. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Sheibani N, Sorenson CM, Cornelius LA and
Frazier WA: Thrombospondin-1, a natural inhibitor of angiogenesis,
is present in vitreous and aqueous humor and is modulated by
hyperglycemia. Biochem Biophys Res Commun. 267:257–261. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Koch M, Hussein F, Woeste A, Gründker C,
Frontzek K, Emons G and Hawighorst T: CD36-mediated activation of
endothelial cell apoptosis by an N-terminal recombinant fragment of
thrombospondin-2 inhibits breast cancer growth and metastasis in
vivo. Breast Cancer Res Treat. 128:337–346. 2011. View Article : Google Scholar
|
|
110
|
Tomii Y, Kamochi J, Yamazaki H, Sawa N,
Tokunaga T, Ohnishi Y, Kijima H, Ueyama Y, Tamaoki N and Nakamura
M: Human thrombospondin 2 inhibits proliferation of microvascular
endothelial cells. Int J Oncol. 20:339–342. 2002.PubMed/NCBI
|
|
111
|
Armstrong LC, Bjorkblom B, Hankenson KD,
Siadak AW, Stiles CE and Bornstein P: Thrombospondin 2 inhibits
microvascular endothelial cell proliferation by a
caspase-independent mechanism. Mol Biol Cell. 13:1893–1905. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Kyriakides TR, Zhu YH, Yang Z, Huynh G and
Bornstein P: Altered extracellular matrix remodeling and
angiogenesis in sponge granulomas of thrombospondin 2-null mice. Am
J Pathol. 159:1255–1262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Calabro NE, Kristofik NJ and Kyriakides
TR: Thrombospondin-2 and extracellular matrix assembly. Biochim
Biophys Acta. 1840.2396–2402. 2014.
|
|
114
|
Krady MM, Zeng J, Yu J, MacLauchlan S,
Skokos EA, Tian W, Bornstein P, Sessa WC and Kyriakides TR:
Thrombospondin-2 modulates extracellular matrix remodeling during
physiological angiogenesis. Am J Pathol. 173:879–891. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Park YW, Kang YM, Butterfield J, Detmar M,
Goronzy JJ and Weyand CM: Thrombospondin 2 functions as an
endogenous regulator of angiogenesis and inflammation in rheumatoid
arthritis. Am J Pathol. 165:2087–2098. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Agah A, Kyriakides TR, Letrondo N,
Bjorkblom B and Bornstein P: Thrombospondin 2 levels are increased
in aged mice: Consequences for cutaneous wound healing and
angiogenesis. Matrix Biol. 22:539–547. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Feige JJ: Thrombospondins: Multimodular
proteins with angiostatic function. Pathol Biol (Paris).
47:339–344. 1999.(In French).
|
|
118
|
Raugi GJ, Mullen JS, Bark DH, Okada T and
Mayberg MR: Thrombospondin deposition in rat carotid artery injury.
Am J Pathol. 137:179–185. 1990.PubMed/NCBI
|
|
119
|
Lemkens P, Boari G, Fazzi G, Janssen G,
Murphy-Ullrich J, Schiffers P and De Mey J: Thrombospondin-1 in
early flow-related remodeling of mesenteric arteries from young
normotensive and spontaneously hypertensive rats. Open Cardiovasc
Med J. 6:50–59. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ichii T, Koyama H, Tanaka S, Shioi A,
Okuno Y, Otani S and Nishizawa Y: Thrombospondin-1 mediates smooth
muscle cell proliferation induced by interaction with human
platelets. Arterioscler Thromb Vasc Biol. 22:1286–1292. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Riessen R, Kearney M, Lawler J and Isner
JM: Immunolocalization of thrombospondin-1 in human atherosclerotic
and restenotic arteries. Am Heart J. 135:357–364. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Yoshida S, Nabzdyk CS, Pradhan L and
LoGerfo FW: Thrombospondin-2 gene silencing in human aortic smooth
muscle cells improves cell attachment. J Am Coll Surg. 213:668–676.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ochoa CD, Yu L, Al-Ansari E, Hales CA and
Quinn DA: Thrombospondin-1 null mice are resistant to
hypoxia-induced pulmonary hypertension. J Cardiothorac Surg.
5:322010. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Satoh M, Nasu T, Osaki T and Hitomi S:
Thrombospondin-1 contributes to slower aortic aneurysm growth by
inhibiting maladaptive remodeling of extracellular matrix. Clin Sci
(Lond). 131:1283–1285. 2017. View Article : Google Scholar
|
|
125
|
Liu Z, Morgan S, Ren J, Wang Q, Annis DS,
Mosher DF, Zhang J, Sorenson CM, Sheibani N and Liu B:
Thrombospondin-1 (TSP1) contributes to the development of vascular
inflammation by regulating monocytic cell motility in mouse models
of abdominal aortic aneurysm. Circ Res. 117:129–141. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Cointe S, Rheaume E, Martel C, Blanc-Brude
O, Dubé E, Sabatier F, Dignat-George F, Tardif JC and Bonnefoy A:
Thrombospondin-1-derived peptide RFYVVMWK improves the adhesive
phenotype of CD34(+) cells from atherosclerotic patients with type
2 diabetes. Cell Transplant. 26:327–337. 2017. View Article : Google Scholar :
|
|
127
|
Kyriakides TR, Rojnuckarin P, Reidy MA,
Hankenson KD, Papayannopoulou T, Kaushansky K and Bornstein P:
Megakaryocytes require thrombospondin-2 for normal platelet
formation and function. Blood. 101:3915–3923. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Kyriakides TR, Leach KJ, Hoffman AS,
Ratner BD and Bornstein P: Mice that lack the angiogenesis
inhibitor, thrombospondin 2, mount an altered foreign body reaction
characterized by increased vascularity. Proc Natl Acad Sci USA.
96:4449–4454. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Roberts DD, Haverstick DM, Dixit VM,
Frazier WA, Santoro SA and Ginsburg V: The platelet glycoprotein
thrombospondin binds specifically to sulfated glycolipids. J Biol
Chem. 260:9405–9411. 1985.PubMed/NCBI
|
|
130
|
Yang S, Song R, Li X, Zhang T, Fu J and
Cui X: Thrombospondin-2 predicts response to treatment with
intravenous immunoglobulin in children with Kawasaki disease. BMJ
Paediatr Open. 2:e0001902018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Reinecke H, Robey TE, Mignone JL, Muskheli
V, Bornstein P and Murry CE: Lack of thrombospondin-2 reduces
fibrosis and increases vascularity around cardiac cell grafts.
Cardiovasc Pathol. 22:91–95. 2013. View Article : Google Scholar
|
|
132
|
Roberts DD: Interactions of thrombospondin
with sulfated glycolipids and proteoglycans of human melanoma
cells. Cancer Res. 48:6785–6793. 1988.PubMed/NCBI
|
|
133
|
Orr AW, Pedraza CE, Pallero MA, Elzie CA,
Goicoechea S, Strickland DK and Murphy-Ullrich JE: Low density
lipoprotein receptor-related protein is a calreticulin coreceptor
that signals focal adhesion disassembly. J Cell Biol.
161:1179–1189. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Li Z, Calzada MJ, Sipes JM, Cashel JA,
Krutzsch HC, Annis DS, Mosher DF and Roberts DD: Interactions of
thrombospondins with alpha4beta1 integrin and CD47 differentially
modulate T cell behavior. J Cell Biol. 157:509–519. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Huang Z, Newcomb CJ, Lei Y, Zhou Y,
Bornstein P, Amendt BA, Stupp SI and Snead ML: Bioactive nanofibers
enable the identification of thrombospondin 2 as a key player in
enamel regeneration. Biomaterials. 61:216–228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Lawler J and Hynes RO: An integrin
receptor on normal and thrombasthenic platelets that binds
thrombospondin. Blood. 74:2022–2027. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Kong P, Cavalera M and Frangogiannis NG:
The role of thrombospondin (TSP)-1 in obesity and diabetes.
Adipocyte. 3:81–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Hogg PJ, Hotchkiss KA, Jimenez BM,
Stathakis P and Chesterman CN: Interaction of platelet-derived
growth factor with thrombospondin 1. Biochem J. 326:709–716. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Qian X, Wang TN, Rothman VL, Nicosia RF
and Tuszynski GP: Thrombospondin-1 modulates angiogenesis in vitro
by up-regulation of matrix metalloproteinase-9 in endothelial
cells. Exp Cell Res. 235:403–412. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Liu A, Garg P, Yang S, Gong P, Pallero MA,
Annis DS, Liu Y, Passaniti A, Mann D, Mosher DF, et al: Epidermal
growth factor-like repeats of thrombospondins activate
phospholipase Cgamma and increase epithelial cell migration through
indirect epidermal growth factor receptor activation. J Biol Chem.
284:6389–6402. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Zheng B and Clemmons DR: Blocking ligand
occupancy of the alphaVbeta3 integrin inhibits insulin-like growth
factor I signaling in vascular smooth muscle cells. Proc Natl Acad
Sci USA. 95:11217–11222. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Saumet A, Slimane MB, Lanotte M, Lawler J
and Dubernard V: Type 3 repeat/C-terminal domain of
thrombospondin-1 triggers caspase-independent cell death through
CD47/alphavbeta3 in promyelocytic leukemia NB4 cells. Blood.
106:658–667. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Isenberg JS, Jia Y, Fukuyama J, Switzer
CH, Wink DA and Roberts DD: Thrombospondin-1 inhibits nitric oxide
signaling via CD36 by inhibiting myristic acid uptake. J Biol Chem.
282:15404–15415. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Lagadec P, Dejoux O, Ticchioni M, Cottrez
F, Johansen M, Brown EJ and Bernard A: Involvement of a
CD47-dependent pathway in platelet adhesion on inflamed vascular
endothelium under flow. Blood. 101:4836–4843. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Mandler WK, Nurkiewicz TR, Porter DW,
Kelley EE and Olfert IM: Microvascular dysfunction following
multiwalled carbon nanotube exposure is mediated by
thrombospondin-1 receptor CD47. Toxicol Sci. 165:90–99. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Gao Q, Chen K, Gao L, Zheng Y and Yang YG:
Thrombospondin-1 signaling through CD47 inhibits cell cycle
progression and induces senescence in endothelial cells. Cell Death
Dis. 7:e23682016. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Yamauchi Y, Kuroki M, Imakiire T, Uno K,
Abe H, Beppu R, Yamashita Y, Kuroki M and Shirakusa T: Opposite
effects of thrombospondin-1 via CD36 and CD47 on homotypic
aggregation of monocytic cells. Matrix Biol. 21:441–448. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Kaur S, Chang T, Singh SP, Lim L, Mannan
P, Garfield SH, Pendrak ML, Soto-Pantoja DR, Rosenberg AZ, Jin S
and Roberts DD: CD47 signaling regulates the immunosuppressive
activity of VEGF in T cells. J Immunol. 193:3914–3924. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Rogers NM, Roberts DD and Isenberg JS:
Age-associated induction of cell membrane CD47 limits basal and
temperature-induced changes in cutaneous blood flow. Ann Surg.
258:184–191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Rogers NM, Sharifi-Sanjani M, Csanyi G,
Pagano PJ and Isenberg JS: Thrombospondin-1 and CD47 regulation of
cardiac, pulmonary and vascular responses in health and disease.
Matrix Biol. 37:92–101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Csanyi G, Feck DM, Ghoshal P, Singla B,
Lin H, Nagarajan S, Meijles DN, Al Ghouleh I, Cantu-Medellin N,
Kelley EE, et al: CD47 and Nox1 mediate dynamic fluid-phase
macropinocytosis of native LDL. Antioxid Redox Signal. 26:886–901.
2017. View Article : Google Scholar :
|
|
152
|
Rogers NM, Sharifi-Sanjani M, Yao M,
Ghimire K, Bienes-Martinez R, Mutchler SM, Knupp HE, Baust J,
Novelli EM, Ross M, et al: TSP1-CD47 signaling is upregulated in
clinical pulmonary hypertension and contributes to pulmonary
arterial vasculopathy and dysfunction. Cardiovasc Res. 113:15–29.
2017. View Article : Google Scholar
|
|
153
|
Kellouche S, Mourah S, Bonnefoy A,
Schoëvaert D, Podgorniak MP, Calvo F, Hoylaerts MF, Legrand C and
Dosquet C: Platelets, thrombospondin-1 and human dermal fibroblasts
cooperate for stimulation of endothelial cell tubulogenesis through
VEGF and PAI-1 regulation. Exp Cell Res. 313:486–499. 2007.
View Article : Google Scholar
|
|
154
|
Murphy-Ullrich JE and Poczatek M:
Activation of latent TGF-beta by thrombospondin-1: Mechanisms and
physiology. Cytokine Growth Factor Rev. 11:59–69. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Schultz-Cherry S, Lawler J and
Murphy-Ullrich JE: The type 1 repeats of thrombospondin 1 activate
latent transforming growth factor-beta. J Biol Chem.
269:26783–26788. 1994.PubMed/NCBI
|
|
156
|
Rosini S, Pugh N, Bonna AM, Hulmes DJS,
Farndale RW and Adams JC: Thrombospondin-1 promotes matrix
homeostasis by interacting with collagen and lysyl oxidase
precursors and collagen cross-linking sites. Sci Signal.
11:eaar25662018. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Yang Z, Strickland DK and Bornstein P:
Extracellular matrix metalloproteinase 2 levels are regulated by
the low density lipoprotein-related scavenger receptor and
thrombospondin 2. J Biol Chem. 276:8403–8408. 2001. View Article : Google Scholar
|
|
158
|
Nakamura M, Oida Y, Abe Y, Yamazaki H,
Mukai M, Matsuyama M, Chijiwa T, Matsumoto H and Ueyama Y:
Thrombospondin-2 inhibits tumor cell invasion through the
modulation of MMP-9 and uPA in pancreatic cancer cells. Mol Med
Rep. 1:423–427. 2008.PubMed/NCBI
|
|
159
|
Murphy-Ullrich JE and Mosher DF:
Interactions of thrombospondin with endothelial cells:
Receptor-mediated binding and degradation. J Cell Biol.
105:1603–1611. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Godyna S, Liau G, Popa I, Stefansson S and
Argraves WS: Identification of the low density lipoprotein
receptor-related protein (LRP) as an endocytic receptor for
thrombospondin-1. J Cell Biol. 129:1403–1410. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Mikhailenko I, Kounnas MZ and Strickland
DK: Low density lipoprotein receptor-related protein/alpha
2-macroglobulin receptor mediates the cellular internalization and
degradation of thrombospondin. A process facilitated by
cell-surface proteoglycans. J Biol Chem. 270:9543–9549. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Daviet L and McGregor JL: Vascular biology
of CD36: Roles of this new adhesion molecule family in different
disease states. Thromb Haemost. 78:65–69. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Ramanathan S, Mazzalupo S, Boitano S and
Montfort WR: Thrombospondin-1 and angiotensin II inhibit soluble
guanylyl cyclase through an increase in intracellular calcium
concentration. Biochemistry. 50:7787–7799. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Trujillo G and Kew RR: Platelet-derived
thrombospondin-1 is necessary for the vitamin D-binding protein
(Gc-globulin) to function as a chemotactic cofactor for C5a. J
Immunol. 173:4130–4136. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Tang Y, Scheef EA, Wang S, Sorenson CM,
Marcus CB, Jefcoate CR and Sheibani N: CYP1B1 expression promotes
the proangiogenic phenotype of endothelium through decreased
intracellular oxidative stress and thrombospondin-2 expression.
Blood. 113:744–754. 2009. View Article : Google Scholar :
|
|
166
|
MacLauchlan S, Yu J, Parrish M, Asoulin
TA, Schleicher M, Krady MM, Zeng J, Huang PL, Sessa WC and
Kyriakides TR: Endothelial nitric oxide synthase controls the
expression of the angiogenesis inhibitor thrombospondin 2. Proc
Natl Acad Sci USA. 108:E1137–E1145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Lee NV, Sato M, Annis DS, Loo JA, Wu L,
Mosher DF and Iruela-Arispe ML: ADAMTS1 mediates the release of
antiangiogenic polypeptides from TSP1 and 2. EMBO J. 25:5270–5283.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Mead TJ and Apte SS: ADAMTS proteins in
human disorders. Matrix Biol. 71–72:225–239. 2018. View Article : Google Scholar
|
|
169
|
Bengtsson E, Hultman K, Duner P, Asciutto
G, Almgren P, Orho-Melander M, Melander O, Nilsson J,
Hultgardh-Nilsson A and Gonçalves I: ADAMTS-7 is associated with a
high-risk plaque phenotype in human atherosclerosis. Sci Rep.
7:37532017. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Pereira A, Palma Dos Reis R, Rodrigues R,
Sousa AC, Gomes S, Borges S, Ornelas I, Freitas AI, Guerra G,
Henriques E, et al: Association of ADAMTS7 gene polymorphism with
cardiovascular survival in coronary artery disease. Physiol
Genomics. 48:810–815. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Bauer RC, Tohyama J, Cui J, Cheng L, Yang
J, Zhang X, Ou K, Paschos GK, Zheng XL, Parmacek MS, et al:
Knockout of Adamts7, a novel coronary artery disease locus in
humans, reduces atherosclerosis in mice. Circulation.
131:1202–1213. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Reilly MP, Li M, He J, Ferguson JF,
Stylianou IM, Mehta NN, Burnett MS, Devaney JM, Knouff CW, Thompson
JR, et al: Identification of ADAMTS7 as a novel locus for coronary
atherosclerosis and association of ABO with myocardial infarction
in the presence of coronary atherosclerosis: Two genome-wide
association studies. Lancet. 377:383–392. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Qin W, Cao Y, Li L, Chen W and Chen X:
Upregulation of ADAMTS7 and downregulation of COMP are associated
with aortic aneurysm. Mol Med Rep. 16:5459–5463. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Kessler T, Zhang L, Liu Z, Yin X, Huang Y,
Wang Y, Fu Y, Mayr M, Ge Q, Xu Q, et al: ADAMTS-7 inhibits
re-endothelialization of injured arteries and promotes vascular
remodeling through cleavage of thrombospondin-1. Circulation.
131:1191–1201. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Pu X, Xiao Q, Kiechl S, Chan K, Ng FL, Gor
S, Poston RN, Fang C, Patel A, Senver EC, et al: ADAMTS7 cleavage
and vascular smooth muscle cell migration is affected by a
coronary-artery-disease-associated variant. Am J Hum Genet.
92:366–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Wang L, Zheng J, Bai X, Liu B, Liu CJ, Xu
Q, Zhu Y, Wang N, Kong W and Wang X: ADAMTS-7 mediates vascular
smooth muscle cell migration and neointima formation in
balloon-injured rat arteries. Circ Res. 104:688–698. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Wu W, Li J, Yu C, Gao Y, Fan S, Ye X, Wang
Y and Zheng J: Association of serum ADAMTS-7 levels with left
ventricular reverse remodeling after ST-elevation myocardial
infarction. Eur J Med Res. 23:152018. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Chan K, Pu X, Sandesara P, Poston RN,
Simpson IA, Quyyumi AA, Ye S and Patel RS: Genetic variation at the
ADAMTS7 locus is associated with reduced severity of coronary
artery disease. J Am Heart Assoc. 6:e0069282017. View Article : Google Scholar :
|
|
179
|
Wu W, Wang H, Yu C, Li J, Gao Y, Ke Y,
Wang Y, Zhou Y and Zheng J: Association of ADAMTS-7 levels with
cardiac function in a rat model of acute myocardial infarction.
Cell Physiol Biochem. 38:950–958. 2016. View Article : Google Scholar : PubMed/NCBI
|