Introduction
Esophageal cancer (EC) is a refractory and common
invasive disease (1,2). Statistically, 17,290 patients were
diagnosed with EC and this led to 15,850 deaths in the United
States in 2018 (3). Esophageal
squamous cell carcinoma (ESCC) is one of the two main pathological
types (ESCC and adenocarcinoma) of EC, accounting for approximately
90% of cases (4). At present,
radical surgery remains the only treatment strategy for patients
with ESCC to achieve long-term survival. However, patients with
ESCC are associated with an extremely poor prognosis due to
recurrence and metastasis (5,6).
Therefore, it is necessary to examine the mechanisms responsible
for the development of ESCC and to provide novel strategies for the
prevention and treatment of ESCC.
Long non-coding RNAs (lncRNAs) are emerging
regulators of gene expression and cell fate (7,8).
Studies have demonstrated that many lncRNAs play differential roles
in the development of various types of tumor via the activation or
inhibition of oncogenes (8,9).
The lncRNA, BRAF activated non-coding RNA (BANCR), was discovered
in melanoma cells and plays a vital role during melanoma cell
migration (10). In recent years,
a substantial amount of evidence has indicated that BANCR plays a
crucial role in the occurrence and progression of cancers, such as
papillary thyroid cancer (11),
colorectal cancer (12) and
hepatocellular cancer (13). In
addition, BANCR was implicated in the progression of ESCC (14,15). However, the underlying molecular
mechanisms of BANCR in the progression of ESCC have not yet been
well explained.
MicroRNAs (miRNAs or miRs) are short non-coding RNAs
(19-25 nucleotides in length) that primarily regulate gene
expression at the post-transcriptional level (16). At present, miRNAs are a promising
tool or target in diagnostics and clinical treatment (17). Li et al reported that
lncRNA ATB regulated kindling-2 expression via miR-200b, which
modulated the malignant behaviors of ESCC cells (18). Liu et al demonstrated that
lncRNA XIST regulated tumor growth via regulating the miR-34a/MET
axis in thyroid cancer (19). In
addition, the enhancement of lncRNA B3GALT5-AS1 has been shown to
impede colon cancer metastasis by suppressing miR-203 expression
(20). A fairly large number of
researches have revealed that miR-338-3p plays a vital role in the
progression of epithelial ovarian cancer (21), prostate cancer (22) and cervical cancer (23). Furthermore, miR-338-3p has been
shown to exert an antitumor effect on ESCC (24). Nevertheless, the potential
molecular regulatory mechanisms of miR-338-3p in ESCC remain
largely unknown.
The receptor tyrosine kinase insulin-like growth
factor 1 receptor (IGF1R) can bind to ligand that can activate the
P13K/AKT/mTOR pathway and the Raf/MEK/ERK pathway (25,26). IGF1R has been shown to be
significantly elevated in the majority of commonly human cancers. A
previous study indicated that IGF1R was also upregulated in ESCC
(27). Considering that BANCR and
IGF1R were upregulated in ESCC, the present study thus investigated
whether BANCR plays a role in the progression of ESCC by regulating
IGF1R expression.
Hence, the present study detected the expression
patterns of BANCR and IGF1R in ESCC tissues and cells. Moreover,
the biological function of BANCR and IGF1R in ESCC cells were
investigated. In addition, the molecular mechanisms by which BANCR
regulates IGF1R expression were investigated.
Materials and methods
Tissue samples
The protocols were ratified by The Affiliated
Huai'an No. 1 People's Hospital of Nanjing Medical University. A
total of 40 paired ESCC tissues and adjacent non-tumor tissues were
obtained from The Affiliated Huai'an No. 1 People's Hospital of
Nanjing Medical University. The excised tissue was immediately
frozen in liquid nitrogen and stored at -80°C until use in
subsequent experiments. The histology and pathology of all biopsy
samples were examined by 2 independent pathologists. The
participating patients did not receive chemotherapy or radiotherapy
prior to surgery. Informed consents were obtained from all the
subjects participating in the study. The characteristics of the
patients with ESCC are presented in Table I.
 | Table ICharacteristics of patients with
esophageal squamous cell carcinoma. |
Table I
Characteristics of patients with
esophageal squamous cell carcinoma.
| Parameter | No. of patients
(n=40) |
|---|
| Sex | |
| Male | 28 |
| Female | 12 |
| Age, years | |
| <60 | 15 |
| ≥60 | 25 |
| Tumor location
(thoracic portion) | |
| Upper | 8 |
| Middle | 27 |
| Lower | 5 |
| Tumor size, cm | |
| <4 | 23 |
| ≥4 | 17 |
| Histological
grade | |
| Well | 13 |
| Moderate | 17 |
| Poor | 10 |
| Tumor stage | |
| I/II | 26 |
| III | 14 |
| Lymph node
metastasis | |
| Yes | 27 |
| No | 13 |
Cells, cell culture and transfection
KYSE450 and KYSE510 cells were obtained from Bena
Culture Collection. The normal human esophageal epithelial cell
line, HET-1A, was purchased from the American Tissue Culture
Collection (ATCC). The above-mentioned cells were maintianed at
37°C in an incubator with 5% CO2. Roswell Park Memorial
Institute (RPMI)-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) was utilized to culture all cells. Moreover, the medium was
replenished with 10% fetal bovine serum (HyClone; GE Healthcare
Life Sciences), and 1% penicillin/streptomycin (Baomanbio).
Small interference RNA (siRNA) targeting BANCR
(si-BANCR-1 and si-BANCR-2), siRNA targeting IGF1R (si-IGF1R-1 and
si-IGF1R-2), and their negative control (si-NC) were obtained from
GenePharma. For BANCR and IGF1R overexpression plasmids, the
full-length sequences of BANCR (NR_047671) and IGF1R (NM_000875)
were inserted into pcDNA3.1 vectors (Invitrogen; Thermo Fisher
Scientific, Inc.), respectively. miR-338-3p mimic (miR-338-3p) and
matching control (miR-NC) were obtained from Guangzhou RiboBio Co.,
Ltd. The transfection of the KYSE450 and KYSE510 cells was
performed using Lipofectamine 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). The concentrations used for transfection
were as follows: si-BANCR-1 (50 nM), si-BANCR-2 (50 nM), si-IGF1R-1
(50 nM), si-IGF1R-2 (50 nM), si-NC (50 nM), miR-338-3p (40 nM),
miR-NC (40 nM), pcDNA (15 nM), BANCR (15 nM), IGF1R (15 nM). The
sequences of the siRNAs were as follows: si-BANCR-1, 5′-GGA CUC CAU
GGC AAA CGU UTT-3′; si-BANCR-2, 5′-GGA AAU AGA CUG CAG CAC CAA
TT-3′; si-IGF1R-1, 5′-CAA CAG UGG UCA UCA UGG AAC UGU TT-3′;
si-IGF1R-2, 5′-UGA CUG UGA AAU CUU CGG CTT-3′; and si-NC, 5′-TTC
TCC GAA CGT GTC ACG T-3′. The cells were collected for subsequent
analysis after 48 h of transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
For the expression levels of BANCR, miR-338-3p and
IGF1R mRNA in ESCC tissues, total RNA was extracted from 40 paired
ESCC tissues (at stages I, II and III disease) and adjacent
non-tumor tissues for RT-qPCR. ESCC tissues or cells were
homogenized using the TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) to extract total RNA. The complementary DNA
(cDNA) of BANCR, miR-338-3p and IGF1R was generated using the Prime
ScriptTM RT reagent kit (Takara Bio, Inc.). The Platinum SYBR-Green
qPCR SuperMix UDG from Invitrogen; Thermo Fisher Scientific, Inc.
was employed for qPCR. The relative expression of BANCR, IGF1R and
miR-338-3p was calculated using the 2−ΔΔCq method
(28), and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or U6 small
nuclear RNA (snRNA) were used as the internal reference genes. The
primers for amplification were as follows: GAPDH forward, 5′-GAC
TCC ACT CAC GGC AAA TTCA-3′ and reverse, 5′-TCG CTC CTG GAA GAT GGT
GAT-3′; BANCR forward, 5′-ACA GGA CTC CAT GGC AAA CG-3′ and
reverse, 5′-ATG AAG AAA GCC TGG TGC AGT-3′; IGF1R forward, 5′-GCG
GTT CTG TTG ATA GTG G-3′ and reverse, 5′-GCC TCG TTC ACC GTC
TTA-3′; U6 snRNA forward, 5′-GCT CGC TTC GGC AGC ACA-3′ and
reverse, 5′-GAG GTA TTC GCA CCA GAG GA-3′; and miR-338-3p forward,
5′-TGC GGT CCA GCA TCA GTG AT-3′ and reverse, 5′-CCA GTG CAG GGT
CCG AGG T-3′.
Western blot analysis
For the levels of IGF1R in ESCC tissues, 3 pairs of
tissue samples were selected for western blot analysis. In brief,
the ESCC tissues or cells were lysed in RIPA lysis buffer (Beyotime
Institute of Biotechnology, Inc.) to obtain total protein.
Subsequently, 8-10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) was utilized for total protein (30
µg) isolation. Thereafter, the isolated proteins were
electrotransferred onto polyvinylidene fluoride (PVDF) membranes
(EMD Millipore). Tris-buffered saline with 0.05% Tween-20 (TBST)
was employed to block the PVDF membranes. The PVDF membranes were
then incubated with primary antibodies at 4°C for overnight.
Subsequently, the PVDF membranes were incubated with a goat
anti-mouse (#7076, 1:1,000) or rabbit IgG (#4414, 1:1,000) (from
Cell Signaling Technology, Inc.) for 1 h at 37°C. The signal was
captured with an enhanced chemiluminescence (ECL) detection system
(Thermo Fisher Scientific, Inc.). Every protein band was analyzed
with the Image Lab™ Software (Bio-Rad Laboratories, Inc.). The
primary anti-bodies used were as follows: Anti-IGF1R (sc-463;
1:500, Santa Cruz Biotechnology, Inc.), anti-p-MEK1/2 (#2338,
1:2,000), anti-MEK1/2 (#9122, 1:1,000) (both from Cell Signaling
Technology, Inc.), anti-E-cadherin (sc-8426, 1:200),
anti-N-cadherin (sc-393933, 1:100), anti-p-ERK1/2 (sc-81492,
1:200), anti-ERK1/2 (sc-81504, 1:200), anti-Vimentin (sc-32322,
1:200), anti-p-Raf-1 (sc-81513, 1:200), anti-Raf-1 (sc-52827,
1:200) and anti-GAPDH (sc-32233, 1:1,000) (all from Santa Cruz
Biotechnology, Inc.).
Cell proliferation assay
Cell proliferation was evaluated using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazo-lium bromide (MTT)
kit (Promega Corporation) according to the manufacturer's protocol.
In brief, the transfected KYSE450 and KYSE510 cells (5,000
cells/well) were seeded into 96-well plates and cultured for 24, 48
or 72 h at 37°C. Thereafter, 20 µl MTT (5 mg/ml) was
supplemented to each well and maintained at 37°C for 4 h.
Subsequently, 200 µl dimethyl sulfoxide (DMSO) was employed
for the dissolution of the formazan crystals. Finally, the color
reaction at 490 nm was measured using a Microplate Absorbance
Reader (Thermo Fisher Scientific, Inc.).
Transwell assay
A Transwell chamber (8 µm pore filter) from
BD Biosciences was employed to evaluate the migration and invasion
of the transfected KYSE450 and KYSE510 cells. Transwell assay was
performed as previously described (29). The transfected KYSE450 and KYSE510
cells (1×106) were placed into the upper chamber
containing RPMI without FBS. Synchronously, RPMI medium with FBS
(10%) was added to the lower chamber. For the invasion assay, the
Matrigel matrix (BD Biosciences) was filled in the upper chamber.
Subsequently, the cells on the lower surface of the membrane were
fixed with glutaraldehyde (5%). Crystal violet (Solarbio, Inc.)
(0.1%) was then added to stain the cells for 10 min at room
temperature. Finally, the number of the migrated and invasive cells
was counted under an inverted microscope (Olympus Corp.).
Dual-luciferase reporter assay
The bioinformatics database starBase2.0 (http://starbase.sysu.edu.cn/starbase2/index.php) was
employed to predict the binding sites between miR-338-3p and BANCR
or IGF1R, as previously described (30). The sequences of wild-type (WT)
BANCR (AUGCUGG), mutant (MUT) BANCR (UACGACC), the 3′-untranslated
region (UTR) of WT IGF1R (UGCUGG), and the 3′UTR of mut IGF1R
(ACGACC) containing assumed miR-338-3p binding sites were
synthesized. The sequences were then inserted into the pGL3-control
luciferase reporter vectors (Promega Corp.) with a restriction
endonuclease for 16 h, respectively. Subsequently, the KYSE450 and
KYSE510 cells were co-transfected with luciferase reporter vectors
and miR-338-3p or miR-NC. Following transfection for 48 h, the
supernatant of the cell lysate was acquired through centrifugation
(5.000 × g) for 5 min at 4°C. The relative luciferase units were
evaluated with luciferase reporter assay kit (Promega Corp.). The
relative fluorescence value was acquired by dividing the relative
luciferase units value determined by Renilla luciferase by
the relative luciferase units determined by Firefly luciferase.
In vivo experiment
The protocols of the animal experiment in the
present study were authorized by The Affiliated Huai'an No. 1
People's Hospital of Nanjing Medical University. The
lentivirus-mediated sh-BANCR was obtained from GenePharma. A total
of 14 female BALB/c nude mice (4-6 weeks old) were obtained from
Shanghai Experimental Animal Center. All mice were randomly divided
into 2 groups (n=7). The growth rate of the transplanted tumors was
positively associated with body weight. However, the weight growth
rate of mice is related to sex; thus, in the present study, female
mice were used in xenotransplantation experiments as they are less
aggressive and are less likely to inflict injury on one another.
The in vivo experiments were performed as previously
described (18,31,32). All mice were kept under specific
pathogen-free conditions (25±2°C, 50-60% relative humidity, and a
12-h light/dark cycle) and food and water were provided ad
libitum. For the stable knockdown of BANCR, the
lentiviral-mediated sh-BANCR was transfected with 293T cells (ATCC)
and then infected with KYSE450 cells with the supernatant of the
293T cells. The KYSE450 (1×107) cells stably transfected
with lentivirus-mediated sh-BANCR or sh-NC were suspended in 100
µl phosphate-buffered saline. The KYSE450 cells were
subcutaneously injected into the right flank of each mouse. Tumor
volume was measured once a week using a caliper. After 35 days, all
mice were euthanized (cervical decapitation) following an injection
of xylazine (10 mg/kg, ChemeGen, Inc.) for subsequent analysis.
Tumor volume was assessed with the equation: Volume = (length x
width2)/2.
Statistical analysis
Data are presented as the means ± standard error.
Prism5 software (GraphPad, Inc.) and SPSS 13.0 software (SPSS,
Inc.) were utilized to conduct the statistical analysis. The
correlation between BANCR and IGF1R was assessed by Spearman's
correlation analysis. The Student's t-test or one-way ANOVA
followed by Tukey's post hoc test were applied to compare
differences between 2 groups and multiple groups, respectively. The
overall survival of the patients with ESCC was determined by
Kaplan-Meier analysis with the log-rank test. A value of P<0.05
was considered to indicate a statistically significant
difference.
Results
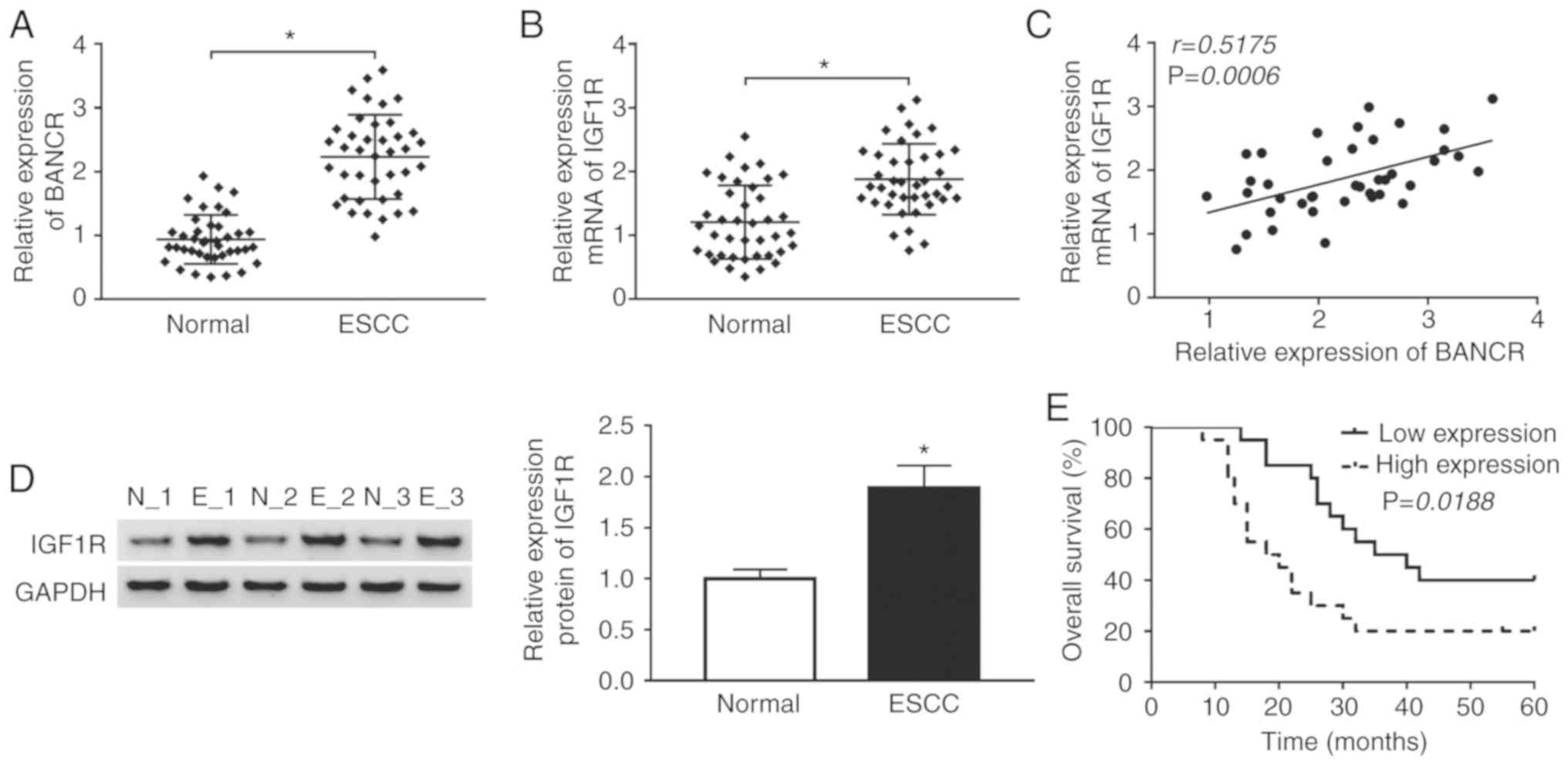
BANCR and IGF1R expression exhibit a
positive association in human ESCC tissues
Given that BANCR and IGF1R have been shown to be
upregulated in ESCC tissues (14,26), the present study thus examined the
association between BANCR and IGF1R in ESCC. The expression levels
of BANCR and IGF1R were first examined in 40 paired ESCC tissues
and adjacent normal tissues by RT-qPCR. The results indicated that
BANCR and IGF1R were upregulated in ESCC tissues when compared with
the adjacent normal tissues (Fig. 1A
and B). Spearman's correlation analysis revealed that the
expression of BANCR positively correlated with that of IGF1R in
ESCC tissues (Fig. 1C). Moreover,
western blot analysis revealed that the protein level of IGF1R was
increased in ESCC tissues compared with adjacent normal tissues
(Fig. 1D). In addition, it was
found that patients with ESCC with a high BANCR expression
exhibited a lower survival rate than patients with ESCC with a low
BANCR expression (Fig. 1E). These
results suggested that BANCR and IGF1R were involved in the
progression of ESCC.
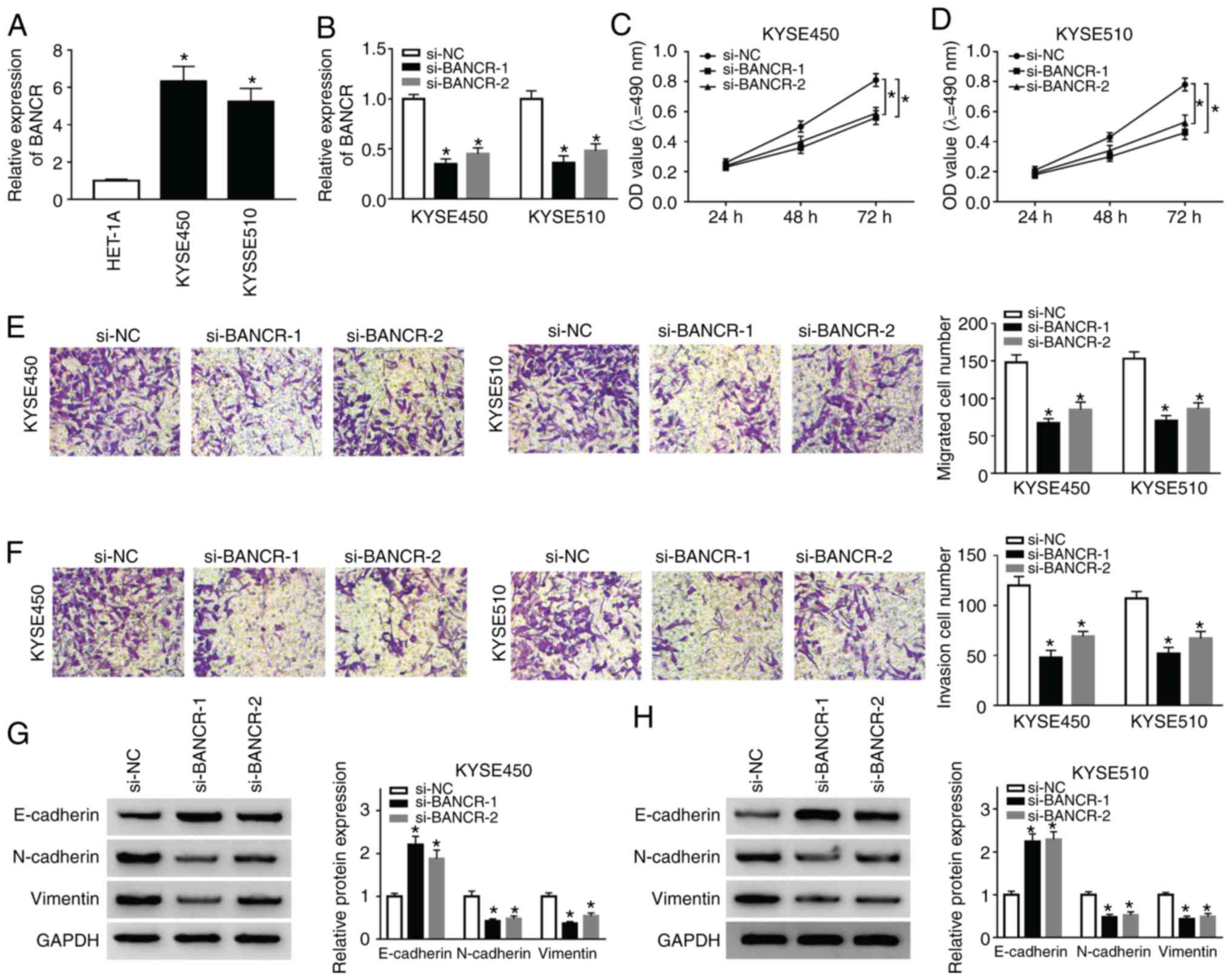
Knockdown of BANCR inhibits the
proliferation, migration, invasion and EMT of human ESCC cells
To investigate the function of BANCR in the
progression ESCC, the expression of BANCR was examined in ESCC
(KYSE450 and KYSE510) and HET-1A cells. The results indicated that
the expression of BANCR was significantly higher in ESCC cells than
in HET-1A cells (Fig. 2A).
Subsequently, the KYSE450 and KYSE510 cells were transfected with
si-BANCR-1 and si-BANCR-2 to silence BANCR. As shown in Fig. 2B, the expression of BANCR was
decreased in the KYSE450 and KYSE510 cells transfected with
si-BANCR-1 and si-BANCR-2. Subsequently, MTT assay was performed
and the results revealed the marked suppression of cell
proliferation by BANCR silencing in both the KYSE450 and KYSE510
cells (Fig. 2C and D). Moreover,
Transwell migration assay revealed that the number of migrated
cells was decreased in the BANCR-silenced KYSE450 and KYSE510 cells
(Fig. 2E). Consistently,
Transwell invasion assay displayed that BANCR silencing inhibited
the invasion of both KYSE450 and KYSE510 cells (Fig. 2F). Studies have indicated that
KYSE450 and KYSE510 cells can simultaneously express EMT-related
markers (E-cadherin, N-cadherin and Vimentin) (33-35); thus, the present study assessed
the effects of BANCR depletion on the levels of E-cadherin,
N-cadherin and Vimentin by western blot analysis. The results
revealed that the level of E-cadherin was distinctly increased in
the KYSE450 and KYSE510 cells by BANCR downregulation, while the
levels of N-cadherin and Vimentin exhibited an opposite trend
(Fig. 2G and H). These results
revealed that BANCR repression inhibited the proliferation,
migration, invasion and EMT of ESCC cells.
IGF1R silencing suppresses the
proliferation, migration, invasion of and EMT of ESCC cells
Subsequently, the role of IGF1R in ESCC was examined
by loss-of-function experiments. As shown in Fig. 3A, the mRNA expression of IGF1R was
markedly elevated in the KYSE450 and KYSE510 cells compared with
the HET-1A cells. Consistently, the protein levels of IGF1R were
markedly enhanced in the KYSE450 and KYSE510 cells when compared to
the HET-1A cells (Fig. 3B).
However, IGF1R mRNA and protein levels were decreased in the
KYSE450 and KYSE510 cells following si-IGF1R-1 or si-IGF1R-2
transfection (Fig. 3C and D). The
results of MTT assay revealed that IGF1R silencing significantly
suppressed the proliferation of hte KYSE450 and KYSE510 cells
compared with the si-NC control (Fig.
3E and F). Furthermore, Transwell assay revealed that the
number of migratory and invasive cells was markedly decreased in
the IGF1R-silenced KYSE450 and KYSE510 cells (Fig. 3G and H). Additionally, IGF1R
knockdown enhanced the E-cadherin level, and reduced N-cadherin and
Vimentin levels in the KYSE450 and KYSE510 cells (Fig. 3I and J). Overall, these results
proved that IGF1R depletion suppressed the proliferation,
migration, invasion and EMT of ESCC cells.
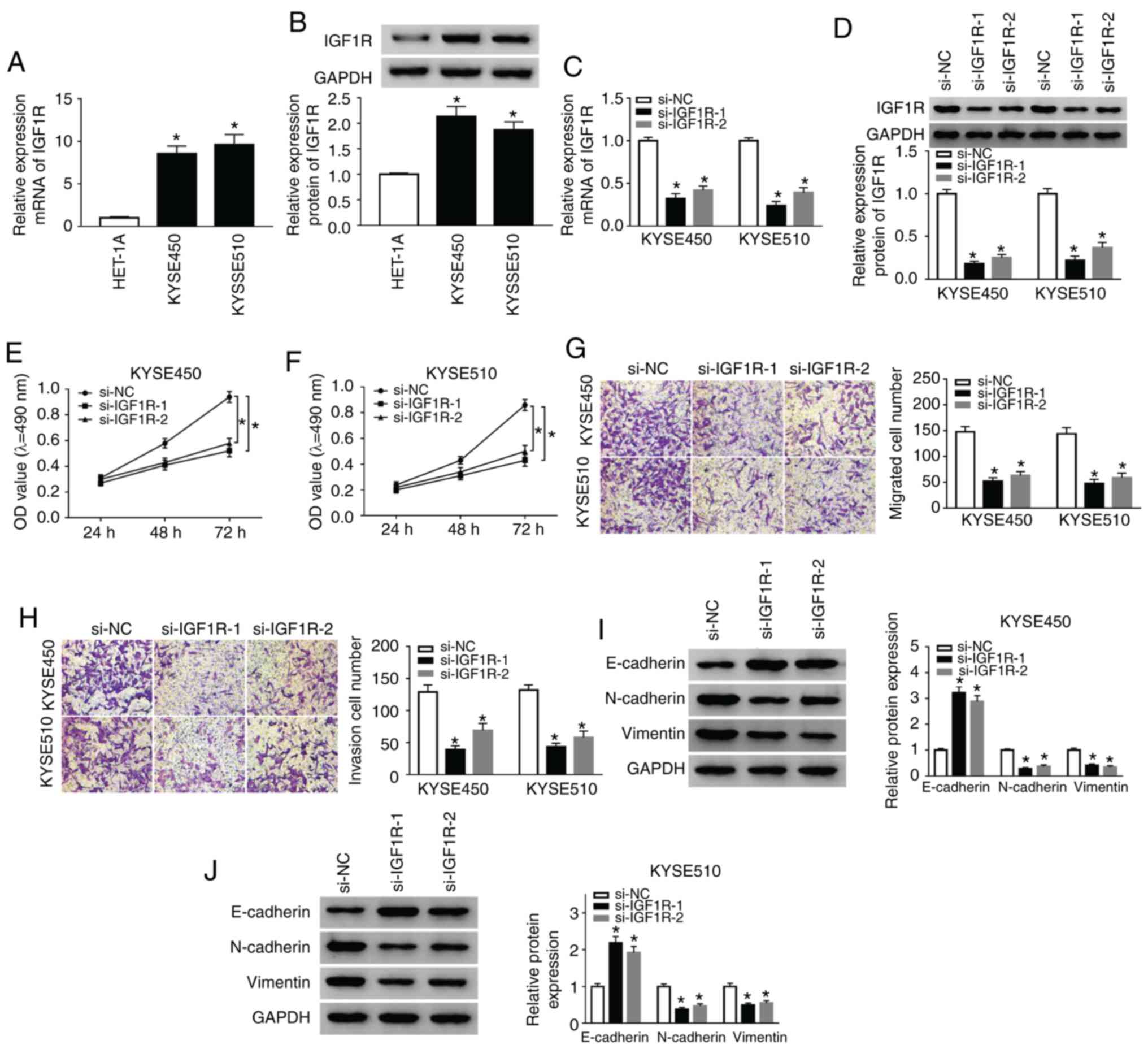 | Figure 3Silencing of IGF1R suppresses the
proliferation, migration, invasion and EMT of ESCC cells. (A)
RT-qPCR was conducted to detect the expression of IGF1R in ESCC and
HET-1A cells. (B) Western blot analysis of IGF1R protein expression
in ESCC and HET-1A cells was implemented. (C-J) KYSE510 cells were
transfected with si-NC, si-IGF1R-1, or si-IGF1R-2. (C and D)
RT-qPCR or western blot analysis were used to assess the expression
of IGF1R mRNA and protein in KYSE450 and KYSE510 cells. (E-H) MTT
or Transwell assays was utilized for the determination of the
proliferation, migration, and invasion of KYSE450 and KYSE510
cells. (I and J) The levels of E-cadherin, N-cadherin, and Vimentin
in KYSE450 and KYSE510 cells were evaluated by western blot
analysis. Data are presented as the means ± standard error of 3
experimental results. One-way ANOVA was applied to compare
differences between groups. *P<0.05. BANCR, BRAF
activated non-coding RNA; IGF1R, insulin-like growth factor 1
receptor; ESCC, esophageal squamous cell carcinoma. |
IGF1R overexpression reverses the effects
of BANCR knock-down on the proliferation, migration, invasion and
EMT of ESCC cells
Based on the above-mentioned results, the present
study further examined whether BANCR carries out its role in ESCC
through IGF1R. The results of RT-qPCR demonstrated that the
expression of BANCR was markedly upregulated in the KYSE450 and
KYSE510 cells transfected with BANCR over-expression plasmid
compared with the control group (Fig.
4A). In addition, the mRNA and protein levels of IGF1R were
markedly elevated in the KYSE450 and KYSE510 cells transfected with
IGF1R overexpression plasmid (Fig. 4B
and C). Moreover, IGF1R mRNA and protein levels were suppressed
by BANCR knockdown and enhanced by BANCR overexpression in the
KYSE450 and KYSE510 cells (Fig.
4D-F). The results of MTT assay also indicated that IGF1R
overexpression markedly attenuated the inhibitory effects of BANCR
knockdown on the proliferation of the KYSE450 and KYSE510 cells
(Fig. 4G and H). As was expected,
the overexpression of IGF1R abrogated the inhibitory effects of
BANCR knockdown on the migration and invasion of the KYSE450 and
KYSE510 cells (Fig. 4I and J). In
addition, IGF1R overexpression reversed the effects of BANCR
knockdown on the levels of E-cadherin, N-cadherin and Vimentin in
the KYSE450 and KYSE510 cells (Fig.
4K and L). Taken together, these data indicated that BANCR
regulated the proliferation, migration, invasion and EMT of ESCC
cells via IGF1R.
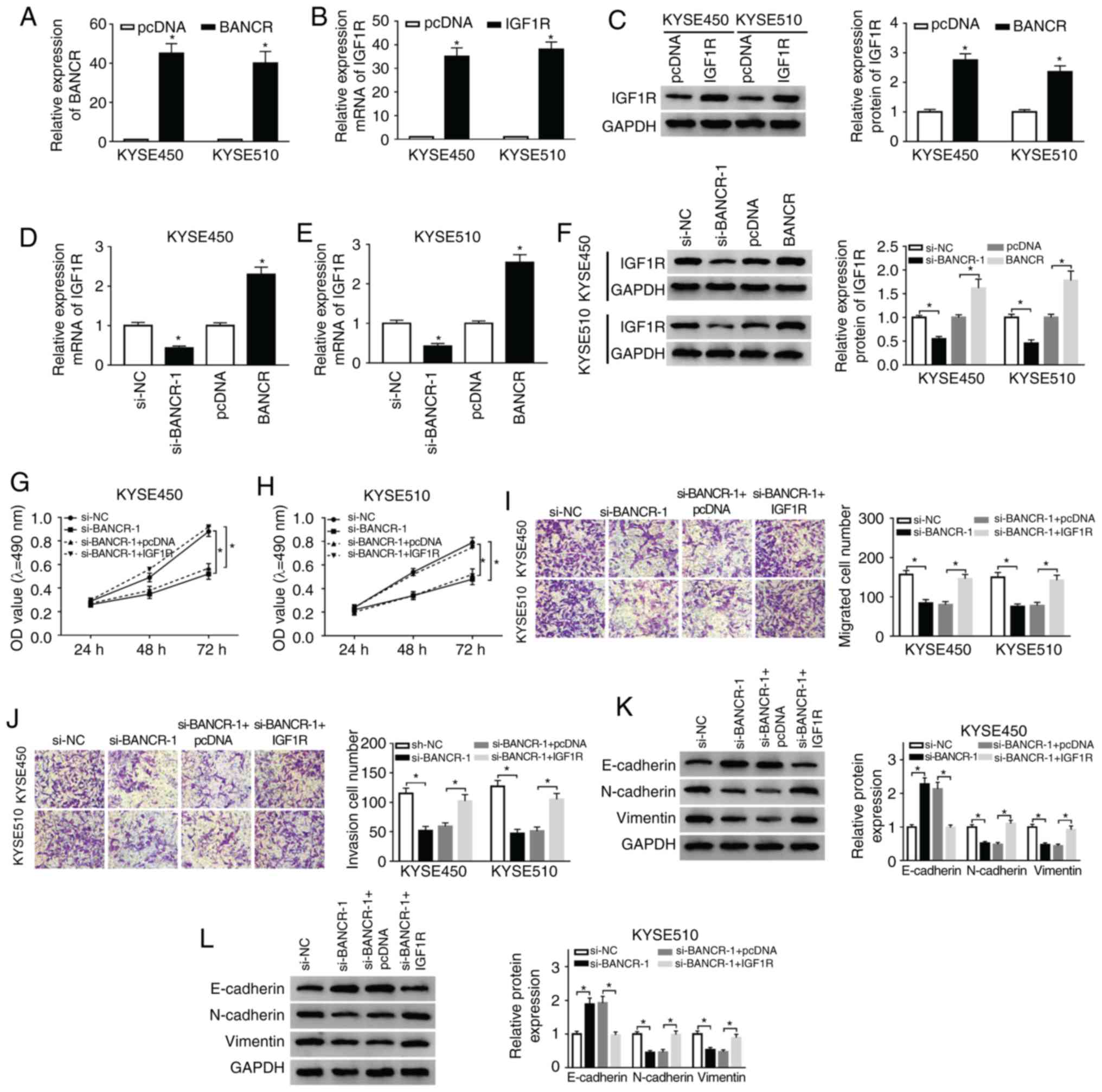 | Figure 4Effects of IGF1R overexpression on
the proliferation, migration, invasion and EMT of ESCC cells
induced by BANCR knockdown. (A) RT-qPCR was performed for the
detection of the expression of BANCR in KYSE450 and KYSE510 cells
transfected with cDNA or BANCR. (B and C) The levels of IGF1R mRNA
and protein in KYSE450 and KYSE510 cells transfected with cDNA or
IGF1R were measured by RT-qPCR or western blot analysis. (D-F)
Effects of BANCR knockdown or overexpression on the levels of IGF1R
mRNA and protein were determined by RT-qPCR or western blot
analysis. (G-L) KYSE450 and KYSE510 cells were transfected with
si-NC, si-BANCR, si-BANCR+pcDNA, or si-BANCR+IGF1R. (G-J) The
proliferation, migration and invasion of KYSE450 and KYSE510 cells
were evaluated by MTT or Transwell assays. (K and L) The levels of
E-cadherin, N-cadherin and Vimentin in KYSE450 and KYSE510 cells
were examined by western blot analysis. Data were presented as the
means ± standard error of 3 experimental results. The Student's
t-test or one-way ANOVA were applied to compare differences between
groups where appropriate. *P<0.05. BANCR, BRAF
activated non-coding RNA; IGF1R, insulin-like growth factor 1
receptor; ESCC, esophageal squamous cell carcinoma. |
BANCR regulates IGF1R expression by
targeting miR-338-3p
In view of the above-mentioned results, the present
study further explored the latent molecular mechanisms between
BANCR and IGF1R. It was discovered that miR-338-3p had
complementary bases pairing with BANCR and IGF1R through
starBase2.0 (Fig. 5A). Moreover,
miR-338-3p expression was markedly decreased in ESCC tissues and
cells (Fig. 5B and C). In
comparison to miR-NC, the expression of miR-338-3p was overtly
increased in the KYSE450 and KYSE510 cells following transfection
with miR-338-3p mimics (Fig. 5D).
The results of dual-luciferase reporter assay revealed that the
luciferase activity of the luciferase reporters containing WT-BANCR
was markedly decreased in the miR-338-3p-overexpressing KYSE450 and
KYSE510 cells, whereas the luciferase reporters containing
MUT-BANCR were not affected (Fig. 5E
and F). Furthermore, miR-338-3p expression was markedly
elevated by BANCR silencing and was overtly reduced by BANCR
over-expression in the KYSE450 and KYSE510 cells (Fig. 5G). The overexpression of
miR-338-3p also suppressed the luciferase intensity of the
luciferase reporters with IGF1R 3′UTR-WT in both KYSE450 and
KYSE510 cells, while there was no marked difference in the
luciferase reporters with IGF1R 3′UTR-MUT (Fig. 5H and I). Furthermore, it was
observed that miR-338-3p overexpression abnormally suppressed the
protein levels of IGF1R in the KYSE450 and KYSE510 cells, while
this reduction was abrogated by BANCR overexpression (Fig. 5J). In addition, the suppressive
effects of miR-338-3p overexpression on the proliferation of the
KYSE450 and KYSE510 cells were attenuated by BANCR overexpression
(Fig. 5K and L). Transwell assay
also revealed that BANCR elevation reversed the inhibitory effects
of miR-338-3p upregulation on the migration and invasion of KYSE450
and KYSE510 cells (Fig. 5M and
N). Taken together, these results indicated that BANCR
regulated the expression of IGF1R by targeting miR-338-3p in ESCC
cells.
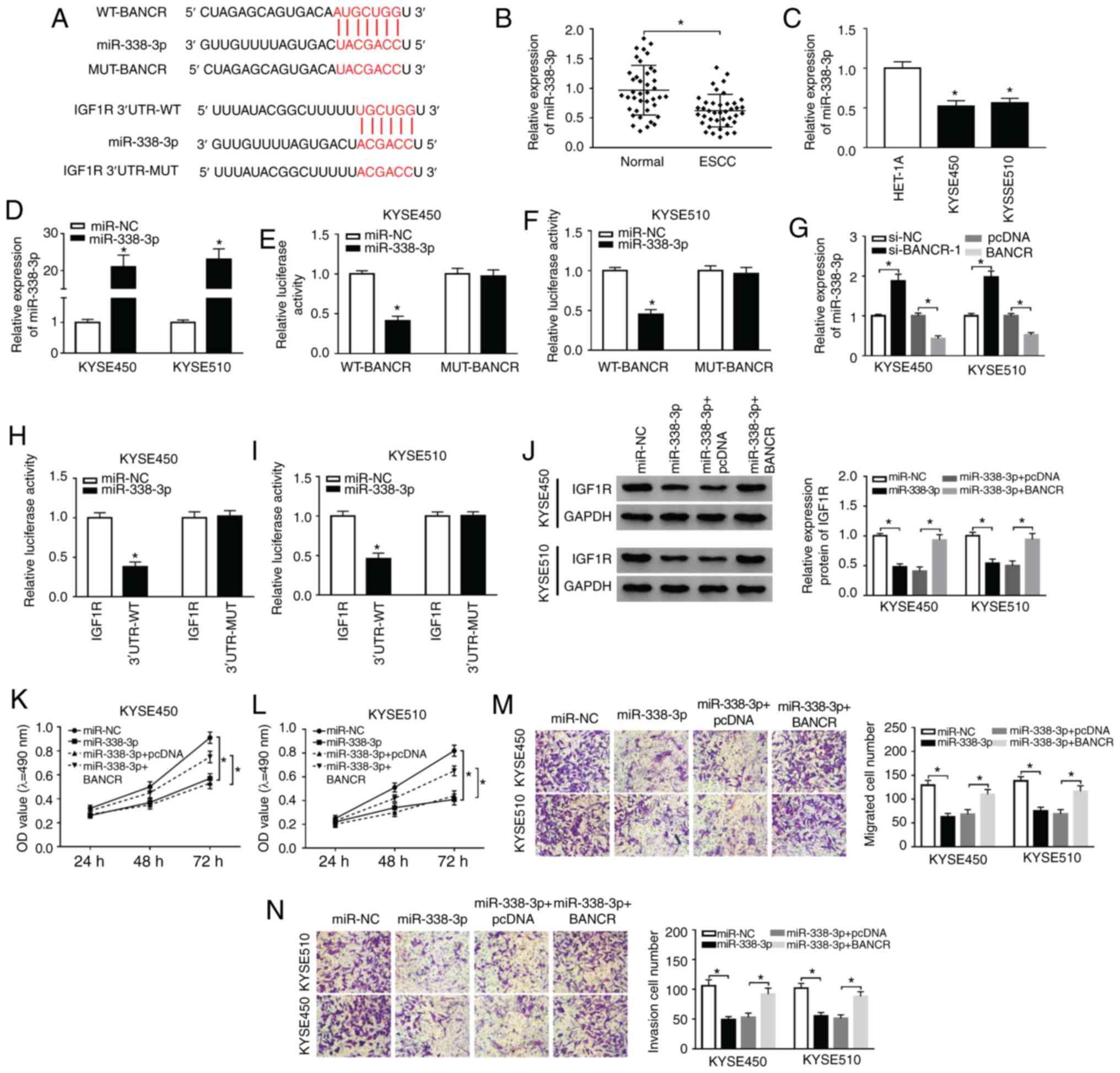 | Figure 5BANCR modulates IGF1R expression via
sponging miR-338-3p. (A) The potential binding sites between
miR-338-3p and BANCR or IGF1R were predicted using starBase2.0. (B
and C) The expression of miR-338-3p in ESCC tissues and adjacent
normal tissues, as well as ESCC cells and HET-1A cells, was
detected by RT-qPCR. (D) Following miR-NC or miR-338-3p
transfection, the levels of miR-338-3p in KYSE450 and KYSE510 cells
were detected by RT-qPCR. (E and F) Dual-luciferase reporter assay
was conducted for the determination of the luciferase activity in
KYSE450 and KYSE510 cells co-trans-fected luciferase reporters
containing WT-BANCR or MUT-BANCR with miR-NC or miR-338-3p. (G)
Effects of BANCR knockdown or overexpression on the expression of
miR-338-3p were determined by RT-qPCR. (H and I) The luciferase
activity of luciferase reporters containing IGF1R 3′UTR-WT or IGF1R
3′UTR-MUT in KYSE450 and KYSE510 cells transfected with miR-NC or
miR-338-3p was detected using dual-luciferase reporter assay. (J-N)
KYSE450 and KYSE510 cells were transfected with miR-NC, miR-338-3p,
miR-338-3p+cDNA, or miR-338-3p+BANCR. (J) The levels of IGF1R
protein in KYSE450 and KYSE510 cells were measured via using
western blotting. (K-N) MTT or Transwell assays was carried out to
analyze the proliferation, migration, and invasion of KYSE450 and
KYSE510 cells. Data are presented as the means ± standard error of
3 experimental results. The Student's t-test or one-way ANOVA were
applied to compare differences between groups where appropriate.
*P<0.05. BANCR, BRAF activated non-coding RNA; IGF1R,
insulin-like growth factor 1 receptor; ESCC, esophageal squamous
cell carcinoma. |
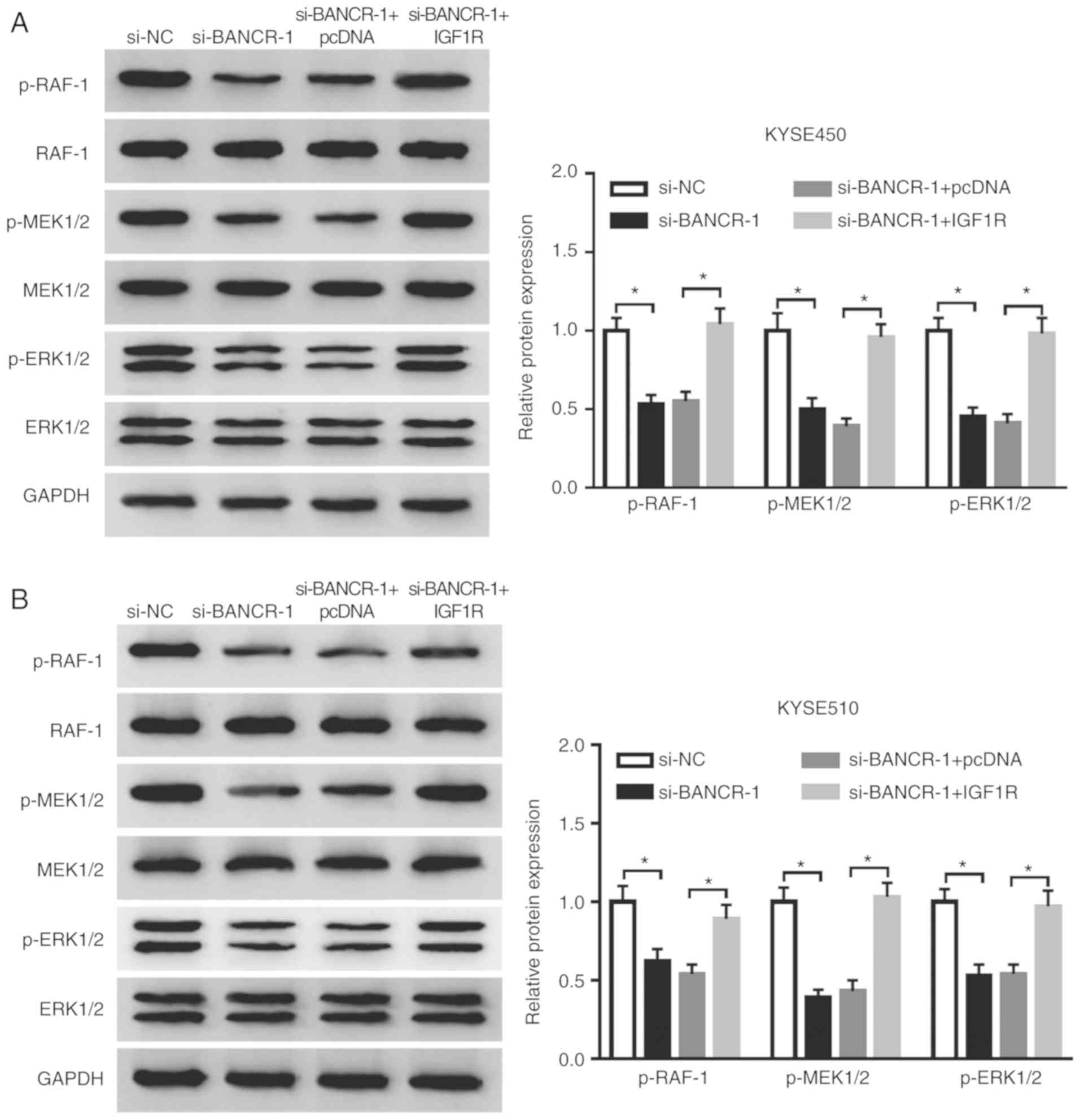
BANCR regulates the Raf/MEK/ERK pathway
by regulating IGF1R expression
Considering that IGF1R was involved in the
Raf/MEK/ERK pathway (36,37), the present study further examined
whether BANCR regulated the Raf/MEK/ERK pathway through IGF1R in
ESCC cells. The results indicated that the inhibition of BANCR
prominently reduced the levels of p-Raf-1, p-MEK1/2 and p-ERK1/2 in
both the KYSE450 and KYSE510 cells. However, this decrease was
attenuated by IGF1R overexpression in the KYSE450 and KYSE510 cells
(Fig. 6A and B). These results
indicated that BANCR regulated the Raf/MEK/ERK pathway by
modulating IGF1R expression.
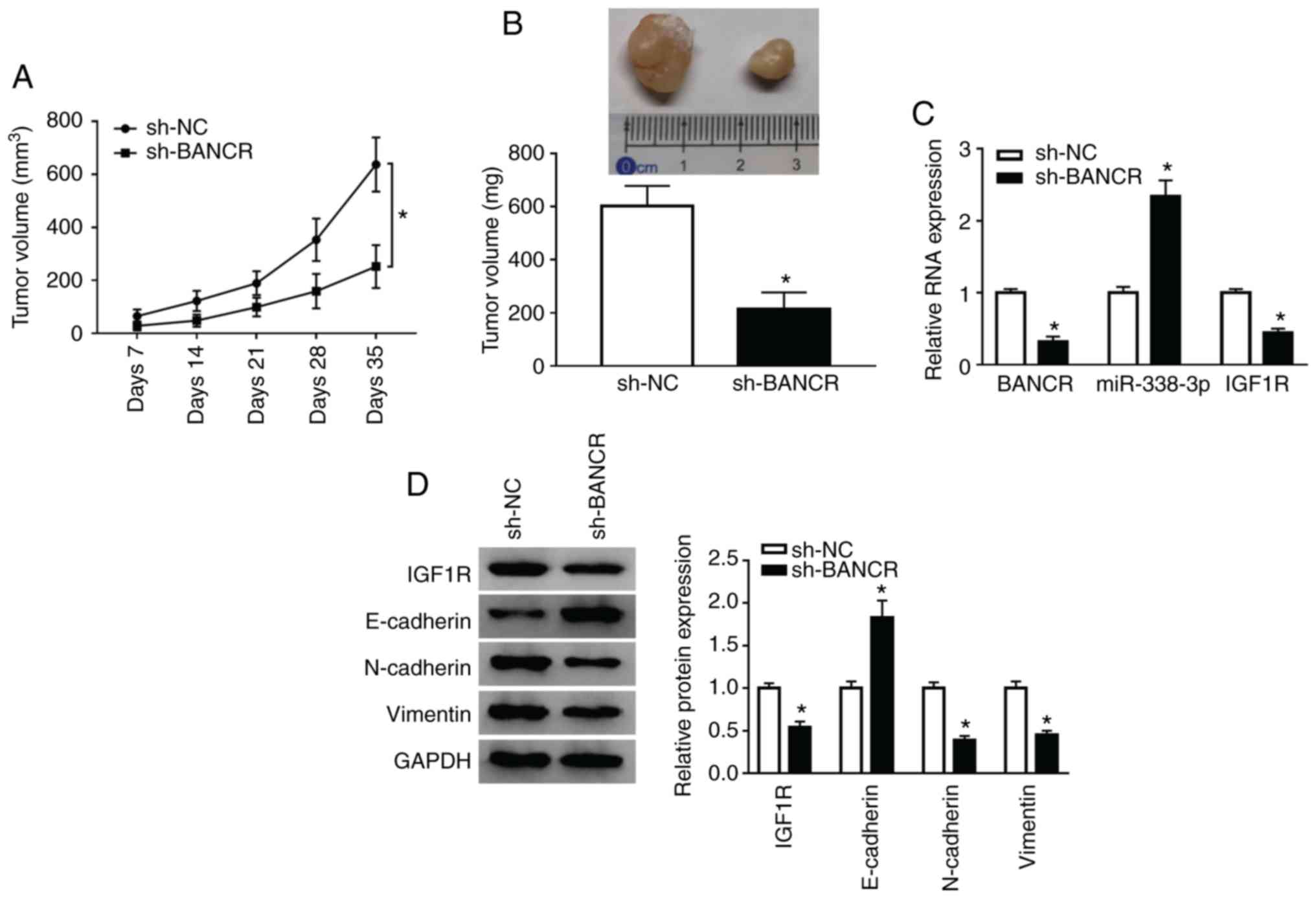
Knockdown of BANCR inhibits tumor growth
in vivo
To confirm the role of BANCR in vivo, KYSE450
cells stably transfected with lentivirus-mediated sh-BANCR or sh-NC
were subcutaneously injected into nude mice. BANCR knockdown
markedly inhibited tumor volume when compared with the control
group (Fig. 7A). In addition,
tumor weight was evidently decreased in the sh-BANCR group when
compared to the control group (Fig.
7B). Moreover, BANCR silencing prominently reduced the BANCR
and IGF1R levels, and enhanced the miR-338-3p levels in the tumor
tissues of mice (Fig. 7C).
Moreover, the levels of IGF1R, N-cadherin and Vimentin were
downregulated in the tumor tissues of mice in the sh-BANCR group,
whereas the E-cadherin levels were upregulated (Fig. 7D). These results indicated that
BANCR inhibition suppressed tumor growth in vivo.
Discussion
ESCC is a gastrointestinal malignant tumor, which is
often invasive and recurrent, has a poor prospect of survival in
patients with advanced disease (38). A substantial amount of evidence
has indicated that lncRNAs play vital roles in the progression of
ESCC (39,40). BANCR has been reported to play a
carcinogenic role in various types of cancer. For instance, Shen
et al indicated that a high BANCR expression was associated
with lymph node metastasis and a poor prognosis in colorectal
cancer (12). Another study also
reported that BANCR upregulation promoted the initiation and
progression of hepatocellular carcinoma (13). In the present study, BANCR
expression was elevated in ESCC tissues and cells. Moreover, BANCR
silencing suppressed tumor growth in vivo, and suppressed
the proliferation, migration, invasion and EMT of ESCC cells in
vitro. For in vivo experiments, only one cell line
(KYSE450) was selected for verification, which is a limitation of
the present study. A previous study indicated that BANCR
enhancement was associated with the development and a poor
prognosis in ESCC (14). Chen
et al demonstrated that BANCR silencing blocked cell
invasion and migration via inactivating the Wnt/β-catenin pathway
in ESCC (15). In addition, the
study of Sadeghpour and Ghorbian demonstrated that the BANCR
expression levels were positively associated with tumor stage,
tumor differentiation and lymph node metastasis (41). However, BANCR plays an antitumor
role in bladder cancer and non-small cell lung cancer, which may be
related to the differences in the tissues and microenvironment
(42,43).
Recently, increasing evidence has indicated that
lncRNAs can function as sponges for miRNAs (44). For example, lncRNA ZEB2-AS1
depletion has been shown to suppress HMGB1 expression via the
enhancement of miR-204 expression, which inhibits the invasion and
growth of pancreatic cancer cells (45). In addition, lncRNA SNHG20
downregulation has been shown to suppress cervical cancer
advancement by regulating ADAM10 expression through the targeting
of miR-140-5p (46). Herein,
BANCR regulated IGF1R expression via targeting miR-338-3p.
Moreover, miR-338-3p expression was decreased in ESCC tissues and
cells.
EMT is the transformation of epithelial cells into
active mesenchymal cells, which leads to invasion and migration
(47). Li et al reported
that miR-338-3p mimic suppressed the growth of tumor in ESCC in
vitro and in vivo (24), and the results of the present
study are consistent with this finding. The study by Cao et
al demonstrated that miR-338-3p overexpression suppressed cell
EMT, migration, proliferation and invasion in osteosarcoma in
vitro (48). Moreover,
miR-338-3p has also been shown to play an antitumor role in in
non-small cell lung cancer (49),
gastric cancer (50) and prostate
cancer (22).
The signaling pathway of Ras/Raf/MEK/ERK is usually
related to the development and progression of diverse types of
cancer (51). It has been
revealed that the RAS/Raf/MEK/ERK signaling pathway is a vital
downstream intracellular signal of IGF1R (36). A previous study demonstrated that
IGF1R played vital roles in cell apoptosis, angiogenesis,
proliferation and invasion in cancers (52). There is a growing body of evidence
to suggest that IGF1R expression is increased in various types of
tumor. For instance, Chen et al demonstrated that IGF1R
expression was markedly enhanced in renal cell cancer cells, and
the silencing of IGF1R suppressed cancer cell proliferation,
migration and invasion (53). In
the present study, it was found that IGF1R expression was augmented
in ESCC tissues and cells. Furthermore, the silencing of IGF1R
expression suppressed the proliferation, migration, invasion and
EMT of ESCC cells. Moreover, BANCR regulated the Raf/MEK/ERK
pathway by regulating IGF1R expression. Mei et al indicted
that IGF1R knockdown blocked the proliferation, migration, invasion
and slug-induced EMT of ESCC cells (33). Another study also indicated that
miR-98 elevation inhibited retinoblastoma cell growth and invasion
by targeting IGF1R/k-RAS/Raf/MEK/ERK pathway (37). Therefore, it was concluded that
BANCR silencing suppressed cell proliferation, migration, invasion
and EMT in ESCC through the inactivation of the IGF1R/Raf/MEK/ERK
pathway via sponging miR-338-3p. However, the present study only
confirmed that IGF1R was the target of miR-338-3p, and whether
miR-338-3p can target RAF, MEK or ERK in ESCC remains to be further
investigated in the future.
In conclusion, the present study demonstrated that
BANCR downregulation suppressed ESCC progression through the
inactivation of the IGF1R/Raf/MEK/ERK pathway by sponging
miR-338-3p, which may provide a possible target for the treatment
of ESCC.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
KW and XY were involved in the conceptualization and
methodology of the study. XY, WD and ZF were involved in formal
analysis and data curation. WS, KW and WD were involved in data
validation and investigation. WS, KW and XY were involved in the
writing and preparation of the original draft, and in the reviewing
and editing of the manuscript. All authors have read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Committee of The Affiliated Huai'an No. 1 People's Hospital of
Nanjing Medical University. Written informed consent was obtained
from all enrolled subjects.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
2
|
Kang X, Chen K, Li Y, Li J, D'Amico TA and
Chen X: Personalized targeted therapy for esophageal squamous cell
carcinoma. World J Gastroenterol. 21:7648–7658. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 68:7–30. 2019. View Article : Google Scholar
|
|
4
|
Abnet CC, Arnold M and Wei WQ:
Epidemiology of esophageal squamous cell carcinoma.
Gastroenterology. 154:360–373. 2018. View Article : Google Scholar :
|
|
5
|
Wang C, Wang J, Chen Z, Gao Y and He J:
Immunohistochemical prognostic markers of esophageal squamous cell
carcinoma: A systematic review. Chin J Cancer. 36:652017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li J, Qi Z, Hu YP and Wang YX: Possible
biomarkers for predicting lymph node metastasis of esophageal
squamous cell carcinoma: A review. J Int Med Res. 47:544–556. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Salehi S, Taheri MN, Azarpira N, Zare A
and Behzad-Behbahani A1: State of the art technologies to explore
long non-coding RNAs in cancer. J Cell Mol Med. 21:3120–3140. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Wu Z, Fu X and Han W: lncRNAs:
Insights into their function and mechanics in underlying disorders.
Mutat Res Rev Mutat Res. 762:1–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Forrest ME and Khalil AM: Review:
Regulation of the cancer epigenome by long non-coding RNAs. Cancer
Lett. 407:106–112. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flockhart RJ, Webster DE, Qu K,
Mascarenhas N, Kovalski J, Kretz M and Khavari PA: BRAFV600E
remodels the melanocyte transcriptome and induces BANCR to regulate
melanoma cell migration. Genome Res. 22:1006–1014. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Lin X, Fu X, Yan W, Lin F, Kuang
P, Luo Y, Lin E, Hong X and Wu G: Long non-coding RNA BANCR
regulates cancer stem cell markers in papillary thyroid cancer via
the RAF/MEK/ERK signaling pathway. Oncol Rep. 40:859–866.
2018.PubMed/NCBI
|
|
12
|
Shen X, Bai Y, Luo B and Zhou X:
Upregulation of lncRNA BANCR associated with the lymph node
metastasis and poor prognosis in colorectal cancer. Biol Res.
50:322017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou T and Gao Y: Increased expression of
lncRNA BANCR and its prognostic significance in human
hepatocellular carcinoma. World J Surg Oncol. 14:82016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Z, Yang T, Xu Z and Cao X:
Upregulation of the long non-coding RNA BANCR correlates with tumor
progression and poor prognosis in esophageal squamous cell
carcinoma. Biomed Pharmacother. 82:406–412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Q, Zheng Y, Wu B, Chen X, Sun F, Ge P
and Wang P: BANCR regulates the cell invasion and migration in
esophageal squamous cell carcinoma through Wnt/β-catenin signaling
pathway. Onco Targets Ther. 12:9319–9327. 2019. View Article : Google Scholar :
|
|
16
|
Anfossi S, Fu X, Nagvekar R and Calin GA:
MicroRNAs, regulatory messengers inside and outside cancer cells.
Adv Exp Med Biol. 1056:87–108. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Abba ML, Patil N, Leupold JH, Moniuszko M,
Utikal J, Niklinski J and Allgayer H: MicroRNAs as novel targets
and tools in cancer therapy. Cancer Lett. 387:84–94. 2017.
View Article : Google Scholar
|
|
18
|
Li Z, Wu X, Gu L, Shen Q, Luo W, Deng C,
Zhou Q, Chen X, Li Y, Lim Z, et al: Long non-coding RNA ATB
promotes malignancy of esophageal squamous cell carcinoma by
regulating miR-200b/Kindlin-2 axis. Cell Death Dis. 8:e28882017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu H, Deng H, Zhao Y, Li C and Liang Y:
lncRNA XIST/miR-34a axis modulates the cell proliferation and tumor
growth of thyroid cancer through MET-PI3K-AKT signaling. J Exp Clin
Cancer Res. 37:2792018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang L, Wei Z, Wu K, Dai W, Zhang C, Peng
J and He Y: Long noncoding RNA B3GALT5-AS1 suppresses colon cancer
liver metastasis via repressing microRNA-203. Aging (Albany NY).
10:3662–3682. 2018. View Article : Google Scholar
|
|
21
|
Zhang R, Shi H, Ren F, Liu Z, Ji P, Zhang
W and Wang W: Down-regulation of miR-338-3p and Up-regulation of
MACC1 indicated poor prognosis of epithelial ovarian cancer
patients. J Cancer. 10:1385–1392. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Zhang D, Lv J, Wang S and Zhang
Q: lncRNA SNHG15 acts as an oncogene in prostate cancer by
regulating miR-338-3p/FKBP1A axis. Gene. 705:44–50. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luan X and Wang Y: lncRNA XLOC_006390
facilitates cervical cancer tumorigenesis and metastasis as a ceRNA
against miR-331-3p and miR-338-3p. J Gynecol Oncol. 29:e952018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Li Z, Yang G and Pan Z:
MicroRNA-338-3p suppresses tumor growth of esophageal squamous cell
carcinoma in vitro and in vivo. Mol Med Rep. 12:3951–3957. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sharmila G, Bhat FA, Arunkumar R, Elumalai
P, Raja Singh P, Senthilkumar K and Arunakaran J: Chemopreventive
effect of quercetin, a natural dietary flavonoid on prostate cancer
in in vivo model. Clin Nutr. 33:718–726. 2014. View Article : Google Scholar
|
|
26
|
Ma W, Zhang T, Pan J, Shi N, Fan Q, Wang L
and Lu SH: Assessment of insulin-like growth factor 1 receptor as
an oncogene in esophageal squamous cell carcinoma and its potential
implication in chemotherapy. Oncol Rep. 32:1601–1609. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kong KL, Kwong DL, Chan TH, Law SY, Chen
L, Li Y, Qin YR and Guan XY: MicroRNA-375 inhibits tumour growth
and metastasis in oesophageal squamous cell carcinoma through
repressing insulin-like growth factor 1 receptor. Gut. 61:33–42.
2012. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Li Y, Chen D, Gao X, Li X and Shi G:
lncRNA NEAT1 regulates cell viability and invasion in esophageal
squamous cell carcinoma through the miR-129/CTBP2 axis. Dis
Markers. 2017:53146492017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen M, Liu P, Chen Y, Chen Z, Shen M, Liu
X, Li X, Li A, Lin Y, Yang R, et al: Long noncoding RNA FAM201A
mediates the radiosensitivity of esophageal squamous cell cancer by
regulating ATM and mTOR expression via miR-101. Front Genet.
9:6112018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chang ZW, Jia YX, Zhang WJ, Song LJ, Gao
M, Li MJ, Zhao RH, Li J, Zhong YL, Sun QZ, et al:
lncRNA-TUSC7/miR-224 affected chemotherapy resistance of esophageal
squamous cell carcinoma by competitively regulating DESC1. J Exp
Clin Cancer Res. 37:562018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu Y, Hu L, Liang Y, Li J, Wang K, Chen X,
Meng H, Guan X, Yang K and Bai Y: Up-regulation of lncRNA CASC9
promotes esophageal squamous cell carcinoma growth by negatively
regulating PDCD4 expression through EZH2. Mol Cancer. 16:1502017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mei LL, Qiu YT, Huang MB, Wang WJ, Bai J
and Shi ZZ: miR-99a suppresses proliferation, migration and
invasion of esophageal squamous cell carcinoma cells through
inhibiting the IGF1R signaling pathway. Cancer Biomark. 20:527–537.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ke K, Sun Z and Wang Z: Downregulation of
long non-coding RNA GAS5 promotes cell proliferation, migration and
invasion in esophageal squamous cell carcinoma. Oncol Lett.
16:1801–1808. 2018.PubMed/NCBI
|
|
35
|
Zhang Q, Zhao X, Zhang C, Wang W, Li F,
Liu D, Wu K, Zhu D, Liu S, Shen C, et al: Overexpressed PKMYT1
promotes tumor progression and associates with poor survival in
esophageal squamous cell carcinoma. Cancer Manag Res. 11:7813–7824.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bertrand FE, Steelman LS, Chappell WH,
Abrams SL, Shelton JG, White ER, Ludwig DL and McCubrey JA: Synergy
between an IGF-1R antibody and Raf/MEK/ERK and PI3K/Akt/mTOR
pathway inhibitors in suppressing IGF-1R-mediated growth in
hematopoietic cells. Leukemia. 20:1254–1260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo L, Bai Y, Ji S and Ma H: MicroRNA98
suppresses cell growth and invasion of retinoblastoma via targeting
the IGF1R/k-Ras/Raf/MEK/ERK signaling pathway. Int J Oncol.
54:807–820. 2019.PubMed/NCBI
|
|
38
|
Sundaram GM and Veera Bramhachari P:
Molecular interplay of pro-inflammatory transcription factors and
non-coding RNAs in esophageal squamous cell carcinoma. Tumour Biol.
39:10104283177057602017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tan D, Wu Y, Hu L, He P, Xiong G, Bai Y
and Yang K: Long noncoding RNA H19 is up-regulated in esophageal
squamous cell carcinoma and promotes cell proliferation and
metastasis. Dis Esophagus. 30:1–9. 2017.
|
|
40
|
Bao J, Zhou C, Zhang J, Mo J, Ye Q, He J
and Diao J: Upregulation of the long noncoding RNA FOXD2-AS1
predicts poor prognosis in esophageal squamous cell carcinoma.
Cancer Biomark. 21:527–533. 2018. View Article : Google Scholar
|
|
41
|
Sadeghpour S and Ghorbian S: Evaluation of
the potential clinical prognostic value of lncRNA-BANCR gene in
esophageal squamous cell carcinoma. Mol Biol Rep. 46:991–995. 2019.
View Article : Google Scholar
|
|
42
|
He A, Liu Y, Chen Z, Li J, Chen M, Liu L,
Liao X, Lv Z, Zhan Y, Zhuang C, et al: Over-expression of long
noncoding RNA BANCR inhibits malignant phenotypes of human bladder
cancer. J Exp Clin Cancer Res. 35:1252016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang L and Liu G: lncRNA BANCR suppresses
cell viability and invasion and promotes apoptosis in
non-small-cell lung cancer cells in vitro and in vivo. Cancer Manag
Res. 11:3565–3574. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gao H, Gong N, Ma Z, Miao X, Chen J, Cao Y
and Zhang G: lncRNA ZEB2-AS1 promotes pancreatic cancer cell growth
and invasion through regulating the miR-204/HMGB1 axis. Int J Biol
Macromol. 116:545–551. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Guo H, Yang S, Li S, Yan M, Li L and Zhang
H: lncRNA SNHG20 promotes cell proliferation and invasion via
miR-140-5p-ADAM10 axis in cervical cancer. Biomed Pharmacother.
102:749–757. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Heery R, Finn SP, Cuffe S and Gray SG:
Long non-coding RNAs: Key regulators of epithelial-mesenchymal
transition, tumour drug resistance and cancer stem cells. Cancers
(Basel). 9. pp. 382017, View Article : Google Scholar
|
|
48
|
Cao R, Shao J, Hu Y, Wang L, Li Z, Sun G
and Gao X: MicroRNA-338-3p inhibits proliferation, migration,
invasion, and EMT in osteosarcoma cells by targeting activator of
90 kDa heat shock protein ATPase homolog 1. Cancer Cell Int.
18:492018. View Article : Google Scholar
|
|
49
|
Ding Z, Zhu J, Zeng Y, Du W, Zhang Y, Tang
H, Zheng Y, Qin H, Liu Z and Huang JA: The regulation of Neuropilin
1 expression by miR-338-3p promotes non-small cell lung cancer via
changes in EGFR signaling. Mol Carcinog. 58:1019–1032. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu S, Suo J, Wang C, Sun X, Wang D, He L,
Zhang Y and Li W: Downregulation of tissue miR-338-3p predicts
unfavorable prognosis of gastric cancer. Cancer Biomark.
21:117–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu F, Yang X, Geng M and Huang M:
Targeting ERK, an Achilles' Heel of the MAPK pathway, in cancer
therapy. Acta Pharm Sin B. 8:552–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pollak MN, Schernhammer ES and Hankinson
SE: Insulin-like growth factors and neoplasia. Nat Rev Cancer.
4:505–518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen J, Deng T, Li X and Cai W: miR-193b
inhibits the growth and metastasis of renal cell carcinoma by
targeting IGF1R. Artif Cells Nanomed Biotechnol. 47:2058–2064.
2019. View Article : Google Scholar : PubMed/NCBI
|