Introduction
Hepatic ischemia-reperfusion injury (HIRI) remains a
major cause of damage that is associated with several liver
diseases and hepatic surgery, which affects the outcomes of hepatic
operations and other liver diseases related to HIRI (1,2).
Complex mechanisms are involved in HIRI and studies indicate that
Kupffer cells (KCs), which are liver-resident macrophages, play a
crucial role in the inflammatory response in several hepatic
diseases (3-6). Two well-known KC phenotypes, M1 and
M2, have different functions in HIRI; M1-polarized KCs exacerbate
IR-induced damage to the liver by producing proin-flammatory
cytokines such as TNF-α and interleukin (IL)-1β, while
anti-inflammatory cytokines, such as IL-10 and TGF-β, are produced
by M2-polarized KCs and protect hepatocytes against IR injury
(7-9). Therefore, increasing the number of
M2 cells or reducing the M1 phenotype of KCs may help to ameliorate
HIRI.
Autophagy is a conserved intracellular
self-digestion process that eliminates damaged organelles and
senescent proteins, in which autophagolysosomes created by the
fusion of autophagosomes and lysosomes play a crucial role
(10). Autophagy plays either a
protective or detrimental role under different conditions, making
it a 'double-edged sword' for stressed cells (11). The role of autophagy in HIRI
remains controversial and depends on the treatment and how the
model is established (warm or cold) (12-14). Studies have shown that autophagy
plays a protective role in orthotopic liver transplantation
(OLT)-induced IRI. Nakamura et al (15) demonstrated that
HO-1/Sirt1-mediated autophagy contributes to ameliorating
OLT-induced IRI in mice and humans. Zaouali et al (16) confirmed the hepatoprotective
effect of AMPK-dependent autophagy in OLT-induced IRI. During HIRI,
classically activated KCs (M1) damage the liver tissue not only by
releasing reactive oxygen species and inflammatory cytokines but
also by attracting other inflammatory cells to amplify these
negative effects, and a previous study showed that autophagy plays
a protective role by downregulating the cellular inflammatory
response (17). In the livers of
high-fat diet-fed mice, the loss of autophagy promotes
lipopolysaccharide (LPS)-induced M1 polarization of KCs (18). Similarly, increased levels of
IL-1β and IL-18 in LC3B knockout macrophages were observed in a
sepsis mouse model, which revealed the protective role of autophagy
in macrophage-related inflammation, but whether autophagy protects
the liver from cold ischemia reperfusion remains to be elucidated
(19).
Suberoylanilide hydroxamic acid (SAHA) is a
pan-histone deacetylase inhibitor that has been applied clinically
for the treatment of cancers for numerous years (20) and has also been shown to have
anti-inflammatory effects on colitis and attenuate con A-induced
acute hepatic injury (21,22).
Choi et al (23)
demonstrated that SAHA downregulates proin-flammatory factor levels
in plasma and inhibits responses of peripheral blood mononuclear
cells to Toll-like receptor 4 (TLR4). Moreover, SAHA protects
cardiomyocytes against IRI in an autophagy-dependent manner
(24). Recent evidence suggests
that SAHA affects the formation of autophagosomes and promotes
autophagy (25). However, the
role of SAHA in cold HIRI remains unclear and so a model of cold
HIRI and SAHA pretreatment was established to investigate its
effect on the IR-injured liver.
Studies have demonstrated that SAHA promotes
autophagy of several cell models by downregulating AKT/mTOR
signaling, which is one of the classical pathways involved in
regulating cellular autophagy (26-29). AKT is a well-studied factor that
functions in several models of diseases and positively regulates
the phosphorylation of NF-κB, thus enhancing M1 polarization of
macrophages (30,31). AKT also phosphorylates glycogen
synthase kinase 3β (GSK3β), which is a conserved kinase that
negatively regulates the activity of NF-κB. Cremer et al
(32) demonstrated that GSK3β
regulates the Burkholderia ceno- cepacia-mediated
inflammatory in phagocytes through the PI3K/AKT/GSK3β/NF-κB
pathway. Therefore, whether SAHA influences the AKT/GSK3β/NF-κB
pathway in KCs and whether AKT/mTOR signaling is involved in
SAHA-induced upregulation of KC autophagy was investigated.
Materials and methods
Animals and OLT in rats
The animal experiments involved in this study were
in accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals and approved by the Ethics
Committee of the Second Affiliated Hospital of Chongqing Medical
University (Chongqing, China). Sprague-Dawley (SD) rats (male,
250-300 g, 8-10 weeks old) were obtained from Chongqing Medical
University Experimental Animal Center (Chongqing, China) and were
selected as both donors and recipients. The animals were housed
under specific pathogen-free conditions with an ambient temperature
of 25°C, a controlled humidity of 50% and a 12 h light-dark cycle
and were provided water and standard chow ad libitum. To
establish an OLT model, phenobarbital sodium was used to
anesthetize both of the donor or recipient rats at a dose of 60
mg/kg body weight via intra-peritoneal injection (33), and the modified Kamada's two cuff
technique (34) was used.
KC isolation
A total of 36 SD rats (male, 250-300 g) were
sacrificed for the isolation of KCs, which was performed as
described previously (35). The
rats were anesthetized with phenobarbital sodium at a dose of 60
mg/kg body weight via intra-peritoneal injection and all of the
rats used in the current research were euthanized by exsanguination
from the inferior vena cava under sevoflurane anesthesia (3-4%).
Electrocardiographic monitoring was used to verify death. After
laparotomy, the livers were softened and yellowed by perfusion of
0.05% collagenase type IV through the portal vein, and then the
liver was homogenized and filtered with a 200 mesh stainless steel
screen. Subsequently, the suspension was centrifuged twice at 300 ×
g (4°C) for 5 min to remove the residual enzymatic solution as
described previously (36). The
supernatant was discarded, and the suspension was centrifuged at 50
× g (4°C) for 3 min to remove hepatocytes and other cells. Finally,
the retained supernatant was centrifuged at 300 × g (4°C) for 5 min
and the supernatant was discarded. The isolated KCs were plated and
cultured with RPMI 1640 (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Hyclone, GE Healthcare Life
Sciences) and 100 U/ml penicillin/streptomycin (Sigma-Aldrich;
Merck KGaA) in a humidified atmosphere at 37°C and 5%
CO2. Primary KCs were identified by CD68 staining using
immunofluorescence and flow cytometry.
Establishment of the rat HIRI model with
ATG5 knockdown in KCs
Adeno-associated virus expressing ATG5-short hairpin
(sh)RNA (AAV-ATG5-shRNA) and a control virus (scramble) were
obtained from HanBio Biotechnology Co., Ltd. To obtain the KCs with
ATG5 knockdown, the wild-type rats were injected with
AAV-ATG5-shRNA or scramble (3x1012 vector genomes/kg) 30
days before liver transplantation via the tail vein. The mRNA and
protein levels of ATG5 were measured by RT-qPCR and western
blotting, respectively. To deplete KCs in the donor livers,
clodronate liposomes (CLs) were used as before (37). Then KCs (2x107 cells)
that were isolated from the liver tissue of AAV-ATG5-shRNA- or
scramble-treated rats, were injected into recipients via the portal
vein during OLT (38).
Experimental groups In vivo
The rats were randomly divided into Sham group
(n=5), IR group (n=5), IR+dimethyl sulfoxide treatment group
(IR+DMSO, n=5), IR+SAHA treatment group (IR+SA, n=5), IR+SAHA+CLs
treatment group (IR+SA+CL, n=5), IR+SAHA+chloroquine treatment
group (IR+SA+CQ, n=5), IR+Scramble-shRNA treatment group
(IR+Scramble, n=5), and IR+SAHA+ATG5-shRNA treatment group
(IR+SA+ATG5-shRNA, n=5). For the Sham group, the rats received an
abdominal incision and exposure of the liver vasculature. For the
IR group, the grafts received 24 h of reperfusion after OLT without
additional treatment. For the IR+SA group, the donors and
recipients were both intraperitoneally injected with a dose of 50
mg/kg SAHA (MedChemExpress) 12 h prior to operation, and the
recipients continued to receive 50 mg/kg SAHA q12 h right from
reper-fusion via intra-peritoneal injection (39). For the IR+SA+CL group, CL
(Encapsula NanoSciences) were injected into the donors at a dose of
4 µl/g body weight via the tail vein 48 h prior to surgery
according to the manufacturer's protocol (37). Depletion of KCs were verified by
using flow cytometry and immunohistochemistry, respectively. For
the IR+SA+CQ group, 60 mg/kg CQ (Sigma-Aldrich; Merck KGaA)
dissolved in PBS was injected into both of the donors and
recipients 1 h prior to surgery (40). For the IR+SA+ATG5-shRNA group, the
technique described above in the 'establishment of the rat HIRI
model with ATG5 knockdown in KCs' was used. IR and SA treatment
were performed as described before. For the vehicle-treatment
groups, equal volumes of DMSO, liposomes or Scramble-shRNA were
administered via the same routes as the respective drug treatments.
No effects of these vehicles on liver function were found.
In vitro assays
KCs were randomly divided into Normal group, LPS
(Sigma-Aldrich; Merck KGaA) treatment group, LPS+DMSO treatment
group, LPS+SAHA treatment group (LPS+SA), LPS+Scramble-siRNA
treatment group (LPS+Scramble), and LPS+SAHA+ATG5-siRNA treatment
group (LPS+SA+ATG5-siRNA). In the Normal group, KCs were cultured
with no additional treatment. In the LPS group, KCs were cultured
with LPS (100 ng/ml) for 6 h (41). In the LPS+SA group, LPS-stimulated
KCs were cultured with SAHA (3 µM) for 24 h (41). In the LPS+SAHA+ATG5-siRNA group,
ATG5-siRNA (sense 5′-CAU GUG UGA AGG AAG CUG ATT-3′, antisense
5′-UCA GCU UCC UUC ACA CAU GTT-3′) or scrambled-siRNA (sense UUC
GCU GAA GGU GCC ACG CTT, antisense ACA CGU CAU GCU CGG CGA GTT) was
mixed with Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at a final concentration of 50 nM according to
the manufacturer's protocol. After transfection at 37°C for 48 h,
KCs were treated with LPS and SAHA as described before, and the
silencing efficiency was checked by western blotting. The sequences
of ATG5-siRNA were determined based on a previous reference
(42). For the vehicle-treatment
group, 0.1% DMSO or scramble-siRNA was added to the medium in the
same volume and at the same time point as the respective reagent
treatments.
Hematoxylin and eosin (H&E)
staining
H&E staining was performed according to the
manufacturer's protocol (Beyotime Institute of Biotechnology).
Paraffin sections (5-µm thick) of liver tissue were dewaxed
and hydrated, followed by hematox-ylin staining at 37°C for 5-10
min. Then, the paraffin sections were stained with eosin at 37°C
for 30 sec-2 min. After dehydration, clearing and sealing, tissue
damage and inflammation were observed under a light microscope
(magnification, x400; Olympus Corporation). A total of six fields
were captured for analysis per sample in blinded fashion. The
severity of IRI was graded using Suzuki criteria (43).
Liver function examination
Serum was isolated from whole blood by centrifuging
at 200 × g at 4°C for 10 min. The levels of serum alanine
aminotransferase (sALT) and serum aspartate aminotransferase (sAST)
were evaluated by using an automatic biochemical meter (Beckman
CX7; Beckman Coulter, Inc.).
TUNEL staining
TUNEL staining was performed according to the
manufacturer's protocol (Wuhan Boster Biological Technology, Ltd.).
Paraffin sections fixed with 4% para-formaldehyde for 1 h at room
temperature were dewaxed, hydrated and digested with proteinase K
for 15 min at 37°C. After washing 3 times for 2 min each, the
tissue sections were incubated with a mixture of TdT, DIG-d-UTP and
labeling buffer for 2 h at 37°C. After blocking for 30 min at 37°C,
the sections were incubated with biotinylated anti-digoxigenin
antibody at 37°C for 30 min, followed by incubation with SABC-FITC
for 30 min. The nuclei were stained with DAPI at 37°C for 10 min.
Finally, the fluorescence was detected by fluorescence microscopy
(magnification, x400; Olympus Corporation). Results were scored
semi-quantitatively by counting the number of positive cells in 6
fields per sample in a blinded fashion.
Flow cytometry analysis
To determine the ratio of M1 phenotype KCs,
1x106 KCs were suspended in cell staining buffer before
incubation with anti-CD68 (cat. no. ab201340; Abcam) and anti-CD86
(cat. no. ab213044; Abcam) antibodies at 4°C for 0.5 h. After
washing 3 times with cell staining buffer, the samples were
incubated with a FITC conjugation kit (Fast) (cat. no. ab188285;
Abcam) or PE/Cy7 conjugation kit (cat. no. ab102903; Abcam) at 4°C
in the dark for 30 min. After washing 3 times with cell staining
buffer, the stained cells were acquired by a BD FACSCalibur and
analyzed by using Cell Quest version 5.1 software (BD
Biosciences).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA of liver tissue and cells was isolated by
TRIzol reagent (Takara Bio, Inc.) and reverse transcribed into cDNA
by using the PrimeScriptVR RT reagent kit with genomic DNA eraser
(Takara Bio, Inc.). Reverse transcription of cDNA was conducted in
a 20 µl reaction system: 10 µl mixture of 2 µl
5X gDNA Eraser Buffer, 1 µl gDNA Eraser, 1 µg Total
RNA and RNase Free dH2O was incubated at 42°C for 2 min
and then stored at 4°C for subsequent reactions. Next, 1 µl
PrimeScript RT Enzyme Mix I, 1 µl RT Primer Mix, 4 µl
5X PrimeScript Buffer 2 and 4 µl RNase Free dH2O
were added to the above mixture. Then the reverse transcription of
cDNA was performed at 37°C for 15 min, 85°C for 5 sec, 4°C for 10
min and finally stored at −20°C. For qPCR, cDNA was mixed with
primers and SYBR-Green (Takara Bio, Inc.) following the
manufacturer's protocol and was then detected by a real-time
detection system (Bio-Rad Laboratories, Inc.). The primers used
were as follows: LC3B forward, -TTAAGCCCCTACCAAGGCAA-3′, reverse,
5′-CACTTGCATGGCACTCAGTTT-3′; P62 forward,
5′-TTGAGGCACCCCGTAAAGTC-3′, reverse, 5′-TACATGGTGGCACCTACTGC-3′;
ATG5 forward, 5′-CAGAAGCTGTTCCGTCCTGT-3′, reverse,
5′-CCGTGAATCATCACCTGGCT-3′; and β-actin forward,
5′-CGTTGACATCCGTAAAGAC-3′, reverse 5′-TGGAAGGTGGACAGTGAG-3′ (Sangon
Biotech Co., Ltd.). qPCR was conducted in a 25 µl reaction
system, including 1 µl forward and 1 µl reverse
primers, 12.5 µl SYBR-Green, 2 µl cDNA, and 8.5
µl dH2O. The thermocycling conditions used for
qPCR were as follows: 95°C for 30 sec, 40 cycles of 95°C for 5 sec,
60°C for 30 sec. All the samples were normalized to β-actin
expression. The comparative method 2−ΔΔCq (44) was used for the relative
quantification of results and all experiments were repeated in
triplicate.
Western blot analysis
KCs and liver tissue were treated with RIPA lysis
buffer supplemented with protease inhibitors and
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology). Protein concentrations were detected by a
bicinchoninic acid protein quantitative kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. A total of
30 µg protein was loaded per lane, separated using 10 or 12%
SDS-PAGE and then electrotransferred onto polyvinylidene difluoride
membranes. After blocking in 5% skim milk at 37°C for 1 h, the
membranes were incubated with primary antibodies at 4°C overnight
and then incubated with secondary antibody at 37°C for 1 h. The
bands were visualized by using an enhanced chemiluminescence kit
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. All images were analyzed using ImageJ software, version
14.8 (National Institutes of Health). The primary antibodies used
were as follows: TNF-α (1:1,000; cat. no. ab66579; Abcam), IL-1β
(1:1,000; cat. no. ab9787; Abcam), IL-6 (1:1,000; cat. no. ab9324;
Abcam), Bax [1:1,000; cat. no. 2772; Cell signaling Technology,
Inc. (CST)], Bcl-2 (1:1,000; cat. no. ab196495; Abcam), caspase3
(1:1,000; cat. no. 9662; CST), cleaved-caspase3 (1:1,000; cat. no.
9664; CST), iNOS (1:1,000; cat. no. ab3523; Abcam), phosphory-lated
(p)-Akt (Ser473) (1:1,000; cat. no. 4060; CST), Akt (1:1,000; cat.
no. 9272; CST), p-NF-κB p65 (1:1,000; cat. no. 3033; CST), NF-κB
p65 (1:1,000; cat. no. 6956; CST), p-GSK3β (Ser9) (1:1,000; cat.
no. 9336; CST), GSK3β (1:1,000; cat. no. ab93926; Abcam), p-mTOR
(Ser2448) (1:1,000; cat. no. 5536; CST), mTOR (1:1,000; cat. no.
2972; CST), ATG5 (1:1,000; cat. no. 12994; CST), LC3B (1:1,000;
cat. no. 3868; CST), P62 (1:1,000; cat. no. 23214; CST), and
ATG16L1 (1:1,000; cat. no. 8089; CST).
Immunofluorescence staining
KCs were incubated with primary antibodies against
LC3B (1:200; cat. no. #3868; CST) and CD68 (1:100; cat. no.
ab201340; Abcam) at 4°C overnight and then treated with
FITC-conjugated goat anti-rabbit IgG (1:1,000; cat. no. ab6717;
Abcam) for 1 h at room temperature in the dark. The nuclei were
stained with DAPI at room temperature in the dark for 10 min, which
was then detected by fluorescence microscopy (magnification, ×200;
Olympus Corporation). Six fields were blindly captured for analysis
per sample. ImageJ software, version 14.8 (National Institutes of
Health) was used to quantitate the levels of immunoreactivity.
Transmission electron microscopy
(TEM)
The autophagic vacuoles in KCs were viewed with TEM
(magnification, x20,000; HITACHI HT-7700, Hitachi, Ltd.) at an
accelerating voltage of 100 kV. After being treated with the
appropriate reagents, the KCs were fixed with 2.5% glutaraldehyde
at 4°C for 24 h, and post-fixed with 1% osmium tetroxide at 4°C for
2 h. Then, the samples were dehydrated with gradient ethanol, and
embedded with 100% epoxy resin (cat. no. 45345-1L-F; Honeywell
Fluka; Honeywell Research Chemicals) at 60°C for 24 h. Next,
ultrathin sections at 70-nm thickness were prepared and stained
with uranyl acetate and lead citrate. The preparation of the
samples and the procedures were performed based on a previous
reference (45). Autophagic
vacuoles in 6 randomly selected fields per sample were counted.
There were 5 sections for each specimen.
Statistical analysis
The statistical significance of differences between
two groups was tested by Student's t test, while comparisons
between more than two groups were performed by using one-way ANOVA
followed by a Bonferroni post hoc test and the data are presented
as the mean ± SD. GraphPad Prism 7 (GraphPad Software, Inc.) was
used for data processing. P<0.05 was considered to indicate a
statistically significant difference. Each experiment was repeated
independently at least 3 times.
Results
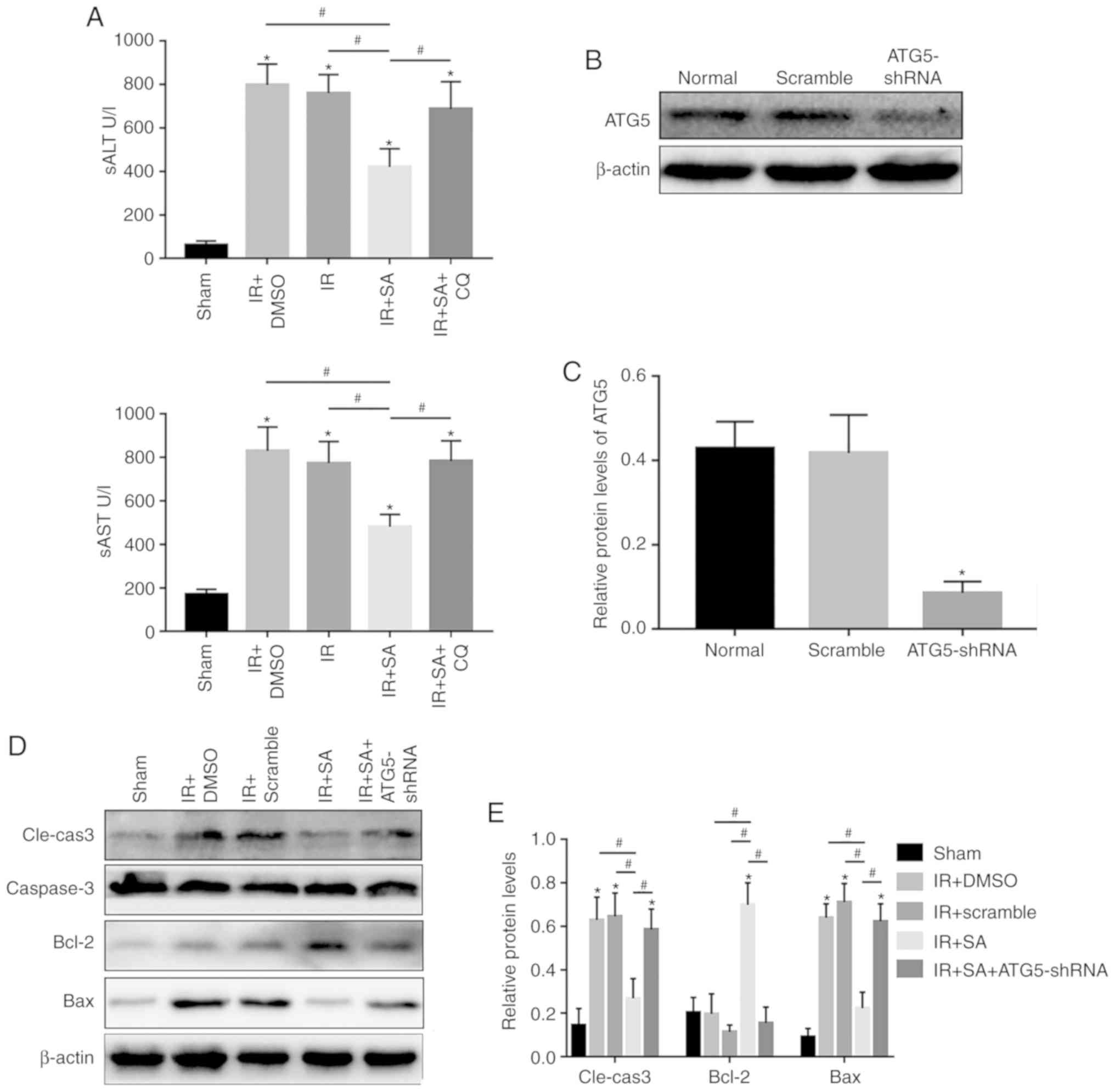
SAHA protects the liver from cold IR in a
KC-dependent manner
In this study, the impact of SAHA on the liver after
cold IR was first investigated. SAHA pretreatment significantly
attenuated damage caused by IRI compared with that of the sham
group at 24 h, as indicated by reduced levels of sALT and sAST
(Fig. 1A), and reduced
hepatocellular damage (Fig. 1B and
C) was found in the SAHA-treated group. Pretreatment with SAHA
protected hepatocytes by reducing hepatocellular apoptosis, as
demonstrated by the protein levels of cleaved-caspase3/caspase3,
Bcl-2 and Bax (Fig. 1D and E).
However, the protective effects of SAHA on IR-injured livers were
diminished after depletion of KCs by CL (Fig. 1). Taken together, SAHA-mediated
protection against cold liver IRI depends on KCs.
 | Figure 1SAHA protects the liver from cold IR
in a KC-dependent manner. (A) The serum concentrations of ALT and
AST at 0, 6, 12, 24 and 48 h after OLT-induced IR in the Sham,
IR+DMSO, IR, IR+SA and IR+SA+CL groups (n=5/group). (B) Images of
hematoxylin and eosin (magnification, ×400) staining showing the
tissue damage in the grafts of different experimental groups at 24
h after IR and the (C) Suzuki score at 24 h after IR. (D) Western
blot analysis of the expression of Cle-caspase3, caspase3, Bcl-2
and Bax. (E) Densitometric analysis of the western blot data.
*P<0.05 vs. the Sham group and #P<0.05
vs. the IR+SA group. OLT, orthotopic liver transplantation; IR,
ischemia reperfusion; DMSO, dimethyl sulfoxide; SA, suberoylanilide
hydroxamic acid; CL, clodronate liposomes; KCs, Kupffer cells;
Cle-caspase3, cleaved caspase3; sALT, serum alanine
aminotransferase; sAST, aspartate transaminase, serum. |
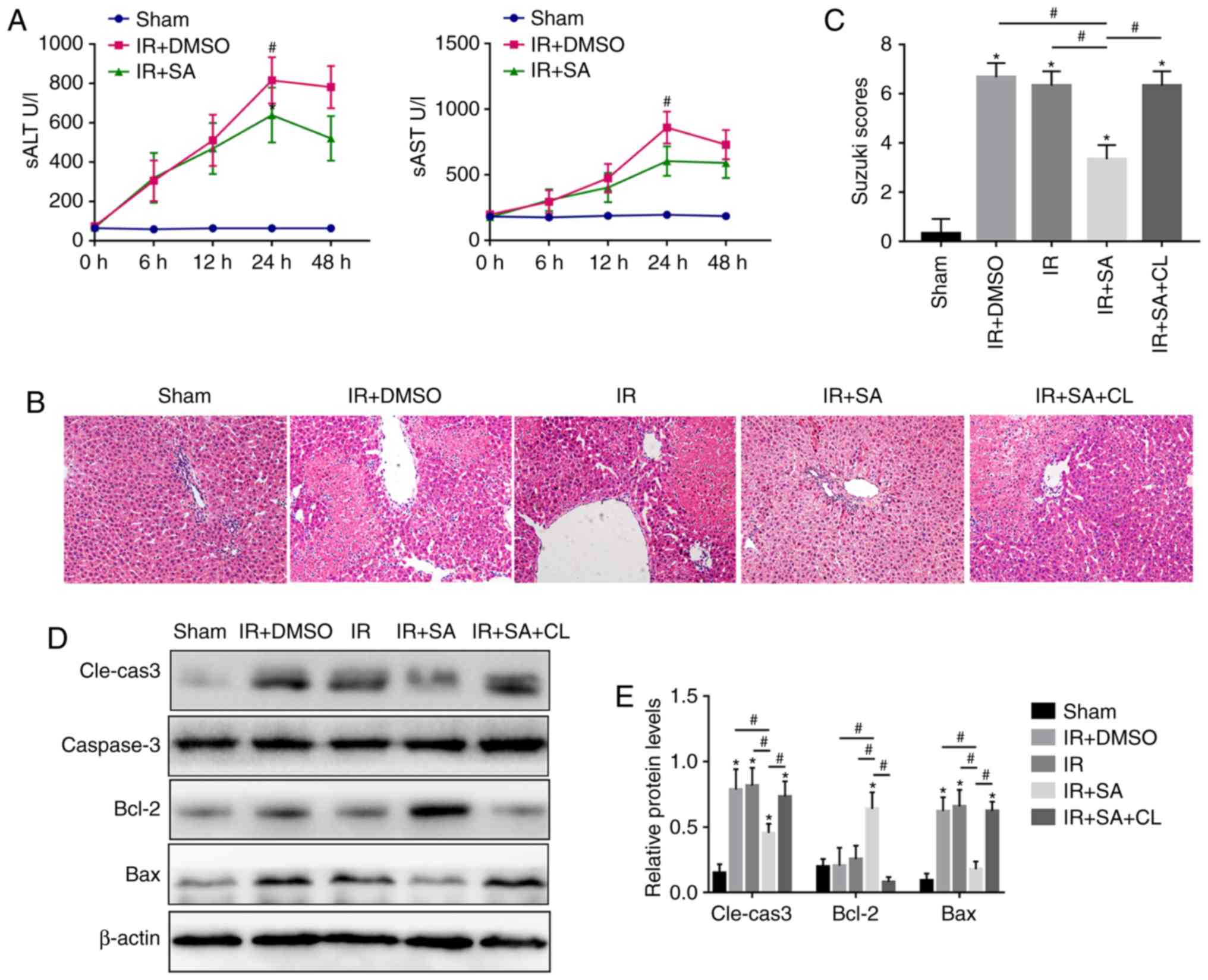
SAHA inhibits inflammation and promotes
autophagy in grafts
To further investigate the effect of SAHA on the
factors related to KC activation, the protein levels of
proin-flammatory factors and AKT/NF-κB p65 were detected, and the
autophagy-related factors AKT/mTOR, ATG5, ATG16L1, LC3B, and P62
were also evaluated after IR insult. As shown in Fig. 2A and B, the proinflammatory
factors IL-1β, TNF-α, and IL-6 were significantly increased after
IR insult but were reversed by SAHA. AKT/NF-κB p65 was upregulated
by IR insult but was reduced by SAHA (Fig. 2C and D). Consistent with the
previous report that SAHA induces autophagy (25), SAHA also promoted autophagy in the
grafts (Fig. 2E and F). As
AKT/mTOR signaling is closely related to autophagy, the activity of
AKT/mTOR was evaluated by western blotting and it was found that
this pathway was downregulated by SAHA (Fig. 2C and D). These results indicate
that SAHA regulates the inflammatory response after IR insult
through the AKT/NF-κB pathway and promotes autophagy in grafts
through the AKT/mTOR pathway, but whether SAHA promotes KC
autophagy remains to be further investigated.
 | Figure 2SAHA inhibits inflammation and
promotes autophagy in grafts. (A) The protein levels of IL-1β,
TNF-α and IL-6 in liver tissue were measured by western blotting.
(B) Quantification of western blot data. (C) Western blot analysis
of the protein levels of p-AKT, AKT, P-P65, P65, p-mTOR and mTOR in
IR-livers. (D) Densitometric analysis of the western blot data. (E)
The protein levels of P62, ATG5 and LC3B in the liver were detected
by western blotting. (F) The relative protein levels of P62, ATG5
and LC3B. (G) The protein levels of ATG16L1 in the liver were
detected by western blotting. (H) The relative protein levels of
ATG16L1. *P<0.05 vs. the Sham group and
#P<0.05 vs. the IR+SA group. IL, interleukin; p-,
phosphorylated; IR, ischemia reperfusion; DMSO, dimethyl sulfoxide;
SA, suberoylanilide hydroxamic acid; CL, clodronate liposomes;
ATG5, autophagy 5 protein. |
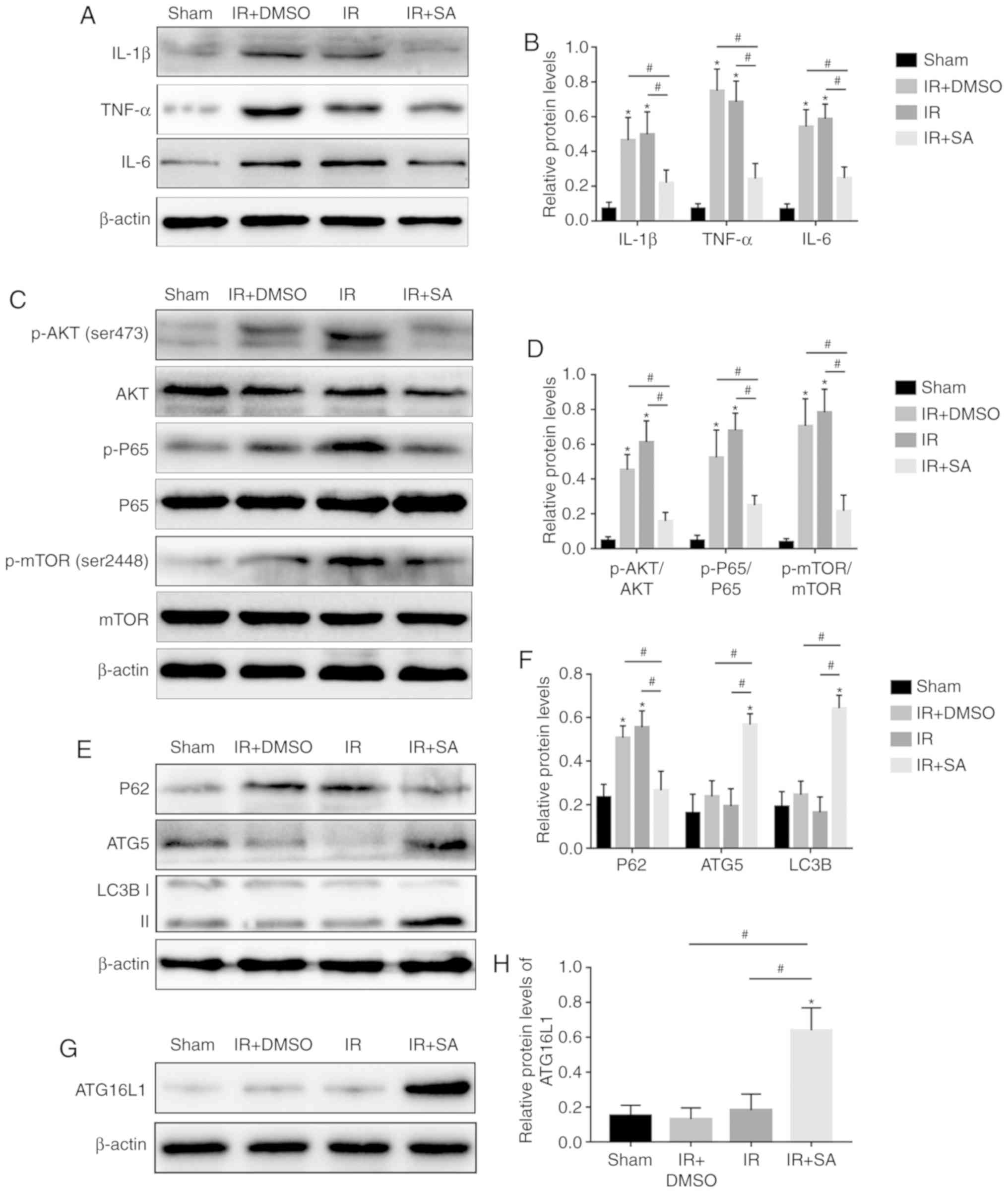
SAHA inhibits KC M1 polarization through
the AKT/GSK3β/NF-κB pathway in vitro
To further investigate the effect of SAHA on the
activation of KCs, LPS was used to induce sterile inflammation in
KCs that were isolated from normal rats. CD68+CD86+ KCs were
detected by flow cytometry and the results showed that the ratio of
M1 KCs in the LPS-treated group was much higher than that in the
normal group but was downregulated in the SAHA-treated group
(Fig. 3A and B). The
downregulated protein level of iNOS, an M1 macrophage polarization
marker, further confirmed that SAHA inhibited M1 polarization of
KCs (Fig. 3C and D). Next,
whether SAHA regulates M1 macrophage polarization through the
AKT/GSK3β/NF-κB pathway was explored. As shown in Fig. 3C and D, the activity of NF-κB p65
was upregu-lated by LPS, which was accompanied by upregulated p-AKT
(Ser473), but both were reduced by SAHA (Fig. 3C and D). Furthermore, LPS
inhibited the activity of GSK3β but was activated by SAHA (Fig. 3C and D). These results suggest
that SAHA reduces KC M1 polarization by inhibiting the
AKT/GSK3β/NF-κB pathway.
 | Figure 3SAHA inhibits KC M1 polarization
through the AKT/GSK3β/NF-κB pathway in vitro. (A) The ratio
of CD68+CD86+ Kupffer cells was measured by flow cytometry and (B)
analyzed. (C) Western blotting and (D) analysis of the protein
levels of p-AKT, AKT, p-GSK3β, GSK3β, p-P65, P65 and iNOS.
*P<0.05 vs. the normal group and
#P<0.05 vs. the LPS+SA group. DMSO, dimethyl
sulfoxide; SA, suberoylanilide hydroxamic acid; CL, clodronate
liposomes; KCs, Kupffer cells; p-, phosphorylated; iNOS, inducible
nitric oxide synthase; LPS, lipopolysaccharide; GSK, glycogen
synthase kinase. |
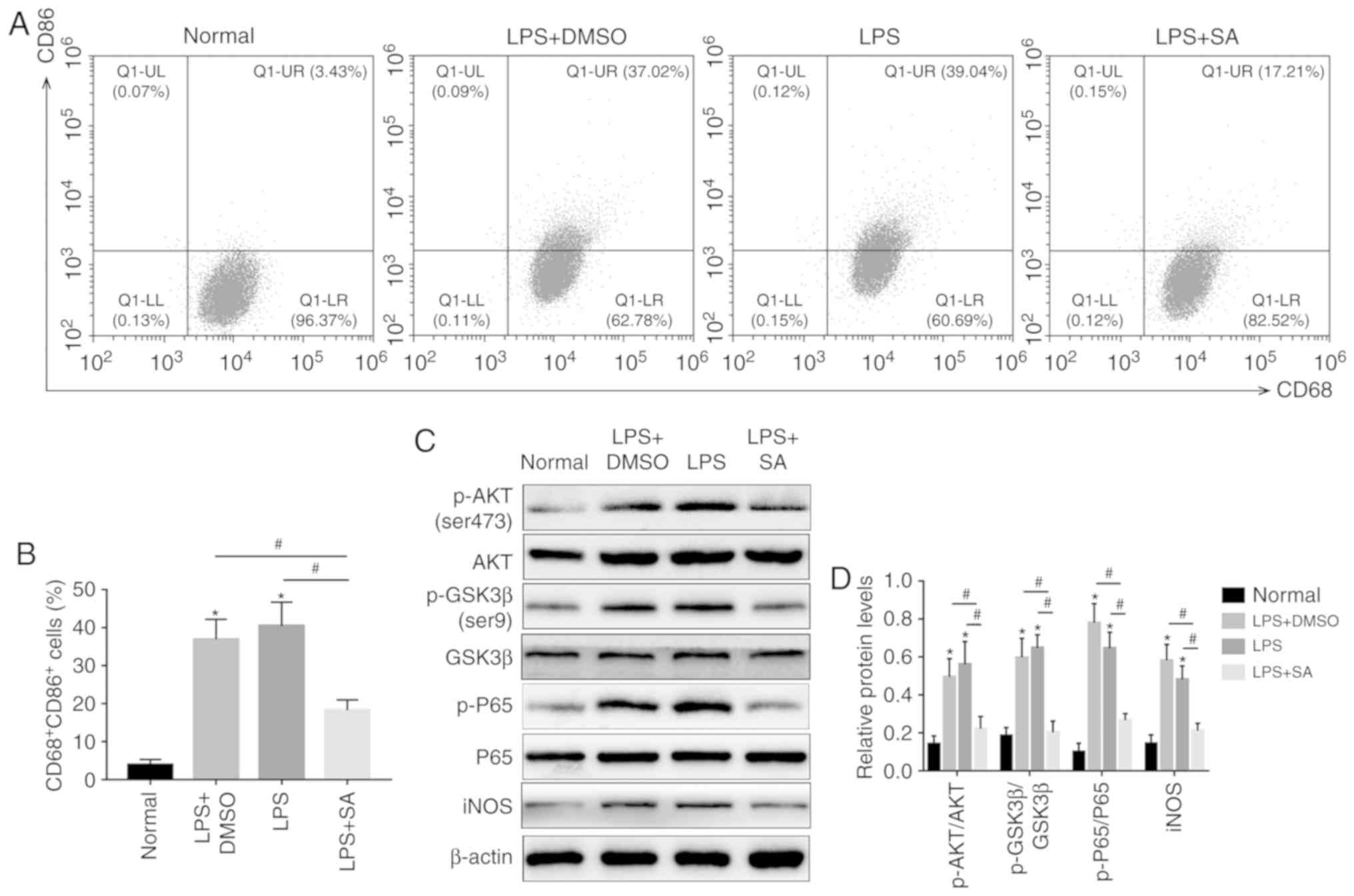
SAHA promotes autophagy in KCs by
inhibiting the AKT/mTOR pathway in vitro
Although SAHA promotes autophagy in liver grafts,
whether it promotes autophagy in KCs remains unknown. Therefore,
autophagy-related factors were detected in isolated KCs from normal
rat livers and the RT-qPCR results showed that ATG5 and LC3B were
both increased in the SAHA-treated group compared with those in the
LPS-treated group, accompanied by a reduction in P62 (Fig. 4A). SAHA-induced autophagy in KCs
was further confirmed by western blotting (Fig. 4B-E). Furthermore, the
immunofluorescence results showed that the fluorescence intensity
in the SAHA-treated group was much higher than that in the
LPS-treated group (Fig. 4F).
Moreover, the TEM results showed that there were significantly more
autophagic vacuoles in the LPS+SA group than in the Normal,
LPS+DMSO and LPS groups (Fig.
5A). AKT/mTOR signaling is one of the classical pathways
(26), and activation of AKT and
mTOR was detected by western blotting, which showed that
LPS-induced upregulation of p-mTOR (Ser2448) and p-AKT (Ser473) was
abrogated by SAHA (Fig. 4B and
C). These results indicate that SAHA promotes KC autophagy by
downregulating the AKT/mTOR pathway.
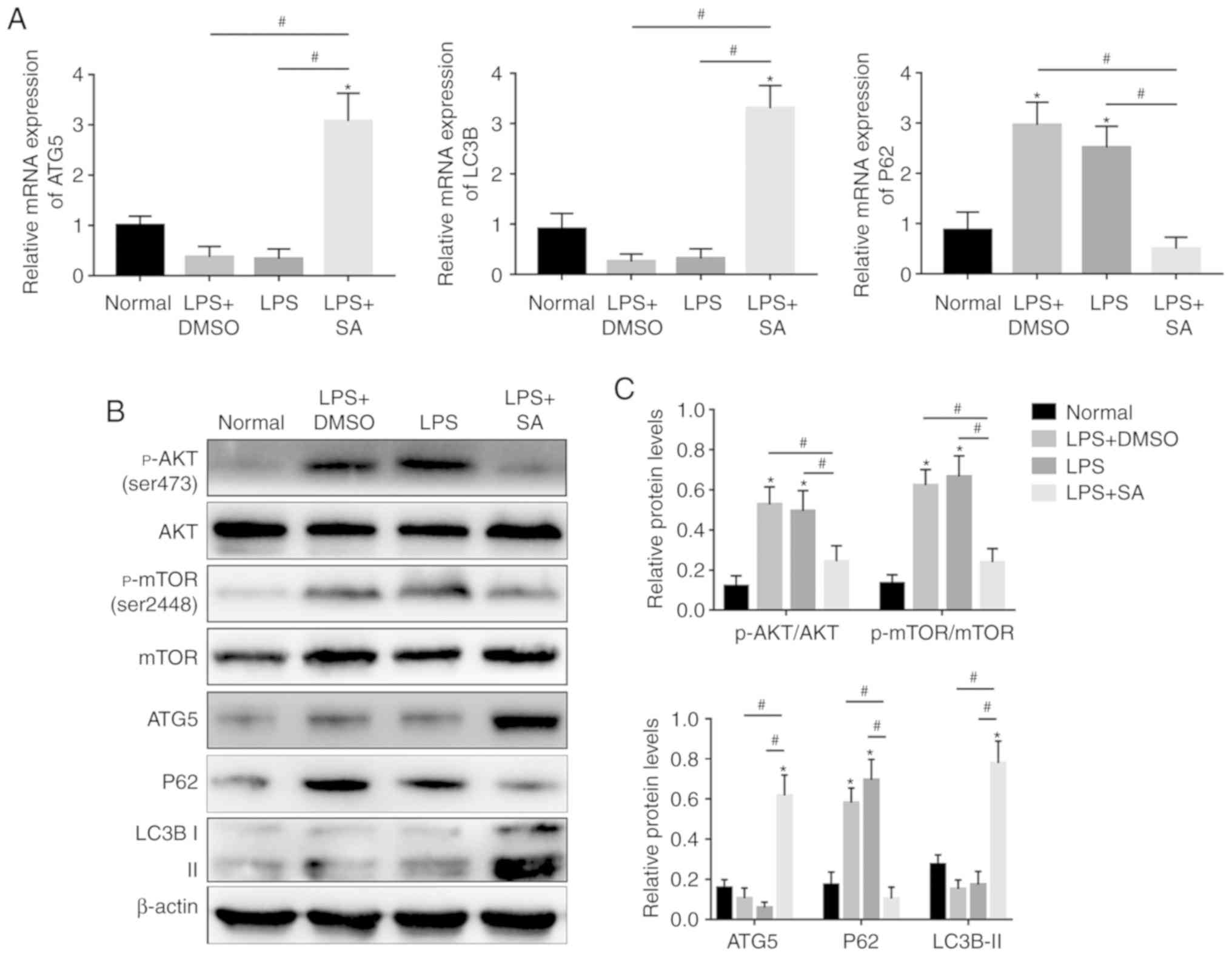 | Figure 4SAHA promotes autophagy in KCs by
inhibiting the AKT/mTOR pathway in vitro. (A) The mRNA
expression of ATG5, LC3B and P62 was analyzed by quantitative PCR.
(B) Western blotting and (C) analysis was used to detect the
expression of p-AKT, AKT, p-mTOR, mTOR, P62, ATG5 and LC3B. (D)
Western blotting and (E) analysis was used to detect the expression
of ATG16L1. (F) The fluorescence intensity of LC3B in KCs was
detected by fluorescence microscopy and DAPI was used for nuclear
staining (magnification, x200). *P<0.05 vs. the
normal group and #P<0.05 vs. the LPS+SA group. DAPI,
4'6-diamidino-2-phenylindole; IR, ischemia reperfusion; SA,
suberoylanilide hydroxamic acid; CL, clodronate liposomes; KCs,
Kupffer cells; LPS, lipopolysaccharide; p, phosphorylated; ATG5,
autophagy 5 protein. |
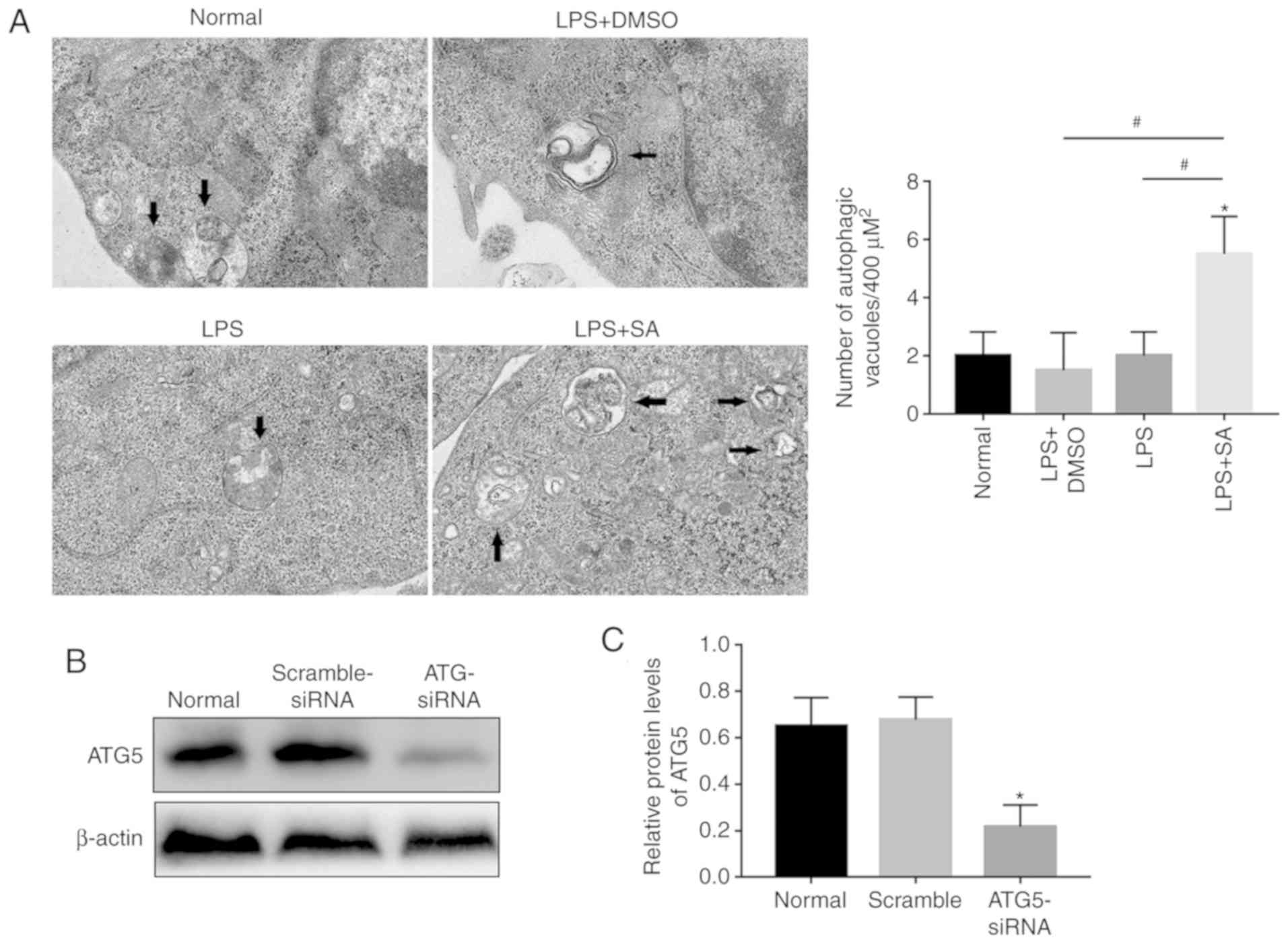 | Figure 5SAHA inhibits inflammatory cytokines
in KCs in an autophagy-dependent manner. (A) Autophagic vacuoles
(arrows) in KCs were observed by transmission electron micrograph
(n=5/group; magnification, x20,000). (B) Western blotting and (C)
analysis of the protein levels of ATG5 in KCs treated with or
without ATG5-siRNA. (D) Western blotting and (E) analysis of the
protein levels of IL-1β, IL6, TNFα, P62, ATG5 and LC3B in KCs. (F)
Western blotting and (G) analysis of the protein levels of ATG16L1.
*P<0.05 vs. the normal group and
#P<0.05 vs. the LPS+SA group. IR, ischemia
reperfusion; DMSO, dimethyl sulfoxide; SA, suberoylanilide
hydroxamic acid; CL, clodronate liposomes; KCs, Kupffer cells; LPS,
lipopolysaccharide; si, small interfering; IL, interleukin. |
To investigate the effect of autophagy on
inflammation, KCs were isolated from normal rat livers and treated
with ATG5-siRNA. As shown in Fig. 5B
and C, the upregu-lated proinflammatory cytokines in
LPS-treated KCs were downregulated by SAHA, and this effect was
reversed by knockdown of ATG5 (Fig.
5B and C), which confirmed that the SAHA-mediated reduction in
inflammation in KCs is partly dependent upon autophagy.
SAHA-mediated amelioration of liver
injury depends on KC autophagy
Published studies have shown that autophagy plays a
controversial role in liver ischemia reperfusion injury (11,46-48); therefore, the present study
investigated the role of autophagy in the protective effect of SAHA
on OLT-induced IRI by using the autophagy inhibitor CQ. The
increased levels of serum ALT and serum AST in the CQ-treated group
indicated that inhibition of autophagy partly impaired the
protective effect of SAHA on OLT-induced IRI (Fig. 6A). Since SAHA protects the liver
from cold IR in a KC-dependent manner, to further investigate the
role of KC autophagy in SAHA-mediated protection against IR-induced
liver injury, KC autophagy was downregulated by AAV-ATG5-shRNA
in vivo as described previously (43) (Fig.
6B and C). The protective effect of SAHA on OLT-induced IRI was
weakened in the AAV-ATG5-shRNA group, as increased levels of
hepatocyte apoptosis were found in the SAHA+AAV-ATG5-shRNA group
compared with those of the SAHA-treated group (Fig. 6D-G).
Discussion
Cold HIRI induced by OLT occurs early in liver
transplantation and seriously decreases the survival rate of liver
transplantation. The results of the current study demonstrate that
the histone deacetylase inhibitor SAHA reduced the levels of
proinflammatory cytokines and attenuated IR-induced liver injury in
a KC-dependent manner. Although SAHA plays an anti-inflammatory
role in various diseases (22,23,49), its role in cold HIRI remains
unclear. The present study showed that SAHA promoted autophagy in
KCs by inhibiting the AKT/mTOR pathway, which contributes to
ameliorating IR-induced liver injury. Moreover, SAHA reduced M1
polarization of KCs by inhibiting the AKT/GSK3β/NF-κB pathway.
Macrophages play a pivotal role in the initiation of
innate and adaptive immune responses by shifting between M1 and M2
phenotypes. M1 macrophages release proinflammatory cytokines such
as IL-1β and TNF-α, while M2 macrophages release anti-inflammatory
cytokines such as TGFβ and IL10 (50). KC, the resident macrophages in the
liver, are tightly associated with IR of the liver by triggering or
suspending inflammation (51). In
the present study, it was found that depletion of KCs impaired the
protective effect of SAHA on the IR-liver, which indicates that
SAHA attenuated OLT-induced IRI in a KC-dependent manner. During
hepatic ischemia reperfusion, TLR4, a surface receptor on KCs,
binds to danger signals such as damage-associated molecular
patterns to activate KCs (52).
In response to danger signal stimulation, activated KCs release
proinflammatory and anti-inflammatory cytokines to play a dual role
in modulating HIRI (53). A study
performed by Leoni et al (22) showed that SAHA decreases
proinflammatory cytokines released by LPS-stimulated peritoneal
macrophages, but its effect on KCs in IRI remains to be
elucidated.
Histone deacetylase inhibitors have been shown to
induce autophagy and anti-exert anti-inflammatory effects in
vitro and in vivo (54,55); SAHA ameliorates the outcomes of
cardiac ischemia reperfusion injury by inducing cardiomyocyte
autophagy (24), but its role in
cold liver IRI remains to be fully investigated. Autophagy is a
highly conserved metabolic process that maintains cellular
homeostasis by forming autophagic lysosomes to remove misfolded
proteins and damaged organelles (56). Evidence indicates that autophagy
is closely related to the inflammatory response; for example,
proinflammatory cytokines IL-1α, IL-1β and type I interferon are
increased in macrophages when autophagy is inhibited (57-59), while attenuated inflammation
occurs in autophagy-overexpressing conditions (60,61). In the present study, it was found
that SAHA reduced the upregulation of p-AKT and p-P65 after OLT,
combined with the downregulation of M1 KC polarization in the
SAHA-treated group in vitro. The current study detected
whether AKT/GSK3β/NF-κB, a signaling pathway that participates in
the inflammatory response of phagocytes (32), changed with SAHA treatment. The
results showed that SAHA downregulated this pathway, which
indicates that SAHA inhibits M1 polarization of KCs through the
AKT/GSK3β/NF-κB signaling pathway. Moreover, SAHA promoted
autophagy in KCs, accompanied by inhibition of AKT/mTOR signaling,
which suggests that SAHA enhances autophagy in KCs through the
AKT/mTOR pathway. Next, to explore the role of enhanced autophagy
by SAHA in cold liver IRI, ATG5 in KCs was knocked down by
AAV-ATG5-shRNA and the results showed that knockdown of ATG5 partly
diminished the protective effect of SAHA on IR-injured livers,
which is consistent with previous studies (57-59).
In conclusion, the present study shows that the
antitumor drug SAHA effectively alleviates OLT-induced IRI. SAHA
induces autophagy and inhibits M1 polarization of KCs, both of
which contribute to ameliorating OLT-induced IRI. These findings
provide evidence that SAHA may be an effective treatment for
OLT-induced IRI.
Acknowledgements
Not applicable.
Funding
This study received financial support from the
National Natural Science Foundation of China (grant no. 81871261),
the National Youth Foundation of China (grant no. 81600504), the
Chongqing Health Committee (grant no. 2018MSXM031), the Yibin
Science and Technology Plan Project (grant no. 2017ZSF007-10), and
the Sichuan Health and Wellness Committee (grant no. 17PJ112).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
JW, MD, JG and SL designed the experiments and wrote
the manuscript. JW, MD, HW and HB performed the experiments and
analyzed the data. YC, JP and YW analyzed the data and revised the
manuscript; they also provided assistance for the acquisition of
the experimental funds. MW and SL contributed to the TEM
experiments, and MW provided advice for the detection of ATG16L1.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments involved in this study were
in accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals and approved by the Ethics
Committee of the Second Affiliated Hospital of Chongqing Medical
University (Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bavarsad K, Riahi MM, Saadat S, Barreto G,
Atkin SL and Sahebkar A: Protective effects of curcumin against
ischemia-reperfusion injury in the liver. Pharmacol Res. 141:53–62.
2019. View Article : Google Scholar
|
|
2
|
Linecker M, Frick L, Kron P, Limani P,
Kambakamba P, Tschuor C, Langiewicz M, Kachaylo E, Tian Y,
Schneider MA, et al: Exercise improves outcomes of surgery on fatty
liver in mice: A novel effect mediated by the AMPK pathway. Ann
Surg. 271:347–355. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ju C and Tacke F: Hepatic macrophages in
homeostasis and liver diseases: From pathogenesis to novel
therapeutic strategies. Cell Mol Immunol. 13:316–327. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takemura S, Azuma H, Osada-Oka M, Kubo S,
Shibata T and Minamiyama Y: S-allyl-glutathione improves
experimental liver fibrosis by regulating Kupffer cell activation
in rats. Am J Physiol Gastrointest Liver Physiol. 314:G150–G163.
2018. View Article : Google Scholar
|
|
5
|
Elsegood CL, Chan CW, Degli-Esposti MA,
Wikstrom ME, Domenichini A, Lazarus K, van Rooijen N, Ganss R,
Olynyk JK and Yeoh GC: Kupffer cell-monocyte communication is
essential for initiating murine liver progenitor cell-mediated
liver regeneration. Hepatology. 62:1272–1284. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zigmond E, Samia-Grinberg S, Pasmanik-Chor
M, Brazowski E, Shibolet O, Halpern Z and Varol C: Infiltrating
monocyte-derived macrophages and resident kupffer cells display
different ontogeny and functions in acute liver injury. J Immunol.
193:344–353. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu Y, Yang C, Shen G, Yang S, Cheng X,
Cheng F, Rao J and Wang X: Hyperglycemia-triggered
sphingosine-1-phosphate and sphingosine-1-phosphate receptor 3
signaling worsens liver ischemia/reperfusion injury by regulating
M1/M2 polarization. Liver Transpl. 25:1074–1090. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng D, Li Z, Wei X, Liu R, Shen A, He D,
Tang C and Wu Z: Role of miR-148a in mitigating hepatic
ischemia-reperfusion injury by repressing the TLR4 signaling
pathway via targeting CaMKIIα in vivo and in vitro. Cell Physiol
Biochem. 49:2060–2072. 2018. View Article : Google Scholar
|
|
9
|
Raptis DA, Limani P, Jang JH, Ungethüm U,
Tschuor C, Graf R, Humar B and Clavien PA: GPR120 on Kupffer cells
mediates hepa-toprotective effects of ω3-fatty acids. J Hepatol.
60:625–632. 2014. View Article : Google Scholar
|
|
10
|
Allaire M, Rautou PE, Codogno P and
Lotersztajn S: Autophagy in liver diseases: Time for translation? J
Hepatol. 70:985–998. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choi Y, Bowman JW and Jung JU: Autophagy
during viral infection-a double-edged sword. Nat Rev Microbiol.
16:341–354. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang JH, Ahn IS, Fischer TD, Byeon JI,
Dunn WA JR, Behrns KE, Leeuwenburgh C and Kim JS: Autophagy
suppresses age-dependent ischemia and reperfusion injury in livers
of mice. Gastroenterology. 141:2188–2199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rickenbacher A, Jang JH, Limani P,
Ungethüm U, Lehmann K, Oberkofler CE, Weber A, Graf R, Humar B and
Clavien PA: Fasting protects liver from ischemic injury through
Sirt1-mediated downregulation of circulating HMGB1 in mice. J
Hepatol. 61:301–308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang JH, Behrns KE, Leeuwenburgh C and Kim
JS: Critical role of autophage in ischemia/reperfusion injury to
aged livers. Autophagy. 8:140–141. 2012. View Article : Google Scholar :
|
|
15
|
Nakamura K, Kageyama S, Yue S, Huang J,
Fujii T, Ke B, Sosa RA, Reed EF, Datta N, Zarrinpar A, et al: Heme
oxygenase-1 regulates sirtuin-1-autophagy pathway in liver
transplantation: From mouse to human. Am J Transplant.
18:1110–1121. 2018. View Article : Google Scholar
|
|
16
|
Zaouali MA, Boncompagni E, Reiter RJ,
Bejaoui M, Freitas I, Pantazi E, Folch-Puy E, Abdennebi HB,
Garcia-Gil FA and Roselló-Catafau J: AMPK involvement in
endoplasmic reticulum stress and autophagy modulation after fatty
liver graft preservation: A role for melatonin and trimetazidine
cocktail. J Pineal Res. 55:65–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Quesnelle KM, Bystrom PV and
Toledo-Pereyra LH: Molecular responses to ischemia and reperfusion
in the liver. Arch Toxicol. 89:651–657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu K, Zhao E, Ilyas G, Lalazar G, Lin Y,
Haseeb M, Tanaka KE and Czaja MJ: Impaired macrophage autophagy
increases the immune response in obese mice by promoting
proinflammatory macrophage polarization. Autophagy. 11:271–284.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nakahira K, Haspel JA, Rathinam VA, Lee
SJ, Dolinay T, Lam HC, Englert JA, Rabinovitch M, Cernadas M, Kim
HP, et al: Autophagy proteins regulate innate immune responses by
inhibiting the release of mitochondrial DNA mediated by the NALP3
inflammasome. Nat Immunol. 12:222–230. 2011. View Article : Google Scholar
|
|
20
|
Sun Y, Sun Y, Yue S, Wang Y and Lu F:
Histone deacetylase inhibitors in cancer therapy. Curr Top Med
Chem. 18:2420–2428. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Glauben R, Batra A, Stroh T, Erben U,
Fedke I, Lehr HA, Leoni F, Mascagni P, Dinarello CA, Zeitz M and
Siegmund B: Histone deacetylases: Novel targets for prevention of
colitis-associated cancer in mice. Gut. 57:613–622. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Leoni F, Zaliani A, Bertolini G, Porro G,
Pagani P, Pozzi P, Donà G, Fossati G, Sozzani S, Azam T, et al: The
antitumor histone deacetylase inhibitor suberoylanilide hydroxamic
acid exhibits antiinflammatory properties via suppression of
cytokines. Proc Natl Acad Sci USA. 99:2995–3000. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi SW, Gatza E, Hou G, Sun Y, Whitfield
J, Song Y, Oravecz-Wilson K, Tawara I, Dinarello CA and Reddy P:
Histone deacetylase inhibition regulates inflammation and enhances
Tregs after allogeneic hematopoietic cell transplantation in
humans. Blood. 125:815–819. 2015. View Article : Google Scholar :
|
|
24
|
Xie M, Kong Y, Tan W, May H, Battiprolu
PK, Pedrozo Z, Wang ZV, Morales C, Luo X, Cho G, et al: Histone
deacetylase inhibition blunts ischemia/reperfusion injury by
inducing cardio-myocyte autophagy. Circulation. 129:1139–1151.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang J, Wang J, Zhou Z, Park JE, Wang L,
Wu S, Sun X, Lu L, Wang T, Lin Q, et al: Importance of TFEB
acetylation in control of its transcriptional activity and
lysosomal function in response to histone deacetylase inhibitors.
Autophagy. 14:1043–1059. 2018.PubMed/NCBI
|
|
26
|
Chiao MT, Cheng WY, Yang YC, Shen CC and
Ko JL: Suberoylanilide hydroxamic acid (SAHA) causes tumor growth
slowdown and triggers autophagy in glioblastoma stem cells.
Autophagy. 9:1509–1526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang P, Guo Z, Wu Y, Hu R, Du J, He X,
Jiao X and Zhu X: Histone deacetylase inhibitors inhibit the
proliferation of gallbladder carcinoma cells by suppressing
AKT/mTOR signaling. PLoS One. 10:e01361932015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu YL, Yang PM, Shun CT, Wu MS, Weng JR
and Chen CC: Autophagy potentiates the anti-cancer effects of the
histone deacetylase inhibitors in hepatocellular carcinoma.
Autophagy. 6:1057–1065. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hrzenjak A, Kremser ML, Strohmeier B,
Moinfar F, Zatloukal K and Denk H: SAHA induces
caspase-independent, autophagic cell death of endometrial stromal
sarcoma cells by influencing the mTOR pathway. J Pathol.
216:495–504. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang X, Ikejima K, Kon K, Arai K, Aoyama
T, Okumura K, Abe W, Sato N and Watanabe S: Ursolic acid
ameliorates hepatic fibrosis in the rat by specific induction of
apoptosis in hepatic stellate cells. J Hepatol. 55:379–387. 2011.
View Article : Google Scholar
|
|
31
|
Fan Z, Li L, Li M, Zhang X, Hao C, Yu L,
Zeng S, Xu H, Fang M, Shen A, et al: The histone methyltransferase
Suv39h2 contributes to nonalcoholic steatohepatitis in mice.
Hepatology. 65:1904–1919. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cremer TJ, Shah P, Cormet-Boyaka E,
Valvano MA, Butchar JP and Tridandapani S: Akt-mediated
proinflammatory response of mononuclear phagocytes infected with
Burkholderia cenocepacia occurs by a novel GSK3β-dependent, IκB
kinase-independent mechanism. J Immunol. 187:635–643. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu A, Guo E, Yang J, Yang Y, Liu S, Jiang
X, Hu Q, Dirsch O, Dahmen U, Zhang C, et al: Young plasma reverses
age-dependent alterations in hepatic function through the
restoration of autophagy. Aging Cell. 17:2018. View Article : Google Scholar
|
|
34
|
Kamada N and Calne RY: Orthotopic liver
transplantation in the rat. Technique using cuff for portal vein
anastomosis and biliary drainage. Transplantation. 28:47–50. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li PZ, Li JZ, Li M, Gong JP and He K: An
efficient method to isolate and culture mouse Kupffer cells.
Immunol Lett. 158:52–56. 2014. View Article : Google Scholar
|
|
36
|
Zeng WQ, Zhang JQ, Li Y, Yang K, Chen YP
and Liu ZJ: A new method to isolate and culture rat kupffer cells.
PLoS One. 8:e708322013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Piao X, Yamazaki S, Komazawa-Sakon S,
Miyake S, Nakabayashi O, Kurosawa T, Mikami T, Tanaka M, Van
Rooijen N, Ohmuraya M, et al: Depletion of myeloid cells
exacerbates hepatitis and induces an aberrant increase in histone
H3 in mouse serum. Hepatology. 65:237–252. 2017. View Article : Google Scholar
|
|
38
|
Pan G, Zhao Z, Tang C, Ding L, Li Z, Zheng
D, Zong L and Wu Z: Soluble fibrinogen-like protein 2 ameliorates
acute rejection of liver transplantation in rat via inducing
Kupffer cells M2 polarization. Cancer Med Cancer Med. 7:3168–3177.
2018. View Article : Google Scholar
|
|
39
|
Yang J, He J, Ismail M, Tweeten S, Zeng F,
Gao L, Ballinger S, Young M, Prabhu SD, Rowe GC, et al: HDAC
inhibition induces autophagy and mitochondrial biogenesis to
maintain mitochondrial homeostasis during cardiac
ischemia/reperfusion injury. J Mol Cell Cardiol. 130:36–48. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kang JW, Cho HI and Lee SM: Melatonin
Inhibits mTOR-Dependent Autophagy during Liver
Ischemia/Reperfusion. Cell Physiol Biochem. 33:23–36. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hsieh IN, Liou JP, Lee HY, Lai MJ, Li YH
and Yang CR: Preclinical anti-arthritic study and pharmacokinetic
properties of a potent histone deacetylase inhibitor MPT0G009. Cell
Death Dis. 5:e11662014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Luo X, Wang D, Zhu X, Wang G, You Y, Ning
Z, Li Y, Jin S, Huang Y, Hu Y, et al: Autophagic degradation of
caveolin-1 promotes liver sinusoidal endothelial cells
defenestration. Cell Death Dis. 9:5762018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li P, Liu H, Zhang Y, Liao R, He K, Ruan X
and Gong J: Endotoxin tolerance inhibits degradation of tumor
necrosis factor receptor-associated factor 3 by suppressing pellino
1 expression and the K48 ubiquitin ligase activity of cellular
inhibitor of apoptosis protein 2. J Infect Dis. 214:906–915. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
45
|
Sun HY, Hu YJ, Zhao XY, Zhong Y, Zeng LL,
chen XB, Yuan J, Wu J, Sun Y, Kong W and Kong WJ: Age-related
changes in mitochondrial antioxidant enzyme Trx2 and
TXNIP-Trx2-ASK1 signal pathways in the auditory cortex of a mimetic
aging rat model: Changes to Trx2 in the auditory cortex. FEBS J.
282:2758–2774. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nakamura K, Kageyama S, Ito T, Hirao H,
Kadono K, Aziz A, Dery KJ, Everly MJ, Taura K, Uemoto S, et al:
Antibiotic pretreatment alleviates liver transplant damage in mice
and humans. J Clin Invest. 129:3420–3434. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu H, Dong J, Song S, Zhao Y, Wang J, Fu
Z and Yang J: Spermidine ameliorates liver ischaemia-reperfusion
injury through the regulation of autophagy by the AMPK-mTOR-ULK1
signalling pathway. Biochem Biophys Res Commun. 519:227–233. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu A, Yang J, Hu Q, Dirsch O, Dahmen U,
Zhang C, Gewirtz DA, Fang H and Sun J: Young plasma attenuates
age-dependent liver ischemia reperfusion injury. FASEB J.
33:3063–3073. 2019. View Article : Google Scholar
|
|
49
|
Ratay ML, Balmert SC, Bassin EJ and Little
SR: Controlled release of an HDAC inhibitor for reduction of
inflammation in dry eye disease. Acta Biomater. 71:261–270. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schmieder A, Michel J, Schönhaar K, Goerdt
S and Schledzewski K: Differentiation and gene expression profile
of tumor-associated macrophages. Semin Cancer Biol. 22:289–297.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang J, Koh HW, Zhou L, Bae UJ, Lee HS,
Bang IH, Ka SO, Oh SH, Bae EJ and Park BH: Sirtuin 2 aggravates
postischemic liver injury by deacetylating mitogen-activated
protein kinas-ephosphatase-1. Hepatology. 65:225–236. 2017.
View Article : Google Scholar
|
|
52
|
Tsung A, Klune JR, Zhang X, Jeyabalan G,
Cao Z, Peng X, Stolz DB, Geller DA, Rosengart MR and Billiar TR:
HMGB1 release induced by liver ischemia involves toll-like receptor
4 dependent reactive oxygen species production and calcium-mediated
signaling. J Exp Med. 204:2913–2923. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tasnim F, Xing J, Huang X, Mo S, Wei X,
Tan MH and Yu H: Generation of mature kupffer cells from human
induced pluripotent stemcells. Biomaterials. 192:377–391. 2019.
View Article : Google Scholar
|
|
54
|
Zhang J, Ng S, Wang J, Zhou J, Tan SH,
Yang N, Lin Q, Xia D and Shen HM: Histone deacetylase inhibitors
induce autophagy through FOXO1-dependent pathways. Autophagy.
11:629–642. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hancock WW, Akimova T, Beier UH, Liu Y and
Wang L: HDAC inhibitor therapy in autoimmunity and transplantation.
Ann Rheum Dis. 71(Suppl 2): i46–i54. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yamamoto A and Yue Z: Autophagy and its
normal and pathogenic states in the brain. Annu Rev Neurosci.
37:55–78. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lee JP, Foote A, Fan H, Peral de Castro C,
Lang T, Jones SA, Gavrilescu N, Mills KH, Leech M, Morand EF and
Harris J: Loss of autophagy enhances MIF/macrophage migration
inhibitory factor release by macrophages. Autophagy. 12:907–916.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Tal MC, Sasai M, Lee HK, Yordy B, Shadel
GS and Iwasaki A: Absence of autophagy results in reactive oxygen
species-dependent amplification of RLR signaling. Proc Natl Acad
Sci USA. 106:2770–2775. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Castillo EF, Dekonenko A, Arko-Mensah J,
Mandell MA, Dupont N, Jiang S, Delgado-Vargas M, Timmins GS,
Bhattacharya D, Yang H, Hutt J, et al: Autophagy protects against
active tuberculosis by suppressing bacterial burden and
inflammation. Proc Natl Acad Sci USA. 109:E3168–E3176. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Eisenberg T, Abdellatif M, Schroeder S,
Primessnig U, Stekovic S, Pendl T, Harger A, Schipke J, Zimmermann
A, Schmidt A, et al: Cardioprotection and lifespan extension by the
natural polyamine spermidine. Nat Med. 22:1428–1438. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sun Y, Yao X, Zhang QJ, Zhu M, Liu ZP, Ci
B, Xie Y, Carlson D, Rothermel BA, Sun Y, et al: Beclin-1-dependent
autophagy protects the heart during sepsis. Circulation.
138:2247–2262. 2018. View Article : Google Scholar : PubMed/NCBI
|