1. Clinical aspects of COVID-19 infections;
acute respiratory distress syndrome (ARDS); known and potential
treatments
In China, the first comprehensive analysis published
on the COVID-19 (1) included
44,672 cases and 1,023 deaths, with an overall case-fatality rate
(CFR) of 2.3%. There were 0 deaths in patients 9 years old or
younger, and the CFR increased with advancing age (70-79 years:
8.0%, 80 years and older: 14.8%) and also with comorbid conditions:
5.6% for cancer, 6.0% for hypertension, 7.3% for diabetes, 10.5%
for cardiovascular disease; for intensive care patients CFR was
49.0%. Similar data were published by other authors on the China
epidemic (2,3); another analysis (4) identified 3 biological markers
associated with the severity of the infection and progression to
ARDS and death: i) neutrophilia; ii) increased lactate
dehydrogenase LDH and iii) coagulation dysfunction (D-dimer). There
were 2 unexpected findings: i) patients with fever ≥39°C were more
likely to develop ARDS but also less likely to die; and ii)
administering methylprednisolone to patients with ARDS was
associated with lower risk of death.
In Europe there was a higher rate of severe cases in
Italy (5,6) where 12% of positive patients were
admitted to ICU vs. 5% in China; this difference was attributed to
a higher proportion of elderly and increased social contact
(7,8). Data from Germany (9) showed that ~1/3 of admitted patients
had leucopenia and most of these (80%) had lymphopenia; C-reactive
protein (CRP) was often increased and very high CRPs were
associated with a less favorable outcome for the patient.
Approximately 40% of admitted patients had increased LDH, D-dimer
and/or thrombocytopenia, and an LDH value of >400 IU/ml was
associated with more severe disease. Troponin was increased in a
small number of patients, with unclear significance.
In the USA, a recent publication showed that a
majority (71%) of COVID-19 patients with ARDS and in intensive
care, also had hypotension (10,11). There is already an extensive
clinical experience with ARDS in the USA, where >180,000
patients annually are admitted with this pathology and ARDS
mortality has improved from 40 to 50% 10 years ago to 20-25% in
recent clinical trials. A group of USA intensive care specialists
led by Dr Calfee has analyzed >30 biomarkers and clinical
variables from two large clinical trials on ARDS (12); subsequently an ARDS patient
population was identified which comprised ~30% of total ARDS cases,
had higher levels of IL-6 and IL-8, more patients with sepsis and
in need for vasopressor treatment, with higher mortality but also
responding better to high positive expiratory-end pressure (PEEP)
therapy. With data from more clinical trials (13), two ARDS populations were
identified, the hypoinflammatory and hyperinflammatory
subphenotypes, with distinctive and opposite characteristics. The
former has low levels of IL-6, IL-8, tumor necrosis factor receptor
1 (TNFr1), lower mortality as well as higher number of
ventilator-free days, high bicarbonate and protein C; the latter
has high levels of IL-6, IL-8, TNFr1, low levels of bicarbonate and
protein C, low number of ventilator-free days and higher
mortality.
Treatment-wise, the same group showed that in the
ARDS hyperinflammatory subphenotype better outcomes were obtained
with administration of high PEEP, liberal fluid strategy (vs.
conservative fluid administration) and simvastatin, while
rosuvastatin administration was of no benefit (12,14-16); Table
I summarizes the data from different clinical trials on ARDS
subphenotypes. The observed difference between statins in ARDS may
be explained by their solubility: while simvastatin is
lipid-soluble, rosuvastatin is water-soluble, with important
consequences on cell membrane interaction.
 | Table IARDS subphenotypes (data from Wilson
and Calfee) (13). |
Table I
ARDS subphenotypes (data from Wilson
and Calfee) (13).
| Parameters | Hypoinflammatory
ARDS | Hyperinflammatory
ARDS |
|---|
| IL-6, IL-8, TNFr1
(plasma levels) | Low | High |
| Bicarbonate, CRP
(plasma levels) | High | Low |
| Mortality | Low | High |
| Ventilator-free
days | High | Low |
| PEEP (positive
end-expiratory pressure) associated with better 90-day mortality
(ALVEOLI study) | Low | High |
| Fluid strategy with
better outcome for 90-day mortality (FACCT study) | Conservative
administration of fluids | Liberal
administration of fluids |
| Simvastatin
administration has benefit on 28- and 90-day survival (HARP-2) | No | Yes |
| Rosuvastatin
administration has benefit on 90-day survival (HARP-2) | No | No |
| ARDS, acute
respiratory distress. | | |
It is safe to say that COVID-19 patients who develop
ARDS (with high levels of interleukins, low blood pressure) belong
to the hyperinflammatory subphenotype and may benefit from the
therapeutic insight associated with this group of patients (higher
PEEP, liberal fluid strategy, and simvastatin). In critically-ill
COVID-19 patients, increased LDH levels are likely associated with
acidosis and if low blood pressure is also present, before
vasopressors they are likely to benefit from administration of
sodium bicarbonate (increments of 100 ml of 8.4% solution).
Treatment-wise, after the SARS and MERS epidemics a
few antivirals were proposed and tested and among these are
mycophenolic acid, cyclosporine at low concentrations, chloroquine;
chlorpromazine, loperamide, and lopinavir, found to be
broad-spectrum coronavirus inhibitors (17). Recently FDA has approved
remdesivir (GS5734 - an RNA polymerase inhibitor) as specific
treatment for COVID-19; different protocols use chloroquine (500 mg
q 12 h); hydroxychloroquine (200 mg q 12 h); lopinavir/ritonavir
(400/100 mg every 12 h), α-interferon (aerosol inhalation 5 million
IUbid), and there are >50 treatments tested in clinical trials
(18).
For ARDS, as the most severe form of COVID-19
infection, there is ample data from proteomics showing highly
activated pathways of inflammation, increased levels of eosinophil-
and neutrophil-derived proteins, epithelial and endothelial injury,
matrix metalloproteinase 7 (MMP7), α- and β-hemoglobin,
apolipoprotein A1, osteopontin and chemokines (19,20). The context of increased
inflammation in ARDS justifies the use of biological agents,
antagonists of the IL-6 receptor (IL-6R), and TNF-α, such as
infliximab, rituximab, ustekinumab, etanercept, adalimumab and
tocilizumab (18). However, the
complexity of living biological systems with redundant pathways and
dual-role modulators means that they do not always conform to
mechanistic determinism, and this is especially true in patients
with chronic diseases or co-morbid conditions, where modifications
of physiological pathways translate into unknown but important
factors with unexpected consequences. One such example is a
clinical trial for sepsis which tested a recombinant IL-1 receptor
antagonist (IL-1Ra) with negative results, but at the same time
benefited patients with higher baseline levels of IL-1Ra (21).
Another important aspect is that multiple cellular
pathways besides inflammation are simultaneously affected in ARDS,
including coagulation, endothelial and epithelial injury pathways
(apoptosis) and healing (fibrosis) in lungs. A unique molecular
signature of lethal infection with another respiratory virus (PR8,
an influenza virus) showed over-activation of pro-inflammatory
pathways, NF-κB, IL-6, TNF and neutrophil chemotaxis (22), associated decreased activation of
genes involved in lung homeostasis and repair. Neutrophils with
high pro-inflammatory activity were numerous in lung infiltrates,
their own chemokines further promoted neutrophil influx in a
positive feedback loop; a dose-dependent survival was seen with
partial neutrophil depletion in this type of lethal infection.
Comparing viral replication in non-lethal vs. lethal infection they
had similar viral replication rates, but the lethal infection had
increased viral titers in the lungs, and also increased early
pro-inflammatory neutrophil activation, resulting in pathologic
neutrophil infiltration and fatal lung damage (23). Similar observations increased
neutrophil- and neutrophil-attracting chemokines, delay in
induction of IFN in airway epitlelial cells resulting in vascular
leakage and lung damage were made in SARS and MERS suggesting a
common pathogenical mechanism (24,25). Indeed, the significant role of
neutrophils in the promotion of ARDS-related alveolal damage was
supported by a report in sepsis ARDS patients, where high
neutrophil counts in bronchiolar lavage material harvested late in
ARDS progression was associated with reduced patient survival
(26). Hence, it has been
proposed that the sulphone dapsone may show efficacy in COVID-19
related ARDS, as the drug is highly and rapidly effective at
inhibiting cytokine-mediated neutrophil chemotaxis and respiratory
burst in a variety of therapeutic contexts (27). Efficacy of Dapsone in the
treatment of Pneumocystis pneumonia suggests it will
penetrate the lung tissue sufficiently to attenuate neutrophil
activity. The haemotoxicity of the drug can be ameliorated with
concomitant cimetidine administration (27,28).
Overall, the major clinical manifestations: sore
throat, cough, breathing difficulties, malaise, fever, chills,
diarrhea, generalized myalgia, drowsiness, dyspnea, and pneumonia
are similar in SARS-CoV2, MERS-CoV, and SARS CoV infections
(29), but COVID-19
differentiates itself from MERS and SARS in infectivity (R0), with
much higher human-to-human transmission rates owing much to the
fact that infectivity is not restricted to symptomatic patients
(30); however, we can improve
therapeutic strategies with inferences from structural comparisons
of these viruses.
2. SARS-CoV2 molecules and proposed
antiviral drugs
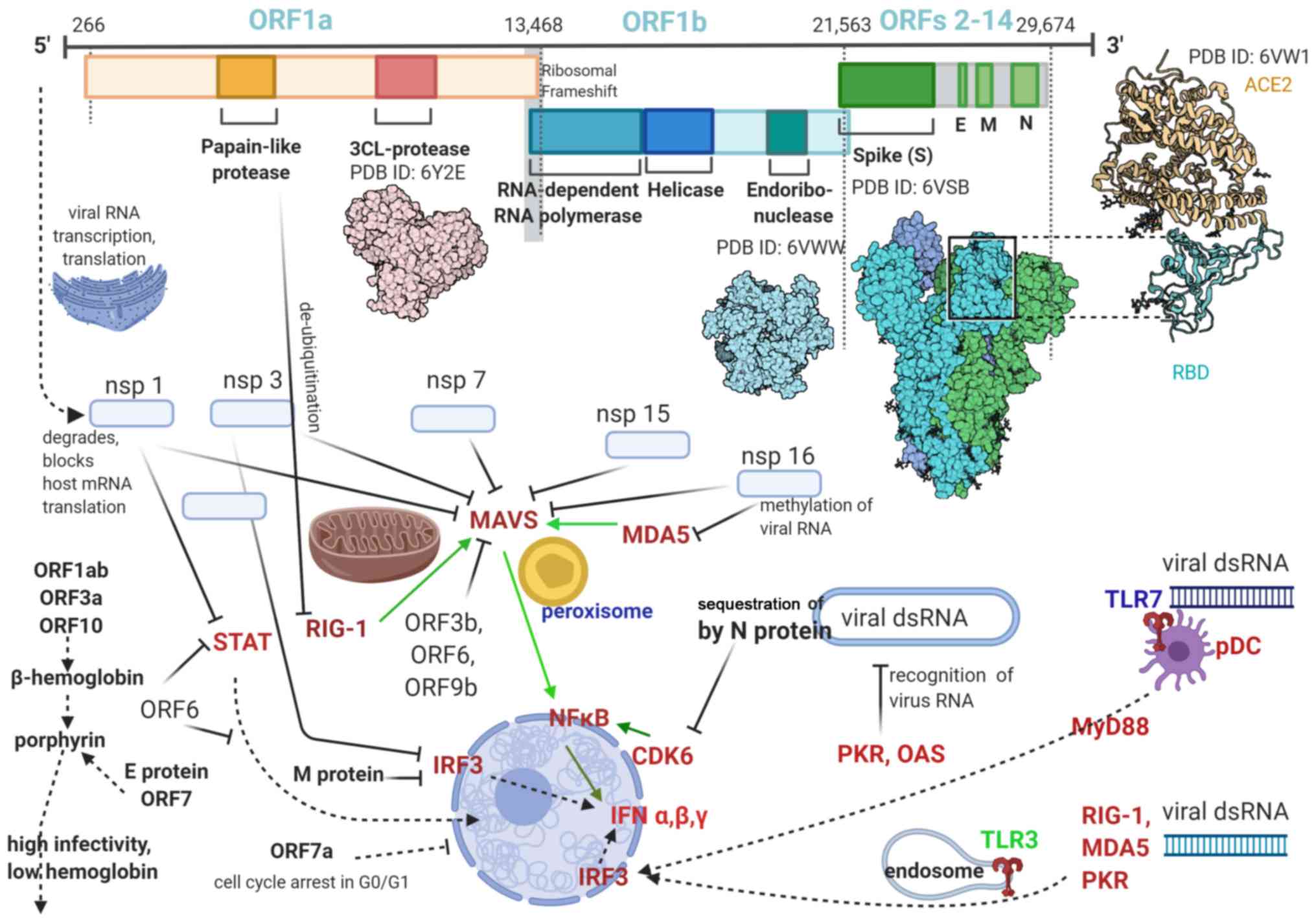
SARS-CoV2 consists of two types of molecules: RNA
and proteins; RNA encodes for 27 viral proteins, of which 16 are
non-structural proteins (nsps), and 11 are accessory and structural
proteins. There are 4 major structural proteins: nucleocapsid
protein (N), spike surface glycoprotein (S), matrix protein (M),
and small envelope protein (E). The N-protein binds to viral
RNA, encloses it in a capsid and assists RNA synthesis and folding
during viral replication; it also modifies host responses, cell
cycle and gene translation, and after RNA replication it guides the
viral membrane proteins for viral assembly. The S-protein is
essential for attachment and fusion of the virus to host cells
(31).
Sequencing the SARS-CoV2 genome in February 2020
(32) showed that
phylogenetically it belongs to the genus Betacoronavirus,
subgenus Sarbecovirus, and has closer genetic similarity to
SARS-CoV than MERS-CoV. The genome consists of a single,
positive-stranded RNA with ~30 k (29,811) nucleotides; from these
only 5 nucleotides were found to be different between SARS-CoV2 and
SARS-CoV. Similar sequencing results (>99.9% similarity) were
obtained by different teams on different patient samples (33,34): GenBank sequences MN988668,
NC_045512, MN938384.1, MN975262.1, MN985325.1 and MN994468.1.
Coronaviruses have the largest known RNA genomes and
their RNA is transcribed via open reading frames (ORFs); this
peculiarity increasing the efficacy of viral replication and
mutagenesis, and making it more difficult to be eliminated by the
immune system. The SARS-CoV2 genome has 14 ORFs of different
lengths; first ORF has ~10,000 nucleotides (~2/3 of viral RNA) and
encodes nsps 1-16, with the structural and accessory proteins being
translated from the remainder 1/3 of RNA (35) (Fig.
1).
Comparing the amino acid sequence of SARS-CoV and
SARS-CoV2 reveals identical sequences in the envelope, matrix,
nsp7, nsp13 and accessory proteins p6 and 8b, while nsp3 and nsp2
had 102 and 61 amino acid substitutions, respectively.
Importantly, orf8 and orf10 are present in SARS-CoV2
but not in SARS-CoV (36). The
orf8 protein from SARS-CoV2 has no known functional domain or motif
and is different from the orf8 or orf8b of SARS-CoV, which does not
have orf10 at all. However, ORF10 binds to heme and ORF8 to
porphyrins (37); heme
inactivation increases viral infectivity through inhibition of the
host innate antiviral response (38) which relies on heme-containing
enzymes such as nitric oxide synthase (detailed below); also
increases viral permeation of tissues via porphirin binding and
tissue destruction (39).
The spike (S) protein of the new virus contains
1,273 amino acids; compared to SARS-CoV it has 27 amino acid
substitutions, 8 in the heptad repeat domain, 6 in the RBD, and 4
in the S1 subunit, modifying two peptides which are antigens for
SARS-CoV (40). The binding
energy between the viral S protein and host ACE2 receptor is higher
in the new virus versus SARS-CoV (-50.6 kcal/mol vs. -78.6
kcal/mol) suggesting a more stable bond (41); at the same time the affinity of
the S protein of the new virus for the ACE2 receptor is in the 15
nM range, which is about 10-20 times higher than SARS-CoV, an
important factor for the differential infectivity (42).
The S protein is a class I viral fusion protein like
the influenza virus hemagglutinin (HA); SARS-CoV2 also has an S1/S2
cleavage site which is recognizable by furin, a ubiquitous cell
protease. When similar modifications (insertion of a polybasic
furin cleavage site) occur in the hemagglutinin proteins of avian
and human influenza viruses, their virulence is greatly increased
(43).
Functionally the S protein has a receptor-binding
subunit (S1), an S2 fusion domain (S2), separated by a cleavage
site (S1/S2) and another cleavage site (S2′) located within S2. The
multiple cleavage sites are activated by a wide variety of
proteases from the host cell: cathepsins, serine proteases from the
trypsin-like transmembrane serine protease family (TTSP), and
furin-like proprotein convertases.
Both SARS-CoV and SARS-CoV2 need S protein priming
through cleavage by a host cellular protease (trans-membrane
protease serine 2 - TMPRSS2, TMPRSS11a,
hypoxanthine-aminopterin-thymidine, trypsin, elastase, cathepsin L)
(44,45), and the cell surface from
respiratory epithelial cells has abundant TMPRSS2 and trypsin-like
protease (46). Interestingly,
human airway epithelial cells can be infected by SARS-CoV even when
proteases from cell surface are absent (44), meaning that a cell fusion process
used by SARS-CoV involves endosomal membranes and a treatment which
blocks only the cell surface proteases or the endosomal protease
will not prevent, but only depress viral cell entry; however, a
combination of two substances, camostat and EST, a cathespsin
inhibitor effectively blocked the viral entry into cells.
SARS-CoV2-infected cells aggregate to form
syncytiums; the S protein of SARS-CoV2 can mediate a cell-cell
syncytium formation without a proteolytic enzyme (trypsin), while
SARS-CoV S protein cannot; thus the membrane fusion capacity of
SARS-CoV2 is much higher than that of SARS-CoV (46).
The S protein of both coronaviruses has another
peculiarity: its dependency on Ca2+ (47). Significant cell membrane ordering
prior to membrane fusion is needed, and it requires the presence of
Ca2+ ions because of the negatively charged residues in
the fusion platform. Consequently the entry of SARS-CoV2 is highly
dependent on the Ca2+ concentration in its cell
environment, and calcium chelators inhibit viral fusion and cell
entry. Thus it was observed that SARS-CoV entry is blocked by
amiodarone, which blocks calcium channels in the endosome
and lysosome (47).
The nsps perform essential functions in immune
antagonism through formation of the viral replication complex and
double membrane vesicles which shield the viral RNA, viral RNA
proofreading, binding of nucleic acid and helicase activity
(48).
Immunosuppressive functions of the SARS-CoV2
proteins. Besides needing to ensure the entry of viral material
into cells and subsequent replication, the viral proteins and RNA
also need to evade the host immune system, and because the success
of viral infection depends on the impairment of the host antiviral
response, this aspect deserves special attention.
Airway epithelial cells are able to prevent viral
infections through multiple mechanisms. The airway surface liquid
(ASL) is secreted by the submucosal glands, goblet and club cells
and mobilized by cilium cells, and contains the gel-forming mucins
MUC5B and MUC5AC, glycosaminoglycan keratin sulfate, and
antimicrobial peptides and proteins (AMPs). ASL forms a
pericilliary layer with volume, fluidity, pH and microbiome
maintained in an optimal range for toxin elimination and the
actions of AMPs, and facilitating mobilization of the innate immune
response by macrophages, monocytes, dendritic cells, innate
lymphoid cells, and γ/δ T cells. Finally an adaptive, specific
immune response is triggered in response to antigens involving T
and B lymphocytes, chemokines, cytokines and antigen-neutralizing
IgAs (49).
After evading or overwhelming this first line of
defense and entering host cells, the virus activates the cellular
antiviral defenses effectuated by hundreds of proteins (cytokines,
chemokines, host restriction factors) which block different steps
in the viral replication and are activated by specific cell
sensors/receptors. Interferons (IFNs) via IFN-stimulated genes
(ISGs) initiate innate and adaptive immune responses which alter
host cell cycle, translation and apoptosis, virus entry, viral RNA
availability, stability and translation, particle and budding
(50).
Molecules involved in virus detection in the
cytoplasm of host cells are: the retinoic acid-inducible gene-I
(RIG-I), a RNA helicase with the respective cell receptors -
RIG-I-like receptors (RLR); and the melanoma
differentiation-associated protein 5 (MDA5), both of which have the
same downstream effector: the mitochondrial antiviral signaling
protein (MAVS). Besides RIG-1 and MDA5, other cellular viral RNA
sensors are known: the protein kinase RNA-activated (PKR),
oligoadenylate synthetase (OAS), latent endoribonuclease (RnaseL),
cyclic GMP-AMP (cGAMP) synthase (cGAS) (51); PKR stops host cell translation and
helps activate the NF-κB.
Early in infection the antiviral response is driven
by the peroxisomal MAVS, while later the mitochondrial MAVS
triggers an interferon (IFN)-dependent, sustained immune response.
MAVS signaling ultimately results in NF-κB (and other
pro-inflammatory pathways) activation which translocates to the
nucleus and activates the transcription of genes encoding IFN-α/β,
cytokines, many antiviral proteins, and RIG-I/MDA5 in a positive
feedback loop (52).
Following activation, IFN-α/β-type I IFNs are
produced in and activate most cells, while type III IFN (IFN-λ)
mostly mucosal cells; they activate very similar IFN-stimulated
genes (ISGs). IFN signaling occurs very fast since it does not
require new protein synthesis; all IFNs use the Janus kinase (JAK)
- signal transducer and activator of transcription (STAT) pathway.
pDCs produce most of the IFN-α during an infection; IFN-γ bridges
the innate and adaptive immune responses, helps regulate immune
function, and is mostly secreted by activated T cells and natural
killer (NK) cells (51).
TLRs from endosomes recognize viruses, while TLRs
from cell membrane typically recognize bacteria. MAVS and the TLR
adaptor MyD88 activate cell-specific transcriptional pathways, with
pro- or anti-inflammatory profiles (ex macrophages vs. fibroblasts)
(53).
All nucleated cells are thought to have RLR and PKR
receptors for viral nucleic acids, while toll-like receptors (TLRs)
are present in myeloid dendritic cells (mDCs) - TLR3/7 and
plasmacytoid dendritic cells (pDC) TLR7/8, and also in endosomes of
most cells (54).
TLR4 interacts with with the adaptor protein MyD88
and activates the mitogen-activated protein kinase (MAPK) and the
NF-κB signaling pathway. The c-Jun NH2-terminal kinases (JNK), the
signal-regulated kinases (ERKs), the p38 MAPKs, are major MAPKs
with important roles in innate immunity (55).
SARS blocks antiviral defenses and the IFN
production in multiple ways, at multiple sites, many involving
RIG-I and MAVS (50): i) after
transcription the coronaviral dsRNA is protected from detection by
storage in double-membrane vesicles; ii) M protein of SARS virus
localizes in membranes associated with the Golgi complex and binds
to the host RIG-I, thus impeding activation of MAVS; iii) ORF3b and
ORF6 move to the mitochondria and block the interaction between
RIG-I and MAVS to block IFN induction; iv) the papain-like protease
(PlPro) removes ubiquitin from RIG-I and down-regulates type-I IFN
response; v) cytoskeleton perturbations in mitochondria, with
modification of the mitochondrial membrane potential and impairment
of MAVS activity; vi) blocking IFN-β production by
de-ubiquitination (DUB) of RIG-1 and other viral sensors and
effectors (56).
Microarrays helped identify the proteins through
which viral particles can antagonize host immune responses: 13
proteins were inhibitors of MAVS (LGP2, A20, SMURF2, etc.) 14 for
RIG-1 (USP3, ARL16 and RNF122) and 6 for MDA5 (USP3, ARL5B,
TRIM59); RNF125 inhibited RIG-1, MDA5 and MAVS (57).
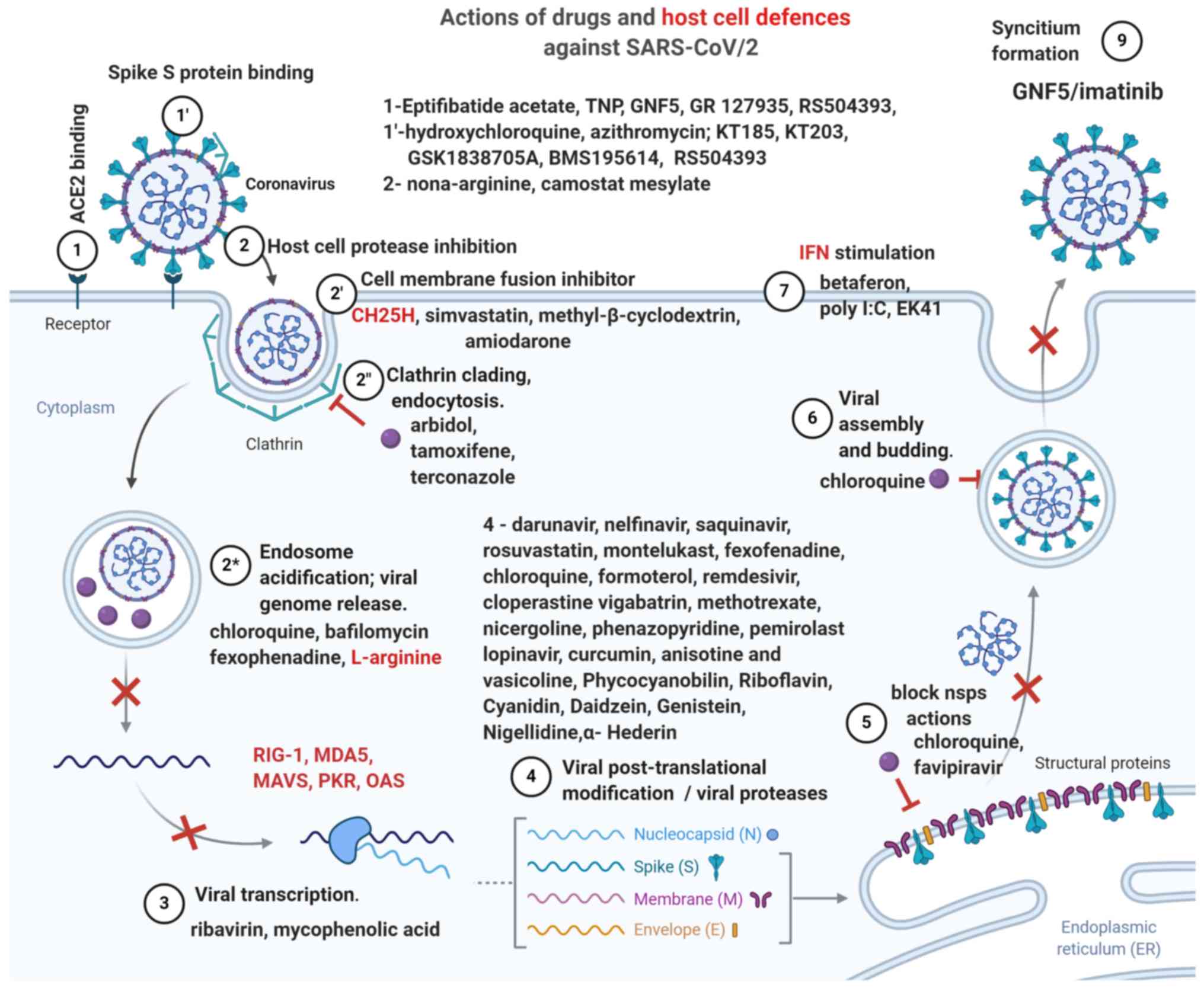
Proposed therapeutic interventions. The
current therapeutic interventions for COVID-19 (18) are derived from SARS treatments;
the first drugs shown in 2005 to be effective in vitro for
SARS, both pre- and post-exposure, are chloroquine and
hydroxychloroquine; since they were extensively studied and shown
to exert pleiotropic antiviral actions (58). Chloroquine is a weakly basic
substance and upon entering cells it is protonated and concentrated
in acidic organelles: lysosomes, Golgi vesicles and endosomes; the
subsequent increase in endosomal pH interferes with the terminal
glycosilation of the ACE2 receptor. Besides inhibiting the S
protein-induced viral fusion (59), and the PlPro protease (60) chloroquine can benefit patients
with porphirin extravasation (Fig.
2) by preventing orf1ab, orf3a, orf10 attack on hemoglobin
(37) with porphirin
formation.
Protease inhibitors target viral nsps needed for its
replication, such as the RNA-dependent RNA polymerase (RdRp)
inhibited by ribavirin (which also inhibits viral mRNA capping);
the 3C-like protease (3CLpro) is inhibited by the
lopinavir-ritonavir combination (48), which seems to have better results
compared to ribavirin, which was associated with anemia, hypoxia
and increased risk of death in SARS patients (61).
FDA-approved drugs and agents re-purposed for
COVID-19, such as ribavirin, IFNs and corticosteroids, were shown,
however, to be ineffective especially in severe CoV infections
(48); recent data on SARS-CoV2
suggest that more specific and efficacious treatments can be
administered to COVID-19 patients; Table II summarizes such proposed
FDA-approved drugs and Fig. 3
offers a graphic presentation.
| Table IIFDA-approved drugs with SARS-CoV2
antiviral actions. |
Table II
FDA-approved drugs with SARS-CoV2
antiviral actions.
| Substance analyzed
(Ref.) | Viral molecule
inhibited; mechanism | Method and/or
software used for testing FDA approved drugs |
|---|
| Hydroxychloroquine
and azithromycin (59) | Spike (S)
Protein - binding motifs of ACE2 receptor; | Virtualized quantum
mechanical modeling |
| Eptifibatide
acetate, TNP, GNF5, GR 127935, RS504393 (62) | ACE2 receptor
protein of host cells binding site | High-throughput
virtual screening used to investigate LOPAC library drugs |
| KT185, KT203
GSK1838705A, BMS195614, RS504393 (62) | S-protein (RBD of
spike protein) binding site anti-inflammatory | Software: PyRx,
Open Babel, AutoDock Vina, PyMol, GROMACS; online resources SWISS
MODEL, HADDOCK, RCSB PDB, NCBI, ProCheck at RCSB validation server,
ProSA-web, SAVES-Verify3D server |
| Darunavir,
Nelfinavir, | Protease: | Structure-based
drug repositioning |
| Saquinavir,
Rosuvastatin (63) | Mpro, central
site | |
| Montelukast,
Fexofenadine (63) | Protease: | Structure-based
drug repositioning |
| Mpro, terminal
site | |
| Chloroquine, | Protease: | Homology model of
the protease based on |
| formoterol, 16
other FDA- | papain-like
protease | SARS-coronavirus
PLpro structure; drugs |
| approved drugs
(60) | (PLpro) | docked in S3/S4
pockets of active site |
| Remdesivir (-13.1
KJ/mol) | Protease | Connectivity map
and the docking configurations used to simulate the docking bonding
energy between antiviral and the respective protease in KJ/mol |
| cloperastine
(-10.4) | 3CLpro (PDB ID
6LU7) | |
| vigabatrin
(-10.2) | | |
| methotrexate
(-6.9) | | |
| Remdesivir (-18.6
KJ/mol) | Protease | Used PyRx for
loading and visualising the SDF files for |
| vigabatrin
(-12.1) | 6Y84 - main
protease with | the ligands and the
AutoDock Vina tool for testing the |
| cloperastine
(-10.1) nicer- | unliganded active
site | docking on two
target proteins 6LU7 and 6Y84 from PDB |
| goline (-9.0)
phenazo- | | |
| pyridine (-8.4)
(64) | | |
| Pemirolast
(65) | Protease; RNA
Replicase inhibitor | Using COVID-19
Docking Server |
| Chloroquine
(37) | Accessory proteins
prevents binding of orf1ab, ORF3a, ORF10 to heme, ORF8, surface
glycoproteins to porphyrins | Simulated molecular
docking of viral proteins with human heme or porphyrins with
(LibDock tool) of Discovery-Studio 2016 |
| Favipiravir
(37) | Accessory proteins:
inhibits binding of E protein, ORF7a to porphyrin, prevent virus
entering host cells | Simulated molecular
docking of viral proteins with human heme or porphyrins with
(LibDock tool) of Discovery-Studio 2016 |
Among COVID-19 proposed drugs already approved for
other pathologies, GR 127935, a known potent entry inhibitor of
Ebola and Marburg viruses, binds the SARS-CoV-2 receptor ACE2 and
also is a selective 5-HT1B/1D receptor antagonist, making it useful
in patients with hypotension and tachyarrhythmia, when
administration of vasopressors is questionable.
Agents that bind the SARS-CoV-2 spike protein and
are known to reduce lung inflammation, include GSK1838705A,
BMS195614, GSK1838705A, which inhibit the insulin like growth
factor-1 receptor and are used in cancer.
For blocking the ACE2 receptor, three FDA-approved
drugs are proposed: TNP, an inhibitor of tyrosine kinase, IP6K and
Akt pathways, which can also shown to inhibit MERS infection;
eptifibatide acetate, an inhibitor of platelet aggregation already
tested in septic shock; and the Abelson (Abl) kinase inhibitor
GNF5, known to inhibit replication of Dengue virus, which deserves
further attention. It was shown that Abl2, but not Abl1 is
necessary for replication of SARS/MERS CoV (66) as the virion and endosomal
membranes fuse via a cathepsin L-dependent mechanism; the Abl2
kinase inhibitors GNF5/GNF2/imatinib inhibit the fusion of the
viral and endosomal membrane, an early stage of viral infection.
GNF5/GNF2/imatinib also inhibit the formation of cell syncitia
before the hemifusion step, effectively blocking membrane fusion
and viral entry (67).
Finally, 3 drugs deserve special attention for
COVID-19 treatment: KT185 and KT203 which bind the S protein and
are potent, selective inhibitors of a/b-hydrolase domain containing
6 (ABHD6), a transmembrane serine which hydrolases the endogenous
cannabinoid 2-arachidonoylglycerol (2-AG), and more importantly
decrease macrophage activation.
The third drug is probably the best viral fusion
inhibitor, RS 504393, which can bind both the SARS-CoV-2 S protein
and its receptor ACE2 and is used for treating lung injury and
bronchial wall thickening; moreover, it is also a selective
antagonist of the monocyte chemoattractant protein-1 (MCP-1)
receptor CCR2, blocks the upregulation of pronociceptive,
pro-inflammatory interleukins IL-1β, IL-18, IL-6, and as such are
very useful for decreasing the 'cytokine storm' and the associated
neuropathic pain (patients hypersensitive to touch).
Besides FDA-approved drugs, a variety of natural
compounds have been investigated for their antiviral actions on
SARS-CoV2, one advantage being that some plants contain more than
one active ingredient (65); in
the case of Nigella sativa its main active substances,
nigellidine and α-Hederin compare positively to either chloroquine
or hydroxychloroquine based on their binding energy with viral
proteases (68), more information
is given in Table III.
| Table IIINatural compounds with antiviral
actions. |
Table III
Natural compounds with antiviral
actions.
| Substance analyzed
(Ref.) | Viral molecule
inhibited; mechanism | Method and/or
software used for testing |
|---|
| Anisotine and
vasicoline of Justicia adhatoda (65) | Protease; RNA
Replicase inhibitor | Using COVID-19
Docking Server |
| Phycocyanobilin,
Riboflavin, Cyanidin, Daidzein, Genistein (69) | Protease (Mpro);
RNA Replicase inhibitors | COVID-19 Docking
Server to inhibit Mpro and RdRp |
| Curcumin (70) | Protease
inhibitor | Structure of
metabolite and COVID_19 protease from PubChem and Protein Data Bank
(PDB); molecular docking by MVD (Molegro Virtual Docker) |
| For 6LU7:
Nigellidine -6.29 Kcal/mol; chloroquine -6.29; OH-chloroquine
-5.57; favipiravir -4.23; For 2GTB: α-Hederin -6.50 kcal/mol;
chloroquine -6.20; OH-chloroquine -5.51; favipiravir -4.12
(68) | Proteases (3CLpro
andMpro) 6LU7 and 2GTB inhibitors | Docking of
compounds from Nigella sativa and drugs performed with
Molecular Operating Environment software (MOE) |
Local treatment and IFNs. Based on the
observation that the host IFN response is delayed in CoV
infections, with subsequent over-activation of neutrophils in the
lung and vascular leakage (54),
IFN-stimulating substances administered by inhalation showed
efficacy against SARS and MERS in vitro and in vivo.
Inhaled substances have the advantage of lower volume of
distribution (ASL has a volume of 20-30 ml) and can act early in
COV infection during the viral fusion process, inhibiting the
binding of its S protein to the host receptor (71).
One such substance is polyI:C, a TLR3 agonist shown
to induce differentiation of mDC, promote Th1 activation with IL-12
and type I IFN production, the activation of the innate immune
response and reversing the PGD2 effects in the lungs (72,73). A shortcoming of the polyI:C may be
its requirement for either RIG-1 or MDA5 activity, which are known
to be inactivated by coronaviruses and render polyI:C ineffective
post-infection, limiting it to a prevention drug (74).
A very potent inhibitor of CoV fusion is
nona-L-arginine with high affinity for furin (effective at
concentrations of 40 nM); hexa- and hepta-peptides of the basic
aminoacids arginine and lysine also showed excellent inhibition of
viral fusion (75).
Another intranasal-administered peptide, EK1, which
contains the heptad-repeat HR2 from the binding motif of the S
protein has inhibited infection by many human coronaviruses
(46); its administration in mice
reduced lung viral titers by 1,000-fold 2014 (76) and its lipidation, EK1C4, resulted
in much greater potency, inhibiting the cell-cell fusion mediated
by S protein with IC50 of 4.3 nM; its protective effect
manifested half an hour after and 6 hours before exposure to
coronavirus (46).
IFN can also be administered directly to patients
via inhalations, however, affinity amino acids have showed limited
benefits depending on the timing of administration and prompts
further analysis of IFN actions.
Similarly to the neutrophilia and lymphopenia
observed in severe COVID-19 patients, fatal SARS infection is
associated with high IFN and ISGs expression and low T cell and
antibody activity, while viral clearance needs increased T cell
responses (25).
The immune response of CoV patients is imbalanced,
with over-stimulation of the monocyte-macrophage line and decreased
T cell activation; while early administration of IFN-1 has
protective effects, in later stages administration of IFN, with the
possible exception of IFN-γ, is associated with increased
neutrophil infiltration in the lungs. This is due to the selective
activation of ISGs in macrophages, but not in B or T cells, since
signaling in these cells requires the myeloid differentiation
factor 88 (MyD88), a key molecule used in all TLR signaling
pathways of adaptive immunity except for TLR3 which is present in
all endosomes (77).
In animal models of lethal COV infection, the
immunopathological events are mostly IFN-I-dependent and
independent of viral replication; IFN-α/β receptor (IFNAR) receptor
ablation or neutrophil depletion avoided lethal infection but did
not affect the viral load. TNF inhibition improved lymphopenia,
which is not unexpected since IFN-I sensitizes T cells to
apoptosis.
It was shown that an important role in modulating
the inflammatory response is played by two proteins containing the
Toll or interleukin (IL)-1 receptor (TIR): MyD88 and the TIR
domain-containing adaptor-inducing interferon (TRIF) (78). For controlling inflammation both
MyD88 and TRIF proteins are required, since the release of the
anti-inflammatory IL10, IL19, endothelial lipase, metalloproteinase
inhibitor 1 (Timp1) and plasminogen activator inhibitor 1 (Serpin1)
is prevented in the absence of either one, resulting in excessive
pro-inflammatory modulation.
During early CoV replication in non-lethal models,
pDCs are the major IFN-I producers, and pDCs sense coronaviral RNA
via TLR7 within endosomes (54).
The same author shows that the coronavirus endonuclease (EndoU)
from nsp 15 has a key role in preventing early induction of innate
responses, by removing the viral dsRNA that would otherwise trigger
host cell dsRNA responses, including IFN, PKR and OAS/RNase L, and
delaying the viral RNA cytoplasmic sensing mediated by MDA5.
During early stages of SARS infection, IFN-γ
presence is associated with viral clearance and infection
resolution; IFN-γ is synthesized mainly by activated T cells and
natural killer (NK) cells (79).
IFN-γ (type II) and IFN-α/β (type I) have synergistic actions;
given alone neither inhibits viral replication, but administration
of both inhibited viral replication by 3000-fold at 24 h, with
stronger inhibition at 48 and 72 h.
Oligonucleotide arrays were used to study the ISGs
regulated by IFNs; all IFNs induced complement component C1r,
IFN-induced nuclear phosphoprotein, vascular endothelial growth
factor (VEGF)-related protein (VRP or VEGF-C), and phospholipid
scramblase, an apoptosis promoter; its levels increased 8-fold by
IFN-α, 10-fold by IFN-β, and 3-fold by IFN-γ (80). Additionally each IFN
preferentially induced specific molecules: IFN-α, somatic
cytochrome c; hepatitis C-associated p44; pyridoxal kinase;
and lysosomal acid lipase (LIPA); IFN-β, clathrin-like protein,
PKR, hypoxia-inducible factor-1 (HIF-1α) which regulates apoptosis
and cell response to hypoxia via expression of VEGF, p53, p21;
IFN-γ, mitochondrial 3-ketoacyl-CoA thiolase; poly (ADP-ribose)
polymerase; proton-ATPase-like protein.
Both type I and type II IFNs increase expression of
the CH25H gene which codes for the enzyme
Cholesterol-25-hydroxylase, which converts cholesterol into
25-hydroxycholesterol (25HC) (50), an oxysterol with antiviral effects
via alteration of membrane properties, inhibition of sterol
biosynthesis and prenylation (priming) of virus and host proteins.
However, 25HC production by macrophages also has a central role in
regulating the immune system, and at nM concentrations it
suppresses the IL-2 mediated proliferation of B cells and inhibits
IgA production (81).
Another possible pathological modification induced
by the CoV in susceptible patients is the activation of
alternatively activated macrophages, M2, which normally have
anti-inflammatory actions and are involved in the wound-healing
processes (82), but were also
found in the lungs of mice lacking STAT1 signaling pathway
(STAT1 knockouts), which after the SARS-CoV infection, had
an increased number of M2 macrophages in their lungs, activated via
the STAT6 pathway, and developed pulmonary fibrosis (83).
The modifications induced by the SARS-Cov infection
in host cells were also investigated via a library screen of small
interfering RNA (siRNA) targeting the human kinome (84). Besides the expected activation of
pro-inflammatory pathways (production of NO and ROS in macrophages,
IL-2, IL-6, IL-8 and IL-17) there was also an increase in the
metabolic pathways of complex lipids (inositol phosphate,
nicotinate, nicotinamide), intermediates needed in SARS-CoV
infection and probably linked to prenylation. The strongest
proviral activation was seen in the β2 subunit of the coatomer
protein complex (COPB2) followed by inositol hexaphosphate kinase 1
(IHPK1); abelson kinase 1 (ABI1); diacylglycerol kinase epsilon
(DGKE); cyclin-dependent kinase 5 regulator (CDK5R2); protein
kinase C ι (PRKCi). Among antiviral molecules were found the
diacylglycerol kinase δ (DGKD), the double-stranded RNA-activated
protein kinase (PKR); cyclin-dependent kinase 6 (CDK6) and most
MAPKs with the exception of MAP3K11, which is weakly proviral; ABl1
inhibitors and molecules acting on lipidic and glucidic metabolism
are thus important.
Another factor influencing IFN actions is the
hemoglobin β-chain which was shown to stimulate IFN-β production
via RIG-1, including potentiating of RIG-I ubiquitination;
hemoglobin is present in lung, kidney, peripheral blood mononuclear
and other cell lines (38,85),
and SARS-CoV2 nsps can bind to and degrade hemoglobin (37).
Indoleamine 2,3-dioxygenase (IDO), stimulated by
IFN-γ and inhibited by TLR activation and allergens plays an
important role in immune suppression (normally prevents
hyper-inflammation and autoimmunity) by inhibiting T cell
activation and polyclonal proliferation of T cells, mostly through
degradation of tryptophan and kynurenines; its role in SARS-CoV2
infection was not explored but likely is important (86). Activation of IDO is strictly
dependent on prior expression of Arginase 1 and the resulting
polyamines, with immunosuppressive actions (87); while NO is an IDO inhibitor; the
role of IDO in the SARS-CoV2 infection and its immune evasion has
not yet been explored.
The data above confirm that in severe CoV infection
the host immune system is mounting an imbalanced response to
infection which powerfully favors neutrophils/macrophages and
inhibits the adaptive cellular response; early events include
hindered pDCs activities and over-activation of macrophages with
induction of IFN-I expression and autocrine over-stimulation; this
may be due to 25-hydroxycholesterol or IDO overactivation, Myd88
inactivation by the virus, hemoglobin degradation and/or
simultaneous actions of other viral molecules (EndoU).
However, it is possible that the delayed IFN
response followed by neutrophil over-activation is simply due to
the lack of activation in the early stage of infection of the
peroxisomal MAVS necessary for early IFN production (52), and this observation gains more
weight as peroxisome function declines significantly in tandem with
cellular senescence, advancing age, and age-related co-morbidities
(88). Peroxisome senescence is
associated with dramatic changes in their protein import
capabilities, functional integrity, ability to process ROS,
regulation of growth and division of other organelles, decreased
ability to oxidize fatty acids, accumulation of free fatty acids
and diacylglycerol, followed by reorganization of protein kinase C
(PKC) signalling (89).
This fact is related to the important metabolic
functions of peroxisomes, which have >50 enzymes in their matrix
performing critical metabolic functions besides production and
degradation of H2O2 including fatty acid
oxidation, synthesis of specialized lipids, including
anti-inflammatory docohexaenoic acid and degradation of
pro-inflammatory leukotrienes and recently it was shown that
multiple viruses interfere with antiviral signaling dependent on
peroxisomes, examples being the accessory protein Vpu of HIV-1
which annihilates peroxisomes (90), the VP16 protein of HSV-1 which
blocks the peroxisomal MAVS (91), also the hepatitis C virus protein
NS3-4A, the capsid of the Dengue and West Nile virus, and the vMIA
protein of cytomegalovirus (92).
Peroxisomal MAVS stimulates ISGs via IRF1 and IRF3
in an IFN-independent manner and does not induce IFN production in
macrophages, which is dependent on mitochondrial MAVS.
Interestingly, cells lacking either peroxisomal or mitochondrial
MAVS had an unusual biphasic behavior, restricting viral
replication in the first 24 h, but not at 72 h, and showing that
both organelles are necessary for viral clearance (53). The functional status of
peroxisomes can be correlated both with age and the severity of
SARS-Cov2 infection, and is impaired by poor oxidative status, low
cellular pH (acidosis) and/or hypoxia which is also seen in
patients with chronic pathologies or genetic deficiencies.
pH-dependent viral actions. At the surface of
the respiratory epithelium low pH favors viral entry and
replication by impairing the actions of the AMPs from the airway
surface liquid (ASL), which have pH-sensitive antimicrobial
functions, with the pH of ASL being actively regulated by
bicarbonate transport (49);
another pH-lowering viral activity is the formation of cell
syncytiums. Lower intracellular pH favors the activity of the
proteases needed for viral replication, decreases the mitochondrial
membrane potential and inactivates MAVS, which is essential for IFN
and NFκB activation and production of antiviral protein switches.
In addition, low pH promotes oxidative phosphorylation to anaerobic
glycolysis with increased mitochondrial ROS production via related
cellular pathways (93).
The modifications in the cellular redox state, ionic
balance and pH which are simultaneous with the actions of specific
molecules in a pathway are not often measured or considered during
lab experiments, but in the context of dual or multiple regulators
they may have important consequences, one such example being the
effect of hemoglobin (Hb) on IFN production (85). While exerting a direct inhibitory
effect on MDA5 which inhibits IFN production, Hb also modifies a
cell redox state, promotes the RIG-I pathway and stimulates IFN
production; the increase in ROS by Hb is stimulated only by the
microbial and not by the host protease.
Additionally, the infectivity of CoV and other
enveloped viruses is known to be impaired by an increase in
endosomal pH in a dose-dependent manner; mild bases such as
lipophilic amines or amphiphilic drugs with a tertiary amino group
can accumulate in acidic organelles via protonation, where they
neutralize internal pH, alter the properties of the adjacent
membranes and inhibit multiple processes occurring in the
respective compartment (94);
this was also observed with administration of bafilomycin or
ammonium chloride (NH4Cl) in cell cultures at 5 mM concentrations
(58,72,95). In case of CoV the elevated
endosomal pH blocks the cysteine proteases cathepsin B and L
(CatB/L) (45) and inhibits the S
protein entry into host cells. Furthermore, NH4Cl and chloroquine
(also a weak base) impair the glycosylation of ACE2, an additional
mechanism for blocking viral fusion.
The pH-dependent fusion of various viruses
endocytosed into host cells by non-coated vesicles (caveolas) or in
association with clathrin is well-documented (96); additionally CoV and influenza
viruses can fuse with host cell membranes via anchoring cell
receptors and internalization in the lipid bi-layer, a process
blocked by constituent hydrolaze CH25H, methyl-β-cyclodextrin, a
cholesterol-sequestering drug, simvastatin and possibly other
lipophilic statins.
Low pH is affecting essential aspects of the native
immune response - neutrophil activation, chemotaxis, production of
ROS, NO and phagocytosis, apoptosis, cytokine production,
activation of transcription factors by binding to DNA, and also
decrease of lymphocyte function, lymphokine secretion and
activation, while hypoxia is potentiating these actions (97-101); acidosis is associated with
increased need for intensive care and hospitalization (102). In addition, a low extra- and
intra-cellular pH not only favors viral infection, but also impedes
the antiviral action of known medication.
pH modifies the activity of antiviral
medications. Many proteins undergo conformational changes of
their tertiary and quaternary structure induced by ionic changes
(Ca2+, H+, Fe2+ and
Mn+) and pH modifications. Such modifications are
documented for the main protease (Mpro) of SARS-CoV2 during its
binding to protease inhibitors at pH 4.0-8.0, and it was shown that
they were pH-dependent, with different binding energy (stability of
protein complex) and specific bonding type/atom interaction
(103).
Small modifications in pH can have critical
consequences as was shown with arbidol, a broad-spectrum antiviral
which binds to the influenza virus haemagglutinin (HA), induce a
0.2 unit pH modification and blocks viral fusion; consequently some
influenza strains developed a HA which disrupted the drug-HA bond
with loss of medication effectiveness (104).
More aspects of pH modifications in viral infections
are discussed below; re-establishing physiological pH levels
(7.35-7.45) is an important therapeutic consideration in
ALI/ARDS.
Genetic and translational factors involved in
severe COVID-19. An important question is whether the host
genetic factors significantly impact the immune functions and the
host answer to viral infection, as it was found that individual
variations exist in expression of RIG-1 and other antiviral
molecules (57).
Studying the bronchoalveolar lavage fluids from ARDS
patients by proteomic analysis during the initial 4 days of care,
survivors and non-survivors showed differences: non-survivors had
decreased expression of proteins involved in immune activation,
coagulation, iron metabolism, and increased protein expression of
glycolysis and collagen pathways (105).
Also, an association was found between four SNPs and
ARDS patients: rs78142040 of ARSD - an arylsulfatase gene involved
in sphingolipid metabolism and associated with 60-day mortality;
rs9605146 in XKR3 gene affecting membrane transport and red cells,
neuromuscular and central nervous systems, was associated with
susceptibility to ARDS; rs3848719 of ZNF335, zinc finger protein
regulating lymphoblast proliferation, was associated with increased
ARDS severity (APACHE II score); nicotinamide
phosphoribosyltransferase (NAMPT) with the -1535T allele associated
with decreased susceptibility to ARDS, and the -1001G variant with
higher susceptibility to ARDS and mortality in a Caucasian
population; other pathways showing differential activation in ARDS
patients were T lymphocyte signalling, calcium-induced apoptosis of
T lymphocytes, B-cell development, and autoimmune thyroid disease
(106).
Other genes associated with the pulmonary injury
type of ARDS are POPDC3 (expressed in cardiac and skeletal muscle,
role in glioma and muscular dystrophy), PDE4B (phosphodiesterase),
ABCC1 (organic anion transporter of oxidized glutatione, cysteinyl
leukotrienes and antiviral drugs), and TNFRS11 (TNF receptor
superfamily member 11) the rs1190286 allele of POPDC3 is associated
with decreased ARDS risk (107).
Using bioinformatic analysis for genetic
associations with ARDS, a total of 201 genes were found
predominantly involved in pathways modulating inflammation,
especially linked to innate immunity, reactive oxygen species, and
endothelial vascular signaling (108); it is worth noting that all these
pathways are modulated by NOS.
ACE2, RAS and ACE inhibitors. First genetic
markers analyzed as risk factors for ARDS pathogenesis were found
in 2002 to be correlated with ACE polymorphism, with the D allele
increasing its actions (109).
The frequency of the DD genotype was significantly increased in the
ARDS group, suggesting a role for the renin-angiotensin system in
ARDS development. A 2004 study in a small Vietnamese population
with SARS found an increase frequency of D allele in patients with
hypoxia (n=22) versus no hypoxia (n=22) (110). However, a larger study (n=168)
evaluated the influence of ACE I/D polymorphism on SARS-CoV
infection susceptibility and development of ARDS in a Chinese
population and found no association between the frequency of the D
allele and presence of infection or severe disease; the only
factors associated with requirement of intensive care was age and
male sex (111).
Measuring serial ACE levels in a small number of
ARDS patients (n=36) revealed that during the first 24 h the ACE
levels were normal or decreased, with no difference between septic
and aseptic ARDS, and after an initial decrease during the first 96
h ACE levels normalized by day 7; persisting low ACE levels tended
to be associated with ARDS aggravation and fibrosis (112).
ACE2 is the main host receptor used by SARS-CoV2,
and together with its homologue ACE, also a non-specific protease,
modulate the activity of RAS, with ACE2 inactivating angiotensin II
(AngII). In animal models, administration of recombinant ACE2
protects mice from lung injury and ACE-deficient mice had much
better outcome in acute lung injury (113). Also in animal models with acute
inflammation it was shown that the renin angiotensin system (RAS)
regulates neutrophil influx and pre-treatment of mice with the RAS
inhibitor enalapril and losartan decreased lung neutrophil
infiltration, concomitant with bradykinin increase and angiotensin
II decrease; agonists of bradykinin receptor also inhibited
neutrophil chemotaxis induced by IL-8 and pulmonary recruitment of
neutrophils (114).
Angiotensin II activates NF-κB in monocytes with
proinflammatory effects, and RAS inhibitors also decrease NF-κB
activation and neutrophil chemotaxis (115). Moreover, angiotensin converting
enzyme (ACE) affects pulmonary vascular tone and permeability,
epithelial cell apoptosis, and lung fibrosis via fibroblast
activation so that RAS inhibitors may be beneficial in ARDS.
ACE2 inactivates AngII and was shown to protect mice
from sepsis-induced ALI, by downregulating RAS which is involved in
pulmonary hypertension and pulmonary fibrosis; ACE2 expression
increases after IFN-I administration. SARS-CoV infection produces a
rapid downregulation of ACE2 expression at cell surface, ACE2
protein levels and RNA expression (83,116).
Another observation is that ACE inhibitors increase
levels of the peptide Ang-(1-7),
which inhibits allergic inflammation by suppressing ERK- and
NF-κB-dependent pathways (117,118).
Translating data from animal models to clinical
practice shows sometimes important differences, and we have
contradicting results in clinical trials involving RAS and ARDS. A
South Korean study with 182 patients (115) showed that mechanically
ventilated patients on RAS inhibitors had better survival rates at
30 days than patients not receiving them; however at 90 days there
was no significant difference in mortality between the 2 groups,
and patients on RAS inhibitor required longer ICU stay and
mechanical ventilation.
More studies are needed to inform decisions on the
use of RAS inhibitors in CoV infected patients; we should note that
ACE inhibitors increase bradykinin levels, responsible for the
associated cough and angio-edema and also a potent vasodepressor
(119), restricting their use in
patients with COVID-19 ARDS and hypotension. The risk for
bradykinin-mediated adverse reactions was found to be linked to the
levels of aminopeptidase P, which catabolizes bradykinin and was
significantly lower in hypertensive patients with a history of
angio-oedema during ACE inhibitor treatment (120); the aminopeptidase N is also a
receptor for some strains of coronaviruses (121).
3. The combination of
arginine-ascorbate
There is ample data suggesting that the severity of
the COVID-19 is less related to the viral replication itself than
to the host responses to the infection: delayed IFN production,
increased neutrophils and cytokines in the lung creating a
pro-inflammatory, pro-apoptotic milieu, combined with lymphopenia,
acidosis, coagulation and vascular endothelium modifications and
defective tissue repair with fibrosis in the lungs. Most treatments
are focused on blocking viral replication, however, in fatal cases
irreversible changes and deterioration occur even though viral
replication is essentially blocked. Therapeutic interventions which
combine agents for pleiotropic actions rather than single agents
acting on well-defined pathways are more likely to improve patient
outcomes.
Analyzing genes with modified expression in ARDS
evidenced 201 genes, predominantly from pathways modulating
inflammation, innate immunity, reactive oxygen species, and
endothelial vascular signaling (108); all these pathways are modulated
by NO generated from arginine, the only physiological substrate for
NO (122).
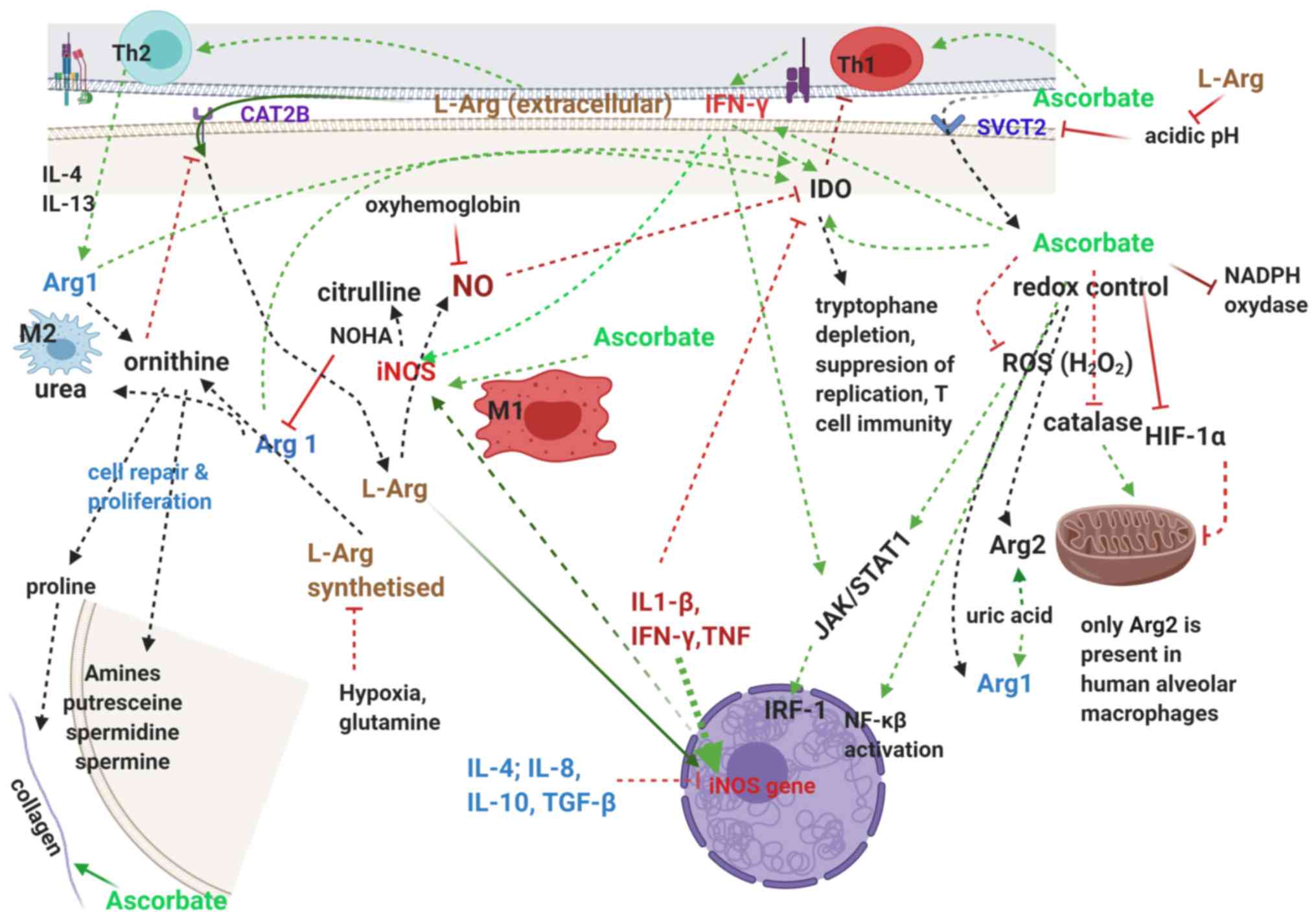
Arginine-ascorbate has several important benefits in
CoV infection: firstly, direct antiviral actions, secondly,
improvement of leukocyte function and number, especially important
in patients with neutrophilia and lymphopenia and finally it
provides essential components and stimulates the mechanisms of
tissue repair.
a) Antiviral actions. During the 2003 SARS
epidemic medicinal NO gas, a mixture of 0.8% NO and 99.2%
N2 was administered by Keyaerts et al (123) and followed by prompt improvement
in oxygenation and sustained patient benefit; following-up on this
observation it was shown that NO donor compounds inhibited SARS-CoV
replication in vitro, a finding replicated by another team
who also observed that besides S-nitroso-N-acetylpenicillamine
(SNAP), IFN also inhibited SARS-CoV replication in vitro
(124), confirming results
obtained by another team who showed that IFN stimulates iNOS and NO
production for prompt antiviral action (125).
Interestingly, IFN-γ stimulates NO production, and
inhibition of NOS in mice resulted in conversion of a resolving
infection by the ectromelia virus into fulminant mousepox (126).
NO production from arginine by the inducible NO
synthase (iNOS) was shown to be essential for native immune
responses in many infections and arginine availability is a
critical factor for host resistance to infection (122). NO can pass through membranes,
unlike complement and antibody, and is especially useful in
syncytia, it can act on multiple targets increasing efficacy and
preventing developing of resistance, and do not require recognition
of infected cell by the immune system, which can be limited by
pathogens or tissue-specific differences (127).
Reducing arginine availability either by nutrient
deprivation or by specific pathogens significantly blunts the
immune response, the former via impaired TLR4/MAPK pathway
signalling, the latter by inducing arginase 1 via other TLRs in
macrophages in an autocrine-paracrine manner involving cytokines,
an immune evasion mechanism shown with mycobacteria (122) and probably also in SARS-CoV2
infection.
Arginine also directly stimulates
transcription/translation of the iNOS gene leading to de
novo NOS protein synthesis, and this sheds light on the
'arginine paradox' where intracellular NO production is directly
related to the extracellular arginine concentration even though
endogenous synthesis of arginine provides approximately half of the
arginine needed by cells (128).
To ensure the increased need for intracellular arginine, the
cationic amino acid transporter CAT2B is induced by IFN-γ, which is
stimulated by ascorbate (129).
Reciprocally, the pH buffering action of arginine is increasing the
cellular uptake of ascorbate by improving the activity of the
cellular transporter of ascorbate, SVCT2, which has a pH optimum of
7.5 and is reduced to ~50% when pH is 5.5 (130).
Ascorbate is actively accumulated in phagocytic
cells where its concentration is 70-100 times higher than in
plasma; it enhances chemotaxis, ROS generation, phagocytosis and
viral clearance, important for minimising necrosis and tissue
damage (131).
Ascorbate stimulates the synthesis and actions of
antiviral NO indirectly in multiple ways, one via IFN-γ stimulation
(132), another via inhibition
of HIF-1α and correction of hypoxia, which significantly inhibits
the availability of intracellular arginine and NO production
(133); and also by synergizing
with the apoptotic effects of NO (134) in virus-infected cells, and thus
inhibiting viral replication. It is also important to note that all
three of NOS as well as IDO (135) are heme-containing proteins which
require ascorbate or another intracellular reducing agent for their
activation. In this context, the observation that three accessory
proteins of SARS-CoV2 - orf1ab, ORF3a, ORF10 - bind to heme
followed by Fe inactivation and sequestration (37) deserve to be further investigated
for specific anti-NOS and other IDO activities.
In the case of SARS-CoV2 infection which directly
antagonizes NO production, arginine provides both the substrate for
iNOS and activates its synthesis; together with ascorbate, which is
necessary for iNOS activation, it prepares a new functional enzyme
necessary for NO production.
b) Improving lymphocyte function and number.
Lymphopenia is present in a significant number of severe COVID-19
patients (9) and its causes are
strongly linked to arginine and ascorbate deficiencies.
Arginine is known to improve lymphocyte-based
immunity (136). Arginine
depletion due to upregulation of Arg1 in myeloid-derived suppressor
cells induces T-cell anergy, decreased proliferation of T cells,
low expression of CD3 ζ-chain T-cell receptor, and impairments of
production of T cell cytokines and of upregulation of cyclin D3 and
cyclin-dependent kinase 4 (cdk4) (137). L-arginine is essential for
maturation of CD3+ and the proliferation of
CD8+ T lymphocytes (138). Arginine is also critical for
B-lymphocyte differentiation; in transgenic mice over-expressing
arginase in intestinal cells there was an impaired transition of
pro- to pre-B cells in the bone marrow with marked decrease in B
cell cellularity, serum IgM levels and number and size of Peyer's
patches; this was reversed by arginine supplementation (139).
Noting the essential role of arginine for immunity,
an arginine deficiency syndrome (ADS) was proposed, defined by
pathological increase in arginase, decreased NO production,
decreased arginine availability, abnormal T cell function including
loss of ζ-chain, and presence of one or more of: trauma, cancer,
chronic infection, liver necrosis, pulmonary hypertension;
treatment for ADS include L-arginine and the arginase inhibitors
N-hydroxy-arginine (NOHA) and COX-2 inhibitors.
Ascorbate is also important for adaptive immune
responses, starting with the observations that reducing
environments make immune responses more efficient (140); it increases the levels of IFN
produced by activated fibroblasts (141); improves neutrophil function by
preserving/restoring function of their myeloperoxidase; is
essential for lymphocyte development, stimulating the proliferation
of NK cells (142); and has
ample epigenetic effects through dioxygenases TET (ten-eleven
translocases) and Jumonji C (JmjC)-domain-containing histone
demethylases (131).
A comprehensive review of the actions of ascorbate
including those on the immune system (131) concludes that enhancement of B-
and T-cell differentiation and proliferation is likely due to its
effects on gene regulation; an epigenetic effect (DNA
demethylation) of ascorbate on the CD8 receptor gene was also
inferred in a series of lab experiments which concluded that
ascorbate is necessary and sufficient for the CD8a gene
transcriptional activation (140). A graphic representation of these
actions are given in Fig. 4.
Multiple authors noted that administration of
ascorbate in chronic granulomatous disease led to consistent
improvements (143,144); more recent research showed that
in human granulomas depletion of arginine by arginases from
macrophages was associated with T-cell suppression without
affecting their viability, via downregulation of T-cells receptors
(128).
Both arginine and ascorbate stimulate production of
NO, which inhibits IDO (135),
and its suppressive effects on lymphocyte proliferation; the
simultaneous action of arginine (NO donor) and ascorbate (catalase
inhibitor) is synergical for leukocytes function and especially
apoptosis (134), yet another
example of the arginine-ascorbate antimicrobial synergy.
Synergistic action of arginine-ascorbate for
inhibiting viral replication and achieving viral clearance is shown
by the combined effect of IFN-α (induced by NO) and IFN-γ (induced
by ascorbate) in inducing the antiviral response via ISGs, which is
much enhanced when compared to either one alone (79,145). This was also seen with IFN-β1
and IFN-γ (146) in improved
apoptosis, RNA degradation and inflammatory response; and also with
TNF-α and IFN-β which together induced a novel synergistic
antiviral state, highly distinct from that induced by either
cytokine alone and involved >850 novel host cell genes (147); this was necessary and sufficient
to completely block the replication and spread of myxoma virus in
human fibroblasts and to block the spread of vaccinia virus and
tanapox virus to neighboring cells.
Nitric oxide from arginine directly enhances the
stimulation of ascorbic acid on T cell proliferation, and this
interaction is especially important in T cell recovery after stem
cell transplantation (147),
where generation and proliferation of T cells is greatly improved
by ascorbate and blocking of NOS diminished this effect (142).
To simplify the arginine-ascorbate actions in the
immune response by the way of dichotomy, arginine is an essential
factor for the native immune response via NO production and
activation of IFN-α/β (type I) while ascorbate is critical for the
efficacy of the adaptive immunity via T cell proliferation and
function via IFN-γ (type II); their synergy ensures both decreased
viral replication and clearance.
c) Stimulation of tissue repair. It is known
for decades that L-arginine supplementation (3-10 g/day) speeds
wound healing and recovery after surgery (136); we also know that ascorbate is an
essential cofactor for collagen synthesis, meaning that both
arginine-ascorbate are required for tissue repair; additional
molecules and pathways are involved in the immune response in viral
infection and individual variability can also have an important
role.
In a first step for tissue repair arginine is
metabolized to ornithine by arginases (Arg1/2), resulting in both
proline (collagen synthesis) and biologically active amines
(putresceine, spermidine, spermine) essential for cell repair and
proliferation; the balance iNOS/Arg1 determines the pro- or
anti-inflammatory environment and intra-cellular use of arginine.
Depending on this, macrophages are polarised in M1
(proinflamamtory, cytotoxic, NO) and M2 (anti-inflammatory,
proliferative, Arg1), and excessive activity of either can create
pathologies; surprisingly the suppression of M2 activity in mice
resulted in reduced lung fibrotic lesions in SARS-CoV infection,
suggesting the virus produces a shift to M2 activities, despite the
high levels of inflammatory cytokines (82,127,148).
Arginine corrects the pH and its increased
extracellular concentration helps cause a shift towards M2, and is
the substrate for arginase, which is stimulated by uric acid.
Arginases are activated by Mn2+ in a pH-dependent
fashion, Arg1/2 have an alkaline optimum activity of 9.0-9.5. NOHA,
the arginase inhibitor, can also be oxidized to NO by many heme
proteins, including hemoglobin, peroxidase, cytochromes P-450, and
catalase, which is inhibited by ascorbate, which is another
mechanism by which ascorbate modulates redox and metabolic
reactions and NO production (133).
While NO donors (SNAP, molsidomine) or iNOS gene
transfer enhance wound collagen synthesis in a dose-dependent
manner, high levels of NO strongly impair collagen synthesis
probably due to inhibition of arginase.
Ascorbate helps reduce inflammation by limiting
expression of pro-inflammatory cytokines, reducing CRP in
individuals with CRP >1.0 mg/l (149); it scavenges ROS and reduces the
oxidative stress on mitochondria (150), improves intracellular pH via
decreasing lactate (151),
decreases cell apoptosis and tissue necrosis also by inhibiting
HIF-1α (152), which is
stimulated by IFN-I (80);
hypoxia by inhibiting arginine intracellular synthesis is a
deleterious factor for tissue regeneration.
To get a good perspective on the dose needed of
arginine-ascorbate, we note that the daily plasma flux of arginine
is about 22-25 g with 5-6 g from diet (153), the majority of arginine is
synthesized in the liver and kidney (15-20 g daily) from citrulline
in the urea cycle, ~10% is used for creatine synthesis and 1% for
NO generation (133).
Pharmacological doses of up to 30 g/day for 2 weeks administered
either intravenously or enteral in multiple clinical studies appear
to be well tolerated by healthy individuals and cancer patients
without major side effects; significant elevation in IGF1 was the
only modified biomarker (153).
Ascorbate is sufficient in normal situations in
doses of 50-250 mg daily for maintaining plasma levels >30
µg/ml, however, during stress and infections the need for
ascorbate intake exceeds 1,000 mg/day and its intestinal absorbtion
is optimal when administration is fractionated at ~250-350 mg per
dose or in extended release forms (calcium ascorbate and
slow-release micronised preparations) (154).
In our experience (155) an effective antiviral regimen is
the combination arginine+extended release vitamin C, in doses of
1,000 mg L-arginine and 250-500 mg calcium/sodium ascorbate
(Ester-C) or Cetebe (GSK) or other modified release vitamin C, at
least 3 times a day each.
We have found this treatment effective in many
types of viral infections, including common cold and flu (many of
which are caused by coronaviruses), herpes labialis, zona zoster,
and hepatitis C, including a chronic hepatitis C patient who had
significant viral loads after 2 treatments with
ribavirin+interferon, and undetectable at 5 years after intravenous
arginine-ascorbate.
Arginine-ascorbate in severely ill patients.
Metabolic acidosis is associated with hyper-inflammation in ARDS
patients, and administration of sodium bicarbonate for balancing pH
can be limited by electrolyte imbalance and/or cardiovascular
pathology. Arginine administration can improve pH via the buffering
action of NH+ groups and additionally can provide an
immunity boost to septic patients, however, the formation of NO
(nitric oxide), which may induce relaxation of the vascular smooth
muscle has raised concerns (156,157).
A very thorough review of arginine use in critical
care has analyzed both the mechanistic concern of possible
NO-induced vasodilatation, alongside other actions that arginine
has in the critically ill patient. This study has importantly
discussed the multiple clinical trials and reports of arginine use
in septic patients (158).
During critical illness there is a severe arginine deficit
following a 2/3 reduction in arginine synthesis and increased
degradation due to a 4-fold increase of arginase activity, so that
arginine becomes an essential need for supplementation (158). Decreased arginine is especially
deleterious for the immune system, which produce excessive myeloid
suppressor cells which induce a pro-inflammatory state and suppress
T lymphocytes, with reduced number of CD4+ cells which
are also functionally deficient, as they lack the ζ-chain of its
receptor.
Clinical trials found that arginine shortened
hospitalizations and decreased infections, and in severe sepsis
(159) improved outcomes and
decreased mortality; it also reversed septic shock without
hemodynamic instability (160).
A better assessment of septic shock patients can be
done by using the ratio of arginine and its metabolite asymmetric
dimethyl arginine (ADMA), which can block NO production and act as
vasoconstrictor; a declining arginine/ADMA ratio was associated
with increased mortality (161);
while an increasing arginine/ADMA ratio in septic patients improved
mortality (162,163). Also important is a report
showing that intravenous administration in critically ill patients
of supra-physiologic doses of arginine did not produce hemodynamic
instability (164); this may be
due to a preferential uptake in septic patients of arginine by
leukocytes compared to endothelial cells.
The review concludes that arginine is safe to
administer and beneficial in critically ill patients, contributing
to resolution of infection and improving morbidity and mortality
(158).
Adding ascorbate makes administration of arginine
even safer; in septic mice ascorbate administration prevented the
impaired vasoconstriction and angiotensin actions and excessive
vasodilation as much as the iNOS gene knockout, and greatly
increased survival (165),
seemingly by inhibiting excessive NO production.
An important fact is that ascorbate, which is known
to be decreased in septic patients, is a cofactor required for
adrenal synthesis of hormones and neurotransmitters including
cathecholamines (norepinephrine) and vasopressin (131), physiological vasopressors which
otherwise need to be administered.
In septic patients ascorbate was shown to decrease
mortality especially in the hyperinflammatory subphenotype
characterized by high leukocyte count (>15,000/ml) and fever
(>37.5°C) (150). A
meta-analysis including 1,210 patients with sepsis showed that
different daily doses of intravenous ascorbate were associated with
different results, and only a dose between 3-10 g/day resulted in
decreased mortality, decreased need for vasopressor and mechanical
ventilation, but not lower or higher doses (166).
In a prospective randomised placebo-controlled
double blind trial - CITRIS-ALI - ascorbate 50 mg/kg or placebo was
given q 6 h intravenously for 96 h to 167 patients with sepsis and
ARDS; no significant difference was seen at 96 h in organ
dysfunction scores (SOFA), inflammation marker CRP or
thrombomodulin (for vascular integrity) (167). However, patients treated with
vitamin C had a significant benefit in 28-day mortality, transfer
from ICU by hour 168, number of ICU-free days by day 28, and number
of hospital-free days.
Hence in ascorbate deficiency, it is likely that
when administered intravenously to hypotensive patients, ascorbate
should be administered first, followed by a lower dose of arginine
(ex 1500 mg ascorbate in 100-200 ml normal saline followed by 1000
mg L-arginine in 100 ml saline administered at 1 ml/min).
Other medication should be added depending on the
clinical and laboratory signs: for the ′ground glass′ image on
imagistics, which signals presence of intra-alveolar- and
intravascular emboli, especially when D-dimer is elevated:
administration of anticoagulant, heparin or fractionated heparin;
for signs of porphyria (spontaneous ecchymosis which do not change
color) - chloroquine, amodiaquine, curcumin; for tachycardic,
tachyarrhythmic patients amiodarone 400 mg qd for a maximum 7 days;
for all patients and especially with high cholesterol, simvastatin
for asthma patients, Montelukast and/or fexofenadine or other
lipophilic antihistaminic and anti- inflammatory, preferably
curcumin; for anti-inflammatory, analgesic actions - PPAR-α/γ
agonists fenofibrate/pioglitasone or ABDH6 inhibitors KT185 and
KT203 (which decrease macrophage activation and pain-inflicting
CCXl-2 with allodynia); rabeprazole for preventing gastritis and
for possible antiviral effects; methylprednisolone was shown to be
of benefit in COVID-19 (4), and
even with associated immune depression favoring viral replication,
it can be considered for short-term in severely ill.
Practical measures - prevention and early
interventions. A majority of SARS-CoV2 patients (85-90%) have
mild to moderate forms of disease, however 10-15% develop rapidly
deteriorating forms with approximately half of those dying, and it
is difficult to predict accurately the individual risk for
developing severe disease besides advanced age and existing
co-morbid conditions.
Unopposed or excessive viral replication is an
important pathogenic mechanism through which the virus overwhelms
the natural defenses of the body; besides a weakened immune
response associated with chronic pathologies (cardiovascular,
renal, hepatic, autoimmune and neoplasic), the virus has an
advantage in tired or stressed individuals (with high cortisol and
lactate/LDH levels) and by repeated exposure which increases the
dose of the virus (doctors, nurses, medical personnel). Protective
measures are very important for limiting exposure to the pathogen
in vulnerable populations and also testing and treating early, as
soon as possible after respiratory symptoms.
After infection and respiratory symptoms, prompt
local treatment is very important: aerosol inhalation, nasal drops,
throat sprays are very useful treatments. Water vapors hydrate the
airway mucosa, increase the volume of ASL and help the natural
clearance of the virus via mucus shedding. Adding to the inhaled
vapors natural antimicrobials such as rosemary, basil, tea tree,
mint, eucalyptus, thyme, onion, in fresh or dry form; throat or
nasal sprays with propolis, argentum, basil, thyme, cumin will also
help viral inactivation and decrease inflammation.
We propose arginine-ascorbate as the mainstay of
antiviral treatment for SARS-CoV2 and other coronaviruses,
administered from the initial signs of infection until its
resolution.
As much as NSAIDs (non-steroidal antiinflammatory
drugs) are not recommended (aspirin, ibuprofen, naproxene and
indomethacin), possibly because of concomitant inhibition of PGD2,
known to be elevated in elderly patients, and further elevated by
SARS-CoV infection and in more severe forms of COVID-19 (73); however PGD2 has protective,
anti-inflammatory actions, and blocking cyclooxygenase (COX) also
inhibits PGD2, which further imbalances an already malfunctioning
immune response.
Interestingly, NSAID-mediated curcumin (also a main
protease inhibitor) is useful for alleviating inflammation and
pain, and if rhinitis and conjunctive inflammation are present, an
antihistaminic can be of help (fexofenadine also inhibits the main
COVID-19 protease). We have successfully used the
curcumin/antihistaminic combination for pain of various types from
sore throat to allodynia, in patients with allergies, increased
PGD2, rheumatoid arthritis, and also for porphyria. Patients who
cannot tolerate curcumin may use instead extracts of Boswellia
serrata (400 mg tid) as anti-inflammatory and analgesic;
artemisinin can be a good substitute for chloroquine or amiodarone
in patients with prolonged QT interval, COPD or lung fibrosis.
Risk evaluation of CoV infected patients.
Assessing the risk for progression to severe forms early on during
CoV infection is important for prompt individual treatment and
medical resource management, and also to avoid imposing restriction
on the whole population, knowing that 90% of infections are
asymptomatic or can be treated outside hospital; this may allow
shifting isolation measures from whole populations to truly at-risk
populations (168).
A two-pronged strategy of risk evaluation can be
employed based on age and exposure risk: in younger individuals
(≤55 years old) a specific panel for genetic risk factors which
predispose to fatal CoV infections (rs78142040, rs9605146,
rs3848719; NAMPT - 1001G allele, POPDC3, PDE4B, ABCC1, TNFRS11 and
DD genotype of ACE) may be more useful, especially in individuals
with potential increased exposure to the virus (eg., healthcare
professionals). However, in patients most at-risk - elderly and
with chronic co-morbidities, evaluation with different testing
panels can reasonably assess their risk for severe COVID19,
starting with the evaluation of oxidative/peroxisomal status
(catalase, malondialdehyde), inflammation - CRP, fibrinogen - and
metabolism - bicarbonate, HgbA1c, lipid profile, and immune balance
- PGD2 and serum protein electrophoresis - for asymptomatic
patients, and adding for symptomatic patients the CD3+,
CD4+, CD8+ and NK counts (predictors of need
for intensive care), LD1 isoensyme, which were good predictors for
intensive care during the SARS epidemic (169); for hyper-inflammatory status
also useful are markers of neutrophil activation - MMP-9, LTB4,
ferritin, CXCl-2 (106).
Another approach to risk evaluation of severe COV
infection is measuring the high-sensitivity cardiac troponin
(hs-cTnT), shown with data from the ARIC study to be a marker for
end-organ damage in patients with chronic pathologies (170); elevated troponin levels were
also seen in some COVID-19 patients (9).
Our efforts as front-line defenders against this
invisible foe, SARS-CoV, and possible others to come, are essential
not only from a medical perspective but also towards fully
regaining our individual and societal freedoms; through risk
evaluation and prompt, effective treatments the burden of disease
can be greatly diminished for both individuals and whole
populations.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
FS conceived and designed the study, tables and
figures and drafted the manuscript. BNI, GZP, DAS, AT, MDC and SK
performed analysis of data, researched the literature and
critically revised the article for important intellectual content.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
All the other authors declare that they have no competing
interests.
Acknowledgements
The present study is dedicated to the memory of
Mihail Coculescu.
References
|
1
|
Wu Z and McGoogan JM: Characteristics of
and important lessons from the coronavirus disease 2019 (COVID-19)
outbreak in China: Summary of a report of 72 314 cases from the
Chinese Center for Disease Control and Prevention. JAMA.
323:1239–1242. 2020. View Article : Google Scholar
|
|
2
|
Yang X, Yu Y, Xu J, Shu H, Xia J, Liu H,
Wu Y, Zhang L, Yu Z, Fang M, et al: Clinical course and outcomes of
critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China:
A single-centered, retrospective, observational study. Lancet
Respir Med. 8:475–481. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li JY, You Z, Wang Q, Zhou ZJ, Qiu Y, Luo
R and Ge XY: The epidemic of 2019-novel-coronavirus (2019-nCoV)
pneumonia and insights for emerging infectious diseases in the
future. Microbes Infect. 22:80–85. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu C, Chen X, Cai Y, Xia J, Zhou X, Xu S,
Huang H, Li Zhang, L Zhou X, Du C, et al: Risk factors associated
with acute respiratory distress syndrome and death in patients with
coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern
Med. Mar 13–2020.2020 (Epub ahead of print). View Article : Google Scholar :
|
|
5
|
Grasselli G, Pesenti A and Cecconi M:
Critical care utilization for the COVID-19 outbreak in Lombardy,
Italy: Early experience and forecast during an emergency response.
JAMA. 323:15452020. View Article : Google Scholar
|
|
6
|
Goumenou M, Sarigiannis D, Tsatsakis A,
Anesti O, Docea AO, Petrakis D, Tsoukalas D, Kostoff R, Rakitskii
V, Spandidos DA, et al: COVID-19 in Northern Italy: An integrative
overview of factors possibly influencing the sharp increase of the
outbreak (Review). Mol Med Rep. 22:20–32. 2020.PubMed/NCBI
|
|
7
|
Petrakis D, Margină D, Tsarouhas K, Tekos
F, Stan M, Nikitovic D, Kouretas D, Spandidos DA and Tsatsakis A:
Obesity - a risk factor for increased COVID-19 prevalence, severity
and lethality (Review). Mol Med Rep. 22:9–19. 2020.PubMed/NCBI
|
|
8
|
Docea AO, Tsatsakis A, Albulescu D,
Cristea O, Zlatian O, Vinceti M, Moschos SA, Tsoukalas D, Goumenou
M, Drakoulis N, et al: A new threat from an old enemy: Re-emergence
of coronavirus (Review). Int J Mol Med. 45:1631–1643.
2020.PubMed/NCBI
|
|
9
|
Kluge S, Janssens U, Welte T,
Weber-Carstens S, Marx G and Karagiannidis C: German
recommendations for critically ill patients with COVID-19. Med Klin
Intensivmed Notfmed. Apr 14–2020.Epub ahead of print. View Article : Google Scholar
|
|
10
|
Bhatraju PK, Ghassemieh BJ, Nichols M, Kim
R, Jerome KR, Nalla AK, Greninger AL, Pipavath S, Wurfel MM, Evans
L, et al: Covid-19 in critically ill patients in the Seattle region
- case series. N Engl J Med. 382:2012–2022. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Farsalinos K, Niaura R, Le Houezec J,
Barbouni A, Tsatsakis A, Kouretas D, Vantarakis A and Poulas K:
Editorial: Nicotine and SARS-CoV-2: COVID-19 may be a disease of
the nicotinic cholinergic system. Toxicol Rep. 7:658–663. 2020.
View Article : Google Scholar :
|
|
12
|
Calfee CS, Delucchi K, Parsons PE,
Thompson BT and Ware LB: Subphenotypes in acute respiratory
distress syndrome: Latent class analysis of data from two
randomised controlled trials. Lancet Respir Med. 2:611–620. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wilson JG and Calfee CS: ARDS
subphenotypes: Understanding a heterogeneous syndrome. Crit Care.
24:1022020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Famous KR, Delucchi K, Ware LB, Kangelaris
KN, Liu KD and Thompson BT: Acute respiratory distress syndrome
subphenotypes respond differently to randomized fluid management
strategy. Am J Respir Crit Care Med. 195:331–338. 2017. View Article : Google Scholar :
|
|
15
|
Calfee CS, Delucchi KL, Sinha P, Matthay
MA, Hackett J, Shankar-Hari M, McDowell C, Laffey JG, O′Kane CM,
McAuley DF, et al Irish Critical Care Trials Group: Acute
respiratory distress syndrome subphenotypes and differential
response to simvastatin: Secondary analysis of a randomised
controlled trial. Lancet Respir Med. 6:691–698. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sinha P, Delucchi KL, Thompson BT, McAuley
DF and Matthay MA: Latent class analysis of ARDS subphenotypes: A
secondary analysis of the statins for acutely injured lungs from
sepsis (SAILS) study. Intensive Care Med. 44:1859–1869. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
de Wilde AH, Jochmans D, Posthuma CC,
Zevenhoven-Dobbe JC, van Nieuwkoop S, Bestebroer TM, van den Hoogen
BG, Neyts J and Snijder EJ: Screening of an FDA-approved compound
library identifies four small-molecule inhibitors of Middle East
respiratory syndrome coronavirus replication in cell culture.
Antimicrob Agents Chemother. 58:4875–4884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cascella M, Rajnik M, Cuomo A, Dulebohn SC
and Di Napoli R: Features, evaluation and treatment coronavirus
(COVID-19). StatPearls [Internet] Treasure Island (FL): StatPearls
Publishing; 2020 Jan. PubMed/NCBIhttps://www.ncbi.nlm.nih.gov/books/NBK554776.
|
|
19
|
Neagu M, Bostan M and Constantin C:
Protein microarray technology: Assisting personalized medicine in
oncology (Review). World Acad Sci J. 1:113–124. 2019.
|
|
20
|
Levitt JE and Rogers AJ: Proteomic study
of acute respiratory distress syndrome: Current knowledge and
implications for drug development. Expert Rev Proteomics.
13:457–469. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meyer NJ, Reilly JP, Anderson BJ,
Palakshappa JA, Jones TK, Dunn TG, Shashaty MGS, Feng R, Christie
JD and Opal SM: Mortality benefit of recombinant human
interleukin-1 receptor antagonist for sepsis varies by initial
interleukin-1 receptor antagonist plasma concentration. Crit Care
Med. 46:21–28. 2018. View Article : Google Scholar
|
|
22
|
Brandes M, Klauschen F, Kuchen S and
Germain RN: A systems analysis identifies a feed forward
inflammatory circuit leading to lethal influenza infection. Cell.
154:197–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bowen JR, Ferris MT and Suthar MS: Systems
biology: A tool for charting the antiviral landscape. Virus Res.
218:2–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kindler E, Thiel V and Weber F:
Interaction of SARS and MERS coronaviruses with the antiviral
interferon response. Adv Virus Res. 96:219–243. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Channappanavar R, Fehr AR, Vijay R, Mack
M, Zhao J, Meyerholz DK and Perlman S: Dysregulated type I
interferon and inflammatory monocyte-macrophage responses cause
lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe.
19:181–193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Baughman RP, Gunther KL, Rashkin MC,
Keeton DA and Pattishall EN: Changes in the inflammatory response
of the lung during acute respiratory distress syndrome: Prognostic
indicators. Am J Respir Crit Care Med. 154:76–81. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kast RE: Dapsone as treatment adjunct in
ARDS. Exp Lung Res. 46:157–161. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Coleman MD, Rhodes LE, Scott AK, Verbov
JL, Friedmann PS, Breckenridge AM and Park BK: The use of
cimetidine to reduce dapsone-dependent methaemoglobinaemia in
dermatitis herpetiformis patients. Br J Clin Pharmacol. 34:244–249.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meo SA, Alhowikan AM, Al-Khlaiwi T, Meo
IM, Halepoto DM, Iqbal M, Usmani AM, Hajjar W and Ahmed N: Novel
coronavirus 2019-nCoV: Prevalence, biological and clinical
characteristics comparison with SARS-CoV and MERS-CoV. Eur Rev Med
Pharmacol Sci. 24:2012–2019. 2020.PubMed/NCBI
|
|
30
|
Nitulescu GM, Paunescu H, Moschos SA,
Petrakis D, Nitulescu G, Ion GND, Spandidos DA, Nikolouzakis TK,
Drakoulis N and Tsatsakis A: Comprehensive analysis of drugs to
treat SARS-CoV-2 infection: Mechanistic insights into current
COVID-19 therapies (Review). Int J Mol Med. 46:467–488. 2020.
|
|
31
|
Tsatsakis A, Petrakis D, Nikolouzakis TK,
Docea AO, Calina D, Vinceti M, Goumenou M, Kostoff RN, Mamoulakis
C, Aschner M, et al: COVID-19, an opportunity to reevaluate the
correlation between long-term effects of anthropogenic pollutants
on viral epidemic/pandemic events and prevalence. Food Chem
Toxicol. 141:1114182020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu R, Zhao X, Li J, Niu P, Yang B, Wu H,
Wang W, Song H, Huang B, Zhu N, et al: Genomic characterisation and
epidemiology of 2019 novel coronavirus: Implications for virus
origins and receptor binding. Lancet. 395:565–574. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu A, Peng Y, Huang B, Ding X, Wang X, Niu
P, Meng J, Zhu Z, Zhang Z, Wang J, et al: Genome composition and
divergence of the novel coronavirus (2019-nCoV) originating in
China. Cell Host Microbe. 27:325–328. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sah R, Rodriguez-Morales AJ, Jha R, Chu
DKW, Gu H, Peiris M, Bastola A, Lal BK, Ojha HC, Rabaan AA, et al:
Complete genome sequence of a 2019 novel coronavirus (SARS-CoV-2)
strain isolated in Nepal. Microbiol Resour Announc. 9:92020.
View Article : Google Scholar
|
|
35
|
Cui J, Li F and Shi ZL: Origin and
evolution of pathogenic coronaviruses. Nat Rev Microbiol.
17:181–192. 2019. View Article : Google Scholar
|
|
36
|
Xu J, Zhao S, Teng T, Abdalla AE, Zhu W,
Xie L, Wang Y and Guo X: Systematic comparison of two
animal-to-human transmitted human coronaviruses: SARS-CoV-2 and
SARS-CoV. Viruses. 12:122020. View Article : Google Scholar
|
|
37
|
Liu W and Li H: COVID-19: Attacks the
1-beta chain of hemoglobin and captures the porphyrin to inhibit
human heme metabolism. ChemRxiv. 2020, Preprint. https://doi.org/10.26434/chemrxiv.11938173.v8.
|
|
38
|
Yang Q, Bai S-Y, Li L-F, Li S, Zhang Y,
Munir M and Qiu H-J: Human hemoglobin subunit beta functions as a
pleiotropic regulator of RIG-I/MDA5-mediated antiviral innate
immune responses. J Virol. 93:932019. View Article : Google Scholar
|
|
39
|
Li T, Lu H and Zhang W: Clinical
observation and management of COVID-19 patients. Emerg Microbes
Infect. 9:687–690. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wan Y, Shang J, Graham R, Baric RS and Li
F: Receptor recognition by the novel coronavirus from Wuhan: An
analysis based on decade-long structural studies of SARS
coronavirus. J Virol. 94:942020. View Article : Google Scholar
|
|
41
|
Xie M and Chen Q: Insight into 2019 novel
coronavirus - An updated interim review and lessons from SARS-CoV
and MERS-CoV. Int J Infect Dis. 94:119–124. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hwang SS, Lim J, Yu Z, Kong P, Sefik E, Xu
H, Harman CCD, Kim LK, Lee GR, Li HB and Flavell RA: mRNA
destabilization by BTG1 and BTG2 maintains T cell quiescence.
Science. 367:1255–1260. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Coutard B, Valle C, de Lamballerie X,
Canard B, Seidah NG and Decroly E: The spike glycoprotein of the
new coronavirus 2019-nCoV contains a furin-like cleavage site
absent in CoV of the same clade. Antiviral Res. 176:1047422020.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kawase M, Shirato K, van der Hoek L,
Taguchi F and Matsuyama S: Simultaneous treatment of human
bronchial epithelial cells with serine and cysteine protease
inhibitors prevents severe acute respiratory syndrome coronavirus
entry. J Virol. 86:6537–6545. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hoffmann M, Kleine-Weber H, Schroeder S,
Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH,
Nitsche A, et al: SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2
and is blocked by a clinically proven protease inhibitor. Cell.
181:271–280.e8. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xia S, Liu M, Wang C, Xu W, Lan Q, Feng S,
Qi F, Bao L, Du L, Liu S, et al: Inhibition of SARS-CoV-2
(previously 2019-nCoV) infection by a highly potent pan-coronavirus
fusion inhibitor targeting its spike protein that harbors a high
capacity to mediate membrane fusion. Cell Res. 30:343–355. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Millet JK and Whittaker GR: Physiological
and molecular triggers for SARS-CoV membrane fusion and entry into
host cells. Virology. 517:3–8. 2018. View Article : Google Scholar
|
|
48
|
Totura AL and Bavari S: Broad-spectrum
coronavirus antiviral drug discovery. Expert Opin Drug Discov.
14:397–412. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Amatngalim GD and Hiemstra PS: Airway
epithelial cell function and respiratory host defense in chronic
obstructive pulmonary disease. Chin Med J (Engl). 131:1099–1107.
2018. View Article : Google Scholar
|
|
50
|
Chiang JJ, Davis ME and Gack MU:
Regulation of RIG-I-like receptor signaling by host and viral
proteins. Cytokine Growth Factor Rev. 25:491–505. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Schneider WM, Chevillotte MD and Rice CM:
Interferonstimulated genes: A complex web of host defenses. Annu
Rev Immunol. 32:513–545. 2014. View Article : Google Scholar
|
|
52
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dixit E, Boulant S, Zhang Y, Lee AS,
Odendall C, Shum B, Hacohen N, Chen ZJ, Whelan SP, Fransen M, et
al: Peroxisomes are signaling platforms for antiviral innate
immunity. Cell. 141:668–681. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kindler E, Gil-Cruz C, Spanier J, Li Y,
Wilhelm J, Rabouw HH, Züst R, Hwang M, V'kovski P, Stalder H, et
al: Early endonuclease-mediated evasion of RNA sensing ensures
efficient coronavirus replication. PLoS Pathog. 13:e10061952017.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cao W, Bao C, Padalko E and Lowenstein CJ:
Acetylation of mitogen-activated protein kinase phosphatase-1
inhibits Toll-like receptor signaling. J Exp Med. 205:1491–1503.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhou Y, He C, Wang L and Ge B:
Post-translational regulation of antiviral innate signaling. Eur J
Immunol. 47:1414–1426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Quicke KM, Diamond MS and Suthar MS:
Negative regulators of the RIG-I-like receptor signaling pathway.
Eur J Immunol. 47:615–628. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Vincent MJ, Bergeron E, Benjannet S,
Erickson BR, Rollin PE, Ksiazek TG, Seidah NG and Nichol ST:
Chloroquine is a potent inhibitor of SARS coronavirus infection and
spread. Virol J. 2:692005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Samarth S and McGregor K: Energetics based
modeling of hydroxychloroquine and azithromycin binding to the
SARS-CoV-2 spike (S)protein - ACE2 complex. ChemRxiv. 2020,
Preprint. https://doi.org/10.26434/chemrxiv.12015792.v2.
|
|
60
|
Arya R, Das A, Prashar V and Kumar M:
Potential inhibitors against papain-like protease of novel
coronavirus (SARS-CoV-2) from FDA approved drugs. ChemrxivOrg. 1–8.
2020. View Article : Google Scholar
|
|
61
|
Dyall J, Gross R, Kindrachuk J, Johnson
RF, Olinger GG Jr, Hensley LE, Frieman MB and Jahrling PB: Middle
East respiratory syndrome and severe acute respiratory syndrome:
Current therapeutic options and potential targets for novel
therapies. Drugs. 77:1935–1966. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Choudhary S, Malik YS and Tomar S:
Identification of SARS-CoV-2 cell entry inhibitors by drug
repurposing using in silico structure-based virtual screening
approach. ChemRxiv. 2020. View Article : Google Scholar
|
|
63
|
Farag A, Wang P, Boys IN, Eitson J, Ohlson
MB, Fan W, McDougal MB, Ahmed M and Schoggins JW: Identification of
atovaquone, quabain and mebendazole as FDA approved drugs
Tar-geting SARS-CoV-2 (Version 4). 2020, ChemRxiv. Preprint
https://doi.org/10.26434/chemrxiv.12003930.v4.
|
|
64
|
Navan C: Possible Drug Candidates for
COVID-19. ChemRxiv. 2020, Preprint. https://doi.org/10.26434/chemrxiv.11985231.v1.
|
|
65
|
Bag A and Bag A: Treatment of COVID-19
patients: Justicia adhatoda leaves extract is a strong remedy for
COVID-19- Case report analysis and docking based study. ChemRxiv.
2020, Preprint. https://doi.org/10.26434/chemrxiv.12038604.v1.
|
|
66
|
Coleman CM, Sisk JM, Mingo RM, Nelson EA,
White JM and Frieman MB: Abelson kinase inhibitors are potent
inhibitors of severe acute respiratory syndrome coronavirus and
Middle East respiratory syndrome coronavirus fusion. J Virol.
90:8924–8933. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sisk JM, Frieman MB and Machamer CE:
Coronavirus S protein-induced fusion is blocked prior to hemifusion
by Abl kinase inhibitors. J Gen Virol. 99:619–630. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Salim B and Noureddine M: Identification
of compounds from Nigella sativa as new potential inhibitors of
2019 novel corona virus (Covid-19): Molecular docking study.
ChemRxiv. 2020, Preprint. https://doi.org/10.26434/chemrxiv.12055716.v1.
|
|
69
|
Pendyala B and Patras A: In silico
screening of food bioactive compounds to predict potential
inhibitors of COVID-19 main protease (Mpro) and RNA-dependent RNA
polymerase (RdRp). ChemRxiv Preprint. https://doi.org/10.26434/chemrxiv.12051927.v2.
|
|
70
|
Mohammadi N: Inhibitory effect of eight
secondary metabolites from conventional medicinal plants on
COVID-19 virus protease by molecular docking analysis. ChemRxiv.
2020, Preprint. https://doi.org/10.26434/chemrxiv.11987475.v1.
|
|
71
|
Bosch BJ, Martina BEE, Van Der Zee R,
Lepault J, Haijema BJ, Versluis C, Heck AJ, De Groot R, Osterhaus
AD and Rottier PJ: Severe acute respiratory syndrome coronavirus
(SARS-CoV) infection inhibition using spike protein heptad
repeat-derived peptides. Proc Natl Acad Sci USA. 101:8455–8460.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yang Z-Y, Huang Y, Ganesh L, Leung K, Kong
WP, Schwartz O, Subbarao K and Nabel GJ: pH-dependent entry of
severe acute respiratory syndrome coronavirus is mediated by the
spike glycoprotein and enhanced by dendritic cell transfer through
DC-SIGN. J Virol. 78:5642–5650. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhao J, Wohlford-Lenane C, Zhao J, Fleming
E, Lane TE, McCray PB Jr and Perlman S: Intranasal treatment with
poly(I•C) protects aged mice from lethal respiratory virus
infections. J Virol. 86:11416–11424. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mian MF, Ahmed AN, Rad M, Babaian A,
Bowdish D and Ashkar AA: Length of dsRNA (poly I:C) drives distinct
innate immune responses, depending on the cell type. J Leukoc Biol.
94:1025–1036. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Cameron A, Appel J, Houghten RA and
Lindberg I: Polyarginines are potent furin inhibitors. J Biol Chem.
275:36741–36749. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Channappanavar R, Lu L, Xia S, Du L,
Meyerholz DK, Perlman S and Jiang S: Protective effect of
intranasal regimens containing peptidic Middle East respiratory
syndrome coronavirus fusion inhibitor against MERS-CoV infection. -
Abstract - Europe PMC. https://europepmc.org/article/med/26164863.
|
|
77
|
Zhou H, Zhao J and Perlman S: Autocrine
interferon priming in macrophages but not dendritic cells results
in enhanced cytokine and chemokine production after coronavirus
infection. MBio. 1:12010. View Article : Google Scholar
|
|
78
|
Iwasaki A and Medzhitov R: Regulation of
adaptive immunity by the innate immune system. Science.
327:291–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sainz B Jr, Mossel EC, Peters CJ and Garry
RF: Interferon-beta and interferon-gamma synergistically inhibit
the replication of severe acute respiratory syndrome-associated
coronavirus (SARS-CoV). Virology. 329:11–17. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Der SD, Zhou A, Williams BRG and Silverman
RH: Identification of genes differentially regulated by interferon
α, β, or γ using oligonucleotide arrays. Proc Natl Acad Sci USA.
95:15623–15628. 1998. View Article : Google Scholar
|
|
81
|
Bauman DR, Bitmansour AD, McDonald JG,
Thompson BM, Liang G and Russell DW: 25-Hydroxycholesterol secreted
by macrophages in response to Toll-like receptor activation
suppresses immunoglobulin A production. Proc Natl Acad Sci USA.
106:16764–16769. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mantovani A, Biswas SK, Galdiero MR, Sica
A and Locati M: Macrophage plasticity and polarization in tissue
repair and remodelling. J Pathol. 229:176–185. 2013. View Article : Google Scholar
|
|
83
|
Gralinski LE and Baric RS: Molecular
pathology of emerging coronavirus infections. J Pathol.
235:185–195. 2015. View Article : Google Scholar
|
|
84
|
de Wilde AH, Wannee KF, Scholte FEM,
Goeman JJ, Ten Dijke P, Snijder EJ, Kikkert M and van Hemert MJ: A
kinome-wide small interfering RNA screen identifies proviral and
antiviral host factors in severe acute respiratory syndrome
coronavirus replication, including double-stranded RNA-activated
protein kinase and early secretory pathway proteins. J Virol.
89:8318–8333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Newton DA, Rao KMK, Dluhy RA and Baatz JE:
Hemoglobin is expressed by alveolar epithelial cells. J Biol Chem.
281:5668–5676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Aldajani WA, Salazar F, Sewell HF, Knox A
and Ghaemmaghami AM: Expression and regulation of immunemodulatory
enzyme indoleamine 2,3-dioxygenase (IDO) by human airway epithelial
cells and its effect on T cell activation. Oncotarget.
7:57606–57617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Mondanelli G, Bianchi R, Pallotta MT,
Orabona C, Albini E, Iacono A, Belladonna ML, Vacca C, Fallarino F,
Macchiarulo A, et al: A relay pathway between arginine and
tryptophan metabolism confers immunosuppressive properties on
dendritic cells. Immunity. 46:233–244. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Nordgren M and Fransen M: Peroxisomal
metabolism and oxidative stress. Biochimie. 98:56–62. 2014.
View Article : Google Scholar
|
|
89
|
Titorenko VI and Terlecky SR: Peroxisome
metabolism and cellular aging. Traffic. 12:252–259. 2011.
View Article : Google Scholar :
|
|
90
|
Xu Z, Lodge R, Power C, Cohen EA and
Hobman TC: The HIV-1 accessory protein vpu downregulates peroxisome
biogenesis. MBio. 11:112020. View Article : Google Scholar
|
|
91
|
Zheng C and Su C: Herpes simplex virus 1
infection dampens the immediate early antiviral innate immunity
signaling from peroxisomes by tegument protein VP16. Virol J.
14:352017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ferreira AR, Marques M and Ribeiro D:
Peroxisomes and innate immunity: Antiviral response and beyond. Int
J Mol Sci. 20:202019. View Article : Google Scholar
|
|
93
|
Skalny AV, Rink L, Ajsuvakova OP, Aschner
M, Gritsenko VA, Alekseenko SI, Svistunov AA, Petrakis D, Spandidos
DA, Aaseth J, et al: Zinc and respiratory tract infections:
Perspectives for COVID-19 (Review). Int J Mol Med. 46:17–26.
2020.
|
|
94
|
Gastaminza P, Whitten-Bauer C and Chisari
FV: Unbiased probing of the entire hepatitis C virus life cycle
identifies clinical compounds that target multiple aspects of the
infection. Proc Natl Acad Sci USA. 107:291–296. 2010. View Article : Google Scholar
|
|
95
|
Simmons G, Gosalia DN, Rennekamp AJ,
Reeves JD, Diamond SL and Bates P: Inhibitors of cathepsin L
prevent severe acute respiratory syndrome coronavirus entry. Proc
Natl Acad Sci USA. 102:11876–11881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Blanchard E, Belouzard S, Goueslain L,
Wakita T, Dubuisson J, Wychowski C and Rouillé Y: Hepatitis C virus
entry depends on clathrin-mediated endocytosis. J Virol.
80:6964–6972. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Trevani AS, Andonegui G, Giordano M, López
DH, Gamberale R, Minucci F and Geffner JR: Extracellular
acidification induces human neutrophil activation. J Immunol.
162:4849–4857. 1999.PubMed/NCBI
|
|
98
|
Rotstein OD, Fiegel VD, Simmons RL and
Knighton DR: The deleterious effect of reduced pH and hypoxia on
neutrophil migration in vitro. J Surg Res. 45:298–303. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Lardner A: The effects of extracellular pH
on immune function. J Leukoc Biol. 69:522–530. 2001.PubMed/NCBI
|
|
100
|
Simchowitz L: Intracellular pH modulates
the generation of superoxide radicals by human neutrophils. J Clin
Invest. 76:1079–1089. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Severin T, Müller B, Giese G, Uhl B, Wolf
B, Hauschildt S and Kreutz W: pH-dependent LAK cell cytotoxicity.
Tumour Biol. 15:304–310. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kellum JA, Song M and Li J: Science
review: extracellular acidosis and the immune response: clinical
and physiologic implications. Crit Care. 8:331–336. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Hamdullah KS, Tanzila A, Zainab Sher M,
Iqra A and Mohtasheemul H: pH dependent differential binding
behavior of protease inhibitor molecular drugs for SARS-COV-2.
ChemRxiv. 2020, Preprint. https://doi.org/10.26434/chemrxiv.12009018.v1.
|
|
104
|
Leneva IA, Russell RJ, Boriskin YS and Hay
AJ: Characteristics of arbidol-resistant mutants of influenza
virus: Implications for the mechanism of anti-influenza action of
arbidol. Antiviral Res. 81:132–140. 2009. View Article : Google Scholar
|
|
105
|
Bhargava M, Becker TL, Viken KJ, Jagtap
PD, Dey S, Steinbach MS, Wu B, Kumar V, Bitterman PB, Ingbar DH, et
al: Proteomic profiles in acute respiratory distress syndrome
differentiates survivors from non-survivors. PLoS One.
9:e1097132014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Shortt K, Chaudhary S, Grigoryev D, Heruth
DP, Venkitachalam L, Zhang LQ and Ye SQ: Identification of novel
single nucleotide polymorphisms associated with acute respiratory
distress syndrome by exome-seq. PLoS One. 9:e1119532014. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Tejera P, Meyer NJ, Chen F, Feng R, Zhao
Y, O′Mahony DS, Li L, Sheu CC, Zhai R, Wang Z, et al: Distinct and
replicable genetic risk factors for acute respiratory distress
syndrome of pulmonary or extrapulmonary origin. J Med Genet.
49:671–680. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Lynn H, Sun X, Casanova N, Gonzales-Garay
M, Bime C and Garcia JGN: Genomic and genetic approaches to
deciphering acute respiratory distress syndrome risk and mortality.
Antioxid Redox Signal. 31:1027–1052. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Marshall RP, Webb S, Bellingan GJ,
Montgomery HE, Chaudhari B, McAnulty RJ, Humphries SE, Hill MR and
Laurent GJ: Angiotensin converting enzyme insertion/deletion
polymorphism is associated with susceptibility and outcome in acute
respiratory distress syndrome. Am J Respir Crit Care Med.
166:646–650. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Itoyama S, Keicho N, Quy T, Phi NC, Long
HT, Ha LD, Ban VV, Ohashi J, Hijikata M, Matsushita I, et al: ACE1
polymorphism and progression of SARS. Biochem Biophys Res Commun.
323:1124–1129. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chan KCA, Tang NLS, Hui DSC, Chung GT, Wu
AK, Chim SS, Chiu RW, Lee N, Choi KW, Sung YM, et al: Absence of
association between angiotensin converting enzyme polymorphism and
development of adult respiratory distress syndrome in patients with
severe acute respiratory syndrome: A case control study. BMC Infect
Dis. 5:262005. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Fourrier F, Chopin C, Wallaert B, Wattre
P, Mangalaboyi J, Durocher A, Dubois D and Wattel F:
Angiotensin-converting enzyme in human adult respiratory distress
syndrome. Chest. 83:593–597. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Imai Y, Kuba K, Rao S, Huan Y, Guo F, Guan
B, Yang P, Sarao R, Wada T, Leong-Poi H, et al:
Angiotensin-converting enzyme 2 protects from severe acute lung
failure. Nature. 436:112–116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Arndt PG, Young SK, Poch KR, Nick JA, Falk
S, Schrier RW and Worthen GS: Systemic inhibition of the
angiotensin-converting enzyme limits lipopolysaccharide-induced
lung neutrophil recruitment through both bradykinin and angiotensin
II-regulated pathways. J Immunol. 177:7233–7241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Kim J, Choi SM, Lee J, Park YS, Lee CH,
Yim JJ, Yoo CG, Kim YW, Han SK and Lee SM: Effect of
renin-angiotensin system blockage in patients with acute
respiratory distress syndrome: A retrospective case control study.
Korean J Crit Care Med. 32:154–163. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wevers BA and van der Hoek L:
Renin-angiotensin system in human coronavirus pathogenesis. Future
Virol. 5:145–161. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Chappell MC: Angiotensin-(17) and the
regulation of anti-fibrotic signaling pathways. J Cell Signal.
2:1342017. View Article : Google Scholar
|
|
118
|
El-Hashim AZ, Renno WM, Raghupathy R,
Abduo HT, Akhtar S and Benter IF: Angiotensin-(17) inhibits
allergic inflammation, via the MAS1 receptor, through suppression
of ERK1/2- and NF-κB-dependent pathways. Br J Pharmacol.
166:1964–1976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Manolis AJ, Marketou ME, Gavras I and
Gavras H: Cardioprotective properties of bradykinin: Role of the
B(2) receptor. Hypertens Res. 33:772–777. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Adam A, Cugno M, Molinaro G, Perez M,
Lepage Y and Agostoni A: Aminopeptidase P in individuals with a
history of angio-oedema on ACE inhibitors. Lancet. 359:2088–2089.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Yeager CL, Ashmun RA, Williams RK,
Cardellichio CB, Shapiro LH, Look AT and Holmes KV: Human
aminopeptidase N is a receptor for human coronavirus 229E. Nature.
357:420–422. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Morris SM Jr: Arginine: Master and
commander in innate immune responses. Sci Signal. 3:pe272010.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Keyaerts E, Vijgen L, Chen L, Maes P,
Hedenstierna G and Van Ranst M: Inhibition of SARS-coronavirus
infection in vitro by S-nitroso-N- acetylpenicillamine, a nitric
oxide donor compound. International Journal of Infectious Diseases
Int J Infect Dis. 8:223–226. 2004. View Article : Google Scholar
|
|
124
|
Akerström S, Mousavi-Jazi M, Klingström J,
Leijon M, Lundkvist A and Mirazimi A: Nitric oxide inhibits the
replication cycle of severe acute respiratory syndrome coronavirus.
J Virol. 79:1966–1969. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Cinatl J, Morgenstern B, Bauer G, Chandra
P, Rabenau H and Doerr HW: Treatment of SARS with human
interferons. Lancet. 362:293–294. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Karupiah G, Xie QW, Buller RML, Nathan C,
Duarte C and MacMicking JD: Inhibition of viral replication by
interferon-γ-induced nitric oxide synthase. Science. 261:1445–1448.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Burrack KS and Morrison TE: The role of
myeloid cell activation and arginine metabolism in the pathogenesis
of virus-induced diseases. Front Immunol. 5:4282014. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Munder M, Schneider H, Luckner C, Giese T,
Langhans CD, Fuentes JM, Kropf P, Mueller I, Kolb A, Modolell M, et
al: Suppression of T-cell functions by human granulocyte arginase.
Blood. 108:1627–1634. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Racké K and Warnken M: L-Arginine
metabolic pathways. The Open Nitric Oxide Journal. 2:pp. 9–19.
2010, https://benthamopen.com/contents/pdf/TONOJ/TONOJ-2-9.pdf.
View Article : Google Scholar
|
|
130
|
Gournas C, Papageorgiou I and Diallinas G:
The nucleobaseascorbate transporter (NAT) family: Genomics,
evolution, structure-function relationships and physiological role.
Mol Biosyst. 4:404–416. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Carr AC and Maggini S: Vitamin C and
immune function. Nutrients. 9:92017. View Article : Google Scholar
|
|
132
|
García-Bailo B, Roke K, Mutch DM,
El-Sohemy A and Badawi A: Association between circulating ascorbic
acid, α-tocopherol, 25-hydroxyvitamin D, and plasma cytokine
concentrations in young adults: A cross-sectional study. Nutr Metab
(Lond). 9:1022012. View Article : Google Scholar
|
|
133
|
Wu G and Morris SM Jr: Arginine
metabolism: Nitric oxide and beyond. Biochem J. 336:1–17. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Scheit K and Bauer G: Synergistic effects
between catalase inhibitors and modulators of nitric oxide
metabolism on tumor cell apoptosis. Anticancer Res. 34:5337–5350.
2014.PubMed/NCBI
|
|
135
|
Thomas SR, Mohr D and Stocker R: Nitric
oxide inhibits indoleamine 2,3-dioxygenase activity in
interferon-gamma primed mononuclear phagocytes. J Biol Chem.
269:14457–14464. 1994.PubMed/NCBI
|
|
136
|
Shi HP, Efron DT, Most D, Tantry US and
Barbul A: Supplemental dietary arginine enhances wound healing in
normal but not inducible nitric oxide synthase knockout mice.
Surgery. 128:374–378. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Rodriguez PC, Quiceno DG and Ochoa AC:
L-arginine availability regulates T-lymphocyte cell-cycle
progression. Blood. 109:1568–1573. 2007. View Article : Google Scholar
|
|
138
|
Ochoa JB, Strange J, Kearney P, Gellin G,
Endean E and Fitzpatrick E: Effects of L-arginine on the
proliferation of T lymphocyte subpopulations. JPEN J Parenter
Enteral Nutr. 25:23–29. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
de Jonge WJ, Kwikkers KL, te Velde AA, van
Deventer SJ, Nolte MA, Mebius RE, Ruijter JM, Lamers MC and Lamers
WH: Arginine deficiency affects early B cell maturation and
lymphoid organ development in transgenic mice. J Clin Invest.
110:1539–1548. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Manning J, Mitchell B, Appadurai DA,
Shakya A, Pierce LJ, Wang H, Nganga V, Swanson PC, May JM, Tantin
D, et al: Vitamin C promotes maturation of T-cells. Antioxid Redox
Signal. 19:2054–2067. 2013. View Article : Google Scholar :
|
|
141
|
Dahl H and Degré M: The effect of
ascorbic acid on production of human interferon and the antiviral
activity in vitro. Acta Pathol Microbiol Scand B. 84B:B280–B284.
1976.
|
|
142
|
Huijskens MJAJ, Walczak M, Sarkar S,
Atrafi F, Senden-Gijsbers BL, Tilanus MG, Bos GM, Wieten L and
Germeraad WT: Ascorbic acid promotes proliferation of natural
killer cell populations in culture systems applicable for natural
killer cell therapy. Cytotherapy. 17:613–620. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Anderson R: Assessment of oral ascorbate
in three children with chronic granulomatous disease and defective
neutrophil motility over a 2-year period. Clin Exp Immunol.
43:180–188. 1981.PubMed/NCBI
|
|
144
|
Patrone F, Dallegri F, Bonvini E,
Minervini F and Sacchetti C: Effects of ascorbic acid on neutrophil
function. Studies on normal and chronic granulomatous disease
neutrophils. Acta Vitaminol Enzymol. 4:163–168. 1982.PubMed/NCBI
|
|
145
|
Moraes MP, de Los Santos T, Koster M,
Turecek T, Wang H, Andreyev VG and Grubman MJ: Enhanced antiviral
activity against foot-and-mouth disease virus by a combination of
type I and II porcine interferons. J Virol. 81:7124–7135. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Peng T, Zhu J, Hwangbo Y, Corey L and
Bumgarner RE: Independent and cooperative antiviral actions of beta
interferon and gamma interferon against herpes simplex virus
replication in primary human fibroblasts. J Virol. 82:1934–1945.
2008. View Article : Google Scholar :
|
|
147
|
Bartee E, Mohamed MR, Lopez MC, Baker HV
and McFadden G: The addition of tumor necrosis factor plus beta
interferon induces a novel synergistic antiviral state against
poxviruses in primary human fibroblasts. J Virol. 83:498–511. 2009.
View Article : Google Scholar :
|
|
148
|
Page C, Goicochea L, Matthews K, Zhang Y,
Klover P, Holtzman MJ, Hennighausen L and Frieman M: Induction of
alternatively activated macrophages enhances pathogenesis during
severe acute respiratory syndrome coronavirus infection. J Virol.
86:13334–13349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Block G, Jensen CD, Dalvi TB, Norkus EP,
Hudes M, Crawford PB, Holland N, Fung EB, Schumacher L and Harmatz
P: Vitamin C treatment reduces elevated C-reactive protein. Free
Radic Biol Med. 46:70–77. 2009. View Article : Google Scholar :
|
|
150
|
Kim WY, Jung JW, Choi JC, Shin JW and Kim
JY: Subphenotypes in patients with septic shock receiving vitamin
C, hydrocortisone, and thiamine: A retrospective cohort analysis.
Nutrients. 11:112019. View Article : Google Scholar
|
|
151
|
Grahame Hardie D: Regulation of
AMP-activated protein kinase by natural and synthetic activators.
Acta Pharm Sin B. 6:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Pagé EL, Chan DA, Giaccia AJ, Levine M
and Richard DE: Hypoxia-inducible factor-1α stabilization in
nonhypoxic conditions: Role of oxidation and intracellular
ascorbate depletion. Mol Biol Cell. 19:86–94. 2008. View Article : Google Scholar
|
|
153
|
Witte MB and Barbul A: Arginine physiology
and its implication for wound healing. Wound Repair Regen.
11:419–423. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Padayatty SJ, Sun H, Wang Y, Riordan HD,
Hewitt SM, Katz A, Wesley RA and Levine M: Vitamin C
pharmacokinetics: Implications for oral and intravenous use. Ann
Intern Med. 140:533–537. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Stancioiu F: Antiviral activity of
L-arginine and extended-release Vitamin C. AASCIT J Health.
3:13–16. 2016.
|
|
156
|
Bertolini G, Iapichino G, Radrizzani D,
Facchini R, Simini B, Bruzzone P, Zanforlin G and Tognoni G: Early
enteral immunonutrition in patients with severe sepsis: Results of
an interim analysis of a randomized multicentre clinical trial.
Intensive Care Med. 29:834–840. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Heyland DK, Novak F, Drover JW, Jain M, Su
X and Suchner U: Should immunonutrition become routine in
critically ill patients? A systematic review of the evidence. JAMA.
286:944–953. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Rosenthal MD, Rosenthal C, Patel J, Jordan
J and Go K: Arginine in the critically ill: Can we finally push
past the controversy? Int J Crit Care Emerg Med. 2:0172016.
View Article : Google Scholar
|
|
159
|
Galbán C, Montejo JC, Mesejo A, Marco P,
Celaya S, Sánchez-Segura JM, Farré M and Bryg DJ: An
immune-enhancing enteral diet reduces mortality rate and episodes
of bacteremia in septic intensive care unit patients. Crit Care
Med. 28:643–648. 2000. View Article : Google Scholar
|
|
160
|
Luiking YC, Poeze M and Deutz NE: Arginine
infusion in patients with septic shock increases nitric oxide
production without haemodynamic instability. Clin Sci (Lond).
128:57–67. 2015. View Article : Google Scholar
|
|
161
|
Gough MS, Morgan MAM, Mack CM, Darling DC,
Frasier LM, Doolin KP, Apostolakos MJ, Stewart JC, Graves BT,
Arning E, et al: The ratio of arginine to dimethylarginines is
reduced and predicts outcomes in patients with severe sepsis. Crit
Care Med. 39:1351–1358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Visser M, Davids M, Verberne HJ, Kok WE,
Tepaske R, Cocchieri R, Kemper EM, Teerlink T, Jonker MA, Wisselink
W, et al: Nutrition before, during, and after surgery increases the
arginine:asymmetric dimethylarginine ratio and relates to improved
myocardial glucose metabolism: a randomized controlled trial. Am J
Clin Nutr. 99:1440–1449. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Visser M, Vermeulen MAR, Richir MC,
Teerlink T, Houdijk AP, Kostense PJ, Wisselink W, de Mol BA, van
Leeuwen PA and Oudemans-van Straaten HM: Imbalance of arginine and
asymmetric dimethylarginine is associated with markers of
circulatory failure, organ failure and mortality in shock patients.
Br J Nutr. 107:1458–1465. 2012. View Article : Google Scholar
|
|
164
|
Arora TK, Malhotra AK, Ivatury R and
Mangino MJ: L-arginine infusion during resuscitation for
hemorrhagic shock: Impact and mechanism. J Trauma Acute Care Surg.
72:397–402. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Wu F, Wilson JX and Tyml K: Ascorbate
protects against impaired arteriolar constriction in sepsis by
inhibiting inducible nitric oxide synthase expression. Free Radic
Biol Med. 37:1282–1289. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Wang Y, Lin H, Lin BW and Lin JD: Effects
of different ascorbic acid doses on the mortality of critically ill
patients: a meta-analysis. Ann Intensive Care. 9:582019. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Fowler AA III, Truwit JD, Hite RD, Morris
PE, DeWilde C, Priday A, Fisher B, Thacker LR II, Natarajan R,
Brophy DF, et al: Effect of vitamin C infusion on organ failure and
biomarkers of inflammation and vascular injury in patients with
sepsis and severe acute respiratory failure: The CITRIS-ALI
Randomized Clinical Trial. JAMA. 322:1261–1270. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Calina D, Docea AO, Petrakis D, Egorov AM,
Ishmukhametov AA, Gabibov AG, Shtilman MI, Kostoff R, Carvalho F,
Vinceti M, et al: Towards effective COVID-19 vaccines: Updates,
perspectives and challenges (Review). Int J Mol Med. 46:3–16.
2020.
|
|
169
|
Chan MHM, Wong VWS, Wong CK, Chan PK, Chu
CM, Hui DS, Suen MW, Sung JJ, Chung SS and Lam CW: Serum LD1
isoenzyme and blood lymphocyte subsets as prognostic indicators for
severe acute respiratory syndrome. J Intern Med. 255:512–518. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Hicks CW, Wang D, Daya NR, Windham BG,
Ballantyne CM, Matsushita K and Selvin E: Associations of cardiac,
kidney, and diabetes biomarkers with peripheral neuropathy among
older adults in the atherosclerosis risk in communities (ARIC)
study. Clin Chem. 66:686–696. 2020. View Article : Google Scholar : PubMed/NCBI
|