Introduction
Idiopathic pulmonary fibrosis (IPF) is a chronic,
progressive, fibrotic interstitial pulmonary disease of unknown
cause, characterized primarily by the excessive deposition of
extracellular matrix (ECM) proteins by activated lung fibroblasts
and myofibroblasts, leading to decreased gas exchange and impaired
pulmonary function (1,2). IPF is one of the most common
idiopathic interstitial pneumonia and the most common interstitial
pulmonary disease, with an estimated incidence of 50/100,000
worldwide every year, with a median survival time after diagnosis
of 3-5 years (3,4).
IPF commonly occurs in older adults, and the median
survival rate of patients with IPF after diagnosis is very poor
(5,6). Previous studies on IPF reported that
some anti-fibrinolytic components, glucocorticoids, antioxidants
and other therapeutic strategies are effective in the laboratory.
However, certain drugs, including glucocorticoids and
immunosuppressants, have serious adverse effects and do not improve
the prognosis of patients. At present, there is no universally
accepted treatment for IPF. Although the new anti-fibrotic drugs
pirfenidone and nintedanib can slow down IPF progression, their
effectiveness and long-term safety remain controversial (7). It is therefore crucial to determine
potential targets in IPF and develop novel treatments for
patients.
Although many traditional Chinese herbs (TCH) have
interesting therapeutic effects in various types of disease
(8,9), only a few TCHs have been reported to
have therapeutic effects in IPF. The potential roles of certain
TCHs in the regulation of IPF remain unexplored. Astragalus
is a plant that has been used in traditional Chinese medicine for
centuries, which is commonly used to treat certain diseases.
Numerous studies reported that Astragalus could have some
immunoregulatory and antitumor effects (10-13). Recent studies demonstrated that
Astragalus polysaccharides (APS), which are the major
components of Astragalus, also have some antitumor and
anti-fibrotic effects (11,14). However, no study evaluated the
role of APS in PF. The present study aimed to explore the potential
role and underlying mechanism of APS in IPF. To do so, a bleomycin
(BLM)-induced PF mouse model and in vitro model of
transforming growth factor β1(TGF-β1)-stimulated human lung
epithelial cells were established. Considering the crucial role of
NF-κB signaling and TGF-β1 activity in PF, the effect of APS on
TGF-β1-induced NF-κB signaling was also assessed. The results from
the present study may provide novel insights into the underlying
mechanism of APS in PF and allow the development of novel
therapeutic options for patients with IPF.
Materials and methods
Animal model and cell culture
The present study was approved by the Institutional
Animal Care and Use Committee of Zhejiang University, Hangzhou,
China. A total of 30 male C57BL/6 mice (6-8 weeks old) purchased
form Vital River Laboratory Animal Technology were used in this
study. Mice were anesthetized by intraperitoneal injection of 0.1
ml of sodium pentobarbital at the dose of 50 mg/kg and received 5
mg/kg BLM (Nippon Kayaku Co., Ltd.) or an equal volume of sterile
saline by intratracheal administration.
APS was obtained from Shanghai Yuanye Bio-Technology
Co., Ltd. and was prepared as described previously (15,16). Mice were treated with either
saline or APS (200 mg/kg/day) daily through gavage seven days prior
to BLM administration. The day following BLM administration, mice
received 200 mg/kg APS daily by gavage until the end of experiment
(15). On days 28, six mice per
group were anesthetized with sodium pentobarbital (75 mg/kg) prior
to cervical dislocation, and lung tissues were collected and
immediately frozen in liquid nitrogen for further experiments.
The primary human bronchial epithelial cells and the
human lung epithelial cell line A549 were obtained from the Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences.
Cells were cultured in F-12K Medium supplemented with 10% fetal
bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin
(all from Gibco; Thermo Fisher Scientific, Inc.) and placed at 37°C
in a humidified incubator containing 5% CO2. A549 cells
was treated with TGF-β1 (10 ng/ml; PeproTech) or 50 µmol/l
of the NF-κB inhibitor pyrrolidine dithiocarbamate (PDTC; cat. no.
P8765; Sigma-Aldrich; Merck KGaA) for 24 h. Cells were cultured in
medium supplemented with 50 mg/ml APS (Shanghai Yuanye
Bio-Technology Co., Ltd.; >98% purity) for 3 days (APS group)
(17).
Hematoxylin and eosin (H&E) staining
and Masson's trichrome staining
Lung tissues from mice were fixed in 4%
paraformaldehyde for 24 h at 4°C and embedded in paraffin. Sections
(4-mm thick) were prepared and subsequently stained with H&E
and Masson's trichrome for histopathological examination. Images
were acquired by an experienced pathologist with an Olympus
microscope (magnification, ×40). The fibrotic area was quantified
using Image Pro Plus 6.0 (Media Cybernetics, Inc.).
Measurement of hydroxyproline (HYP)
content
The content in matrix proteins in lung tissues was
determined by using the Hydroxyproline detection kit (cat. no.
A030-3; Nanjing Jiancheng Bioengineering Institute) according to
the manufacturer's instructions. The lung tissues were
prepared for hydrolysis, adjusting the PH value to 6.0-6.8. Double
distilled water was added and tissues were incubated at room
temperature for 20 min. Subsequently, the developing solution was
added to the tissues that were incubated at 37°C for 5 min.
Absorbance was read at 550 nm using a microplate reader. Data were
expressed as micrograms (µg) of HYP per mg of dry lung
tissue.
Western blotting, reverse
transcription-quantitative (RT-q) PCR and immunofluorescence
(IF)
Western blotting, RT-qPCR and IF were performed as
described previously (18-20).
The primary antibodies for western blotting and IF are provided in
Table SI.
Western blotting was performed as described
previously (18,19). Proteins were extracted from lung
tissues and A549 cells using RIPA lysis buffer at 4°C. Proteins (10
µg) proteins were separated by 10% SDS-PAGE and transferred
onto polyvinylidene fluoride membranes. Membranes were blocked with
3% BSA in TBST at room temperature for 60 min. Membranes were
incubated with primary antibodies overnight at 4°C and with
secondary antibody for 1 h at room temperature. β-actin was used as
a control and Lamin B1 was used as a nuclear control. Signals were
visualized by using an enhanced chemiluminescence detection kit
(Auragene Technology, Co., Inc.). Densitometric analysis was
performed using Image Pro Plus v 6.0 (Media Cybernetics, Inc.).
RT-qPCR was performed as described previously
(18,19). Total RNA was extracted from mice
lung samples with TRIzol reagent. The sequences of the primers were
as follows: Collagen-1a1, forward 5′-GCC CGA ACC CCA AGG AAA AGA
AGC-3′, reverse 5′-CTG GGA GGC CTC GGT GGA CAT TAG-3′;
collagen-3a1, forward 5′-GCC CAC AGC CTT CTA CAC CT-3′, reverse
5′-GCC AGG GTC ACC ATT TCT C-3′; fibronectin, forward 5′-GAT GTC
CGA ACA GCT ATT TACC A-3′, reverse 5′-CCT TGC GAC TTC AGC CAC T-3′;
and GAPDH, forward 5′-TGC AC CAC CAA CTG CTT AG C-3′ and reverse
5′-GGC ATG GAC TGT GGT CAT GAG-3′. The relative expression levels
were normalized to endogenous control GAPDH and were expressed as
2−ΔΔCq (21) using ABI
software v2.3 (Applied Biosystems).
IF was performed as described previously (18,20). Briefly, lung tissues and A549
cells were fixed, permeabilized and incubated for 1 h at 37°C with
primary antibody. The appropriate Alexa Fluor linked secondary
antibody was applied for 30 min at room temperature. Cells were
counterstained with DAPI and images were acquired with a
fluorescence microscope.
Immunohistochemistry (IHC)
IHC and subsequent analyses were performed as
described previously (19,20).
The primary antibodies used for IHC are presented in Table SI. Briefly, formalin-fixed and
paraffin embedded mice lung samples were cut into 5-µm-thick
sections. Sections were incubated with the primary antibodies at
4°C overnight. Images were acquired on an Olympus microscope. The
EnVision kit (Dako; Agilent Technologies, Inc.) was used to detect
the signal.
Ethynyl deoxyuridine (EdU) assay
A Click-iT Ethynyl deoxyuridine Imaging Kit (cat.
no. C10086; Invitrogen; Thermo Fisher Scientific, Inc.) was used
for EdU labelling to assess A549 cell proliferation as previously
described (18).
Wound healing assay
The wound healing assay was performed as described
previously (19). A549 cells were
cultured in 6-well plates until they reach 100% confluence. The
wound was made with a 200 µl sterile pipet tip and 1% FBS
was added to the medium.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software version 5.0 (GraphPad Software, Inc.) and SPSS
version 19.0 (IBM Corp.). Unpaired Student's t-test was used for
comparison between two groups. One-way ANOVA followed by
Student-Newman-Keuls post hoc test was used for comparison between
≥3 groups. The data were presented as the means ± standard
deviation of three independent experiments. P<0.05 was
considered to indicate a statistically significant difference.
Results
APS reduces BLM-induced PF in mice
To evaluate the effect of APS on PF, a BLM-induced
PF mouse model was established. The establishment of the PF model
was verified by H&E staining and Masson's trichrome assay, and
the results confirmed the collagen deposition (Fig. S1A) and the increase in fibrotic
area (Fig. S1B). Furthermore,
the results from RT-qPCR demonstrated that the expression level of
collagen-1a1, collagen-3a1 and fibronectin was increased in lung
tissues from BLM-treated mice compared with lung tissues from the
saline control group mice (Fig.
S1C-E). Furthermore, results from IHC and IF demonstrated an
increase in the fibrosis-related proteins collagen and fibronectin
in the PF mouse model (Fig. S1F and
G). These results confirmed that the BLM-induced PF model was
successfully established.
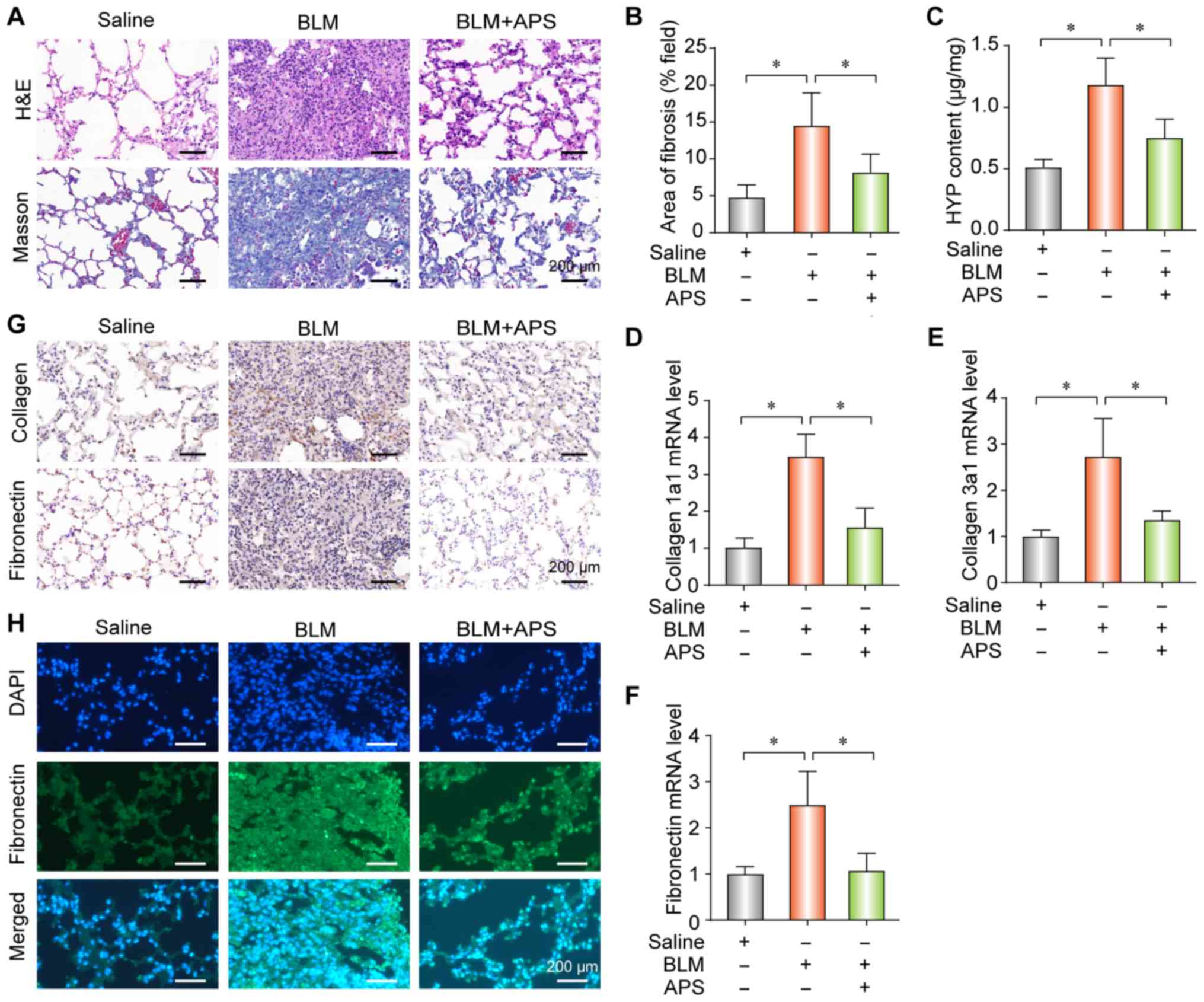
To explore the potential therapeutic effects of APS
in PF, APS was administered to the BLM-induced PF mouse model by
gavage. The results demonstrated that APS reduced collagen
deposition (Fig. 1A), fibrotic
area (Fig. 1B) and HYP content
(Fig. 1C), which all together are
indicators of PF. Furthermore, APS alleviated the BLM-induced
up-regulation of collagen-1a1 (Fig.
1D), collagen-3a1 (Fig. 1E)
and fibronectin (Fig. 1F) at the
mRNA level. In addition, the results from IHC and IF demonstrated
that APS inhibited the BLM-induced upregulation of collagen and
fibronectin (Fig. 1G and 1H, respectively). These findings
suggested that APS may attenuate BLM-induced PF.
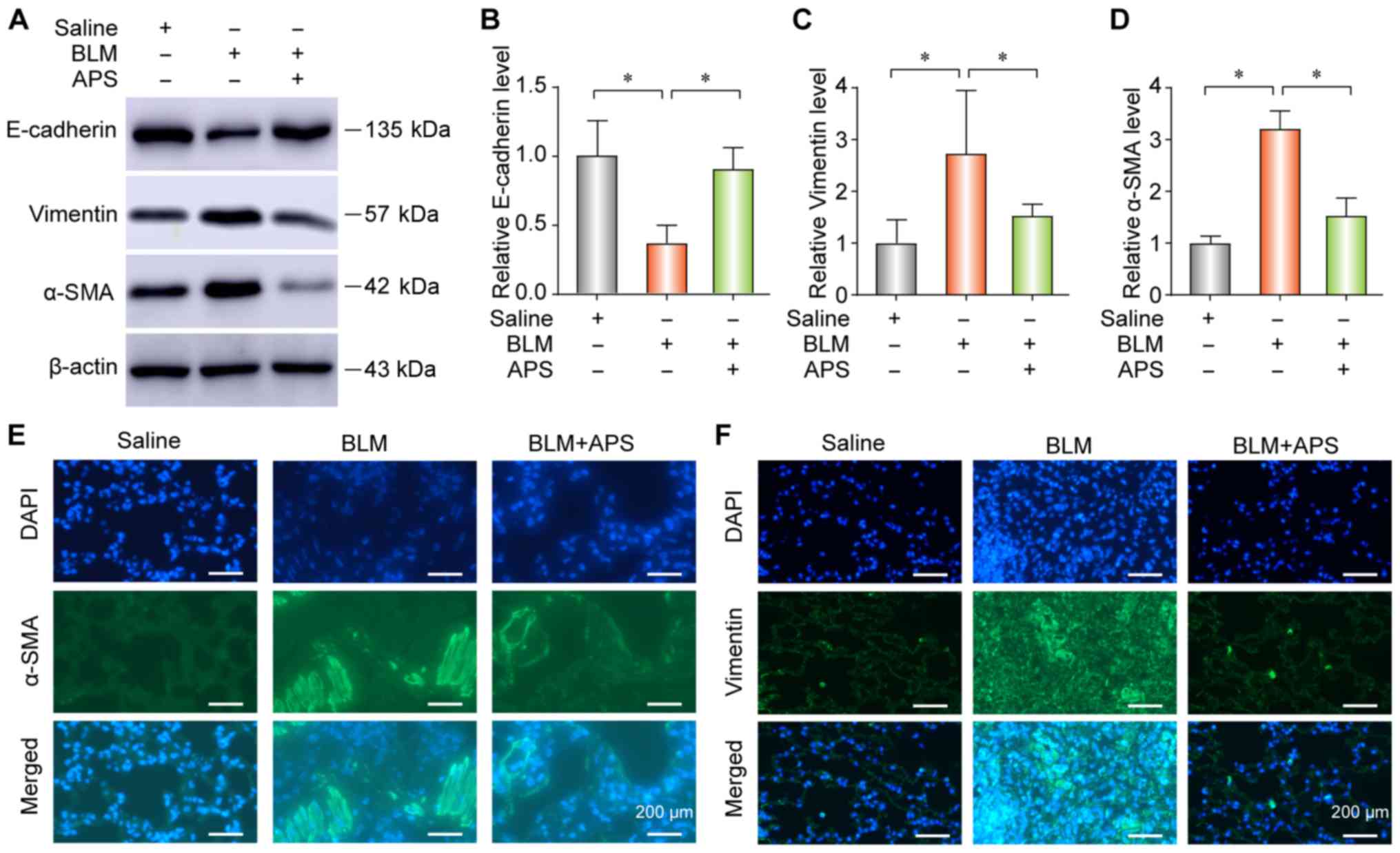
APS inhibits BLM-induced EMT in mice with
PF
To explore the occurrence of EMT during PF, the
expression of the epithelial marker E-cadherin and of the
mesenchymal markers vimentin and alpha smooth muscle actin (α-SMA)
were determined by western blotting or IF. The results from western
blotting (Fig. 2A-D) and IF
(Fig. 2E and F) demonstrated that
BLM treatment significantly reduced E-cadherin expression and
upregulated the expression of vimentin and α-SMA in lung tissues.
These data suggested that epithelial cells decreased and
myofibroblasts increased during PF. However, the results from
western blotting and IF demonstrated that treatment with APS could
reverse BLM-induced EMT, along with a decrease in vimentin and
α-SMA expression and an increase in the E-cadherin expression
(Fig. 2A-D, E and F).
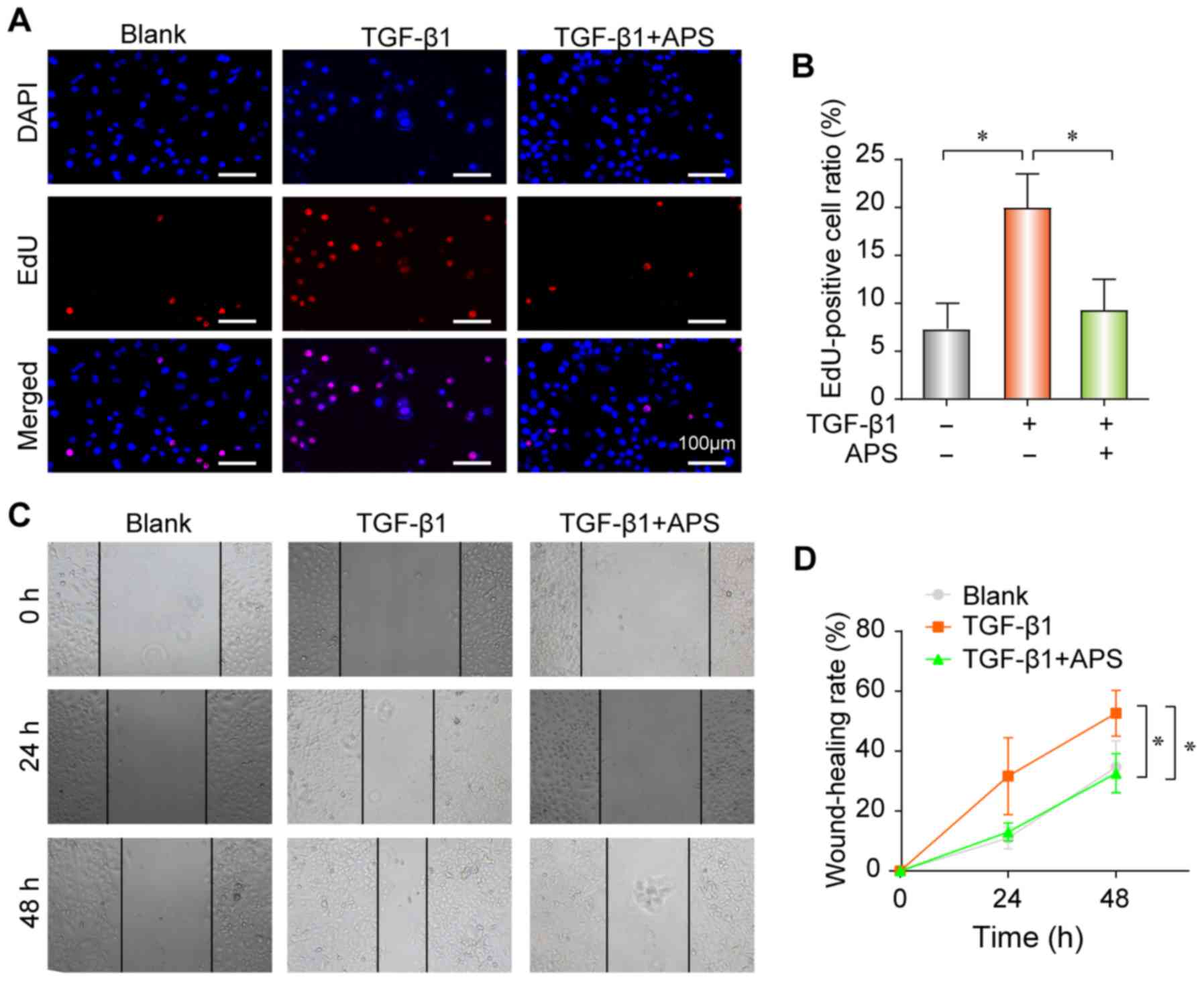
APS decreases TGF-β1-induced fibrogenesis
in pulmonary epithelial cells
The proliferation and differentiation of fibroblasts
serve crucial roles in PF, and can promote extracellular matrix
(ECM) deposition and aggravate therefore fibrosis. The results from
the present study demonstrated that TGF-β1 promoted the
proliferation (Fig. 3A and B) and
migratory ability (Fig. 3C and D)
of pulmonary epithelial cells in vitro, whereas these
effects were nearly completely reversed following treatment with
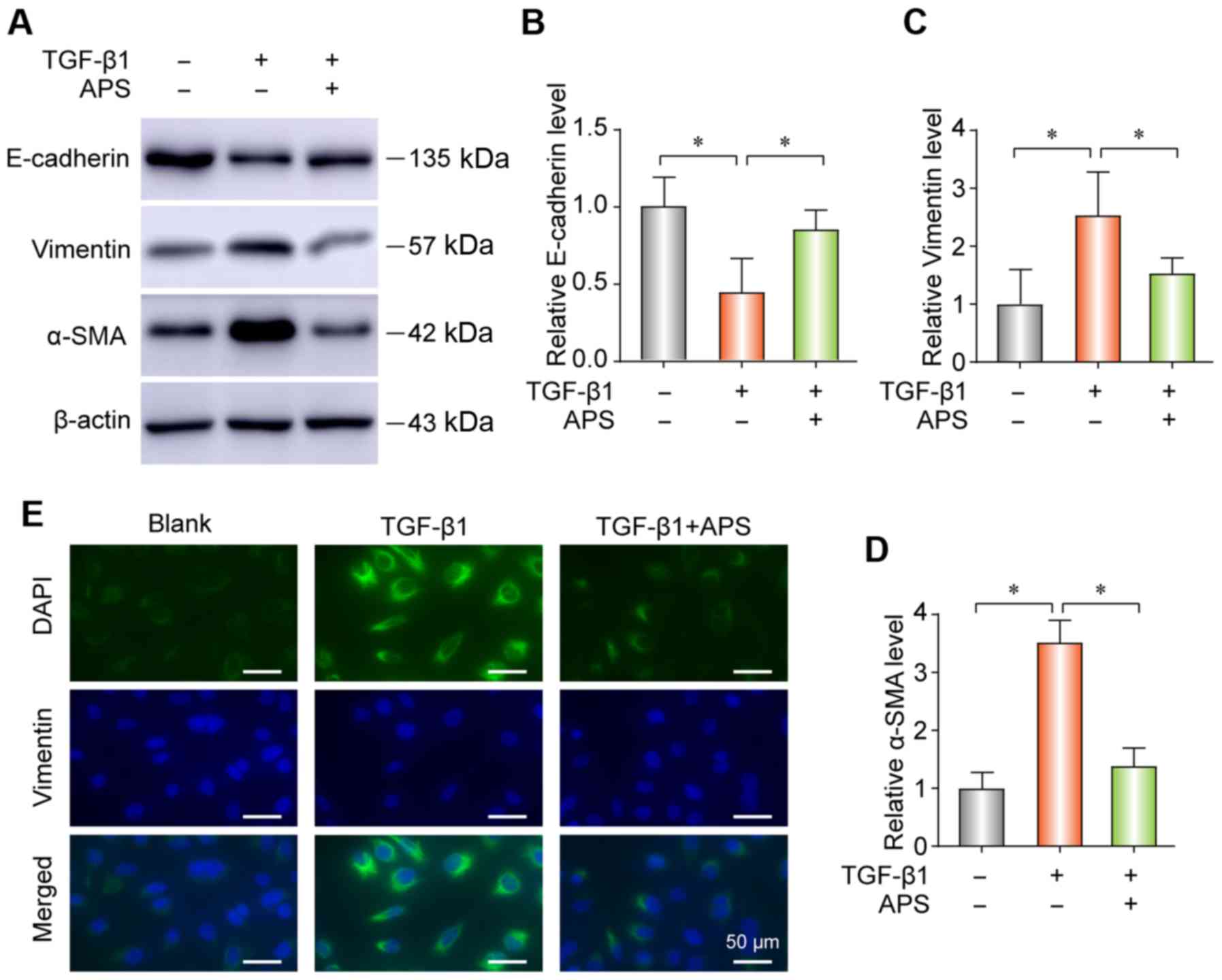
APS. EMT is commonly known to be induced by TGF-β1 (22,23). The results from western blotting
indicated that TGF-β1 could upregulate the expression of α-SMA and
vimentin and reduce the expression of E-cadherin in A549 cells
(Fig. 4A-D). However, cell
treatment with APS reversed these changes. The results from IF
demonstrated that TGF-β1 could increase vimentin expression in A549
cells (Fig. 4E).
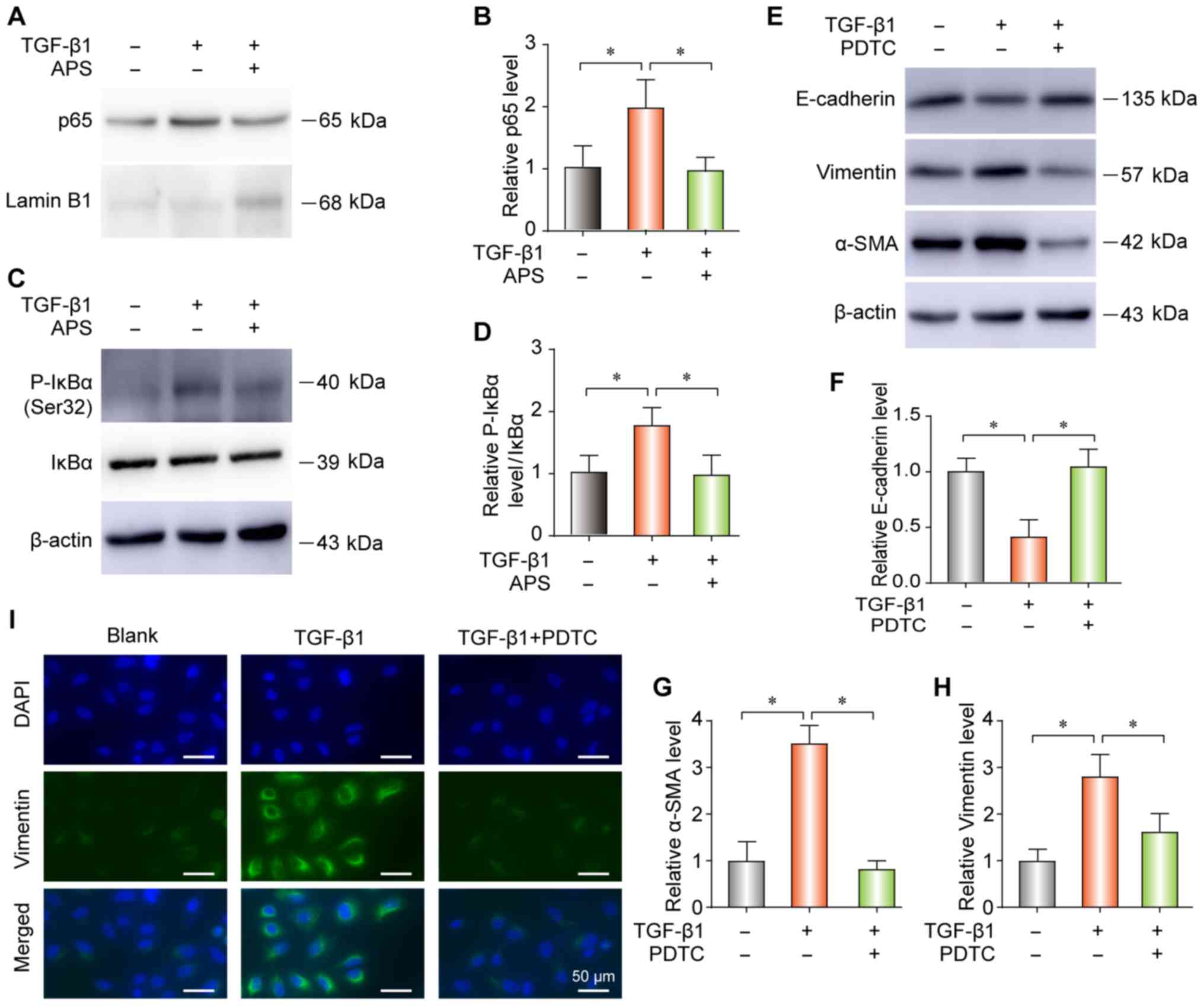
APS inhibits TGF-β1-induced NF-κB pathway
activation in pulmonary epithelial cells
Considering the essential role of TGF-β1 activity
and NF-κB signaling in PF, the effect of APS on TGF-β1-induced
NF-κB signaling was evaluated in the present study. Cell treatment
with TGF-β1 increased the nuclear translocation of p65 (Fig. 5A and B) and the expression level
of phospho (p-) IκBα (Fig. 5C and
D). Furthermore, APS blocked the NF-κB pathway activation by
TGF-β1 (Fig. 5A-D). In addition,
PDTC (24) neutralized the
TGF-β1-induced increase in E-Cadherin, vimentin and α-SMA
expression by western blot (Fig.
5E-H) or IF (Fig. 5I).
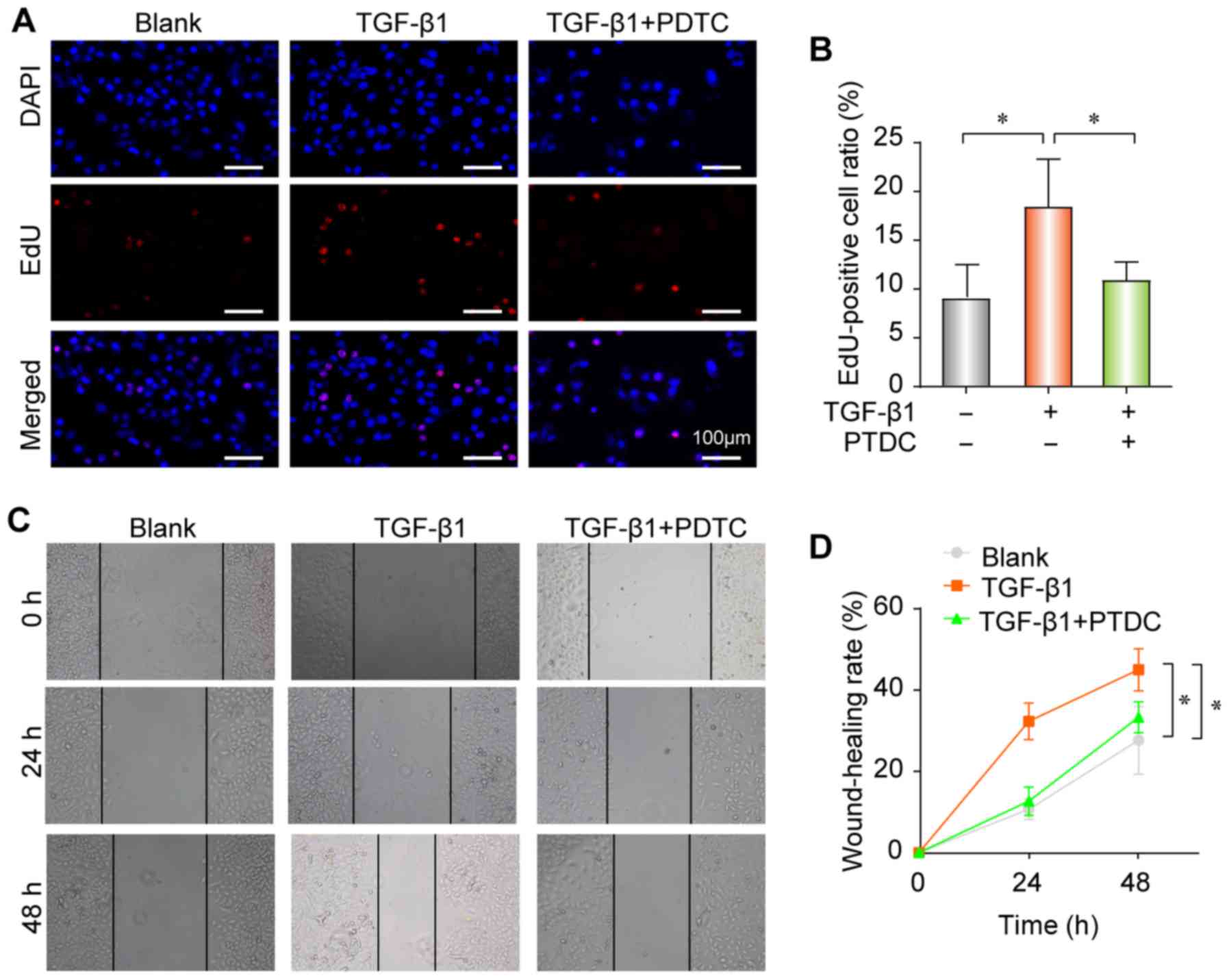
Furthermore, TGF-β1 increased the proliferation (Fig. 6A and B) and migratory ability
(Fig. 6C and D) of pulmonary
epithelial cells; however, these effects were reversed following
treatment with PDTC. Taken together, these findings indicate that
the NF-κB signaling pathway may serve a crucial role in
TGF-β1-induced EMT in A549 cells, and that TGF-β1-induced PF
progression may be inhibited by APS via the NF-κB pathway.
Discussion
EMT serves a key role in the occurrence and
development of PF. During EMT, the expression in epithelial cells
of adherens junction proteins, including E-cadherin, is decreased
whereas the expression of fibroblast markers, including vimentin
and α-SMA, is increased (25,26). One of the histological
characteristics of PF is the ECM deposition. The cytokine TGF-β1 is
an important regulator of fibrogenesis. TGF-β1 activates
fibroblasts and transforms them into myofibroblasts (2,3).
Numerous studies reported that TGF-β1 is a key fibrogenic factor,
which can induce EMT in PF (27).
Subsequently, inhibition of EMT through the TGF-β1 pathway may be
considered as a potential therapeutic option for IPF.
Previous studies reported the antitumor (28), anti-diabetic (29), antioxidant and immunomodulatory
properties of APS (30,31). Several studies demonstrated that
Astragalus has strong anti-fibrotic activities (12,15). Astragalus was found to
inhibit the EMT of peritoneal mesothelial cells by decreasing
β-catenin expression (32).
Furthermore, it was reported that Astragalus has some
therapeutic effects on BLM-induced PF by reducing Jagged1/Notch1
expression (33). Furthermore,
Astragalus saponins could reduce liver fibrosis by
decreasing TGF-β1/Smad signaling pathway (34). Astragaloside IV isolated from
Astragalus can reduce the hyperphosphorylation of forkhead
box O3a induced by TGF-β1/PI3K/Akt and reverse EMT involved in the
process of fibrosis (35). In the
present study, APS attenuated BLM-induced PF and EMT in mice. These
results were similar to those from a previous study demonstrating
that APS can inhibit excessive collagen accumulation in a
BLM-induced scleroderma mouse model (36).
Toll-like receptor 4 (TLR4)/NF-κB pathway activation
is a key mechanism involved in the inflammatory response observed
in pulmonary disease (37,38).
NF-κB signaling is activated by TLR4 via the stimulation of
released inflammatory cytokines, which promote the proinflammatory
response. Under normal conditions, NF-κB is isolated in the
cytoplasm and binds to IκB. Once activated, NF-κB translocates to
the nucleus and regulates the production of inflammatory cytokines
(39,40).
In the present study, treatment with PDTC alleviated
PF in BLM-treated mice. These findings suggested that NF-κB may
serve an important role in the progression of PF, which may be
reversed by inhibiting NF-κB signaling. A previous study reported
that APS reduces inflammation by inhibiting the activation of the
TLR-4/NF-κB pathway and protects mice from coxsackievirus
B3-induced viral myocarditis (15). In the present study, APS
attenuated BLM-induced PF and EMT in mice. Furthermore, APS reduced
the TGF-β1-induced EMT and NF-κB pathway activation in pulmonary
epithelial cells. Taken together, these findings suggested that APS
may have an anti-fibrotic effect by repressing the activation of
the NF-κB signaling pathway, preventing therefore EMT progression
and ameliorating PF.
In conclusion, the present study demonstrated that
APS could alleviate PF in vitro and in vivo and
reduce the TGF-β1-induced NF-κB pathway activation. To the best of
our knowledge, this study was the first to demonstrate the
APS-mediated alleviation of PF in vitro and in vivo.
In addition, this study elucidated some mechanism of action of APS
in PF, suggesting that APS may be considered as a therapeutic agent
to treat IPF.
Supplementary Data
Funding
This study was supported by the Zhejiang Provincial
Natural Science Foundation of China (grant nos. LY19H160057 to SL,
LY19H030013 to RM and LY15H030011 to LZ.), the Zhejiang Provincial
Public Welfare Technology Research Plan of China (grant no.
LGD19H030003 to XA) and the Chinese Medicine Science and Technology
Plan of Zhejiang Province (grant no. 2019ZA070 to XA).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
SL, XS and XA designed the study. RZ performed all
experiments. LX analyzed the data. SL and XS wrote the article. All
authors discussed the results and revised the article. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by The Institutional
Research Board of the First Affiliated Hospital, College of
Medicine, Zhejiang University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Lederer DJ and Martinez FJ: Idiopathic
pulmonary fibrosis. N Engl J Med. 378:797–798. 2018. View Article : Google Scholar
|
|
2
|
Richeldi L, Collard HR and Jones MG:
Idiopathic pulmonary fibrosis. Lancet. 389:1941–1952. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hutchinson J, Fogarty A, Hubbard R and
McKeever T: Global incidence and mortality of idiopathic pulmonary
fibrosis: A systematic review. Eur Respir J. 46:795–806. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wolters PJ, Collard HR and Jones KD:
Pathogenesis of idiopathic pulmonary fibrosis. Annu Rev Pathol.
9:157–179. 2014. View Article : Google Scholar :
|
|
5
|
Raghu G: Idiopathic pulmonary fibrosis:
Combating on a new turf. Lancet Respir Med. 4:430–432. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu YM, Nepali K and Liou JP: Idiopathic
pulmonary fibrosis: Current status, recent progress and emerging
targets. J Med Chem. 60:527–553. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vancheri C, Kreuter M, Richeldi L, Ryerson
CJ, Valeyre D, Grutters JC, Wiebe S, Stansen W, Quaresma M,
Stowasser S, et al: Nintedanib with add-on pirfenidone in
idiopathic pulmonary fibrosis. Results of the INJOURNEY trial. Am J
Respir Crit Care Med. 197:356–363. 2018. View Article : Google Scholar
|
|
8
|
Wang R, Sun Q, Wang F, Liu Y, Li X, Chen
T, Wu X, Tang H, Zhou M, Zhang S, et al: Efficacy and safety of
chinese herbal medicine on ovarian cancer after reduction surgery
and adjuvant chemotherapy: A systematic review and meta-analysis.
Front Oncol. 9:7302019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhai B, Zhang N, Han X, Li Q, Zhang M,
Chen X, Li G, Zhang R, Chen P, Wang W, et al: Molecular targets of
β-elemene, a herbal extract used in traditional Chinese medicine,
and its potential role in cancer therapy: A review. Biomed
Pharmacother. 114:1088122019. View Article : Google Scholar
|
|
10
|
Lin S, An X, Guo Y, Gu J, Xie T, Wu Q and
Sui X: Meta-analysis of Astragalus-containing traditional chinese
medicine combined with chemotherapy for colorectal cancer: Efficacy
and safety to tumor response. Front Oncol. 9:7492019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu C, Li H, Wang K, Zhuang J, Chu F, Gao
C, Liu L, Feng F, Zhou C, Zhang W and Sun C: Identifying the
antiproliferative effect of Astragalus polysaccharides on breast
cancer: Coupling network pharmacology with targetable screening
from the cancer genome atlas. Front Oncol. 9:3682019. View Article : Google Scholar :
|
|
12
|
Huang KC, Su YC, Sun MF and Huang ST:
Chinese herbal medicine improves the long-term survival rate of
patients with chronic kidney disease in taiwan: A nationwide
retrospective population-based cohort study. Front Pharmacol.
9:11172018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen X, Chen X, Gao J, Yang H, Duan Y,
Feng Y, He X, Gong X, Wang H, Wu X and Chang J: Astragaloside III
enhances anti-tumor response of NK cells by elevating NKG2D and
IFN-γ. Front Pharmacol. 10:8982019. View Article : Google Scholar
|
|
14
|
Ren L, Guo XY, Gao F, Jin ML and Song XN:
Identification of the perturbed metabolic pathways associating with
renal fibrosis and evaluating metabolome changes of pretreatment
with astragalus polysaccharide through liquid chromatography
quadrupole time-of-flight mass spectrometry. Front Pharmacol.
10:16232020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu T, Zhang M, Niu H, Liu J, Ruilian M,
Wang Y, Xiao Y, Xiao Z, Sun J, Dong Y and Liu X: Astragalus
polysaccharide from Astragalus Melittin ameliorates inflammation
via suppressing the activation of TLR-4/NF-κB p65 signal pathway
and protects mice from CVB3-induced virus myocarditis. Int J Biol
Macromol. 126:179–186. 2019. View Article : Google Scholar
|
|
16
|
Li XT, Zhang YK, Kuang HX, Jin FX, Liu DW,
Gao MB, Liu Z and Xin XJ: Mitochondrial protection and anti-aging
activity of Astragalus polysaccharides and their potential
mechanism. Int J Mol Sci. 13:1747–1761. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang L, Luo Y, Lu Z, He J, Wang L, Zhang
L, Zhang Y and Liu Y: Astragalus polysaccharide inhibits ionizing
radiation-induced bystander effects by regulating MAPK/NF-κB
signaling pathway in bone mesenchymal stem cells (BMSCs). Med Sci
Monit. 24:4649–4658. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin S, Zhang R, An X, Li Z, Fang C, Pan B,
Chen W, Xu G and Han W: LncRNA HOXA-AS3 confers cisplatin
resistance by interacting with HOXA3 in non-small-cell lung
carcinoma cells. Oncogenesis. 8:602019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin S, Zhou S, Jiang S, Liu X, Wang Y,
Zheng X, Zhou H, Li X and Cai X: NEK2 regulates stem-like
properties and predicts poor prognosis in hepatocellular carcinoma.
Oncol Rep. 36:853–862. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhai S, Lin S, Lin Z, Xu J, Ji T, Chen K,
Wu K, Liu H, Ying H, Fei W, et al: eIF4EBP3 was downregulated by
methylation and acted as a tumor suppressor by targeting
eIF4E/β-catenin in gastric cancer. Gastric Cancer. 2019.Epub ahead
of print.
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Zheng Q, Tong M, Ou B, Liu C, Hu C and
Yang Y: Isorhamnetin protects against bleomycin-induced pulmonary
fibrosis by inhibiting endoplasmic reticulum stress and
epithelial-mesenchymal transition. Int J Mol Med. 43:117–126.
2019.
|
|
23
|
Zhu X, Li Q, Hu G, Wang J, Hu Q, Liu Z, Wu
G and Zhong Y: BMS-345541 inhibits airway inflammation and
epithelial-mesenchymal transition in airway remodeling of asthmatic
mice. Int J Mol Med. 42:1998–2008. 2018.PubMed/NCBI
|
|
24
|
Liu F, Zhang X, Zhang B, Mao W, Liu T, Sun
M and Wu Y: TREM1: A positive regulator for inflammatory response
via NF-κB pathway in A549 cells infected with mycoplasma
pneu-moniae. Biomed Pharmacother. 107:1466–1472. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin HY, Liang YK, Dou XW, Chen CF, Wei XL,
De Zeng, Bai JW, Guo YX, Lin FF, Huang WH, et al: Notch3 inhibits
epithelial-mesenchymal transition in breast cancer via a novel
mechanism, upregulation of GATA-3 expression. Oncogenesis.
7:592018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li N, Babaei-Jadidi R, Lorenzi F,
Spencer-Dene B, Clarke P, Domingo E, Tulchinsky E, Vries RGJ, Kerr
D, Pan Y, et al: An FBXW7-ZEB2 axis links EMT and tumour
microenvironment to promote colorectal cancer stem cells and
chemoresistance. Oncogenesis. 8:132019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu HF, Liu YP, Liu DL, Ma YD, Hu ZY, Wang
XY, Gu CS, Zhong Y, Long T, Kan HP and Li ZG: Role of
TGFβ3-Smads-Sp1 axis in DcR3-mediated immune escape of
hepatocellular carcinoma. Oncogenesis. 8:432019. View Article : Google Scholar
|
|
28
|
Bao WR, Li ZP, Zhang QW, Li LF, Liu HB, Ma
DL, Leung CH, Lu AP, Bian ZX and Han QB: Astragalus polysaccharide
RAP selectively attenuates paclitaxel-induced cytotoxicity toward
RAW 264.7 cells by reversing cell cycle arrest and apoptosis. Front
Pharmacol. 9:15802019. View Article : Google Scholar :
|
|
29
|
Liu P, Peng QH, Tong P and Li WJ:
Astragalus polysaccharides suppresses high glucose-induced
metabolic memory in retinal pigment epithelial cells through
inhibiting mitochondrial dysfunction-induced apoptosis by
regulating miR-195. Mol Med. 25:212019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jia N, Qiao H, Zhu W, Zhu M, Meng Q, Lu Q
and Zu Y: Antioxidant, immunomodulatory, oxidative stress
inhibitory and iron supplementation effect of Astragalus
membranaceus polysaccharide-iron (III) complex on iron-deficiency
anemia mouse model. Int J Biol Macromol. 132:213–221. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jin M, Zhao K, Huang Q and Shang P:
Structural features and biological activities of the
polysaccharides from Astragalus membranaceus. Int J Biol Macromol.
64:257–266. 2014. View Article : Google Scholar
|
|
32
|
Yu M, Shi J, Sheng M, Gao K, Zhang L, Liu
L and Zhu Y: Astragalus inhibits epithelial-to-mesenchymal
transition of peritoneal mesothelial cells by down-regulating
β-catenin. Cell Physiol Biochem. 51:2794–2813. 2018. View Article : Google Scholar
|
|
33
|
Zhou Y, Liao S, Zhang Z, Wang B and Wan L:
Astragalus injection attenuates bleomycin-induced pulmonary
fibrosis via down-regulating Jagged1/Notch1 in lungs. J Pharm
Pharmacol. 68:389–396. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou Y, Tong X, Ren S, Wang X, Chen J, Mu
Y, Sun M, Chen G, Zhang H and Liu P: Synergistic anti-liver
fibrosis actions of total Astragalus saponins and glycyrrhizic acid
via TGF-β1/Smads signaling pathway modulation. J Ethnopharmacol.
190:83–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qian W, Cai X, Qian Q, Zhang W and Wang D:
Astragaloside IV modulates TGF-β1-dependent epithelial-mesenchymal
transition in bleomycin-induced pulmonary fibrosis. J Cell Mol Med.
22:4354–4365. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hao ZF, Su YM, Liu JY, Wang CM and Yang
RY: Astragalus polysaccharide suppresses excessive collagen
accumulation in a murine model of bleomycin-induced scleroderma.
Int J Clin Exp Med. 8:3848–3854. 2015.PubMed/NCBI
|
|
37
|
Tian Y, Li H, Qiu T, Dai J, Zhang Y, Chen
J and Cai H: Loss of PTEN induces lung fibrosis via alveolar
epithelial cell senescence depending on NF-κB activation. Aging
Cell. 18:e128582019.
|
|
38
|
Chae U, Lee H, Kim B, Jung H, Kim BM, Lee
AH, Lee DS and Min SH: A negative feedback loop between XBP1 and
Fbw7 regulates cancer development. Oncogenesis. 8:122019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tian B, Zhao Y, Sun H, Zhang Y, Yang J and
Brasier AR: BRD4 mediates NF-κB-dependent epithelial-mesenchymal
transition and pulmonary fibrosis via transcriptional elongation.
Am J Physiol Lung Cell Mol Physiol. 311:L1183–L1201. 2016.
View Article : Google Scholar
|
|
40
|
Ishak Gabra MB, Yang Y, Lowman XH, Reid
MA, Tran TQ and Kong M: IKKβ activates p53 to promote cancer cell
adaptation to glutamine deprivation. Oncogenesis. 7:932018.
View Article : Google Scholar
|