Introduction
Oral squamous cell carcinoma (OSCC) is the sixth
most prevalent type of human cancer, and approximately 275,000
cases are newly diagnosed, annually (1). Despite tremendous efforts being made
in treatment modalities, the overall 5-year survival rates
(<50%) remain poor due to the high recurrence and metastasis
rate following surgery (2,3).
Thus, there is an urgent need for the identification of key
molecules which are responsible for and are involved in the
progression of OSCC.
MicroRNAs (miRNAs or miRs) are short, non-coding
RNAs that can bind to the 3'-untranslated region (3'-UTR) of target
mRNAs to regulate gene expression, leading to the degradation of
target mRNAs or translational inhibition of functional proteins
(4). Emerging evidence indicates
that miRNAs are extensively involved in the development of OSCC,
functioning as oncogenes or tumor suppressors, depending on the
target genes (5-7). For instance, Chou et al
demonstrated that miR-486 overexpression led to growth inhibition
and apoptosis induction by targeting discoidin domain receptor-1
(DDR1) in oral cancer cells (8).
Another study revealed that miR-10a promoted tumor cell
proliferation by regulating the glucose transporter 1 (GLUT1)
oncogene in OSCC (9). Peng and
Pang found that miR-140-5p overexpression suppressed the growth of
OSCC tumor xenografts in mice by downregulating p21-activated
kinase 4 (PAK4) (10). In
addition, recent studies have reported that changes in miRNA
profiles in cancer cells have the potential to serve as diagnostic
markers for OSCC (11,12). These previous findings suggest
that the manipulation of miRNAs may serve as a novel therapeutic
approach for OSCC. However, to date, only a limited number of
studies on the roles of miRNAs in OSCC have been conducted, at
least to the best of our knowledge, and thus further extensive
investigations are required.
In the present study, miRNA profiles were examined
in tumor tissues from patients with OSCC using a microarray and
miR-144-3p was found to be one of the most significantly
downregulated miRNAs. Subsequently, gain-of-function experiments
were performed to determine the roles of miR-144-3p. The findings
suggest that miR-144-3p functions as a tumor suppressor by directly
targeting enhancer of zeste homolog 2 (EZH2), and may thus be a
novel target for the diagnosis and treatment of OSCC.
Materials and methods
Clinical specimens
The OSCC tissues and adjacent non-cancerous were
collected from 50 patients with OSCC between May, 2017 and July,
2018 at the Clinical Research Center of Shaanxi Province for Dental
and Maxillofacial Diseases, College of Stomatology, Xi'an Jiaotong
University. These patients included 26 males and 24 females, and
their age ranged from 22 to 72 years, with an average age of 44±6.9
years. The inclusion criteria were histologically established
squamous cell carcinoma within the oral cavity that had primary
surgical treatment with curative intent. Patients treated for OSCC
prior to 2017, and those with recurrent tumors and distant
metastasis at the time of diagnosis were excluded from the study.
All patient characteristics are presented in Table I. The experimental protocol was
approved by the Ethics Committee of the Xi'an Jiaotong University.
Written informed consents for tissue donation were obtained from
each patient for the research only.
 | Table IAssociation between miR-144-3p and
clinicopathological features of patients with oral squamous cell
carcinoma (OSCC). |
Table I
Association between miR-144-3p and
clinicopathological features of patients with oral squamous cell
carcinoma (OSCC).
| Clinical
parameters | All cases
(n=50) | miR-144-3p
expression
| P-value |
|---|
| High (n=21) | Low (n=29) |
|---|
| Sex | | | | 0.5356 |
| Male | 26 | 12 | 14 | |
| Female | 24 | 9 | 15 | |
| Age (years) | | | | 0.1973 |
| ≥0 | 40 | 15 | 25 | |
| <50 | 10 | 6 | 4 | |
| Site | | | | 0.3399 |
| Buccal mucosa | 27 | 13 | 14 | |
| Non-buccal
mucosa | 23 | 8 | 15 | |
| Alcohol
consumption | | | | 0.7381 |
| Yes | 32 | 14 | 18 | |
| No | 18 | 7 | 11 | |
| Smoking habits | | | | 0.1452 |
| Yes | 31 | 11 | 21 | |
| No | 19 | 10 | 8 | |
| Tumor size
(cm) | | | | 0.0025b |
| ≥2 | 22 | 4 | 18 | |
| <2 | 28 | 17 | 11 | |
|
Differentiation | | | | 0.021a |
| Well and
moderate | 15 | 10 | 5 | |
| sPoor | 35 | 11 | 24 | |
| Lymph node
metastasis | | | | 0.044a |
| Present | 34 | 11 | 23 | |
| Absent | 16 | 10 | 6 | |
| cTNM stage | | | | 0.2907 |
| I + II | 21 | 7 | 14 | |
| III + IV | 29 | 14 | 15 | |
miRNA microarray analysis
Total RNA was extracted from the OSCC tissues using
the miRNeasy mini kit (Qiagen, Inc.). The samples were assessed
using the miRCURY™ LNA Array (v.16.0). The procedure and imaging
processes were as previously described (13).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the OSCC tissues and
cells using TRIzol Reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The RNA was reverse transcribed into cDNA using the
PrimeScript RT reagent kit (Takara Bio, Inc.) and the MicroRNA
Reverse Transcription kit (Thermo Fisher Scientific, Inc.),
respectively. miR-144a-3p and EZH2 expression levels were measured
using a SYBR® PrimeScript™ RT-PCR kit (Takara Bio, Inc.)
on a Light Cycler instrument (Bio-Rad Laboratories, Inc.). The
primers of miR-144a-3p and EZH2 were as follows: miR-144-3p
forward, 5′-GCC CCT ACA GTA TAG ATG ATG TA-3′ and reverse, 5′-GTG
CAG GGT CCG AGG T-3′; U6 forward, 5′-GCT TCG GCA GCA CAT ATA CTA
AAA T-3′ and reverse, 5′-CGC TTC ACG AAT TTG CGT GTC AT-3′; EZH2
forward, 5′-TTG TTG GCG GAA GCG TGT AAA ATC-3′ and reverse, 5′-TCC
CTA GTC CCG CGC AAT GAG C-3′; GAPDH forward, 5′-GTG GTG AAG ACG CCA
GT G GA-3′ and reverse, 5′-CGA GCC ACA TCG CTC AGA CA-3′. U6 and
GAPDH were used as internal controls for detecting miR-144-3p and
EZH2, respectively and fold changes were calculated using the
2-∆∆Cq method (14).
Cells and cell culture
HSC-2, CAL-27 and SCC-4 cell lines were purchased
from ATCC and were cultured in DMEM (Invitrogen; Thermo Fisher
Scientific, Inc.) containing with 10% fetal bovine serum at 37°C in
a 5% CO2 incubator. The normal human oral keratinocyte
(NHOK) cells which served as control cells were obtained from the
ScienCell Research Laboratories, Inc. (cat. no. 2610) and cultured
as previously described (15).
Cell transfection
The miR-144-3p mimics and mimics negative control
(mimics NC) were obtained from GenePharma. The EZH2 overexpression
vector, pcDNA-EZH2, and pcDNA vector were constructed by
GenePharma. In addition, EZH2 siRNA (si-EZH2) and si-Scramble were
also purchased from GenePharma.
CAL-27 and SCC-4 cells (8.0×105/well) in
a 6-well plate grown to approximately 80% confluence, and then
respectively transfected with miR-144-3p mimics (20 nM), mimics NC
(20 nM), si-EZH2 (50 nM), or 2 µg pcDNA-EZH2 using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). In addition, inhibition experiments were
performed using the EZH2 inhibitor, GSK126 (a compound competes
with S-adenosyl-methionine for binding to EZH2, thereby inhibiting
histone methyltransferase activity without affecting EZH2 protein
expression), as previously described (16). Briefly, the CAL-27 and SCC-4 cells
were treated with 5 µM GSK126 (Shanghai HanXiang Life
Technology Limited Corporation) or DMSO for 48 h and then collected
for use in further experiments. The concentration of DMSO used was
≤1% to ensure the lack of cytotoxicity.
Cell proliferation assay
The anti-proliferative effect was measured using
Cell Counting kit-8 (CCK-8) assay. Briefly, the CAL-27 and SCC-4
cells were prepared in 96-well plates containing a final volume of
100 µl/well. Following transfection with miR-144-3p mimics
for 24 and 48 h, 10 µl CCK-8 solution was added to the cells
followed by culture for a further 2 h. The absorbance value at 490
nm was read using a spectrophotometer (SpectraMax 190; Molecular
Devices).
Cell apoptosis assay
The cells were harvested at 48 h post-transfection.
After washing in ice-cold PBS, 100 µl binding buffer was
added to the cells, followed by staining with 5 µl
AnnexinV-FITC/PI (Roche Diagnostics GmbH) at room temperature.
Following maintenance for 15 min in the dark, data were collected
on an EPICS XL-MCL FACScan (BD Biosciences) and analyzed using
FlowJo 8.7.1 software (FlowJo, LLC).
Cell invasion assays
For the invasion assay, a Transwell chamber (8
µm; BD Biosciences) coated with Matrigel was used. At 48 h
post-transfection, 8×104 transfected cells in DMEM
without serum were added to the upper chamber for incubation, while
DMEM with 20% FBS was added to the lower chamber at 37°C for 24 h.
The cells at the bottom chamber were then fixed with 4%
paraformaldehyde for 30 min and stained with 0.5% crystal violet
(Solarbio) for 30 min at room temperature. Subsequently, those
cells that had invaded to the lower side of the membrane were
counted as the number of cells under a 10/20X inverted microscope
(Olympus Corp.) in 5 different fields and analyzed using ImageJ
software (version 1.46; Rawak Software, Inc.).
Wound healing assay
The CAL-27 and SCC-4 cells were added in 6-well
plates at a density of 2×105 per well and cultured until
~90% confluency was reached. Following transfection for 24 h, gaps
were made using a 10 µl pipette tip. After washing with PBS
to remove non-adherent cells, the scratches were photographed using
a microscope (Carl Zeiss MicroImaging GmbH). The cells were then
cultured with serum-free medium at 37°C for 24 h, and the scratches
were photographed again. Distances were measured and analyzed using
ImageJ software (version 1.46; Rawak Software, Inc.).
Luciferase assay
miRNA target prediction tools, including MiRanda
(http://miranda.org.uk) and TargetScan Release 7.0
(http://targetscan.org/) were used to search for
the putative targets of miR-144-3p. CAL-27 cells (8×104)
were added to 24-well plates until ~90% confluency was reached, and
the cells were then co-transfected with the luciferase reporter
plasmids (wt-EZH2-UTR-pGL3 or mt-EZH2-UTR-pGL3) with or without
miR-144-3p mimics using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). At 48 h post-transfection, the activity
of luciferase was measured using the dual-luciferase reporter assay
system (Promega Corporation). Renilla luciferase was used
for normalization. The experiments were independently performed in
triplicate.
Western blot analysis
Western blot analysis was performed as previously
described (17). Briefly, total
protein was isolated and quantitated using BCA assay at 48 h
post-transfection. The protein lysates (40 µg) were
electrophoretically transferred onto PVDF membranes (EMD
Millipore), followed blocking with 5% skim milk at 4°C overnight.
The membranes were probed with primary antibodies against EZH2
(1:1,000; cat. no. 5246), E-cadherin (1:1,000; cat. no. 3195),
N-cadherin (1:1,000; cat. no. 13116) and β-actin (1:1,000; cat. no.
4970), followed by incubation with secondary antibody (anti-rabbit
IgG, 1:10,000; cat. no. 7074) for 1 h at room temperature. All
antibodies were obtained from Cell Signaling Technology, Inc.
Proteins bands were detected using an ECL detection system (GE
Healthcare Life Sciences) and blot bands were quantified using
ImageJ software (version 1.46; Rawak Software, Inc.).
Statistical analysis
SPSS 13.0 software package (SPSS, Inc.) was applied
to analyze the data. All data are presented as the means ± SD. When
only 2 groups were compared, the Student's t-test was used. One-way
analysis of variance followed by Tukey's post hoc test was applied
to compare differences between multiple groups. Categorical data
were compared using the Pearson's Chi-squared test. P<0.05 was
considered to indicate statistically significant differences.
Spearman's correlation analysis was used for correlation
analysis.
Results
miR-144-3p expression is downregulated in
OSCC tissues and cell lines
To investigate the role of miRNA expression in OSCC,
the profiles of miRNAs were compared between OSCC tumors and
adjacent non-cancerous tissues using microarray analysis. Following
normalization of the raw data, 50 significant differentially
expressed miRNAs (expression of 31 miRNAs was increased and that of
19 miRNAs was decreased) were observed in the OSCC group compared
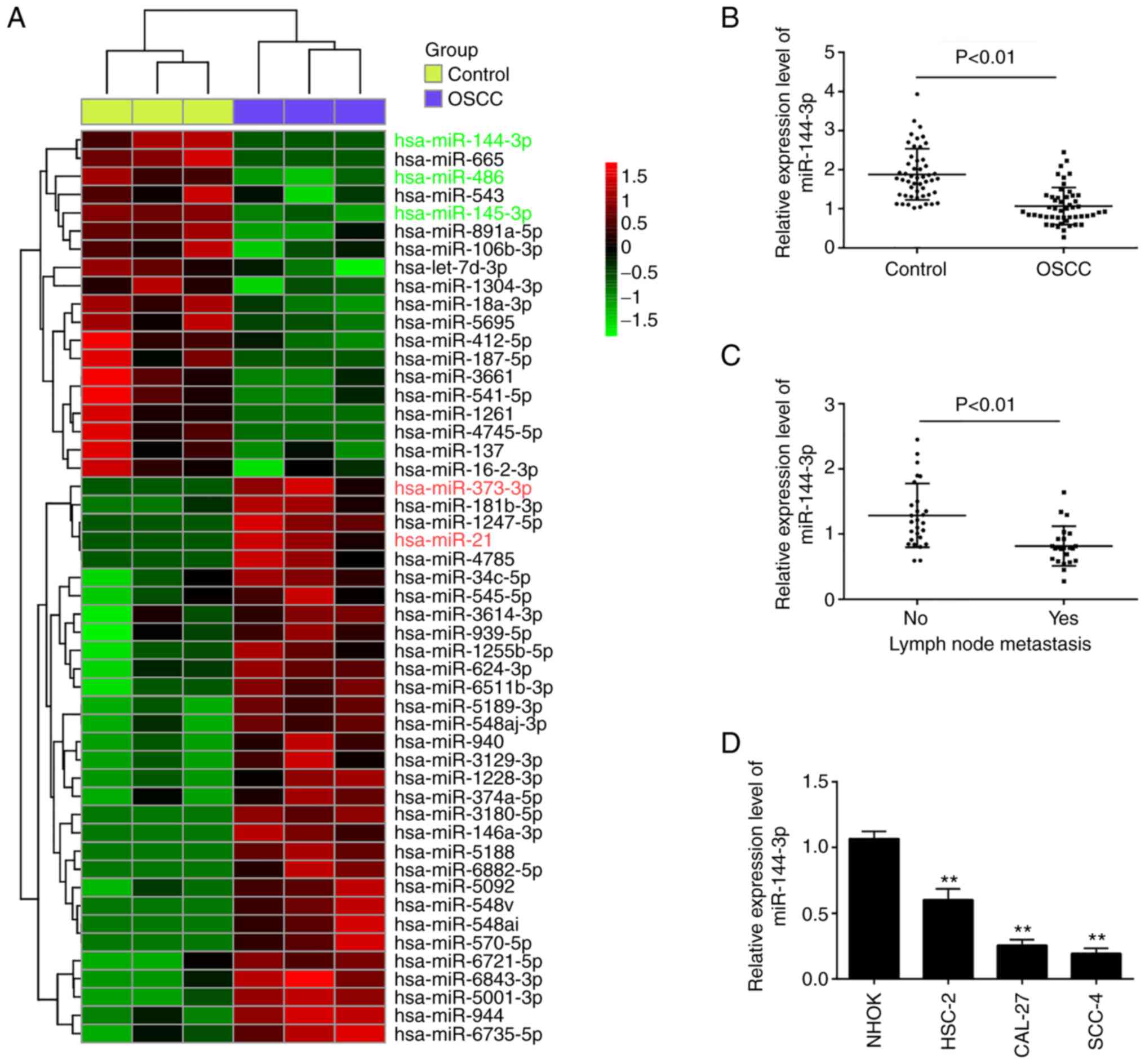
with the control group (Fig. 1A).
Of these aberrant miRNAs, the expression levels of miR-486 and
miR-145-3p were decreased, while those of miR-373-3p and miR-21
were increased; these findings are consistent with those of
previous studies (5,12,17), indicating the reliability of the
microarray in the present study. Among the downregulated miRNAs,
miR-144-3p exhibited the lowest expression levels in the OSCC group
compared with the control group, and some studies have demonstrated
the potent suppressive roles of miR-144-3p in various types of
human cancer (18,19). Therefore, this miRNA was selected
for analysis in further experiments.
To validate the microarray results, the expression
of miR-144-3p was compared between tumor and adjacent non-cancerous
tissues (n=50) by RT-qPCR. It was found that the miR-143-3p levels
in the OSCC tissues were much lower than those in adjacent
non-cancerous tissues (Fig. 1B).
The levels of miR-144-3p were also detected in OSCC tissues from
patients with lymph node metastasis. It was observed that
miR-144-3p expression was notably decreased in tumor tissues with
lymph node metastasis, suggesting that miR-144-3p level was closely
associated with a higher incidence of metastasis (Fig. 1C). In addition, the levels of
miR-144-3p in OSCC cells were measured. Consistent with the results
obtained with the clinical samples, miR-143-3p expression was
significantly downregulated in OSCC cells compared with that in the
normal human oral keratinocyte (NHOK) cells (Fig. 1D).
Subsequently, the clinicopathological significance
of miR-144-3p downregulation in OSCC was analyzed (Table I). It was found that a low
expression of miR-144-3p was closely associated with tumor size,
differentiation and lymph node metastasis. However, other
clinicopathological parameters, such as sex, age, site, alcohol
consumption, smoking habits and TNM stage exhibited no significant
association with miR-144-3p expression. These data indicate that
miR-144-3p may be a potential biomarker for the diagnosis of
OSCC.
Overexpression of miR-144-3p suppresses
cell proliferation and induces cell apoptosis
The frequent downregulation of miR-144-3p in OSCC
cell lines and OSCC tissues suggests that miR-144-3p plays a role
in OSCC carcinogenesis. To prove this hypothesis, the effects of
the ectopic expression of miR-144-3p on cell growth were
investigated in 2 OSCC cell lines (CAL-27 and SCC-4). The rationale
of using these 2 cell lines is that both cell lines exhibited the
lowest expression levels of miR-144-3p among the 3 cell lines
examined (Fig. 1D). As shown in
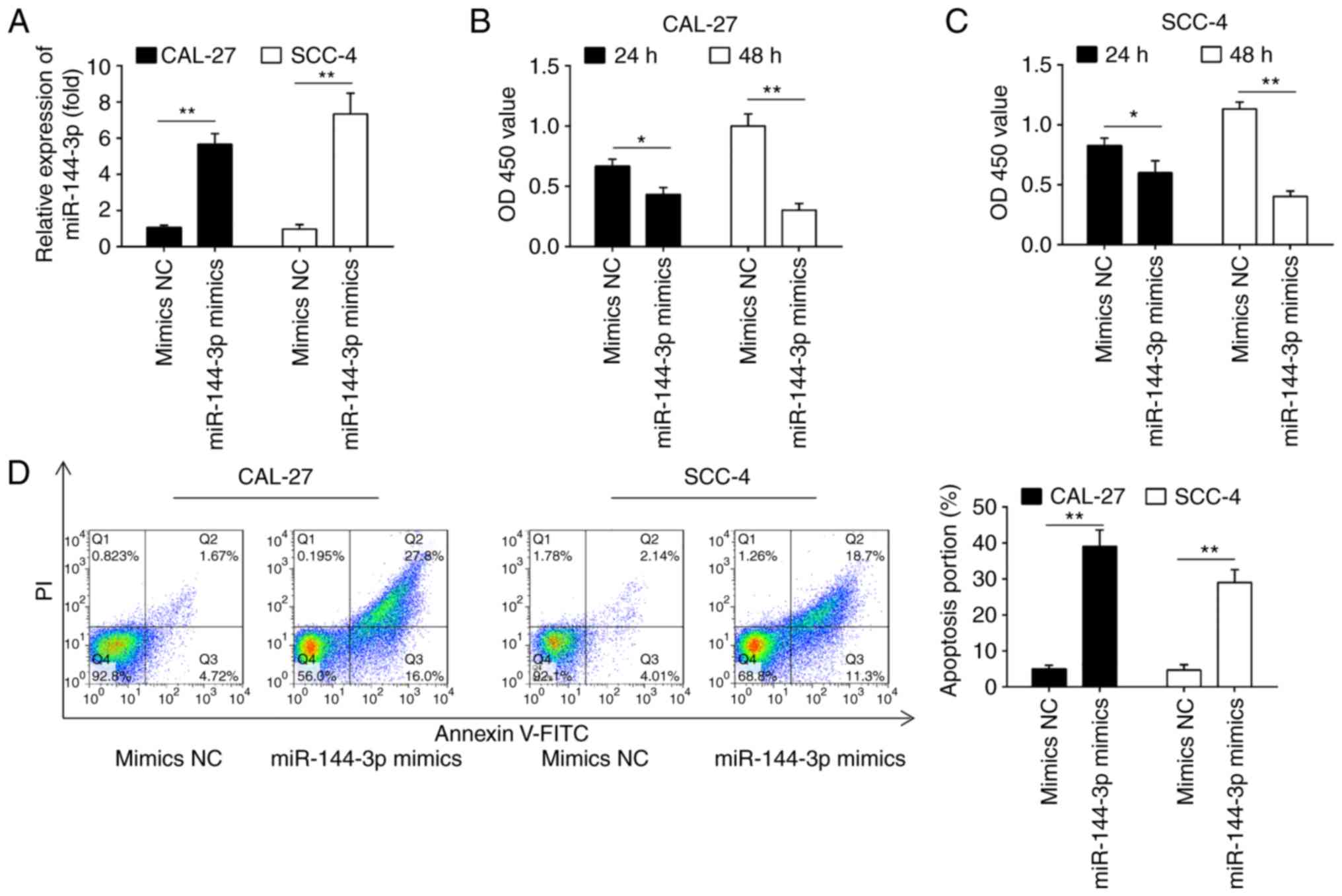
Fig. 2A, miR-144-3p expression
was notably increased in the CAL-27 and SCC-4 cells following
transfection with miR-144-3p mimics, which confirmed the
transfection efficiency. CCK-8 assay revealed that miR-144-3p
upregulation markedly suppressed the proliferation of the CAL-27
and SCC-4 cells compared with the cells transfected with the mimics
control (Fig. 2B and C). Cell
apoptosis analysis was then performed to further examine the
effects of miR-144a-3p on cell proliferation. It was observed that
miR-144-3p upregulation significantly increased cell apop-tosis
compared to the mimics control (Fig.
2D). Thus, it was concluded that miR-144-3p may suppress cell
proliferation by promoting cell apoptosis.
Overexpression of miR-144-3p inhibited
cell invasion and migration
miR-144-3p has previously been found to regulate the
epithelial-mesenchymal transition (EMT) process in gastric cancer
(20). Therefore, it was
hypothesized that the tumor suppressive effects of miR-144-3p may
involve EMT. To verify this hypothesis, the effects of miR-144-3p
on the expression levels of EMT markers were examined at the
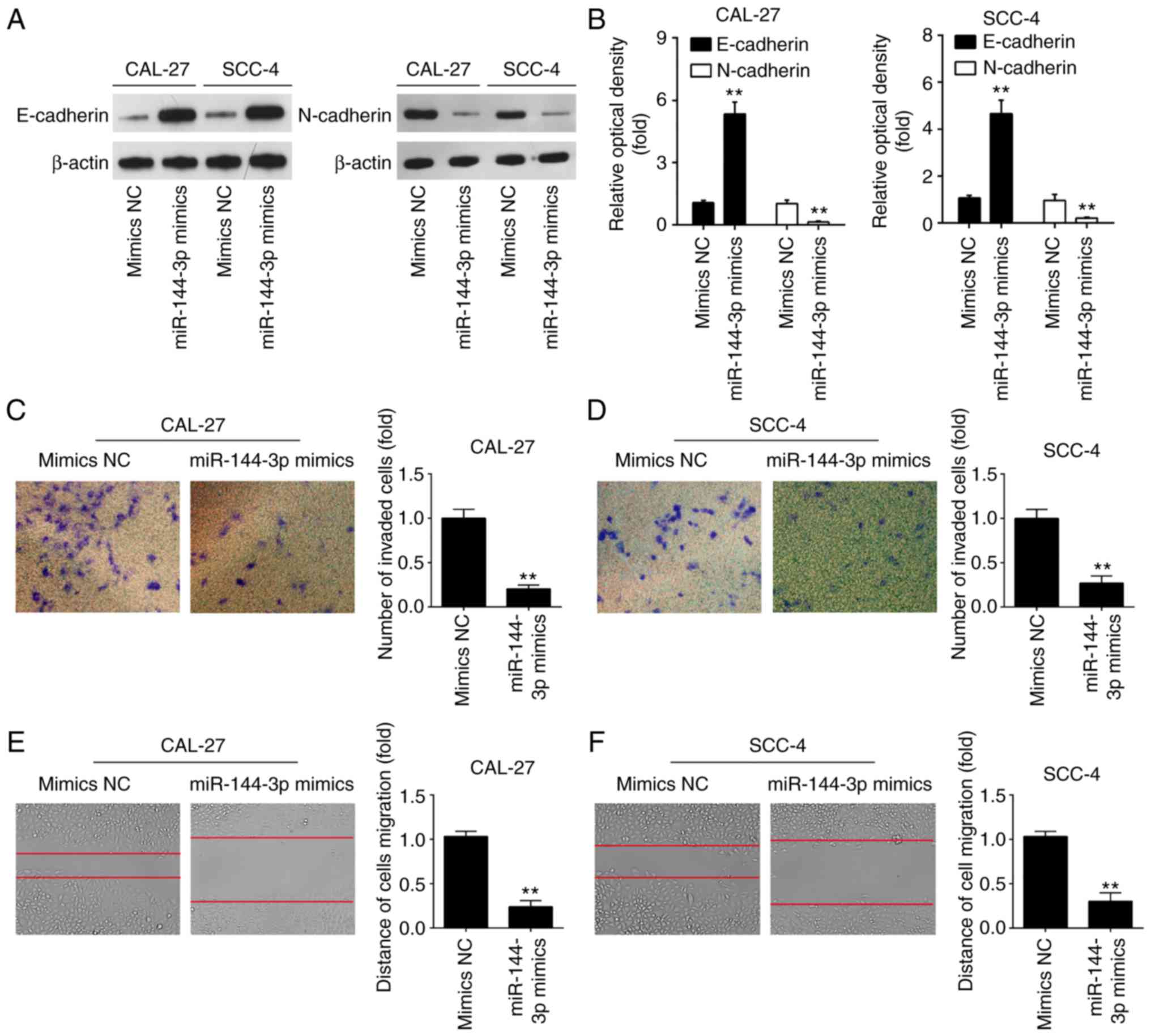
protein level in OSCC cells. As was expected, miR-144-3p mimics
markedly upregulated the protein level of the epithe-lial marker,
E-cadherin, and downregulated the expression of the mesenchymal
marker, N-cadherin, in the CAL-27 and SCC-4 cells, suggesting that
miR-144-3p suppressed the EMT process (Fig. 3A and B). To further examine the
effects of miR-144-3p in the migratory and invasive abilities of
OSCC cells, Transwell assay and wound healing assay were conducted.
miR-144-3p upregulation suppressed the invasion and migration of
CAL-27 and SCC-4 cells compared to the mimics NC group (Fig. 3C-F). The above-mentioned data
indicate that miR-144-3p suppresses the migration and invasion of
OSCC cells.
EZH2 is a direct target of miR-144-3p in
OSCC cells
To better understand the mechanisms through which
miR-144-3p exerts its tumor suppressive effects, the present study
sought to identify the mRNA targets of miR-144-3p. Thus, TargetScan
7.0 (http://targetscan.org/) and miRanda
(http://miranda.org.uk) were used to search for
the predict targets of miR-144-3p, particularly for those that were
associated with cell growth or invasion. Bioinformatics analysis
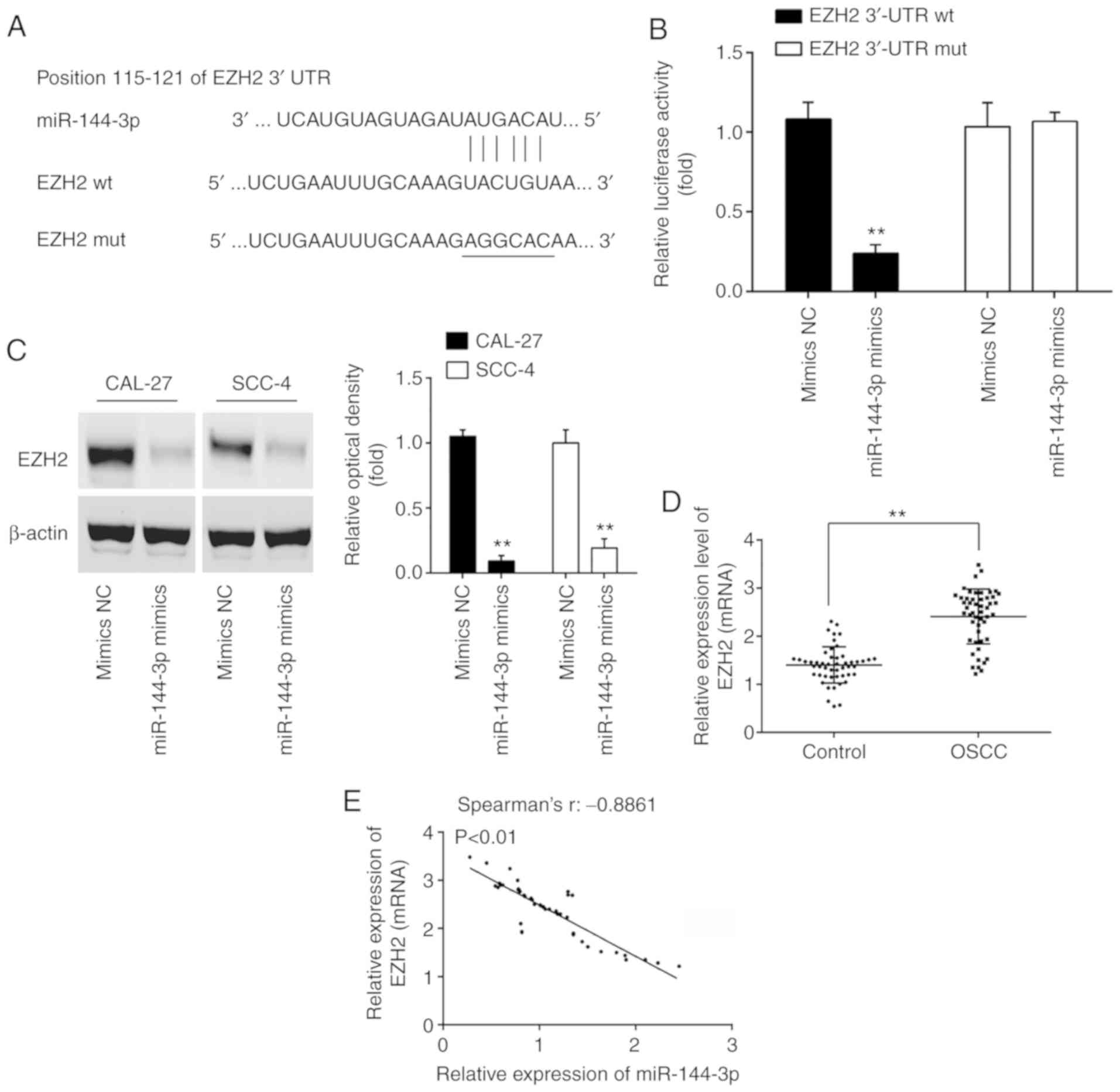
predicted that EZH2 may be a potential target of miR-144-3p, since
the EZH2 3'UTR possesses a miR-144-3p binding site (Fig. 4A). In order to confirm the
interaction between miR-144-3p and EZH2, a luciferase assay was
performed. It was found that miR-144-3p mimics decreased the
relative luciferase activity of EZH2 3'-UTR wt. However, no
significant differences were observed in the luciferase activity of
EZH2-3'UTR mut reporter by transfection with miR-144-3p mimics
(Fig. 4B). Western blot analysis
was then performed to explore the association between miR-144-3p
and EZH2 protein expression. Compared to the mimics NC group,
miR-144-3p overexpression significantly decreased the expression of
EZH2 in both the CAL-27 and SCC-4 cells (Fig. 4C). In addition, the expression
levels of EZH2 in 50 pairs of OSCC clinical samples were measured
by RT-qPCR. In accordance with the findings of previous studies
(21,22), EZH2 was highly expressed in the
OSCC group compared with the control group (Fig. 4D). Furthermore, Spearman's
correlation analysis revealed a significant inverse correlation
between the EZH2 and miR-144-3p expression levels in the tumor
tissues (Fig. 4E). These data
suggest that miR-144-3p suppresses the expression of EZH2 oncogene
in OSCC.
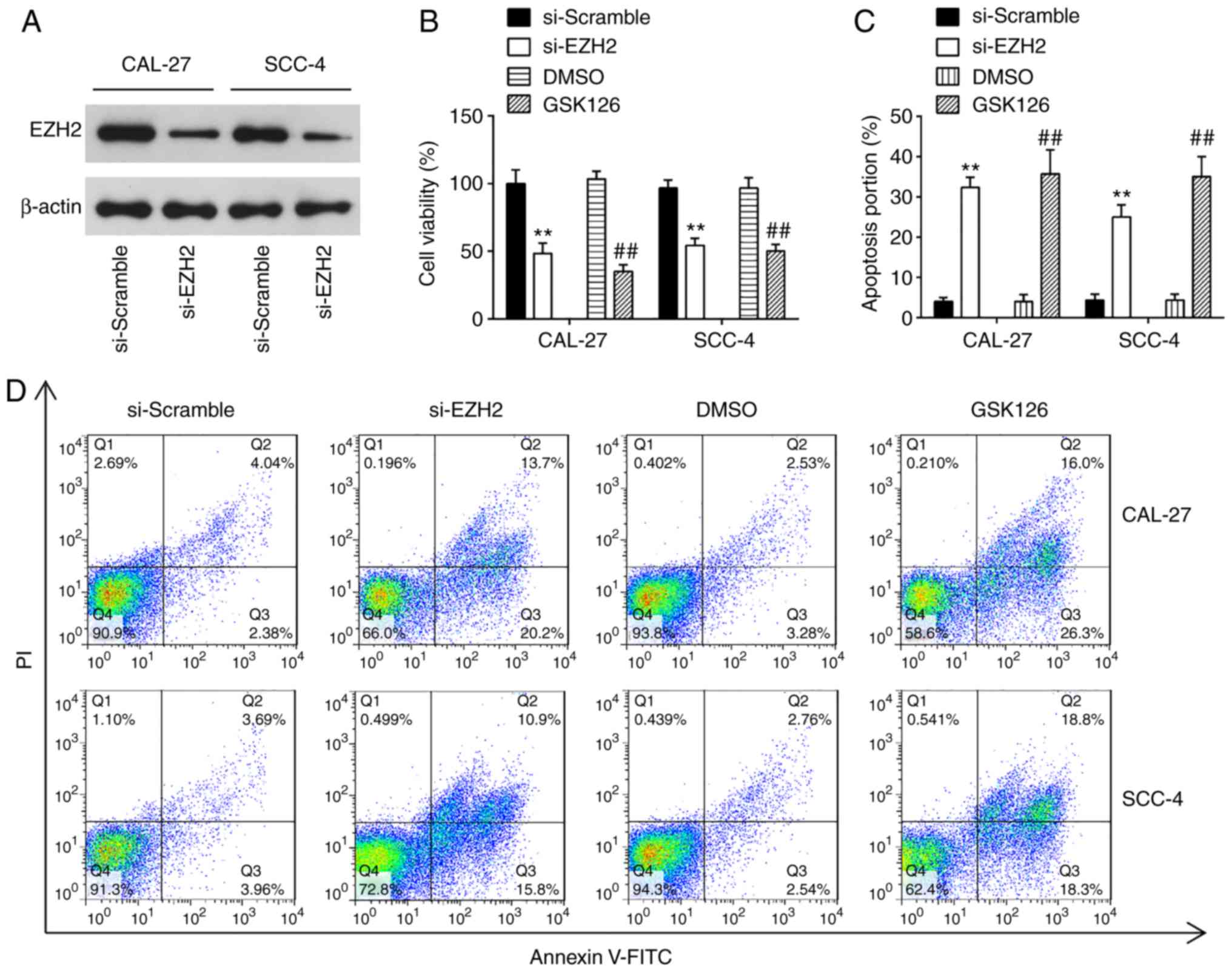
Knockdown of EZH2 inhibits cell
proliferation, promotes cell apoptosis and suppresses cell invasion
and migration
EZH2 is a well-known oncogene in several human
cancers, including prostate cancer (23), bladder cancer (24) and renal cell carcinomas (25). In the present study, to examine
the effects of EZH2 on OSCC cells, cell proliferation, apoptosis,
invasion and migration were examined in the CAL-27 and SCC-4 cells
following transfection with si-NC or si-EZH2 for 48 h. Western blot
analysis revealed that EZH2 expression was significantly decreased
in the CAL-27 and SCC-4 cells transfected with si-EZH2 for 48 h
compared with the si-Scramble group (Fig. 5A). The results of CCK-8 assay
revealed that the knock-down of EZH2 4 significantly suppressed the
proliferation of OSCC cells, compared with the si-Scramble group
(Fig. 5B). Additionally, EZH2
inhibition markedly promoted cell apoptosis compared with the
si-Scramble group (Fig. 5C and
D). Furthermore, Transwell assay suggested that the invasive
abilities of the CAL-27 and SCC-4 cells were suppressed by the
knockdown of EZH2 (Fig. 5E).
Consequently, EZH2 silencing exerted similar effects to miR-144-3p
overexpression on OSCC cells.
To further confirm the inhibitory effect of EZH2
knock-down on OSCC cells, the CAL-27 and SCC-4 cells were treated
with 5 µM GSK126, an Ezh2 inhibitor, for 48 h. The results
revealed that GSK126 treatment suppressed the proliferation,
promoted the apoptosis and inhibited the invasion and migration of
CAL-27 and SCC-4 cells; these findings were similar to those
observed with si-EZH2 on the cellular functions of OSCC cells.
Taken together, these findings suggest the feasibility of EZH2 for
use in clinical trials for the control of OSCC.
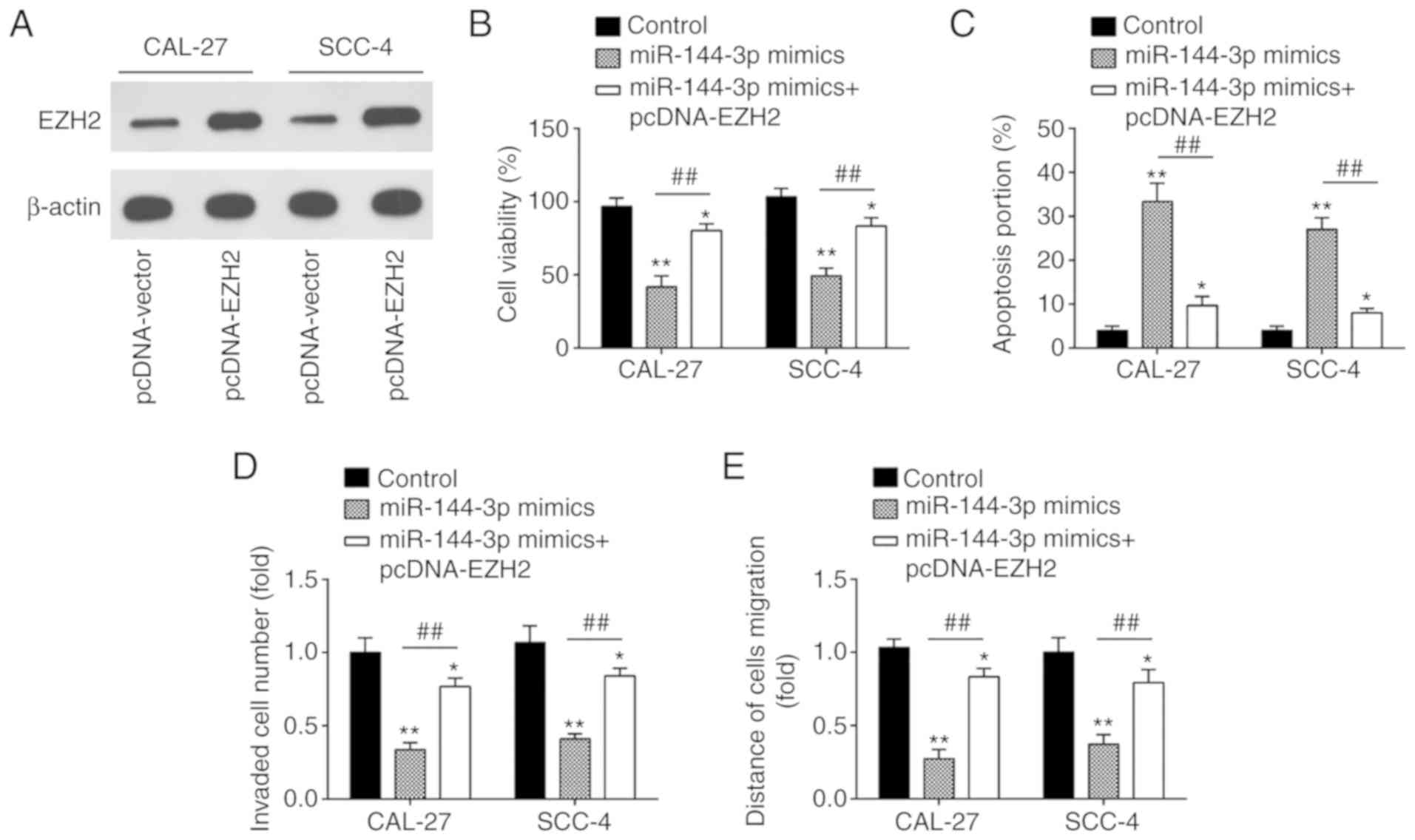
miR-144-3p regulates the biological
behavior of OSCC cells by targeting EZH2
To determine whether the EZH2 oncogene is
responsible for the inhibitory effects of miR-144-3p in OSCC cells,
pcDNA-EZH2 combined with miR-144-3p mimics or mimics NC were added
to the CAL-27 and SCC-4 cells. Western blot analysis revealed that
EZH2 expression was upregulated by transfection with pcDNA-EZH2
plasmid in the CAL-27 and SCC-4 cells (Fig. 6A). Subsequently, CCK-8 assay
demonstrated that the concomitant overexpression of miR-144-3p
mimics and pcDNA-EZH2 abrogated this inhibitory effect of
miR-144-3p mimics on cell proliferation (Fig. 6B). Consistently, the pro-apoptotic
effect by miR-144-3p was markedly abolished by pcDNA-EZH2 (Fig. 6C). Furthermore, the overexpression
of EZH2 partially reversed the inhibitory effects of miR-144-3p on
the cell invasion and migration (Fig.
6D and E). These results suggest that EZH2 is a functional
mediator of miR-144-3p in OSCC cells.
Discussion
In the present study, it was demonstrated that
miR-144-3p was expressed at low level in OSCC tissues and cells.
Clinical association analysis indicated that the low expression of
miR-144-3p was associated with tumor size, differentiation and
lymph node metastasis. Moreover, miR-144-3p inhibited the growth
and invasive ability of OSCC cells by suppressing the EZH2
oncogene. This suggests that miR-144-3p has the potential to
function as a novel therapeutic target for OSCC.
Several studies have demonstrated that miRNAs play
essential roles in controlling multiple steps of OSCC occurrence
and development, including proliferation, apoptosis, invasion and
metastasis. For example, miR-188 overexpression has been reported
to inhibit OSCC cell growth and invasion by targeting SIX1
(26). Cui et al revealed
that miR-378-3p/5p suppressed OSCC metastasis by inhibiting
kallikrein-related peptidase 4 (KLK4) expression (6). Ding et al demonstrated that
miR-145 overexpression suppressed the growth of OSCC xenograft
tumors in vivo (27).
Therefore, the identification of novel oncogenic or tumor
suppressive miRNA involved in OSCC progression is beneficial for
the discovery of novel therapeutic targets for OSCC. In the present
study, using a microarray, a number of miRNAs were found to be
aberrantly expressed in OSCC tissues; in particular, miR-144-3p
expression displayed the most downregulated changes, which was
supported by a previous study (28). Notably, low expression of
miR-144-3p was observed in lymph node metastasis tissues,
suggesting the association with higher incidence of lymph node
metastasis. Of note, the low expression of miR-144-3p was
associated with the tumor size, differentiation and lymph node
metastasis. Thus, it is worthy to further detect the serum
miR-144-3p level in OSCC patients to clarify its significance as a
diagnostic biomarker.
It is worth mentioning that miR-144-3p is mainly
considered as a tumor suppressor in many solid tumors (29). For example, miR-144-3p has been
demonstrated to function as a tumor suppressor in gastric cancer by
activating enhancer-binding protein 4 (AP4) regulation (30). Wu et al illustrated that
miR-144-3p targets MAPK6 to suppress cervical cancer cell growth of
cervical cancer (19).
Furthermore, miR-144-5p, another isoform of miR-144, directly
targets SDC3 in renal cell carcinoma (RCC) to inhibit the cancer
cell growth (31). Additionally,
the study by Chen et al demonstrated that the low expression
of miR-144-3p contributes to the prediction in lung cancer patients
(32). These previous studies
support the important roles of miR-144-3p in different types of
cancer. Although there is evidence to indicate the suppressive role
of miR-144-3p in OSCC (28,33), the unique role of miR-144-3p and
its exact mechanisms in OSCC remain largely unclear. In the present
study, it was found that miR-143-3p upregulation significantly
suppressed OSCC cell proliferation, induced apoptosis, and
effectively suppressed the migration and invasion of OSCC cells by
gain-of-function experiments, which is consistent with the findings
of a previous study (28). These
data indicate that miR-144-3p functions as a tumor suppressor in
OSCC progression.
EZH2 belongs to the polycomb-group (PcG) family, and
affects embryonic stem cell pluripotency and self-renewal (34,35). Additionally, increasing evidences
have revealed that EZH2 was overexpressed in multiple tumors, such
as hepatocellular carcinoma, colorectal cancer, and lung cancer
(36). Furthermore, EZH2 has been
indicated to function as an oncogene by decreasing the activity of
histone methyltransferase and silencing the downstream
anti-oncogene (37,38). For example, EZH2 expression is
associated with the clinical characteristics of patients with
colorectal cancer (CC), and the inhibition of EZH2 reduces the
proliferation and invasion of CC cells (39). Furthermore, EZH2 has also been
identified as an oncogene in breast and prostate cancer (40). Moreover, the use of inhibitors of
EZH2 for cancer treatment has been an active area (41,42), and have moved to clinical trials
(ClinicalTrials.gov Identifier:
NCT02395601 and NCT02082977). Moreover, a number of researchers
have reported that EZH2 is well regulated by miRNAs. For instance,
Sun et al found that EZH2 was a direct target gene of
miR-4465 in non-small cell lung cancer (43). Fan et al discovered miR-217
can regulate EZH2 in nasopharyngeal carcinoma (44). Herein, it was confirmed that EZH2
was directly targeted by miR-144-3p. It was also found that the
EZH2 expression level was increased, and inversely correlated with
the miR-144-3p levels in OSCC tissues. Finally, it was demonstrated
that EZH2 inhibition exhibited a similar role with miR-144-3p
mimics, whereas the overexpression of EZH2 abolished the tumor
suppressive effect of miR-144-3p mimics on OSCC cells, suggesting
that the EZH2 oncogene mediates the role of miR-144-3p in OSCC.
Although the results of recent genomic studies have
enhanced the understanding of the role of EZH2 in cancer
development, the detailed mechanisms underlying EZH2 function are
still complex in distinct cancer types. Generally, EZH2 is the
enzymatic subunit of polycomb repressive complex 2 (PRC2), which
methylates lysine 27 of histone H3 (H3K27) to promote
transcriptional silencing (45).
Given its role as a transcriptional regulator, several efforts have
been dedicated to the identification of downstream targets or
pathways that are driven by EZH2. Researchers have revealed that
EZH2 contributes to carcinogenesis by functioning as an oncogene
through the silencing of E-cadherin and DNA-damage repair pathways
(46,47). The ectopic expression of EZH2 also
promotes cancers driven by the loss of SMARCB1 in malignant
rhabdoid tumor (48). On the
contrary, EZH2 has been shown to interact with PCNA-associated
factor (PAF) to the β-catenin complex, and promoting
transcriptional activation Wnt target genes in colon cancer cells
(49). In addition, the
phosphorylation of EZH2 by AKT functions as a co-activator for
critical transcription factors, such as androgen receptor (AR) in
prostate cancer cells (50).
Therefore, EZH2 functions as a double-facet molecule in regulation
of gene expression via repression or activation mechanism,
depending on the different cellular contexts. In the present study,
it was proven that miR-144-3p functions as a tumor suppressor by
suppressing EZH2. However, the detailed mechanisms underlying the
inhibition of cell viability, and cell invasion and migration by
the knockdown of EZH2 warrant further investigation in the
future.
However, there are still some limitations to the
present study. Due to the limitation in experimental conditions and
funds, further research is required in the future to investigate
the expression levels of miR-144-3p in more clinical samples.
Furthermore, the present study investigated the cellular function
of miR-144-3p and its underlying mechanisms in OSCC; however, in
vivo studies and clinical trial data are required to validate
the preliminary in vitro results obtained in the present
study. Therefore, the function of miR-144-3p in OSCC warrants
further investigation in vivo.
In conclusion, the findings of the present study
demonstrate that miR-144-3p is frequently downregulated in OSCC and
serves as a potential tumor suppressor in OSCC. The findings
suggest that the miR-144-3p/EZH2 axis may be a promising target for
the treatment of OSCC.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH and LL performed the experiments, contributed to
data analysis and wrote the manuscript. LH and LL analyzed the
data. LD conceptualized the study design, contributed to data
analysis and experimental materials. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All individuals provided informed consent for the
use of human specimens for clinical research. The present study was
approved by the College of Stomatology, Xi'an Jiaotong University
Ethics Committee.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Warnakulasuriya S: Global epidemiology of
oral and oropharyngeal cancer. Oral Oncol. 45:309–316. 2009.
View Article : Google Scholar
|
|
2
|
Sasahira T, Kurihara M, Bhawal UK, Ueda N,
Shimomoto T, Yamamoto K, Kirita T and Kuniyasu H: Downregulation of
miR-126 induces angiogenesis and lymphangiogenesis by activation of
VEGF-A in oral cancer. Br J Cancer. 107:700–706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marsh D, Suchak K, Moutasim KA, Vallath S,
Hopper C, Jerjes W, Upile T, Kalavrezos N, Violette SM, Weinreb PH,
et al: Stromal features are predictive of disease mortality in oral
cancer patients. J Pathol. 223:470–481. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li T and Cho WC: MicroRNAs: Mechanisms,
functions and progress Genomics Proteomics. Bioinformatics.
10:237–238. 2012.
|
|
5
|
Zhang XJ, Jin Y, Song JL and Deng F:
MiR-373 promotes proliferation and metastasis of oral squamous cell
carcinoma by targeting SPOP. Eur Rev Med Pharmacol Sci.
23:5270–5276. 2019.PubMed/NCBI
|
|
6
|
Cui Z, Sun S, Liu Q, Zhou X, Gao S, Peng P
and Li Q: MicroRNA-378-3p/5p suppresses migration and invasion of
oral squamous carcinoma cells by inhibiting KLK4 expression.
Biochem Cell Biol. 98:154–163. 2019. View Article : Google Scholar
|
|
7
|
Zhang C, Hao Y, Sun Y and Liu P: Quercetin
suppresses the tumorigenesis of oral squamous cell carcinoma by
regulating microRNA-22/WNT1/β-catenin axis. J Pharmacol Sci.
140:128–136. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chou ST, Peng HY, Mo KC, Hsu YM, Wu GH,
Hsiao JR, Lin SF, Wang HW and Shiah SG: MicroRNA-486-3p functions
as a tumor suppressor in oral cancer by targeting DDR1. J Exp Clin
Cancer Res. 38:2812019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen YH, Song Y, Yu YL, Cheng W and Tong
X: miRNA-10a promotes cancer cell proliferation in oral squamous
cell carcinoma by upregulating GLUT1 and promoting glucose
metabolism. Oncol Let. 17:5441–5446. 2019.
|
|
10
|
Peng M and Pang C: MicroRNA-140-5p
inhibits the tumorigenesis of oral squamous cell carcinoma by
targeting p21-activated kinase 4. Cell Biol Int. 2019.Ahead of
print.
|
|
11
|
Parpart S and Wang XW: microRNA regulation
and its consequences in cancer. Curr Pathobiol Rep. 1:71–79. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shao Y, Qu Y, Dang S, Yao B and Ji M:
MiR-145 inhibits oral squamous cell carcinoma (OSCC) cell growth by
targeting c-Myc and Cdk6. Cancer Cell Int. 13:512013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei D, Wang W, Shen B, Zhou Y, Yang X, Lu
G, Yang J and Shao Y: MicroRNA199a-5p suppresses migration and
invasion in oral squamous cell carcinoma through inhibiting the
EMTrelated transcription factor SOX4. Int J Mol Med. 44:185–195.
2019.PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Wei D, Shen B, Wang W, Zhou Y, Yang X, Lu
G, Yang J and Shao Y: MicroRNA199a-5p functions as a tumor
suppressor in oral squamous cell carcinoma via targeting the
IKKβ/NF-κB signaling pathway. Int J Mol Med. 43:1585–1596.
2019.PubMed/NCBI
|
|
16
|
Huang S, Wang Z, Zhou J, Huang J, Zhou L,
Luo J, Wan Y, Long H and Zhu B: EZH2 Inhibitor GSK126 suppresses
anti-tumor immunity by driving production of myeloid-derived
suppressor cells. Cancer Res. 79:2009–2020. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan Y, Wang X, Veno MT, Bakholdt V,
Sørensen JA, Krogdahl A, Sun Z, Gao S and Kjems J: Circulating
miRNAs as biomarkers for oral squamous cell carcinoma recurrence in
operated patients. Oncotarget. 8:8206–8214. 2017. View Article : Google Scholar :
|
|
18
|
Jiang W, Xu Z, Yu L, Che J, Zhang J and
Yang J: MicroRNA-144-3p suppressed TGF-B1-induced lung cancer cell
invasion and adhe-sion by regulating the Src-Akt-Erk pathway. Cell
Biol Int. 2019.Ahead of print.
|
|
19
|
Wu J, Zhao Y, Li F and Qiao B: MiR-144-3p:
A novel tumor suppressor targeting MAPK6 in cervical cancer. J
Physiol Biochem. 75:143–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li B, Zhang S, Shen H and Li C:
MicroRNA-144-3p suppresses gastric cancer progression by inhibiting
epithelial-to-mesenchymal transition through targeting PBX3.
Biochem Biophys Res Commun. 484:241–247. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao L, Yu Y, Wu J, Bai J, Zhao Y, Li C,
Sun W and Wang X: Role of EZH2 in oral squamous cell carcinoma
carcinogenesis. Gene. 537:197–202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Wan Q, Wang W, Mai L, Sha L, Mashrah
M, Lin Z and Pan C: LncRNA ADAMTS9-AS2 promotes tongue squamous
cell carcinoma proliferation, migration and EMT via the
miR-600/EZH2 axis. Biomed Pharmacother. 112:1087192019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Labbe DP, Sweeney CJ, Brown M, Galbo P,
Rosario S, Wadosky KM, Ku SY, Sjostrom M, Alshalalfa M, Erho N, et
al: TOP2A and EZH2 provide early detection of an aggressive
prostate cancer subgroup. Clin Cancer Res. 23:7072–7083. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu D, Li Y, Luo G, Xiao X, Tao D, Wu X,
Wang M, Huang C, Wang L, Zeng L and Jiang G: LncRNA SPRY4-IT1
sponges miR-101-3p to promote proliferation and metastasis of
bladder cancer cells through up-regulating EZH2. Cancer Lett.
388:281–291. 2017. View Article : Google Scholar
|
|
25
|
Wang Y, Chen Y, Geng H, Qi C, Liu Y and
Yue D: Overexpression of YB1 and EZH2 are associated with cancer
metastasis and poor prognosis in renal cell carcinomas. Tumour
Biol. 36:7159–7166. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang L and Liu H: microRNA-188 is
downregulated in oral squamous cell carcinoma and inhibits
proliferation and invasion by targeting SIX1. Tumour Biol.
37:4105–4113. 2016. View Article : Google Scholar
|
|
27
|
Ding J, Sun D and Xie P: Elevated
microRNA-145 inhibits the development of oral squamous cell
carcinoma through inactivating ERK/MAPK signaling pathway by
down-regulating HOXA1. Biosci Rep. 39:BSR201822142019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li X, Li Y, Jiang C, Chen L and Gan N:
MicroRNA-144-3p inhibits tumorigenesis of oral squamous cell
carcinoma by downregulating ERO1L. J Cancer. 11:759–768. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu M, Lin Y, Zhou Y, Jin H, Hou B, Wu Z,
Li Z, Jian Z and Sun J: MiR-144 suppresses cell proliferation,
migration, and invasion in hepatocellular carcinoma by targeting
SMAD4. Onco Targets Ther. 9:4705–4714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mushtaq F, Zhang J and Li J: miR-144
suppresses cell proliferation and invasion in gastric cancer
through downregulation of activating enhancer-binding protein 4.
Oncol Lett. 17:5686–5692. 2019.PubMed/NCBI
|
|
31
|
Yamada Y, Arai T, Kojima S, Sugawara H,
Kato M, Okato A, Yamazaki K, Naya Y, Ichikawa T and Seki N:
Regulation of antitumor miR-144-5p targets oncogenes: Direct
regulation of syndecan-3 and its clinical significance. Cancer Sci.
109:2919–2936. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen N, Feng L, Lu K, Li P, Lv X and Wang
X: STAT6 phosphor-ylation upregulates microRNA-155 expression and
subsequently enhances the pathogenesis of chronic lymphocytic
leukemia. Oncol Lett. 18:95–100. 2019.PubMed/NCBI
|
|
33
|
Pedersen NJ, Jensen DH, Lelkaitis G, Kiss
K, Charabi BW, Ullum H, Specht L, Schmidt AY, Nielsen FC and
Buchwald C: MicroRNA-based classifiers for diagnosis of oral cavity
squamous cell carcinoma in tissue and plasma. Oral Oncol. 83:46–52.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gu W, Zhang E, Song L, Wang Z, Tu L, Tian
F, Aikenmu K, Chu G and Zhao J: Long noncoding RNA HOXD-AS1
aggravates osteosarcoma carcinogenesis through epigenetically
inhibiting p57 via EZH2. Biomed Pharmacother. 106:890–895. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pasini D, Bracken AP, Hansen JB, Capillo M
and Helin K: The polycomb group protein Suz12 is required for
embryonic stem cell differentiation. Mol Cell Biol. 27:3769–3779.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kondo Y: Targeting histone
methyltransferase EZH2 as cancer treatment. J Biochem. 156:249–257.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Matsukawa Y, Semba S, Kato H, Ito A,
Yanagihara K and Yokozaki H: Expression of the enhancer of zeste
homolog 2 is correlated with poor prognosis in human gastric
cancer. Cancer Sci. 97:484–491. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Raman JD, Mongan NP, Tickoo SK, Boorjian
SA, Scherr DS and Gudas LJ: Increased expression of the polycomb
group gene, EZH2, in transitional cell carcinoma of the bladder.
Clin Cancer Res. 11:8570–8576. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song-Bing H, Hao Z, Jian Z, Guo-Qiang Z,
Tuo H, Dai-Wei W, Wen G, Lin G, Yi Z, Xiao-Feng X, et al:
Inhibition of EZH2 expression is associated with the proliferation,
apoptosis and migration of SW620 colorectal cancer cells in vitro.
Exp Biol Med. 240:546–555. 2015.
|
|
40
|
Yoo KH and Hennighausen L: EZH2
methyltransferase and H3K27 methylation in breast cancer. Int J
Biol Sci. 8:59–65. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim KH and Roberts CW: Targeting EZH2 in
cancer. Nat Med. 22:128–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim W, Bird GH, Neff T, Guo G, Kerenyi MA,
Walenssky LD and Orkin SH: Targeted disruption of the EZH2-EED
complex inhibits EZH2-dependent cancer. Nat Chem Biol. 9:643–650.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sun J, Tian X, Lu SQ and Hu HB:
MicroRNA-4465 suppresses tumor proliferation and metastasis in
non-small cell lung cancer by directly targeting the oncogene EZH2.
Biomed Pharmacother. 96:1358–1362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fan DC, Zhao YR, Qi H, Hou JX and Zhang
TH: MiRNA-506 presents multiple tumor suppressor activities by
targeting EZH2 in nasopharyngeal carcinoma. Auris Nasus Larynx.
2020.Ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gall Troselj K, Novak Kujundzic R and
Ugarkovic D: Polycomb repressive complex's evolutionary conserved
function: The role of EZH2 status and cellular background. Clin
Epigenetics. 8:552016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao K, He J, Wang YF, Jin SD, Fan Y, Fang
N, Qian J, Xu TP and Guo RH: EZH2-mediated epigenetic suppression
of EphB3 inhibits gastric cancer proliferation and metastasis by
affecting E-cadherin and vimentin expression. Gene. 686:118–124.
2019. View Article : Google Scholar
|
|
47
|
Ito T, Teo YV, Evans SA, Neretti N and
Sedivy JM: Regulation of cellular senescence by polycomb chromatin
modifiers through distinct DNA damage- and histone
methylation-dependent path-ways. Cell Rep. 22:3480–3492. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Knutson SK, Warholic NM, Wigle TJ, Klaus
CR, Allain CJ, Raimondia A, Scott MP, Chesworth R, Moyer MP,
Copeland RA, et al: Durable tumor regression in genetically altered
malignant rhab tumors by inhibition of methyltransferase EZH2. Proc
Natl Acad Sci USA. 110:7922–7927. 2013. View Article : Google Scholar
|
|
49
|
Jung HY, Jun S, Lee M, Kim HC, Wang X, Ji
H and Park J II: PAF and EZH2 induce Wnt/β-catenin signaling
hyperactivation. Mol Cell. 52:193–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu Q, Wang G, Li Q, Jiang W, Kim JS, Wang
R, Zhu S, Wang X, Yan L, Yi Y, et al: Polycomb group proteins EZH2
and EED directly regulate androgen receptor in advanced prostate
cancer. Int J Cancer. 145:415–426. 2019. View Article : Google Scholar : PubMed/NCBI
|