Introduction
Hepatocellular carcinoma (HCC) has been recognized
as one of the major causes of cancer-associated death worldwide,
affecting ~600,000 people annually (1,2).
The high incidence of postoperative recurrence and metastasis
contribute to a low survival rate for HCC patients (3,4);
therefore, exploring new and effective therapeutic approaches
against HCC metastasis is urgently needed.
Sevoflurane (Sevo) is a volatile anesthetic agent
clinically, which exerts a suppressive role in varying cancers. For
example, Liang et al (5)
have shown that Sevo inhibited the proliferation, induced apoptosis
and blocked cell cycle progression of lung carcinoma cells. Liu
et al (6) have found that
Sevo exerts a suppressive role on breast cancer cell proliferation
and cell cycle. Yang et al (7) simulated the effects of clinical use
of Sevo on colon cancer cells in vitro and found that Sevo
inhibited tumor cell growth and induced apoptosis. In contrast,
several preclinical studies reported that Sevo could promote breast
cancer growth and enhances renal cancer survival (8,9).
However, few studies have addressed the influence of Sevo on HCC;
therefore, this study explored the role of Sevo on the various
biological aspects of HCC cells.
MicroRNAs (miRNAs/miRs) are a family of short,
small, noncoding RNAs (an average size of 22 nucleotides), which
negatively regulate target gene expression (10,11). Increasing evidence has
demonstrated the important role of miRNAs in Sevo-mediated mediated
processes in numerous cancers. For example, Sevo inhibited the
migration and invasion of colorectal cancer cells by upregulating
miR-203 (12). Another study from
Sun et al (13) showed
that Sevo inhibited migration and invasion of colorectal cancer
cells by regulating miR-34a. Notably, Song et al (14) found that Sevo restored the
expression of miR-29 and in turn miR-29a inhibition abolished the
antitumor property of Sevo in HCC cells. Nonetheless, whether Sevo
exerts its antitumor effect by regulating miRNA in HCC is not fully
clear.
The present study analyzed the miRNA expression
profile following exposure to Sevo using microarray assay and
investigated the roles of Sevo in HCC cells. Subsequently, the
regulatory role and relevant mechanism of miR-25-3p in the
antitumor effect of Sevo were explored. The present findings may
provide a potential theoretical basis for the development of new
therapies for HCC.
Materials and methods
Cell culture and drugs
The human HCC cell lines HCCLM3 and Huh7, and 293T
cells were obtained from the American Type Culture Collection. All
cells were grown in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich; Merck
KGaA), 100 IU/ml penicillin and 100 mg/ml streptomycin at 37°C and
5% CO2 incubator. Sevo was obtained from Sigma-Aldrich;
Merck KGaA.
Sevo treatment
HCCLM3 and Huh7 cells were divided into 4 groups:
Control group, 1.7% Sevo group, 3.4% Sevo group and 5.1% Sevo
group. According to the experimental protocol as previously
described (15,16), cultured HCCLM3 and Huh7 cells were
placed in an air-tight glass chamber with inflow and outflow
connectors. The chamber atmosphere was kept continuously saturated
with water at 37°C. The entrance port of the chamber was connected
to anesthetic machine (Cicero-EM 8060, Drägerwerk AG & Co.
KGaA). Sevoflurane was delivered into the chamber by a Sevo
vaporizer (SEVORANE®; Abott Pharmaceutical Co. Ltd.)
attached to the anesthesia machine. The concentrations of Sevo in
the chamber were detected at the chamber exit port by a gas monitor
(PM 8060, Drägerwerk AG & Co. KGaA) that inlayed with the
anesthetic machine. The control group was exposed to 95% air/5%
CO2 at 6 l/min for 6 h. The sevoflurane group was
exposed to 1.7, 3.4, or 5.1% of sevoflurane mixed with 95% air/5%
CO2 at 6 l/min for 6 h. A stable sevoflurane
concentration was achieved within 5 min.
Cell proliferation
The antiproliferative effect of Sevo against HCC
cells was measured using MTT assay. At the end of transfection, 20
µl MTT solution (Sigma-Aldrich; Merck KGaA) was added to
each well (1×105/well) and cultured for 4 h.
Subsequently, MTT solution was aspirated and dimethylsulfoxide (200
µl/well) was added. The optical density absorbance of the
samples at 570 nm was detected by a micro-plate reader (Bio-Rad
Laboratories, Inc.).
Lactate dehydrogenase (LDH) release
assay
A colorimetric assay kit (Nanjing Jiancheng
Bioengineering Institute; http://www.njjcbio.com/) was used to quantify the LDH
released from the cultured HCCLM3 and Huh7 cells. After treated
with Sevo, HCCLM3 and Huh7 cells were collected, then centrifuged
at 400 × g for 5 min at 4°C. The supernatant was removed and 150
µl LDH release reagent was added, mixed completely, then
incubated at 37°C 5% CO2 for 1 h. Finally, 120 µl
supernatant was added to 96-well plates and cell cytotoxicity was
measured by the absorbance at 490 nm by a micro-plate reader
(Bio-Rad Laboratories, Inc.).
Transwell invasion assay
Transwell chambers (8-µm pore; BD
Biosciences; Becton, Dickinson and Company) coated with Matrigel
(BD Biosciences; Becton, Dickinson and Company) were used for
invasion assay. Briefly, 1×105 HCCLM3 and Huh7 cells
were added in the top chamber with DMEM, while the lower chamber
added DMEM containing 20% FBS. After 24 h incubation, the invasion
cells was counted and images were captured of five independent
visual fields under the fluorescence microscope (Olympus Corp.) at
×200 magnification.
Wound-healing assay
When HCCLM3 and Huh7 cells reached ~80% confluence,
cells were serum starved overnight and the monolayer was scratched
with a 10 µl pipette tip, and then the wound area was
measured at 0 and 24 h under the fluorescence microscope (Olympus
Corp.) and Image J analysis software 1.46 (National Institute of
Health) was used to calculated the migration distances.
Transfection assay
When HCCLM3 and Huh7 cells in six-well plate grown
to ~80% confluence, miR-25-3p mimics (20 nmol/l), miR-25-3p
inhibitor (20 nmol/l) and their corresponding negative controls
(NC-mimic/NC-inhibitor; 20 nmol/l) were transfected into cells at
37°C for 24 h, using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). miR-25-3p mimics, mimics NC,
miR-25-3p inhibitor and inhibitor NC were obtained from Guangzhou
RiboBio Co., Ltd. The miR-25 mimics sequence is 5′-CAU UGC ACU UGU
-CUC GGU CUG A-3′. The control RNA mimics sequence is 5′-UCA CAA
CCU CCU AGA AAG AGU AGA-3′. The miR-25 inhibitor sequence is 5′-UCA
GAC CGA GAC AAG UGC A AU G-3′. The control miRNA inhibitor sequence
is 5′-UUU GUA CUA CAC AAA AGU ACU G-3′.
miRNA microarray analysis
Total RNA from HCCLM3 cells treated with or without
3.4% Sevo were extracted using the miRNeasy Mini kit (Qiagen,
Inc.). The samples were assessed using the miRCURY LNA™ Array v.
18.0 (Agilent Technologies, Inc.). The procedure and imaging
processes were as described previously (17).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from cells were isolated by using TRIzol
reagent (Takara Biotechnology, Co., Ltd.). Reverse transcription of
miR-25-3p was performed at 42°C using the miScript II RT kit (cat.
no. 4366597; Thermo Fisher Scientific, Inc.). miR-25-3p expression
was measured using the iCycler iQ Multicolor RT-qPCR System
(Bio-Rad Laboratories, Inc.). The primers for RT-qPCR analysis were
as follows: MiR-25-3p forward, 5′-TCT GGT CTC CCT CAC AGG AC-3′ and
reverse 5′-CAT GGG TCG CCT ACT CAC-3′; U6 forward, 5′-TGC GGG TGC
TCG CTT CGC AGC-3′ and reverse, 5′-CCA GTG CAG GGT CCG AGGT-3′. The
PCR thermocycling conditions were as follows: 5 min at 95°C, and 36
cycles of 10 sec at 95°C, 10 sec at 58°C and 20 sec at 72°C. The
miRNA relative expression was analyzed using the 2−ΔΔCq
method (18) and determined by
normalization to U6.
Target gene analyses of miR-25-3p
Bioinformatics tools, including TargetScan 7.0
(targetscan.org/) and miRanda (microrna.org/), were used to predict the potential
target genes of miR-25-3p.
Luciferase reporter assay
miRNA target prediction tools, including PicTar
version 2007 (https://pictar.mdc-berlin.de/), Miranda (http://miranda.org.uk) and TargetScan Release 7.0
(http://targetscan.org/) were used to search for
the putative targets of miR-25-3p. The dual-luciferase reporter
assay was performed as described previously (19). 293T cells were co-transfected with
miR-25-3p mimics, miR-25-3p inhibitor and the luciferase reporter
plasmids using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 48 h post-transfection, luciferase activities
were measured with the dual luciferase reporter kit (Beyotime
Institute of Biotechnology). Normalization of firefly luciferase
activity to Renilla luciferase activity was subsequently
performed.
Western blotting
Western blot was performed as previously described
(19,20). Briefly, 40 µg extracted
protein samples were transferred onto a polyvinylidene difluoride
(EMD Millipore) membrane and then blocked with 5% skim milk for 2 h
at 4°C. Then each membrane was probed with primary antibodies
against PTEN, phosphorylated-protein kinase B (p-Akt) (S473) (cat.
no. OMA1-03061; 1:1,000), Akt (cat. no. 44-609G; 1:1,000),
p-glycogen synthase kinase (GSK)-3β (S9) (cat. no. MA5-14873;
1:1,000), GSK-3β (cat. no. MA3-038; 1:1,000), β-catenin (cat. no.
71-2700; 1:1,000), c-Myc (cat. no. MA5-27025; 1:1,000), matrix
metallopro-teinase-9 (MMP-9; cat. no. MA5-15886; 1:1,000) and
β-actin (cat. no. MA5-15739-D800; 1:1,000) (all primary antibodies
were obtained from Thermo Fisher Scientific, Inc.) at 4°C for 20 h,
followed by horseradish peroxidase-conjugated goat anti-rabbit
immunoglobulin G (1:10,000; cat. no. 205718; Abcam). β-actin served
as the loading control and for normalization of protein expression.
The protein bands were developed using ECL kit (GE Healthcare) and
blot bands were quantified with ImageJ (version 1.46; Rawak
Software, Inc.).
Statistical analyses
Statistical analysis was performed by SPSS 18.0
(SPSS, Inc.). Each experiment was repeated three times. All data
were presented as mean ± standard deviation. The comparisons among
data were calculated by one-way analysis of variance followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
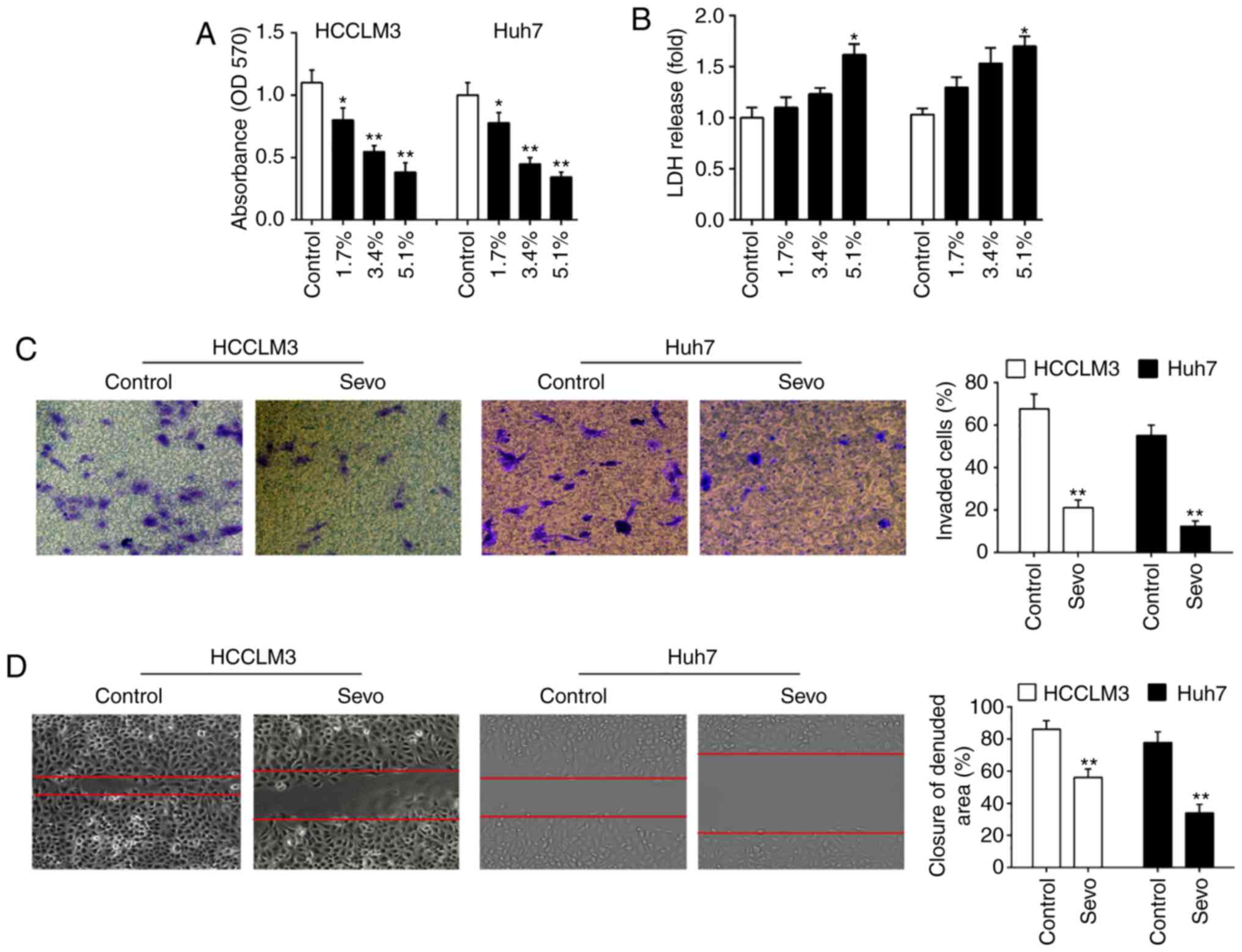
Sevo inhibits the proliferation, invasion
and migration of HCC cells
To explore the effects of Sevo on HCC cells, HCCLM3
and Huh7 cells were exposed to different concentrations of Sevo
(1.7, 3.4, and 5.1%) for 6 h, and then the cell viability was
assessed. The results showed that Sevo treatment suppressed the
HCCLM3 and Huh7 cells viability in a dose-dependent manner
(Fig. 1A). Cytotoxicity of Sevo
on HCCLM3 and Huh7 cells was measured by LDH assays. It was shown
that LDH release in 5.1% Sevo group was slightly increased,
compared with that in control group (Fig. 1B) indicating that the decreased
cell viability induced by Sevo is associated with the some degree
of toxicity. The invasiveness of both HCCLM3 and Huh7 cells was
further tested using the Transwell assay. It was found that Sevo
treatment significantly suppressed the invasive ability of both
HCCLM3 and Huh7 cells (Fig. 1C).
Moreover, the wound healing assay showed that Sevo treatment
significantly inhibited wound closures compared with the control
group (Fig. 1D). Collectively,
these data indicated that Sevo displays the anti-tumor activity on
HCC cells.
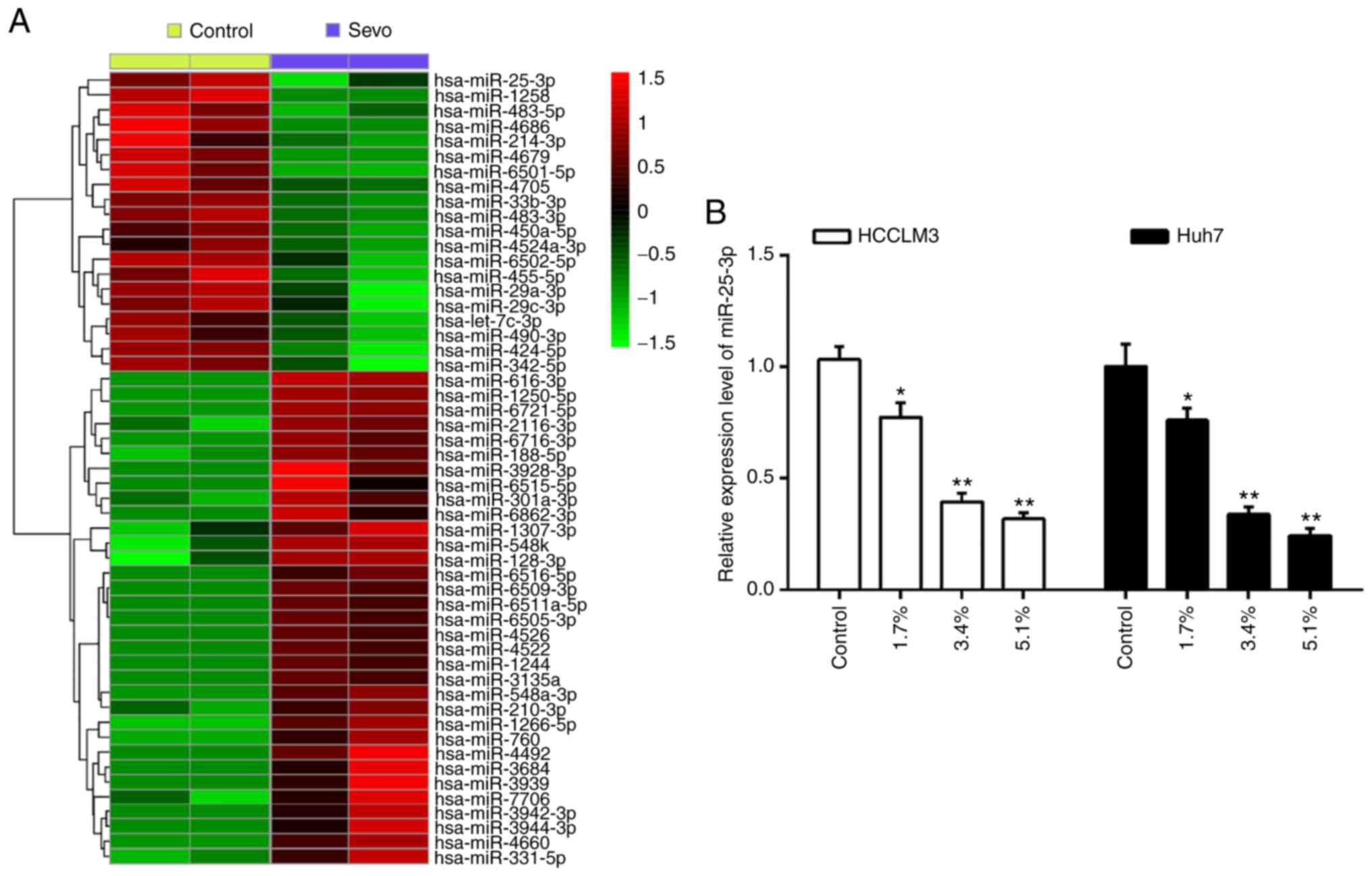
Sevo downregulates the expression of
miR-25-3p in hepatocellular carcinoma cells
Several studies have addressed the pharmacological
activities of Sevo, especially its anticancer effects, through the
regulation of cell miRNAs (6,21).
To explore the potential role of miRNAs in the anti-tumor activity
of Sevo, the differentially expressed miRNAs in HCCLM3 cells
treated with or without Sevo were identify using a miRNA
microarray. As shown in Fig. 2A,
of the 53 miRNAs screened, 20 were downregulated and 33 were
upregulated by Sevo in HCCLM3 cells. Of interest, miR-25-3p
displays a significant decrease after Sevo treatment. Additionally,
miR-25-3p is well known oncogenic miRNA in several types of human
cancers, including HCC (22-24); therefore, it appears plausible
that Sevo's observed anticancer effects might be exerted through
miR-25-3p in HCC cells.
Using RT-qPCR, the expression of miR-25-3p was
detected in Sevo treated HCC cells. The results showed that Sevo
significantly downregulated miR-25-3p expression in both HCCLM3 and
Huh7 cells, and this effect was dose-dependent (Fig. 2B). These results further suggested
that miR-25-3p may play important roles in the anticancer effects
of Sevo on HCC.
miR-25-3p is involved in the anticancer
effects of Sevo on HCC cells
To determine whether miR-25-3p was required for
Sevo-mediated processes in HCC cells, miR-25-3p mimics and
miR-25-3p inhibitor were added into the HCCLM3 and Huh7 cells at 6
h after administering Sevo, and incubated for 48 h. miR-25-3p
levels were significantly increased or decreased in HCCLM3 and Huh7
cells after transfection with the mimic or inhibitor, respectively
(Fig. 3A and B). Subsequently,
cell viability, invasion and migration were assessed. It was
observed that 3.4% Sevo treatment significantly inhibited the
proliferation, invasion and migration of HCCLM3 and Huh7 cells, but
these inhibitory effects were attenuated by miR-25-3p mimics
(Fig. 3C-E). In contrast, Sevo's
inhibitory effects on cell proliferation, invasion and migration
were enhanced by the miR-25-3p inhibitor (Fig. 3F-H). Overall, these data support
miR-25-3p as an essential target of Sevo for mediating the
anticancer effects of HCC cells in vitro.
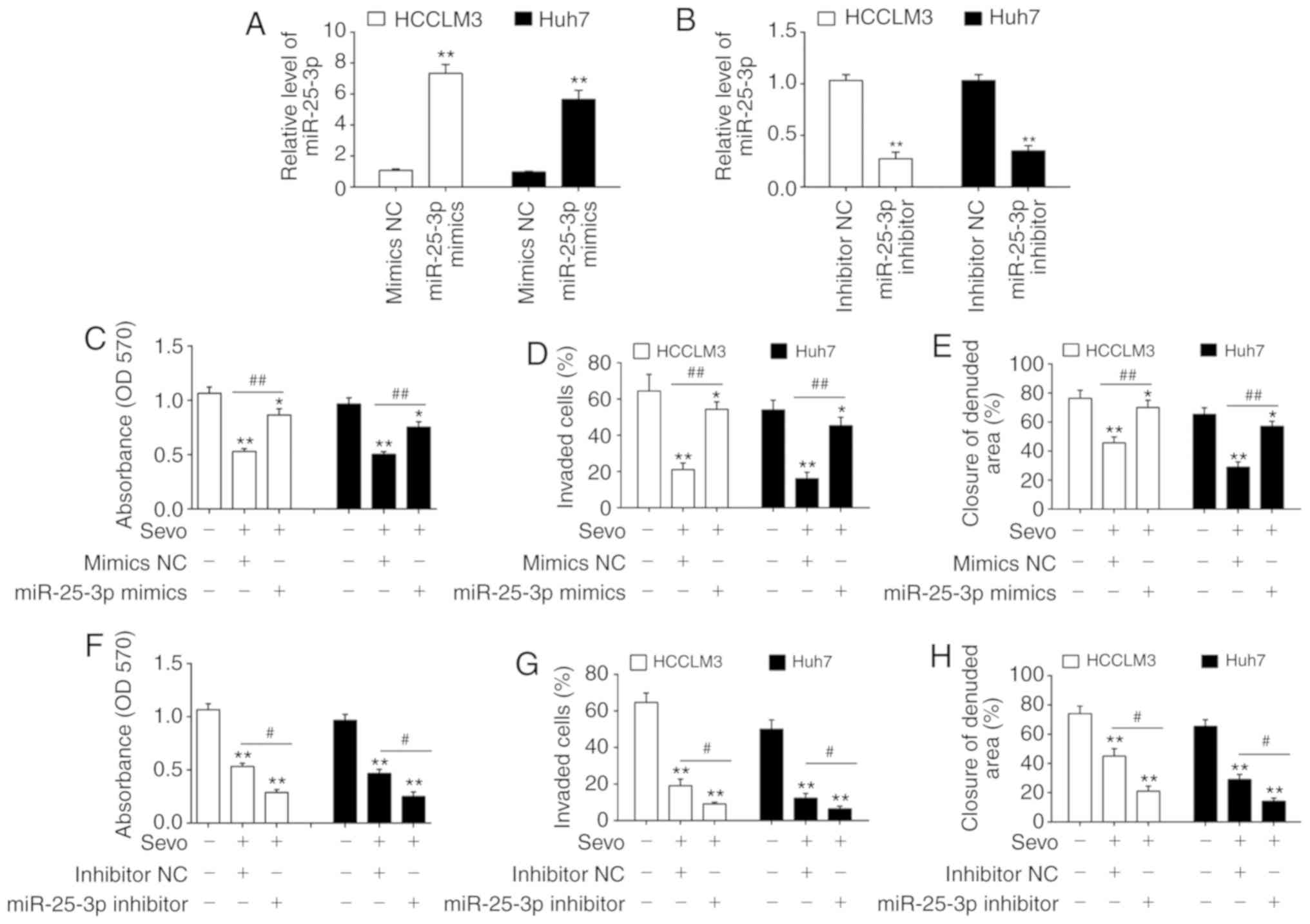 | Figure 3miR-25-3p is involved in the
anticancer effects of Sevo on hepatocellular carcinoma cells. (A)
miR-25-3p mimics or (B) miR-25-3p inhibitor were transfected into
HCCLM3 and Huh7 cells for 24 h, and then cells were treated with
Sevo for 6 h. The expression of miR-25-3p was determined by reverse
transcription quantitative-PCR. Data are presented as the mean ±
SD. (n=3) of one representative experiment, **P<0.01
vs. mimics NC or inhibitor NC group. The effect of the miR-25-3p
mimic on (C) cell proliferation was determined by MTT assay, (D)
cell invasion was measured by Transwell assay and (E) cell
migration was detected by wound healing assay. The effect of the
inhibitor was tested on (F) cell proliferation, (G) cell invasion
and (H) Cell migration. Data are the mean ± SD. (n=3) of three
representative experiment, *P<0.05 and
**P<0.01 vs. mimics NC or inhibitor NC group.
#P<0.05 and ##P<0.01 vs. Sevo + mimics
NC or Sevo + inhibitor NC. Sevo, sevoflurane; NC, negative control;
SD, standard deviation; miR/miRNA, microRNA; OD, optical
density. |
PTEN is a direct target of miR-25-3p
To examine the molecular mechanism by which
miR-25-3p mediated the anticancer effects of Sevo in HCC, candidate
target genes of miR-25-3p were computationally screened using
TargetScan 7.0 and miRanda. The bioinformatics analysis showed that
a putative target site of miR-25-3p in the 3'-UTR of PTEN (Fig. 4A). To validate whether miR-25-3p
targeted PTEN, a luciferase reporter assay was performed. The
luciferase reporter assay showed that the miR-25-3p mimics
significantly inhibited the luciferase activity in the PTEN-3'UTR
wild type reporter and that the miR-25-3p inhibitor caused an
increased luciferase activity; however, no changes were observed in
the cells co-transfected with PTEN 3'-UTR-mutant with miR-25-5p
(Fig. 4B). Western blot analyses
demonstrated that PTEN was decreased in miR-25-3p mimics
transfected HCC cells, while increased in miR-25-3p inhibitor
transfected HCC cells (Fig. 4C).
These data suggest that PTEN is a direct target of miR-25-3p.
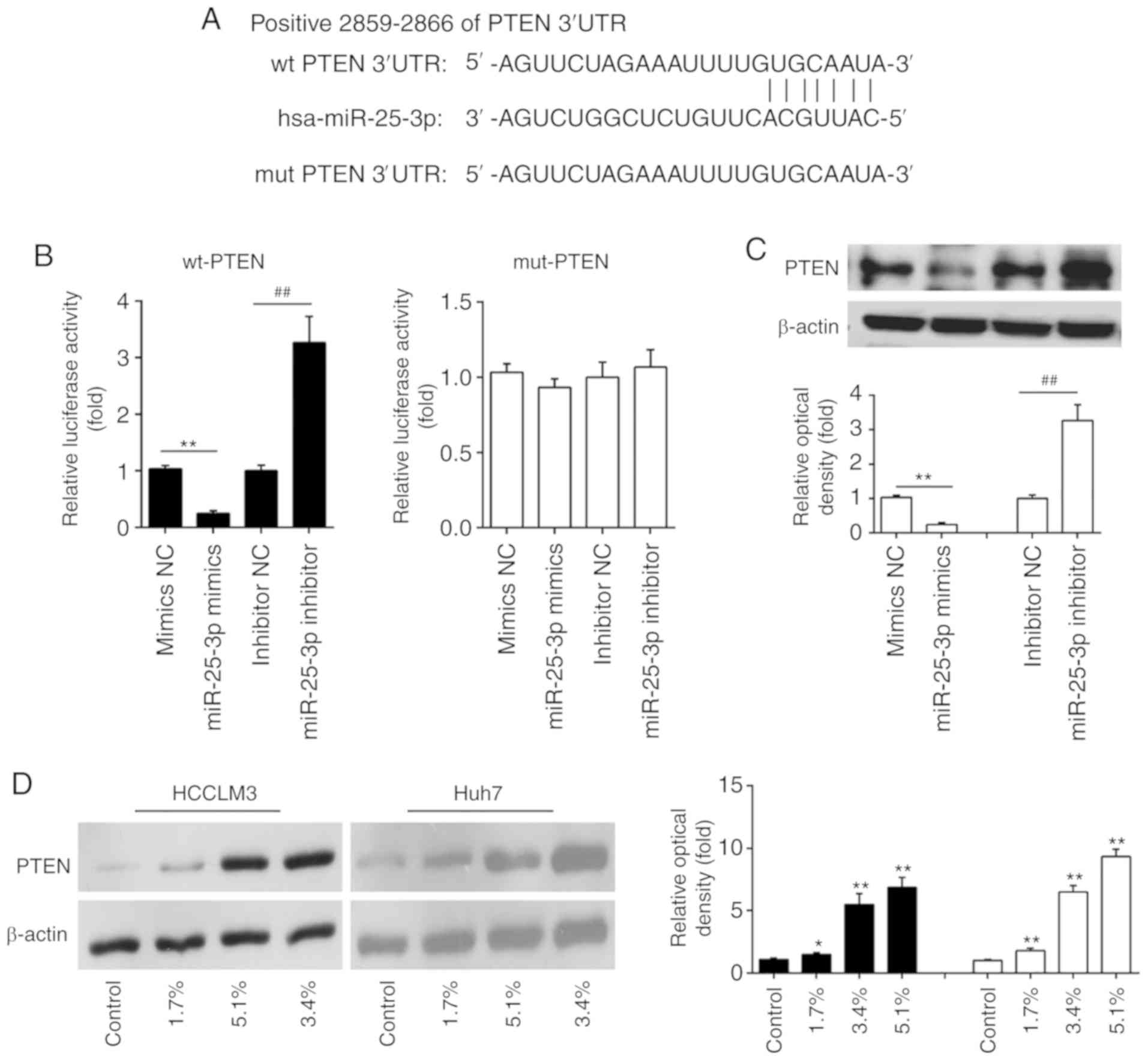 | Figure 4PTEN is a direct target of miR-25-3p
in hepatocellular carcinoma cells. (A) The putative binding site of
miR-25-3p and PTEN is shown. (B) 293T cells were cotransfected with
the PTEN wt or mut 3'-UTRs and miR-25-3p mimics and miR-25-3p
inhibitor, and then the luciferase activities were quantified
(n=3). (C) The expression of PTEN was measured using western
blotting. Data are the mean ± SD. (n=3) of one representative
experiment, **P<0.01 vs. mimics NC,
##P<0.01 vs. inhibitor NC. (D) HCCLM3 and Huh7 cells
were treated with different concentrations (0, 1.7, 3.4, and 5.1%)
of Sevo for 6 h and the expression of PTEN was measured using
western blotting. β-actin served as the loading control. Data are
presented as the mean ± SD. (n=3) of three representative
experiment, *P<0.01 and **P<0.01 vs.
NC. Sevo, sevoflurane; NC, negative control; SD, standard
deviation; miR/miRNA, microRNA; OD, optical density; PTEN,
phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase and
dual-specificity protein phosphatase PTEN; UTR, untranslated
region; wt, wild type; mut, mutant. |
It is well-known that PTEN acts as a tumor
suppressor in various human cancers (25,26). Given the relationship between Sevo
and miR-25-3p, the present study further examined whether the
expression of PTEN is regulated by Sevo. The expression of PTEN in
HCCLM3 and Huh7 cells treated with Sevo was measured by western
blotting. As shown in Fig. 4D,
Sevo treatment dose-dependently upregulated the expression levels
of PTEN in HCCLM3 and Huh7 cells. These findings support the
possible roles of the miR-25-3p/PTEN axis in the anticancer effects
of Sevo on HCC cells.
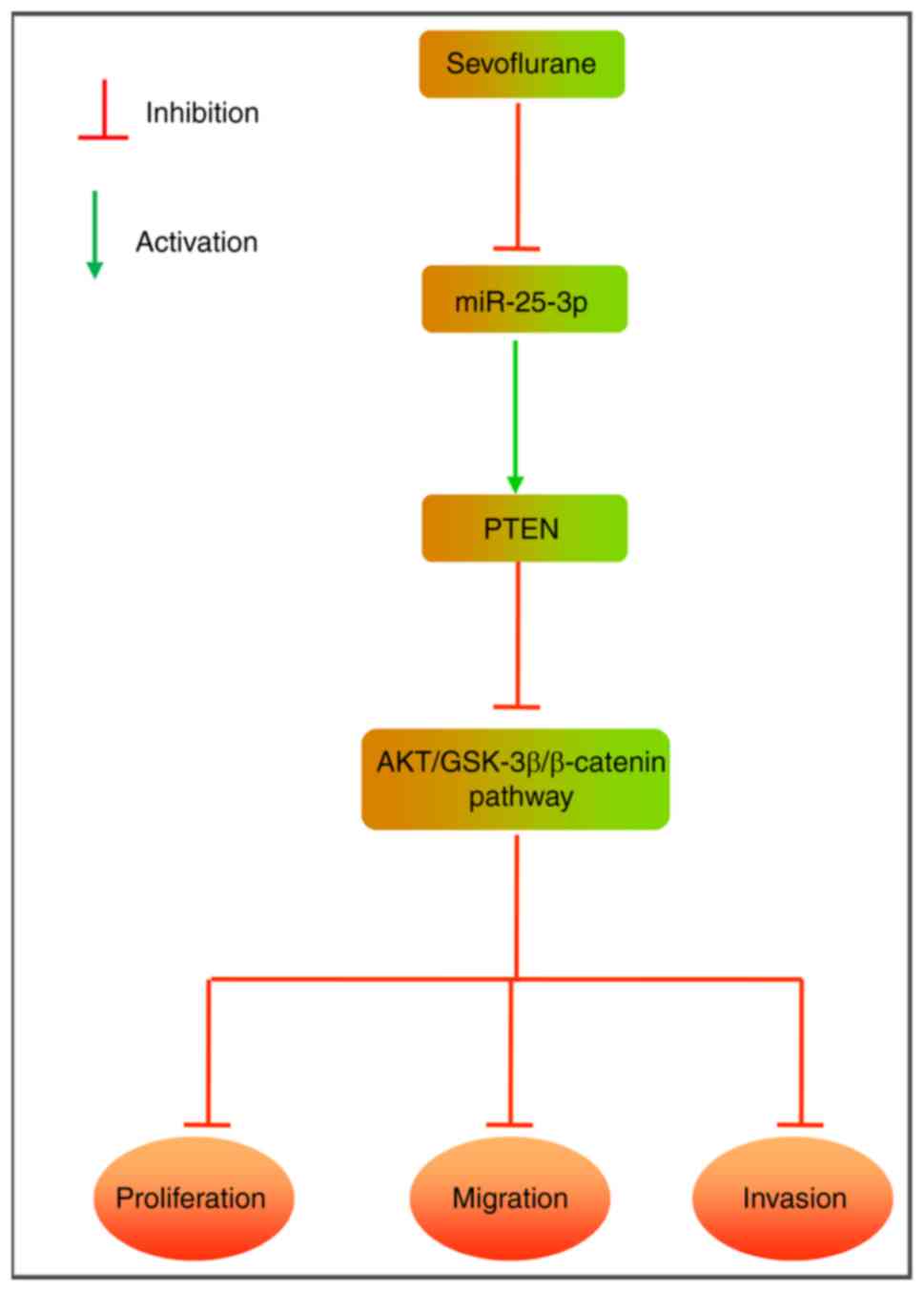
Sevo blocks the PTEN/Akt/GSK-3β/β-catenin
signaling pathway by modulating the expression of miR-25-3p in HCC
cells
As the Akt/GSK-3β/β-catenin pathway is associated
with cell migration and invasion in numerous cancers (20,27), the effect of Sevo on the
Akt/GSK-3β/β-catenin pathway was investigated in HCC cells. The
results of western blot-ting showed that Sevo treatment reduced the
expression of key proteins, including p-Akt (S473), p-GSK3β (S9),
c-Myc, β-catenin and MMP9, but the inhibitory effects of Sevo on
protein expression were partly restored by miR-25-3p overexpression
(Fig. 5A and B). These data
suggest that Sevo blocks the PTEN/Akt/GSK-3β/β-catenin pathway by
downregulating the expression of miR-25-3p in HCC cells.
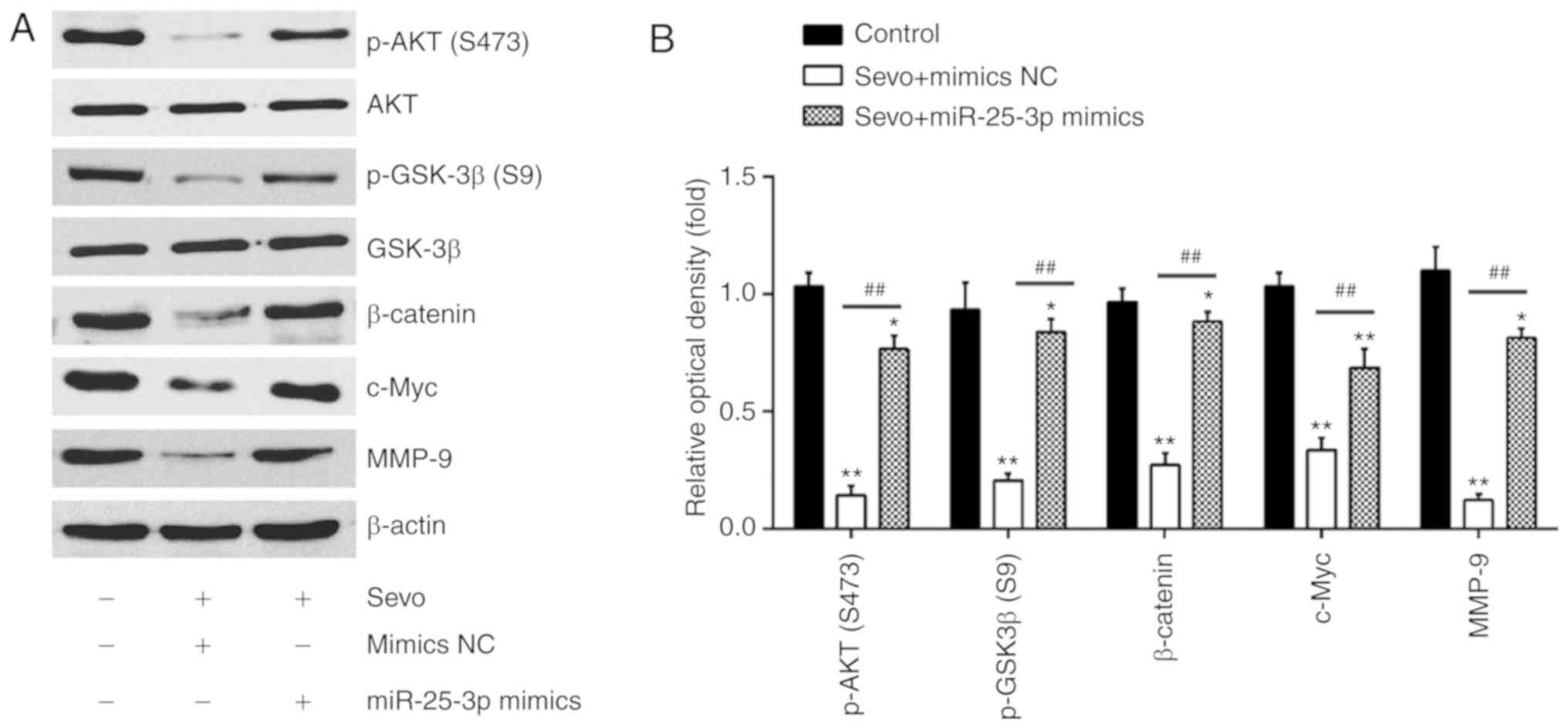 | Figure 5Sevo blocks the
PTEN/Akt/GSK-3β/β-catenin signaling pathway by modulating the
expression of miR-25-3p in HCC cells. HCCLM3 and Huh7 cells were
transfected with miR-25-3p mimics or miR-25-3p inhibitor for 24 h
followed by Sevo treatment for 6 h. (A) Protein levels of p-Akt,
Akt, p-GSK-3β, GSK-3β, β-catenin, c-Myc and MMP9 were measured
using western blotting. β-actin served as the loading control. (B)
The bands were semi quantitatively analyzed using ImageJ and
normalized to β-actin density. Data are presented as the mean ±
standard deviation. (n=3) of three representative experiments.
*P<0.01 and **P<0.01 vs. NC group.
##P<0.01 vs. Sevo + mimics NC. PTEN,
phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase and
dual-specificity protein phosphatase PTEN; Sevo, sevoflurane; NC,
negative control; SD, standard deviation; miR/miRNA, microRNA; MMP,
matrix metalloproteinase; PTEN, phosphatidylinositol
3,4,5-trisphosphate 3-phosphatase and dual-specificity protein
phosphatase PTEN; GSK, glycogen synthase kinase; p-Akt,
phosphorylated-protein kinase B. |
Sevo exerts its antitumor effects by
blocking the PTEN/Akt/GSK-3β/β-catenin signaling pathway in HCC
cells
To validate the role of the
PTEN/Akt/GSK-3β/β-catenin pathway on Sevo's observed anticancer
effects on HCC cells, HCCLM3 and Huh7 cells were treated with 1
µM PI3K inhibitor LY294002 and 100 ng/ml PI3K signaling
activator IGF-1. The results indicated that LY294002 enhanced
Sevo's inhibitory effects on cell proliferation, invasion and
migration in HCCLM3 and Huh7 cells. In contrast, IGF-1 reversed
these inhibitory effects (Fig.
6A-C). Taken together, the present findings suggest that Sevo's
anticancer effects on HCC cells might be modulated by blocking the
PTEN/Akt/GSK-3β/β-catenin signaling pathway (Fig. 7).
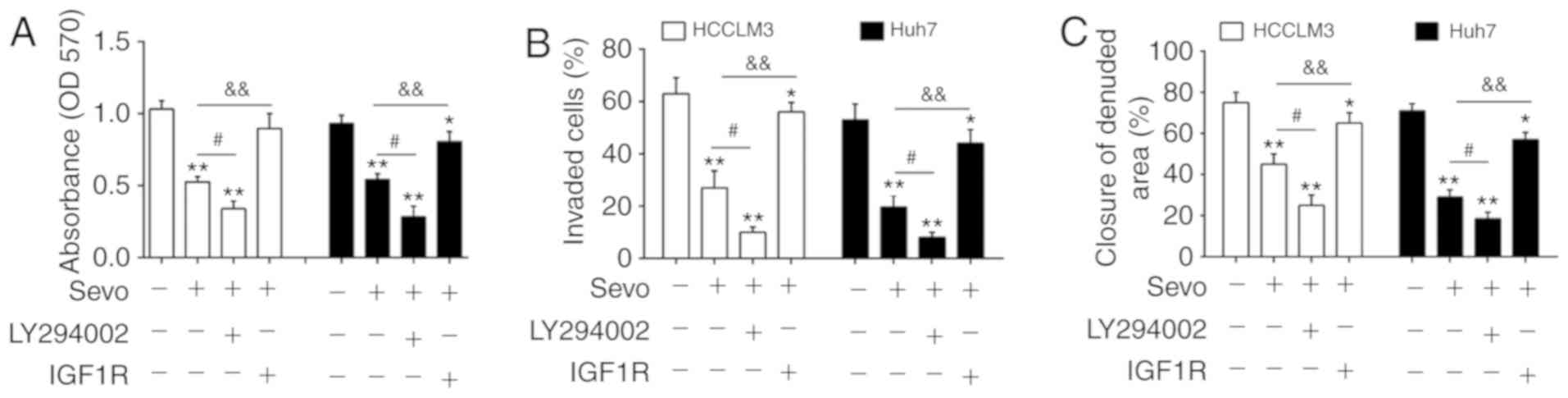 | Figure 6Sevo exerts its antitumor effects by
blocking the PTEN/Akt/GSK-3β/β-catenin signaling pathway through
the downregulation of miR-25-3p in hepatocellular carcinoma cells.
HCCLM3 and Huh7 cells were treated with 1 µM PI3K inhibitor
LY294002 or 100 ng/ml PI3K signaling activator insulin-like growth
factor-1 for 24 h followed by Sevo treatment for 6 h. (A) Cell
proliferation was determined by MTT assay. (B) Cell invasion was
measured by Transwell assay. (C) Cell migration was detected by
wound healing assay. Data are the mean ± standard deviation. (n=3)
of three representative experiments, *P<0.01 and
**P<0.01 vs. negative control group.
#P<0.05 vs. Sevo, &&P<0.01 vs.
Sevo. Sevo, sevoflurane; NC, negative control; miR/miRNA, microRNA;
OD, optical density; PTEN, phosphatidylinositol 3,4,5-trisphosphate
3-phosphatase and dual-specificity protein phosphatase PTEN;GSK,
glycogen synthase kinase; p-Akt, phosphorylated-protein kinase B;
PI3K, phosphatidylinositol 3 kinase. |
Discussion
The present study found that Sevo inhibited the
proliferation, invasion and migration of HCC cells. Moreover, Sevo
was found to downregulate the level of miR-25-3p in HCC cell lines
and proved that the miR-25-3p/PTEN/Akt/GSK-3β/β-catenin axis was
responsible for the antitumor actions of Sevo in HCC cells. It is
of clinical significance for anesthesiologists to select volatile
anesthetics for surgical resection of HCC.
Previous studies have indicated that Sevo displays
its anticancer activity in numerous human cancers. For example,
Liang et al (28) have
shown that Sevo actually inhibited lung cancer cell invasion by
suppressing the activation of the p38 mitogen associated protein
kinase pathway. Liang et al (5) have also found that Sevo inhibited
proliferation by inducing cell apoptosis in A549 cells.
Müller-Edenborn et al (29) found that Sevo reduced the
invasiveness of colon cancer cells in vitro by
downregulating MMP9. Sevo could suppress hypoxia-induced growth and
metastasis of lung cancer cells via inhibiting hypoxia-inducible
factor-1α (30). Sevo has an
inhibitory effect on the migration and MMP-2 activity in glioma
cells (31). In addition, Sevo
exerts its antiproliferative effect on colon cancer cells (32,33); therefore, taking into account the
inhibitory properties of Sevo against the tumor cells, Sevo is a
better choice of anesthetics during cancer surgery. However, few
reports have addressed the roles of Sevo in HCC. The present
results indicate that Sevo markedly suppressed cell proliferation,
invasion and migration of HCCLM3 and Huh7 cells, which supports the
efficacy of Sevo on tumor growth and invasion.
Several studies have also reported that miRNAs are
implicated in processes affected by Sevo in numerous conditions
(34,35). For example, Shao and Xia (36) showed that Sevo repressed
neurogenesis by regulating miR-183 expression in newborn rats. Zhao
et al (34) found that
Sevo upregulated the expression of miR-19-3p and inhibited the
expression of CCNA2, thus resulting in the impairment of learning
and memory in neonatal rats. Otsuki et al (37) found that Sevo exhibited a
protective effect against endotoxin-induced acute injury in rats by
regulating inflammation-associated miRNAs. Moreover, increasing
evidence has indicated that Sevo exerts antitumor effects on
several cancers by modulating the expression of several miRNAs. For
example, Sevo inhibited cell migration and invasion by the
induction of miR-637 in glioma (21). Liu et al (6) found that Sevo suppresses breast
cancer cell growth by modulating miR-203 expression. Therefore the
present study hypothesized that Sevo inhibited cell migration and
invasion of HCC cells by regulating miRNAs. This study found that a
large number of miRNAs were differently expressed following
exposure to Sevo in HCC cells and the most downregulated miR-25-3p
were selected for further study.
A number of miRNAs have been revealed to be involved
in the pathogenesis of HCC (38).
For example, upregulation of miR-21 could enhance cell
proliferation, reduce cell apoptosis and favor invasion (39). These findings suggest that miRNAs
could become novel molecular targets for HCC treatment. The
oncogenic roles of miR-25-3p in numerous types of cancers have
previously been investigated (40,41). In HCC, one previous study
demonstrated that miR-25-3p was upregulated in HCC tissues when
compared with adjacent normal tissues and that the upregulation of
miR-25-3p is of predictive value on poor prognosis (22). Another study from Wang et
al (23) revealed that
miR-25-3p dramatically stimulated HCC cell growth and activated the
epithelial-mesenchymal transition. Thus, the present study
hypothesized that miR-25-3p has an important role in Sevo's
observed anticancer effects on HCC cells. As expected, the current
results showed that the antitumor effects by Sevo was suppressed by
miR-25-3p overexpression, while enhanced by miR-25-3p inhibition.
These findings suggest that Sevo's anti-HCC effects are mediated by
downregulating miR-25-3p.
PTEN is a tumor suppressor gene, which could
negatively regulate the Akt/GSK-3β/β-catenin pathway (42). In addition, PTEN could regulate
proliferation, migration and invasion of HCC (43,44). Moreover, Wan et al
(45) showed that miR-25-3p
promoted malignant phenotypes of retinoblastoma by targeting PTEN
and activating Akt signaling pathway. In the present study, PTEN
was directly targeted by miR-25-3p in HCC cells and its expression
was upregulated after exposure to Sevo. Moreover, it was also found
that Sevo decreased p-Akt, p-GSK3β, β-catenin, c-Myc and MMP-9
expression; whereas, overexpression of miR-25-3p obviously reversed
this inhibitory effect, which suggested that Sevo suppressed this
signaling pathway by downregulating the miR-25-3p/PTEN axis. In
addition, the PI3K inhibitor LY294002 enhanced Sevo's inhibitory
effects on HCC cell proliferation, invasion and migration, while
these inhibitory effects were reversed with the PI3K signaling
activator IGF-1. The above results suggest that Sevo inhibits the
proliferation, invasion and migration of HCC cells in part by
downregulating miR-25-3p through suppressing the
PTEN/Akt/GSK-3β/β-catenin signaling pathway.
There are still some limitations in the present
study. For example, this exploratory study summarizes the novel
role of Sevo in regulating the miR-25-3p/PTEN/Akt/GSK-3β/β-catenin
pathway and subsequently exerts its anti-tumor activity in
vitro. However, more experiments in vivo should be
performed to confirm that miR-25-3p mediated the anti-tumor
activity of Sevo in HCC. Moreover, further study is needed to
explore the regulatory functions of miR-25-3p in the other
signaling pathways, such as Rho GDP dissociation inhibitor
α/Wnt/β-catenin pathway (23).
Thus, future research is needed to gain deeper insight into these
questions.
The current study demonstrates that Sevo exerts its
anti-tumor activity through inactivation of
PTEN/Akt/GSK-3β/β-catenin pathway by the downregulation of
miR-25-3p. The present study provides new clinical implications for
the potential role of Sevo in HCC surgery.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YC, WL and WD performed the experiments, contributed
to data analysis and wrote the paper. YC, WL and WD analyzed the
data. JL conceptualized the study design, contributed to data
analysis and experimental materials. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the No. 6 Medical
Center, General Hospital of PLA Ethics Committees.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ghouri YA, Mian I and Rowe JH: Review of
hepatocellular carcinoma: Epidemiology, etiology, and
carcinogenesis. J Carcinog. 16:12017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McGlynn KA and London WT: The global
epidemiology of hepatocellular carcinoma: Present and future. Clin
Liver Dis. 15:223–243. vii–x. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tang ZY: Hepatocellular carcinoma-cause,
treatment and metastasis. World J Gastroenterol. 7:445–454. 2001.
View Article : Google Scholar
|
|
4
|
Tang Z, Zhou X, Lin Z, Yang B, Ma Z, Ye S,
Wu Z, Fan J, Liu Y, Liu K, et al: Surgical treatment of
hepatocellular carcinoma and related basic research with special
reference to recurrence and metastasis. Chin Med J (Engl).
112:887–891. 1999.
|
|
5
|
Liang H, Gu MN, Yang CX, Wang HB, Wen XJ
and Zhou QL: Sevoflurane inhibits proliferation, induces apoptosis,
and blocks cell cycle progression of lung carcinoma cells. Asian
Pac J Cancer Prev. 12:3415–3420. 2011.PubMed/NCBI
|
|
6
|
Liu J, Yang L, Guo X, Jin G, Wang Q, Lv D,
Liu J, Chen Q, Song Q and Li B: Sevoflurane suppresses
proliferation by upregulating microRNA-203 in breast cancer cells.
Mol Med Rep. 18:455–460. 2018.PubMed/NCBI
|
|
7
|
Yang X, Zheng YT and Rong W: Sevoflurane
induces apoptosis and inhibits the growth and motility of colon
cancer in vitro and in vivo via inactivating Ras/Raf/MEK/ERK
signaling. Life Sci. 239:1169162019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ciechanowicz S, Zhao H, Chen Q, Cui J, Mi
E, Mi E, Lian Q and Ma D: Differential effects of sevoflurane on
the metastatic potential and chemosensitivity of non-small-cell
lung adenocarcinoma and renal cell carcinoma in vitro. Br J
Anaesth. 120:368–375. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ecimovic P, McHugh B, Murray D, Doran P
and Buggy DJ: Effects of sevoflurane on breast cancer cell function
in vitro. Anticancer Res. 33:4255–4260. 2013.PubMed/NCBI
|
|
10
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fan L, Wu Y, Wang J, He J and Han X:
Sevoflurane inhibits the migration and invasion of colorectal
cancer cells through regulating ERK/MMP-9 pathway by up-regulating
miR-203. Eur J Pharmacol. 850:43–52. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun SQ, Ren LJ, Liu J, Wang P and Shan SM:
Sevoflurane inhibits migration and invasion of colorectal cancer
cells by regulating microRNA-34a/ADAM10 axis. Neoplasma.
66:887–895. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song G, Tian L, Cheng Y, Liu J, Wang K, Li
S and Li T: Antitumor activity of sevoflurane in HCC cell line is
mediated by miR-29a-induced suppression of Dnmt3a. J Cell Biochem.
120:18152–18161. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roesslein M, Frick M, Auwaerter V, Humar
M, Goebel U, Schwer C, Geiger KK, Pahl HL, Pannen BH and Loop T:
Sevoflurane-mediated activation of p38-mitogen-activated
stresskinase is independent of apoptosis in Jurkat T-cells. Anesth
Analg. 106:1150–1160, table of contents. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loop T, Scheiermann P, Doviakue D,
Musshoff F, Humar M, Roesslein M, Hoetzel A, Schmidt R, Madea B,
Geiger KK, et al: Sevoflurane inhibits
phorbolmyristate-acetate-induced activator protein-1 activation in
human T lymphocytes in vitro: Potential role of the p38-stress
kinase pathway. Anesthesiology. 101:710–721. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mei LL, Wang WJ, Qiu YT, Xie XF, Bai J and
Shi ZZ: MiR-125b-5p functions as a tumor suppressor gene partially
by regulating HMGA2 in esophageal squamous cell carcinoma. PLoS
One. 12:e01856362017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Yao Y, Sun F and Lei M: MiR-25 inhibits
sepsis-induced cardiomyocyte apoptosis by targetting PTEN. Biosci
Rep. 38:BSR201715112018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Q, Xu HX, Li JP, Wang S, Fu Z, Jia J,
Wang L, Zhu ZF, Lu R and Yao Z: Growth differentiation factor 15
induces growth and metastasis of human liver cancer stem-like cells
via AKT/GSK-3β/β-catenin signaling. Oncotarget. 8:16972–16987.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yi W, Li D, Guo Y, Zhang Y, Huang B and Li
X: Sevoflurane inhibits the migration and invasion of glioma cells
by upregulating microRNA-637. Int J Mol Med. 38:1857–1863. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su ZX, Zhao J, Rong ZH, Geng WM, Wu YG and
Qin CK: Upregulation of microRNA-25 associates with prognosis in
hepatocellular carcinoma. Diagn Pathol. 9:472014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang C, Wang X, Su Z, Fei H, Liu X and Pan
Q: MiR-25 promotes hepatocellular carcinoma cell growth, migration
and invasion by inhibiting RhoGDI1. Oncotarget. 6:36231–36244.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Gong X, Tian K, Chen D, Sun J,
Wang G and Guo M: MiR-25 promotes glioma cell proliferation by
targeting CDKN1C. Biomed Pharmacother. 71:7–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu LF, Wu ZP, Chen Y, Zhu QS, Hamidi S and
Navab R: MicroRNA-21 (miR-21) regulates cellular proliferation,
invasion, migration, and apoptosis by targeting PTEN, RECK and
Bcl-2 in lung squamous carcinoma, Gejiu City, China. PLoS One.
9:e1036982014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song MS, Salmena L and Pandolfi PP: The
functions and regu-lation of the PTEN tumour suppressor. Nat Rev
Mol Cell Biol. 13:283–296. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park NR, Cha JH, Jang JW, Bae SH, Jang B,
Kim JH, Hur W, Choi JY and Yoon SK: Synergistic effects of CD44 and
TGF-β1 through AKT/GSK-3β/β-catenin signaling during
epithelial-mesenchymal transition in liver cancer cells. Biochem
Biophys Res Commun. 477:568–574. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang H, Gu M, Yang C, Wang H, Wen X and
Zhou Q: Sevoflurane inhibits invasion and migration of lung cancer
cells by inactivating the p38 MAPK signaling pathway. J Anesth.
26:381–392. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Müller-Edenborn B, Roth-Z'graggen B,
Bartnicka K, Borgeat A, Hoos A, Borsig L and Beck-Schimmer B:
Volatile anesthetics reduce invasion of colorectal cancer cells
through down-regulation of matrix metalloproteinase-9.
Anesthesiology. 117:293–301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang H, Yang CX, Zhang B, Wang HB, Liu
HZ, Lai XH, Liao MJ and Zhang T: Sevoflurane suppresses
hypoxia-induced growth and metastasis of lung cancer cells via
inhibiting hypoxia-inducible factor-1α. J Anesth. 29:821–830. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hurmath FK, Mittal M, Ramaswamy P,
Umamaheswara Rao GS and Dalavaikodihalli Nanjaiah N: Sevoflurane
and thiopental preconditioning attenuates the migration and
activity of MMP-2 in U87MG glioma cells. Neurochem Int. 94:32–38.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kvolik S, Glavas-Obrovac L, Bares V and
Karner I: Effects of inhalation anesthetics halothane, sevoflurane,
and isoflurane on human cell lines. Life Sci. 77:2369–2383. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kvolik S, Dobrosevic B, Marczi S, Prlic L
and Glavas-Obrovac L: Different apoptosis ratios and gene
expressions in two human cell lines after sevoflurane anaesthesia.
Acta Anaesthesiol Scand. 53:1192–1199. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao X, Jin Y, Li H, Jia Y and Wang Y:
Sevoflurane impairs learning and memory of the developing brain
through post-transcriptional inhibition of CCNA2 via
microRNA-19-3p. Aging (Albany NY). 10:3794–3805. 2018. View Article : Google Scholar
|
|
35
|
Xu C, Niu JJ, Zhou JF and Wei YS:
MicroRNA-96 is responsible for sevoflurane-induced cognitive
dysfunction in neonatal rats via inhibiting IGF1R. Brain Res Bull.
144:140–148. 2019. View Article : Google Scholar
|
|
36
|
Shao CZ and Xia KP: Sevoflurane anesthesia
represses neurogenesis of hippocampus neural stem cells via
regulating microRNA-183-mediated NR4A2 in newborn rats. J Cell
Physiol. 234:3864–3873. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Otsuki T, Ishikawa M, Hori Y, Goto G and
Sakamoto A: Volatile anesthetic sevoflurane ameliorates
endotoxin-induced acute lung injury via microRNA modulation in
rats. Biomed Rep. 3:408–412. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gramantieri L, Fornari F, Callegari E,
Sabbioni S, Lanza G, Croce CM, Bolondi L and Negrini M: MicroRNA
involvement in hepatocellular carcinoma. J Cell Mol Med.
12:2189–2204. 2008. View Article : Google Scholar
|
|
39
|
Liu C, Yu J, Yu S, Lavker RM, Cai L, Liu
W, Yang K, He X and Chen S: MicroRNA-21 acts as an oncomir through
multiple targets in human hepatocellular carcinoma. J Hepatol.
53:98–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen H, Pan H, Qian Y, Zhou W and Liu X:
MiR-25-3p promotes the proliferation of triple negative breast
cancer by targeting BTG2. Mol Cancer. 17:42018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao H, Wang Y, Yang L, Jiang R and Li W:
MiR-25 promotes gastric cancer cells growth and motility by
targeting RECK. Mol Cell Biochem. 385:207–213. 2014. View Article : Google Scholar
|
|
42
|
Wang F, Qi X, Li Z, Jin S, Xie Y and Zhong
H: lncRNA CADM1-AS1 inhibits cell-cycle progression and invasion
via PTEN/AKT/GSK-3β axis in hepatocellular carcinoma. Cancer Manag
Res. 11:3813–3828. 2019. View Article : Google Scholar :
|
|
43
|
He J, Mu M, Luo Y, Wang H, Ma H, Guo S,
Fang Q, Qian Z, Lu H and Song C: MicroRNA-20b promotes
proliferation of H22 hepatocellular carcinoma cells by targeting
PTEN. Oncol Lett. 17:2931–2936. 2019.PubMed/NCBI
|
|
44
|
Han Y, Chen M, Wang A and Fan X:
STAT3-induced upregulation of lncRNA CASC11 promotes the cell
migration, invasion and epithelial-mesenchymal transition in
hepatocellular carcinoma by epigenetically silencing PTEN and
activating PI3K/AKT signaling pathway. Biochem Biophys Res Commun.
508:472–479. 2019. View Article : Google Scholar
|
|
45
|
Wan W, Wan W, Long Y, Li Q, Jin X, Wan G,
Zhang F, Lv Y, Zheng G, Li Z and Zhu Y: MiR-25-3p promotes
malignant phenotypes of retinoblastoma by regulating PTEN/Akt
pathway. Biomed Pharmacother. 118:1091112019. View Article : Google Scholar : PubMed/NCBI
|