Introduction
Acute myocardial infarction (AMI) is a major cause
of mortality worldwide and the incidence of AMI has increased in
China in recent years. Revascularization to restore blood flow is
currently the most effective treatment for AMI (1). However, while early reperfusion is
effective in decreasing mortality, the perfusion of previously
ischemic myocardial tissue results in a second wave of injury,
which is referred to as myocardial ischemia-reperfusion injury
(MIRI) (2,3). MIRI may lead to severe adverse
events, including acute left ventricular failure and malignant
arrhythmia (4). Therefore,
preventing MIRI in the early stages of AMI may improve patient
prognosis.
The mitochondrial permeability transition pore
(MPTP), which is located in the inner membrane, plays an important
role in apoptosis (5). Following
ischemia, reperfusion may result in the generation of reactive
oxygen species and Ca2+ influx, which may lead to the
opening of the MPTP (6,7). The opening of the MPTP, which is
known as the mitochondrial permeability transition, decreases the
inner mitochondrial membrane potential (ΔΨm). It also leads to the
release of cytochrome c into the cytoplasm and activates
caspase, resulting in mitochondrial dysfunction and swelling,
eventually leading to apoptosis (8–11).
Previous studies have demonstrated that the inhibition of the
opening of the MPTP with an inhibitor, such as cyclosporin A, may
alleviate MIRI (12,13), while promoting MPTP opening, with
for example, atractyloside, may aggravate MIRI (14). Therefore, the MPTP may serve as an
important target for modulating MIRI.
Certain natural compounds are known to exert
pharmacological actions and are generally well-tolerated by the
human body. Curculigoside, extracted from Curculigo
orchioides Gaertn, is a phenolic glycoside antioxidant that
exerts anti-inflammatory and antitumor effects (15–17). Curculigoside has been demonstrated
to inhibit H2O2-induced oxidative stress in
human umbilical vein endothelial cells (18). Furthermore, several studies have
reported that curculigoside attenuates experimental cerebral
ischemia injury in vitro and ex vivo (19–21). However, whether curculigoside
exerts a cardioprotective effect following I/R injury remains
unclear.
The aim of the present study was to determine
whether curculigoside exerts cardioprotective effects following I/R
or H/R injury in vitro and ex vivo and to investigate
whether the underlying mechanisms involve the inhibition of MPTP
opening.
Materials and methods
Drugs
Curculigoside (CAS no. 85643-19-2; formula,
C22H26O11; the chemical structure
of curculigoside can be found at: http://www.chemfaces.com/natural/Curculigoside-CFN97419.html)
and atractyloside were purchased from ChemFaces® and
Nantong Feiyu Biological Technology Co. Ltd., respectively. Both
drugs were dissolved in 1% dimethyl sulfoxide (DMSO; Solarbio
Science & Technology Co.) and stored at −20°C until further
use.
Cell culture
H9c2 rat cardiomyocytes were purchased from the Cell
Bank of the Chinese Academy of Sciences. Cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
fetal bovine serum and 1% penicillin/streptomycin at 37°C in a
humidified atmosphere containing 5% CO2.
Induction of the H/R model in vitro
Upon reaching 80–90% confluence, the H9c2 cells were
pre-treated with curculigoside (5, 10 and 15 μM) or the equivalent
volume of 1% DMSO for 8 h. The spent DMEM was replaced with
glucose- and FBS-free Earle’s medium in all groups, except for the
control group. Cells were subsequently incubated in a tri-gas
chamber containing 95% (v/v) N2, 5% (v/v) CO2
and 5% O2 at 37°C for 12 h to induce hypoxia. Earle’s
medium was replaced with DMEM supplemented with 10% FBS and the
cells were incubated at 37°C and 5% CO2 for 1 h to allow
reoxygenation.
Cell experiment protocol
H9c2 cells were divided into 6 groups as follows: i)
The control group; ii) the hypoxia/reoxygenation group (H/R); iii)
the vehicle group; iv) the 5 μM curculigoside-treated group; v) the
10 μM curculigoside-treated group; and vi) the 15 μM
curculigoside-treated group. Based on the results obtained,
curculigoside was used at a concentration of 10 μM in subsequent
experiments. H9c2 cells were subsequently divided into 5 groups as
follows: i) The control group; ii) the H/R group; iii) the vehicle
group; iv) the 10 μM curculigoside-treated group; v) the combined
treatment group (10 μM curculigoside and 20 μM atractyloside). The
curculigoside-treated and combined treatment groups were treated
with curculigoside and curculigoside and atractyloside,
respectively, for 8 h prior to H/R.
Assessment of cell proliferation
The proliferation of the H9c2 cells was assessed
using a Cell Counting kit-8 (BD Biosciences) assay, according to
the manufacturer’s protocol. The H9c2 cells (3×103
cells/well) were cultured in 96-well plates. Following treatment,
10 μl CCK-8 solution were added to each well and incubated for 1 h
at 37°C in a 5% CO2 incubator. The optical density was
measured at a wavelength of 490 nm using a microplate reader
(Biotek Instruments, Inc.).
Assessment of lactate dehydrogenase (LDH)
activity
LDH activity was assessed to evaluate the
cytoprotective effects of curculigoside. An LDH assay (Nanjing
Jiancheng Bioengineering Institute) was performed according to the
manufacturer’s instructions. The optical density was measured at a
wavelength of 450 nm using a microplate reader (Biotek Instruments,
Inc.) and was used to calculate the concentration of LDH.
Assessment of H9c2 cell apoptosis
The apoptotic rate of the H9c2 cells was evaluated
by flow cytometry using an Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) kit (BD Biosciences). Cells in 6-well
plates were trypsinized and washed twice in ice-cold PBS. Cells
were suspended in binding buffer and 5 μl Annexin V-FITC and 5 μl
PI were added to each sample. The cells were incubated at room
temperature in the dark for 15 min and analyzed using a flow
cytometer (BD Biosciences).
Assessment of ΔΨm
The change in ΔΨm was measured by rhodamine 123
staining following the manufacturer’s protocol (Beyotime Institute
of Biotechnology). Cell apoptosis leads to a decrease in ΔΨm and a
reduction in the fluorescence intensity in cells. The H9c2 cells
were incubated with rhodamine 123 for 30 min at 37°C in the dark.
Cells were visualized by fluorescence microscopy (IX73, Olympus)
and the staining was analyzed using Image-Pro software.
Assessment of MPTP opening
MPTP opening was determined using the
CaCl2-induced method (22). Mitochondria were isolated from the
H9c2 cells using the Mitochondrial Extract kit (BestBio Co.),
according to the manufacturer’s instructions. The sensitivity of
MPTP opening to Ca2+ was determined using the Purified
Mitochondrial Membrane Pore Channel Colorimetric Assay kit
(Shanghai Genmed Pharmaceutical Technology Co., Ltd.), according to
the manufacturer’s protocol.
Animals and animal care
Male Wistar rats (age, 8 weeks; weight, 300±30 g)
were purchased from Liaoning Changsheng Biotechnology Co. and
housed in environmentally-controlled conditions (temperature,
22±2°C; relative humidity, 60±5%; 12-h light/dark cycles). All rats
were allowed to acclimatize to their environment prior to the
experiments and fed a standard diet. Food and bedding were changed
for the rats, and the health of the animals was monitored every
other day. The present study was approved by the Ethics Committee
of China Medical University and procedures for animals handling and
care adhered to the Guide for the Care and Use of Laboratory
Animals (23).
Induction of the ischemia-reperfusion
group (I/R) model
Krebs-Henseleit (K-H) buffer (0.15 mol/l NaCl, 0.006
mol/l KCl, 0.002 mol/l CaCl2, 0.002 mol/l
NaHCO3 and 0.011 mol/l glucose) was prepared and the pH
was adjusted to 7.35–7.45 by the addition of NaOH. The rats were
anesthetized by an intraperitoneal injection of 30 mg/kg
pentobarbital sodium. An incision was made into the thoracic cavity
and the heart was rapidly removed and placed in ice-cold
heparinized K-H buffer with continuous oxygenation (oxygen flow
rate, 3 l/min). The heart connective tissue was resected to expose
the aorta, which was subsequently connected to a Langendorff
system. The isolated hearts were retrogradely perfused with K-H
buffer containing 95% O2 and 5% CO2 for 15
min at 37°C and at a constant pressure of 76 mmHg. Rat hearts were
subjected to global ischemia for 30 min, followed by 60 min of
reperfusion to generate the MIRI model as described in a previous
study by the authors (24). All
rats used in this study (total, 165 rats) were subjected to
euthanasia. As for the details of euthanasia, each of the rats was
anesthetized in an isolated room individually by intraperitoneal
injection of pentobarbital sodium at a dose of 30 mg/kg. After
confirming that the rat was completely anesthetized and unconscious
by determining whether the rats exhibited nerve reflexes or not,
the chest of the rat was opened, the aorta was disconnected and the
heart was rapidly isolated to induce the permanent cessation of
circulation. The experiment described herein is an acute,
non-survival experiment. Euthanasia was achieved by exsanguination
during the harvesting of the heart as previously described
(25).
Animal experiment protocol
The rats were randomly divided into 6 groups (n=15
rats per group) as follows: i) The control group; ii) the I/R
group; iii) the vehicle group; iv) the 5 mg/kg
curculigoside-treated group; v) the 10 mg/kg curculigoside-treated
group; and vi) the 15 mg/kg curculigoside-treated group. Prior to
isolation of the hearts, the rats in the curculigoside-treated
groups received a daily intraperitoneal injection of curculigoside
for 7 days. The animals in the vehicle group received the
equivalent volume of 1% DMSO.
Based on the results obtained, 10 mg/kg
curculigoside was selected for further experimentation. The rats
were randomly divided into 5 groups (n=15 rats per group) as
follows: i) The control group; ii) the I/R group; iii) the vehicle
group; iv) the 10 mg/kg curculigoside-treated group; v) the
combined treatment group (10 mg/kg curculigoside and 5 mg/kg
atractyloside). Similar to the curculigoside-treated group, the
animals in the combined treatment group received a daily
intraperitoneal injection of curculigoside and atractyloside for 7
days prior to surgery.
Assessment of infarct size
Triphenyltetrazolium chloride (TTC) staining was
performed to quantify the infarct size in myocardial tissues. The
heart was removed and the left ventricular tissue was frozen at
−70°C and subsequently cut into 2-mm-thick sections. The sections
were incubated with 1% TTC (Solarbio Science & Technology Co.)
at 37°C for 30 min at 37°C in the dark. Images of the staining were
acquired using a digital camera (EOS 90D, Canon) and the staining
was quantified by ImageJ2x software.
Assessment of myocardial morphological
changes
Hearts were removed from the Langendorff system
after 1 h of reperfusion and fixed in 4% paraformaldehyde overnight
at 4°C. Following dehydration with ethanol, the left ventricular
tissue was embedded in paraffin and cut into 5-μm-sections using a
rotary microtome. Following dewaxing and rehydration, the sections
were immersed in hematoxylin solution for 5 min, 1% hydrochloric
acid alcohol for 3 sec, and eosin solution for 3 min at room
temperature for staining. Myocardial morphological characteristics
were subsequently observed using a light microscope (CX23,
Olympus).
Assessment of myocardial apoptosis
Terminal deoxynucleotidyl-transferase-mediated dUTP
nick-end labeling (TUNEL) staining was performed to assess
myocardial apoptosis using the In Situ Cell Death Detection
kit (Roche Diagnostics), according to the manufacturer’s
instructions. TUNEL-positive cells were stained dark grey and were
visible under a light microscope (CX23, Olympus).
Assessment of protein expression
Total protein was extracted from myocardial tissue
and H9c2 cells using radioimmunoprecipitation assay buffer and
protease inhibitors. Total protein was quantified using a
bicinchoninic acid assay and 30 μg protein/lane was separated via
SDS-PAGE on 10% gels. The separated proteins were subsequently
transferred onto polyvinylidene difluoride membranes and blocked
for 1 h at room temperature with 5% skim milk dissolved in TBST.
The membranes were then incubated with primary antibodies against
B-cell lymphoma 2 (Bcl-2; 1:500; Wan Biotechnology; cat. no.
WL01556), Bcl-2-associated X protein (Bax; 1:500; Wan
Biotechnology; cat. no. WL01637), apoptotic protease activating
factor-1 (APAF-1; 1:1,000; Proteintech; cat. no. 21093-1-AP),
cytochrome c (1:1,000; Proteintech; cat. no. 66264-1-Ig),
cleaved caspase-9 (1:500; Wan Biotechnology; cat. no. 66264-1-Ig),
cleaved caspase-3 (1:500; Wan Biotechnology; cat. no. WL01992),
β-actin (1:500; Wan Biotechnology; cat. no. WL01372) overnight at
4°C. Following incubation with the primary antibodies, the
membranes were incubated with horseradish peroxidase-labeled
anti-mouse and anti-rabbit secondary antibodies (both 1:4,000;
EarthOx Life Sciences; cat. no. E030110-02 and E030120-02) for 0.5
h at room temperature. Protein bands were visualized using enhanced
chemiluminescence substrates. Protein expression was quantified
using ImageJ software (version 1.51).
Assessment of mRNA expression
Total RNA was extracted from myocardial tissue and
H9C2 cells using TRIzol® reagent (Solarbio Science &
Technology Co.), according to the manufacturer’s protocol. The
concentration and the purity of the RNA samples were analyzed using
the Nanodrop 2000 System (Thermo Fisher Scientific, Inc.). Total
RNA was reverse transcribed into cDNA using the PrimeScript RT
reagent kit (Takara Bio), according to the manufacturer’s protocol.
The following temperature protocol was used: 37°C for 15 min, 85°C
for 5 sec and cooling down to 4°C. qPCR was subsequently performed
using the SYBR Premix Ex Taq II (Takara Bio), according to the
manufacturer’s instructions. The primer pairs used for qPCR were
designed by Biotechnology Co., Ltd. and are presented in Table I. The following thermocycling
conditions were used: i) Step 1, 5 min at 95°C; ii) step 2, 8
cycles at 95°C for 30 sec, at 60°C for 45 sec, at 72°C for 20 sec;
iii) step 3, 35 cycles at 95°C for 30 sec, at 56°C for 45 sec, at
72°C for 20 sec; and iv) step 4, 1 min at 95°C, 30 sec at 55°C, 30
sec at 95°C. GAPDH was used as the internal reference gene. Target
gene mRNA levels were quantified using the 2−ΔΔCq method
(26).
 | Table IPrimer sequences used for
quantitative PCR. |
Table I
Primer sequences used for
quantitative PCR.
| Primers | Primer sequences
(5′-3′) |
|---|
| Cytochrome
c | Forward:
AGGGTGTCGCCTCAAACCTA |
| Cytochrome
c | Reverse:
ACTGAAGCACGGGTGAGTCT |
| APAF1 | Forward:
CAAGGACACAGACGGTGGAA |
| APAF1 | Reverse:
TGAATCGCACTGACCAGCTT |
| Caspase-9 | Forward:
CAGGTGGAGGTCAGGTGTGA |
| Caspase-9 | Reverse:
TCCGTGAGAGAGGATGACCA |
| Caspase-3 | Forward:
CCATCCTTCAGTGGTGGACA |
| Caspase-3 | Reverse:
TTGAGGCTGCTGCATAATCG |
| Bax | Forward:
GGCGATGAACTGGACAACAA |
| Bax | Reverse:
CAGTTGAAGTTGCCGTCTGC |
| Bcl-2 | Forward:
CACGGTGGTGGAGGAACTCT |
| Bcl-2 | Reverse:
TCCACAGAGCGATGTTGTCC |
Statistical analysis
To ensure the consistency and repeatability of the
results, individual experiments were performed in triplicate. The
experimental results are presented as the means ± standard
deviation. One-way analysis of variance followed by Tukey’s
multiple comparison test was used to compare the groups. P<0.05
was considered to indicate a statistically significant difference.
All data were analyzed using SPSS software (version 20.0; IBM
Corp.).
Results
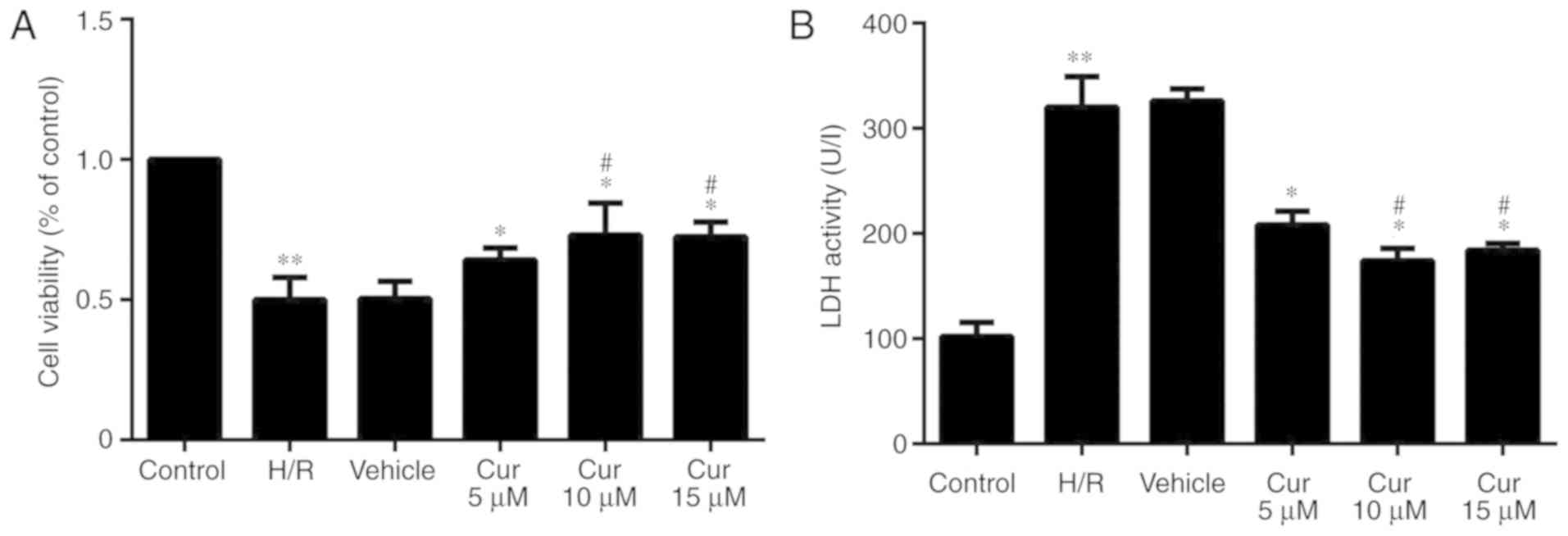
Curculigoside inhibits H/R injury in H9c2
cells in vitro
Following H/R injury, the proliferation of the H9c2
cells was significantly decreased. Curculigoside pre-treatment
significantly increased cell proliferation following H/R injury.
The protective effects of 10 and 15 μM curculigoside on cell
viability were significantly more prominent compared with those of
5 μM curculigoside. No significant differences were observed
between the 10 and 15 μM curculigoside groups (Fig. 1A). The results of LDH assay
revealed that LDH activity in the H/R group was significantly
increased compared with that in the control group. The activity of
LDH was significantly decreased following curculigoside
pre-treatment. LDH activity in the 10 and 15 μM
curculigoside-treated groups was significantly lower compared with
that in the 5 μM curculigoside-treated group. However, there was no
significant difference in LDH activity between the 10 and 15 μM
curculigoside-treated groups (Fig.
1B). Therefore, the concentration of 10 μM curculigoside was
selected for use in subsequent experiments.
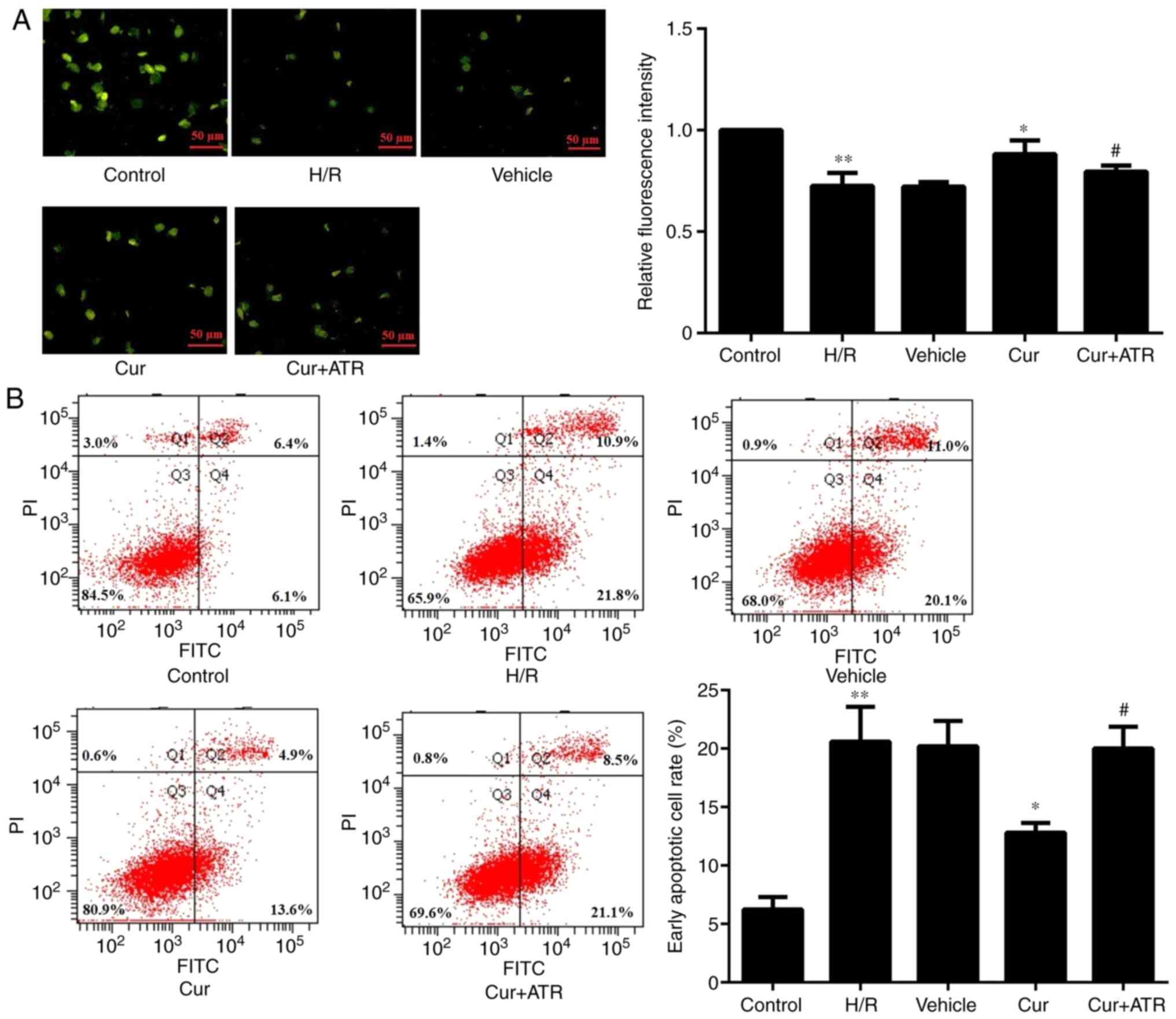
Curculigoside decreases H9c2 cell
apoptosis and inhibits the decrease in ΔΨm
Following rhodamine 123 staining, the florescence
intensity was significantly increased in the H9c2 cells in the
curculigoside-treated group compared with the H/R group,
demonstrating that curculigoside pre-treatment effectively
prevented the decrease in ΔΨm and maintained a high ΔΨm. On the
other hand, the H/R group exhibited a loss in ΔΨm (Fig. 2A). Flow cytometric analysis
revealed that following H/R injury, apoptosis was significantly
increased in the untreated H9c2 cells compared with the control
group. Furthermore, pre-treatment with curculigoside significantly
inhibited H9c2 cell apoptosis following H/R compared with the H/R
group (Fig. 2B).
Curculigoside inhibits
mitochondrial-mediated apoptosis
RT-qPCR analysis revealed that H/R injury
significantly upregulated the mRNA expression of cytochrome
c, APAF-1, caspase-9 and caspase-3 in the H9c2 cells
compared with the control group in vitro. Curculigoside
pre-treatment significantly attenuated the increased mRNA
expression of cytochrome c, APAF-1, caspase-9 and caspase-3
in vitro (Fig. 3A).
Consistent with the changes observed in mRNA expression, western
blot analysis revealed that the protein expression of the
aforementioned proteins in the H/R group was significantly
increased compared with the control group and treatment with
curculigoside prior to H/R injury significantly attenuated these
increased expression levels (Fig.
3B).
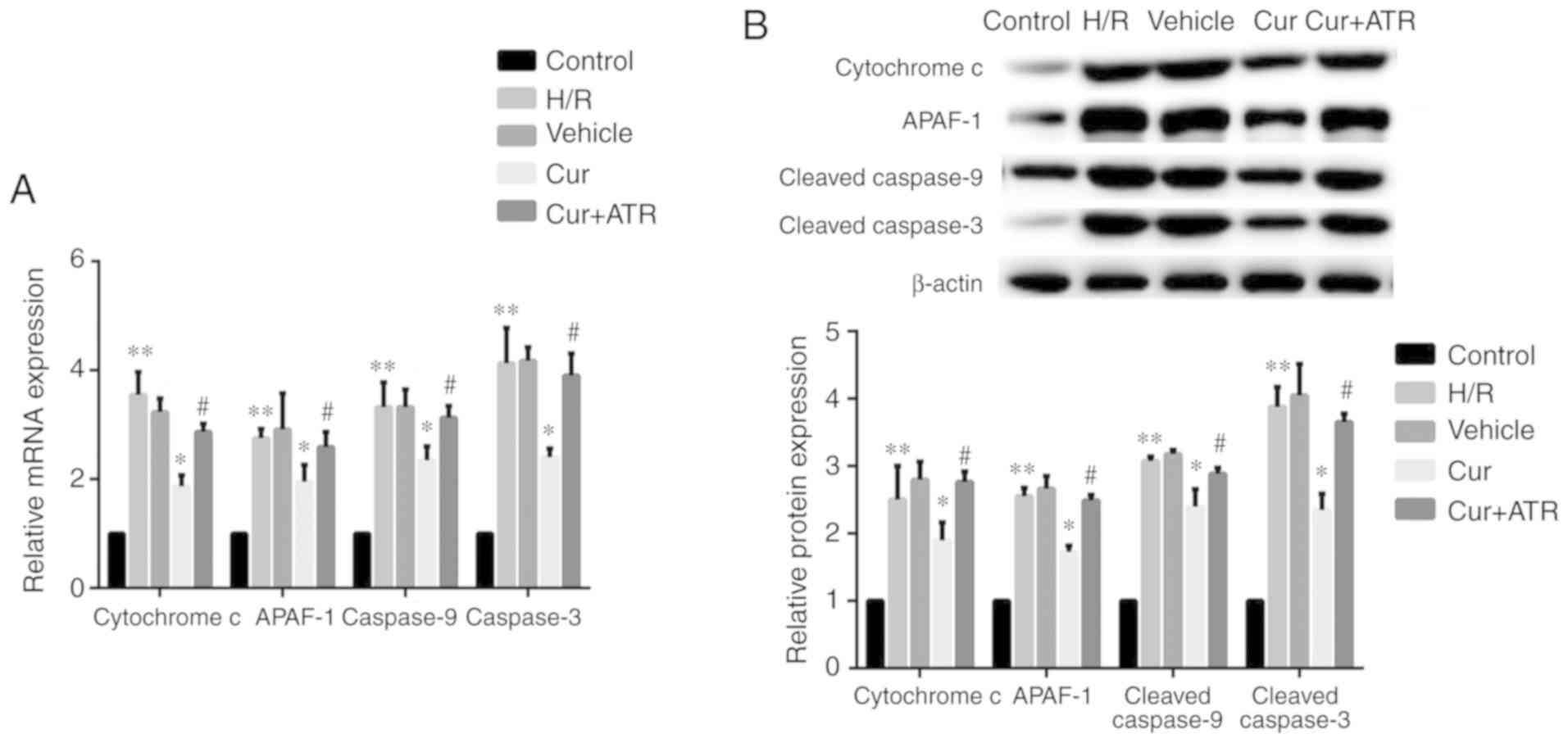 | Figure 3Curculigoside reduces the expression
of cytochrome c, APAF-1, caspase-9 and caspase-3 at the gene
and protein levels in vitro. (A) The mRNA expression of
cytochrome c, APAF-1, caspase-9 and caspase-3 was determined
by RT-qPCR. (B) The protein expression of cytochrome c,
APAF-1, cleaved caspase-9 and cleaved caspase-3 was evaluated by
western blot analysis. **P<0.05 vs. the control
group, *P<0.05 vs. the H/R group,
#P<0.05 vs. the curculigoside-treated group. APAF-1,
apoptotic protease activating factor-1; H/R,
hypoxia/re-oxygenation; Cur, curculigoside; ATR, atractyloside. |
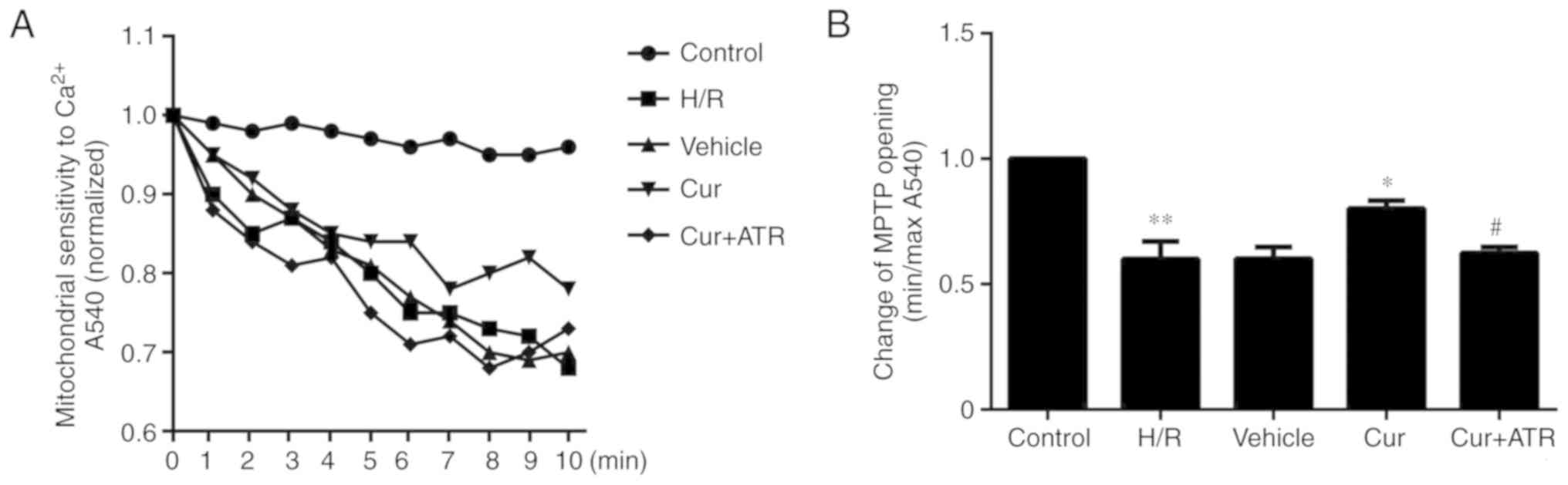
Curculigoside inhibits the opening of the
MPTP
The results revealed that the sensitivity of MPTP
opening to Ca2+ was significantly increased in the H/R
group as compared with the control group, meanwhile, pre-treatment
with curculigoside significantly decreased the sensitivity of MPTP
opening to Ca2+ compared with the H/R group, indicating
that curculigoside inhibited MPTP opening following H/R injury
(Fig. 4).
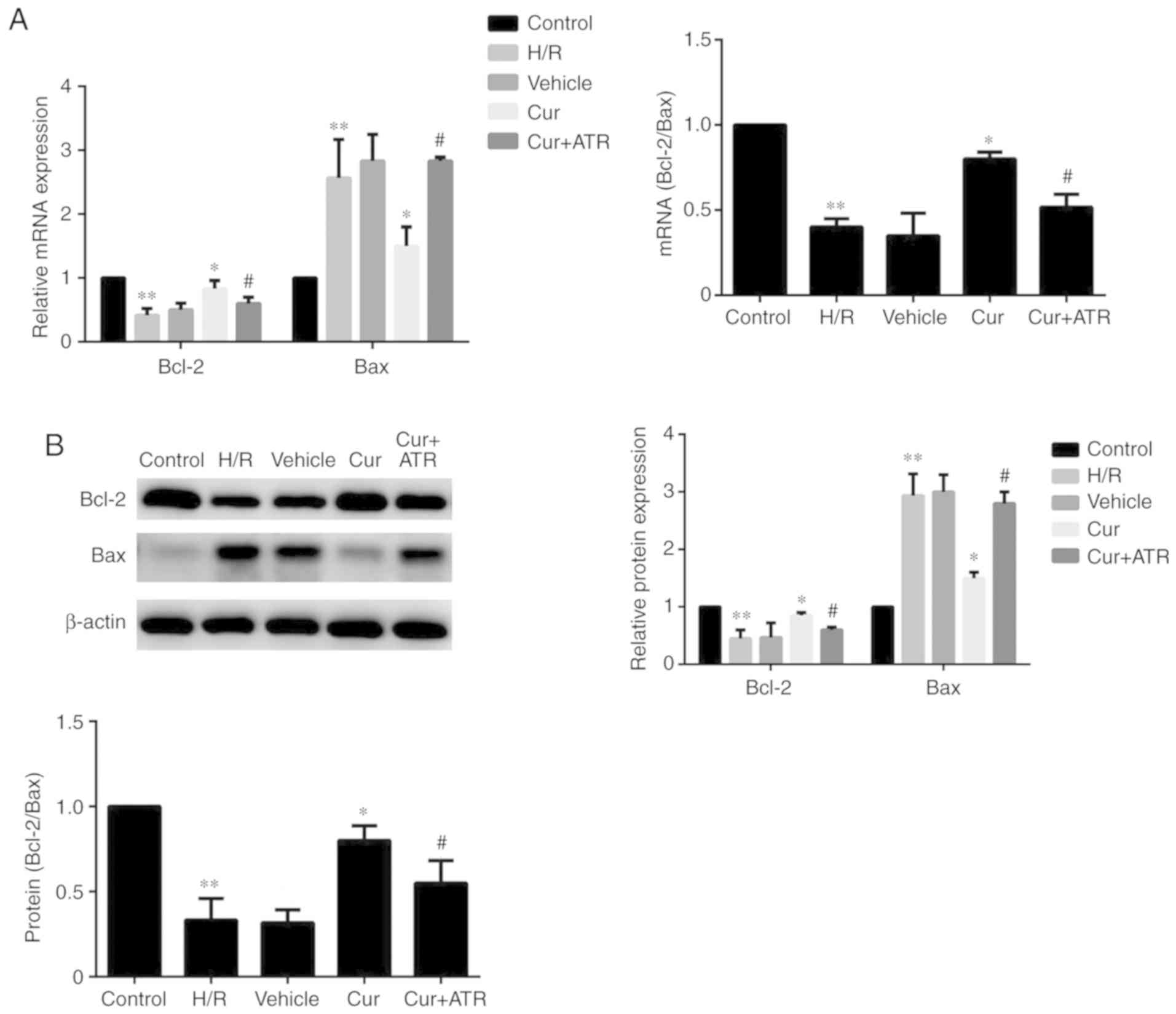
Curculigoside regulates the expression of
Bax and Bcl-2 at the gene and protein levels
RT-qPCR analysis indicated that H/R injury
significantly upregulated the mRNA expression of Bax, downregulated
Bcl-2 expression and decreased the ratio of Bcl-2/Bax compared with
the control group. Curculigoside treatment significantly attenuated
the changes in mRNA expression and increased the ratio of Bcl-2/Bax
(Fig. 5A). Similarly, western
blot analysis revealed that the protein expression levels of Bax
and Bcl-2 were significantly increased and decreased, respectively,
in the H/R group compared with the control group. Pre-treatment
with curculigoside significantly attenuated the increased protein
expression of Bax, the decreased protein expression of Bcl-2 and
the decreased ratio of Bcl-2/Bax (Fig. 5B).
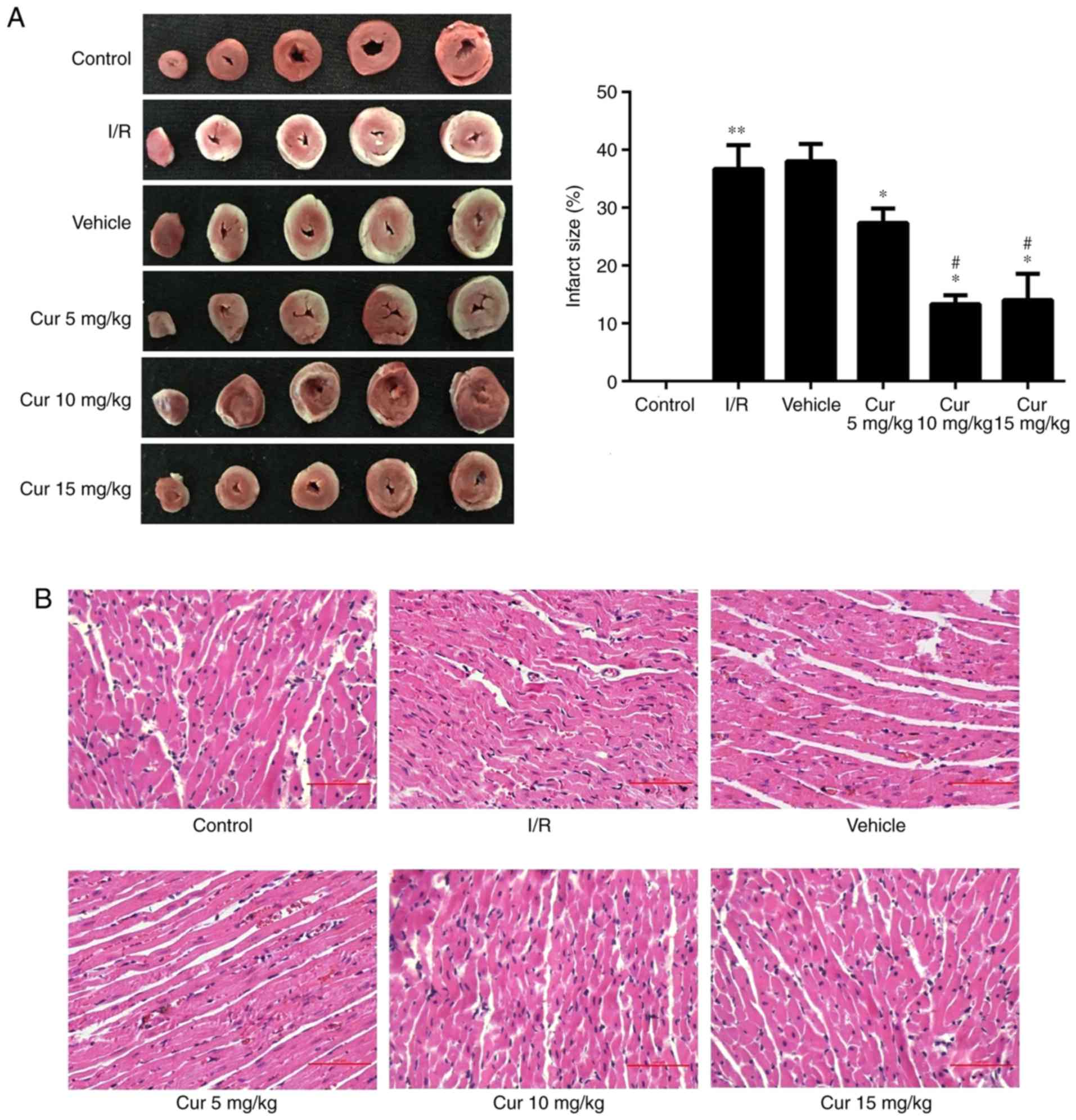
Curculigoside attenuates I/R-induced
myocardial injury ex vivo
In order to examine the potential cardioprotective
effects of curculigoside, rat hearts were isolated and subjected to
I/R with or without pre-treatment with 5, 10 and 15 mg/kg
curculigoside. Curculigoside pre-treatment significantly decreased
the infarct size compared with the I/R group. Furthermore, the
infarct size was decreased in the 10 and 15 mg/kg
curculigoside-treated groups compared with the 5 mg/kg
curculigoside-treated group. However, there was no statistically
significant difference between the infarct sizes in the 10 and 15
mg/kg curculigoside-treated groups (Fig. 6A). H&E staining revealed that
the myocardial fibers in the control group were of a normal size
and highly aligned. In the I/R group, the myocardial fibers were
loosely and irregularly arranged and inflammatory cell infiltration
and myocardial cell edema were observed. However, these
pathological changes were less pronounced in the
curculigoside-treated groups, particularly in the 10 mg/kg
curculigoside-treated group (Fig.
6B). Accordingly, 10 mg/kg curculigoside was selected as the
optimal dose and was used for subsequent experimentation.
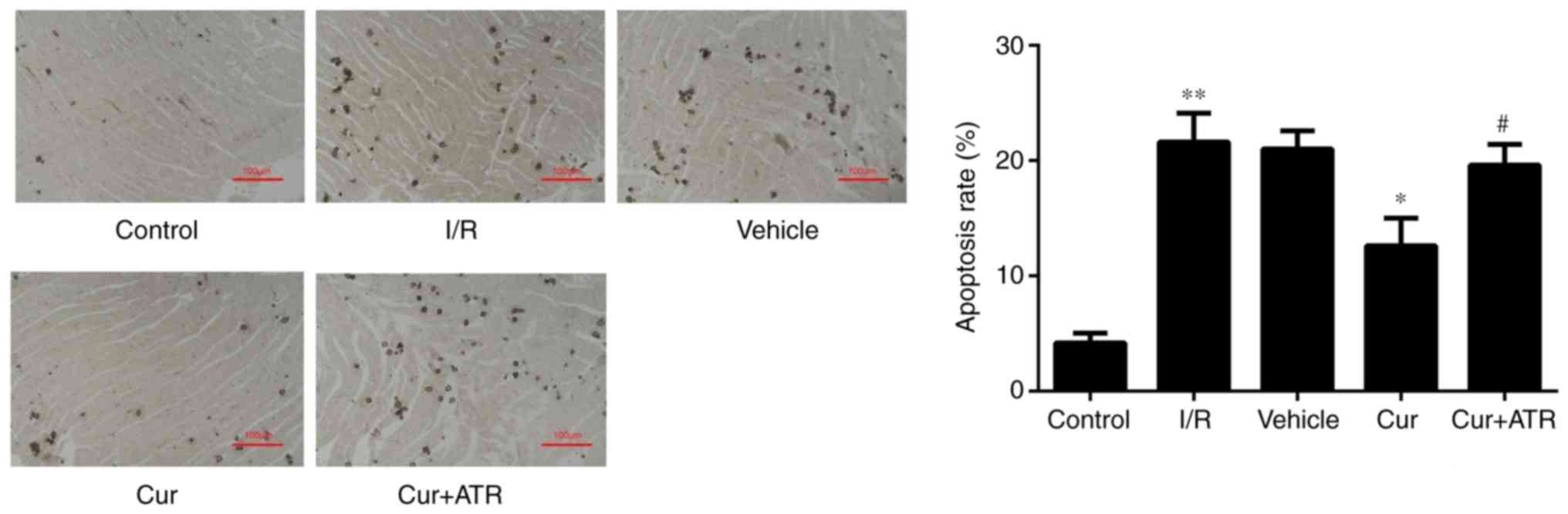
Curculigoside inhibits myocardial cell
apoptosis and decreases the expression of cytochrome c, APAF-1,
caspase-9 and caspase-3 at the gene and protein levels ex vivo
TUNEL staining revealed that myocardial apoptosis
following I/R injury was significantly decreased by pre-treatment
with curculigoside (Fig. 7).
RT-qPCR analysis revealed that I/R injury significantly upregulated
the mRNA expression of cytochrome c, APAF-1, caspase-9 and
caspase-3 compared with the control group ex vivo.
Furthermore, curculigoside pre-treatment significantly attenuated
the increased mRNA expression of the aforementioned proteins ex
vivo (Fig. 8A). Additionally,
western blot analysis revealed that the expression of these
proteins in the I/R group was significantly upregulated compared
with the control group. Curculigoside treatment prior to I/R injury
significantly decreased the upregulated protein expression of
cytochrome c, APAF-1, cleaved caspase-9 and cleaved
caspase-3 (Fig. 8B).
 | Figure 8Curculigoside downregulates the
protein expression of cytochrome c, APAF-1, caspase-9 and
caspase-3 at the gene and protein levels ex vivo. (A) The
mRNA expression of cytochrome c, APAF-1, caspase-9 and
caspase-3 was evaluated by reverse-transcription quantitative PCR.
(B) The protein expression of cytochrome c, APAF-1, cleaved
caspase-9 and cleaved caspase-3 was examined by western blot
analysis. n=6. **P<0.05 vs. the control group,
*P<0.05 vs. the H/R group, #P<0.05 vs.
the curculigoside-treated group. APAF-1, apoptotic protease
activating factor-1; I/R, ischemia/reperfusion; Cur, curculigoside;
ATR, atractyloside. |
Atractyloside abrogates the
cardioprotective effects of curculigoside following I/R or H/R
injury
In order to investigate whether curculigoside exerts
cardioprotective effects following I/R or H/R injury by inhibiting
the opening of the MPTP, the effects of atractyloside, a known MPTP
opener, were examined. Apoptosis was significantly increased in the
myocardium ex vivo and in H9c2 cells in vitro in the
combination treatment group compared with the curculigoside-treated
group (Figs. 2B and 7). Furthermore, the effects of the
combination treatment on mitochondrial-mediated apoptosis were
investigated. The cardioprotective effects of curculigoside
following I/R or H/R injury were attenuated by atractyloside, and
this resulted in decreased ΔΨm, the upregulated expression of
cytochrome c, APAF-1, cleaved caspase-9 and cleaved
caspase-3, increased MPTP opening, the decreased expression of
Bcl-2 and the increased expression of Bax (Figs. 2A, 3A and B, 4, 5A and
B, and 8A and B).
Discussion
MIRI occurs in a number of heart diseases, and while
several therapeutic strategies have been proposed, there is
currently no standard treatment protocol. Curculigo
orchioides Gaertn is a traditional herb listed in Pharmacopeia
of China (2015 edition) (27),
which has been used to enhance kidney yang, strengthen bones and
muscles, and alleviate coldness and wetness in the body in China
for many centuries (28). A
recent study also reported that the extracts from the rhizomes of
the plant Curculigo orchioides Gaertnits exhibited potent
antioxidant activities (29).
Curculigoside is a major active constituent of Curculigo
orchioides Gaertn and a number of previously published studies
have revealed that curculigoside inhibits oxidative damage and I/R
injury in mouse cortical neurons, rat calvarial osteoblasts and
human umbilical vein endothelial cells (18,30,31). Thus, it was hypothesized that
curculigoside may exert protective effects against MIRI. As
expected, the results of the present study demonstrated that
curculigoside attenuated MIRI in a dose-dependent manner in
vitro and ex vivo. To the best of the authors’
knowledge, the present study was the first to reveal the
cardioprotective effects of curculigoside on MIRI. These findings
suggest that curculigoside may have a potential use for the
prevention of MIRI.
Curculigoside has been demonstrated to exhibit a
protective effect in nerve tissue following oxidative stress by
inhibiting mitochondria-mediated apoptosis (32). Furthermore, mitochondrial-mediated
apoptosis has previously been implicated in MIRI (33,34). Mounting evidence has demonstrated
that MPTP plays an important role in modulating the mitochondrial
apoptotic pathway, as MPTP opening releases cytochrome c and
APAF-1, activates caspase-9 and caspase-3, and ultimately induces
apoptosis (35,36). A previous study revealed that MPTP
opening played an important role during the course of cardiac I/R
injury (37). Therefore, the
present study investigated whether curculigoside exerted
cardioprotective effects by inhibiting MPTP opening.
In the present study, the sensitivity of the MPTP to
Ca2+ increased and ΔΨm decreased in H9c2 cells following
H/R injury. This resulted in MPTP opening, the increased expression
of apoptosis-associated proteins and cardiomyocyte apoptosis
(38). A decrease in ΔΨm and an
increased sensitivity of MPTP to Ca2+ are key features
of mitochondrial-mediated apoptosis (39–41). In the present study, following
curculigoside pre-treatment, the sensitivity of MPTP to
Ca2+ and the loss in ΔΨm were decreased, suggesting the
inhibition of MPTP opening, and the expression of
apoptosis-associated proteins and cardiomyocyte apoptosis were
decreased. Atractyloside, a known MPTP opener, abrogated the
inhibitory effects of curculigoside on the decreased expression of
apoptosis-associated proteins and myocardial apoptosis. The results
of the present study indicated that curculigoside targeted the MPTP
and protected the rat myocardium and H9c2 cells against I/R or H/R
injury by inhibiting MPTP opening.
Previous studies have revealed that Bax and Bcl-2
are key upstream proteins in the regulation of MPTP opening
(42,43). Bax binds to voltage dependent
anion channels (VDAC) and leads to MPTP opening (44). However, Bcl-2 inhibits the binding
of Bax to VDAC and p53 to peptidylprolyl isomerase D, and decreases
MPTP opening (44,45). Therefore, the ratio of Bax/Bcl-2
regulates MPTP opening (46). The
present study revealed that curculigoside increased the expression
of Bcl-2, and decreased the expression of Bax, therefore increasing
the Bcl-2/Bax ratio. This suggested that curculigoside inhibited
MPTP opening by decreasing the Bax/Bcl-2 ratio.
The present study had two important limitations.
Firstly, the isolated rat heart model lacks humoral and neural
regulation and does not accurately represent the physiological
progression of MIRI in situ. Future studies are required to
investigate the cardioprotective effects of curculigoside in
situ following I/R injury. Secondly, the present study
demonstrated that curculigoside inhibited mitochondria-mediated
apoptosis by acting on the MPTP. However, other apoptotic signaling
pathways, such as death receptor-mediated apoptosis, were not
investigated. Therefore, further investigation of the mechanism of
action of curculigoside is required.
In conclusion, the present study revealed a
previously unknown, at least to the best of our knowledge,
cardioprotective effect of curculigoside. The results obtained in
the present study may serve as the basis for the development of
novel therapeutic agents for MIRI.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81670320 and 81800232) and
the Natural Science Foundation of Liaoning Province (grant no.
201602826).
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors’ contributions
DJ and NW conceived and designed the experiments. YZ
conducted the experiments. YG, YC, and SL participated in the
completion of the experiments. YZ and NW analyzed the data. YZ and
NW wrote the manuscript. All authors read and approved the final
version of this manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of China Medical University and procedures for animals
handling and care adhered to the Guide for the Care and Use of
Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Writing Group Members. Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Executive summary: Heart disease and
stroke statistics-2016 update: A report from the American heart
association. Circulation. 133:447–454. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Turer AT and Hill JA: Pathogenesis of
myocardial ischemia-reperfusion injury and rationale for therapy.
Am J Cardiol. 106:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kalogeris T, Baines CP, Krenz M and
Korthuis RJ: Cell biology of ischemia/reperfusion injury. Int Rev
Cell Mol Biol. 298:229–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fraccarollo D, Galuppo P and Bauersachs J:
Novel therapeutic approaches to post-infarction remodelling.
Cardiovasc Res. 94:293–303. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morciano G, Bonora M, Campo G, Aquila G,
Rizzo P, Giorgi C, Wieckowski MR and Pinton P: Mechanistic role of
mPTP in ischemia-reperfusion injury. Adv Exp Med Biol. 982:169–189.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sanada S, Komuro I and Kitakaze M:
Pathophysiology of myocardial reperfusion injury: Preconditioning,
postconditioning, and translational aspects of protective measures.
Am J Physiol Heart Circ Physiol. 301:H1723–H1741. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Verma S, Fedak PW, Weisel RD, Butany J,
Rao V, Maitland A, Li RK, Dhillon B and Yau TM: Fundamentals of
reperfusion injury for the clinical cardiologist. Circulation.
105:2332–2336. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nazari A, Sadr SS, Faghihi M, Azizi Y,
Hosseini MJ, Mobarra N, Tavakoli A and Imani A: Vasopressin
attenuates ischemia-reperfusion injury via reduction of oxidative
stress and inhibition of mitochondrial permeability transition pore
opening in rat hearts. Eur J Pharmacol. 760:96–102. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murphy E and Steenbergen C: Mechanisms
underlying acute protection from cardiac ischemia-reperfusion
injury. Physiol Rev. 88:581–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bernardi P and Di Lisa F: The
mitochondrial permeability transition pore: Molecular nature and
role as a target in cardioprotection. J Mol Cell Cardiol.
78:100–106. 2015. View Article : Google Scholar :
|
|
11
|
Shires SE and Gustafsson ÅB: Mitophagy and
heart failure. J Mol Med (Berl). 93:253–262. 2015. View Article : Google Scholar
|
|
12
|
Piot C, Croisille P, Staat P, Thibault H,
Rioufol G, Mewton N, Elbelghiti R, Cung TT, Bonnefoy E, Angoulvant
D, et al: Effect of cyclosporine on reperfusion injury in acute
myocardial infarction. N Engl J Med. 359:473–481. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morin D, Pires F, Plin C and Tillement JP:
Role of the permeability transition pore in cytochrome C release
from mitochondria during ischemia-reperfusion in rat liver. Biochem
Pharmacol. 68:2065–2073. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Winkler HH, Bygrave FL and Lehninger AL:
Characterization of the atractyloside-sensitive adenine nucleotide
transport system in rat liver mitochondria. J Biol Chem. 243:20–28.
1968.PubMed/NCBI
|
|
15
|
Murali VP and Kuttan G: Curculigoside
augments cell-mediated immune responses in metastatic tumor-bearing
animals. Immunopharmacol Immunotoxicol. 38:264–269. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kubo M, Namba K, Nagamoto N, Nagao T,
Nakanishi J, Uno H and Nishimura H: A new phenolic glucoside,
curculigoside from rhizomes of Curculigo orchioides. Planta Med.
47:52–55. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu FB, Wang JY, Zhang YL, Quan RF, Yue
ZS, Zeng LR, Zheng WJ, Hou Q, Yan SG and Hu YG: Curculigoside
regulates proliferation, differentiation, and pro-inflammatory
cytokines levels in dexamethasone-induced rat calvarial
osteoblasts. Int J Clin Exp Med. 8:12337–12346. 2015.PubMed/NCBI
|
|
18
|
Wang YK, Hong YJ, Wei M, Wu Y, Huang ZQ,
Chen RZ and Chen HZ: Curculigoside attenuates human umbilical vein
endothelial cell injury induced by H2O2. J Ethnopharmacol.
132:233–239. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu H, He J, Ye L, Lin F, Hou J, Zhong Y
and Jiang W: Mechanisms of angiogenesis in a curculigoside
A-treated rat model of cerebral ischemia and reperfusion injury.
Toxicol Appl Pharmacol. 288:313–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang W, Fu F, Tian J, Zhu H and Hou J:
Curculigoside A attenuates experimental cerebral ischemia injury in
vitro and vivo. Neuroscience. 192:572–579. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang Z, Zhu H, Luan H, Han F and Jiang W:
Curculigoside A induces angiogenesis through VCAM-1/Egr-3/CREB/VEGF
signaling pathway. Neuroscience. 267:232–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu H, Ding Y, Xu X, Li M, Fang Y, Gao B,
Mao H, Tong G, Zhou L and Huang J: Prostaglandin E1 protects
coronary microvascular function via the glycogen synthase kinase
3β-mitochondrial permeability transition pore pathway in rat hearts
subjected to sodium laurate-induced coronary microembolization. Am
J Transl Res. 9:2520–2534. 2017.
|
|
23
|
Kastenmayer RJ, Moore RM, Bright AL,
Torres-Cruz R and Elkins WR: Select agent and toxin regulations:
Beyond the eighth edition of the guide for the care and use of
laboratory animals. J Am Assoc Lab Anim Sci. 51:333–338.
2012.PubMed/NCBI
|
|
24
|
Wu N, Li W, Shu W and Jia D: Protective
effect of picroside II on myocardial ischemia reperfusion injury in
rats. Drug Des Devel Ther. 8:545–554. 2014.PubMed/NCBI
|
|
25
|
Herr DJ, Aune SE and Menick DR: Induction
and assessment of ischemia-reperfusion injury in
langendorff-perfused rat hearts. J Vis Exp. 27:e529082015.
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Chinese Pharmacopoeia Commission.
Pharmacopoeia of the People Republic of China. Chin Med Sci Technol
Press; Beijing: pp. 189–191. 2015
|
|
28
|
Miao M, Tian S, Guo L, Bai M, Fang X and
Liu S: The effect of curculigoside on mouse model of perimenopausal
depression. Saudi J Biol Sci. 24:1894–1902. 2017. View Article : Google Scholar
|
|
29
|
Hejazi II, Khanam R, Mehdi SH, Bhat AR,
Rizvi MMA, Thakur SC and Athar F: Antioxidative and
anti-proliferative potential of Curculigo orchioides Gaertn in
oxidative stress induced cytotoxicity: In vitro, ex vivo and in
silico studies. Food Chem Toxicol. 115:244–259. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tian Z, Yu W, Liu HB, Zhang N, Li XB, Zhao
MG and Liu SB: Neuroprotective effects of curculigoside against
NMDA-induced neuronal excitoxicity in vitro. Food Chem Toxicol.
50:4010–4015. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Zhao L, Wang Y, Xu J, Nie Y, Guo
Y, Tong Y, Qin L and Zhang Q: Curculigoside isolated from Curculigo
orchioides prevents hydrogen peroxide-induced dysfunction and
oxidative damage in calvarial osteoblasts. Acta Biochim Biophys Sin
(Shanghai). 44:431–441. 2012. View Article : Google Scholar
|
|
32
|
Changa HT, Jan CR and Liang WZ: Protective
effects of a phenolic glycoside compound curculigoside on
h2o2-induced oxidative stress and
cytotoxicity in normal human breast epithelial cells. J Funct Food.
41:171–182. 2018. View Article : Google Scholar
|
|
33
|
Yang HX, Wang P, Wang NN, Li SD and Yang
MH: Tongxinluo ameliorates myocardial ischemia-reperfusion injury
mainly via activating parkin-mediated mitophagy and downregulating
ubiquitin-proteasome system. Chin J Integr Med. Jun 21–2019.(Epub
ahead of print). View Article : Google Scholar
|
|
34
|
Li Y, Xiang Y, Zhang S, Wang Y, Yang J,
Liu W and Xue F: Intramyocardial injection of thioredoxin
2-expressing lentivirus alleviates myocardial ischemia-reperfusion
injury in rats. Am J Transl Res. 9:4428–4439. 2017.PubMed/NCBI
|
|
35
|
Karch J and Molkentin JD: Identifying the
components of the elusive mitochondrial permeability transition
pore. Proc Natl Acad Sci USA. 111:10396–10397. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baines CP, Song CX, Zheng YT, Wang GW,
Zhang J, Wang OL, Guo Y, Bolli R, Cardwell EM and Ping P: Protein
kinase cepsilon interacts with and inhibits the permeability
transition pore in cardiac mitochondria. Circ Res. 92:873–880.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu Z, Alloush J, Beck E and Weisleder N: A
murine model of myocardial ischemia-reperfusion injury through
ligation of the left anterior descending artery. J Vis Exp.
86:513292014.
|
|
38
|
Kim JS, Wang JH and Lemasters JJ:
Mitochondrial permeability transition in rat hepatocytes after
anoxia/reoxygenation: Role of Ca2+-dependent
mitochondrial formation of reactive oxygen species. Am J Physiol
Gastrointest Liver Physiol. 302:G723–G731. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Scarffe LA, Stevens DA, Dawson VL and
Dawson TM: Parkin and pink1: Much more than mitophagy. Trends
Neurosci. 37:315–324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Skulachev VP: Bioenergetic aspects of
apoptosis, necrosis and mitoptosis. Apoptosis. 11:473–485. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim R, Emi M, Tanabe K, Murakami S, Uchida
Y and Arihiro K: Regulation and interplay of apoptotic and
non-apoptotic cell death. J Pathol. 208:319–326. 2006. View Article : Google Scholar
|
|
42
|
Low IC, Kang J and Pervaiz S: Bcl-2: A
prime regulator of mitochondrial redox metabolism in cancer cells.
Antioxid Redox Signal. 15:2975–2987. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Takahashi A, Masuda A, Sun M, Centonze VE
and Herman B: Oxidative stress-induced apoptosis is associated with
alterations in mitochondrial caspase activity and Bcl-2-dependent
alterations in mitochondrial pH (pHm). Brain Res Bull. 62:497–504.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Martel C, Wang Z and Brenner C: Vdac
phosphorylation, a lipid sensor influencing the cell fate.
Mitochondrion. 19:69–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Green DR and Evan GI: A matter of life and
death. Cancer Cell. 1:19–30. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Burguillos MA, Hajji N, Englund E, Persson
A, Cenci AM, Machado A, Cano J, Joseph B and Venero JL:
Apoptosis-inducing factor mediates dopaminergic cell death in
response to LPS-induced inflammatory stimulus: Evidence in
Parkinson’s disease patients. Neurobiol Dis. 41:177–188. 2011.
View Article : Google Scholar
|