Introduction
Colorectal cancer (CRC) was the third most common
cancer and the third most common cause of cancer-associated
mortality in 2019 in the USA (1).
Patients with stage I and II CRC who undergo colectomy, proctectomy
or proctocolectomy often receive neoadjuvant radiation or
chemotherapy treatments to lower the risk of recurrence, whereas
patients with stage III and IV CRC are typically treated with
chemo-therapy or radiotherapy (2). However, studies have suggested that
drug resistance developed by patients with CRC limits the
therapeutic efficacy of anticancer agents (3,4).
Therefore, developing novel effective chemotherapy drugs will
benefit patients with CRC.
Centella asiatica, a herb of the apiaceae
family (5), contains a variety of
active ingredients, such as triterpenoids and saponins (6). AC and madecassoside are the
principal monomer components of saponins in C. asiatica
(7). Previous studies have
revealed that AC possesses a wide range of biological functions,
such as facilitating angiogenesis by increasing collagen synthesis
(8,9), inhibiting inflammatory process
(10,11), suppressing scar formation
(12,13) and promoting wound healing
(12,13). Additionally, AC has a crucial
function in inflammatory lung diseases (14), neurological disorders (15) and osteogenic differentiation of
human periodontal ligament cells (16). The clinical applicability of AC in
the treatment of malignant tumors has been extensively studied over
the last few years. Studies have confirmed that AC has a particular
therapeutic effect on breast cancer (17), multiple myeloma and other tumors
(18), and can improve the
sensitivity of cancer cells to chemotherapy drugs (18,19). However, to the best of our
knowledge, the biological function of AC in CRC cells remains
largely unknown.
AC reduces the incidence of DMBA-induced breast
cancer in rats by inhibiting expression of TNF-α and IL-1β
(20). AC treatment considerably
suppresses the proliferation of human breast cancer MCF-7 cells,
and induces cell apoptosis in vitro (17). A previous study on multiple
myeloma cancer cells indicated that AC inhibits cellular
proliferation by inducing autophagy coupled with elevated
expression of LC3-II (18).
Notably, AC increases sensitivity of tumor cells to vincristine by
promoting apoptosis and inducing cell cycle arrest (19).
Therefore, the aim of the present study was to
improve understanding of the biological effects of AC on CRC cells.
Using several functional experiments in vitro and in mice
models in vivo, the present findings revealed that AC could
inhibit activation of the NF-κB signaling pathway by suppressing
IκBα phosphorylation, which may be one of the primary mechanisms of
AC-induced CRC cell apoptosis and cell cycle arrest. Overall, the
present data suggest that AC exhibits antitumor effects in CRC.
Materials and methods
Cell culture and solution
preparation
Normal human intestinal FHC cells and human CRC cell
lines (HCT116, SW480 and LoVo) were obtained from the Cell Bank of
Shanghai Institutes for Biological Sciences, Chinese Academy of
Sciences. The cells were cultured in Dulbecco's modified Eagle's
medium (Hyclone; GE Healthcare Life Sciences) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.) in
an incubator at 37°C, containing 5% CO2. Culture medium
was changed every 24 h.
A total of 95.90 mg AC (Santa Cruz Biotechnology,
Inc.) was weighed and dissolved in 100 µl dimethyl sulfoxide
(DMSO; Santa Cruz Biotechnology, Inc.) to prepare a 1 mM storage
solution. Subsequently, different concentrations of the working
solutions (0.1, 0.5 and 2 µM) were prepared using culture
medium. Cells at the logarithmic growth phase were treated with the
working solutions. A quantity of 0.2% DMSO was used as the
control.
Cell Counting Kit-8 (CCK-8) assay
Cell viability was assessed using a CCK-8 assay. CRC
cells and FHC cells in the logarithmic growth phase were digested
and seeded into 96-well plates at a density of 5×103
cells/well followed by treatment with various concentrations of AC
(0.1, 0.5 and 2 µM) at 37°C. Cells treated with 0.3% DMSO
were used as the negative controls. Subsequently, 10% CCK-8 reagent
(Beijing Zoman Biotechnology Co., Ltd.) was added to each well at
different time intervals (24, 48 and 72 h), and cells incubated for
2 h at 37°C. The absorbance was then measured using a microplate
reader (Bio-Rad Laboratories, Inc.) at a wavelength of 450 nm.
Colony formation assay
CRC cells in the logarithmic growth phase were
seeded in 6-well plates (1×103 cells/well) and then
treated with different concentrations of AC (0.1, 0.5 and 2
µM) at 37°C. After 7 days, the cell colonies were fixed with
4% paraformaldehyde solution (Beijing Solarbio Biotechnology Co.,
Ltd.) for 15 min at room temperature. Cells were rinsed three times
with PBS and then stained with 0.1% crystal violet solution
(Beijing Solarbio Biotechnology Co., Ltd.) according to the
manufacturer's instructions. The number of colonies and the number
of cells within the colonies were counted in five randomly selected
fields under an optical microscope (magnification, ×40).
Cell cycle analysis
CRC cells in the logarithmic growth phase were
seeded into 6-well plates (5×105 cells/well), and then
incubated with various concentrations of AC (0.1, 0.5 and 2
µM) for 24 and 72 h. The cells were collected, digested with
0.25% trypsin (Gibco; Thermo Fisher Scientific, Inc.) and
centrifuged at 200 × g for 4 min at 4°C. Subsequently, the cells
were fixed with 70% pre-cooled ethanol (prepared in PBS) overnight
at 4°C, followed by centrifugation at 700 × g for 4 min at 4°C.
Cells were rinsed with 1 ml pre-cooled PBS and resuspended in
pre-cooled PBS at a density of 1×106 cells/ml. Next, 100
µl cell suspension was treated with 10 µg/ml RNase A.
Cells were incubated with 20 µg/ml propidium iodide (PI)
solution at room temperature for 10 min and then centrifuged at 700
× g for 4 min at 4°C. The cells were then rinsed with pre-chilled
PBS and resuspended in pre-chilled PBS. Flow cytometry was
performed to assess the cell cycle using a CytoFLEX flow cytometer
(Beckman Coulter, Inc.). Data were analyzed using FlowJo (version
7.6.1; FlowJo LLC).
Cell apoptosis analysis
Cell apoptosis analysis was performed using the
Annexin V-Alexa Fluor 488/PI kit (cat. no. CA1040, Beijing Solarbio
Biotechnology Co., Ltd.) according to the standard procedures. CRC
cells were treated with various concentrations of AC (0.1, 0.5 and
2 µM) for 24 and 72 h. Cells were collected, rinsed twice in
PBS and then resuspended in a binding buffer. The cells were
subsequently stained with Annexin V-Alexa Fluor 488/PI for 5 min at
room temperature following the manufacturer's instructions, and
cell apoptosis detected using a CytoFLEX flow cytometer (Beckman
Coulter, Inc.). Data analysis was performed using FlowJo (version
7.6.1; FlowJo LLC).
Analysis of mitochondrial membrane
potential
Mitochondrial Membrane Potential assay kit with JC-1
(cat. no. M8650; Beijing Solarbio Biotechnology Co., Ltd.) was used
to evaluate the mitochondrial membrane potential of CRC cells
following treatment with AC. CRC cells in the logarithmic growth
phase were seeded into 6-well plates (5×105 cells/well),
and then treated with various concentrations of AC (0.1, 0.5 and 2
µM) for 72 h. The cells were harvested, rinsed twice with
PBS and incubated with 1.2 µg/ml phenol red-free medium
containing JC-1 probes for 30 min at 37°C. Subsequently, the cells
were collected, rinsed and resuspended in PBS. Fluorescence of the
cells was measured using a CytoFLEX flow cytometer (Beckman
Coulter, Inc.) at wavelengths of 590 and 530 nm. Data were analyzed
using FlowJo (version 7.6.1; FlowJo LLC).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
TRIzol reagent (Thermo Fisher Scientific, Inc.) was
used to isolate the total RNA from CRC cells treated with AC. The
concentration and purity of RNA was detected with a UV
spectrophotometer (A260/A280). A ratio of A260/A280 between 1.8 and
2.0 was regarded as a sufficient quality. A quantity of 500 ng
total RNA was reverse transcribed into cDNA using a PrimeScript™ RT
Master mix (cat. no. RR036A; Takara Bio, Inc.). The RT conditions
were 37°C for 15 min, followed by 85°C for 5 sec, and storage at
4°C. qPCR was performed on a LightCycler® 96 system
(Roche Diagnostics) using One Step TB Green®
PrimeScript™ RT-PCR kit (cat. no. RR066A; Takara Bio, Inc.). The
thermocycling conditions were as follows: Pre-denaturation at 95°C
for 2 min; 40 cycles of denaturation at 95°C for 30 sec, annealing
at 60°C for 30 sec, and extension at 72°C for 30 sec. The relative
expression levels of genes were calculated using the
2−ΔΔCq method (21),
and β-actin was used as an internal control. The experiment was
performed in triplicate. All primer sequences used in the present
study are listed in Table I.
 | Table IPrimers for reverse
transcription-quantitative PCR. |
Table I
Primers for reverse
transcription-quantitative PCR.
| Gene | Forward
sequence | Reverse
sequence |
|---|
| β-actin |
5′-CCTCGCCTTTGCCGATCC-3′ |
5′-GGATCTTCATGAGGTAGTCAGTC-3′ |
| IκBα |
5′-CTCCGAGACTTTCGAGGAAATAC-3′ |
5′-GCCATTGTAGTTGGTAGCCTTCA-3′ |
| P65 |
5′-ATGTGGAGATCATTGAGCAGC-3′ |
5′-CCTGGTCCTGTGTAGCCATT-3′ |
| Caspase-9 |
5′-CTCAGACCAGAGATTCGCAAAC-3′ |
5′-GCATTTCCCCTCAAACTCTCAA-3′ |
| Caspase-3 |
5′-AGAGGGGATCGTTGTAGAAGTC-3′ |
5′-ACAGTCCAGTTCTGTACCACG-3′ |
| P21 |
5′-TGTCCGTCAGAACCCATGC-3′ |
5′-AAAGTCGAAGTTCCATCGCTC-3′ |
| Bax |
5′-CCCAGAGTTTGAGCCGAGTG-3′ |
5′-CCCATCCCTTCGTCGTCCT-3′ |
| Cyclin D1 |
5′-CAATGACCCCGCACGATTTC-3′ |
5′-CATGGAGGGCGGATTGGAA-3′ |
| CDK4 |
5′-CTGGTGTTTGAGCATGTAGACC-3′ |
5′-GATCCTTGATCGTTTCGGCTG-3′ |
| Bcl-2 |
5′-CCAGCGTATATCGGAATGTGG-3′ |
5′-CCATGTGATACCTGCTGAGAAG-3′ |
| P53 |
5′-GTTTCCGTCTGGGCTTCTTG-3′ |
5′-CACAACCTCCGTCATGTGCT-3′ |
Western blotting
Total protein was extracted from CRC cells or tumor
tissues using RIPA lysis buffer (Beyotime Institute of
Biotechnology) containing 1% protease inhibitor (Beyotime Institute
of Biotechnology). Protein concentration was deter-mined using a
BCA protein assay kit (Nanjing KeyGEN Biotech, Co., Ltd.) according
to the manufacturer's protocol. Subsequently, proteins (30
µg per lane) were separated by 10% SDS-PAGE (Nanjing KeyGEN
Biotech, Co., Ltd.) and then transferred onto polyvinylidene
fluoride membranes (EMD Millipore). The membranes were blocked with
5% skimmed milk (prepared in TBST containing 0.1% Tween-20) for 2 h
at room temperature and incubated with corresponding primary
antibodies overnight at 4°C. The following primary antibodies were
used for incubation: Anti-NF-kB P65 (1:1,000; cat. no. ab16502;
Abcam), anti-phosphorylated (p)-P65 (1:1,500; cat. no. ab86299;
Abcam), anti-caspase-3 (1:1,000; cat. no. ab4051, Abcam),
anti-cleaved caspase-3 (1:1,000; cat. no. ab32042; Abcam),
anti-caspase-9 (1:1,000; cat. no. ab32539; Abcam), anti-cleaved
caspase-9 (1:1,000; cat. no. ab32539; Abcam), anti-Bcl-2 (1:1,000;
cat. no. ab32124; Abcam), anti-Bax (1:1,000; cat. no. ab32503;
Abcam), anti-CDK4 (1:1,000; cat. no. ab199728, Abcam), anti-Cyclin
D1 (1:2,000; cat. no. ab205718; Abcam), anti-IκBα (1:1,000; cat.
no. 9242; Cell Signaling Technology, Inc.), anti-p-IκBα (1:1,000;
cat. no. 2859; Cell Signaling Technology, Inc.), anti-Histone H3
(1:3,000; cat. no. ab1791; Abcam) and anti-β-actin (1:5,000; cat.
no. ab8227; Abcam). Subsequently, the membranes were incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG
H&L (1:5,000; cat. no. ab205718; Abcam) or HRP-conjugated goat
anti-mouse IgG H&L (1:5,000; cat. no. ab205719; Abcam)
secondary antibodies for 2 h at room temperature. The
immunoreactive proteins were then visualized using immobilon
western chemiluminescent HRP substrate (EMD Millipore). The western
blots were analyzed with Image Lab 3.0 software (Bio-Rad
Laboratories, Inc.).
Nuclear/cytosol fractionation kit
assay
Nuclear and cytosolic fractions were prepared using
the nuclear/cytosol extraction kit (cat. no. K266-25; BioVision,
Inc.) according to the manufacturer's instructions. All buffers
were stored in ice throughout the experimental period. All
centrifugation steps were performed at 4°C. CRC cells were treated
with AC (2 µM) for 72 h at 37°C, harvested and rinsed twice
with pre-cooled PBS. Cells were subsequently centrifuged at 600 × g
for 5 min. A total of 1×106 cells were treated with 200
µl pre-chilled CEB-A mix (1 ml CEB-A mix containing 1
µl DTT and 2 µl protease inhibitor), vortexed
vigorously for 15 sec to fully resuspend the cell pellets and
incubated on ice for 10 min. Subsequently, 11 µl pre-cooled
cytosol extraction buffer-B was added, vortexed vigorously for 5
sec, and incubated on ice for 1 min. The tube was continuously
vortexed vigorously for 5 sec, followed by centrifugation at 14,000
× g for 5 min. The supernatant (cytoplasmic extract) was
immediately transferred to a clean pre-cooled tube and the tube was
placed on ice.
Next, 100 µl ice-cold nuclear extraction
buffer mix (1 ml NEB containing 2 µl protease inhibitor
cocktail and 1 µl DTT) was added to the cell pellets
(nuclei), and then vortexed vigorously for 10 sec. The cell pellets
were placed on ice for 40 min, vortexed for 15 sec and the samples
placed on ice after 10 min. Finally, following centrifugation at
16,000 × g for 5 min, the supernatant (nuclear extract) was
immediately transferred into a pre-cooled clean tube and stored at
-80°C. The cytosol and nuclear extracts were then used for western
blotting to measure P65 expression level.
Animal experiments
A total of 18 5-week-old female BALB/c-nude mice
(weight, 20-25 g) were purchased from the Model Animal Research
Center of Nanjing University and maintained according to
regulations of the Animal Care Committee of The First Affiliated
Hospital of Xiamen University. The housing conditions of mice were
as follows: The temperature was 22-24°C, humidity was 45-60%,
ventilation was 15 times/h, and 12 h of light and 12 h of dark.
Mice were provided food and water ad libitum. In the present
study, mice were randomly divided into three groups (n=6 per
group): i) Control group; ii) 5 mg/kg AC treatment group; and iii)
10 mg/kg AC treatment group. HCT116 cells (5×106) were
subcutaneously injected into the right hind legs of the mice.
Subsequently, 1 week later, 5 or 10 mg/kg AC or PBS (10 mg/kg) was
administered to mice by oral gavage every 2 days for 6 consecutive
weeks. Tumor diameters were measured using calipers every week.
Mice were euthanized at 42 days after the first administration of
AC or PBS by cervical dislocation, and the tumors were carefully
excised and weighed. All mice were in good health throughout the
experimental period. All in vivo experiments were performed
at the Animal Center of The First Affiliated Hospital of Xiamen
University (Xiamen, China) according to the guidelines of the
National Institutes of Health and approved by the Animal Care and
Use Committee of The First Affiliated Hospital of Xiamen University
(approval no. XMU-AEA-20180137).
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 6.0 software (GraphPad Software, Inc.) and SPSS software
version 21.0 (IBM Corp.). Comparisons between groups were performed
using one-way analysis of variance with Bonferroni's post hoc
analysis. Data from three independent experiments are expressed as
the mean ± standard deviation. P<0.05 was considered to indicate
a statistically significant difference.
Results
AC inhibits CRC cell proliferation and
induces CRC cell cycle arrest at the G0/G1 phase
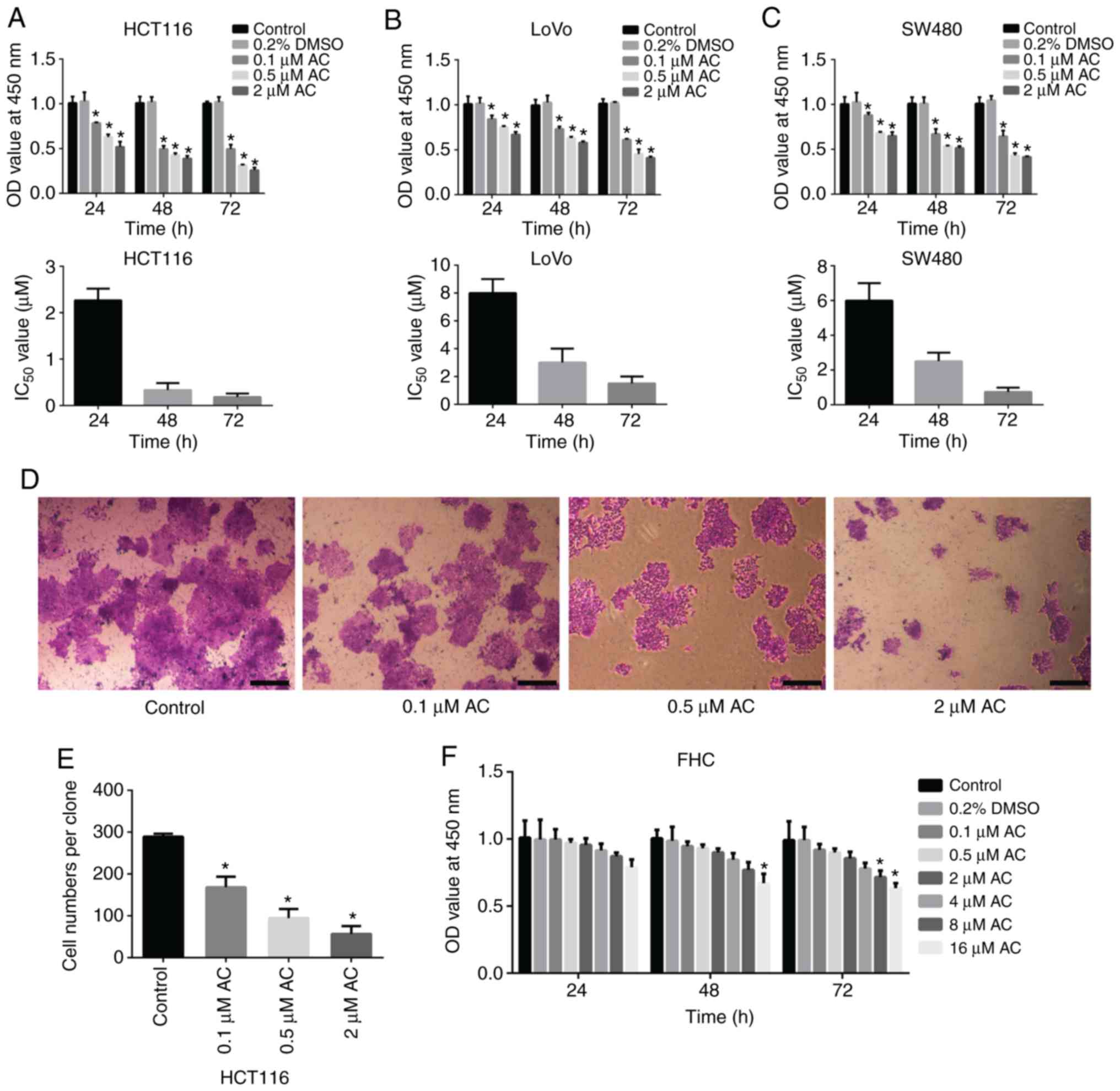
CCK-8 assay and colony formation assay were
performed to determine the effect of AC on the viability of CRC
cells. Results of the CCK-8 assay demonstrated that AC
significantly reduced the viability of HCT116, SW480, and LoVo
cells in a time and dose-dependent manner (Fig. 1A-C), whereas AC had no significant
effects on normal human intestinal FHC cells at a given range of
concentrations (0.1-8 µM) (Fig. 1F). However, the viability of FHC
cells was significantly reduced after treating with 16 µM AC
for 72 h, indicating that high concentration of AC has an adverse
effect on human normal epithelial cells. Additionally, HCT116
cells, with the lowest IC50 value, were selected for
subsequent analyses. Colony formation assay revealed that AC
significantly inhibited the colony formation of HCT116 cells in a
dose-dependent manner (Fig. 1D and
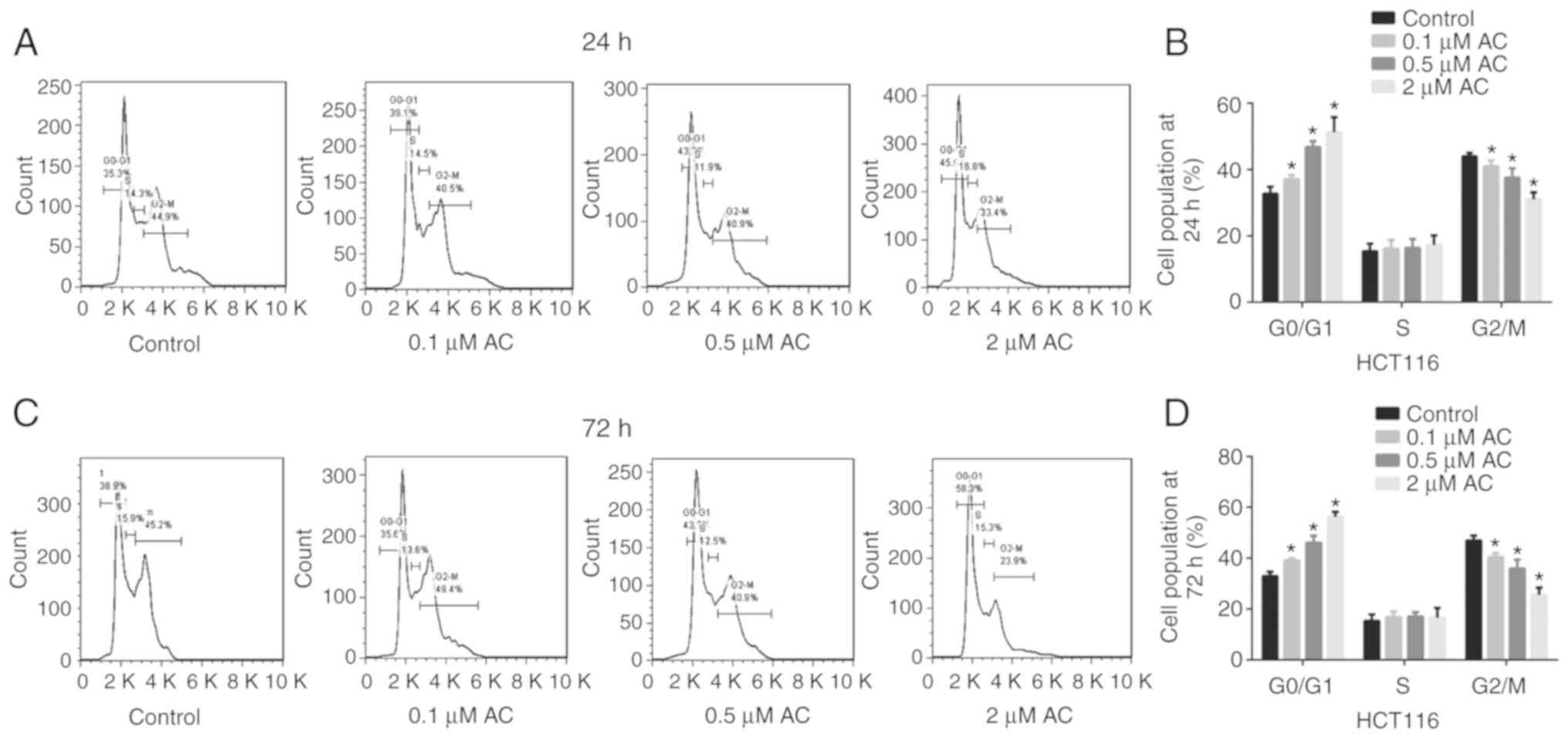
E). Furthermore, flow cytometry analysis indicated that AC
treatment for 24 or 72 h significantly induced CRC cell cycle
arrest at the G0/G1 phase. The percentage of cells in the G2/M
phase significantly decreased following treatment with AC for 24 or
72 h and no difference was observed for the percentage of cells in
the S phase (Fig. 2).
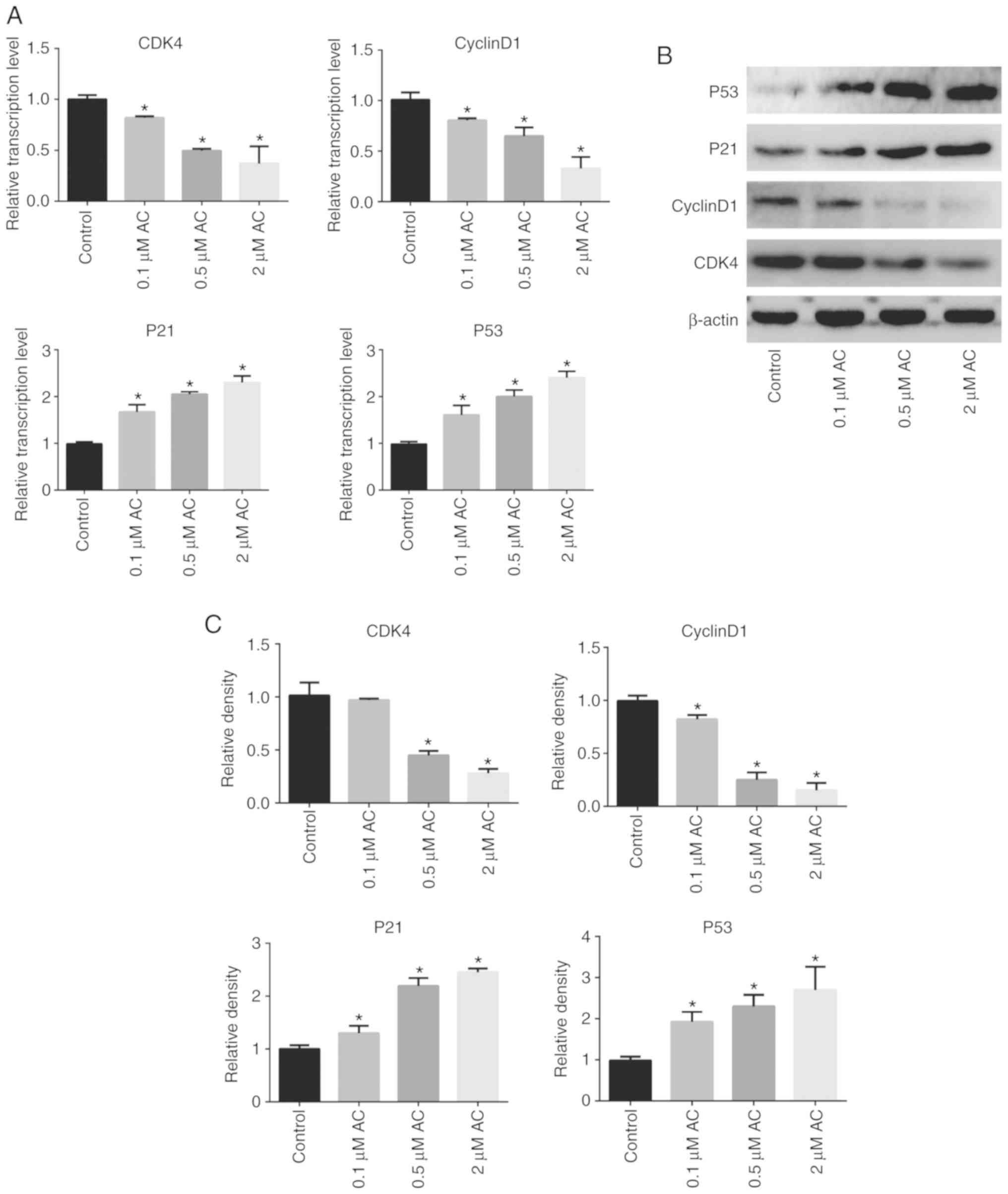
AC suppresses the expression levels of
cell cycle-associated genes
RT-qPCR and western blotting assays were performed
to further understand the specific mechanism of the effects of AC
on the CRC cell cycle. The findings revealed that AC significantly
decreased the expression levels of CDK4 and Cyclin D1, whereas the
expression levels of P53 and P21 were significantly higher in
AC-treated cells compared with the control cells (Fig. 3).
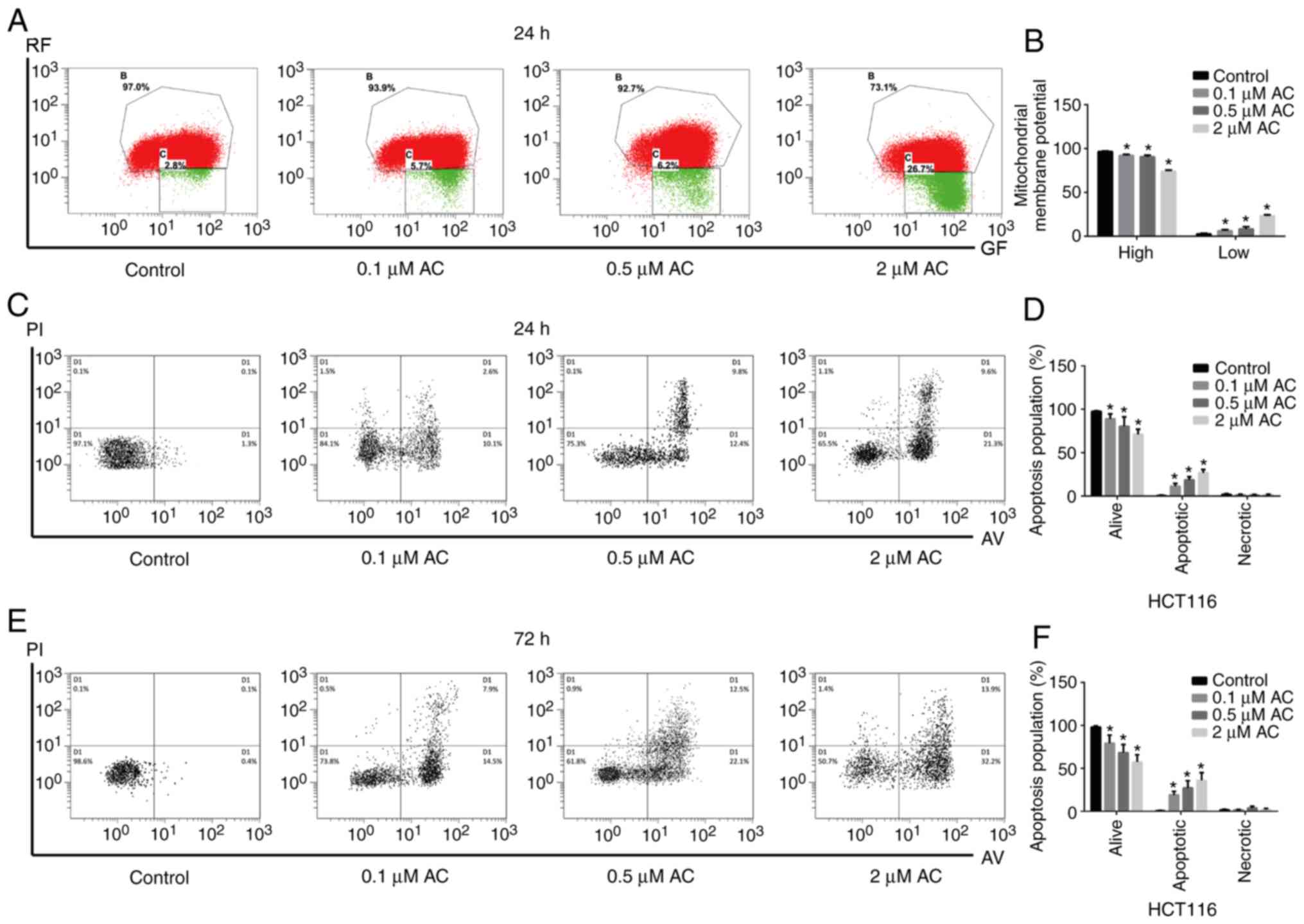
AC decreases mitochondrial membrane
potential and promotes CRC cell apoptosis
In the present study, JC-1 probes were used to
measure mitochondrial membrane potential, and cell apoptosis was
detected using flow cytometry. JC-1, a cyanine dye, promotes
discrimination of energized and deenergized mitochondria, where the
normally green fluorescent dye forms red fluorescent aggregates
when concentrated in energized mitochondria (22). However, when cell apoptosis
occurs, the mitochondrial transmembrane potential is decreased,
JC-1 is released from the mitochondria, decreasing in concentration
and maintaining the monomeric form with green fluorescence.
Therefore, it is widely used to determine the apoptosis of the cell
by detecting green (low) and red (high) fluorescence. The present
results indicated that AC significantly decreased the mitochondrial
membrane potential of HCT116 cells in a dose-dependent manner
(Fig. 4A and B). Furthermore, AC
significantly increased the apoptosis of CRC cells compared with
the untreated cells (Fig.
4C-F).
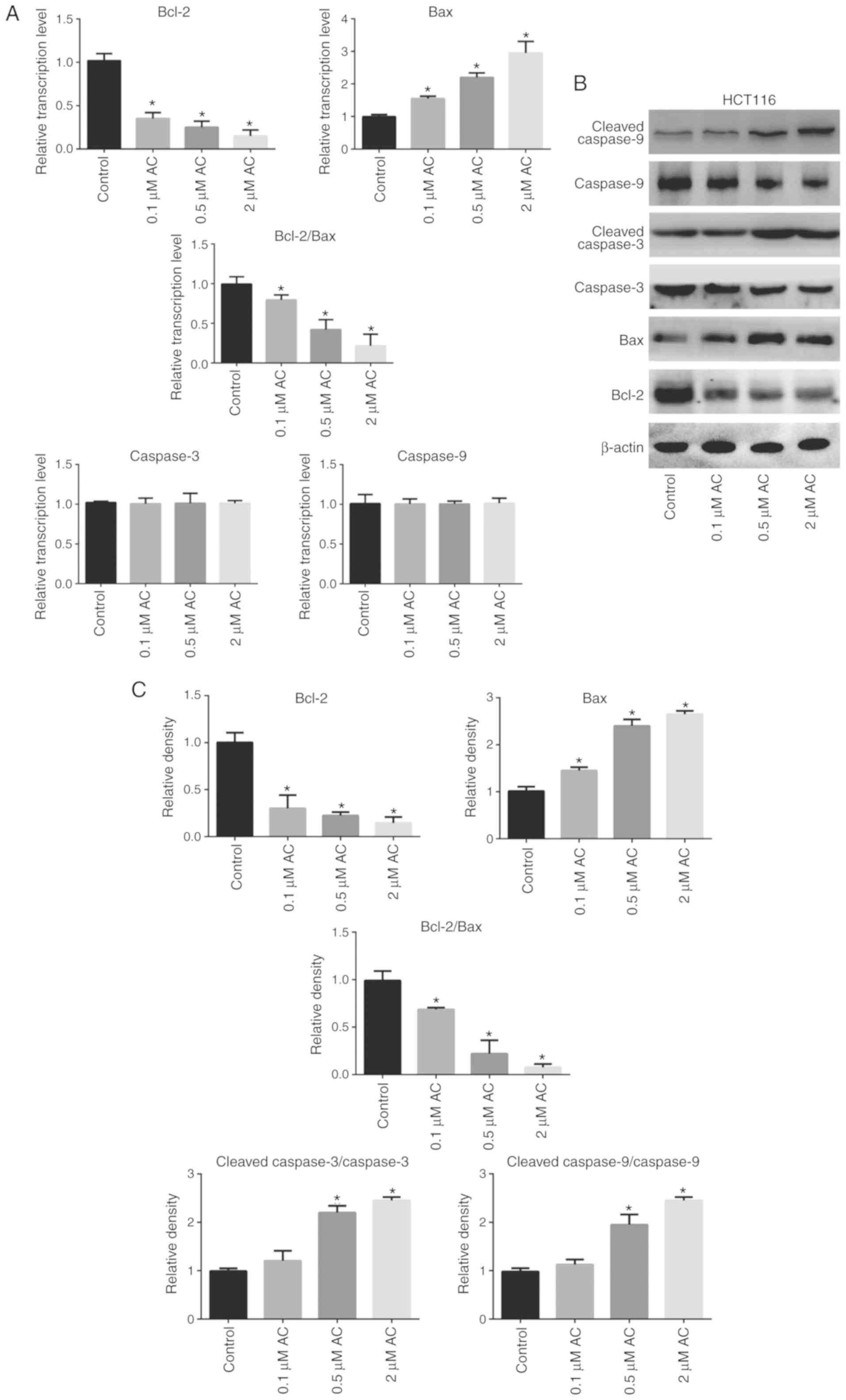
AC regulates the expression of
apoptosis-related genes in CRC cells
RT-qPCR and western blotting were performed to
further elucidate the biological function of AC in CRC cells
(Fig. 5). The results
demonstrated that AC significantly increased the activation of
caspase-9 and caspase-3 in HCT116 cells in a dose-dependent manner
(Fig. 5C), but had no effect on
the mRNA levels of caspase-9 and caspase-3 (Fig. 5A). In addition, AC significantly
downregulated Bcl-2 expression and upregulated Bax expression at
both the mRNA and protein levels (Fig. 5A-C. The ratio of Bcl-2/Bax protein
expression was significantly decreased following AC treatment
compared with the control cells (Fig.
5C).
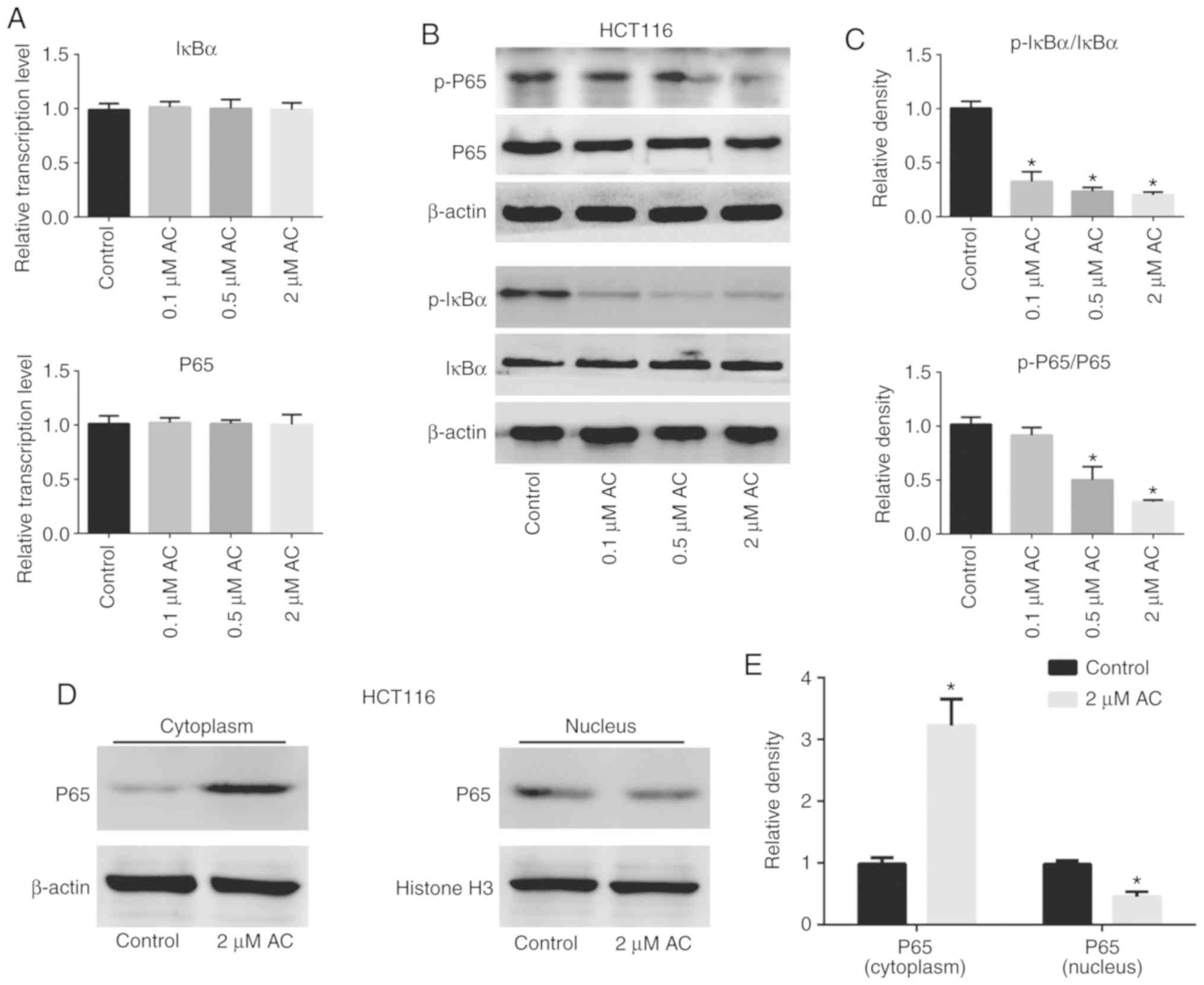
AC suppresses phosphorylation of IκBα and
NF-κB p65, inhibiting the NF-κB signaling pathway
The NF-κB signaling pathway is crucial for the
regulation of various biological activities of eukaryotic cells,
such as cell proliferation, survival and apoptosis (23,24). Inhibitors of the NF-κB signaling
pathway are potential anticancer drug candidates (25). The present study assessed the
activation and expression of molecules associated with the NF-κB
pathway in HCT116 cells treated with AC using RT-qPCR and western
blotting (Fig. 6). The results
revealed that treatment of HCT116 cells with AC significantly
decreased phosphorylation of IκBα and P65 (Fig. 6B and C), but had no effect on the
mRNA levels of total IκBα and P65 compared with the control cells
(Fig. 6A). Furthermore, AC
significantly increased the content of P65 in the nucleus, but
decreased P65 content in the cytoplasm, suggesting that AC
treatment inhibited nuclear translocation of P65 (Fig. 6D and E).
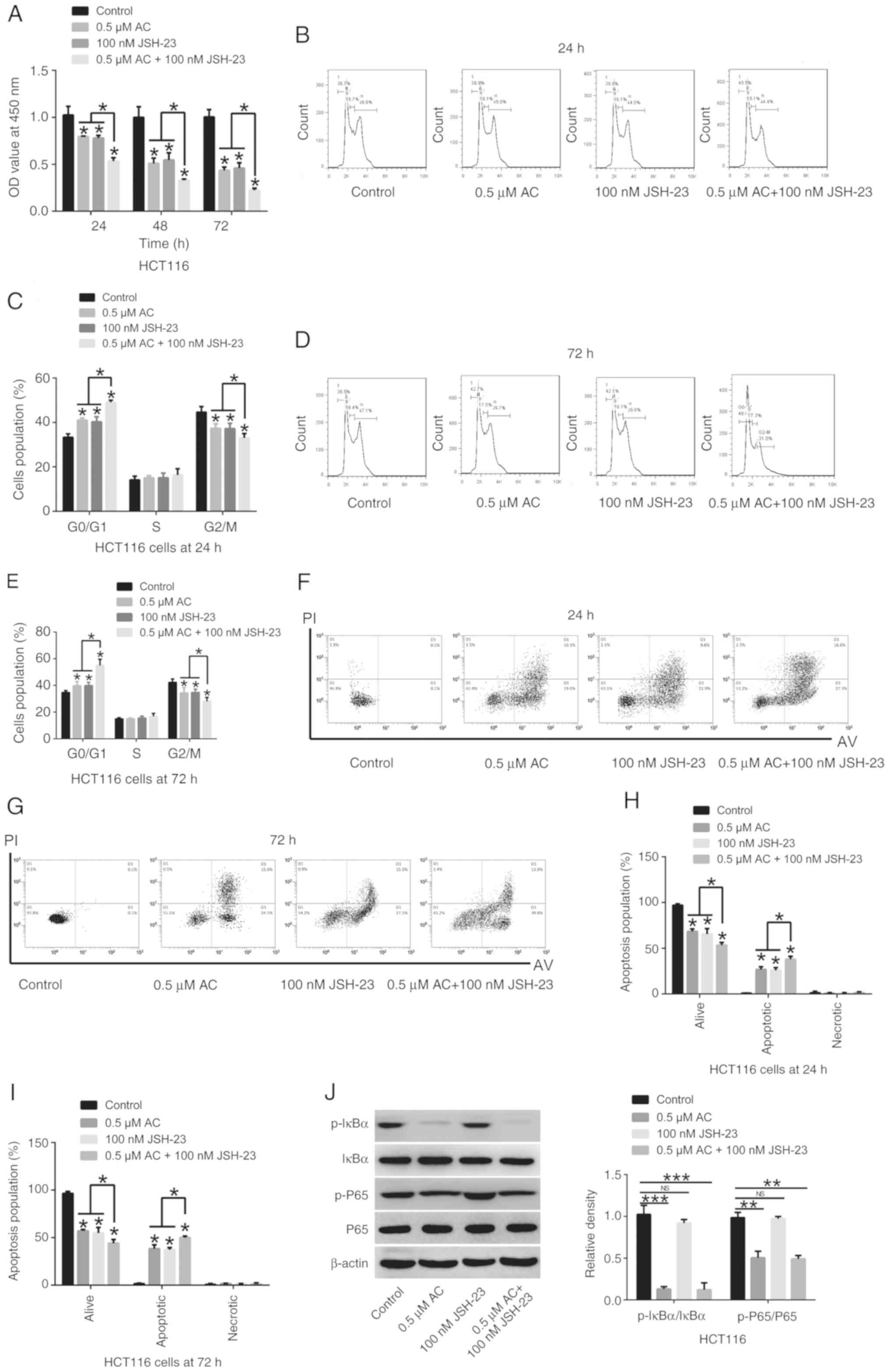
Synergistic inhibitory effects of AC and
the NF-κB signaling pathway inhibitor JSH-23 on CRC cells
Administration of 0.5 µM AC and the NF-κB
signaling pathway inhibitor JSH-23 (100 nM) significantly
suppressed CRC cell viability, significantly induced cell cycle
arrest at the G0/G1 phase, significantly promoted cell apoptosis
and significantly deceased the phosphorylation of IκBα and P65 in
HCT116 cells in comparison to HCT 116 cells treated either with AC
or JSH-23 (Fig. 7). A synergistic
inhibitory effect of AC and JSH-23 on CRC cells was observed,
indicating that AC may be a promising adjuvant agent in treatment
of CRC.
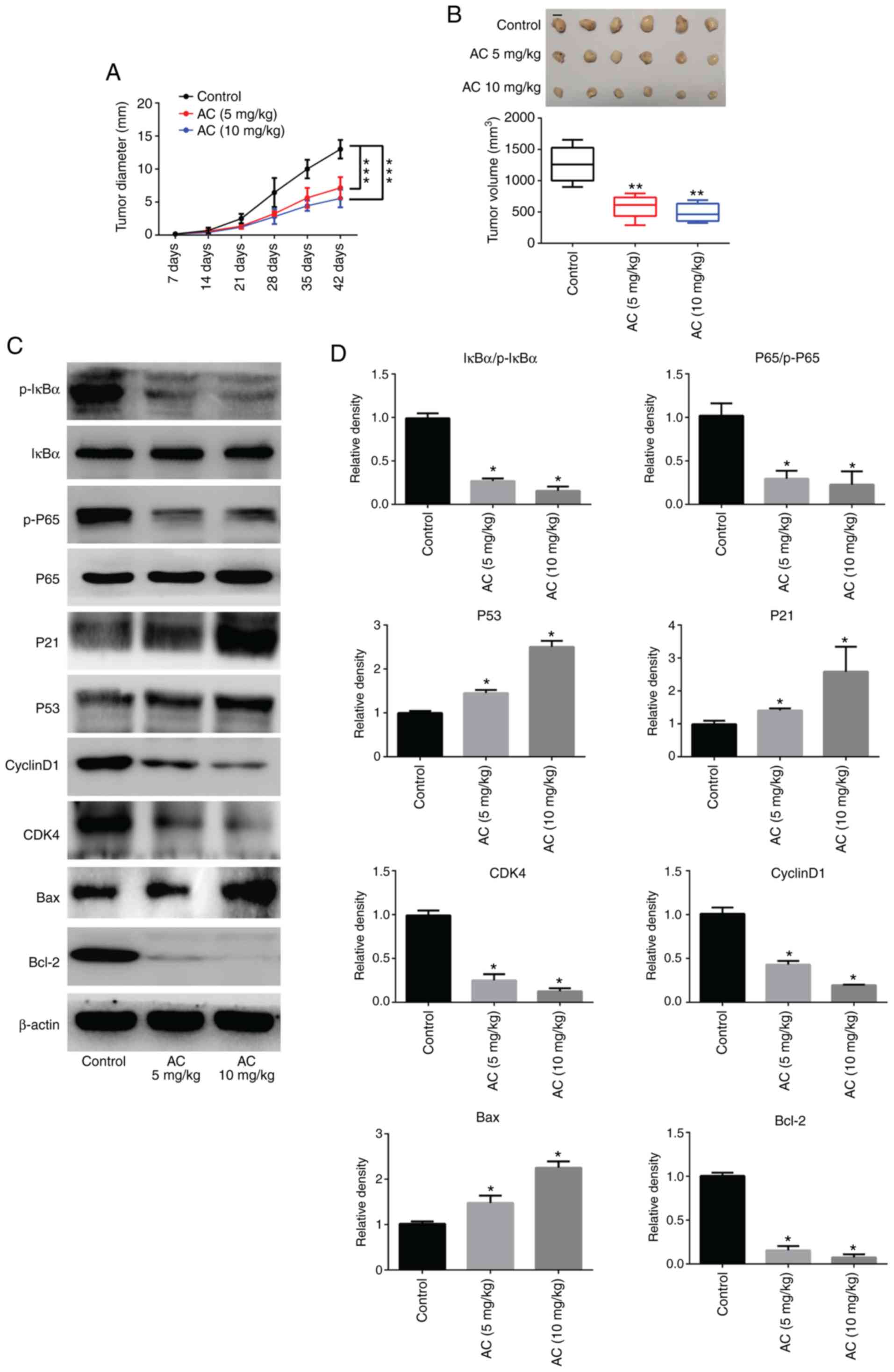
AC inhibits CRC tumor cell growth in
vivo
The present findings revealed that administration of
AC in mice with tumors notably inhibited tumor growth; tumor
diameters and tumor volumes were significantly decreased in
comparison with the control group (Fig. 8A and B). Furthermore, RT-qPCR and
western blotting revealed that the protein expression levels of
P53, P21 and Bax were significantly upregulated, whereas expression
levels of CDK4, Cyclin D1 and Bcl-2 were significantly
downregulated in tumor tissues following treatment with AC compared
with untreated controls. Moreover, administration of AC caused a
significant decrease in the phosphorylation of IκBα and P65
(Fig. 8C and D). These results
were consistent with those of the in vitro analysis.
Discussion
The present study revealed that AC, a plant-derived
triterpenoid, induced cell cycle arrest at the G0/G1 phase,
promoted CRC cell apoptosis and suppressed cell viability. AC
treatment contributed to the aberrant expression of the cell
cycle-related genes CDK4, Cyclin D1 and P53 and P21. AC increased
the activation of caspase-9 and caspase-3 (apoptosis-related
genes), upregulated Bax level, and downregulated Bcl-2 expression.
Furthermore, AC inhibited the phosphorylation of IκBα and NF-κB
p65, resulting in the inhibition of the NF-κB signaling pathway.
This consequently affects various biological activities in
eukaryotic cells, including cell proliferation, survival and
apoptosis. Furthermore, a synergistic inhibitory effect of AC and
the NF-κB signaling pathway inhibitor, JSH-23 on CRC was observed.
AC suppressed proliferation of CRC cells both in vitro and
in vivo, implying that it can be a potential adjuvant agent
for the treatment of CRC.
A number of studies have demonstrated that AC
exhibits antitumor activity (17,18,20). AC increases the sensitivity of the
human oral epithelial cancer cells and breast cancer cells to
chemotherapy drugs by increasing intracellular reactive oxygen
species and activating apoptotic pathways in vitro (19). AC inhibits breast cancer
progression in mice by downregulating IL-1β expression and
activating downstream signaling pathways in vivo (20). The present study demonstrated that
AC significantly suppressed proliferation and colony formation in
CRC cells. Decreased mitochondrial membrane potential is a common
phenomenon of mitochondrial apoptosis in cancer cells (4), which results in the release of
mitochondrial apoptogens to initiate the caspase cascade, causing
programmed cell death (26,27). The present study identified that
AC treatment for 24 h decreased the mitochondrial membrane
potential in HCT116 cells and induced CRC cell apoptosis. At this
point, the proportion of cell apoptosis in the AC-treated group was
not so high, but the mitochondrial membrane potential was
significantly altered, indicating that the change of mitochondrial
membrane potential occurred earlier than cell apoptosis. When
HCT116 cells were treated with AC for 72 h, cell apoptosis was
significantly elevated in comparison with the untreated cells.
Hence, the mitochondrial membrane potential was not detected.
Previous studies have demonstrated that the
decreased ratio of Bcl-2/Bax regulates apoptosis in cells (28,29). In the present study, AC-induced
cell apoptosis was coupled with upregulation of Bax, cleaved
caspase-3 and cleaved caspase-9, and the downregulation of Bcl-2.
Additionally, the cell cycle transition from G0/G1 to S is effected
by multiple genes, such as CDK4/6 and Cyclin D1 (30). A study has indicated that
downregulation of CDK4 expression causes cell cycle arrest at the
G0/G1 phase (31), which may
explain the mechanism of antitumor effects of several drugs. The
present results indicated that AC significantly inhibited the
proliferation of CRC cells by interrupting the cell cycle at the
G0/G1 phase. AC treatment substantially reduced the expression of
CDK4 and Cyclin D1. However, the specific mechanism involved in the
regulation of CDK4 and Cyclin D1 gene expression remains
unknown.
AC suppresses the inflammatory response after injury
via regulation of the NF-κB signaling pathway (14,32,33). IκBα is an inhibitory protein of
the NF-κB signaling pathway that normally binds to P65 resulting in
the inhibition of P65 phosphorylation and its nuclear translocation
(34). Conversely, IκBα
phosphorylation promotes its own ubiquitination and degradation,
thereby eliminating the inhibitory effect on P65 (35,36). Once P65 enters the nucleus, it
triggers a series of gene expressions such as Bcl-2 and CDK4
(37). Results of the present
study revealed that AC treatment significantly reduced IκBα and P65
phosphorylation, resulting in inhibition of P65 nuclear
translocation and interfering with the expression of downstream
molecules. Although results of the in vivo experiments
demonstrated that AC administration significantly inhibited tumor
growth without evident adverse effects on mice, the clinical use of
AC as an antitumor drug requires further study.
In conclusion, AC inhibited activation of the NF-κB
signaling pathway and subsequently modulated the expression of its
downstream genes by attenuating IκBα and P65 phosphorylation, which
may induce apoptosis and cell cycle arrest in CRC cells. Thus,
proliferation of CRC cells in vitro and tumor growth in
vivo were suppressed. Therefore, AC is a promising adjuvant
with antitumor effects in CRC.
Acknowledgments
Not applicable.
Funding
This study was financed by the National Social
Science Foundation of China (grant no. 81871877), National Major
Scientific and Technological Special Project for 'Significant New
Drugs Development' (grant no. 2020ZX09201005) and Youth Foundation
of Xiamen Cancer Center (grant nos. ZLYYA201710 and
ZLYYA201704).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YM and CK conceived and designed the study. XZ, YL
and YM analyzed the experimental data. YM and CK acquired the
funding. XZ, YL, CR, TL and FD performed the experiments. XZ, YL,
CR, YM and CK participated in the design of the experimental
methods. XZ, YM and CK wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Care and Use Committee of The First Affiliated Hospital of Xiamen
University (Xiamen, China; approval no. XMU-AEA-20180137) and
carried out in accordance with the Guide for the Care and Use of
Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Nogueira L, Mariotto AB,
Rowland JH, Yabroff KR, Alfano CM, Jemal A, Kramer JL and Siegel
RL: Cancer treatment and survivorship statistics, 2019. CA Cancer J
Clin. 69:363–385. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van der Jeught K, Xu HC, Li YJ, Lu XB and
Ji G: Drug resistance and new therapies in colorectal cancer. World
J Gastroenterol. 24:3834–3848. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bahrami A, Amerizadeh F, Hassanian SM,
ShahidSales S, Khazaei M, Maftouh M, Ghayour-Mobarhan M, Ferns GA
and Avan A: Genetic variants as potential predictive biomarkers in
advanced colorectal cancer patients treated with oxaliplatin-based
chemotherapy. J Cell Physiol. 233:2193–2201. 2018. View Article : Google Scholar
|
|
5
|
Bylka W, Znajdek-Awiżeń P,
Studzińska-Sroka E, Dańczak-Pazdrowska A and Brzezińska M: Centella
asiatica in dermatology: An overview. Phytother Res. 28:1117–1124.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chandrika UG, Prasad Kumarab and Peramune
AAS: Gotu Kola (Centella asiatica): Nutritional properties and
plausible health benefits. Adv Food Nutr Res. 76:125–157. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
da Rocha PBR, Santos Souza BD, Andrade LM,
Marreto RN, Lima EM and Taveira SF: Development of a
high-performance liquid chromatographic method for asiaticoside
quantification in different skin layers after topical application
of a Centella asiatica extract. Planta Med. 83:1431–1437. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nowwarote N, Osathanon T, Jitjaturunt P,
Manopattanasoontorn S and Pavasant P: Asiaticoside induces type I
collagen synthesis and osteogenic differentiation in human
periodontal ligament cells. Phytother Res. 27:457–462. 2013.
View Article : Google Scholar
|
|
9
|
Lee J, Jung E, Kim Y, Park J, Park J, Hong
S, Kim J, Hyun C, Kim YS and Park D: Asiaticoside induces human
collagen I synthesis through TGFbeta receptor I kinase (TbetaRI
kinase)-independent Smad signaling. Planta Med. 72:324–328. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Phaechamud T, Yodkhum K, Charoenteeraboon
J and Tabata Y: Chitosan-aluminum monostearate composite sponge
dressing containing asiaticoside for wound healing and angiogenesis
promotion in chronic wound. Mater Sci Eng C Mater Biol Appl.
50:210–225. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang J, Zhou X, Shen Y, Li H, Zhou G,
Zhang W, Zhang Y and Liu W: Asiaticoside loading into
polylactic-co-glycolic acid electrospun nanofibers attenuates host
inflammatory response and promotes M2 macrophage polarization. J
Biomed Mater Res A. 108:69–80. 2020. View Article : Google Scholar
|
|
12
|
Zhang CZ, Niu J, Chong YS, Huang YF, Chu
Y, Xie SY, Jiang ZH and Peng LH: Porous microspheres as promising
vehicles for the topical delivery of poorly soluble asiaticoside
accelerate wound healing and inhibit scar formation in vitro &
in vivo. Eur J Pharm Biopharm. 109:1–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qi SH, Xie JL, Pan S, Xu YB, Li TZ, Tang
JM, Liu XS, Shu B and Liu P: Effects of asiaticoside on the
expression of Smad protein by normal skin fibroblasts and
hypertrophic scar fibroblasts. Clin Exp Dermatol. 33:171–175. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu J, Yu L, Zhang X, Wu Q, Wang D, Wang
X, Xia C and Feng H: Asiaticoside attenuates
lipopolysaccharide-induced acute lung injury via down-regulation of
NF-kB signaling pathway. Int Immunopharmacol. 26:181–187. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luo Y, Fu C, Wang Z, Zhang Z, Wang H and
Liu Y: Asiaticoside attenuates the effects of spinal cord injury
through antioxidant and antiinflammatory effects, and inhibition of
the p38MAPK mechanism. Mol Med Rep. 12:8294–8300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fitri AR, Pavasant P, Chamni S and
Sumrejkanchanakij P: Asiaticoside induces osteogenic
differentiation of human peri-odontal ligament cells through the
Wnt pathway. J Periodontol. 89:596–605. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Al-Saeedi FJ, Bitar M and Pariyani S:
Effect of asiaticoside on 99mTc-tetrofosmin and 99mTc-sestamibi
uptake in MCF-7 cells. J Nucl Med Technol. 39:279–283. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yingchun L, Huihan W, Rong Z, Guojun Z,
Ying Y and Zhuogang L: Antitumor activity of asiaticoside against
multiple myeloma drug-resistant cancer cells is mediated by
autophagy induction, activation of effector caspases, and
inhibition of cell migration, invasion, and STAT-3 signaling
pathway. Med Sci Monit. 25:1355–1361. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang YH, Zhang SH, Zhen RX, Xu XD and
Zhen YS: Asiaticoside inducing apoptosis of tumor cells and
enhancing anti-tumor activity of vincristine. Ai Zheng.
23:1599–1604. 2004.In Chinese. PubMed/NCBI
|
|
20
|
Al-Saeedi FJ: Study of the cytotoxicity of
asiaticoside on rats and tumour cells. BMC Cancer. 14:2202014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Perelman A, Wachtel C, Cohen M, Haupt S,
Shapiro H and Tzur A: JC-1: Alternative excitation wavelengths
facilitate mitochondrial membrane potential cytometry. Cell Death
Dis. 3:e4302012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Soleimani A, Rahmani F, Ferns GA, Ryzhikov
M, Avan A and Hassanian SM: Role of the NF-κB signaling pathway in
the pathogenesis of colorectal cancer. Gene. 726:1441322020.
View Article : Google Scholar
|
|
24
|
Zhang Q, Lenardo MJ and Baltimore D: 30
Years of NF-κB: A blossoming of relevance to human pathobiology.
Cell. 168:37–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
de Castro Barbosa ML, da Conceicao RA,
Fraga AGM, Camarinha BD, de Carvalho Silva GC, Lima AGF, Cardoso
EA, de Oliveira Freitas and Lione V: NF-kappaB signaling pathway
inhibitors as anticancer drug candidates. Anticancer Agents Med
Chem. 17:483–490. 2017. View Article : Google Scholar
|
|
26
|
Kocyigit A and Guler EM: Curcumin induce
DNA damage and apoptosis through generation of reactive oxygen
species and reducing mitochondrial membrane potential in melanoma
cancer cells. Cell Mol Biol (Noisy-le-grand). 63:97–105. 2017.
View Article : Google Scholar
|
|
27
|
Yun X, Rao W, Xiao C and Huang Q:
Apoptosis of leukemia K562 and Molt-4 cells induced by emamectin
benzoate involving mitochondrial membrane potential loss and
intracellular Ca(2+) modulation. Environ Toxicol Pharmacol.
52:280–287. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Y, Zong M, Xu W, Zhang Y, Wang B,
Yang M and Tao L: Natural pyrethrins induces apoptosis in human
hepatocyte cells via Bax- and Bcl-2-mediated mitochondrial pathway.
Chem Biol Interact. 262:38–45. 2017. View Article : Google Scholar
|
|
29
|
Tang S, Hu J, Meng Q, Dong X, Wang K, Qi
Y, Chu C, Zhang X and Hou L: Daidzein induced apoptosis via
down-regulation of Bcl-2/Bax and triggering of the mitochondrial
pathway in BGC-823 cells. Cell Biochem Biophys. 65:197–202. 2013.
View Article : Google Scholar
|
|
30
|
Zhu J, Chen M, Chen N, Ma A, Zhu C, Zhao
R, Jiang M, Zhou J, Ye L, Fu H and Zhang X: Glycyrrhetinic acid
induces G1-phase cell cycle arrest in human nonsmall cell lung
cancer cells through endo-plasmic reticulum stress pathway. Int J
Oncol. 46:981–988. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bonelli M, Monica SL, Fumarola C and
Alfieri R: Multiple effects of CDK4/6 inhibition in cancer: From
cell cycle arrest to immunomodulation. Biochem Pharmacol.
170:1136762019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He L, Hong G, Zhou L, Zhang J, Fang J, He
W, Tickner J, Han X, Zhao L and Xu J: Asiaticoside, a component of
Centella asiatica attenuates RANKL-induced osteoclastogenesis via
NFATc1 and NF-kB signaling pathways. J Cell Physiol. 234:4267–4276.
2019. View Article : Google Scholar
|
|
33
|
Yin Z, Yu H, Chen S, Ma C, Ma X, Xu L, Ma
Z, Qu R and Ma S: Asiaticoside attenuates diabetes-induced
cognition deficits by regulating PI3K/Akt/NF-kB pathway. Behav
Brain Res. 292:288–299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meshram SN, Paul D, Manne R, Choppara S,
Sankaran G, Agrawal Y and Santra MK: FBXO32 activates NF-kB through
IkBα degradation in inflammatory and genotoxic stress. Int J
Biochem Cell Biol. 92:134–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tsuchiya Y, Osaki K, Kanamoto M, Nakao Y,
Takahashi E, Higuchi T and Kamata H: Distinct B subunits of PP2A
regulate the NF-kB signalling pathway through dephosphorylation of
IKKβ, IkBα and RelA. FEBS Lett. 591:4083–4094. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu J, Haseebuddin M, Young M and Colburn
NH: Suppression of p65 phosphorylation coincides with inhibition of
IkappaBalpha polyubiquitination and degradation. Mol Carcinog.
44:274–284. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thoms HC, Dunlop MG and Stark LA:
p38-mediated inactivation of cyclin D1/cyclin-dependent kinase 4
stimulates nucleolar translocation of RelA and apoptosis in
colorectal cancer cells. Cancer Res. 67:1660–1669. 2007. View Article : Google Scholar : PubMed/NCBI
|