Introduction
Myocardial ischemia is a major cause of
cardiovascular disease, and acute myocardial ischemia is the most
common form (1). Ischemia always
gives rise to oxidative stress (2,3).
Studies have demonstrated that sevoflurane preconditioning can
lessen myocardial oxidative stress and reduce myocardial damage
(4,5). Moreover, sevoflurane has been shown
to reduce the myocardial infarct size in animal models and in mice
with diabetes mellitus, as it protects the heart via
AMPK-independent activation (6,7).
Several randomized, controlled trials have demonstrated that
sevoflurane reduces the levels of biomarkers of myocardial injury
(6,8). In rat models of myocardial
ischemia-reperfusion, sevoflurane preconditioning has been shown to
alleviate ischemia-reperfusion damage by inhibiting transcription
factor SP1 (9). However, the
molecular mechanisms of sevoflurane preconditioning during
oxidative stress in cardiomyocytes remain unclear.
The thioredoxin interaction protein (TXNIP) is a
small-molecule protein with redox activity (5,10).
TXNIP can bind to various proteins, such as Trx, and has various
physiological functions, including regulating glucose metabolism
and angiogenesis, inducing oxidative stress, and promoting
apoptosis and inflammation (11-13). TXNIP plays a crucial role in
ameliorating oxidative injury in diabetic kidneys (14). The inhibition of TXNIP suppresses
oxidative stress and inflammation in lipopolysaccharide-induced
acute lung injury (15). However,
the role of TXNIP in the oxidative stress of cardiomyocytes remains
unclear, and no available studies to date have evaluated whether
TXNIP reduces oxidative stress following sevoflurane
preconditioning in cardiomyocytes, at least to the best of our
knowledge. In the present study, cardiomyocytes were preconditioned
with sevoflurane, and were then exposed oxygen-glucose deprivation
(OGD). Subsequently, the possible molecular mechanisms underlying
the protective effects of sevoflurane against oxidative stress were
investigated.
TXNIP exerts a regulatory effect on PKB (protein
kinase B/AKT) phosphorylation in pancreatic β-cells, and AKT
signaling can cause p27 downregulation in tumor cells (16,17). AKT is a serine/threonine protein
kinase that plays an important role in oxidative stress (18). AKT activation promotes
curcumin-mediated resistance to oxidative stress in neurons
(19). As a member of the
cyclin-dependent kinase inhibitor family, p27 (p27kip1) regulates
the cell cycle, apoptosis and cellular proliferation (20,21), and it reportedly induces oxidative
stress in liver cancer (22). The
inhibition of oxidative stress is accompanied by a decreased p27
expression (23), and p27 has
been demonstrated to protect cardiomyocytes from sepsis by
suppressing apoptosis (24).
The effect of p27 on oxidative stress in
cardiomyocytes remains unknown. It was hypothesized that TXNIP may
regulate p27 by activating AKT and then modulating oxidative stress
in cardiomyocytes that have been preconditioned with sevoflurane.
The present study focused on identifying the role and molecular
mechanisms of TXNIP by assessing the expression of TXNIP in
cardiomyocytes following sevoflurane preconditioning. The data
revealed that the downregulation of TXNIP protected H9c2 cells
against injury induced by oxidative stress by modulating AKT and
p27 following sevoflurane preconditioning.
Materials and methods
Cells and cell culture
H9c2 cardiomyocytes (ATCC) were cultured in
Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% fetal bovine serum, G418 (80 µg/ml)
and hygromycin B (80 µg/ml) in an incubator with 5%
CO2 at 37°C. The sevoflurane preconditioning of the H9c2
cells was achieved by culture with 0, 1.5, 2%, 3.5, 5 or 6%
sevoflurane dissolved in DMEM for 3 h. To inhibit AKT, H9c2 cells
were incubated with 10 µM of AKT inhibitor, LY294002 (S1105;
Selleck Chemicals) for 12 h.
OGD
H9c2 cells exposed to OGD were cultured with
Earle's balanced salt solution in a humid atmosphere (95%
N2 and 5% CO2) for 4 h and then cultured with
Earle's balanced salt solution supplemented with sugar in a
non-hypoxic atmosphere (95% air and 5% CO2) at 37°C for
9 h. Subsequently, all media from the H9c2 cells were changed to
DMEM and cultured in 5% CO2 at 37°C for 15 h, as
previously described (25). For
indicated experiments, cells were preconditioned with 0, 1.5, 2,
3.5, 5 or 6% sevoflurane for 3 h and then subjected to OGD.
Western blot analysis
Cells were treated with lysis buffer (Beyotime
Institute of Biotechnology). Proteins were extracted and then
quantified using a BCA kit (Perbio Science). Proteins (25
µg) were loaded and separated in 10% sodium dodecyl sulfate
polyacrylamide gel via electrophoresis. The separated proteins were
electro-transferred to a polyvinylidene-fluoride membrane (Bio-Rad
Laboratories, Inc.). The membrane was blocked with 5% non-fat milk
(Bio-Rad Laboratories, Inc.) in phosphate-buffered saline for 1 h
and blotted with anti-TXNIP (1:800, sc-166234), anti-pan-AKT
antibody (1:500, sc-5298), phosphor-AKT (Ser473; 1:800, sc-293125),
and anti-p27 (1:800, sc-56338) antibodies (all from Santa Cruz
Biotechnology Inc.). The membrane then was incubated overnight at
4°C, followed by exposure to horseradish-peroxidase-conjugated
secondary antibodies (1:1,000, sc-2005; Santa Cruz Biotechnology
Inc.) for 1.5 h at room temperature. GAPDH was used as the internal
control (1:2,000, sc-32233; Santa Cruz Biotechnology). The
immunoblot signals were developed using an enhanced
chemiluminescence kit (ECL kit, EMD Millipore) and the intensities
of each band were measured by ImageJ 1.4.1 software (NIH).
MTT and lactate dehydrogenase (LDH)
detection
Cells were cultured in a 96-well cell plate
(1x105 cells per well) in a
humidified atmosphere of 5% CO2 at 37°C for 12 h and
then subjected to a
3-(4,5-dimethyl-2-thiazolyl)-2,5-di-phenyl-2-H-tetrazolium bromide
(MTT) assay. Briefly, the old medium was replaced with fresh medium
plus 20 µl of MTT solution (0.5 mg/ml) and incubated at 37°C
for 5 h. The formazan crystals were dissolved using 160 µl
of dimethyl-sulfoxide per well. Optical density was measured at 490
nm. The solution was tested using a microplate analyzer model MR
600 (Dynatech Laboratories, Inc.). Using an LDH cytotoxicity
detection kit (Takara Biotechnology, Inc.), the membrane integrity
of the treated as was detected. Briefly, the transfected cells were
incubated with chrysophanol for 24 h, and LDH in the culture medium
was detected according to the standard instruction.
Caspase-3 activity assay
The caspase-3 activity of the cells was measured in
accordance with the instructions provided with the Caspase-3
activity assay kit (Shanghai Haoran Bio-Technology Co., Ltd.).
Measurement of reactive oxygen species
(ROS)
ROS release was determined using
2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA)
(Molecular Probes, Inc.) according to the specifications of the
Reactive Oxygen Species Assay kit. Briefly, the cells were
incubated with 60 µM of DCFH-DA at 37°C for 45 min in the
dark. Dichlorodihydrofluorescein fluorescence was determined using
a flow cytometer (FACSCalibur; BD Biosciences) with an excitation
wavelength of 485 nm and an emission wavelength of 530 nm.
Detection of the malondialdehyde (MDA)
concentration
The MDA concentration in the culture medium was
quantified using MDA assay kits (Beyotime Institute of
Biotechnology) according to the manufacturer's instructions.
The absorbance was determined using an ELISA reader (MRX Microplate
Reader, Dynatech Laboratories, Inc.) at 532 nm.
Plasmid construction and cell
transfection
The full-length DNA of the TXNIP gene (GenBank
accession no. NM023719) and p27 gene (GenBank accession no.
NM004064) were amplified from the cDNA of cardiomyocytes using PCR.
The DNA was subsequently digested with restriction endonucleases
EcoRI and BamHI and inserted into the pcDNA.3.1
(Invitrogen; Thermo Fisher Scientific, Inc.), which was cut with
the same enzymes. Following transduction into Escherichia
coli DH5α (Gibco; Thermo Fisher Scientific, Inc.), the
recombinant plasmid was amplified overnight at 37°C. The plasmids
were then sequenced, and the correct ones were designated as
pcDNA.3.1-TXNIP and pcDNA.3.1-p27.
TXNIP C247S-pcDNA3.1 was obtained by in vitro
mutagenesis using a Site Directed Mutagenesis kit (Beyotime
Institute of Biotechnology), and full-length TXNIP-pcDNA3.1 was
used as the template. All constructs were confirmed by
sequencing.
For cell transfection, the cells were seeded into
24-well plates in a humidified atmosphere with 5% CO2 at
37°C, in accordance with the manufacturer's instructions for
TurboFect (Thermo Fisher Scientific, Inc.). A total of 1.0
µg of the pcDNA.3.1-TXNIP, pcDNA.3.1-p27, mutant
pcDNA.3.1-TXNIP or pcDNA.3.1 were separately transfected into the
cells with 2 µl of TurboFect until cell fusion reached 80%.
The cells were then cultured in 5% CO2 at 37°C for 24
h.
Statistical analysis
Statistical analyses were performed using SPSS
version 22.0 software (SPSS, Inc.). The Mann-Whitney U test was
used to determine significant differences between 2 groups, and
one-way ANOVA followed by a Bonferroni test was used for multiple
groups. In all the figures, the data points and bar graphs
represent the means of independent biological replicates. The error
bars represent the standard deviation in graphs. A P-value <0.05
was considered to indicate a statistically significant difference.
Each experiment was performed at least in triplicate in 3
independent experiments.
Results
Sevoflurane preconditioning increases
cell viability, and inhibits apoptosis and LDH leakage in H9c2
cells exposed to OGD
The H9c2 cells were cultured with 0, 1.5, 2, 3.5, 5
or 6% sevoflurane in DMEM for 3 h, followed by incubation under OGD
conditions. Cell viability and LDH leakage were detected by an MTT
assay and an LDH cytotoxicity detection kit, respectively. The
results revealed that compared with the control (H9c2 cells without
sevoflurane and OGD treatment), OGD induced a decrease in cell
viability (Fig. 1A) and an
increase in LDH leakage (Fig. 1B)
and caspase-3 activity (Fig. 1C).
However, preconditioning with sevoflurane at 3.5, 5 and 6%
increased cell viability (Fig.
1A) and suppressed LDH leakage (Fig. 1B) compared to 0% sevoflurane
preconditioning. Caspase-3 activity also decreased by sevoflurane
preconditioning at 3.5, 5 and 6% compared with 0% sevoflurane
preconditioning (Fig. 1C).
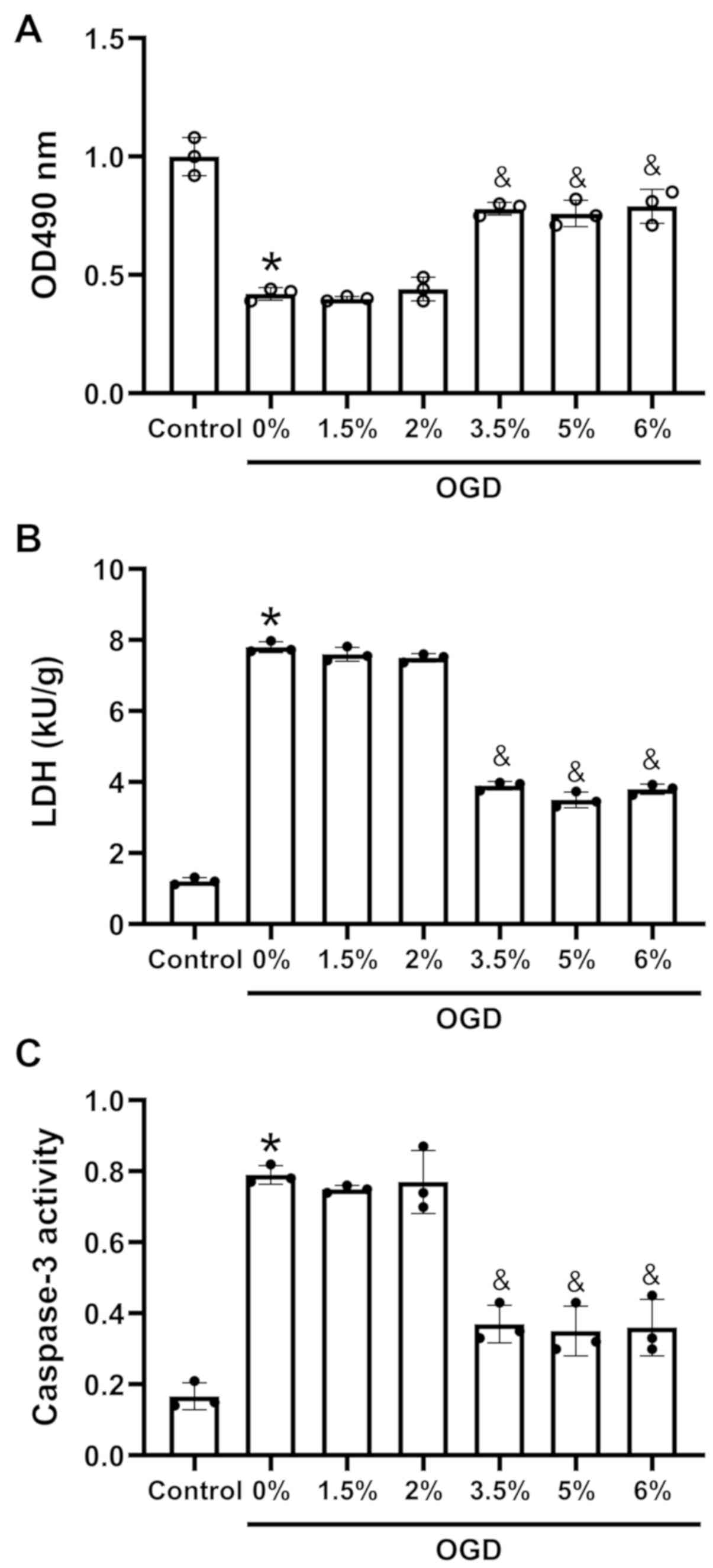 | Figure 1Effects of sevoflurane
preconditioning on cell viability, lactate dehydrogenase leakage
and apoptosis of H9c2 cells under exposed to OGD. (A) Cell
viability following sevoflurane preconditioning and OGD, as
measured by MTT assay. (B) LDH leakage following sevoflurane
preconditioning and oxygen and glucose deprivation, as measured
using an LDH cytotoxicity detection kit. (C) Apoptosis following
sevoflurane preconditioning and OGD, as measured by caspase-3
activity detection. H9c2 cells were cultured in 0, 1.5, 2, 3.5, 5
or 6% sevoflurane for 3 h and then exposed to OGD; n=3;
*P<0.01 vs. the control group,
&P<0.05 vs. the 0% group. OGD, oxygen-glucose
deprivation; LDH, lactate dehydrogenase. |
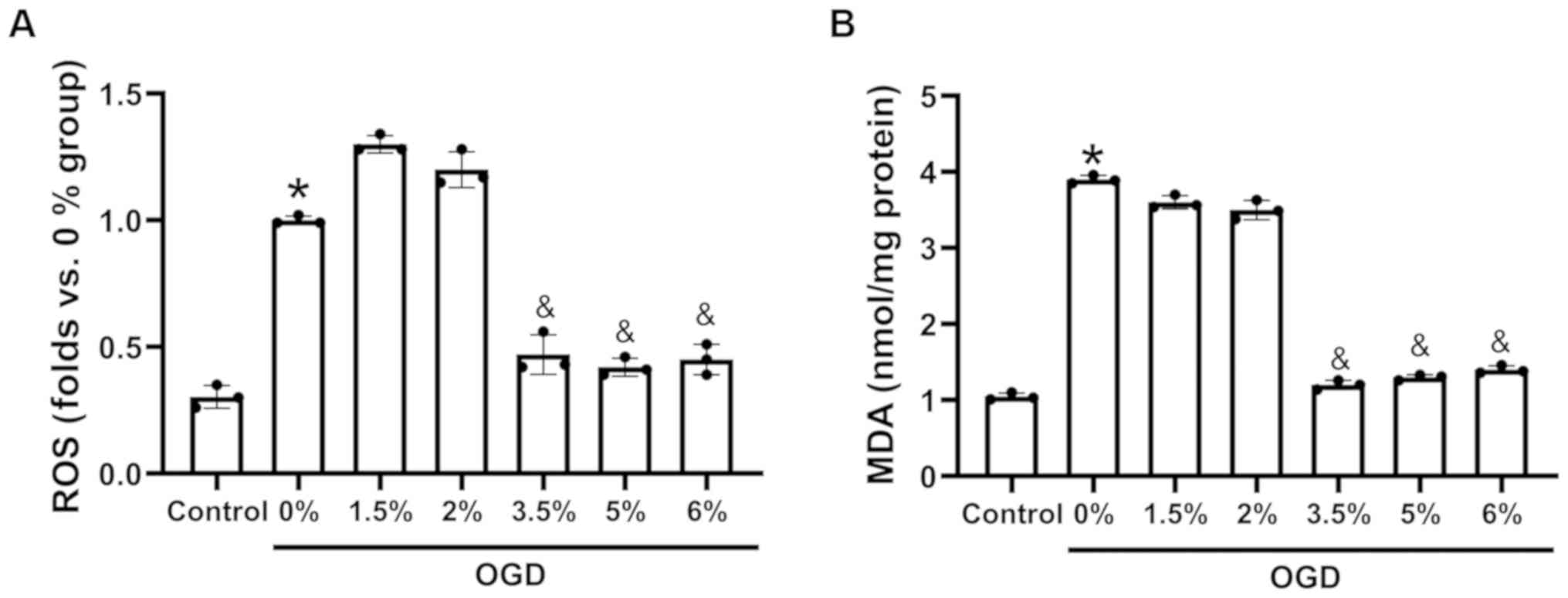
Sevoflurane preconditioning reduces ROS
production and the MDA content in H9c2 cells exposed to OGD
H9c2 cells were cultured under OGD conditions and
then exposed to 3 h of preconditioning with 0, 1.5, 2, 3.5, 5 or 6%
sevoflurane. The data demonstrated that OGD induced an increase in
ROS production (Fig. 2A) and the
MDA content (Fig. 2B). However,
sevoflurane preconditioning at 3.5, 5 and 6% significantly
decreased the ROS levels (Fig.
2A) and the MDA content (Fig.
2B), compared with 0% sevoflurane preconditioning. Thus,
sevoflurane preconditioning at 3.5% was selected for use in
subsequent experiments.
Sevoflurane preconditioning inhibits
TXNIP expression in H9c2 cells exposed to OGD
The protein expression of TXNIP in the H9c2 cells
exposed to sevoflurane preconditioning and OGD was assessed. It was
found that the protein expression (Fig. 3A) of TXNIP increased following
exposure to OGD compared with the control. However, TXNIP
expression decreased in the H9c2 cells subjected to sevoflurane
preconditioning compared with the cells not subjected to
sevoflurane preconditioning.
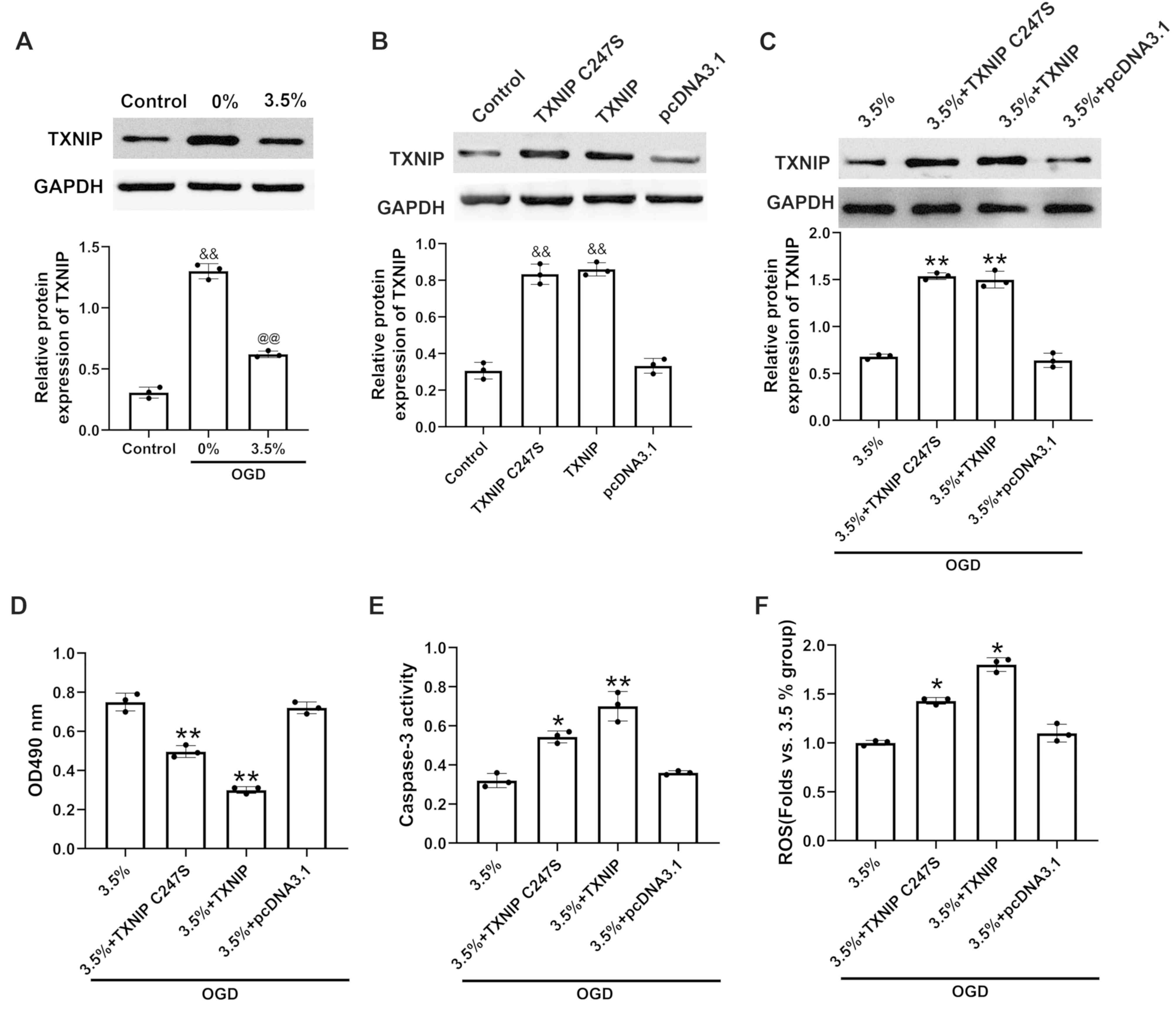 | Figure 3Effects of TXNIP upregulation on cell
viability, apoptosis and ROS in H9c2 cells exposed to sevoflurane
preconditioning and oxygen and glucose deprivation. (A) TXNIP
protein expression in H9c2 cells with sevoflurane preconditioning
and oxygen and glucose deprivation, as measured by western blot
analysis. (B) Relative protein expression of TXNIP following
transfection with pcDAN3.1-TXNIP in H9c2 cells under normal
conditions or (C) in H9c2 cells subjected to sevoflurane
preconditioning at 3.5% and OGD measured by western blot analysis.
(D) Cell viability following transfection in H9c2 cells subjected
to sevoflurane preconditioning at 3.5% and OGD, as measured by MTT
assay. (E) Apoptosis following transfection of H9c2 cells subjected
to sevoflurane preconditioning at 3.5% and OGD, as measured by
caspase-3 activity detection. (F) ROS levels following transfection
of H9c2 cells subjected to sevoflurane preconditioning at 3.5% and
OGD, as measured using a ROS assay kit. 3.5%+TXNIP, H9c2 cells
transfected with pcDNA.3.1-TXNIP, TXNIP C247S mutant or empty
plasmid pcDNA3.1 following sevoflurane preconditioning at 3.5% and
OGD; 3.5%+pcDNA.3.1, H9c2 cells transfected with pcDNA.3.1
following sevoflurane preconditioning at 3.5% and OGD; n=3,
&&P<0.01 vs. the control group,
@@P<0.01 vs. the 0% group; *P<0.05 and
**P<0.01 vs. the 3.5% + pcDNA.3.1 group. TXNIP,
thioredoxin interaction protein; OGD, oxygen-glucose deprivation;
ROS, reactive oxygen species. |
TXNIP elevation promotes injury in H9c2
cells exposed to sevoflurane preconditioning and OGD
TXNIP was overexpressed by transfection with
pcDNA.3.1-TXNIP or a C247S TXNIP mutant plasmid under normal
conditions or following sevoflurane preconditioning and OGD. The
data indicated that the protein expression of TXNIP (Fig. 3B and C) following transfection was
significantly upregulated in the H9c2 cells. It was also found that
the overexpression of wild-type TXNIP or the C247S mutant TXNIP
significantly decreased cell viability (Fig. 3D) and significantly elevated
caspase-3 activity (Fig. 3E) and
ROS levels (Fig. 3F).
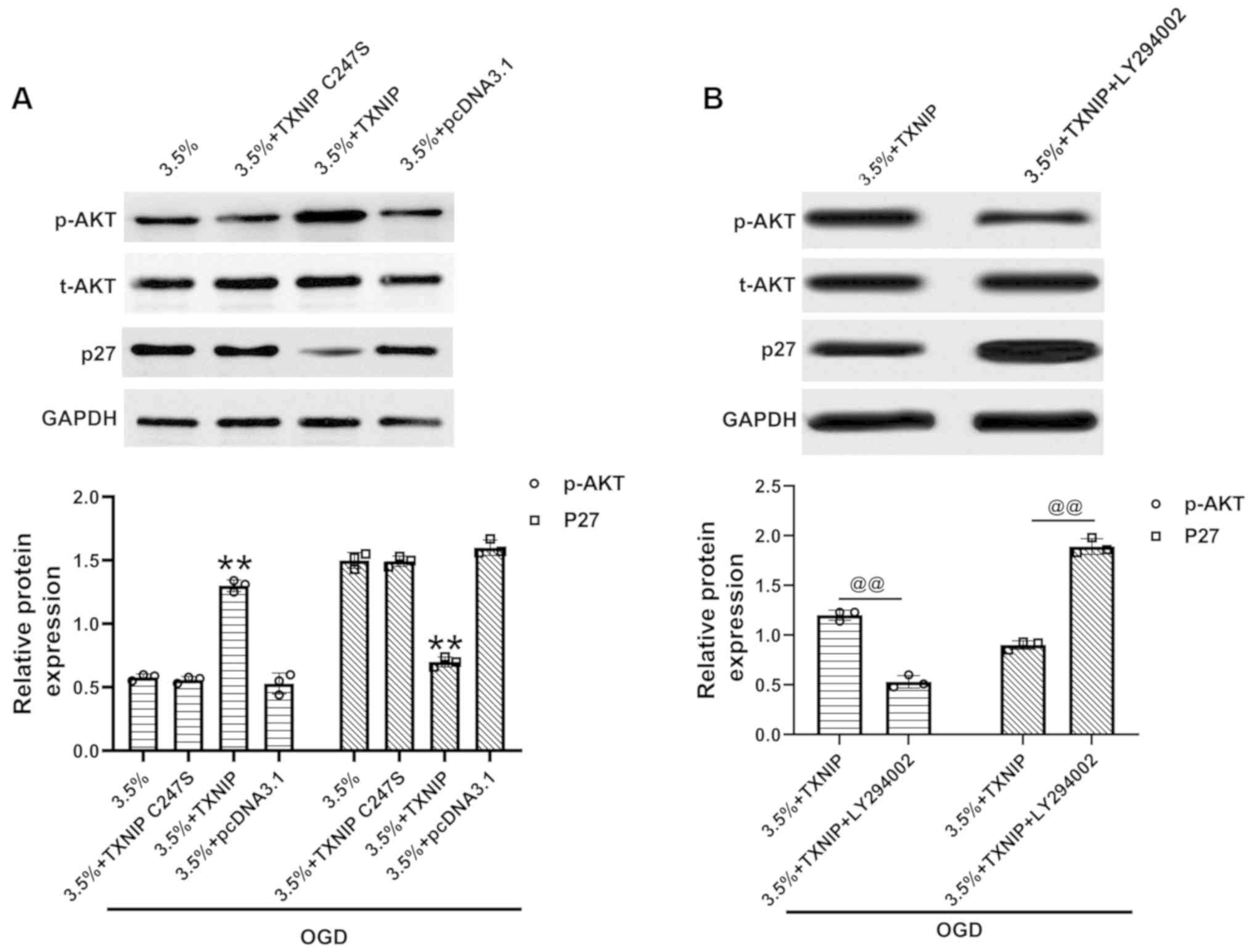
TXNIP regulates p27 by modulating AKT
activation in H9c2 cells exposed to sevoflurane preconditioning and
OGD
AKT phosphorylation and p27 expression were measured
following the overexpression of TXNIP in H9c2 cells subjected to
sevoflurane preconditioning and OGD. The data demonstrated the
elevated phosphorylation of AKT and the decreased expression of p27
following the overexpression of wild-type TXNIP (Fig. 4A), while the overexpression of the
C247S TXNIP mutant exhibited no interaction with AKT (Fig. 4A). Incubation with the AKT
inhibitor, LY294002 (10 µM) (26,27), suppressed AKT phosphorylation and
upregulated p27 expression (Fig.
4B), indicating that the inhibition of AKT activation reversed
the effects of TXNIP on p27 expression in these cells. Thus, TXNIP
regulates p27 expression via AKT activation in H9c2 cells exposed
to sevoflurane preconditioning and OGD.
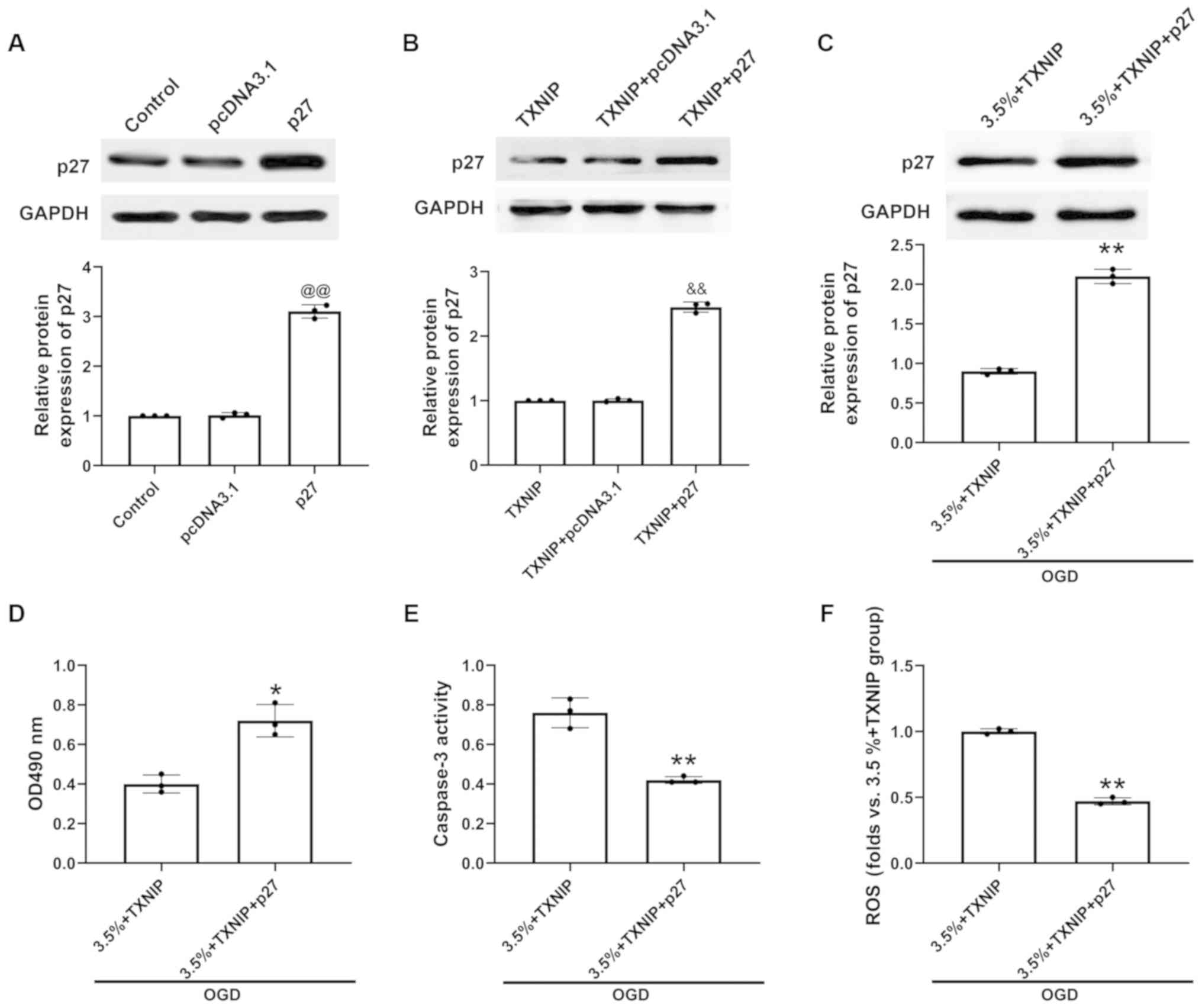
TXNIP expression mediated by sevoflurane
preconditioning protects H9c2 cells against injury induced by OGD
by modulating p27 expression
The p27 transfection efficiency was examined by
western blot analysis. The results revealed that compared with the
pcDNA3.1-transfected cells, the level of p27 was markedly increased
in the p27-transfected H9c2 cells (Fig. 5A). In the H9c2 cells or H9c2 cells
with sevoflurane preconditioning and OGD exposure, p27
overexpression was induced by pcDNA.3.1-p27 transfection following
the overexpression of TXNIP. It was found that compared with the
TXNIP- and pcDNA3.1-co-transfected cells, the level of p27 was
markedly increased in the TXNIP- and p27-co-transfected H9c2 cells,
indicating the successful overexpression of p27 (Fig. 5B). In addition, p27 protein
expression increased in the H9c2 cells subjected to sevoflurane
preconditioning and OGD exposure (Fig. 5C). The upregulation of p27
promoted cell viability (Fig.
5D), and inhibited caspase-3 activity (Fig. 5E) and ROS production (Fig. 5F) to a greater extent in
TXNIP-overexpressing H9c2 cells subjected to sevoflurane
preconditioning and OGD injury than in H9c2 cells subjected to
sevoflurane preconditioning and OGD exposure only. These results
indicated that sevoflurane preconditioning protects H9c2 cells
against injury induced by OGD by modulating TXNIP, AKT activation
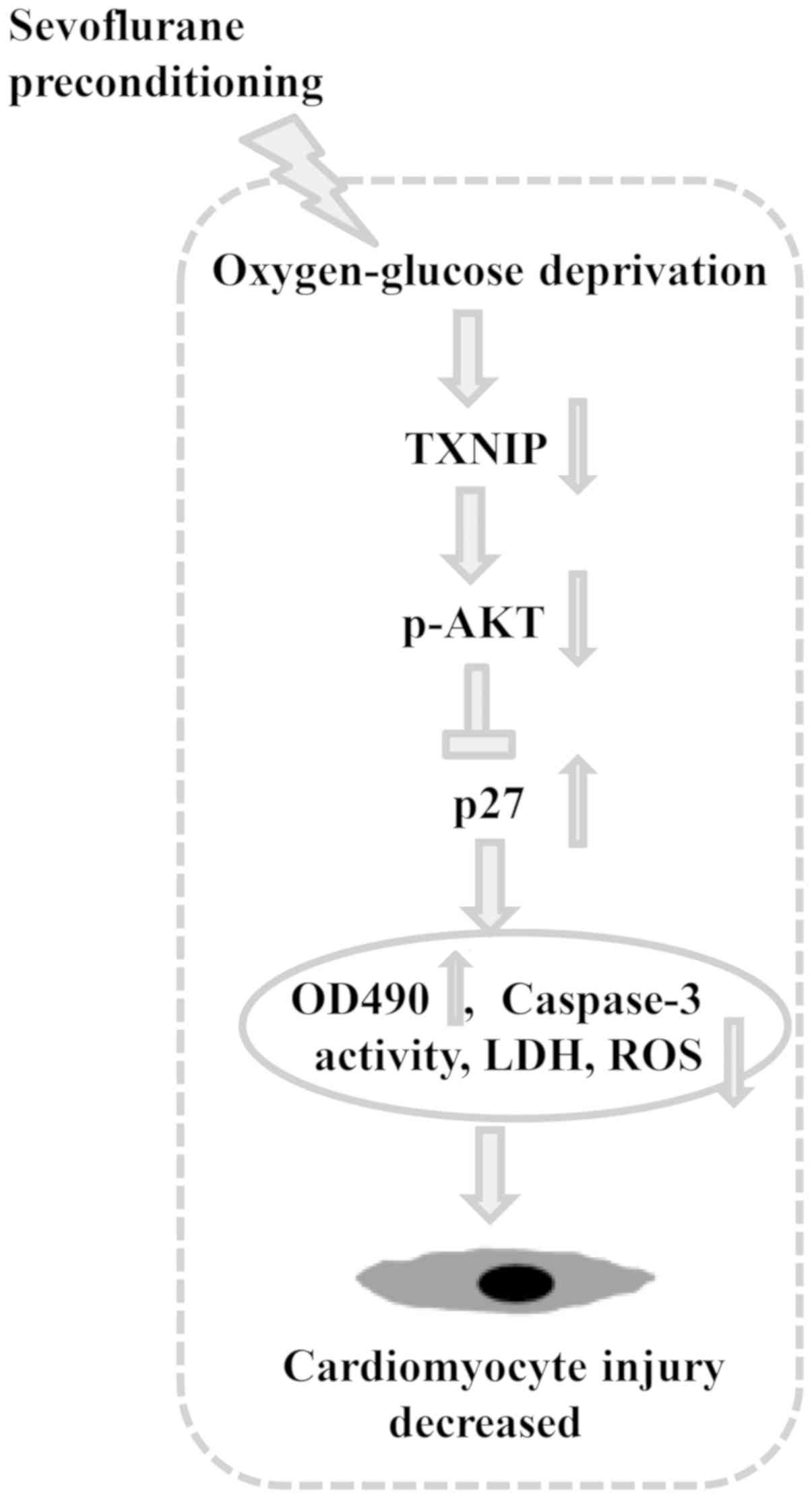
and p27 signaling (Fig. 6).
Discussion
Myocardial ischemia induces oxidative stress and
widespread damage to cells (28-30). Sevoflurane preconditioning may
protect the heart against this type of injury (31,32). In the present study, cells were
incubated with 0, 1.5, 2, 3.5, 5 or 6% sevoflurane and then exposed
to OGD. The data indicated that sevoflurane preconditioning at 3.5%
markedly inhibited caspase-3 activity, LDH leakage, and MDA and ROS
production, and increased cell viability. The molecular mechanisms
underlying the protective effects of sevoflurane preconditioning
were then investigated.
TXNIP plays a role in salidroside-mediated
protection against high-glucose-induced oxidative stress in rat
glomerular mesangial cells (33).
Hou et al demonstrated that the inhibition of TXNIP
suppressed lipopolysaccharide-induced oxidative stress and the
apoptosis of vascular endothelial cells (34). TXNIP overexpression has also been
shown to induce cardiomyocyte apoptosis and injury (35). Currently, to the best of our
knowledge, there is no available study to date on the role of TXNIP
in the OGD-induced oxidative stress of cardiomyocytes following
sevoflurane preconditioning. In the present study, the results
indicated that sevoflurane preconditioning at 3.5% suppressed TXNIP
expression in cardiomyocytes exposed to OGD. Furthermore, the
overexpression of TXNIP by pcDNA.3.1-TXNIP transfection
significantly increased caspase-3 activity and ROS production, and
it decreased the viability of cardiomyocytes subjected to
sevoflurane preconditioning at 3.5% and OGD exposure. Previous
studies have reported that N-methyl-D-aspartic acid (NMDA) receptor
is an important target for analgesia of sevoflurane (36). In addition, the blockade of NMDA
receptor has been shown to dephosphorylate Forkhead box O1 (FOXO1)
at Thr24 and induce its nuclear translocation, thus increasing the
transcription of TXNIP (37).
NMDA receptor blockade upregulates TXNIP, where it binds
thioredoxin and boosts antioxidant defenses via the
thioredoxin-peroxiredoxin system (38). Thus, it was hypothesized that
sevoflurane inhibits TXNIP expression through the NMDA receptor and
FOXO1. The present study, to the best of our knowledge, is the
first to demonstrate that the downregulation of TXNIP is closely
related to the protective effects of sevoflurane preconditioning
against oxidative stress in cardiomyocytes.
TXNIP regulates AKT phosphorylation in pancreatic
β-cells, and AKT signaling can downregulate p27 in tumor cells
(16,17). Mahmoud et al demonstrated
that AKT assists in the prevention of lipid-induced endothelial
damage and oxidative stress (39). Zeng et al found that AKT is
crucial in alleviating H2O2-induced oxidative
stress caused by thyroid hormones in cardiomyocytes (40). In addition, p27 reportedly lessens
the toxic effects of β-amyloid 42 during oxidative stress (41). Zhao et al demonstrated that
p27 protects cardiomyocytes from sepsis by inhibiting apoptosis
(24).
The effects of p27 on oxidative stress in
cardiomyocytes subjected to sevoflurane preconditioning and OGD
exposure remain unclear, however. It was hypothesized that TXNIP
would regulate OGD-induced oxidative stress in cardiomyocytes
subjected to sevoflurane preconditioning via AKT/p27 signaling. In
the present study, TXNIP overexpression elevated the
phosphorylation of AKT and decreased the expression of p27 in H9c2
cells exposed to sevoflurane preconditioning and OGD. The
suppression of AKT phosphorylation by LY294002 (10 µM)
markedly increased the expression of p27 in H9c2 cells with TXNIP
overexpression. The results indicated that TXNIP regulated p27
expression via AKT in cardiomyocytes exposed to sevoflurane
preconditioning and OGD. Moreover, p27 over-expression induced by
pcDNA.3.1-p27 promoted cell viability and inhibited caspase-3
activity and ROS production, indicating that p27 overexpression
abolished the effects of TXNIP on cell viability, caspase-3 and ROS
in H9c2 cells subjected to sevoflurane preconditioning and OGD
exposure. Therefore, it can be concluded that following sevoflurane
preconditioning, sevoflurane protected cardiomyocytes against
OGD-induced oxidative stress by regulating TXNIP/p27
expression.
In conclusion, the present study found that
sevoflurane preconditioning at 3.5% significantly inhibited TXNIP
expression, caspase-3 activity, LDH leakage, and MDA and ROS
production, and it increased the viability of H9c2 cells that were
exposed to OGD. TXNIP upregulation effectively reversed the effects
of sevoflurane preconditioning at 3.5% on caspase-3 activity, ROS
production and cell viability. Moreover, the results indicated that
TXNIP regulated p27 expression via AKT. Thus, sevoflurane
preconditioning may protect cardiomyocytes against oxidative
stress, and this effect may be modulated by TXNIP, AKT and p27
signaling (Fig. 6).
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
All authors (MM, RL, WS, QW, HongY and HongmeiY)
participated in the design, interpretation of the studies and
analysis of the data and review of the manuscript. MM designed,
prepared and performed the experiments. RL, WS, QW, HongY and
HongmeiY contributed to the provision of
reagents/materials/analysis tools. MM wrote the manuscript, and
modified and revised the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
TXNIP
|
thioredoxin interaction protein
|
|
AKT/PKB
|
protein kinase B
|
|
OGD
|
oxygen-glucose deprivation
|
|
LDH
|
lactate dehydrogenase
|
|
DCFH-DA
|
2',7'-dichlorodihydrofluorescein diacetate
|
|
MTT
|
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H- tetrazolium
bromide
|
|
MDA
|
malondialdehyde
|
Acknowledgments
Not applicable.
References
|
1
|
Neri M, Riezzo I, Pascale N, Pomara C and
Turillazzi E: Ischemia/reperfusion injury following acute
myocardial infarction: A critical issue for clinicians and forensic
pathologists. Mediators Inflamm. 2017:70183932017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li P, Stetler RA, Leak RK, Shi Y, Li Y, Yu
W, Bennett MVL and Chen J: Oxidative stress and DNA damage after
cerebral ischemia: Potential therapeutic targets to repair the
genome and improve stroke recovery. Neuropharmacology. 134:208–217.
2018. View Article : Google Scholar
|
|
3
|
Cheng YC, Sheen JM, Hu WL and Hung YC:
Polyphenols and oxidative stress in atherosclerosis-related
ischemic heart disease and stroke. Oxid Med Cell Longev.
2017:85264382017. View Article : Google Scholar
|
|
4
|
Li YQ, Zhan ZL and Li QF: Exploration of
protective effect of Sevoflurane preconditioning on hypoxia
reoxygenation injury of myocardial cells in rats and related
molecular mechanisms. J Hainan Med Univ. 22:8–11. 2016.
|
|
5
|
Wen T, Wang L, Sun XJ, Zhao X, Zhang GW
and Ling LJ: Sevoflurane preconditioning promotes activation of
resident CSCs by transplanted BMSCs via miR-210 in a rat model for
myocardial infarction. Oncotarget. 8:114637–114647. 2017.
View Article : Google Scholar
|
|
6
|
Pagel PS and Crystal GJ: The discovery of
myocardial preconditioning using volatile anesthetics: A history
and contemporary clinical perspective. J Cardiothorac Vasc Anesth.
32:1112–1134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie D, Zhao J, Guo R, Jiao L, Zhang Y, Lau
WB, Lopez B, Christopher T, Gao E, Cao J, et al: Sevoflurane
pre-conditioning ameliorates diabetic myocardial
ischemia/reperfusion injury via differential regulation of p38 and
ERK. Sci Rep. 10:232020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kunst G and Klein AA: Peri-operative
anaesthetic myocardial preconditioning and protection - cellular
mechanisms and clinical relevance in cardiac anaesthesia.
Anaesthesia. 70:467–482. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang SB, Liu TJ, Pu GH, Li BY, Gao XZ and
Han XL: MicroRNA-374 exerts protective effects by inhibiting SP1
through activating the PI3K/Akt pathway in rat models of myocardial
ischemia-reperfusion after sevoflurane preconditioning. Cell
Physiol Biochem. 46:1455–1470. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Huang J, Yang X, Sun X, Xu Q,
Wang B, Zhong P and Wei X: Altered Expression of TXNIP in the
peripheral leukocytes of patients with coronary atherosclerotic
heart disease. Medicine (Baltimore). 96:e91082017. View Article : Google Scholar
|
|
11
|
Wang CY, Xu Y, Wang X, Guo C, Wang T and
Wang ZY: Dl-3-n-butylphthalide inhibits NLRP3 inflammasome and
mitigates alzheimer's-Like pathology via Nrf2-TXNIP-TrX axis.
Antioxid Redox Signal. 30:1411–1431. 2019. View Article : Google Scholar
|
|
12
|
Friedemann T, Schumacher U, Tao Y, Leung
AK and Schröder S: Neuroprotective activity of coptisine from
coptis chinensis (Franch). Evid Based Complement Alternat Med.
2015:8273082015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ye X, Zuo D, Yu L, Zhang L, Tang J, Cui C,
Bao L, Zan K, Zhang Z, Yang X, et al: ROS/TXNIP pathway contributes
to thrombin induced NLRP3 inflammasome activation and cell
apoptosis in microglia. Biochem Biophys Res Commun. 485:499–505.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ji L, Wang Q, Huang F, An T, Guo F, Zhao
Y, Liu Y, He Y, Song Y and Qin G: FOXO1 overexpression attenuates
tubulointerstitial fibrosis and apoptosis in diabetic kidneys by
ameliorating oxidative injury via TXNIP-TRX. Oxid Med Cell Longev.
2019:32869282019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han X, Wu YC, Meng M, Sun QS, Gao SM and
Sun H: Linarin prevents LPSinduced acute lung injury by suppressing
oxidative stress and inflammation via inhibition of TXNIP/NLRP3 and
NFκB pathways. Int J Mol Med. 42:1460–1472. 2018.PubMed/NCBI
|
|
16
|
Chen J, Hui ST, Couto FM, Mungrue IN,
Davis DB, Attie AD, Lusis AJ, Davis RA and Shalev A:
Thioredoxin-interacting protein deficiency induces Akt/Bcl-xL
signaling and pancreatic beta-cell mass and protects against
diabetes. FASEB J. 22:3581–3594. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Narita Y, Nagane M, Mishima K, Huang HJ,
Furnari FB and Cavenee WK: Mutant epidermal growth factor receptor
signaling down-regulates p27 through activation of the
phosphatidylinositol 3-kinase/Akt pathway in glioblastomas. Cancer
Res. 62:6764–6769. 2002.PubMed/NCBI
|
|
18
|
Oyagbemi AA, Omobowale TO, Asenuga ER,
Ochigbo GO, Adejumobi AO, Adedapo AA and Yakubu MA: Sodium
arsenite-induced cardiovascular and renal dysfunction in rat via
oxidative stress and protein kinase B (Akt/PKB) signaling pathway.
Redox Rep. 22:467–477. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui Q, Li X and Zhu H: Curcumin
ameliorates dopaminergic neuronal oxidative damage via activation
of the Akt/Nrf2 pathway. Mol Med Rep. 13:1381–1388. 2016.
View Article : Google Scholar
|
|
20
|
Wang L, Shen S, Xiao H, Ding F, Wang M, Li
G and Hu F: ARHGAP24 inhibits cell proliferation and cell cycle
progression and induces apoptosis of lung cancer via a
STAT6-WWP2-P27 axis. Carcinogenesis (bjz144). Aug 20–2019.Epub
ahead of print. View Article : Google Scholar
|
|
21
|
Wang Z, Yu C and Wang H: HOXA5 inhibits
the proliferation and induces the apoptosis of cervical cancer
cells via regulation of protein kinase B and p27. Oncol Rep.
41:1122–1130. 2019.
|
|
22
|
Williams V, Brichler S, Khan E, Chami M,
Dény P, Kremsdorf D and Gordien E: Large hepatitis delta antigen
activates STAT-3 and NF-κB via oxidative stress. J Viral Hepat.
19:744–753. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Smith RS Jr, Agata J, Xia CF, Chao L and
Chao J: Human endothelial nitric oxide synthase gene delivery
protects against cardiac remodeling and reduces oxidative stress
after myocardial infarction. Life Sci. 76:2457–2471. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao X, Qi H, Zhou J, Xu S and Gao Y: P27
protects cardiomyocytes from sepsis via activation of autophagy and
inhibition of apoptosis. Med Sci Monit. 24:8565–8576. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma L, Liu H, Xie Z, Yang S, Xu W, Hou J
and Yu B: Ginsenoside Rb3 protects cardiomyocytes against
ischemia-reperfusion injury via the inhibition of JNK-mediated
NF-kappaB pathway: A mouse cardiomyocyte model. PLoS One.
9:e1036282014. View Article : Google Scholar
|
|
26
|
Kim DE, Kim B, Shin HS, Kwon HJ and Park
ES: The protective effect of hispidin against hydrogen
peroxide-induced apoptosis in H9c2 cardiomyoblast cells through
Akt/GSK-3β and ERK1/2 signaling pathway. Exp Cell Res. 327:264–275.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jun HO, Kim DH, Lee SW, Lee SH, Seo JH,
Kim JH, Kim JH, Yu YS, Min BH and Kim KW: Clusterin protects H9c2
cardiomyocytes from oxidative stress-induced apoptosis via
Akt/GSK-3β signaling pathway. Exp Mol Med. 43:53–61. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhai M, Li B, Duan W, Jing L, Zhang B,
Zhang M, Yu L, Liu Z, Yu B, Ren K, et al: Melatonin ameliorates
myocardial ischemia reperfusion injury through SIRT 3-dependent
regulation of oxidative stress and apoptosis. J Pineal Res.
63:e124192017. View Article : Google Scholar
|
|
29
|
Zhao D, Yang J and Yang L: Insights for
oxidative stress and mTOR signaling in myocardial
ischemia/reperfusion injury under diabetes. Oxid Med Cell Longev.
2017:64374672017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu L, Li Q, Yu B, Yang Y, Jin Z, Duan W,
Zhao G, Zhai M, Liu L, Yi D, et al: Berberine attenuates myocardial
ischemia/reperfusion injury by reducing oxidative stress and
inflammation response: Role of silent information regulator 1. Oxid
Med Cell Longev. 2016:16896022016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qian B, Yang Y, Yao Y, Liao Y and Lin Y:
Upregulation of vascular endothelial growth factor receptor-1
contributes to sevoflurane preconditioning-mediated
cardioprotection. Drug Des Devel Ther. 12:769–776. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pasqualin RC, Mostarda CT, de Souza LE,
Vane MF, Sirvente R, Otsuki DA, Torres MLA, Irigoyen MCC and Auler
JOC Jr: Sevoflurane preconditioning during myocardial
ischemia-reperfusion reduces infarct size and preserves autonomic
control of circulation in rats. Acta Cir Bras. 31:338–345. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang S, Zhao X, Yang S, Chen B and Shi J:
Salidroside alleviates high glucose-induced oxidative stress and
extracellular matrix accumulation in rat glomerular mesangial cells
by the TXNIP-NLRP3 inflammasome pathway. Chem Biol Interact.
278:48–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hou X, Yang S and Yin J: Blocking the
REDD1/TXNIP axis ameliorates LPS-induced vascular endothelial cell
injury through repressing oxidative stress and apoptosis. Am J
Physiol Cell Physiol. 316:C104–C110. 2019. View Article : Google Scholar
|
|
35
|
Yao YL, Yang X, Xue XW, Fan LF and Jiao
XY: Effect of adenovirus-mediated TXNIP overexpression on apoptosis
and injury of H9C2 cardiomyocytes. Sheng Li Xue Bao. 65:309–318.
2013.In Chinese. PubMed/NCBI
|
|
36
|
Petrenko AB, Yamakura T, Sakimura K and
Baba H: Defining the role of NMDA receptors in anesthesia: Are we
there yet? Eur J Pharmacol. 723:29–37. 2014. View Article : Google Scholar
|
|
37
|
Yamaguchi F, Hirata Y, Akram H, Kamitori
K, Dong Y, Sui L and Tokuda M: FOXO/TXNIP pathway is involved in
the suppression of hepatocellular carcinoma growth by glutamate
antagonist MK-801. BMC cancer. 13:4682013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Papadia S, Soriano FX, Léveillé F, Martel
M-A, Dakin KA, Hansen HH, Kaindl A, Sifringer M, Fowler J,
Stefovska V, et al: Synaptic NMDA receptor activity boosts
intrinsic antioxidant defenses. Nature neuroscience. 11:476–487.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mahmoud AM, Wilkinson FL, McCarthy EM,
Moreno-Martinez D, Langford-Smith A, Romero M, Duarte J and
Alexander MY: Endothelial microparticles prevent lipid-induced
endothelial damage via Akt/eNOS signaling and reduced oxidative
stress. FASEB J. 31:4636–4648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zeng B, Liu L, Liao X, Zhang C and Ruan H:
Thyroid hormone protects cardiomyocytes from
H2O2 -induced oxidative stress via the
PI3K-AKT signaling pathway. Exp Cell Res. 380:205–215. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hussien-Ali A and Alifragis P: P27
Minimising the toxic effects of beta-Amyloid 42 on oxidative
stress. Biochem Pharmacol. 139:133–134. 2017. View Article : Google Scholar
|