Introduction
Esophageal cancer is the seventh most common type of
human cancer and the 6th leading cause of cancer-associated
mortality worldwide (1). More
than 455,800 newly diagnosed esophageal cancer cases and 400,200
related deaths are reported worldwide each year (2), and the majority of these cases are
detected in Asia, Northern France, and Eastern and Southern Africa
(3). Esophageal squamous cell
carcinoma (ESCC), the major histological type of esophageal cancer,
accounts for approximately 90% of all esophageal cancer cases
(4). Approximately half of all
patients with ESCC present with advanced-stage disease at the time
of the initial diagnosis, and these patients thus have poor
therapeutic responses following surgical resection (5). Despite tremendous advances being
made in multidisciplinary therapies involving surgical excision and
chemoradiotherapy, the clinical efficacy of treatment for ESCC
remains largely unfavorable, and the 5-year survival rate is
<25%. Multiple factors, including the excessive consumption of
alcohol, smoking, the consumption of very hot tea and red meat, and
a low intake of fruits and vegetables, have been identified to be
closely associated with the pathogenesis of ESCC (6,7);
however, the complex mechanistic events remain largely unknown.
Hence, the determination of the detailed mechanisms closely
associated with the pathogenesis of ESCC would represent a highly
significant step toward improving the diagnostic techniques and
therapeutics for ESCC.
Long non-coding RNAs (lncRNAs) are a class of long
nucleotide transcripts (>200 nucleotides in length) that are
incapable of coding proteins (8).
lncRNAs contribute to the control of gene expression at the
transcriptional, post-transcriptional and chromosomal levels
(9). A number of studies have
verified the regulatory effects of lncRNAs in numerous normal
physiological and pathological processes (10-12). In recent decades, the significance
of lncRNAs in carcinogenesis and progression has been recognized by
researchers (13-15). The differential expression of
various lncRNAs in ESCC has been identified, and some lncRNAs have
been demonstrated to be crucial contributors to the oncogenicity of
ESCC via their cancer-inhibiting or cancer-promoting activities
(16-18).
MicroRNAs (miRNAs or miRs) are a class of
endogenous, highly conserved and non-coding RNA transcripts with
lengths of 17-23 nucleotides (19). miRNAs participate in the
regulation of gene expression at the post-transcriptional level by
binding directly to the 3′-untranslated regions (3′-UTRs) of target
genes and consequently triggering the degradation of mRNAs and/or
inhibiting translation (20).
Research has highlighted the necessary role of miRNAs in the onset
and progression of ESCC (21).
Existing evidence has demonstrated the dysregulation of miRNAs in
ESCC and the capacities of these molecules to regulate various
malignant phenotypes (22-24).
The competing endogenous (ce)RNA hypothesis suggests that lncRNAs
function predominantly as molecular sponges to decrease the
availability of target miRNAs, thereby reducing the suppressive
effects of miRNAs on their downstream mRNA targets (25). Therefore, an exploration of the
lncRNAs and miRNAs involved in the oncogenesis and development of
ESCC may help identify potential targets for the diagnosis,
prevention, and treatment of this malignancy.
Long intergenic non-coding RNA LINC01232 has been
identified as a critical regulator of pancreatic adenocarcinoma
oncogenesis (26). However, the
involvement of LINC01232 in ESCC has not yet been thoroughly
elucidated. In the present study, the pattern of LINC01232
expression in ESCC was examined, and the regulatory roles of this
lncRNA in ESCC cells, as well as the underlying mechanisms were
investigated.
Materials and methods
Clinical tissues, cell culture and cell
transfection
A total of 55 pairs of ESCC tissues and matched
normal tissues were collected from patients (34 males, 21 females;
age range, 57-74 years) with ESCC at the Affiliated Hospital of
Inner Mongolia University for the Nationalities. Follow-up was
performed through outpatient visits or via telephone (every 1-2
months). All tissues were obtained from patients who had not
undergone chemotherapy or radiotherapy prior to surgical excision.
Immediately following excision, all tissue specimens were
snap-frozen and stored in liquid nitrogen prior to RNA isolation.
Human clinical tissues were collected and used in accordance with
the Institutional Ethics Committee of the Affiliated Hospital of
Inner Mongolia University for the Nationalities (#17-0602). Written
informed consent was provided by all patients prior to tissue
collection.
A total of 3 ESCC cell lines, Eca109, TE-1 and
KYSE150, were acquired from the Type Culture Collection of the
Chinese Academy of Sciences. The Eca109 and TE-1 cells were grown
in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.), while
the KYSE150 cells were cultured in RPMI-1640/F12 medium (Gibco;
Thermo Fisher Scientific, Inc.). Both types of basal media were
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Gibco; Thermo
Fisher Scientific, Inc.). The normal human esophageal epithelial
cell line, HET-1A, was obtained from the American Type Culture
Collection (ATCC) and was cultured in BEGM medium (Lonza/Clonetics
Corporation) supplemented with 10% FBS. All cells were cultured at
37°C in a humidified incubator containing 5% CO2.
Small interfering RNAs (siRNAs) specific for
LINC01232 (si-LINC01232#1, si-LINC01232#2 and si-LINC01232#3) and
scrambled siRNAs (si-ctrl) were generated by Ribobio (Guangzhou
RiboBio Co., Ltd.). The si-LINC01232 sequences were as follows:
si-LINC01232 #1, 5′-GTG TAA TTT CAC TTG AAT AAA T-3′; si-LINC01232
#2, 5′-GTG CGT TTT GTC CAA TTA TTT GT-3′; and si-LINC01232 #3,
5′-GAC TTC TTA GAA CTA AAT TGA AA-3′. The si-NC sequence was 5′-CAC
GAT AAG ACA ATG TAT TT-3′. miR-654-3p mimic, negative control (NC)
miRNA mimic, miR-654-3p inhibitor and NC inhibitor were purchased
from GenePharma. The plasmid used to induce HDGF overexpression
(pcDNA3.1-HDGF) was produced by GenePharma, and the empty pcDNA3.1
plasmid vector was used as the negative control. Cells were
cultured in 6-well plates and transfected with miRNA mimics (100
pmol), miRNA inhibitors (100 pmol), siRNAs (100 pmol), or plas-mids
(4 µg) using Lipofectamine 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR), flow cytometric analysis, and
migration and invasion assays were conducted at 48 h
post-transfection. Cell Counting kit-8 (CCK-8) assay and western
blot analysis were performed following 24 and 72 h of incubation,
respectively.
Isolation of cell cytosolic/nuclear
fractions
The nuclear and cytosolic fractions of ESCC cells
were separated using the Cytoplasmic & Nuclear RNA Purification
kit (Norgen Biotek Corp.) in accordance with the manufacturer's
protocol. RNA was isolated from the nuclear and cytosolic fractions
and used in RT-qPCR assays to determine the distribution of
LINC01232 in ESCC cells.
RT-qPCR
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract total RNA. To quantify the
expression of LINC01232 and HDGF, reverse transcription was
conducted using the PrimeScript™ RT reagent kit with gDNA Eraser
(Takara, Dalian, China). The resultant first-strand cDNA was used
as a template for qPCR, which was performed using TB
Green® Premix Ex Taq™ II (Takara Biotechnology Co.,
Ltd.). To analyze the expression of miR-654-3p, the Mir-X miRNA
First-Strand Synthesis kit (Takara Biotechnology Co., Ltd.) was
used to prepare cDNA, and the Mir-X miRNA qRT-PCR TB
Green® Kit was then used to perform qPCR (Takara
Biotechnology Co., Ltd.). GAPDH and U6 small nuclear RNA
were used as the reference transcripts for lncRNA/mRNA and miRNA,
respectively. All data were analyzed using the 2−ΔΔCq
method (27). All primer
sequences are listed in Table
I.
 | Table ISequences of primers used in
RT-qPCR. |
Table I
Sequences of primers used in
RT-qPCR.
| Gene | Sequences
(5′-3′) |
|---|
| LINC01232 | Forward:
AAAACCTTGAAATCCCTTAATACCA |
| Reverse:
CCTTACCCGTGGAATTCACATATA |
| HDGF | Forward:
AATCAACAGCCAACAAATACCAAGT |
| Reverse:
AGCCTTGACAGTAGGGTTGTTCTC |
| GAPDH | Forward:
CGGAGTCAACGGATTTGGTCGTAT |
| Reverse:
AGCCTTCTCCATGGTGGTGAAGAC |
| miR-122-5p | Forward:
TCGGCAGGUGGAGUGUGACAAUG |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-181b-5p | Forward:
TCGGCAGGGGUCACAAUCAACAUUC |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-181c-5p | Forward:
TCGGCAGGGGUUUGGGGGAACAUUC |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-181d-5p | Forward:
TCGGCAGGGGUCACAAUCAACAUUC |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-211-5p | Forward:
TCGGCAGGUUCCCUUUGUCAUCC |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-370-5p | Forward:
TCGGCAGGCAGGUCACGUCUC |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-493-5p | Forward:
TCGGCAGGUUGUACAUGGUAGG |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-506-5p | Forward:
TCGGCAGGUAUUCAGGAAGGUG |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-590-5p | Forward:
TCGGCAGGGAGCUUAUUCAUAA |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-642b-5p | Forward:
TCGGCAGGGGUUCCCUCUCCA |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| miR-654-3p | Forward:
TCGGCAGGUGGUGGGCCGCAG |
| Reverse:
CACTCAACTGGTGTCGTGGA |
| U6 | Forward:
GCTTCGGCAGCACATATACTAAAAT |
| Reverse:
CGCTTCACGAATTTGCGTGTCAT |
CCK-8 assay
The CCK-8 assay was conducted to determine cell
proliferation. At 48 h following transfection, cells were collected
and seeded into 96-well plates. Each well contained 100 µl
of cell suspension containing 2×103 cells. The cells
were incubated in a humidified incubator with 5% CO2 at
37°C. At 0, 24, 48 and 72 h following cell inoculation, the culture
medium was carefully discarded, and 100 µl of fresh cultured
medium supplemented with 10 µl of CCK-8 solution (Dojindo
Molecular Technologies, Inc.) was added to each well. Following
incubation at 37°C for 2 h, the absorbance was detected in each
well at a wavelength of 450 nm using a microplate reader (BioRad
Laboratories, Inc.).
Flow cytometry
Following 48 h of culture, the transfected cells
were treated with ethylenediaminetetraacetic acid (EDTA)-free
trypsin (Gibco; Thermo Fisher Scientific, Inc.) and washed with
precooled phosphate-buffered saline (Gibco; Thermo Fisher
Scientific, Inc.), followed by centrifugation at 4°C at 12,000 × g
for 5 min. Subsequently, an Annexin V-fluorescein isothiocyanate
(FITC) apoptosis detection kit (BioLegend, Inc.) was used to assess
cell apoptosis quantitatively. The collected cells were suspended
in 100 µl of binding buffer and subsequently mixed with 10
µl of Annexin V-FITC and 5 µl of propidium iodide.
Following 15 min of incubation at room temperature in the dark, the
apoptotic rate was determined using a flow cytometer (FACScan; BD
Biosciences).
Migration and invasion assays
For the invasion assays, the transfected cells were
rinsed with phosphate-buffered saline and resuspended in FBS-free
culture medium. Subsequently, 200 µl of cell suspension
containing 5×104 cells was plated into each upper
compartment of a Biocoat Matrigel Invasion Chamber (BD
Biosciences), while the lower compartments were loaded with 600
µl of culture medium containing 20% FBS to serve as a
chemoattractant. After 24 h, the invaded cells were fixed with 4%
paraformaldehyde at 37°C for 0.5 h and stained with 0.1% crystal
violet at 37°C for 0.5 h. The residual cells remaining on the upper
surfaces were carefully wiped off with a cotton bud. For the
migration assay, all experimental steps were performed as described
above, except the chambers were not pre-coated with Matrigel. An
inverted microscope (IX31; Olympus Corporation) was used to image
the migrated and invaded cells. In total, 5 fields were randomly
selected per insert, and the transmigrated cells were counted
manually under a microscope as mentioned above.
Xenograft mouse model
A short hairpin RNA (shRNA) targeting LINC01232
(sh-LINC01232) and scrambled shRNA (sh-ctrl) were designed and
generated by GenePharma and inserted into the GenePharma
Supersilencing Vector. Subsequently, the vectors were transfected
into 293T cells (the Type Culture Collection of the Chinese Academy
of Sciences) along with lentiviral packaging plasmids. The
resulting lenti-viruses were collected following 72 h of incubation
at 37°C, mixed with polybrene (5 µg/ml; Sigma-Aldrich; Merck
KGaA) and used to infect the Eca109 cells at an MOI of 5. Eca109
cells exhibiting stable sh-LINC01232 or sh-ctrl expression were
selected using puromycin.
The animal experiments were conducted with the
approval of the Institutional Animal Care and Use Committee of the
Affiliated Hospital of Inner Mongolia University for the
Nationalities. BALB/c nude mice (n=6; 4-6 weeks old) were purchased
from Beijing Vital River Laboratory Animal Technology Co., Ltd.,
and kept in a specific pathogen-free environment at 25°C with 50%
humidity, with a 10/14-h light/dark cycle and ad libitum food/water
access. Mice were subcutaneously injected with Eca109 cells that
stably expressed sh-LINC01232 or sh-ctrl. Each group included 3
mice. The sizes of the resulting subcutaneous xenografts were
monitored by measuring the tumor length (L) and width (W) every 6
days. The tumor volume was calculated using the formula: Volume=0.5
× (L × W2). All mice were euthanized at the end of day
30, and the tumor xenografts were imaged and weighed.
Bioinformatics analysis
GEPIA (http://gepia.cancer-pku.cn/) contained the TCGA and
GTEx databases, and was used to analyze the LINC01232 expression
profile in ESCC tumors (n=182) and normal healthy tissues (n=286).
lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/) was
applied to predict the localization of LINC01232.
StarBase 3.0 (http://starbase.sysu.edu.cn/) was used to identify the
potential miRNAs that may be adsorbed by LINC01232. The putative
target of miR-654-3p was predicted by StarBase 3.0, TargetScan
(http://www.targetscan.org/), and miRDB
(http://mirdb.org/).
RNA immunoprecipitation (RIP) assay
To assess the interaction between LINC01232 and
miR-654-3p in ESCC cells, RIP assay was performed using the Magna
RIP RNA-Binding Protein Immunoprecipitation kit (EMD Millipore).
ESCC cells were treated with RIP buffer. An anti-Argonaute2 (Ago2)
antibody or normal mouse immunoglobulin G (IgG) antibody (1:5,000
dilution; both cat. no. 03-110; EMD Millipore) was conjugated to
magnetic beads, which were then incubated overnight with whole-cell
extracts at 4°C. The magnetic beads were then harvested and treated
with proteinase K, and the immunoprecipitated RNA was extracted.
The enrichment of LINC01232 and miR-654-3p in immunoprecipitated
RNA was evaluated by RT-qPCR.
Luciferase reporter assay
The wild-type (wt) fragments of LINC01232 and HDGF
3′-UTR containing the miR-654-3p binding site were amplified by
RT-qPCR and inserted down-stream of the pmirGLO dual-luciferase
vector (Promega Corporation) to yield the LINC01232-wt and HDGF-wt
reporter plasmids, respectively. Binding site-directed mutagenesis
was conducted using the GeneTailor™ Site-Directed Mutagenesis
System (Invitrogen; Thermo Fisher Scientific, Inc.) to generate
LINC01232-mutant (LINC01232-mut) and HDGF-mut. ESCC cells were
seeded in 24-well plates and co-transfected with miR-654-3p mimic
or NC mimic and wt or mut reporter plasmids. Luciferase activity
was measured 48 h following transfection using the Dual-Luciferase
Reporter Assay System (Promega Corporation), and normalized to
Renilla luciferase activity.
Western blot analysis
Total proteins were isolated from the cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology, Inc.), and
the concentration in each lysate was measured using a BCA Protein
Assay kit (Beyotime Institute of Biotechnology, Inc.). Equal
amounts of total protein (30 µg) were subjected to
electrophoresis on 10% sodium dodecyl sulphate-polyacrylamide gel
electrophoresis gels and trans-ferred to polyvinylidene difluoride
membranes. Membranes were blocked for 2 h in 5% non-fat milk at
room temperature and then probed with primary antibodies overnight
at 4°C. Subsequently, the membranes were incubated with a
horse-radish peroxidase-conjugated goat anti-rabbit IgG secondary
antibody (1:5,000 dilution; cat. no. ab205718; Abcam) at room
temperature for 1 h. Finally, Immobilon® ECL Ultra
Western HRP Substrate (EMD Millipore) was used to visualize the
signals corresponding to the labeled proteins. Primary anti-bodies
specific for HDGF (cat. no. ab128921; Abcam) and GAPDH (cat. no.
ab181603; Abcam) were used at a dilution of 1:1,000. Densitometry
was performed using Quantity One software version 4.62 (Bio-Rad
Laboratories, Inc.).
Statistical analysis
All data are presented as the means ± standard
deviation. Data from 2 groups were compared using a Student's
t-test, whereas data from multiple groups were compared using
one-way analysis of variance, followed by Tukey's test. Pearson's
correlation coefficient was used to analyze the correlation among
LINC01232, miR-654-3p and HDGF expression in ESCC tissues. The
overall survival of ESCC patients was analyzed via the Kaplan-Meier
method, and compared with the log-rank test. All statistical
analyses were performed using SPSS version 19.0 software package
(IBS SPSS, Inc.), and P-values <0.05 were considered to indicate
statistically significant differences.
Results
Interference of LINC01232 inhibits the
malignant characteristics of ESCC cells
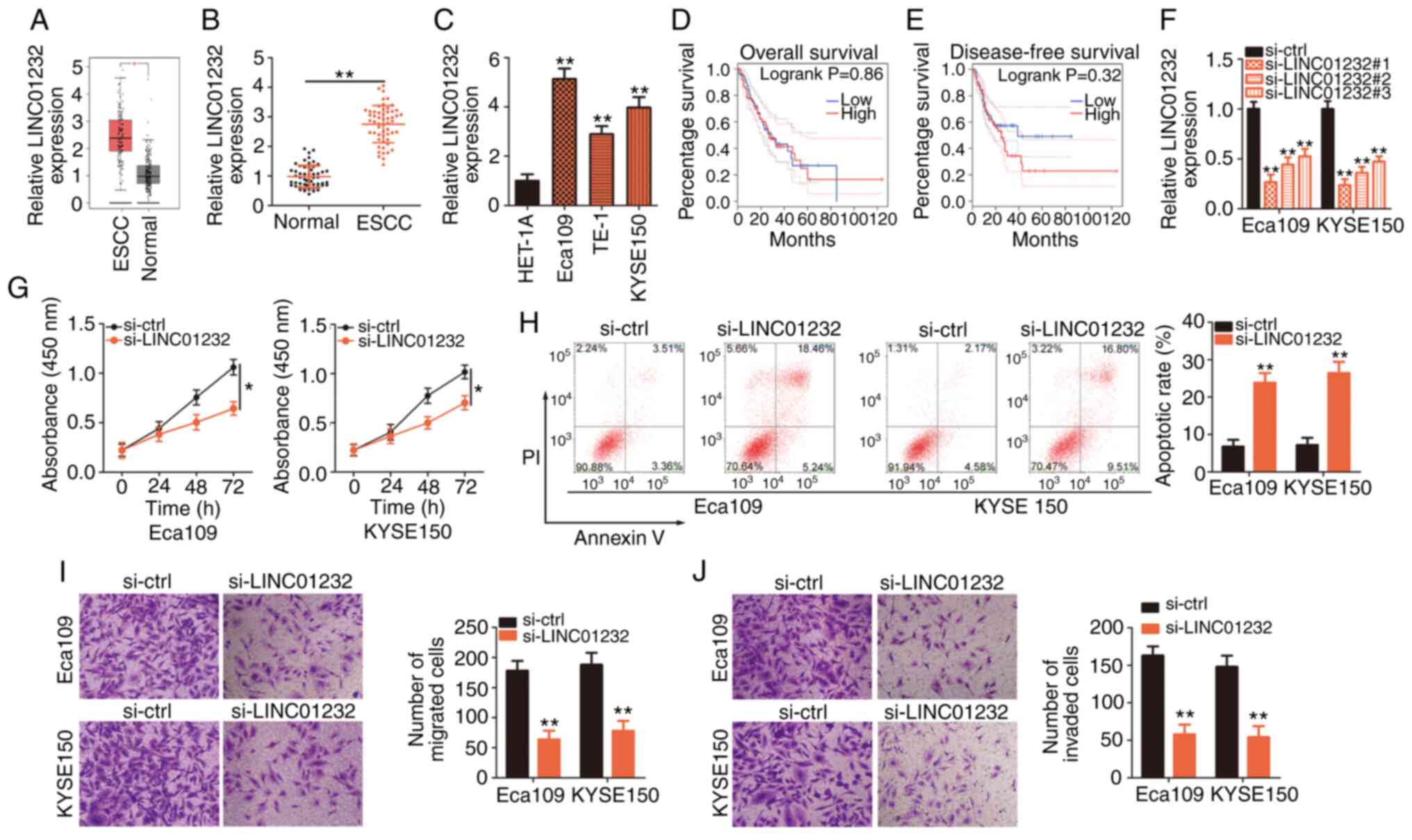
Initially, GEPIA (http://gepia.cancer-pku.cn/) was used to analyze the
LINC01232 expression profile in ESCC. Data from the TCGA and GTEx
databases revealed high levels of LINC01232 expression in ESCC
tissues compared to normal tissues (Fig. 1A). To confirm this observation,
RT-qPCR was performed to determine the abundance of LINC01232 in 55
pairs of ESCC tissues and matched normal tissues. RT-qPCR validated
the upregulation of LINC01232 in ESCC tissues compared with that in
matched normal tissues (Fig. 1B).
Additionally, RT-qPCR was used to determine the expression of
LINC01232 in the ESCC cell lines, Eca109, TE-1 and KYSE150, with
the HET-1A normal human esophageal epithelial cells used as the
control. LINC01232 was expressed at higher levels in all 3 ESCC
cell lines compared with the control cells (Fig. 1C). The TCGA and GTEx data were
analyzed to determine the clinical relevance of LINC01232 in ESCC
and it was revealed that its expression was not associated with the
overall survival (Fig. 1D) or
disease-free survival (Fig. 1E)
of patients with ESCC.
LINC01232 expression was most abundant in the Eca109
and KYSE150 cell lines; thus, these cell lines were selected for
further experimental use. To determine the functional roles of
LINC01232 in ESCC cellular processes, si-LINC01232 was designed to
silence LINC01232 expression in the Eca109 and KYSE150 cells. The
silencing efficiency was confirmed by RT-qPCR. Of the 3 siRNAs,
si-LINC01232#1 exhibited the most efficient interference and was
thus used in subsequent experiments (Fig. 1F). Transfection with si-LINC01232
inhibited the proliferation (Fig.
1G) and promoted the apoptosis (Fig. 1H) of both the Eca109 and KYSE150
cells. Moreover, LINC01232 knockdown evidently impaired the
migratory (Fig. 1I) and invasive
capabilities (Fig. 1J) of the
Eca109 and KYSE150 cells, as demonstrated by migration and invasion
assays, respectively. In summary, LINC01232 expression was found to
be upregulated in ESCC and to exert pro-oncogenic effects during
cancer progression.
miR-654-3p is sponged by LINC01232 and
serves as a tumor inhibitor in ESCC cells
To explore the mechanisms underlying the effects of
LINC01232, the distribution of LINC01232 in ESCC cells was first
analyzed. Using lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/), it
was determined that LINC01232 was distributed primarily in the
cytoplasm (Fig. 2A), which was
consistent with the results of cell cytosolic/nuclear fraction
isolation and RT-qPCR analysis (Fig.
2B). Previous studies have revealed that cytoplasmic lncRNAs
can serve as molecular sponges to prevent the suppression of target
mRNAs by specific miRNAs (28-30). Therefore, it was hypothesize that
LINC01232 may also serve as a sponge for miRNA and would thus
regulate the expression of specific miRNAs.
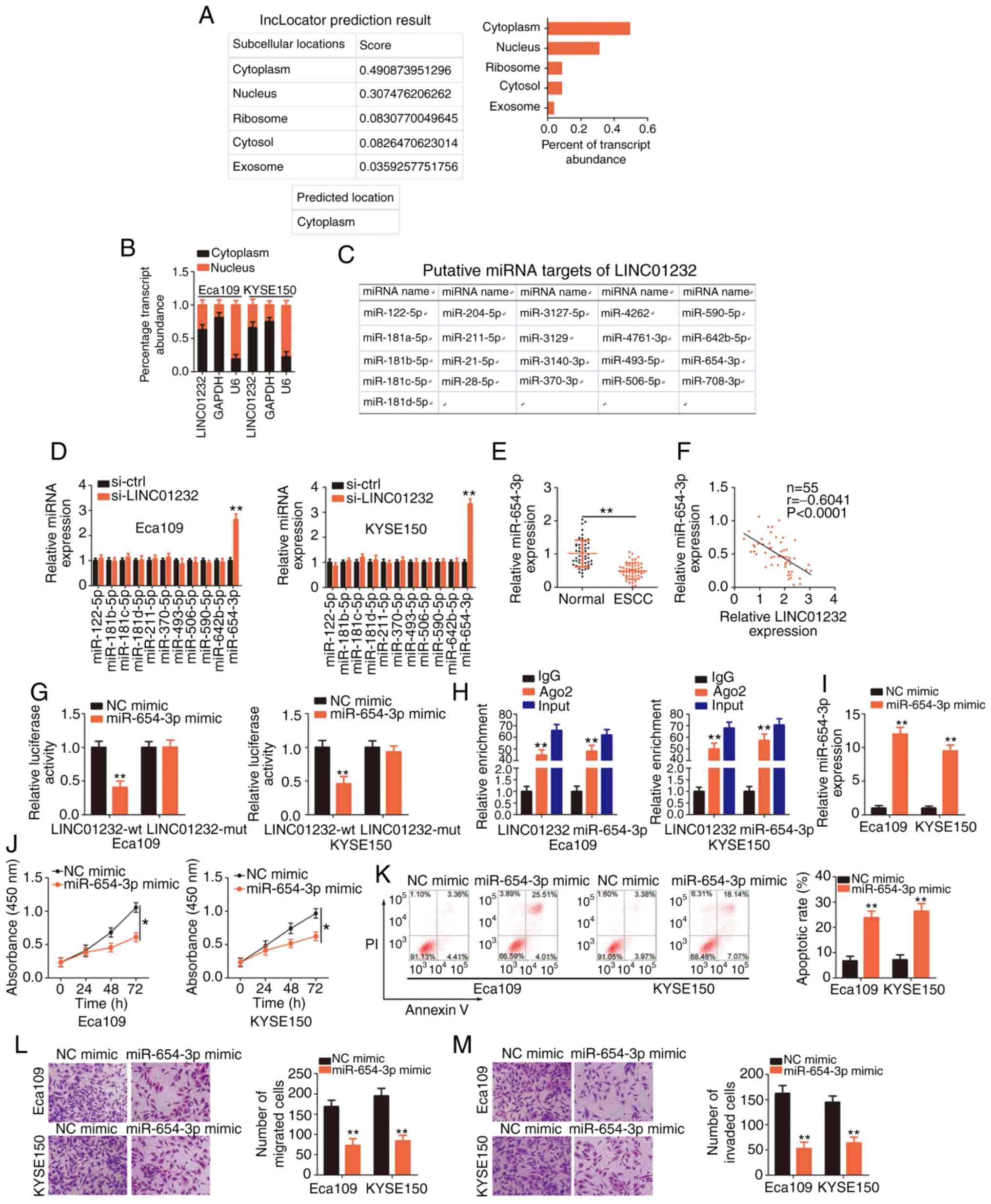 | Figure 2MicroRNA (miR)-654-3p is sponged by
long intergenic non-coding RNA (LINC)01232 and plays an inhibitory
role in esophageal squamous cell carcinoma (ESCC). (A) The
lncLocator database was used to examine the subcellular location of
LINC01232. (B) The subcellular location of LINC01232 was evaluated
in cell cytosolic/nuclear fractions isolated from Eca109 and
KYSE150 ESCC cells. (C) Potential LINC01232-interacting miRNAs
identified through a StarBase 3.0 search. (D) The expression levels
of miR-122-5p, miR-181b-5p, miR-181c-5p, miR-181d-5p, miR-211-5p,
miR-370-5p, miR-493-5p, miR-506-5p, miR-590-5p, miR-642b-5p and
miR-654-3p in LINC01232-deficient Eca109 and KYSE150 cells were
detected by reverse transcription-quantitative poly-merase chain
reaction (RT-qPCR). (E) RT-qPCR was performed to detect miR-654-3p
expression in 55 pairs of ESCC tissues and matched normal tissues.
(F) Pearson's correlation coefficient illustrates the correlation
between LINC01232 and miR-654-3p in 55 ESCC tissues. (G) Luciferase
activity corresponding to LINC01232-wt or LINC01232-mut was
detected in Eca109 and KYSE150 cells following transfection with
the miR-654-3p mimic or NC mimic. (H) RT-qPCR was conducted to
determine the enrichment of LINC01232 and miR-654-3p in Eca109 and
KYSE150 cells after RNA immunoprecipitation. (I) Eca109 and KYSE150
cells were transfected with miR-654-3p mimic or NC mimic, and the
overexpression efficiency was determined by RT-qPCR. (J-M) CCK-8
assay, flow cytometric analysis, and migration and invasion assays
were performed to detect the proliferation, apoptosis, and
migration and invasion of miR-654-3p-overexpressing Eca109 and
KYSE150 cells. *P<0.05 and **P<0.01 vs.
NC mimic or si-ctrl. |
A StarBase 3.0 search predicted 21 miRNAs as
candidate targets of LINC01232 (Fig.
2C). Of these, miR-181a-5p (31), miR-21-5p (32) and miR-708-5p (33) have been found to be upregulated in
ESCC, while miR-28-5p has been shown to exhibit a stable expression
(34); therefore, these 4 miRNAs
were excluded from subsequent analyses. Of the remaining
candidates, miR-122-5p, miR-181b-5p, miR-181c-5p, miR-181d-5p,
miR-211-5p, miR-370-5p, miR-493-5p, miR-506-5p, miR-590-5p,
miR-642b-5p and miR-654-3p were selected, and their expression in
LINC01232-deficient Eca109 and KYSE150 cells was evaluated. RT-qPCR
analysis clearly affirmed the upregulation of miR-654-3p in Eca109
and KYSE150 cells following the silencing of LINC01232, whereas the
expression of the other miRNAs remained unaffected (Fig. 2D). Subsequently, miR-654-3p
expression was detected in 55 pairs of ESCC tissues and matched
normal tissues. The downregulation of miR-654-3p in ESCC tissues
(Fig. 2E) indicated a negative
correlation with LINC01232 expression (Fig. 2F; r=-0.6041, P<0.0001).
Additionally, luciferase reporter assay, which was performed to
examine the binding between miR-654-3p and LINC01232 in ESCC cells,
revealed a marked decrease in the luciferase activity of
LINC01232-wt in response to miR-654-3p upregulation in the Eca109
and KYSE150 cells, whereas miR-654-3p over-expression had no effect
on the luciferase activity associated with LINC01232-mut (Fig. 2G). Moreover, the considerable
enrichment of LINC01232 and miR-654-3p was observed in
AGO2-containing immunoprecipitation complexes when compared with
IgG immunoprecipitates in RIP assay (Fig. 2H).
Subsequently, to assess the roles of miR-654-3p in
ESCC cells, Eca109 and KYSE150 cells were transfected with
miR-654-3p mimic or NC mimic and subjected to RT-qPCR analyses to
detect miR-654-3p expression. Transfection with miR-654-3p mimic
notably increased the expression of miR-654-3p in both the Eca109
and KYSE150 cells (Fig. 2I). The
results of CCK-8 assay revealed that the upregulation of miR-654-3p
significantly hindered the proliferation of Eca109 and KYSE150
cells (Fig. 2J). The ectopic
expres-sion of miR-654-3p also markedly induced the apoptosis of
the Eca109 and KYSE150 cells (Fig.
2K). Furthermore, miR-654-3p overexpression led to a marked
decrease in the migration (Fig.
2L) and invasion (Fig. 2M) of
the Eca109 and KYSE150 cells. In summary, these results indicated
that miR-654-3p functioned as a cancer-inhibiting miRNA that was
sponged by LINC01232 in ESCC cells.
LINC01232 functions as a ceRNA to
positively regulate HDGF expression in ESCC cells by competitively
binding to miR-654-3p
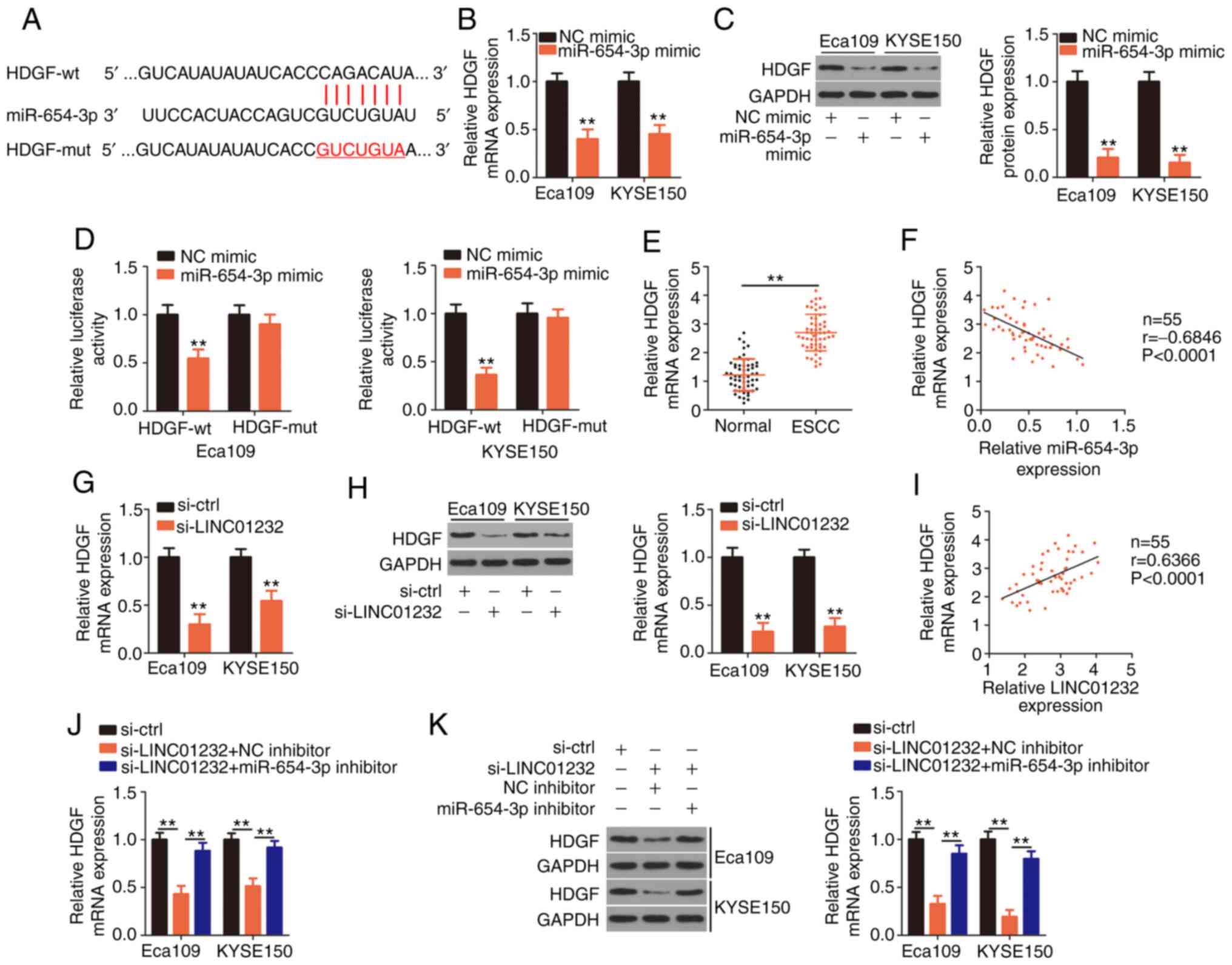
The TargetScan, miRDB and StarBase 3.0 databases
were searched to identify the putative targets of miR-654-3p. HDGF
(Fig. 3A) was selected for
further evaluation given its important role in the oncogenicity of
ESCC (35-37). In the present study, the results
of RT-qPCR and western blot analyses demonstrated that the
upregulation of miR-654-3p downregulated the expression of HDGF
mRNA (Fig. 3B) and protein
(Fig. 3C) in the Eca109 and
KYSE150 cells. To further verify these findings, HDGF-wt and
HDGF-mut reporter plasmids were generated and used in a luciferase
reporter assay. Notably, miR-654-3p mimic mark-edly blocked the
luciferase activity associated with HDGF-wt in the Eca109 and
KYSE150 cells. However, the luciferase activity of HDGF-mut
remained unaffected in response to miR-654-3p overexpression
(Fig. 3D). Furthermore, a
substantially upregulated HDGF mRNA expression was observed in ESCC
tissues compared with matched normal tissues (Fig. 3E), and this expression negatively
correlated with that of miR-654-3p (Fig. 3F; r=-0.6846, P<0.0001).
To investigate the association among LINC01232,
miR-654-3p and HDGF in ESCC cells, the HDGF mRNA and protein levels
in LINC01232-deficient Eca109 and KYSE150 cells were evaluated by
RT-qPCR and western blot analysis, respectively. The silencing of
LINC01232 led to a significant decrease in HDGF expression at the
mRNA (Fig. 3G) and protein level
(Fig. 3H) in the Eca109 and
KYSE150 cells. Notably, HDGF mRNA expression positively correlated
with LINC01232 expression in ESCC tissues (Fig. 3I; r=0.6366, P<0.0001). Rescue
experiments were also conducted using miR-654-3p inhibitor in
Eca109 and KYSE150 cells subjected to LINC01232 silencing.
LINC01232 knockdown induced decreases in the expression of both
HDGF mRNA (Fig. 3J) and protein
(Fig. 3K) in Eca109 and KYSE150
cells, which were restored by co-transfection with miR-654-3p
inhibitor. These results indicate that LINC01232 positively
regulates HDGF expression by competitively binding to miR-654-3p in
ESCC cells.
LINC01232 functions by targeting the
miR-654-3p/HDGF axis in ESCC cells
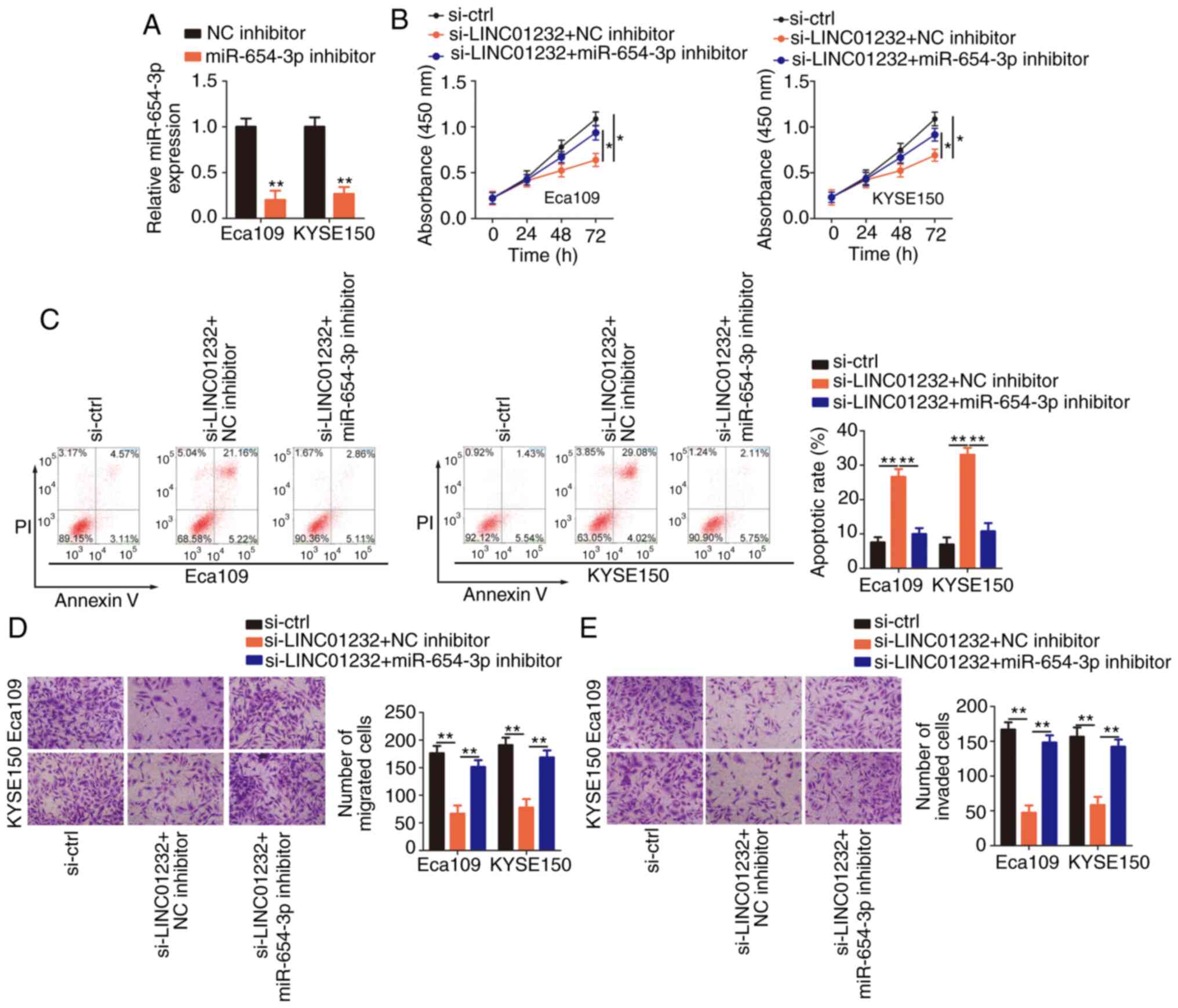
Rescue experiments were then performed to further
verify the effects of the LINC01232/miR-654-3p/HDGF pathway on the
malignant behaviors of ESCC cells. Briefly, si-LINC01232 and
miR-654-3p inhibitor or NC inhibitor were co-transfected into the
Eca109 and KYSE150 cells, which were then subjected to functional
assays. The efficiency of miR-654-3p inhibitor in the Eca109 and
KYSE150 cells was confirmed by RT-qPCR analysis (Fig. 4A). The inhibition of miR-654-3p
was sufficient to abolish the changes in Eca109 and KYSE150 cell
proliferation (Fig. 4B),
apoptosis (Fig. 4C), migration
(Fig. 4D) and invasion (Fig. 4E) induced by the silencing of
LINC01232.
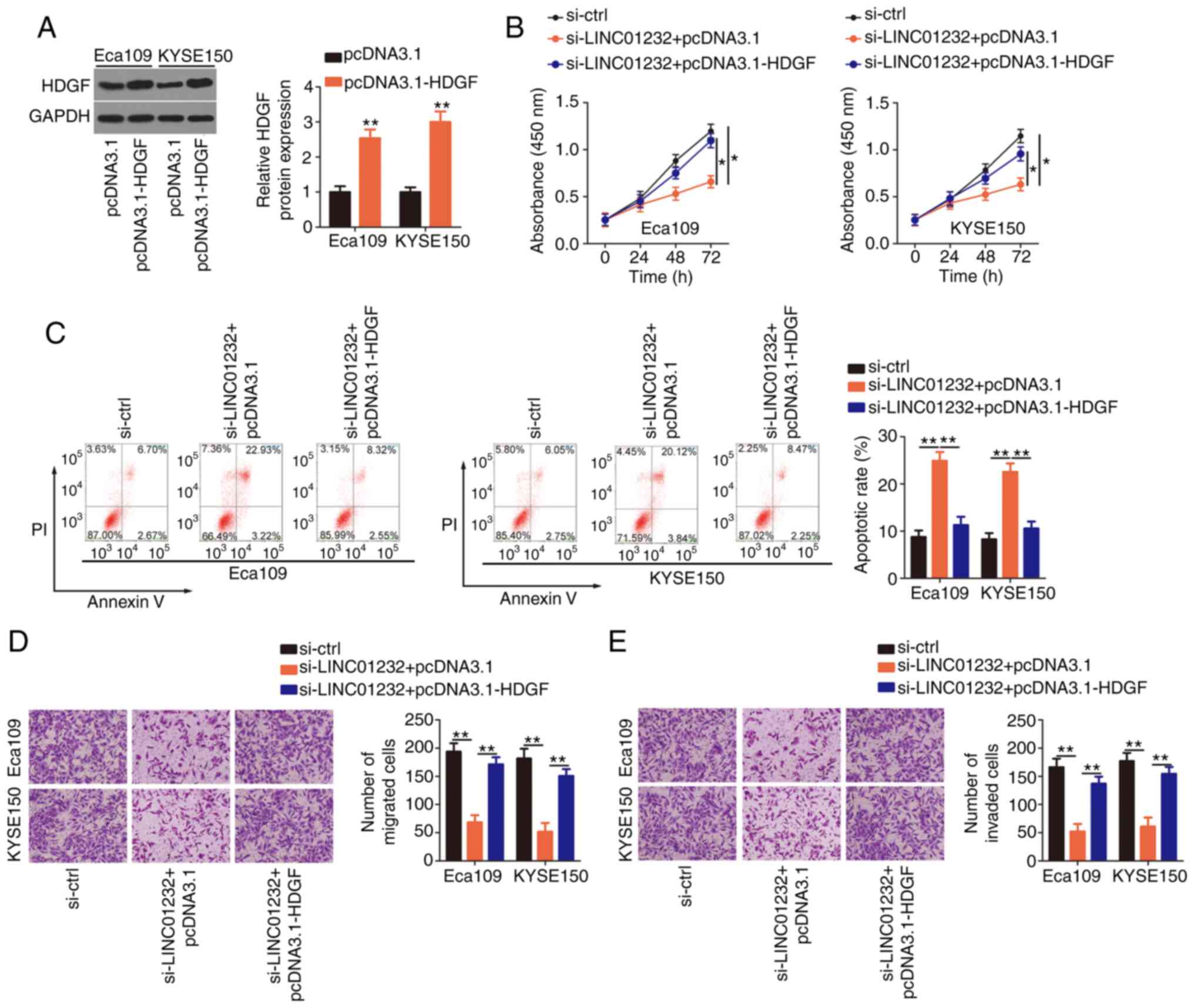
To examine whether HDGF is needed to achieve the
effects of LINC01232 on the malignant phenotype of ESCC cells,
either the HDGF-overexpressing plasmid pcDNA3.1-HDGF or the empty
pcDNA3.1 vector were co-transfected with si-LINC01232 into the
Eca109 and KYSE150 cells. The transfection efficiency of
pcDNA3.1-HDGF was determined by western blot analysis (Fig. 5A). Indeed, the overexpression of
HDGF partially reverse the impairment of Eca109 and KYSE150 cell
proliferation induced by LINC01232 knockdown (Fig. 5B). Additionally, the apoptosis of
Eca109 and KYSE150 cells induced by si-LINC01232 was partially
abolished by co-transfection with pcDNA3.1-HDGF (Fig. 5C). Furthermore, the upregulation
of HDGF restored the migratory (Fig.
5D) and invasive capabilities (Fig. 5E) of the Eca109 and KYSE150 cells,
which had been suppressed by LINC01232 silencing. Collectively,
these results indicate that LINC01232 regulates ESCC cell
activities at least partly by regulating the miR-654-3p/HDGF
axis.
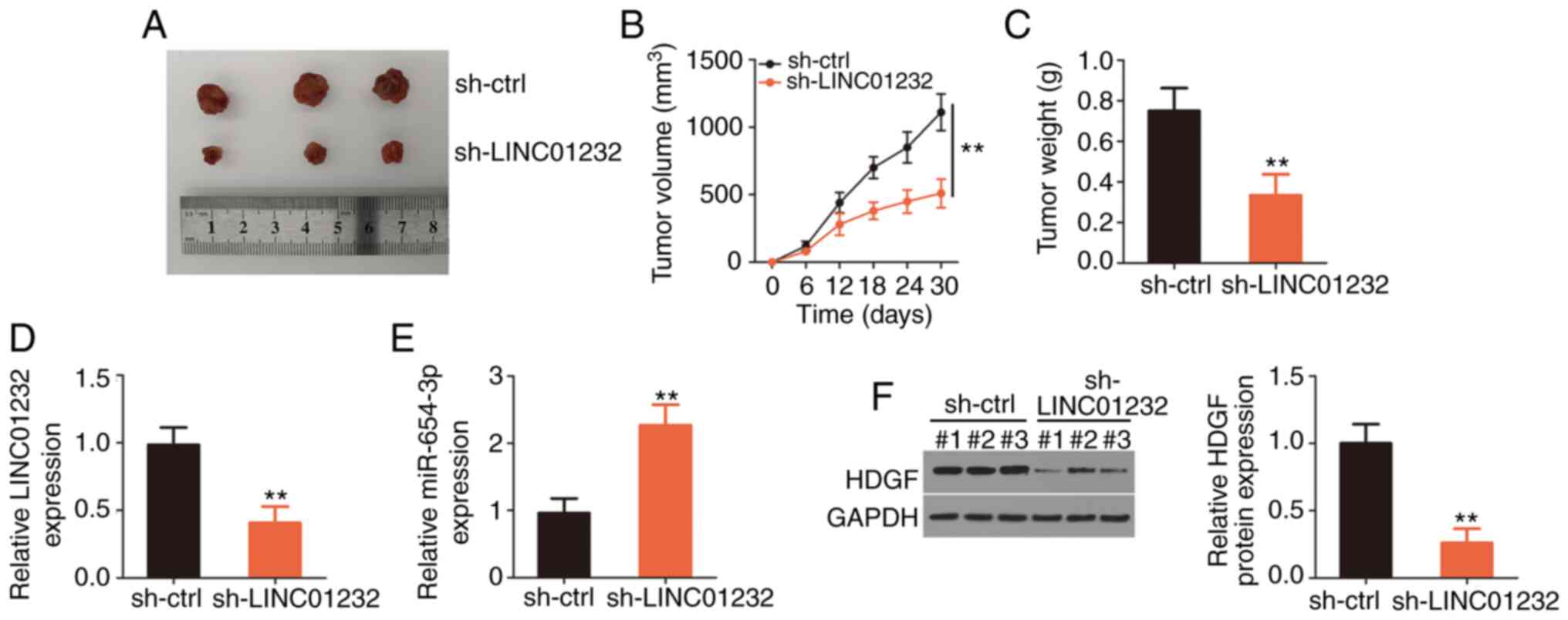
LINC01232 knockdown suppresses ESCC tumor
growth in vivo
Finally, the effects of LINC01232 on ESCC tumor
growth in vivo were examined using a xenograft mouse model.
Eca109 cells were stably transfected with sh-LINC01232 or sh-ctrl
and subcutaneously inoculated into nude mice. The subcutaneous
xenografts grew at a markedly slower rate in the si-LINC01232 group
than in the sh-ctrl group (Fig. 6A
and B). The weights of the tumor xenografts were notably lower
in the sh-LINC01232 group than in the sh-ctrl group (Fig. 6C). Following excision, the tumor
xenografts were subjected to RT-qPCR and western blot analyses to
determine the changes in the expression of LINC01232, miR-654-3p
and HDGF. RT-qPCR analysis revealed the downregulation of LINC01232
(Fig. 6D) and the upregulation of
miR-654-3p (Fig. 6E) in tumors
originating from the sh-LINC01232-transfected Eca109 cells. Western
blot analysis also revealed the decreased HDGF protein level in the
tumor xenografts collected from the sh-LINC01232 mice (Fig. 6F). On the whole, these results
demonstrate the inhibitory effects of LINC01232 knockdown on ESCC
tumor growth in vivo.
Discussion
Recent studies on lncRNAs have attracted increasing
attention due to the involvement of these molecules in cancer
oncogenesis and progression (38-40). Studies have reported the
expression of hundreds of lncRNAs in ESCC, and differentially
expressed lncRNAs have been shown to be closely related to ESCC
malignancy through their effects on a wide range of tumor behaviors
(41,42). Therefore, it is necessary to
clarify the functions of lncRNAs in ESCC and thus identify
effective diagnostic markers and treatment targets. In the present
study, the regulatory functions of LINC01232 in ESCC cells were
examined and the mechanisms through which this molecule affects the
malignant characteristics of this tumor type were investigated.
Using both in vitro and in vivo functional
experiments, the pro-oncogenic functions of LINC01232 and its
interactions with miR-654-3p and HDGF in ESCC were examined.
LINC01232 expression is upregulated in pancreatic
adenocarcinoma, and a high level of expression is associated with a
poor clinical outcome (26).
LINC01232 functions as an oncogenic lncRNA in pancreatic
adenocarcinoma and regulates cell proliferation, apoptosis,
migration, invasion and the epithelial-mesenchymal transition
(26). However, the expression
and roles of LINC01232 in ESCC have not yet been elucidated.
Herein, expression pattern analyses of lncRNAs in
ESCC were conducted using the TCGA and GTEx databases. LINC01232
was identified as one of the most dysregulated lncRNAs in ESCC.
Subsequently, RT-qPCR was performed to measure the expression of
LINC01232 in ESCC tissues and cell lines. LINC01232 was highly
expressed in ESCC tissues and cell lines, suggesting that this
molecule plays crucial roles in ESCC progression. However, the
expression high of LINC01232 was not found to be associated with
the overall survival or disease-free survival of patients with
ESCC. This finding suggests that LINC01232 has no potential to be
investigated as a prognostic marker for ESCC. Through a series of
functional in vitro experiments, it was determined that
LINC01232 played carcinogenic roles in ESCC and that LINC01232
interference suppressed cell proliferation, migration, and invasion
and promoted cell apoptosis. Moreover, a xenograft mouse model was
established and the cancer-promoting roles of LINC01232 in ESCC
tumor growth were examined in vivo.
In recent years, studies have increasingly
demonstrated that lncRNAs function as ceRNAs that may sponge miRNAs
to decrease their expression and activity, which would thereby
regulate the suppressive effects on miRNA targets (43-45). In the present study, lncLocator
analysis revealed that LINC01232 was mostly localized in the
cytoplasm, consistent with the results of the cell
cytosolic/nuclear fraction isolation experiment. This observation
suggests that LINC01232 interacts with miRNAs in the cytoplasm.
Initially, the target prediction algorithm indicated a potential
interaction between LINC01232 and miR-654-3p. Herein, RT-qPCR was
first used to confirm that LINC01232 silencing led to an increased
miR-654-3p expression in ESCC cells. Additionally, it was observed
that miR-654-3p and LINC01232 exhibited a negative correlation in
ESCC tissues. Furthermore, luciferase reporter assay revealed that
miR-654-3p could bind directly to the 'seed site' of LINC01232 in
ESCC cells. RIP assays revealed that both LINC01232 and miR-654-3p
were enriched in AGO2-containing immunoprecipitation complexes in
ESCC cells. Furthermore, LINC01232 controlled the expression of
HDGF, a direct target of miR-654-3p, by sponging miR-654-3p in ESCC
cells. Accordingly, the results of the present study indicate a
ceRNA process that involves LINC01232, miR-654-3p and HDGF in ESCC
cells.
miR-654-3p is aberrantly expressed in several types
of human cancers (46,47). However, its expression status and
roles in ESCC remain to be elucidated. In the present study, a weak
miR-654-3p expression was observed in ESCC and it was determined
that its upregulation suppressed the oncogenicity of ESCC cells.
Subsequent mechanistic analyses validated HDGF as a downstream
target of miR-654-3p in ESCC cells. HDGF, which is encoded by a
gene located on the q21-q23 region of chromosome 1, is highly
expressed in ESCC (36). Patients
with ESCC having a high HDGF expression manifest a poorer
disease-free and overall survival compared with patients with a low
HDGF expression (36).
Furthermore, HDGF is identified as an independent prognostic factor
for ESCC patients with early stage (36). Functionally, HDGF exerts
cancer-promoting roles, and contributes to tumor progression
through its effects on various aggressive processes (35-37). The results of the rescue
experiments reconfirmed that the effects of LINC01232 deficiency on
ESCC cells could be reversed by increasing the output of the
miR-654-3p/HDGF axis. The findings indicate that LINC01232
competitively binds to and acts as a molecular sponge for
miR-654-3p, which in turn activates HDGF and affects ESCC
malignancy.
LINC01232 is upregulated in ESCC and plays a
tumor-promoting role during cancer progression by sequestering
miR-654-3p to increase HDGF expression. Accordingly, the
LINC01232/miR-654-3p/HDGF pathway may provide a novel theoretical
basis for the management of ESCC.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
MZ and HC designed the study. All authors (MZ, HC,
BZ, ML and HM) performed the functional experiments. MZ and HC
wrote the manuscript. All authors reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Human clinical tissues were collected and used in
accordance with the Institutional Ethics Committee of the
Affiliated Hospital of Inner Mongolia University for the
Nationalities. Additionally, written informed consent was provided
by all patients before tissue collection. The animal experiments
were conducted with approval from the Institutional Animal Care and
Use Committee of Affiliated Hospital of Inner Mongolia University
for the Nationalities.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Funakawa K, Uto H, Sasaki F, Nasu Y,
Mawatari S, Arima S, Nakazawa J, Taguchi H, Hashimoto S, Kanmura S,
et al: Effect of endoscopic submucosal dissection for superficial
esophageal neoplasms and risk factors for postoperative stricture.
Medicine (Baltimore). 94:e3732015. View Article : Google Scholar
|
|
4
|
Smyth EC, Lagergren J, Fitzgerald RC,
Lordick F, Shah MA, Lagergren P and Cunningham D: Oesophageal
cancer. Nat Rev Dis Primers. 3:170482017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Y: Epidemiology of esophageal
cancer. World J Gastroenterol. 19:5598–5606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song Y, Li L, Ou Y, Gao Z, Li E, Li X,
Zhang W, Wang J, Xu L, Zhou Y, et al: Identification of genomic
alterations in oesophageal squamous cell cancer. Nature. 509:91–95.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bach DH and Lee SK: Long noncoding RNAs in
cancer cells. Cancer Lett. 419:152–166. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cao W, Liu JN, Liu Z, Wang X, Han ZG, Ji
T, Chen WT and Zou X: A three-lncRNA signature derived from the
Atlas of ncRNA in cancer (TANRIC) database predicts the survival of
patients with head and neck squamous cell carcinoma. Oral Oncol.
65:94–101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sarfi M, Abbastabar M and Khalili E: Long
noncoding RNAs biomarker-based cancer assessment. J Cell Physiol.
234:16971–16986. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun Y and Ma L: New insights into long
non-coding RNA MALAT1 in cancer and metastasis. Cancers (Basel).
11:2162019. View Article : Google Scholar
|
|
12
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Javed Z, Khan K, Sadia H, Raza S, Salehi
B, Sharifi-Rad J and Cho WC: LncRNA and Wnt signaling in colorectal
cancer. Cancer Cell Int. 20:3262020. View Article : Google Scholar
|
|
14
|
Tang WW, Wu Q, Li SQ, Tong YS, Liu ZH,
Yang TX, Xu Y and Cao XF: Implication of lncRNAs in pathogenesis of
esophageal cancer. Onco Targets Ther. 8:3219–3226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luo F, Wen Y, Zhou H and Li Z: Roles of
long non-coding RNAs in cervical cancer. Life Sci. 256:1179812020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Du F, Guo T and Cao C: Restoration of
UPK1A-AS1 expression suppresses cell proliferation, migration, and
invasion in esophageal squamous cell carcinoma cells partially by
sponging microRNA-1248. Cancer Manag Res. 12:2653–2662. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren P, Zhang H, Chang L, Hong XD and Xing
L: LncRNA NR2F1-AS1 promotes proliferation and metastasis of ESCC
cells via regulating EMT. Eur Rev Med Pharmacol Sci. 24:3686–3693.
2020.PubMed/NCBI
|
|
18
|
Liang X, Wu Z, Shen S, Niu Y, Guo Y, Liang
J and Guo W: LINC01980 facilitates esophageal squamous cell
carcinoma progression via regulation of miR-190a-5p/MYO5A pathway.
Arch Biochem Biophys. 686:1083712020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
21
|
Sharma P and Sharma R: miRNA-mRNA
crosstalk in esophageal cancer: From diagnosis to therapy. Crit Rev
Oncol Hematol. 96:449–462. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du G, Zhou J, Cheng L, Ma X, Gui Y and Tan
B: High expression of miR-206 predicts adverse outcomes: A
potential therapeutic target for esophageal cancer. Comb Chem High
Throughput Screen. 22:599–611. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang W, Fu S, Lin X, Zheng J, Pu J, Gu Y,
Deng W, Liu Y, He Z, Liang W and Wang C: miR-92b-3p functions as a
key gene in esophageal squamous cell cancer as determined by
co-expression analysis. Onco Targets Ther. 12:8339–8353. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu X, Wang M, Cao L, Cong L, Gao Y, Lu J,
Feng J, Shen B and Liu D: miR-4319 suppresses the growth of
esophageal squamous cell carcinoma via targeting NLRC5. Curr Mol
Pharmacol. 13:144–149. 2020. View Article : Google Scholar
|
|
25
|
Ye Y, Shen A and Liu A: Long non-coding
RNA H19 and cancer: A competing endogenous RNA. Bull Cancer.
106:1152–1159. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Q, Lei C, Lu C, Wang J, Gao M and Gao
W: LINC01232 exerts oncogenic activities in pancreatic
adenocarcinoma via regulation of TM9SF2. Cell Death Dis.
10:6982019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Bayoumi AS, Sayed A, Broskova Z, Teoh JP,
Wilson J, Su H, Tang YL and Kim IM: Crosstalk between long
noncoding RNAs and microRNAs in health and disease. Int J Mol Sci.
17:3562016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abdollahzadeh R, Daraei A, Mansoori Y,
Sepahvand M, Amoli MM and Tavakkoly-Bazzaz J: Competing endogenous
RNA (ceRNA) cross talk and language in ceRNA regulatory networks: A
new look at hallmarks of breast cancer. J Cell Physiol.
234:10080–10100. 2019. View Article : Google Scholar
|
|
30
|
Dong BS, Shi MJ, Su SB and Zhang H:
Insight into long noncoding competing endogenous RNA networks in
hepatic fibrosis: The potential implications for mechanism and
therapy. Gene. 687:255–260. 2019. View Article : Google Scholar
|
|
31
|
Xiang Z, Dong X, Sun Q, Li X and Yan B:
Clinical significance of up-regulated miR-181a in prognosis and
progression of esophageal cancer. Acta Biochim Biophys Sin
(Shanghai). 46:1007–1010. 2014. View Article : Google Scholar
|
|
32
|
Xia Y, Wang Y, Wang Q, Ghaffar M, Wang Y,
Sheng W and Zhang F: Increased miR-203-3p and reduced miR-21-5p
synergistically inhibit proliferation, migration, and invasion in
esophageal cancer cells. Anticancer Drugs. 30:38–45. 2019.
View Article : Google Scholar
|
|
33
|
Yang H, Su H, Hu N, Wang C, Wang L, Giffen
C, Goldstein AM, Lee MP and Taylor PR: Integrated analysis of
genome-wide miRNAs and targeted gene expression in esophageal
squa-mous cell carcinoma (ESCC) and relation to prognosis. BMC
Cancer. 20:3882020. View Article : Google Scholar
|
|
34
|
Chen L, Jin Y, Wang L, Sun F, Yang X, Shi
M, Zhan C, Shi Y and Wang Q: Identification of reference genes and
miRNAs for qRT-PCR in human esophageal squamous cell carcinoma. Med
Oncol. 34:22017. View Article : Google Scholar
|
|
35
|
Matsuyama A, Inoue H, Shibuta K, Tanaka Y,
Barnard GF, Sugimachi K and Mori M: Hepatoma-derived growth factor
is associated with reduced sensitivity to irradiation in esophageal
cancer. Cancer Res. 61:5714–5717. 2001.PubMed/NCBI
|
|
36
|
Yamamoto S, Tomita Y, Hoshida Y, Morii E,
Yasuda T, Doki Y, Aozasa K, Uyama H, Nakamura H and Monden M:
Expression level of hepatoma-derived growth factor correlates with
tumor recurrence of esophageal carcinoma. Ann Surg Oncol.
14:2141–2149. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bao CH, Wang XT, Ma W, Wang NN, Nesa EU,
Wang JB, Wang C, Jia YB, Wang K, Tian H and Cheng YF: Irradiated
fibroblasts promote epithelial-mesenchymal transition and HDGF
expression of esophageal squamous cell carcinoma. Biochem Biophys
Res Commun. 458:441–447. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang Z, Zhou JK, Peng Y, He W and Huang
C: The role of long noncoding RNAs in hepatocellular carcinoma. Mol
Cancer. 19:772020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mozdarani H, Ezzatizadeh V and Rahbar
Parvaneh R: The emerging role of the long non-coding RNA HOTAIR in
breast cancer development and treatment. J Transl Med. 18:1522020.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Khajehdehi M, Khalaj-Kondori M, Ghasemi T,
Jahanghiri B and Damaghi M: Long noncoding RNAs in gastrointestinal
cancer: Tumor suppression versus tumor promotion. Dig Dis Sci Mar.
17:2020.Epub ahead of print. View Article : Google Scholar
|
|
41
|
Feng Q, Zhang H, Yao D, Chen WD and Wang
YD: Emerging role of non-coding RNAs in esophageal squamous cell
carcinoma. Int J Mol Sci. 21:2582019. View Article : Google Scholar
|
|
42
|
Talebi A, Masoodi M, Mirzaei A,
Mehrad-Majd H, Azizpour M and Akbari A: Biological and clinical
relevance of metastasis-associated long noncoding RNAs in
esophageal squamous cell carcinoma: A systematic review. J Cell
Physiol. 235:848–868. 2020. View Article : Google Scholar
|
|
43
|
Wang JJ, Huang YQ, Song W, Li YF, Wang H,
Wang WJ and Huang M: Comprehensive analysis of the lncRNA
associated competing endogenous RNA network in breast cancer. Oncol
Rep. 42:2572–2582. 2019.PubMed/NCBI
|
|
44
|
Pan H, Guo C, Pan J, Guo D, Song S, Zhou Y
and Xu D: Construction of a competitive endogenous RNA network and
identification of potential regulatory axis in gastric cancer.
Front Oncol. 9:9122019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liang W and Sun F: Identification of
pivotal lncRNAs in papillary thyroid cancer using lncRNA-mRNA-miRNA
ceRNA network analysis. PeerJ. 7:e74412019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang J, Zhang Z, Chen S, Dou W, Xie R and
Gao J: miR-654-3p predicts the prognosis of hepatocellular
carcinoma and inhibits the proliferation, migration, and invasion
of cancer cells. Cancer Biomark. 28:73–79. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li P, Cai JX, Han F, Wang J, Zhou JJ, Shen
KW and Wang LH: Expression and significance of miR-654-5p and
miR-376b-3p in patients with colon cancer. World J Gastrointest
Oncol. 12:492–502. 2020. View Article : Google Scholar : PubMed/NCBI
|