Introduction
Breast cancer is the most common malignant tumor
among women worldwide and represents one of the major life
threatening diseases in women (1). In 2019, ~268,600 new cases of breast
cancer were diagnosed and 41,760 women died from this disease in US
(2). Breast cancer mostly occurs
in women between the ages of 55 and 60 years (1). Long menstruation, no history of
pregnancy, long-term use of hormonal contraceptives and obesity are
the main factors contributing to breast cancer occurrence (1). Epidemiological studies reported that
abnormal amplification of certain genes and mutations as well as
genetic susceptibility modifications also serve crucial roles in
the development of breast cancer (3,4).
It has been reported that 5-10% of breast cancer cases are due to
genetic disorders, such as BRCA1 and BRCA2 mutations, which are
associated with the hereditary breast-ovarian cancer syndrome
(5). According to global
statistics released by the International Agency for Research on
Cancer in 2018, the incidence of breast cancer accounted for 11.6%
of all cancers and represented the second most common cancer
worldwide after lung cancer (6).
In addition, breast cancer is a leading cause of cancer-associated
mortality in women. In addition, the incidence of breast cancer has
significantly grown worldwide, and has increased by ~10 times in
the last decade (7). It is
therefore urgent to determine some genes that could be associated
with the proliferation, metastasis and apoptosis of breast cancer
cells, in order to develop novel treatment.
With the continuous expansion of the Human Genome
Project and the development of innovative technologies, including
transcriptome and high-throughput gene sequencing, genetic
mysteries have been gradually revealed. In 2012, the Encyclopedia
of DNA Elements project has confirmed that non-coding RNAs are no
longer RNAs of low interest, but are instead largely involved in
numerous biological processes, including cell proliferation,
differentiation and apoptosis (8). Long non-coding RNAs (lncRNAs) are
defined as transcripts of >200 nucleotides in length that are
not translated into proteins and that account for much of the
transcribed genome. Previous studies reported that lncRNAs serve
crucial roles in the development, diagnosis and treatment of
certain types of tumor, including non-small cell lung cancer and
colorectal cancer (9-12). Tissue differentiation inducing
non-protein coding RNA (TINCR) is a key lncRNA of ~3,700 bp in
length, which induces human epidermal differentiation following
transcription (13). TINCR is a
human skin disease-mutated gene, and its deletion can cause
epidermal formation disorder (13). The mechanism of TINCR action is
related to the binding of staufen1 (STAU1) protein and the
stabilization of relevant differentiation genes mRNAs (14). It has been demonstrated that TINCR
is abnormally expressed in a variety of malignant tumors, including
gastric cancer (15), colon
cancer (16), bladder cancer
(17) and hepatocellular
carcinoma (18), indicating that
TINCT might be used as a biomarker for early tumor diagnosis. Gao
et al (19) reported that
the transcription factor SP1-induced upregulation of TINCR inhibits
cell migration and invasion by regulating miR-107 and miR-1286
expression in lung adenocarcinoma. Furthermore, TINCR interacts
with miR-335, and silencing TINCR inhibits epithelial ovarian
cancer progression in vitro and in vivo by reducing
fibroblast growth factor 2 expression (20). A meta-analysis demonstrated that
TINCR overexpression might increase tumor size and worsen prognosis
of patients with cancer (such as breast cancer and liver cancer)
(21). In breast cancer,
activation of TINCR by H3K27 acetylation promotes cell resistance
to trastuzumab and epithelial-mesenchymal transition by targeting
miR-125b (22). A previous study
reported that upregulation of the competing endogenous RNA TINCR by
transcription factor SP1 contributes to the tumorigenesis of breast
cancer (23). Furthermore, the
results from co-expression network analysis reported that TINCR
expression is associated with breast cancer prognosis (24). By using the Gene Expression
Omnibus (GEO) and The Cancer Genome Atlas (TCGA), it has been
demonstrated that TINCR is significantly elevated in breast cancer
cells (25). However, the
underlying mechanisms remain unknown, and treatment for patients
with breast cancer must be developed.
MicroRNAs (miRNAs) are a class of highly conserved
single-stranded non-coding small RNAs that serve crucial roles in
the growth and development of organisms (26). Although miRNAs account for only
~2% of human genomes, they can regulate ~21,000 protein-coding
genes (27). In-depth study of
miRNAs will benefit cancer treatment and prognosis, by allowing
early clinical diagnosis and therefore suggesting the most
appropriate treatment for patients with breast cancer. Previous
studies reported that TINCR can sponge certain miRNAs to promote
cancer progression. For example, TINCR has been demonstrated to
sponge miR-214-5p in order to upregulate Rho associated coiled-coil
containing protein kinase 1 in hepatocellular carcinoma cells,
leading therefore to the promotion cancer cell invasion and
migration (18). Chen et
al (28) reported that TINCR
sponges miR-375 to upregulate pyruvate dehydrogenase kinase 1 that
leads to gastric cancer progression. The present study used the
breast cancer cell lines MCF-7 and MDA-MB-231 in order to explore
the targeted relationship between TINCR and miR-589-3p, and to
determine the underlying mechanism of miR-589-3p in breast cancer.
The findings from this study may provide a reliable experimental
basis for miRNA treatment of breast cancer.
Materials and methods
Cell culture
The MCF-7 (HTB-22) and MDA-MB-231 (HTB-26) cell
lines used in the present study were purchased from the American
Type Culture Collection. The cells were cultured in DMEM-H medium
(cat. no. 12110-500; Beijing Solarbio Science & Technology Co.,
Ltd.) containing 10% fetal bovine serum (FBS; cat. no. 0025;
ScienCell Research Laboratories, Inc.) and placed at 37°C in a
humidified incubator containing 5% CO2.
Specimen collection form patients with
breast cancer
Breast cancer and adjacent normal tissues of 68
patients with breast cancer (age range, 26-58 years; mean age, 42
years) treated at the Affiliated Zhongshan Hospital of Dalian
University for breast surgery were collected between January 2012
and June 2018, whether they had received chemotherapy or not. Fresh
specimens obtained during the surgery were immediately washed with
physiological saline, frozen in liquid nitrogen and stored at
−80°C. The present study has been approved by the Ethics Committee
of Affiliated Zhongshan Hospital of Dalian University (approval no.
201112013RXW) and all patients signed informed consent.
Target gene prediction
The miRNAs targeted by TINCR were predicted by the
StarBase database (http://starbase.sysu.edu.cn/index.php). TargetScan
database (http://www.targetscan.org) was used
to predict targeted genes for miRNAs.
Cell transfection
TINCR overexpression vector was constructed using
pcDNA3.2 (cat. no. 12489019; Gibco; Thermo Fisher Scientific,
Inc.). The primer sequences for PCR of TINCR cDNA were as follows:
TINCR, forward 5′-CCC AAG CTTG GTC TGG GCT CCC AGG TGG ACC-3′ and
reverse 5′-AGC GAT ATC CTA TAG TTG TTT TCA AAC ATG TAA TCTT-3′.
After obtaining the full-length sequence of TINCR by reverse
transcription-quantitative (RT-q)PCR, the TINCR gene was cloned
into the pcDNA3.2 to obtain pc-TINCR. MCF-7 and MDA-MB-231 cells in
the logarithmic growth stage were seeded in 6-well plates at the
density of 2×105 cells/well one day prior to
transfection. Cell transfection was performed when cells reached
70~80% confluence by using LipofectamineTM 3000 kit
(cat. no. L3000015; Thermo Fisher Scientific, Inc.). Briefly, 1
µg of pc-TINCR, pcDNA3.2 vector, TINCR small-interfering RNA
(siTINCR; cat. no. SR316884; Origene), negative control siRNA (cat.
no. SR30004; Origene), miR-589-3p mimic (cat. no. 4464066; Thermo
Fisher Scientific, Inc.), miR-589-3p inhibitor (cat. no. 4464084,
Thermo Fisher Scientific, Inc.), mimic control (cat. no. 4464058;
Thermo Fisher Scientific, Inc.), inhibitor control (cat. no.
4464076; Thermo Fisher Scientific, Inc.), IGF1R (cat. no. RC214928;
Origene) or IGF1R small interfering RNA (siIGF1R; cat. no.
siB05109161217-1-5; RiboBio) were diluted in 50 µl of
Opti-MEM® medium (cat. no. 31985062; Invitrogen; Thermo
Fisher Scientific, Inc.). In addition, 3 µl
LipofectamineTM 3000 reagent was diluted in 50 µl
Opti-MEM. The two solutions were then mixed together and were
incubated for 15 min at room temperature. Subsequently, pc-TINCR,
pcDNA3.2 vector, miR-589-3p mimic, mimic control or siIGF1R was
added to MCF-7 cells seeded in a 6-well plate at the density of
5×105/well (500 µl). Furthermore, siTINCR,
negative control siRNA, miR-589-3p inhibitor, inhibitor control or
siIGF1R was added to MDA-MB-231 cells seeded in a 6-well plates at
the density of 5×105/well (500 µl). In addition,
MCF-7 cells were co-transfected with pc-TINCR and miR-589-3p mimic,
or IGF1R and miR-589-3p mimic. MDA-MB-231 cells were also
co-transfected with siTINCR and miR-589-3p inhibitor, or siIGF1R
and miR-589-3p inhibitor. Expression of TINCR, miR-589-3p and IGF1R
was measured after 24 h transfection.
Luciferase assay
PmirGLO reporter plasmids (cat. no. CL414-01;
Biomed) containing wild type TINCR (TINCR-wt) or mutant TINCR
(TINCR-mut), or wild type IGF1R (IGF1R-wt) or mutant IGF1R
(IGF1R-mut) were constructed. MCF-7 and MDA-MB-231 cells in
logarithmic growth phase were washed twice with PBS, and harvested
by using 6 ml 0.25% trypsin (cat. no. 25200072; Invitrogen; Thermo
Fisher Scientific, Inc.) from a 10-cm culture dish. Once cells were
evenly distributed into the dish, the culture was terminated by
adding medium containing 15% fetal calf serum (cat. no. C0251;
Beyotime Institute of Biotechnology). Cells were seeded in 6-well
plates at the density of 5×105 cells/well and placed in
an incubator at 37°C. After 24 h, the co-transfection was initiated
by adding 50 ng pmirGLO plasmid and 100 pmol miR-589-3p mimic (or
miR-589-3p mimic control) in each well. After 48 h, luciferase
activity was detected by using the dual-luciferase reporter kit
(cat. no. 16184; Thermo Fisher Scientific, Inc.). After aspirating
the culture medium, cells were washed once by PBS. Subsequently,
500 µl PLB was added to the cells that were gently shaken
for 15 min at room temperature. Firefly luciferase activity
(firefly) was measured after adding 100 µl LARII to 20
µl sample lysate. Eventually, 100 µl Stop&Glo
reagent was added to the mixture and the activity of Renilla
luciferase was detected by using the luminometer GloMax®
20/20 (Promega Corporation). The experimental results were obtained
by calculating the ratio of firefly/renilla.
Total RNA extraction from tissues and
cells and RT-qPCR
Total RNA was extracted from tissues and cells by
using TRIzol™ reagent (cat. no. 15596-026; Invitrogen; Thermo
Fisher Scientific, Inc.). Briefly, frozen tissues were sliced into
pieces and placed in a centrifuge tube and TRIzol™ reagent (1 ml)
was added. The mixture was grounded in a TissuePrep homogenizer
(Gering Scientific Instruments) for 5 min at 30 times/sec and was
centrifuged for 10 min (4°C; 1,600 × g). The supernatant was
collected, mixed with 200 µl chloroform and centrifuged for
10 min (4°C; 1,600 × g) again. The supernatant was mixed with 500
µl isopropyl alcohol and a white precipitate was obtained
following centrifugation for 10 min (4°C; 1,600 × g). The
precipitate was washed twice with 75% ethanol and was dissolved by
adding 20 µl diethyl pyrocarbonate (DEPC)-treated water
(cat. no. 750023; Thermo Fisher Scientific, Inc.) in order to
obtain the total RNA. Total RNA was extracted from breast cancer
cells by using TRIzol™ reagent according to the manufacturers'
instructions. Subsequently, RNA concentration was measured by using
a NanoDrop2000 (cat. no. YQ1633128263; Thermo Fisher Scientific,
Inc.). RNA was reverse transcribed according to the manufacturers'
protocol. The RNAs of TINCR, IGF1R, AKT and GAPDH (internal
reference) were reverse transcribed by using RevertAid First Strand
cDNA Synthesis kit (cat. no. k1622; Thermo Fisher Scientific,
Inc.). The RNA of miR-589-3p was formulated by using
All-in-OneTM miRNA First-Strand cDNA Synthesis kit (cat.
no. QP013; GeneCopoeia, Inc.) and TaqManTM Universal PCR
Master Mix (cat. no. 4304437; ABI). U6 served as an internal
reference. The reaction mixture for RT-qPCR contained 2 µl
cDNA (diluted 10-fold), 1 µl of forward and reverse primers
at 10 µM, 6 µl DEPC-treated water and 10 µl
FastStart Universal SYBR Green Master (cat. no. 0491391400; Roche
Diagnostics). RT-qPCR reactions were performed as follows:
Pre-denaturation at 95°C for 10 min, denaturation at 95°C for 15
sec, annealing at 60°C for 1 min, for a total of 40 cycles. The
relative expression levels were normalized to endogenous controls
and were expressed as 2−ΔΔCq (29). The sequences of the primers used
were as follows: TINCR, forward 5′-CCC AAG CTT GGT CTG GGC TCC CAG
GTG GACC-3′, reverse 5′-AGC GAT ATC CTA TAG TTG TTT TCA AAC ATG TAA
TCTT-3′; IGF1R, forward 5′-TGC GTG AGA GGA TTG AGT TTC-3′, reverse
5′-CTT ATT GGC GTT GAG GTA TGC-3′; AKT, forward 5′-TGT GGA TTT ACC
TTA TCC CCTCA-3′, reverse 5′-GTT TGG CTT TGG TCG TTC TGT-3′; GAPDH,
forward 5′-TGT GGG CAT CAA TGG ATTTGG-3′, reverse 5′-ACA CCA TGT
ATT CCG GGT CAAT-3′; miR-589-3p, forward 5′-AAC AAA TGC CGG TTC
CCAGA-3′, reverse 5′-TGT CGT GGA GTC GGC AATTG-3′; and U6, forward
5′-TCTG CTC CTA TCC CAA TTA CCTG-3′ and reverse 3′-ACT CCC GGA TCT
CTT CTA AGTTG-3′.
Patients survival analysis
A monthly follow-up was conducted on the 68 patients
with breast cancer up to five years. The survival rate was
calculated by using the Kaplan-Meier cumulative survival curve.
Cell proliferation
MCF-7 and MDA-MB-231 cell proliferation was detected
by using MTT reagent (cat. no. ST316; Beyotime Institute of
Biotechnology). Briefly, cells in the logarithmic growth phase were
harvested by using 0.25% trypsin. The cell concentration was
adjusted to 5×103 cells/well, and 100 µl
suspension was seeded in a 96-well plate and incubated at 37°C for
48 h. Subsequently, supernatant was removed, and 90 µl
culture medium and 10 µl MTT reagent were added to each
well. After 4 h incubation, supernatant was aspirated, 100
µl DMSO was added to each well and the plate was gently
shaken for 10 min to fully dissolve formazan crystals. Absorbance
was read at 490 nm on a microplate reader.
Cell apoptosis
Flow cytometry was used to detect the apoptosis of
MCF-7 and MDA-MB-231 cells following transfection with different
plasmids. Cells in the logarithmic growth phase were washed three
times with PBS, harvested by using 0.25% trypsin, collected and
centrifuged for 5 min (1,000 × g, at 4°C). MCF-7 and MDA-MB-231
cells were washed twice with PBS and centrifuged for 5 min (1,000 ×
g; 4°C), and then incubated with 5 µl Annexin V-FITC (cat.
no. KGA108; Nanjing KeyGen Biotech Co., Ltd.) and 10 µl
propidium iodide solution (cat. no. C1062M; Beyotime Institute of
Biotechnology) for 20 min at room temperature in the dark. Cell
apoptosis was detected by flow cytometry (cat. no. 322457; Bio-Rad
Laboratories, Inc.) and data analysis was performed using FACSdiva
software version 6.1.2 (BD Biosciences).
Wound healing assay
A marker was used to evenly draw a 0.5~1 cm
horizontal line under the 6-well plate. MCF-7 and MDA-MB-231 cells
in the logarithmic growth phase were seeded at the density of
5×105 cells/well and incubated overnight. The next day,
the tip of a pipette was used to scratch the cell layer, and cells
were washed three times with PBS. Serum-free medium was added and
cells were cultured at 37°C with 5% CO2 for 48 h. Cell
migration was imaged using a light microscope (magnification, ×100)
at 0 and 48 h.
Cell invasion assay
Transwell chambers (8-µm pore size; BD
Biosciences) were used to detect the invasion ability of MCF-7 and
MDA-MB-231 cells. The two cell lines were serum-starved for 12 h
and harvested by using 0.25% trypsin. Cells were seeded at the
density of 1×104 cells/per well in the upper chamber of
the Transwell that was pre-coated with Matrigel (cat. no.
YZ-354234; Beijing Solarbio Science & Technology Co., Ltd.),
and DMEM containing 10% FBS was added to the lower chamber.
Following 48 h incubation, cells that have invaded the bottom
membrane were fixed with 4% paraformaldehyde for 15 min at room
temperature and stained by 0.1% crystal violet stain. Cell invasion
was observed under a light microscope (magnification, ×200) and the
cell number was counted by Image J software (version 1.8.0;
National Institutes of Health).
Western blotting
Proteins were extracted from MCF-7 and MDA-MB-231
cells. Cells were lysed using RIPA (cat. no. 89900; Thermo Fisher
Scientific, Inc.). Cell lysate was centrifuged at 1,600 × g, 4°C
for 10 min. The supernatant was collected and protein concentration
was measured using the BCA Protein Assay kit (cat. no. 23227;
Thermo Fisher Scientific, Inc.). Proteins (25 µg) were
separated by 12% SDS-PAGE and transferred onto PVDF membranes (cat.
no. RPN303F; GE Healthcare). Membranes were washed three times with
TBST (cat. no. T1085; Beijing Solarbio Science & Technology
Co., Ltd.) and blocked with a blocking buffer containing 5% bovine
serum albumin (cat. no. SW3015-500 ml; Beijing Solarbio Science
& Technology Co., Ltd.) for 2 h at room temperature. Membranes
were incubated with primary antibodies against IGF1R (cat. no.
ab39675; 155 KD; 1:1,000; Abcam), phosphor (p)-Akt (cat. no.
ab38449; 56 kD; 1:1,000; Abcam), Akt (cat. no. ab8805; 55 kD;
1:500; Abcam), GAPDH (internal control; cat. no. ab8245; 36 KD;
1:10,000, Abcam) overnight at 4°C. Membranes were washed three
times with TBST and incubated with the goat anti-rabbit IgG
secondary antibody (cat. no. ab6721, 1:10,000, Abcam) for 1 h at
room temperature. Immobilon Western Chemiluminescent HRP Substrate
(cat. no. WBKLS0100; Merck KGaA) was used to detect the signal on
the membrane. Bands were imaged by using the Gel Imager System
(1708265, Bio-Rad) and data were analyzed via densitometry using
ImageJ software (version 1.46) and normal-ized to expression of the
internal control.
Statistical analysis
Data were analyzed using SPSS 23.0 software (IBM
Corp.) and were expressed as the means ± standard deviation of the
mean. Student's two-tailed t-test was used to compare differences
between two groups. ANOVA followed by Turkey test was used to
compare differences between ≥3 groups. Correlation analysis was
preformed using Spearman correlation coefficient algorithm.
P<0.05 was considered to indicate a statistically significant
difference.
Results
TINCR overexpression in breast cancer
tissues is negatively correlated with miR-589-3p expression
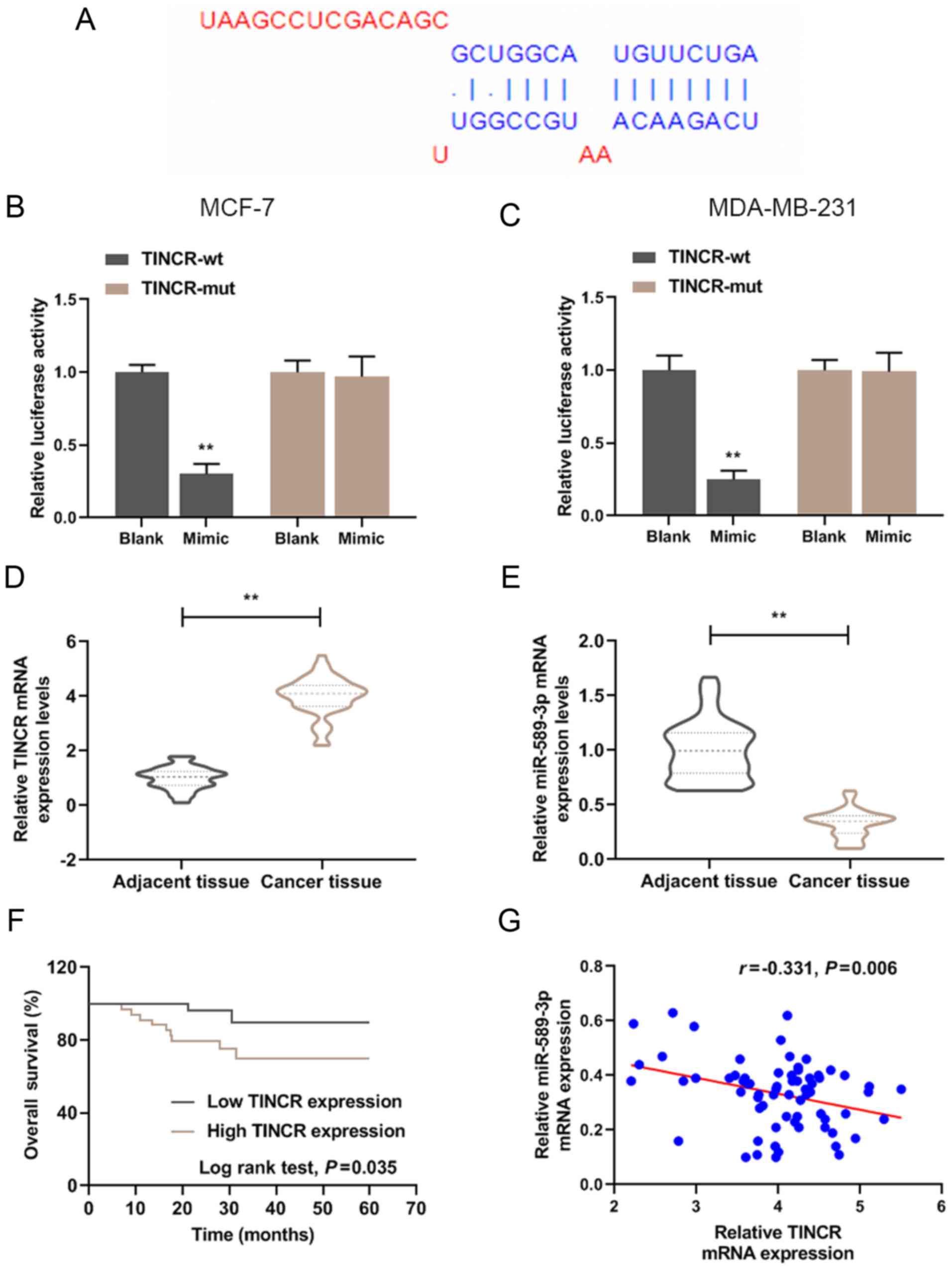
A targeted relationship between TINCR and miR-589-3p
was predicted by Starbase and results from luciferase assay
(Fig. 1A-C). To examine the role
of TINCR and miR-589-3p in breast cancer, the expression level of
TINCR and miR-589-3p in breast cancer tissues was determined by
RT-qPCR (Fig. 1D and E). The
results demonstrated that TINCR expression level was significantly
higher in breast cancer tissues compared with adjacent tissues
(P<0.001); however, miR-589-3p expression level was
significantly lower in cancer tissues compared with normal tissues
(P<0.001). Furthermore, the results demonstrated that TINCR and
miR-589-3p expression levels were negatively correlated (Fig. 1G; r=-0.331; P=0.006). According to
the mean as the segmentation point, the expression of miR-589-3p
was divided into high expression and low expression. In addition,
the results from Kaplan-Meier overall survival curves demonstrated
that patients with high TINCR expression level had a lower survival
rate at five years (P<0.05; Fig.
1F).
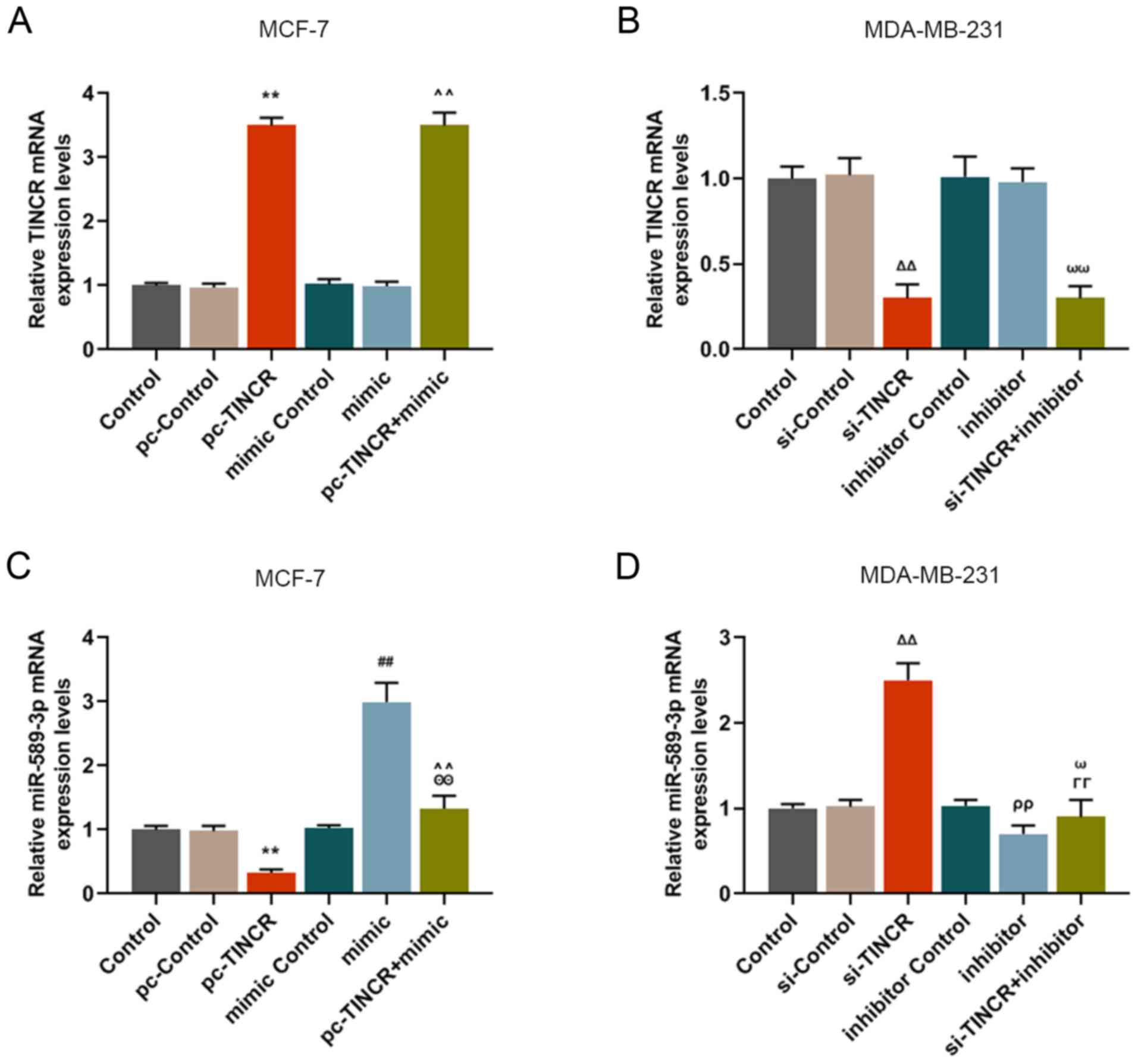
TINCR overexpression downregulates
miR-589-3p expression in breast cancer cells
The regulatory relationship between TINCR and
miR-589-3p was further investigated by RT-qPCR. The results
demonstrated that cell transfection with miR-589-3p mimic and
inhibitor did not affect TINCR expression (Fig. 2A and B); however, the effect of
pc-TINCR on miR-589-3p expression was reversed by miR-589-3p mimic
(P<0.001; Fig. 2C).
Furthermore, miR-589-3p inhibitor reversed miR-589-3p expression
level that was promoted by silencing TINCR (P<0.001; Fig. 2D). These results suggested that
TINCR may be able to downregulate miR-589-3p, which may be reversed
by miR-589-3p mimic.
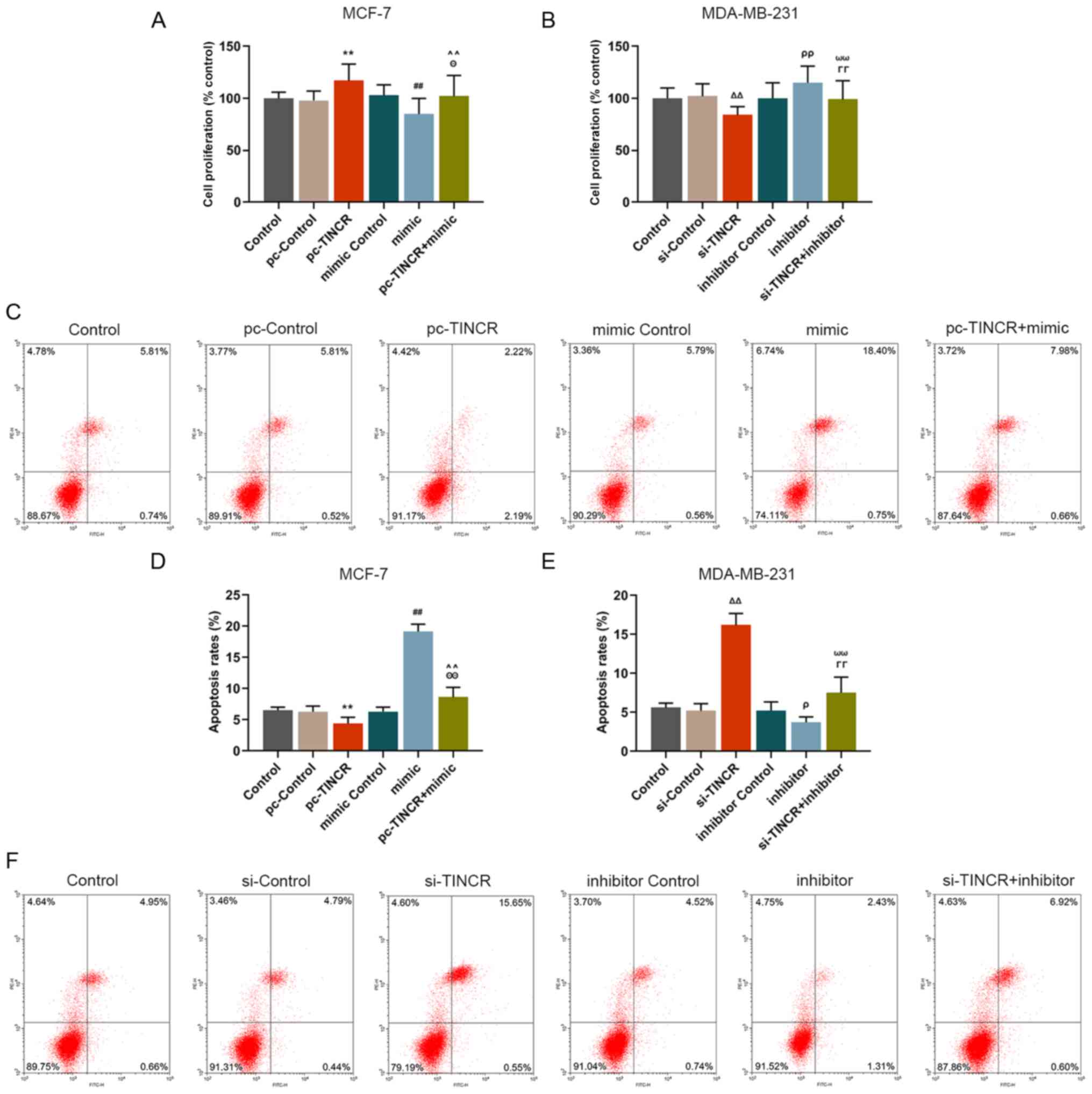
miR-589-3p mimic partially reverses the
promoting and inhibiting effects of TINCR on breast cancer cell
proliferation and apoptosis, respectively
The effect of TINCR on breast cancer cell
proliferation and apoptosis was investigated. The results
demonstrated that pc-TINCR promoted MCF-7 cell proliferation
(P<0.001; Fig. 3A) and
inhibited MCF-7 cell apoptosis (P<0.001; Fig. 3C and D); however, siTINCR
significantly inhibited MDA-MB-231 cell proliferation (P<0.001;
Fig. 3B) and promoted cell
apoptosis (P<0.001; Fig. 3E and
F). These findings suggested that TINCR could promote breast
cancer cell proliferation and inhibit their apoptosis. In addition,
further results demonstrated that miR-589-3p mimic reversed the
effect of pc-TINCR on breast cancer cell proliferation (P<0.001;
Fig. 3A) and inhibited cell
apoptosis (P<0.001; Fig. 3C and
D). Furthermore, miR-589-3p inhibitor reversed the effect of
siTINCR on breast cancer cell proliferation and apoptosis
(P<0.001; Fig. 3B, E and F).
These data indicated that the effects of TINCR on the promotion and
the inhibition of breast cancer cell proliferation and apoptosis,
respectively, may be reversed by miR-589-3p mimic.
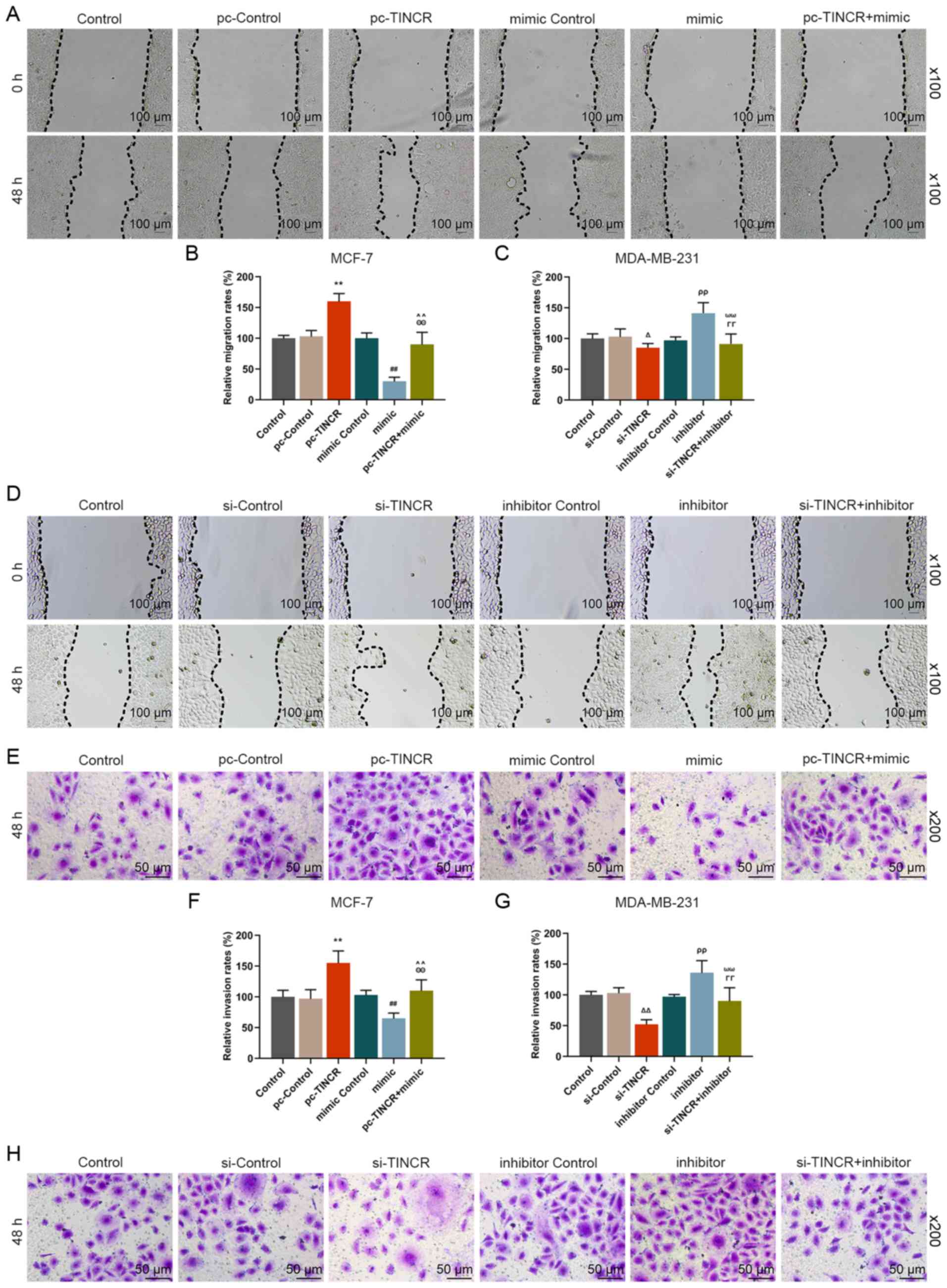
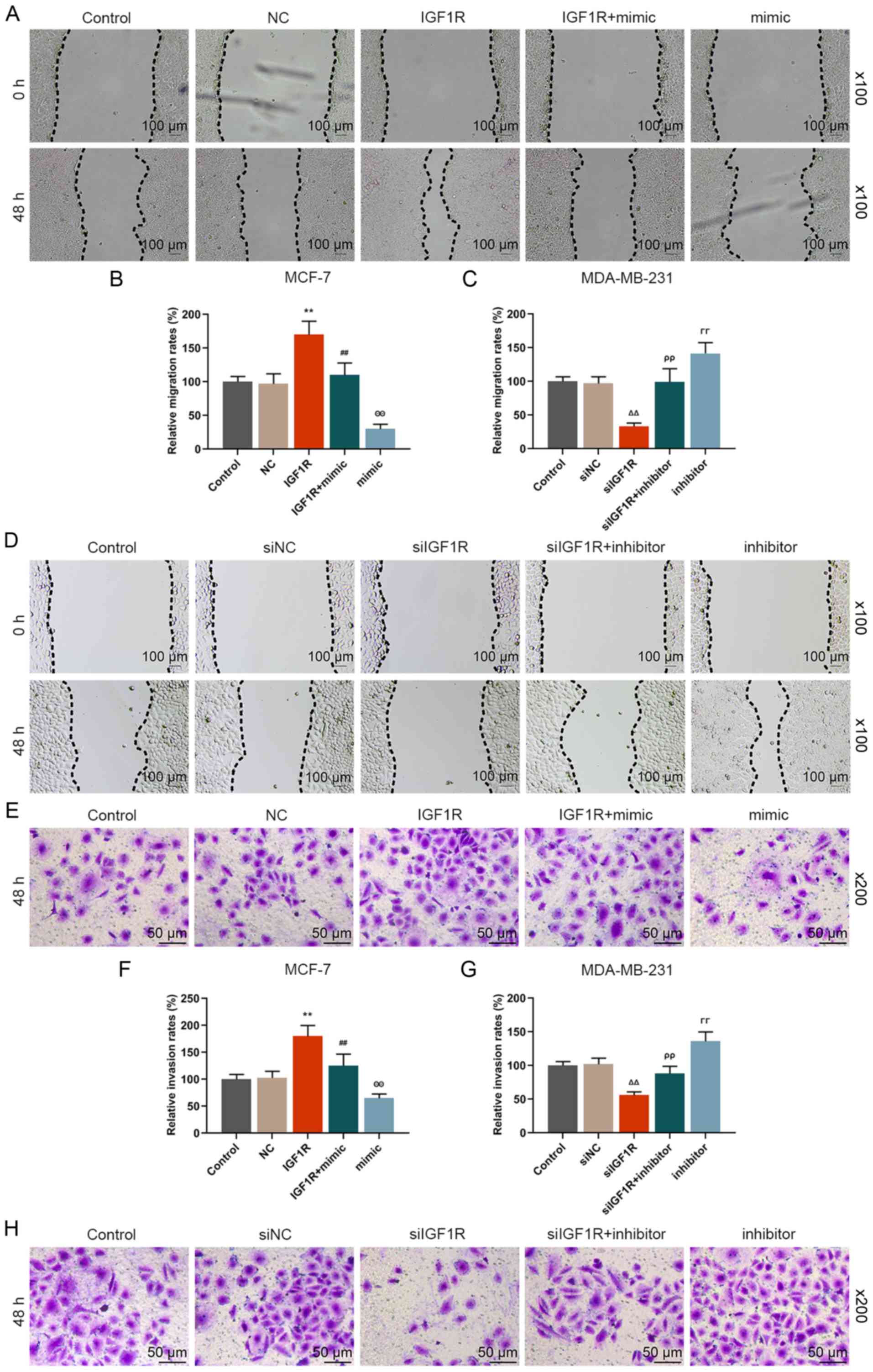
miR-589-3p mimic partially reverses the
promoting effect of TINCR on the migratory and invasive abilities
of breast cancer cells
The effect of TINCR and miR-589-3p mimic on the
migratory and invasive abilities of breast cancer cells was
examined. The results from the wound healing assay demonstrated
that the migratory rate of breast cancer cells in the pc-TINCR
group was significantly increased compared with the pc-control
group (P<0.001; Fig. 4A and
B). The results from Transwell assay demonstrated that the
invasive ability of breast cancer cells the pc-TINCR group was
significantly increased compared with the pc-control group
(P<0.001; Fig. 4E and F);
however, siTINCR transfection had the opposite effects (P<0.001;
Fig. 4C, D, G and H). In
addition, the results demonstrated that miR-589-3p mimic partially
reversed the effects of pc-TINCR on the migratory and invasive
abilities of breast cancer cells (P<0.001; Fig. 4A, B, E and F). Furthermore,
miR-589-3p inhibitor also partially reversed the inhibiting effects
of siTINCR on the migratory and invasive abilities of breast cancer
cells (P<0.001; Fig. 4C, D, G and
H). These findings suggested that TINCR may promote breast
cancer cell migratory and invasive abilities; however, these
effects may by partially reversed by miR-589-3p mimic.
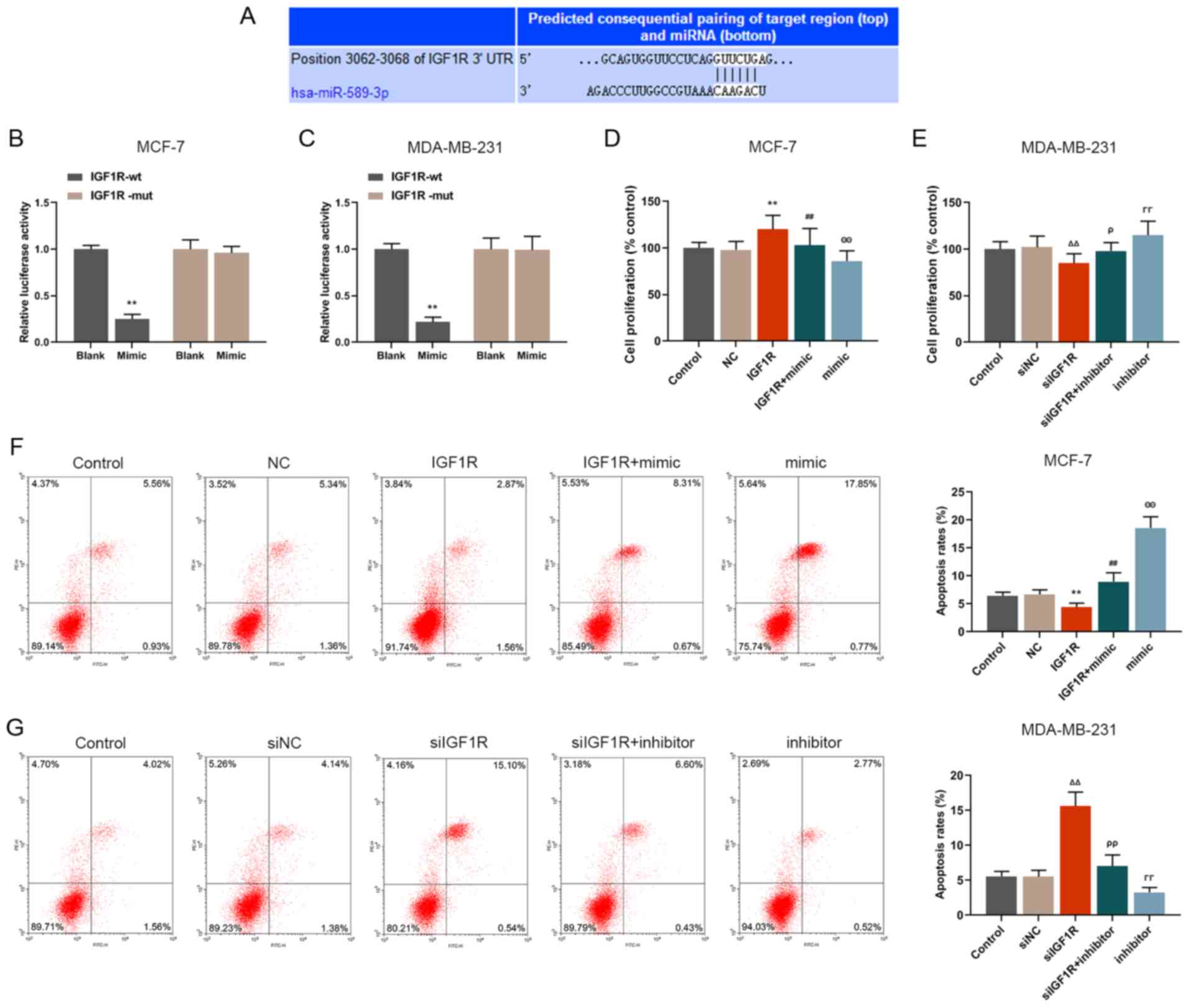
IGF1R is a target gene of miR-589-3p
The results from TargetScan database revealed that
IGF1R had a binding site to miR-589-3p at the 3′UTR position
(Fig. 5A), indicating that IGF1R
may be considered as a target gene for miR-589-3p. For further
validation, luciferase assay was performed and the results
demonstrated that IGF1R luciferase activity in the miR-589-3p mimic
group was lower compared with the control group (P<0.001;
Fig. 5B and C) in both breast
cancer cell lines. Not only IGF1R was a target gene of miR-589-3p
but it may also be downregulated by miR-589-3p mimic.
miR-589-3p mimics can partially reverse
the effects of IGF1R on cell proliferation, migration, invasion and
apoptosis
Proliferation, apoptosis, and migratory and invasive
abilities of breast cancer cells were detected. The results
demonstrated that IGF1R significantly promoted the proliferation,
migration and invasion of MCF-7 cells (P<0.001; Figs. 5D, 6A and B, E and F); however, these
effects were partially reversed by miR-589-3p mimic (P<0.001;
Figs. 5D, 6A and B, E and F). Furthermore, IGF1R
silencing inhibited MDA-MB-231 cell proliferation, migration and
invasion (P<0.001; Figs. 5E,
6C and D, G and H), which was
reversed by miR-589-3p inhibitor. In addition, the results from
flow cytometry demonstrated that the inhibitory effect of IGF1R on
the apoptosis of MCF-7 cells was reversed by miR-589-3p mimic
(P<0.001; Fig. 5F), and the
promoting effects of siIGF1R on MDA-MB-231 cell apoptosis were
reversed by miR-589-3p inhibitor (P<0.001; Fig. 5G). These findings demonstrated
that IGF1R may promote cell proliferation, migration and invasion,
and inhibit cell apoptosis; however these effects may be partially
reversed by miR-589-3p mimic.
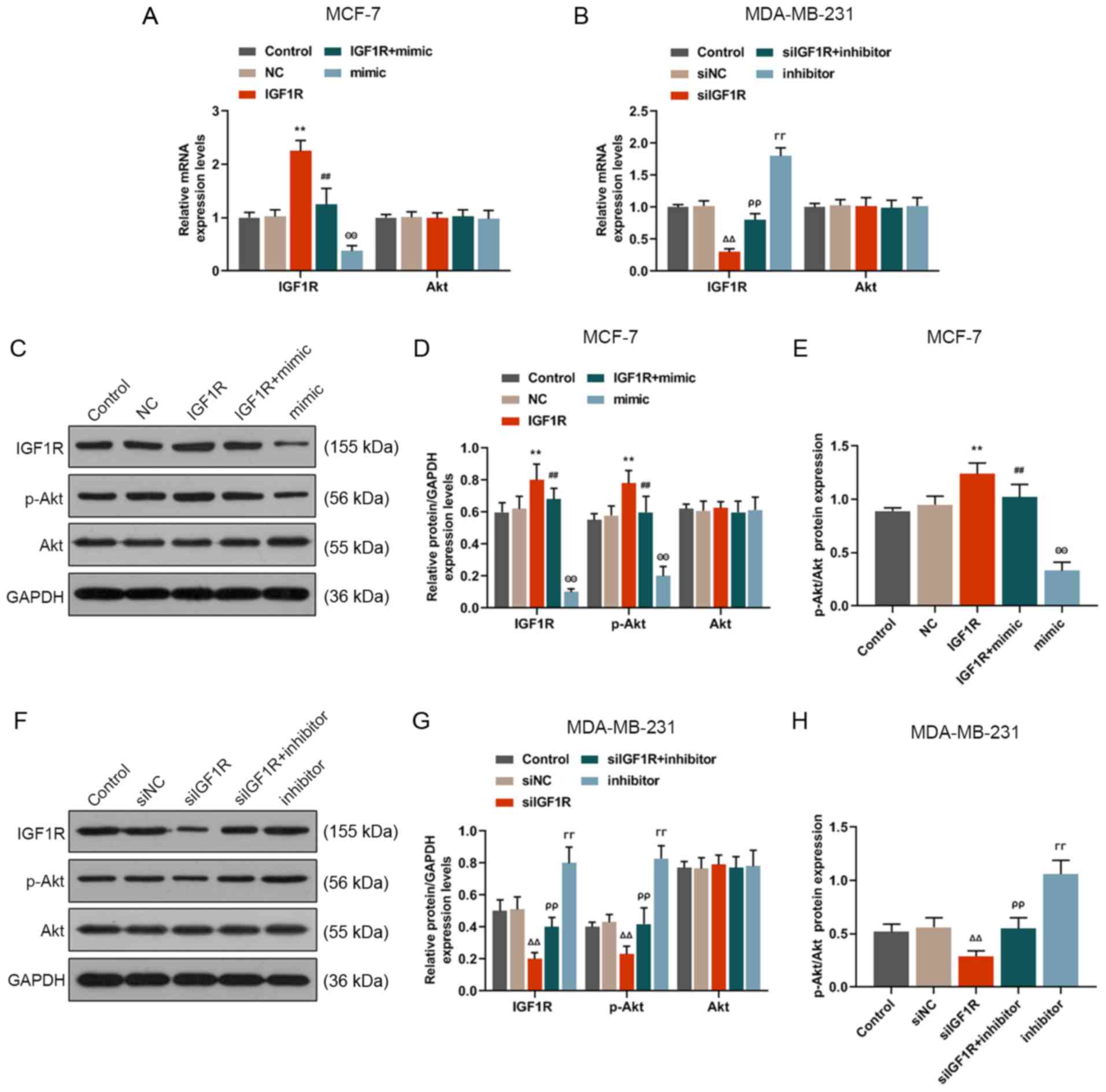
miR-589-3p can downregulate IGF1R and Akt
protein expression in breast cancer cells
The underlying mechanism of miR-589-3p in breast
cancer cells was investigated by RT-qPCR and western blotting. The
mRNA expression of IGF1R and Akt (Fig. 7A and B) was detected by RT-qPCR.
The results demonstrated that IGF1R expression was downregu-lated
following co-transfection with IGFR1 and miR-589-3p mimic compared
with IGF1R group (P<0.001), whereas IGF1R expression was
significantly upregulated following co-trans-fection with siIGF1R
and miR-589-3p inhibitor compared with siIGF1R group (P<0.001).
No change in Akt expression level was observed in any condition.
Subsequently, western blotting was performed to analyse the protein
expression of IGF1R, p-Akt and Akt. The results were similar to
those from RT-qPCR. In particular, miR-589-3p mimic partially
reversed IGF1R and p-Akt high protein expression in the IGF1R group
(P<0.001; Fig. 7C-E), whereas
the Akt protein expression did not significantly change.
Subsequently, the ratio p-Akt/Akt was downregulated in siIGF1R
group (P<0.001), whereas the ratio p-Akt/Akt was increased in
the siIGF1R + inhibitor and inhibitor groups (P<0.001; Fig. 7F-H). These findings suggested that
miR-589-3p may downregulate IGF1R and p-Akt expression in breast
cancer cells.
Discussion
The results from the present study demonstrated that
TINCR was highly expressed in MCF-7 and MDA-MB-231 breast cancer
cells, and that TINCR promoted the proliferation and migratory and
invasive abilities of breast cancer cells and inhibited cancer cell
apoptosis. These findings were consistent with a previous study
(21). Furthermore, to the best
of our knowledge, the present study was the first to demonstrate
that miR-589-3p may act as a targeted miRNA for TINCR, and that its
expression was inhibited by TINCR. In breast cancer cells that were
co-transfected with miR-589-3p mimic and pc-TINCR, the promoting
effects of TINCR on cancer cell proliferation and migratory and
invasive abilities were attenuated by miR-589-3p mimic. In
addition, this study demonstrated that the target gene IGF1R of
miR-589-3p promoted the proliferation and migratory and invasive
abilities of breast cancer cells and inhibited their apoptosis,
which may be partially reversed by miR-589-3p mimic. These effects
were obtained via inhibition of the IGF1R-Akt pathway.
In the last decades, the prognosis of patients with
breast cancer has greatly improved thanks to the constant advances
in medical technologies and techniques. However, it remains crucial
to make progress on the development of novel diagnosis tools and
therapeutic options for patients. Tumor presentation varies among
molecular subtypes, suggesting that tumors of different subtypes
may be useful in selecting local therapy (30). Targeted therapeutic drugs could
aim different genes and signalling pathways in breast cancer. For
example, agents targetin estrogen receptor (ER) and HER2, such as
tamoxifen and trastuzumab, are the most extensively used
therapeutics in breast cancer (31,32). These therapeutic drugs have
therefore been designed for different molecular subtypes. However,
the existence of chemoresistance and the lack of therapeutic drugs
for triple-negative breast cancer remain to be resolved (33,34). It is therefore crucial to explore
the underlying mechanisms of breast cancer metastasis and
recurrence, in order to determine potential novel intervention
targets and develop more effective treatment strategies. miRNAs
represent a class of endogenous single-stranded non-coding RNAs of
~20-24 nt in length, which are able to bind to the 3′UTR regions of
mRNA, leading to gene silencing at post-transcriptional levels.
miRNAs are widely involved in cell proliferation, organ
development, immune response and tumor formation among other
processes. Previous studies reported that miRNAs are closely
related to the occurrence and development of certain types of tumor
(35-37). Not only they can function as tumor
promoters but they can also function as oncogenes. miRNAs in the
plasma can also be used as biomarkers for early diagnosis of
certain tumors. It has been reported that exosomal microRNA-21
could be considered as a potential biomarker for the early
diagnosis of pancreatic cancer using a tethered cationic lipoplex
nanoparticle biochip (38).
Furthermore, exosomal microRNA-210 is a potentially non-invasive
biomarker for the diagnosis and prognosis of glioma (39). In breast cancer, >30 miRNAs
have been confirmed to be associated with breast cancer (40). For example, miR-145 can inhibit
cancer cell proliferation and invasion via the regulation of N-RAS
and vascular endothelial growth factor A in triple-negative breast
cancer, miR-145 can inhibit angiogenesis in breast cancer tissues
(41). Furthermore, miR-203,
which is lowly expressed in metastatic breast cancer tissues, can
promote breast cancer cell metastasis via upregulation of snail
family transcriptional repressor 2 (42). In addition, it was reported that
the proto-oncogene miR-210 is highly expressed in breast cancer
tissues, which is associated with poor prognosis of patients with
breast cancer (43). The present
study demonstrated that miR-589-3p overexpression could partially
reverse the effects of TINCR on breast cancer cell proliferation
and migratory and invasive abilities and on the inhibition of
breast cancer cell apoptosis. Similarly, Cesarini et al
(44) reported that miR-589-3p
can inhibit the proliferation and migratory and invasive abilities
of glioblastoma cells, suggesting its anti-cancer effect.
Insulin-like growth factors (IGFS) represent a class
of growth factors that serve crucial roles in the growth and
differentiation of bones and muscles (45,46). The main biological activities of
IGFS are mediated by IGFR1. IGF1R is a trans-membrane tyrosine
kinase receptor that plays a vital role in cell mitosis,
proliferation, differentiation and apoptosis (47). Loughran et al (48) confirmed that IGF1R overexpression
can cause cell division and proliferation, increasing therefore the
risk of tumorigenesis; however, cell malignant transformation is
suppressed following IGF1R downregulation (49). In 1992, De Leon et al
(50) reported that IGF-I and
IGF-II, which are effective mitogens in the breast cancer cells
MCF-7, can promote cancer cell proliferation. Turner et al
(51) reported that IGF1R
expression is increased in resected breast cancer tumor tissues and
is associated with early recurrence. Powell et al (52) demonstrated that the commonly
inherited IGF1R variant (rs2016347) reduces breast cancer risk by
enhancing mammary gland involution. In addition, triptolide and
IGF1R inhibitor (AG1024) can synergistically inhibit the
proliferation and induce the apoptosis of triple-negative breast
cancer cells (53). These studies
indicated that IGF1R serve some essential roles in the occurrence
and development of breast cancer. IGF1R is widely expressed in
various cells of the body, and PI3K-Akt and ERK-MAPK signalling
pathways are associated with the occurrence and development of
malignant tumors. The signalling pathways regulated by IGF1R
include PI3K-Akt and ERK-MAPK pathways (54). Akt is a serine/threonine protein
kinase that is the most important target in the PI3K-Akt signalling
pathway. The effects of Akt on regulatory mechanisms are dependent
on p-Akt that promotes Akt activation, serving therefore a central
role in the transmission of information (55). Previous studies confirmed that Akt
can regulate the proliferation, migration, invasion and apoptosis
of tumor cells via multiple signalling pathways, and also mediate
tumor angiogenesis and tumor resistance (56,57). Blocking or inhibiting Akt
expression could therefore be effective in cancer treatment.
Similarly, the present study demonstrated that miR-589-3p may
inhibit breast cancer cell proliferation by decreasing the
expression of IGF1R and p-AKT.
Some inhibitors of IGF1R and Akt have been used in
the treatment of breast cancer, including NVP-AEW541 (58), OSI-906 (59) and MK-2206 (60). Inhibitors could be used in
combination, or multi-target drugs should be developed. de Lint
et al (59) reported that
the combination of IGF1R inhibitors and PI3K inhibitors has a
therapeutic effect on certain patients with triple-negative breast
cancer. In the present study, not only miR-589-3p could partially
reverse the effect of TINCR on breast cancer cells but it also
inhibited the proliferation and migratory and invasive abilities of
breast cancer cells by inhibiting the IGF1R-Akt pathway, promoting
therefore cancer cell apoptosis, which showed its dual regulatory
effect. These findings provided a reliable basis for miRNA
application in the treatment of breast cancer, which could improve
the prognosis of patients with breast cancer. However, this study
presented some limitations, and whether IGF1R or Akt inhibitors
could have therapeutic roles in animal models of breast cancer
requires further investigation. In addition, the stimulating
effects of TINCR on the proliferation and migratory and invasive
abilities of breast cancer cells through regulation of the
miR-589-3p/IGF1R/Akt axis should be further confirmed by in
vivo experiments.
Abbreviations:
|
GEO
|
Gene Expression Omnibus
|
|
lncRNA
|
long non-coding RNA
|
|
miRNA
|
microRNA
|
|
STAU1
|
staufen1
|
|
TCGA
|
The Cancer Genome Atlas
|
|
TINCR
|
tissue differentiation inducing
non-protein coding RNA
|
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FG and XZ provided substantial contributions to the
conception and design of the study. QZ and QH acquired, analyzed
and interpreted data. FG and XZ drafted and critically revised the
manuscript for important intellectual content. All authors agreed
to be accountable for all aspects of the work in ensuring that
questions related to the accuracy or integrity of the work were
appropriately investigated and resolved. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study has been approved by the Ethics
Committee of Affiliated Zhongshan Hospital of Dalian University
(approval no. 201112013RXW) and all procedures were performed in
accordance with the 1964 Declaration of Helsinki and its later
amendments or comparable ethical standards. All patients signed
informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Woolston C: Breast cancer. Nature.
527:S1012015. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Ma J, Gaudet MM, Newman LA,
Miller KD, Goding Sauer A, Jemal A and Siegel RL: Breast cancer
statistics, 2019. CA Cancer J Clin. 69:438–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ng CK, Martelotto LG, Gauthier A, Wen HC,
Piscuoglio S, Lim RS, Cowell CF, Wilkerson PM, Wai P, Rodrigues DN,
et al: Intra-tumor genetic heterogeneity and alternative driver
genetic alterations in breast cancers with heterogeneous HER2 gene
amplification. Genome Biol. 16:1072015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Carmichael H, Matsen C, Freer P, Kohlmann
W, Stein M, Buys SS and Colonna S: Breast cancer screening of
pregnant and breastfeeding women with BRCA mutations. Breast Cancer
Res Treat. 162:225–230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nik-Zainal S, Davies H, Staaf J,
Ramakrishna M, Glodzik D, Zou X, Martincorena I, Alexandrov LB,
Martin S, Wedge DC, et al: Landscape of somatic mutations in 560
breast cancer whole-genome sequences. Nature. 534:47–54. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
ENCODE Project Consortium: An integrated
encyclopedia of DNA elements in the human genome. Nature.
489:57–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Renganathan A and Felley-Bosco E: Long
noncoding RNAs in cancer and therapeutic potential. Adv Exp Med
Biol. 1008:199–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Q, Zhu C, Jin Y, Si X, Jiao W, He W,
Mao W, Li M and Luo G: Plasma long non-coding RNA RP11-438N5.3 as a
novel biomarker for non-small cell lung cancer. Cancer Manag Res.
12:1513–1521. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen X, Xue Y, Cong H, Wang X, Fan Z, Cui
X and Ju S: Circulating lncRNA DANCR as a potential auxillary
biomarker for the diagnosis and prognostic prediction of colorectal
cancer. Biosci Rep. 40:BSR201914812020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y and Tang L: The application of
lncRNAs in cancer treatment and diagnosis. Recent Pat Anticancer
Drug Discov. 13:292–301. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kretz M, Siprashvili Z, Chu C, Webster DE,
Zehnder A, Qu K, Lee CS, Flockhart RJ, Groff AF, Chow J, et al:
Control of somatic tissue differentiation by the long non-coding
RNA TINCR. Nature. 493:231–235. 2013. View Article : Google Scholar :
|
|
14
|
Kretz M: TINCR, staufen1, and cellular
differentiation. RNA Biol. 10:1597–1601. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu TP, Wang YF, Xiong WL, Ma P, Wang WY,
Chen WM, Huang MD, Xia R, Wang R, Zhang EB, et al: E2F1 induces
TINCR transcriptional activity and accelerates gastric cancer
progression via activation of TINCR/STAU1/CDKN2B signaling axis.
Cell Death Dis. 8:e28372017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang ZY, Lu YX, Zhang ZY, Chang YY, Zheng
L, Yuan L, Zhang F, Hu YH, Zhang WJ and Li XN: Loss of TINCR
expression promotes proliferation, metastasis through activating
EpCAM cleavage in colorectal cancer. Oncotarget. 7:22639–22649.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Z, Liu Y, He A, Li J, Chen M, Zhan Y,
Lin J, Zhuang C, Liu L, Zhao G, et al: Theophylline controllable
RNAi-based genetic switches regulate expression of lncRNA TINCR and
malignant phenotypes in bladder cancer cells. Sci Rep. 6:307982016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu M, Han Y, Zhang Y, Zhou Y and Ye L:
lncRNA TINCR sponges miR-214-5p to upregulate ROCK1 in
hepatocellular carcinoma. BMC Med Genet. 21:22020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao YW, Ma F, Xie YC, Ding MG, Luo LH,
Jiang S, Rao L and Liu XL: Sp1-induced upregulation of the long
noncoding RNA TINCR inhibits cell migration and invasion by
regulating miR-107/miR-1286 in lung adenocarcinoma. Am J Transl
Res. 11:4761–4775. 2019.PubMed/NCBI
|
|
20
|
Li R, Wang Y, Xu Y, He X and Li Y:
Silencing the long noncoding RNA, TINCR, a molecular sponge of
miR-335, inhibits the malignant phenotype of epithelial ovarian
cancer via FGF2 suppression. Int J Oncol. 55:1110–1124.
2019.PubMed/NCBI
|
|
21
|
Li S, Li J, Li H, Gao M, Li N, Wang Y,
Tong L, Song M and Yin Z: Clinicopathological and prognostic
significance of TINCR in caner: A meta-analysis. Pathol Res Pract.
215:1525962019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong H, Hu J, Zou K, Ye M, Chen Y, Wu C,
Chen X and Han M: Activation of LncRNA TINCR by H3K27 acetylation
promotes Trastuzumab resistance and epithelial-mesenchymal
transition by targeting MicroRNA-125b in breast cancer. Mol Cancer.
18:32019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Du Y, Hu X, Zhao L and Xia W:
Up-regulation of ceRNA TINCR by SP1 contributes to tumorigenesis in
breast cancer. BMC Cancer. 18:3672018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li J, Gao C, Liu C, Zhou C, Ma X, Li H, Li
J, Wang X, Qi L, Yao Y, et al: Four lncRNAs associated with breast
cancer prognosis identified by coexpression network analysis. J
Cell Physiol. 234:14019–14030. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu S, Kong D, Chen Q, Ping Y and Pang D:
Oncogenic long noncoding RNA landscape in breast cancer. Mol
Cancer. 16:1292017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mah SM, Buske C, Humphries RK and
Kuchenbauer F: miRNA*: A passenger stranded in RNA-induced
silencing complex? Crit Rev Eukaryot Gene Expr. 20:141–148. 2010.
View Article : Google Scholar
|
|
27
|
Backes C, Meese E and Keller A: Specific
miRNA disease biomarkers in blood, serum and plasma: Challenges and
prospects. Mol Diagn Ther. 20:509–518. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Z, Liu H, Yang H, Gao Y, Zhang G and
Hu J: The long noncoding RNA, TINCR, functions as a competing
endogenous RNA to regulate PDK1 expression by sponging miR-375 in
gastric cancer. Onco Targets Ther. 10:3353–3362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Wiechmann L, Sampson M, Stempel M, Jacks
LM, Patil SM, King T and Morrow M: Presenting features of breast
cancer differ by molecular subtype. Ann Surg Oncol. 16:2705–2710.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nagini S: Breast cancer: Current molecular
therapeutic targets and new players. Anticancer Agents Med Chem.
17:152–163. 2017. View Article : Google Scholar
|
|
32
|
Kast K, Schoffer O, Link T, Forberger A,
Petzold A, Niedostatek A, Werner C, Klug SJ, Werner A, Gatzweiler
A, et al: Trastuzumab and survival of patients with metastatic
breast cancer. Arch Gynecol Obstet. 296:303–312. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Saw PE, Park J, Jon S and Farokhzad OC: A
drug-delivery strategy for overcoming drug resistance in breast
cancer through targeting of oncofetal fibronectin. Nanomedicine.
13:713–722. 2017. View Article : Google Scholar
|
|
34
|
DeMichele A, Yee D and Esserman L:
Mechanisms of resistance to neoadjuvant chemotherapy in breast
cancer. N Engl J Med. 377:2287–2289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qadir MI and Faheem A: miRNA: A diagnostic
and therapeutic tool for pancreatic cancer. Crit Rev Eukaryot Gene
Expr. 27:197–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ganapathy K, Staklinski S, Hasan MF,
Ottman R, Andl T, Berglund AE, Park JY and Chakrabarti R:
Multifaceted function of MicroRNA-299-3p fosters an antitumor
environment through modulation of androgen receptor and VEGFA
signaling pathways in prostate cancer. Sci Rep. 10:51672020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Indrieri A, Carrella S, Carotenuto P,
Banfi S and Franco B: The pervasive role of the miR-181 Family in
development, neurode-generation, and cancer. Int J Mol Sci.
21:E20922020. View Article : Google Scholar
|
|
38
|
Pu X, Ding G, Wu M, Zhou S, Jia S and Cao
L: Elevated expression of exosomal microRNA-21 as a potential
biomarker for the early diagnosis of pancreatic cancer using a
tethered cationic lipoplex nanoparticle biochip. Oncol Lett.
19:2062–2070. 2020.PubMed/NCBI
|
|
39
|
Lan F, Yue X and Xia T: Exosomal
microRNA-210 is a potentially non-invasive biomarker for the
diagnosis and prognosis of glioma. Oncol Lett. 19:1967–1974.
2020.PubMed/NCBI
|
|
40
|
Nassar FJ, Nasr R and Talhouk R: MicroRNAs
as biomarkers for early breast cancer diagnosis, prognosis and
therapy prediction. Pharmacol Ther. 172:34–49. 2017. View Article : Google Scholar
|
|
41
|
Zou C, Xu Q, Mao F, Li D, Bian C, Liu LZ,
Jiang Y, Chen X, Qi Y, Zhang X, et al: MiR-145 inhibits tumor
angiogenesis and growth by N-RAS and VEGF. Cell Cycle.
11:2137–2145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Z, Zhang B, Li W, Fu L, Fu L, Zhu Z
and Dong JT: Epigenetic silencing of miR-203 upregulates SNAI2 and
contributes to the invasiveness of malignant breast cancer cells.
Genes Cancer. 2:782–791. 2011. View Article : Google Scholar
|
|
43
|
Tang Y, Zhou X, Ji J, Chen L, Cao J, Luo J
and Zhang S: High expression levels of miR-21 and miR-210 predict
unfavorable survival in breast cancer: A systemic review and
meta-analysis. Int J Biol Markers. 30:e347–e358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cesarini V, Silvestris DA, Tassinari V,
Tomaselli S, Alon S, Eisenberg E, Locatelli F and Gallo A:
ADAR2/miR-589-3p axis controls glioblastoma cell
migration/invasion. Nucleic Acids Res. 46:2045–2059. 2018.
View Article : Google Scholar :
|
|
45
|
Duan C, Ren H and Gao S: Insulin-like
growth factors (IGFs), IGF receptors, and IGF-binding proteins:
Roles in skeletal muscle growth and differentiation. Gen Comp
Endocrinol. 167:344–351. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Patil AS, Sable RB and Kothari RM: Role of
insulin-like growth factors (IGFs), their receptors and genetic
regulation in the chondrogenesis and growth of the mandibular
condylar cartilage. J Cell Physiol. 227:1796–1804. 2012. View Article : Google Scholar
|
|
47
|
Simpson A, Petnga W, Macaulay VM,
Weyer-Czernilofsky U and Bogenrieder T: Insulin-like growth factor
(IGF) pathway targeting in cancer: Role of the IGF Axis and
opportunities for future combination studies. Target Oncol.
12:571–597. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Loughran G, Huigsloot M, Kiely PA, Smith
LM, Floyd S, Ayllon V and O'Connor R: Gene expression profiles in
cells transformed by overexpression of the IGF-I receptor.
Oncogene. 24:6185–6193. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Werner H and Sarfstein R: Transcriptional
and epigenetic control of IGF1R gene expression: Implications in
metabolism and cancer. Growth Horm IGF Res. 24:112–118. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
De Leon DD, Wilson DM, Powers M and
Rosenfeld RG: Effects of insulin-like growth factors (IGFs) and IGF
receptor antibodies on the proliferation of human breast cancer
cells. Growth Factors. 6:327–336. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Turner BC, Haffty BG, Narayanan L, Yuan J,
Havre PA, Gumbs AA, Kaplan L, Burgaud JL, Carter D, Baserga R and
Glazer PM: Insulin-like growth factor-I receptor overexpression
mediates cellular radioresistance and local breast cancer
recurrence after lumpectomy and radiation. Cancer Res.
57:3079–3083. 1997.PubMed/NCBI
|
|
52
|
Powell MJ, Dufault SM, Henry JE, Allison
AC, Cora R and Benz CC: Pregnancy hypertension and a commonly
inherited IGF1R variant (rs2016347) reduce breast cancer risk by
enhancing mammary gland involution. J Oncol. 2019:60184322019.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wu H, Sun T and Bi R: Inhibition of
insulin-like growth factor 1 signaling synergistically enhances the
tumor suppressive role of triptolide in triple-negative breast
cancer cells. Oncol Lett. 18:822–829. 2019.PubMed/NCBI
|
|
54
|
Riedemann J and Macaulay VM: IGF1R
signalling and its inhibition. Endocr Relat Cancer. 13(Suppl 1):
S33–S43. 2006. View Article : Google Scholar
|
|
55
|
Jonassen AK, Sack MN, Mjos OD and Yellon
DM: Myocardial protection by insulin at reperfusion requires early
administration and is mediated via Akt and p70s6 kinase
cell-survival signaling. Circ Res. 89:1191–1198. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Nitulescu GM, Van De Venter M, Nitulescu
G, Ungurianu A, Juzenas P, Peng Q, Olaru OT, Grădinaru D, Tsatsakis
A, Tsoukalas D, et al: The Akt pathway in oncology therapy and
beyond (Review). Int J Oncol. 53:2319–2331. 2018.PubMed/NCBI
|
|
57
|
Mundi PS, Sachdev J, McCourt C and
Kalinsky K: AKT in cancer: New molecular insights and advances in
drug development. Br J Clin Pharmacol. 82:943–956. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hartog H, Van Der Graaf WT, Boezen HM and
Wesseling J: Treatment of breast cancer cells by IGF1R tyrosine
kinase inhibitor combined with conventional systemic drugs.
Anticancer Res. 32:1309–1318. 2012.PubMed/NCBI
|
|
59
|
de Lint K, Poell JB, Soueidan H,
Jastrzebski K, Vidal Rodriguez J, Lieftink C, Wessels LF and
Beijersbergen RL: Sensitizing triple-negative breast cancer to PI3K
inhibition by cotargeting IGF1R. Mol Cancer Ther. 15:1545–1556.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen X, Cui D, Bi Y, Shu J, Xiong X and
Zhao Y: AKT inhibitor MK-2206 sensitizes breast cancer cells to
MLN4924, a first-in-class NEDD8-activating enzyme (NAE) inhibitor.
Cell Cycle. 17:2069–2079. 2018. View Article : Google Scholar : PubMed/NCBI
|