Introduction
Peripheral nerve injury (PNI) is a common condition
caused by trauma, burn damage or surgical intervention, and often
results in permanent motor function disability and sensory
perception (1). The peripheral
nervous system (PNS) differs from the central nervous system (CNS)
as it exhibits a certain capacity for axonal self-regeneration.
However, the spontaneous regeneration of PNS is always incomplete
and results in poor clinical outcomes (2). Schwann cells (SCs) have gained
increasing attention in the field of peripheral nerve regeneration
(PNR) owing to their vital role in the process (3). Following PNI, axons of distal
impaired segments exhibit dystrophy, leading to axonal
demyelination and Wallerian degeneration, while the expression of
SC genes alter rapidly, switching the neuro-transmitter state to a
pro-regenerative state and forming Bungner's bands (4). SCs proliferate and signal to cells
to aggregate in the lesions to clear axons and myelin debris
(5). They also secrete
neurotrophic factors (NTFs) to prevent the apoptosis of damaged
neurons and provide an enabling environment for axonal regeneration
(6). However, the detailed
mechanisms underlying SCs PNR are complex; they are not fully
understood and require further investigation.
Glycoprotein non-metastatic melanoma protein B
(GPNMB), also known as hematopoietic growth factor inducible
neurokinin-1 (HGFIN), osteoactivin (rat orthologue) and DC-HIL
(mouse orthologue), is widely expressed in various tissues and
cells, including breast cancer cells, macrophages, melanoma cells,
bladder cancer cells and osteoclasts (7). It has been demonstrated that GPNMB
is involved in various biological processes, including malignant
tumor cell differentiation, inflammation, regeneration, invasion
and metastasis (8). GPNMB also
promotes the proliferation, migration and differentiation of adult
bone marrow mesenchymal stem cells and dental pulp cells (9,10).
Additionally, GPNMB has been reported to have neuroprotective
functions in amyotrophic lateral sclerosis (11) and cerebral ischemia reperfusion
injury (12), and protective
effects against mutant TDP-43-induced motor neuron cell death
(13) in the CNS. Recently, it
has been determined that the expression levels of GPNMB in SCs
harvested from injured sciatic nerves are increased in vitro
(14). However, the expression of
GPNMB in vivo, as well as its role following PNI, remains
unknown.
The present study investigated the expression of
GPNMB in the distal sciatic nerve following transection via
microarray analysis, the results of which were validated by reverse
transcription-quantitative PCR (RT-qPCR), western blot analysis
(WB) and immunohistochemistry (IHC). Furthermore, denervated SCs
harvested from distal stumps post-sciatic nerve transection were
treated with various doses of recombinant human GPNMB (rhGPNMB).
Then, cell proliferation, expression and secretion of NTFs and
neural adhesion molecules (NAMs) were detected in vitro. The
results revealed that GPNMB promoted the proliferation of SCs, as
well as the expression and secretion of NTFs and NAMs. The results
indicated that GPNMB may be utilized as a novel treatment strategy
for PNR.
Materials and methods
Animals and tissue preparation
Adult male Sprague-Dawley (SD) rats were obtained
from the Animal Center of Second Military Medical University and
all animal experiments were approved and supervised by the Animal
Care and Use Committee of the Second Military Medical University
(permit no. SYXK-2002-042). All rats were kept in cages under
controlled conditions at 22°C with ad libitum access to food
and water and a 12-12 h day/night cycle.
A total of 84 rats (weight, 180-200 g) were randomly
divided into seven groups of 12 rats each. Following anesthesia by
intra-peritoneal injection of pentobarbital sodium (40 mg/kg), the
bilateral sciatic nerves were exposed 2 mm from distal ischial
tuberosity and transected with fine scissors, after which the
incision sites were closed. No rats died following sciatic nerve
transection. Then, the distal injured sciatic nerves of each rat
were obtained in 5 min at day 0, 1, 3, 7, 14, 21 and 28
post-transection. After distal sciatic nerves were obtained, rats
remained under anesthesia and were immediately sacrificed by
cervical dislocation. Following confirmation of an absence of
respiration, the rats were collected and handed over to the Animal
Center of Second Military Medical University for disposal.
Microarray analysis
Microarray experiments (n=3) were performed by OE
Biotech Co., Ltd. Total RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) and quantified using
NanoDrop ND-2000 (Thermo Fisher Scientific, Inc.). RNA integrity
was assessed using the Agilent Bioanalyzer 2100 (Agilent
Technologies, Inc.). Sample labeling (Quick Amp Labeling kit; cat.
no. 5190-2305), microarray hybridization (Agilent Gene Expression
Hybridization kit; cat. no. 5188-5242) and washing (Gene Expression
Wash Buffer; cat. no. 5188-5325 and 5188-5326; all Agilent
Technologies, Inc.) were performed based on the manufacturer's
standard protocols. Total RNA was transcribed to double-stranded
cDNA, then synthesized into cRNA and labeled with Cyanine-3-CTP.
Labeled cRNAs were subsequently hybridized onto the 8x60 K Agilent
Rat long non-coding RNA (lncRNA) Array. Following washing, the
arrays were scanned using the Agilent Scanner G2505C (Agilent
Technologies, Inc.). Feature Extraction software (v.10.7.1.1;
Agilent Technologies, Inc.) was used to analyze array images to
obtain raw data. Genespring software (v.13.1; Agilent Technologies,
Inc.) was utilized to conclude basic analysis using raw data.
Search for specific mRNA profiles
Short time-series expression miner (STEM) analysis
(15) was performed to obtain
LncRNA and mRNA expression profiles, which were subsequently
compared for significance and false discovery rate. Differentially
expressed genes at certain time points were obtained via screening.
Using the mRNAs with significant expression levels, hierarchical
clustering to determine the distinguishable gene expression
patterns among samples, Gene ontology (GO) (16,17) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) (18) enrichment
analyses were conducted to determine the roles of these
differentially expressed mRNAs.
Culture of denervated SCs from distal
stumps of resected sciatic nerve
Denervated SCs were harvested as previously
described (19). A total of 4
weeks after sciatic nerve transection, distal stumps were harvested
and washed with PBS, after which the epineurium and perineurium
were stripped and the tissue was sliced into 1x1x1 mm fragments.
The fragments were then cultured in DMEM Nutrient Mixture F-12
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 5% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). After 7
days of preparation, nerve segments were seeded in
poly-L-ornithine/Laminin (Gibco; Thermo Fisher Scientific, Inc.)
coated 6- or 24-well plates and cultured in culture medium
consisting of DMEM/F12, 10% FBS, 1% Glutamine, 100 U/ml penicillin
and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.)
and 5 µM forskolin (Sigma-Aldrich; Merck KGaA). After 48 h
incubation, SCs were released from the nerve segments. The medium
was replaced with culture medium supplemented with 2 M cytarabine
(Sigma-Aldrich; Merck KGaA) for purification. After 72 h, the
medium with cytarabine was replaced with culture medium. When cells
grew to 90% confluence, they were passaged. Cells from passage 4
were used for all experiments.
rhGPNMB treatment
rhGPNMB (R&D Systems, Inc.), which included the
sequences of human GPNMB from Lysine 23 to Asparagine 486, was
dissolved in PBS. Passage 4 SCs were divided into four groups and
treated with 0, 10, 50 and 100 nM rhGPNMB, respectively. The 0 nM
rhGPNMB-treated group served as the control. SCs were treated for
48 h, after which they were examined.
SC proliferation assay
SC proliferation was determined using a Cell
Counting Kit 8 (CCK8; Dojindo Molecular Technologies, Inc.) assay.
rhGPNMB treated SCs were seeded into 96-well plates at 3,000
cells/well and incubated for 24 h in a humidified atmosphere with
5% CO2. Cells were then incubated with 10 µl CCK8
solution per well at 37°C for 1 h, after which the absorbance was
measured using a microplate reader (BioTek Instruments, Inc.) at
450 nm. Results were presented as mean optical density values
(n=6).
RT-qPCR
Total RNA (n=6) was extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.) and
concentration was detected using a microplate reader (BioTek
Instruments, Inc.). cDNA was subsequently synthesized using the
PrimeScript™ RT Reagent kit (Takara Bio, Inc.) in accordance with
the manufacturer's protocol. The expression of mRNA was determined
by SYBR-Green I-based qPCR using the Bio-Rad 3000 Quantitative
Real-time PCR System (Bio-Rad Laboratories, Inc.). The primers used
for PCR are presented in Table I.
The efficiency and specificity values of each primer set were
confirmed by comparing the standard curves of the threshold cycle
values to the RNA serial dilutions and melting curve evaluations.
GAPDH was used as an internal control to normalize gene expression
levels and the relative expression of target mRNA was calculated
using the 2−ΔΔCq method (20).
 | Table ISequences of primers used for RT-qPCR
analysis. |
Table I
Sequences of primers used for RT-qPCR
analysis.
| Target | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| GPNMB |
ATGTCCTCATTCACGATCC |
TGAAGGTTCCATTGAGCAC |
| NGF |
CATCACTGTGGACCCCAAACTGT |
GTCCGTGGCTGTGGTCTTATCTC |
| BDNF |
TTGATGAGACCGGGTTCCCT |
GTCCGTGGACGTTTGCTTCTT |
| NT-3 |
GCGATGCTACCGCAAAGAAC |
ATGGCGTCTCCCACACTACC |
| NCAM |
CAAGTCCCTAGACTGGAACGC CC |
TTGGATTTTCCTTGCTGGT |
| N-cadherin CC |
AAATCCTAAAATCATTCGCC |
CACAGGGTCTATTTTCAGCCAGT |
| GAPDH |
GCCATCAACGACCCCTTCAT |
TTCACACCCATCACAAACA |
WB
Distal sciatic nerves (n=6) were harvested at day 0,
1, 3, 7, 14, 21 and 28 post-transection and SCs that were treated
with rhGPNMB for 48 h were washed with PBS, lysed using RIPA assay
buffer (cat. no. G2002-100; Service Biotechnology Co., Ltd.) and
centrifuged at 4°C at 12,000 x g for 10 min to harvest the
supernatant. Protein concentration was then detected using a
Bicinchoninic Acid kit (Shanghai Yeasen Biotechnology Co., Ltd.).
The supernatant was boiled at 100°C and centrifuged at 1,000 x g
for 5 min for later use. Following preparation of the separation
gel, SDS-PAGE was performed. An equal quantity (10 µg/lane) of
extracted protein was resolved on a (10-15%) polyacrylamide
gradient. Proteins were then transferred to a nitrocellulose
membrane. The membrane was blocked in 5% skimmed milk/PBS for 1 h
at 4°C and then incubated overnight at 4°C with the following
primary antibodies: Rabbit polyclonal anti-GPNMB (cat. no.
GXP212342; 1:2,000; GenXspan, Inc.), rabbit monoclonal anti-nerve
growth factor (NGF; cat. no. ab6199; 1:2,000; Abcam), rabbit
monoclonal anti-brain-derived neurotrophic factor (BDNF; cat. no.
ab108319; 1:2,000; Abcam), rabbit polyclonal anti-neurotrophin 3
(NT-3; cat. no. ab16640; 1:2,000; Abcam), mouse monoclonal
anti-neural cell adhesion molecule (NCAM; cat. no. ab9018; 1:2,000;
Abcam), mouse monoclonal anti-neural cadherin (N-cadherin; cat. no.
ab98952; 1:2,000; Abcam) and GAPDH (cat. no. WB2197; 1:2,000;
Biotech Well Co., Ltd.). The sample was then incubated at 4°C for 1
h with horseradish peroxidase (HRP)-labeled secondary antibodies
which were diluted in 0.01 M TBST; goat anti-rabbit secondary
antibody (cat. no. 111-035-003; 1:5,000) and goat anti-mouse
secondary antibody (cat. no. 115-035-003; 1:5,000, both Jackson
ImmunoResearch Laboratories, Inc.). Subsequently, the membrane was
incubated with the ECL hypersensitive chemiluminescence kit (cat.
no. WB0164; Biotech Well Co., Ltd.) and chemiluminescence was
detected following exposure to X-rays. The bands were quantified by
measuring the band intensity of each group and normalizing it to
GAPDH using Odyssey v.1.2 software (LI-COR Biosciences).
IHC
An IHC kit (Wuhan Boster Biological Technology,
Ltd.) was used to perform IHC in accordance with the manufacturer's
protocol. Distal segments of transected sciatic nerve were fixed in
4% paraformaldehyde at 4°C for 24 h, dehydrated, at room
temperature using a gradient alcohol series (75% for 2 h, 85% for
1.5 h, 95% for 1 h, and 100% for 30 min), vitrified by
dimethylbenzene for 2 min, embedded in paraffin, and sectioned.
Sections were subsequently baked at 60°C for 1 h, deparaffinated at
room temperature by dimethylbenzene (2 times for 15 min each),
treated in a gradient alcohol series (100% for 5 min, 95% for 5
min, 85% for 5 min and 75% for 5 min), rehydrated, subjected to
antigen retrieval by heating sections in 0.01 M sodium citrate
buffer (pH 6.0) for 15 min at 96–98°C and cooled to room
temperature. Following peroxidase blocking with 3% hydrogen
peroxide at room temperature for 10 min, samples were further
blocked with blocking solution containing 10% goat serum (cat. no.
AR1009; Wuhan Boster Biological Technology, Ltd.) at 37°C for 30
min. Sections were then incubated with rabbit polyclonal anti-GPNMB
primary antibody (1:50; GenXspan, Inc.) at 4°C overnight followed
by incubation at room temperature for 30 min. Following washing
with 0.01 M PBS, sections were incubated with biotin-labeled
anti-rabbit IgG secondary antibody (1:200; cat. no. SA1052; Wuhan
Boster Biological Technology, Ltd.) for 1 h at 37°C. Then, 0.01%
DAB solution (Wuhan Boster Biological Technology, Ltd.) was added
according to the manufacturer's protocol, sections were rinsed with
distilled water and sealed with neutral resin. Images were captured
under a microscope (magnification, x200; Leica Microsystems GmbH)
(n=3).
ELISA
ELISA kits was used to measure levels of NGF (cat.
no. ER0143), BDNF (cat. no. ER0102), NT-3 (cat. no. ER0144; all
Biotech Well Co., Ltd.), NCAM (cat. no. EK1562) and N-cadherin
(cat. no. EK1156, both Wuhan Boster Biological Technology, Ltd.) in
culture medium following the manufacturer's protocol. SCs were
seeded into 6-well plates at 5×105 cells/well and
cultured in serum-free medium. After 48 h induction, supernatants
were collected and used to measure NGF, BDNF, NT-3, NCAM and
N-cadherin levels secreted by SCs (n=8).
Statistical analysis
All the aforementioned tests were performed at least
in triplicate and were repeated at least 3 times. The results were
expressed as the mean ± standard deviation and analyzed using SPSS
v.21.0 software (IBM Corp.). Figures were generated using Prism
v.6.0 software (GraphPad Software, Inc.). A one way analysis of
variance followed by Tukey's post hoc test was used for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Bioinformatics analyses of microarray
data
Microarray data were divided into 50 profiles to
summarize the expression of mRNAs and long non-coding RNAs
(lncRNAs) via STEM analysis. The expression tendency of profile 41,
which increased, peaked and then decreased was similar to the
proliferation tendency of distal acute denervated SCs. Considering
the vital roles of SCs in PNR, profile 41 was selected for further
analyses.
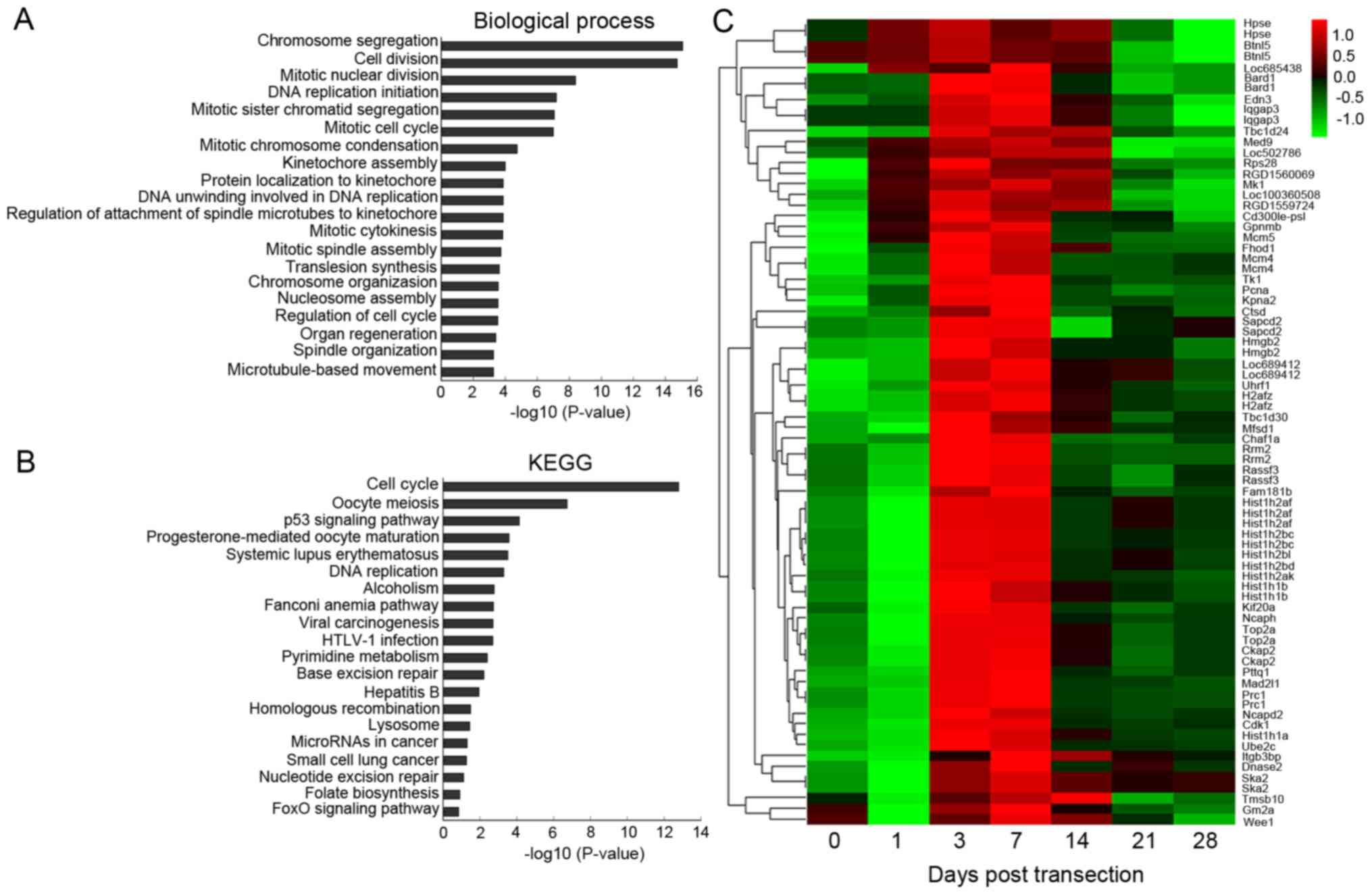
Profile 41 consisted of 76 mRNAs and 282 lncRNAs,
and was selected for bioinformatics analyses. The results of GO
analysis (Table SI) revealed
that profile 41 was associated with cell division, mitotic nuclear
division and DNA replication initiation (Fig. 1A). KEGG analysis (Table SII) confirmed the results of GO
analysis, demonstrating that profile 41 was associated with the
cell cycle, the p53 signaling pathway and DNA replication (Fig. 1B). The results of GO and KEGG
analyses indicated that profile 41 may be involved in cell
proliferation following transection. In addition, hierarchical
clustering analysis was performed to reveal systematic variations
in the 76 markedly altered mRNA expression profiles of profile 41
(Fig. 1C). The expression level
of GPNMB, the most significantly altered mRNA, increased from day
1, peaked at day 7, which was ~48-fold higher compared with that of
day 0, and then gradually decreased from day 14.
Altered expression of GPNMB in distal
sciatic nerves post-transection
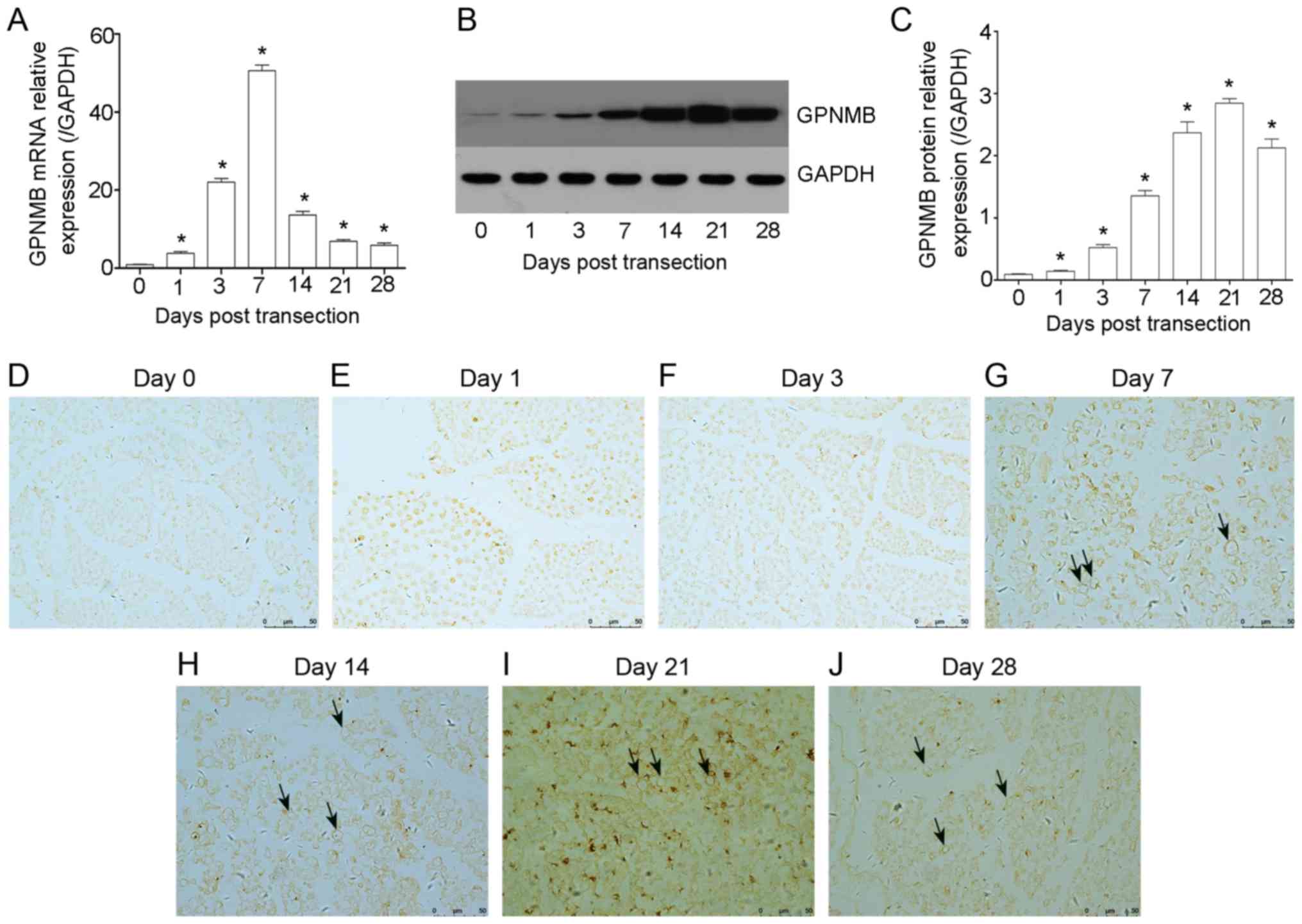
GPNMB, the most significantly altered mRNA, was
selected for analysis via RT-qPCR, WB and IHC in distal stumps at
certain time points post-transection. The results revealed that the
expression of GPNMB increased at day 1 (3.87±0.43-fold increase
compared with that of day 0; P<0.05), rapidly rose from day 3,
peaked at day 7 (48.63±3.71-fold increase compared with that of day
0, P<0.05) and gradually decreased from day 14 to day 28
(Fig. 2A). The alteration
tendency of GPNMB at the transcriptional level was consistent with
microarray data. Furthermore, similar results in GPNMB protein
expression were obtained by WB (Fig.
2B and C). The results revealed that the expression of GPNMB
rose at day 1 and 3 post injury, significantly increased from day
7, peaked at day 21 and then gradually decreased at day 28.
Although the time point at which GPNMB reached its peak varied
following RT-qPCR and WB, the results were concomitant with the
microarray data.
IHC was performed to clarify the expression and
localization of GPNMB in transverse (Fig. 2D-J) and longitudinal (Fig. S1) sections following transection.
The results revealed that the expression of GPNMB was consistent
with WB. Additionally, it was demonstrated that GPNMB was expressed
around the nerve fibers, indicating that GPNMB may be expressed in
SCs (Fig. 2).
These results demonstrated that the expression of
GPNMB was altered in the distal sciatic nerves
post-transection.
GPNMB promotes the proliferation of
SCs
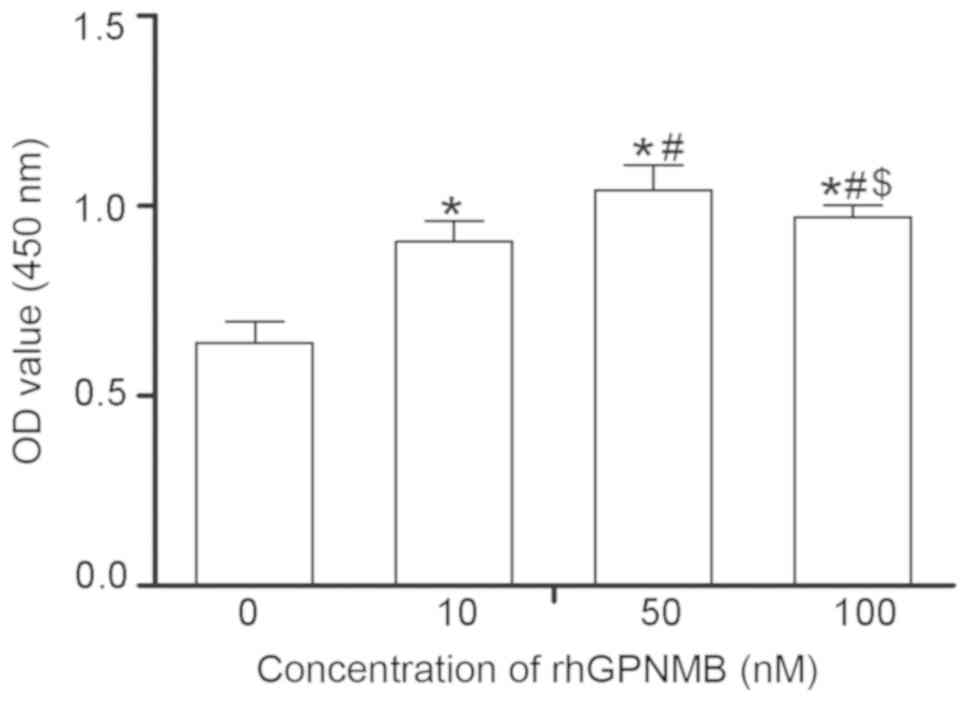
Following exposure to various doses of rhGPNMB for
48 h, the proliferation of SCs was detected using CCK-8 (Fig. 3). The results revealed that SCs
proliferated at an increased rate compared with the control when
cultured with rhGPNMB (P<0.05). Additionally, SCs treated with
50 nM rhGPNMB grew more rapidly compared with the other 2 groups
(10 and 100 nM), indicating that the proliferation of SCs increased
in a dose-dependent manner (P<0.05).
GPNMB accelerates the expression and
secretion of NTFs
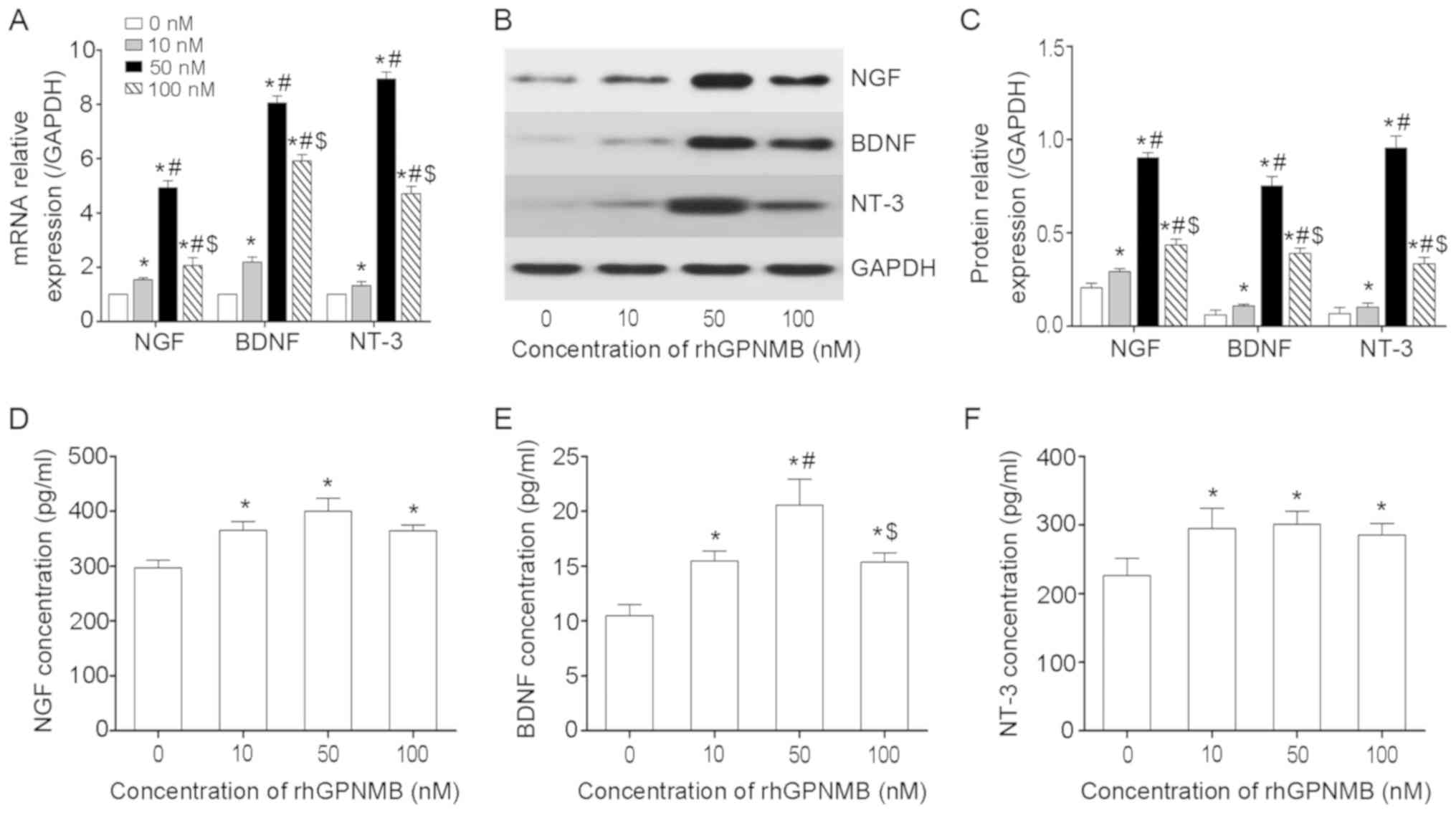
NTFs, including NGF, BDNF and NT-3, serve
significant roles in nerve regeneration. Following treatment with
various doses of rhGPNMB, RT-qPCR and WB were performed to measure
the expression of NGF, BDNF and NT-3 in SCs. The results determined
that SCs treated with rhGPNMB expressed increased levels of NGF,
BDNF and NT-3 compared with the control (P<0.05; Fig. 4A). Furthermore, the expression of
NGF was 4-5-fold higher in the 50 nM rhGPNMB treated group compared
with the control. It was also higher compared with the other two
rhGPNMB treated groups (P<0.05). In addition, the expression
levels of BDNF and NT-3 were significantly increased in the 50 nM
rhGPNMB-treated group compared with the other three groups
(P<0.05). These results indicated that the expression of NGF,
BDNF and NT-3 at the transcriptional level increased significantly
in a dose-dependent manner in SCs following treatment with rhGPNMB.
Similar results were also obtained via WB (Fig. 4B and C). The expression of NGF,
BDNF and NT-3 at the protein level was also significantly increased
in the 50 nM rhGPNMB-treated group compared with the other three
groups (P<0.05). Additionally, the expression of NGF, BDNF and
NT-3 at the protein level was increased in the 10 and 100 nM
rhGPNMB-treated groups compared with the control (P<0.05).
ELISAs were performed to measure the secretion
levels of NGF, BDNF and NT-3 in SCs. The results revealed that SCs
secreted an increased level of NGF, BDNF and NT-3 compared with the
control following treatment with rhGPNMB (P<0.05; Fig. 4D-F). However, there were no
significant differences in the secretions of NGF and NT-3 among the
three rhGPNMB-treated groups (P>0.05; Fig. 4D and F). The secretion of BDNF was
increased in the 50 nM rhGPNMB treated group compared with the
other rhGPNMB treated groups (P<0.05; Fig. 4E).
The aforementioned results indicated that GPNMB
accelerated the expression and secretion of NGF, BDNF and NT-3 in
SCs.
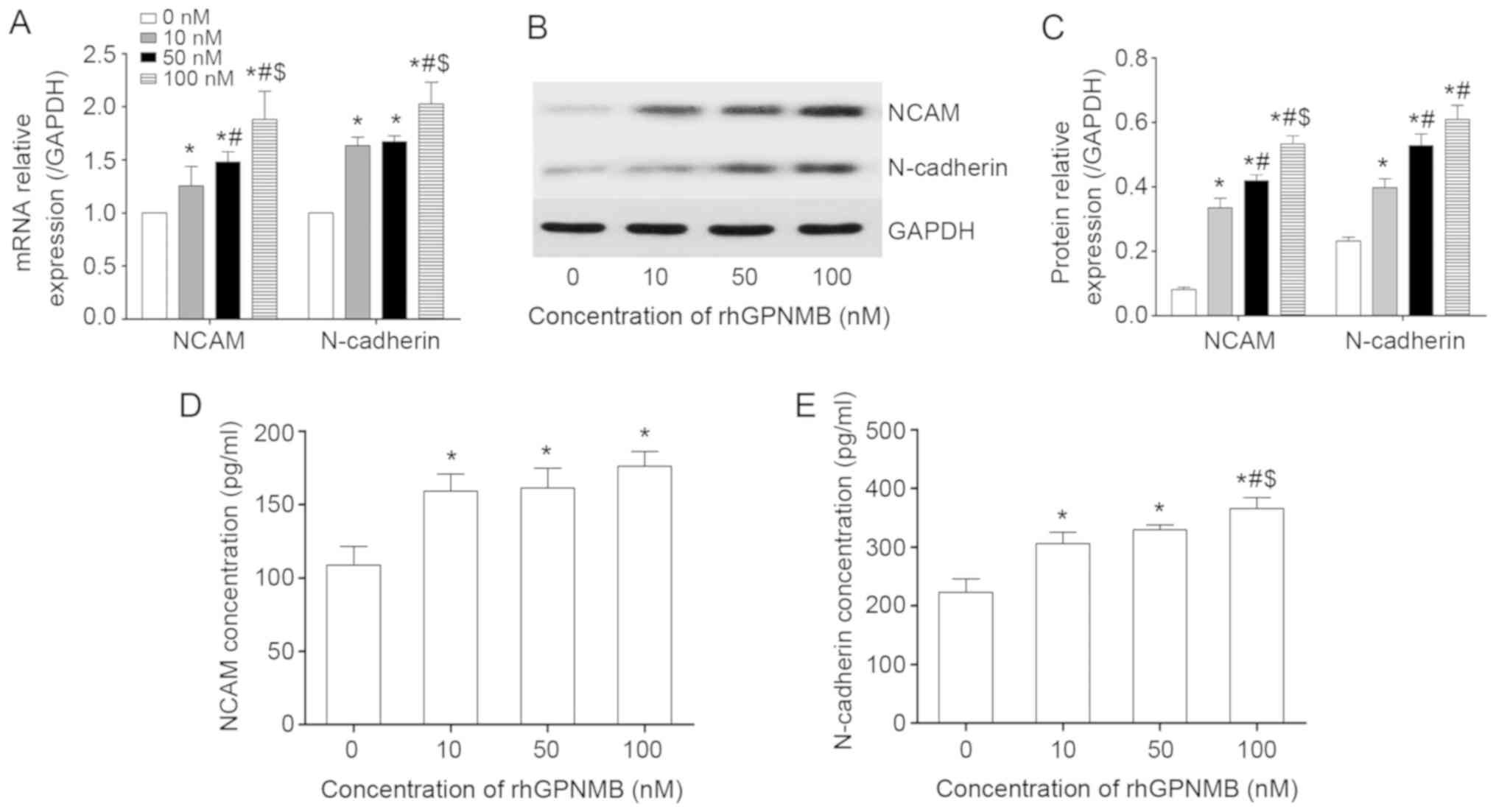
GPNMB triggers the expression and
secretion of NAMs
The expression and secretion of certain NAMs,
including NCAM and N-cadherin, were also detected by RT-qPCR, WB
and ELISA. These NAMs were selected as they can promote SC
migration during the nerve regeneration processes (21). The results revealed that the SCs
expressed increased levels of NCAM and N-cadherin in
rhGPNMB-treated groups compared with the control group at the
transcriptional level (P<0.05; Fig. 5A). The expression of NCAM in the
100 nM rhGPNMB-treated group was increased compared with the other
two rhGPNMB-treated groups (P<0.05), indicating that its
expression increased in a dose-dependent manner. However, there was
no significant difference in the expression of N-cadherin between
the 10 and 50 nM rhGPNMB treated groups (P>0.05). The results of
WB were consistent with those of RT-qPCR (Fig. 5B and C). It was revealed that the
expression of NCAM at the protein level increased in a
dose-dependent manner when SCs were treated with rhGPNMB.
Additionally, SCs expressed increased levels of N-cadherin in
rhGPNMB treated groups compared with the control. However, this was
not dose-dependent.
Using ELISAs, the present study obtained similar
results regarding the secretion of NCAM and N-cadherin in SCs
following treatment with rhGPNMB (Fig. 5D and E). It was also revealed that
SCs secreted increased levels of NCAM and N-cadherin when compared
with the control (P<0.05). However, there were no significant
differences in the secretion of NCAM and N-cadherin among the three
rhGPNMB-treated groups (P>0.05).
The results of the RT-qPCR, WB and ELISA analyses
revealed that GPNMB triggered the expression and secretion of NCAM
and N-cadherin in SCs.
Discussion
GPNMB, first cloned and described as a regulator
protein of tumor growth (22), is
ubiquitously expressed in the CNS (8) and exerts protective effects in
amyotrophic lateral sclerosis by regulating the amelioration of
skeletal muscle lesions, motor neuron cell death and PI3K/Akt and
MEK/ERK pathway activation (11,13,23,24). It also has been revealed that
GPNMB exerts neuroprotective effects against cerebral ischemia
reperfusion injury via ERK1/2 and Akt pathways (12). Recently, it has been reported that
the expression of GPNMB in SCs harvested from injured sciatic nerve
increases in vitro (14).
However, no reports have determined the expression of GPNMB in
vivo or have assessed the associated importance of PNS injuries
and diseases.
The present study used a sciatic nerve transection
model to investigate gene expression profiles in distal segments
via microarray analysis. The results of the STEM analysis revealed
that the expression of profile 41, which consisted of 76 mRNAs and
282 lncRNAs, increased, peaked and then gradually decreased at
serial time points post-injury. The results also demonstrated the
altered expression of GPNMB in vivo following injury, which
was different to previous research regarding GPNMB expression in
vitro (14). These results
may serve an important role in the study of PNR.
PNR is a complex biological process that involves
interactions among multiple cells, NTFs and extracellular matrix
proteins (25). During PNR, SCs
shed their myelin sheaths, dedifferentiate to a progenitor/stem
cell-like state, proliferate, migrate and produce a favorable
environment for axonal regeneration by helping to clear myelin
debris and by forming cellular conduits for the growth and
pathfinding of axons (26). In
the present study, the results of the STEM analysis revealed that
the expression of profile 41 was similar to that of the
proliferation tendency of distal acute denervated SCs (27). Profile 41 was therefore selected
for further bioinformatics analyses.
The results of the present study revealed that
profile 41 was associated with cell division, mitotic nuclear
division, DNA replication initiation, cell cycle, the p53 signaling
pathway and DNA replication. In addition, GPNMB was the most
significantly varied mRNA among the 76 included in profile 41.
These results were consistent with previous studies, which revealed
that the level of cellular p53 is associated with the activity of
GPNMB, also termed HGFIN, and the cell cycle, and that the p53
signaling pathway is involved in the regulation of GPNMB expression
and activity (28,29). Therefore, it was hypothesized that
GPNMB may be involved in cell proliferation following PNI from
bioinformatics analyses. The results of the present study also
revealed that GPNMB promoted denervated SC proliferation in
vitro, which confirmed these hypotheses.
GPNMB was selected for validation via RT-qPCR, WB
and IHC. The results indicated that the altered expression of GPNMB
at the transcriptional and protein levels were in concordance with
the microarray analysis. Furthermore, transverse section IHC
analysis revealed that GPNMB was expressed around nerve fibers,
indicating that GPNMB may be expressed in SCs. These results were
consistent with those of a previous study, in which the increased
expression of GPNMB in SCs following injury was observed in
vitro (14). However, the
time point at which GPNMB peaked in WB was different to that
observed following microarray analysis and RT-qPCR. The present
study hypothesized that this may due to protein translation from
mRNA and post-translational modification.
The microenvironment, consisting of NTFs and NAMs,
has been proven to promote nerve regeneration in vitro and
in vivo and serve neuroregenerative and neuroprotective
roles (30). Recently, it has
been demonstrated that NGF promotes the regeneration and functional
recovery of impaired peripheral nerves (31). Furthermore, it has also been
confirmed that BDNF (32), NT-3
(33) and N-cadherin (34) serve a vital role in promoting
nerve regeneration after injury. In the present study, denervated
SCs were harvested from injured nerves, as described in our
previous study (19) and were
identified to exhibit a long bipolar or tripolar elongated shape
with a small oval-shaped nucleus and expressed protein S100-β in
the membrane of the nucleus and cytoplasm. The results of the
present study revealed that denervated SCs expressed and secreted
increased levels of NGF, BDNF, NT-3, NCAM and N-cadherin following
rhGPNMB treatment in vitro, indicating that GPNMB may
promote PNR.
The expression profile of GPNMB observed in the
present study was consistent with other study by proteomics
published recently (14). The
altered expression tendency of GPNMB, which increased, peaked, and
then decreased was also similar to the proliferation of distal
acute denervated SCs (27). In
addition, the results revealed that GPNMB promoted the
proliferation of denervated SCs as well as the expression and
secretion of NTFs and NAMs in vitro. However, whether GPNMB
promotes the proliferation and secretion of SCs in vivo
remains unknown. Furthermore, there may be bias and statistical
error in the study due to the small sample size. Therefore, future
studies should assess the functions of GPNMB in vivo,
particularly on the proliferation and secretion of SCs, and
determine the underlying mechanisms involved in PNR.
The results of the present study revealed that GPNMB
was expressed and altered in the distal sciatic nerve following
transection in vivo and that GPNMB also promoted the
proliferation of denervated SCs as well as the expression and
secretion of NTFs and NAMs in vitro.
In conclusion, the present study demonstrated the
altered expression of GPNMB in distal sciatic nerve following
transection in vivo. It was also demonstrated that GPNMB
promoted the proliferation of denervated SCs as well as the
expression and secretion of NTFs and NAMs in vitro. The
results revealed that GPNMB may be used as a novel strategy for
PNR.
Supplementary Data
Acknowledgements
The authors would like to thank Professor Fang Liu
and Professor Haiyan Lin (Department of Anatomy, Second Military
Medical University, Shanghai, China) for helpful suggestions and
technical assistance.
Funding
This study was supported by the Funds of the
National Natural Science Foundation of China (grant no.
81271396).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ and XY designed the study. YZ and CH performed
the experiments. YZ and CH analyzed data. ZZ, XY, YZ and CH wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved and supervised
by the Animal Care and Use Committee of the Second Military Medical
University (permit no. SYXK-2002-042).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ray WZ and Mackinnon SE: Management of
nerve gaps: Autografts, allografts, nerve transfers, and
end-to-side neurorrhaphy. Exp Neurol. 223:77–85. 2010. View Article : Google Scholar :
|
|
2
|
Du J, Liu J, Yao S, Mao H, Peng J, Sun X,
Cao Z, Yang Y, Xiao B, Wang Y, et al: Prompt peripheral nerve
regeneration induced by a hierarchically aligned fibrin nanofiber
hydrogel. Acta Biomater. 55:296–309. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scheib J and Höke A: Advances in
peripheral nerve regeneration. Nat Rev Neurol. 9:668–676. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Allodi I, Udina E and Navarro X:
Specificity of peripheral nerve regeneration: Interactions at the
axon level. Prog Neurobiol. 98:16–37. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vargas ME and Barres BA: Why is Wallerian
degeneration in the CNS so slow? Annu Rev Neurosci. 30:153–179.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han B, Zhao JY, Wang WT, Li ZW, He AP and
Song XY: Cdc42 promotes schwann cell proliferation and migration
through Wnt/β-catenin and p38 MAPK signaling pathway after sciatic
nerve injury. Neurochem Res. 42:1317–1324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taya M and Hammes SR: Glycoprotein
non-metastatic melanoma protein B (GPNMB) and cancer: A novel
potential therapeutic target. Steroids. 133:102–107. 2018.
View Article : Google Scholar :
|
|
8
|
Huang JJ, Ma WJ and Yokoyama S: Expression
and immunolocalization of Gpnmb, a glioma-associated glycoprotein,
in normal and inflamed central nervous systems of adult rats. Brain
Behav. 2:85–96. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu X, Zhang P, Xu Z, Chen H and Xie X:
GPNMB enhances bone regeneration by promoting angiogenesis and
osteogenesis: Potential role for tissue engineering bone. J Cell
Biochem. 114:2729–2737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang YL, Hu YJ and Zhang FH: Effects of
GPNMB on proliferation and odontoblastic differentiation of human
dental pulp cells. Int J Clin Exp Pathol. 8:6498–6504.
2015.PubMed/NCBI
|
|
11
|
Ono Y, Tsuruma K, Takata M, Shimazawa M
and Hara H: Glycoprotein nonmetastatic melanoma protein B
extracellular fragment shows neuroprotective effects and activates
the PI3K/Akt and MEK/ERK pathways via the Na+/K+-ATPase. Sci Rep.
6:232412016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakano Y, Suzuki Y, Takagi T, Kitashoji A,
Ono Y, Tsuruma K, Yoshimura S, Shimazawa M, Iwama T and Hara H:
Glycoprotein nonmetastatic melanoma protein B (GPNMB) as a novel
neuroprotective factor in cerebral ischemia-reperfusion injury.
Neuroscience. 277:123–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nagahara Y, Shimazawa M, Ohuchi K, Ito J,
Takahashi H, Tsuruma K, Kakita A and Hara H: GPNMB ameliorates
mutant TDP-43-induced motor neuron cell death. J Neurosci Res.
95:1647–1665. 2017. View Article : Google Scholar
|
|
14
|
Shi GD, Cheng X, Zhou XH, Fan BY, Ren YM,
Lin W, Zhang XL, Liu S, Hao Y, Wei ZJ and Feng SQ: iTRAQ-based
proteomics profiling of Schwann cells before and after peripheral
nerve injury. Iran J Basic Med Sci. 21:832–841. 2018.PubMed/NCBI
|
|
15
|
Kang W, Sun T, Tang D, Zhou J and Feng Q:
Time-course transcriptome analysis of gingiva-derived mesenchymal
stem cells reveals that fusobacterium nucleatum triggers oncogene
expression in the process of cell differentiation. Front Cell Dev
Biol. 7:3592020. View Article : Google Scholar :
|
|
16
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
The Gene Ontology Consortium: The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47(D1): D330–D338. 2019. View Article : Google Scholar :
|
|
18
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47(D1): D590–D595. 2019. View Article : Google Scholar :
|
|
19
|
Zheng Y, Huang C, Liu F, Lin H, Niu Y,
Yang X and Zhang Z: Reactivation of denervated Schwann cells by
neurons induced from bone marrow-derived mesenchymal stem cells.
Brain Res Bull. 139:211–223. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). method Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Ren T, Yu S, Mao Z and Gao C: A
complementary density gradient of zwitterionic polymer brushes and
NCAM peptides for selectively controlling directional migration of
Schwann cells. Biomaterials. 56:58–67. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weterman MA, Ajubi N, van Dinter IM, Degen
WG, van Muijen GN, Ruitter DJ and Bloemers HP: nmb, a novel gene,
is expressed in low-metastatic human melanoma cell lines and
xenografts. Int J Cancer. 60:73–81. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tanaka H, Shimazawa M, Kimura M, Takata M,
Tsuruma K, Yamada M, Takahashi H, Hozumi I, Niwa J, Iguchi Y, et
al: The potential of GPNMB as novel neuroprotective factor in
amyotrophic lateral sclerosis. Sci Rep. 2:5732012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagahara Y, Shimazawa M, Tanaka H, Ono Y,
Noda Y, Ohuchi K, Tsuruma K, Katsuno M, Sobue G and Hara H:
Glycoprotein nonmetastatic melanoma protein B ameliorates skeletal
muscle lesions in a SOD1G93A mouse model of amyotrophic lateral
sclerosis. J Neurosci Res. 93:1552–1566. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gu X, Ding F, Yang Y and Liu J:
Construction of tissue engineered nerve grafts and their
application in peripheral nerve regeneration. Prog Neurobiol.
93:204–230. 2011. View Article : Google Scholar
|
|
26
|
Parrinello S, Napoli I, Ribeiro S,
Wingfield Digby P, Fedorova M, Parkinson DB, Doddrell RD, Nakayama
M, Adams RH and Lloyd AC: EphB signaling directs peripheral nerve
regeneration through Sox2-dependent Schwann cell sorting. Cell.
143:145–155. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hall SM: The biology of chronically
denervated Schwann cells. Ann N Y Acad Sci. 883:215–233. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Metz RL, Yehia G, Fernandes H, Donnelly RJ
and Rameshwar P: Cloning and characterization of the 5′ flanking
region of the HGFIN gene indicate a cooperative role among p53 and
cytokine-mediated transcription factors: Relevance to cell cycle
regulation. Cell Cycle. 4:315–322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Metz RL, Patel PS, Hameed M, Bryan M and
Rameshwar P: Role of human HGFIN/nmb in breast cancer. Breast
Cancer Res. 9:R582007. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu QG, Midha R, Martinez JA, Guo GF and
Zochodne DW: Facilitated sprouting in a peripheral nerve injury.
Neuroscience. 152:877–887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li R, Li Y, Wu Y, Zhao Y, Chen H, Yuan Y,
Xu K, Zhang H, Lu Y, Wang J, et al: Heparin-poloxamer
thermosensitive hydrogel loaded with bFGF and NGF enhances
peripheral nerve regeneration in diabetic rats. Biomaterials.
168:24–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hei WH, Almansoori AA, Sung MA, Ju KW, Seo
N, Lee SH, Kim BJ, Kim SM, Jahng JW, He H and Lee JH: Adenovirus
vector-mediated ex vivo gene transfer of brain-derived neurotrophic
factor (BDNF) tohuman umbilical cord blood-derived mesenchymal stem
cells (UCB-MSCs) promotescrush-injured rat sciatic nerve
regeneration. Neurosci Lett. 643:111–120. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sahenk Z, Nagaraja HN, McCracken BS, King
WM, Freimer ML, Cedarbaum JM and Mendell JR: NT-3 promotes nerve
regeneration and sensory improvement in CMT1A mouse models and in
patients. Neurology. 65:681–689. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Clements MP, Byrne E, Camarillo Guerrero
LF, Cattin AL, Zakka L, Ashraf A, Burden JJ, Khadayate S, Lloyd AC,
Marguerat S and Parrinello S: The wound microenvironment reprograms
schwann cells to invasive mesenchymal-like cells to drive
peripheral nerve regeneration. Neuron. 96:98–114.e7. 2017.
View Article : Google Scholar : PubMed/NCBI
|