Introduction
The processes that induce the restoration of injured
tissues immediately after wound healing comprise three interrelated
dynamic phases, namely, the inflammatory, proliferative and
regeneration phases (1,2). A number of studies have demonstrated
that the primary resident fibroblasts in the dermis are the key
players that maintain skin homeostasis during wound healing
(3-5). Dermal fibroblasts migrate to the
wound site and are induced to proliferate by a variety of growth
factors, including platelet-derived growth factor (PDGF), epidermal
growth factor (EGF) and transforming growth factor β (3,6,7).
Therefore, the migration and proliferation of fibroblasts need to
be precisely regulated during wound healing.
Previous studies have reported that a variety of
chemokines are expressed at the wound site, indicating the
important role of chemokines in recruiting inflammatory cells to
the wound site (6,8-10).
Notably, the monocyte chemoattractant protein-1, also referred to
as C-C motif chemokine ligand 2 (CCL2), is one of the chemokines
that mediate the infiltration of monocytes and macrophages during
wound healing (11-15). Additionally, treatment with CCL2
improves wound healing by enhancing the recruitment of myeloid
cells in db/db mice, a Toll-like receptor 3-deficient mouse
model in which the secretion of CCL2 is impaired (16,17). Therefore, understanding the role
of CCL2 is important for aiding wound healing and developing novel
therapeutic agents.
The erythroid differentiation regulator 1 (Erdr1) is
a highly conserved autocrine factor that can induce the synthesis
of hemoglobin in both human and murine erythroleukemia (18). Erdr1 is expressed in various
tissues, including the placenta, liver, brain, lung, intestine,
bone marrow, thymus, sebaceous glands, vessels, nerves, normal
human epidermis and human keratinocytes (18-20). A number of studies have reported
that Erdr1 exhibits anticancer effects in various types of cancer,
including gastric cancer and melanoma. For example, Erdr1 is an
antimetastatic factor that is negatively regulated by IL-18 via
downregulation of the expression of heat shock protein 90 in
melanoma (21). Additionally,
recombinant Erdr1 has been reported to suppress the invasiveness
and motility of gastric cancer cells via the JNK pathway (19). Erdr1 exhibits a therapeutic
potential for various inflammatory diseases, including psoriasis,
rosacea, hair loss disorders and rheumatoid arthritis (20,22-26). We therefore hypothesized that
Erdr1 may promote the migration and proliferation of fibroblasts
involved in wound healing. The mechanisms underlying the
therapeutic effects of Erdr1 in wound healing are yet to be
elucidated. The present study aimed to investigate the effects of
Erdr1 on wound healing.
Materials and methods
Mice and cell culture
Female BALB/c mice (7-week-old; weight, 20-22 g)
were purchased from orient Bio, Inc. All experiments were performed
following the ethical guidelines of the Korea university
Institutional Animal Care and use Committee (Seoul, Korea; approval
no. KUIACUC-2018-0045). Human dermal fibroblasts (HDFs; Biosolution
Co., Ltd.) were cultured in a mixture (3:1) of DMEM (Thermo Fisher
scientific, Inc) and F-12K with 10% fetal bovine serum (Capricorn
scientific GmbH), 100 u/ml penicillin and 0.1 mg/ml streptomycin
(Thermo Fisher scientific, Inc.) in an incubator at 37°C with 5%
CO2. The passage number was <13 for all
experiments.
Preparation of recombinant proteins
The recombinant mouse Erdr1 protein was prepared as
previously reported (19,21). Briefly, the Erdr1-pCMv-SPORT6
plasmid was purchased from open Biosystems, Inc. The region of the
coding sequence was transferred into the bacterial expression
plasmid pET22B (Merck KGaA). The 177 amino acid encoded Erdr1
protein was expressed in the Escherichia coli Top10 system
(Invitrogen; Thermo Fisher scientific, Inc.), purified using a
Ni-NTA purification system according to the manufacturer's
instructions (Invitrogen; Thermo Fisher scientific, Inc.),
quantified by Pierce™ BCA protein assay kit (Thermo Fisher
scientific, Inc.), separated by 10% SDS-PAGE and visualized by
Coomassie blue staining (sigma-Aldrich; Merck KGaA) (purity
>95%). The endotoxin level of the purified protein (<0.1
EU/ml) was measured using the LAL system (Associated of Cape Cod
International, Inc.). The recombinant human EGF protein (purity
>98%) was purchased from PeproTech, Inc.
Reagents and antibodies
Antibodies against tubulin (cat. no. sc-69969),
JNK1/2 (cat. no. sc-571), phosphorylated (p-) JNK1/2 (cat. no.
sc-6254), ERK1/2 (cat. no. sc-153), p-ERK1/2 (cat. no. sc-7383) and
p38 (cat. no. sc-535) were purchased from Santa Cruz Biotechnology,
Inc. Rabbit-HRP (cat. no. 7074), mouse-HRP (cat. no. 7076) and
p-p38 (cat. no. 9216) antibodies were purchased from Cell signaling
Technology, Inc. Human CCL2 ELISA pair set (cat. no. SEK10134) was
purchased from Sino Biological, Inc. Pluronic® F-127
(cat. no. P2443) for the preparation of 22% (w/v) hydrogel was
purchased from sigma Aldrich; Merck KGaA and dissolved in saline
overnight on the rotator at 4°C. Inhibitors for ERK (u0126; cat.
no. 662005), p38 (SB203580; cat. no. S8307) and JNK (sP600125; cat.
no. S5567) were purchased from Merck KGaA.
Proliferation assay
HDFs were seeded and pre-cultured into 96-well
plates at a density of 5×103 cells/well for 24 h at 37°C
in a 5% CO2 incubator. Subsequently, the cells were
cultured without serum for 16 h at 37°C and pre-treated with 1, 10
or 25 µg/ml mitomycin-C (MMC) or PBs for 1 h at 37°C. HDFs
were washed and treated with 1, 10 or 50 ng/ml Erdr1 or EGF as a
positive control for 24 h at 37°C in serum-free culture medium. To
evaluate the effects of the CCL2-neutralizing antibody (Sino
Biological, Inc.) on cell proliferation, HDFs were treated with 50
ng/ml Erdr1 or PBs for 24 h following pretreatment with 1, 10 or 30
µg/ml CCL2-neutralizing antibody. After 24 h, the cells were
treated with 20 µl of 100 µg/ml
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
solution for 2 h, and the medium was removed. The precipitated
formazan was dissolved in 200 µl DMSO, and the absorbance at
560 nm was determined by a microplate reader.
In vitro migration assay
HDFs were seeded into 12-well plates at a density of
8×104 cells/well and pre-cultured for 24 h, followed by
culture with serum-free medium for 16 h at 37°C in 5%
Co2 incubator. The monolayers of HDFs at 80-90%
confluency were scratched with a 200-µl pipette tip, and the
plates were washed with PBs to remove the cell debris. HDFs were
pre-treated with 1, 10 or 25 ng/ml MMC for 1 h at 37°C, washed and
stimulated with 50 ng/ml Erdr1 or 50 ng/ml EGF. To evaluate the
effects of the CCL2-neutralizing antibody on cell migration, HDFs
were treated with 50 ng/ml Erdr1 or PBs for 24 h following
pretreatment with 1, 10 or 30 µg/ml CCL2-neutralizing
antibody. After 24 h, the cells were fixed with 4% paraformaldehyde
for 10 min, permeabilized with 0.1% Triton x-100 in PBs for 5 min
and incubated with Rhodamine phalloidin for 1 h at room
temperature. Fluorescence images (magnification, ×100) were
acquired using an olympus I× 71 fluorescence microscope (olympus
Corporation).
Wound healing assay in vivo
Mouse dorsal hair was shaved and wiped with 70%
ethanol before wounding. The mice were anesthetized with 65%
N2, 30% O2 and 5% isoflurane and maintained
with 2% isoflurane while a 5-mm wound puncture was performed. HG
was not used for delivering the compounds to the wound sites after
4 days. The wounds were directly treated with saline, 1 µg
EGF or 1 µg Erdr1 in 22% hydrogel (HG) once a day for 5
days. The wound closure was monitored by capturing images at the
same position with a fixed height stand to ensure consistent image
size. Measurement of wound area in the images was performed using
ImageJ software (version 1.45; National Institutes of Health). The
analysis of area measurement was performed by one person who was
blinded to the grouping of the mice and the sampling day for each
wound being measured to avoid measurement bias, and the percentage
of wound size was calculated as follows (4,27):
wound closure (%)=(wound area on day 0-wound area on day of
post-wounding)/wound area on day 0×100%.
Reverse transcription (RT)-PCR
HDFs were stimulated with 1, 10, 25, 50 or 100 ng/ml
Erdr1 or EGF for 1, 3, 6, 9 or 12 h, and total RNA was extracted
using a RiboEx total RNA kit (GeneAll Biotechnology Co., Ltd.).
Total RNA (1 µg) was reverse-transcribed into cDNA by the
AccuPower® RT Premix kit (BioNeer Corporation) according
to the manufacturer's instructions. The cDNAs were amplified with
specific primers by PCR using Genie™ 32 Thermal Block (BioNeer
Corporation). The specific primer sequences were as follows: CCL2,
sense, 5′-CCTTGCCTTGCTGCTCTACC-3′ and antisense,
5′-CCTATGTGCTGGCCTTGGTG-3′; and GAPDH sense,
5′-ACATCAAGAAGGTGGTGAAG-3′ and antisense,
5′-ATTCAAGAGAGTAGGGAGGG-3′. The PCR products were separated on 1.5%
agarose gels and stained using the StainingSTAR solution (Dynebio,
Inc.).
ELISA
Human dermal fibroblasts were seeded at
2×105 cells/well into 6-well plates and cultured
overnight in serum-free medium for 24 h at 37°C in a 5%
Co2 incubator. The cells were subsequently treated with
1, 10 or 50 ng/ml EGF or Erdr1 for 24 h at 37°C. CCL2 protein
levels were determined using a CCL2 matched ELISA kit (cat. no.
SEK10134; Sino Biological, Inc.) according to the manufacturer's
protocol. The absorbance at 450 nm was measured by a microplate
reader.
Western blot analysis
Whole cell lysates were prepared using RIPA lysis
and extraction buffer (Thermo Fisher scientific, Inc.), and the
protein concentration was quantified by BCA assay kit (Thermo
Fisher scientific, Inc.). A total of 30 µg of samples per
lane were separated on 10% SDS-PAGE gels and transferred to a PVDF
membrane. The membrane was blocked with 5% skim milk (Neogen
Corporation) in 1× TBS-T (0.1% Tween-20) buffer. The membrane was
washed twice with 1× TBS and incubated with the primary antibodies
(1:1,000) in 5% BSA for 2 h at room temperature (sigma-Aldrich;
Merck KGaA)-TBS buffer. The membrane was washed six times with
TBS-T buffer and incubated for 1 h at room temperature with the
corresponding HRP-conjugated mouse or rabbit IgG secondary
antibodies (1:5,000) in 5% skim milk-TBS. The target proteins were
visualized using Amersham ECL Prime western blotting Detection
Regent (Cytiva).
Histologic analysis of wounds
Mice (n=5 mice/group) were sacrificed and skin
tissues were harvest at day 6 post-wounding. Cross-sections (5
µm) of paraffin-embedded skin tissues were stained with
hematoxylin for 5 min and eosin for 2 min (H&E). Stained
tissues were mounted with mounting media (Thermo Fisher scientific,
Inc.) and observed under light microscopy (x100 magnification). The
granulation tissue was identified by the presence of tissue matrix,
immune cells, vascular tissue and fibroblasts, and the formation of
granulation tissue area was defined as the area between underneath
the neo-epithelium and above the subcutaneous fat tissue. The
relative granulation tissue area ratios were calculated on the
digital images using ImageJ based on the wound closure area, area
of dermis and epidermis.
Statistical analysis
The data are presented as the mean ± standard
deviation. All experiments were repeated at least three times
independently. Statistical differences between the control and the
treated groups were assessed by one-way analysis of variance
followed by Tukey's post hoc test using SPSS 22 software (IBM
Corp.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Erdr1 promotes wound repair in vivo
To examine the effects of Erdr1 on wound closure,
the wounds were treated with Erdr1-, EGF-, or saline-HG and
analyzed. As demonstrated in Fig.
1A, the sizes of wounds in the Erdr1-HG or EGF-HG-treated mice
were decreased between days 4 and 10 compared with those in the
mice that were treated with saline-HG. The wound closure of the
mice treated with Erdr1-HG or saline-HG was ~100% on day 10
(Fig. 1B). As previous studies
have reported that the formation of granulation tissue in the early
stage of a wound is an important step for supporting
re-epithelialization and dermal reconstitution (28,29), the tissues from the wound site
were collected on day 6 and stained with H&E for histological
analysis; the results demonstrated that the formation of
granulation tissues in the group that was treated with Erdr1-HG was
similar to that of the group treated with EGF-HG and higher
compared with that in the group treated with saline-HG (Fig. 1C). These results demonstrated that
the promotion of wound healing of the Erdr1-HG-treated group was
similar to that of EGF-HG treated group.
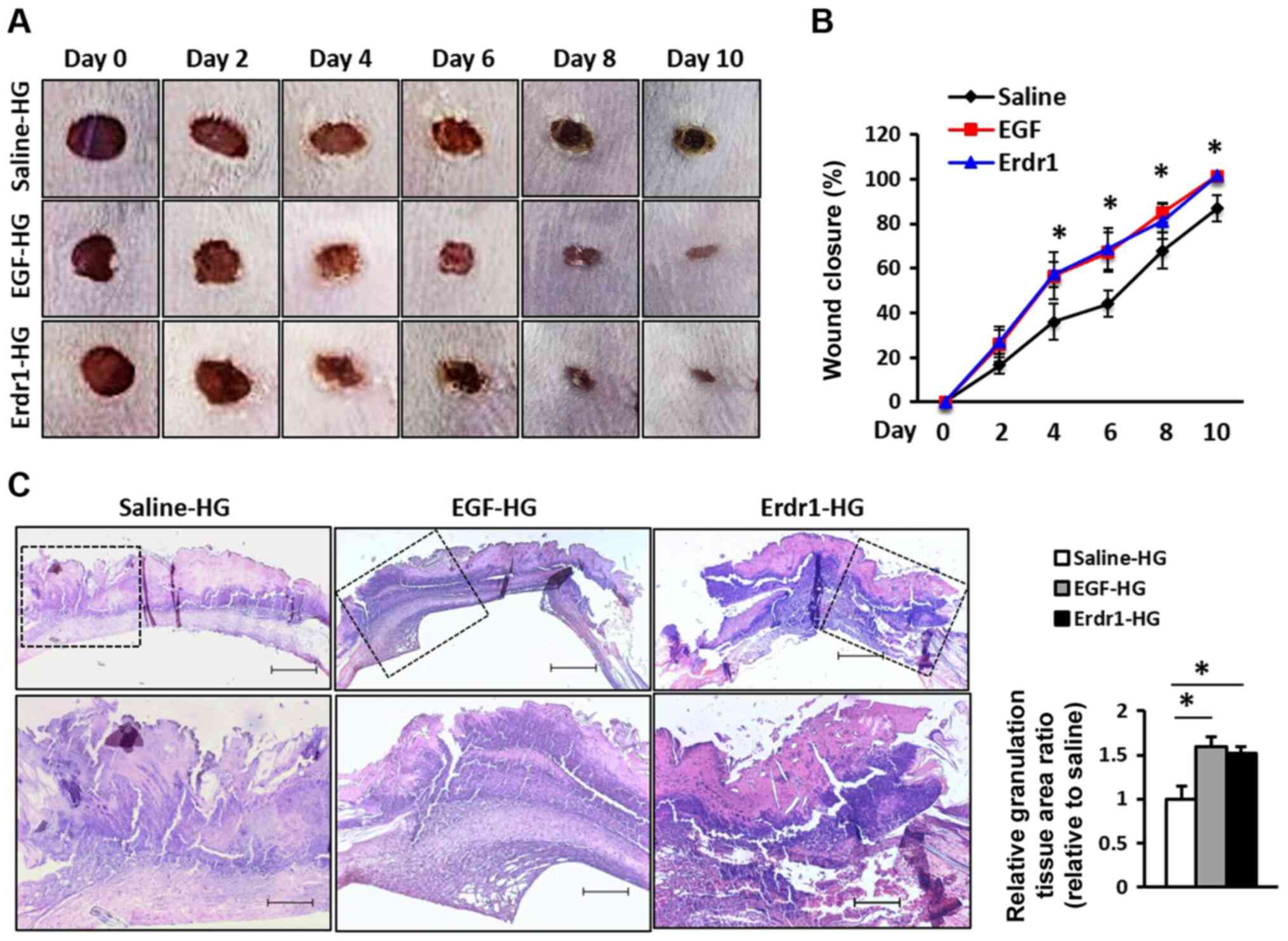 | Figure 1Treatment with Erdr1 accelerates
wound healing in vivo. (A) Representative images of the
wounds in mice treated with Erdr1-HG, EGF-HG or control saline-HG
every 24 h for 5 days. The images represent the status of wound
repair during 10 days of wound healing. (B) The wound closure in
mice treated with saline-HG, Erdr1-HG, or EGF-HG was measured
digitally on the indicated days after the infliction of injury, and
the wound closure was calculated as a percentage. (C) Tissues
obtained from the wound sites on day 6 after inflicting the wounds
were stained with H&E, and the formation of granulation tissues
was analyzed. The relative granulation in the tissues was
calculated and compared with that of the group treated with
saline-HG. Magnification, ×40 (upper panel) and ×100× (lower
panel); scale bar, 200 µm. n=6. *P<0.01 vs.
Saline-HG. Erdr1, erythroid differentiation regulator 1; EGF,
epidermal growth factor; HG, hydrogel. |
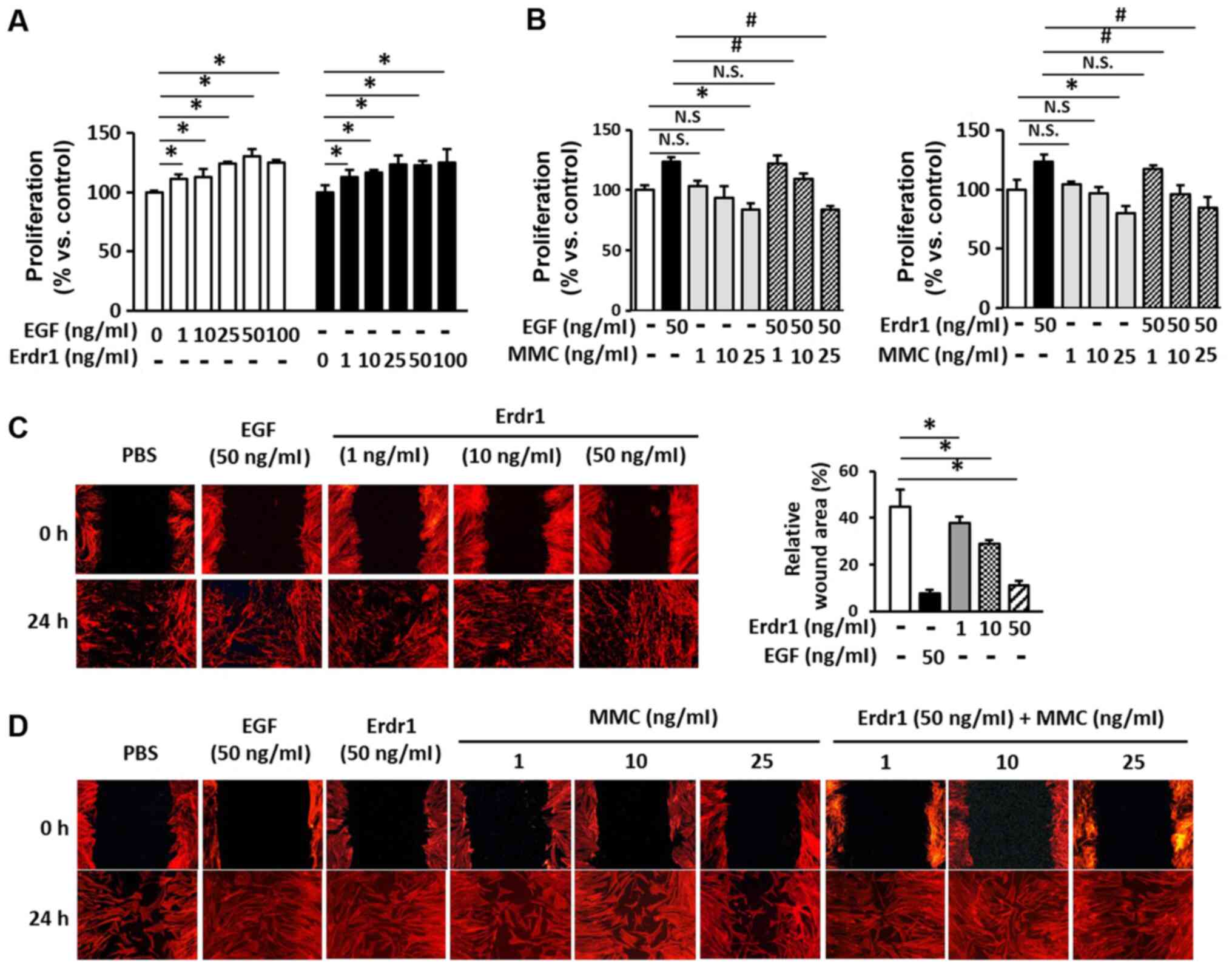
Erdr1 induces HDF proliferation and
migration
To investigate the effects of Erdr1 on the
proliferation and migration of HDFs, MTT and wound healing assays
were performed. As demonstrated in Fig. 2A, 25-100 ng/ml Erdr1 significantly
increased the proliferation of HDFs by ~25% compared with that of
the PBs-treated control group. The proliferation of the HDFs that
were treated with Erdr1 was comparable to that of the HDFs that
were treated with 50 ng/ml EGF in the positive control group. To
exclude the effects of Erdr1 on the proliferation of HDFs while
evaluating the Erdr1-induced migration of HDF wound healing,
subsequent assays were performed in the presence of MMC to block
cell proliferation. As demonstrated in Fig. 2B, 25 µg/ml MMC completely
inhibited cell proliferation, which was not observed at
concentrations of 1 and 10 µg/ml. In addition, the
proliferation of HDFs induced by 50 ng/ml EFG or 50 ng/ml Erdr1 was
decreased by MMC (Fig. 2B).
The results of the wound healing assay demonstrated
that Erdr1 increased the migration of HDFs (Fig. 2C). To further examine the effects
of Erdr1 on the migration of HDFs, the wound healing assay was
performed in the presence of MMC; as presented in Fig. 2D, 1 and 10 µg/ml MMC did
not inhibit the migration of Erdr1-treated HDFs. However, 25
µg/ml MMC slightly inhibited the Erdr1-induced migration of
HDFs (Fig. 2D). These results
suggested that Erdr1 may promote wound healing by inducing the
proliferation and migration of HDFs.
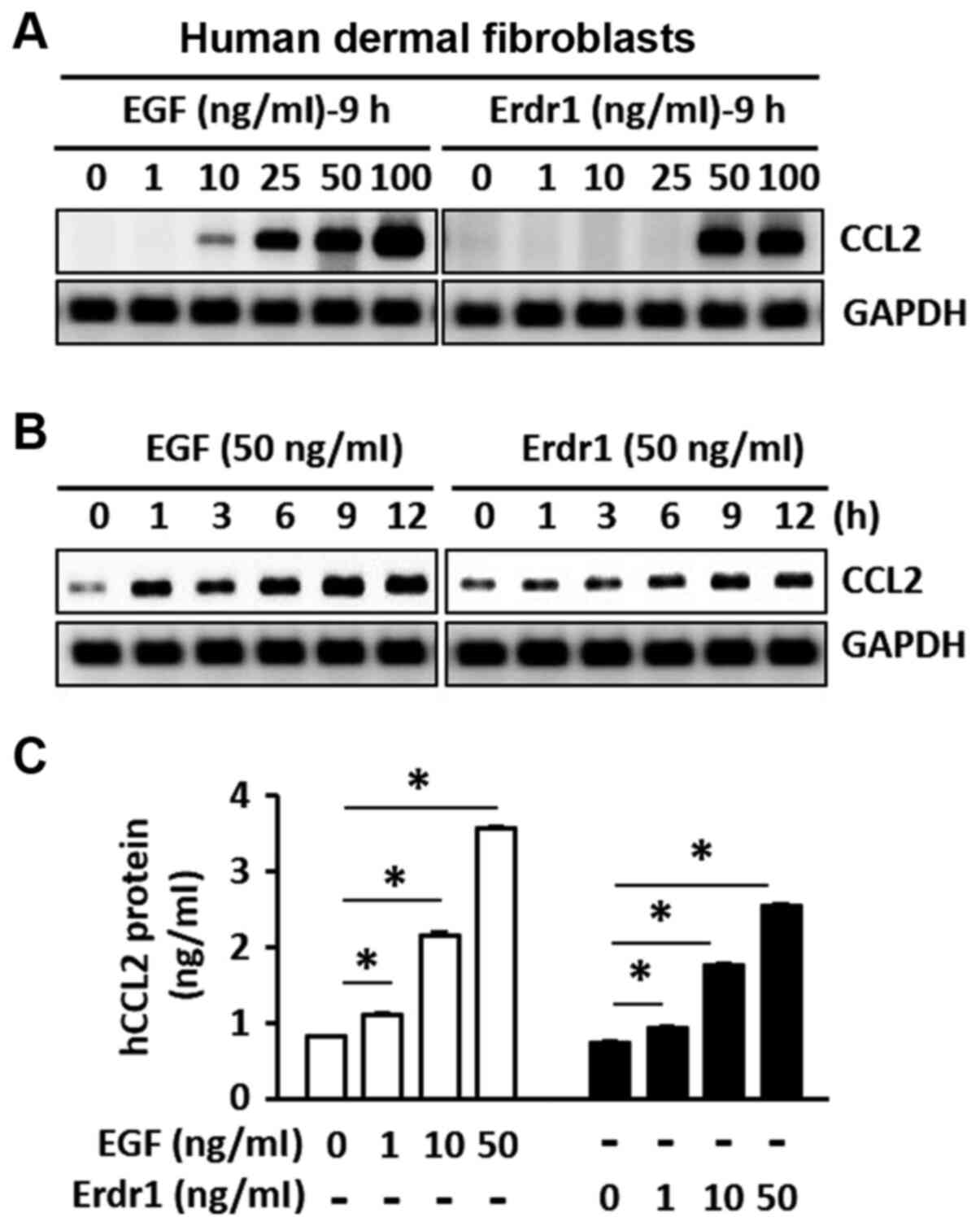
Erdr1 increases the production of CCL2 in
HDFs
To examine the effects of Erdr1 on the expression of
CCL2, HDFs were cultured with various concentrations of Erdr1 or
EGF (positive control) for different durations. The mRNA and
protein levels of CCL2 were determined using RT-PCR and ELISA,
respectively. As presented in Fig.
3A, treatment with 50 and 100 ng/ml Erdr1 increased the mRNA
expression levels of CCL2. In addition, the mRNA expression of CCL2
induced by 50 ng/ml Erdr1 appeared to increase over time (Fig. 3B). In order to confirm the effects
of Erdr1 on the production of CCL2, ELISA was performed to detect
the production of CCL2 in HDFs following stimulation with Erdr1 or
EGF. As demonstrated in Fig. 3C,
following HDF stimulation with Erdr1, the secretion of CCL2
significantly increased compared with that in the control cells.
These results suggested that the Erdr1-induced production of CCL2
may serve a crucial role in wound healing in HDFs.
Erdr1 induces CCL2 production via the MAP
kinase pathway in HDFs
Previous studies have demonstrated that the MAP
kinase signaling pathway serves an important role in the migration
and proliferation of skin fibroblasts and keratinocytes (5,30-33). Therefore, the effects of Erdr1 on
the activation of the ERK1/2, p38 and JNK1/2 MAP kinases in HDFs
were assessed in the present study. As demonstrated in Fig. 4A, treatment with Erdr1 induced the
phosphorylation of ERK1/2, p38 and JNK1/2, and the phosphorylation
of MAP kinases induced by Erdr1 at 5 min was comparable to that in
the EGF-treated HDFs (Fig.
4A).
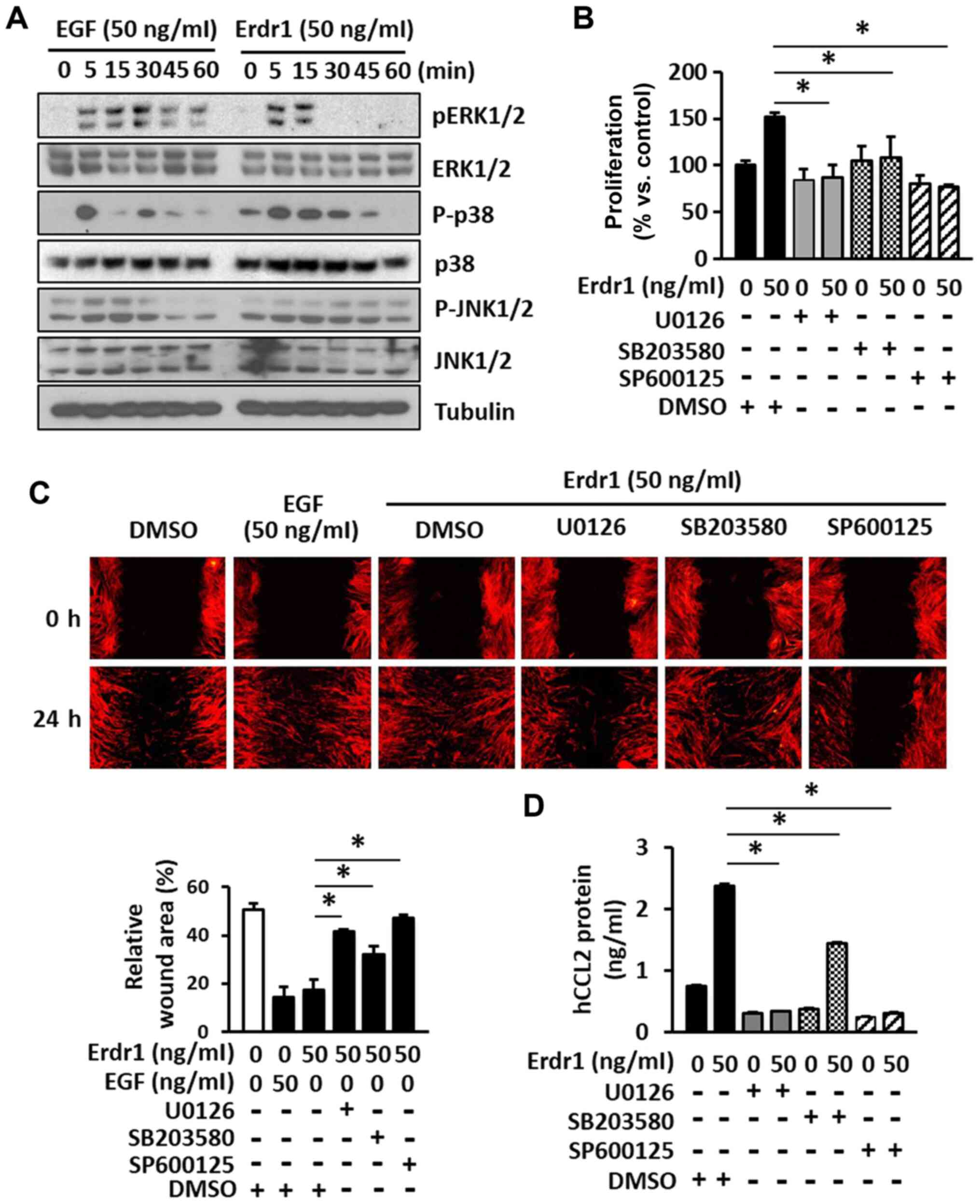 | Figure 4Erdr1 enhances the proliferation and
migration of HDFs by inducing the production of CCL2 via the
activation of MAPK kinases. (A) The levels of ERK1/2, p-ERK1/2,
p38, p-p38, JNK1/2 and p-JNK1/2 in HDFs treated with 50 ng/ml
Erdr1, EGF or PBs for 5, 15, 30, 45 and 60 min were analyzed by
western blotting. Tubulin was used as the loading control. The (B)
proliferation and (C) migration of HDFs treated with 50 ng/ml Erdr1
or DMSO (control) for 24 h after pretreatment with the ERK1/2
inhibitor u0126 (20 µM), the p38 inhibitor sB203580 (20
µM) or the JNK1/2 inhibitor sP600125 (20 µM) for 1 h
was determined using the MTT and wound healing assays,
respectively. n=6. (D) The levels of CCL2 protein were determined
in HDFs pre-treated with 20 µM u0126, sB203580 or sP600125
for 1 h and subsequently treated with 50 ng/ml Erdr1 for 24 h using
ELISA. n=6. *P<0.01 vs. Erdr1. Erdr1, erythroid
differentiation regulator 1; EGF, epidermal growth factor; HDFs,
human dermal fibroblasts; CCL2, C-C motif chemokine ligand 2. |
In order to further investigate the role of the
Erdr1-induced phosphorylation of ERK1/2, p38, and JNK1/2 in the
proliferation and migration of HDFs, specific inhibitors of ERK1/2
(u0126), p38 (sB203580) and JNK1/2 (sP600125) were used. As
presented in Fig. 4B and C, the
proliferation and migration of HDFs induced by Erdr1 were
significantly inhibited by the specific inhibitors of ERK1/2, p38
and JNK1/2 MAP kinases. To determine whether these MAP kinases were
involved in the Erdr1-induced production of CCL2, HDFs were
pretreated with specific MAP kinase inhibitors, and the production
of CCL2 protein was measured by ELISA. The results demonstrated
that the MAP kinase inhibitors significantly inhibited the
production of CCL2 in Erdr1-treated HDFs compared with that in the
Erdr1-HG and DMSO-treated group (Fig.
4D), consistent with the results of the proliferation and
migration assays. Taken together, these results suggested that the
Erdr1-induced production of CCL2 enhanced the proliferation and
migration of HDFs by phosphorylating ERK1/2, p38 and JNK1/2.
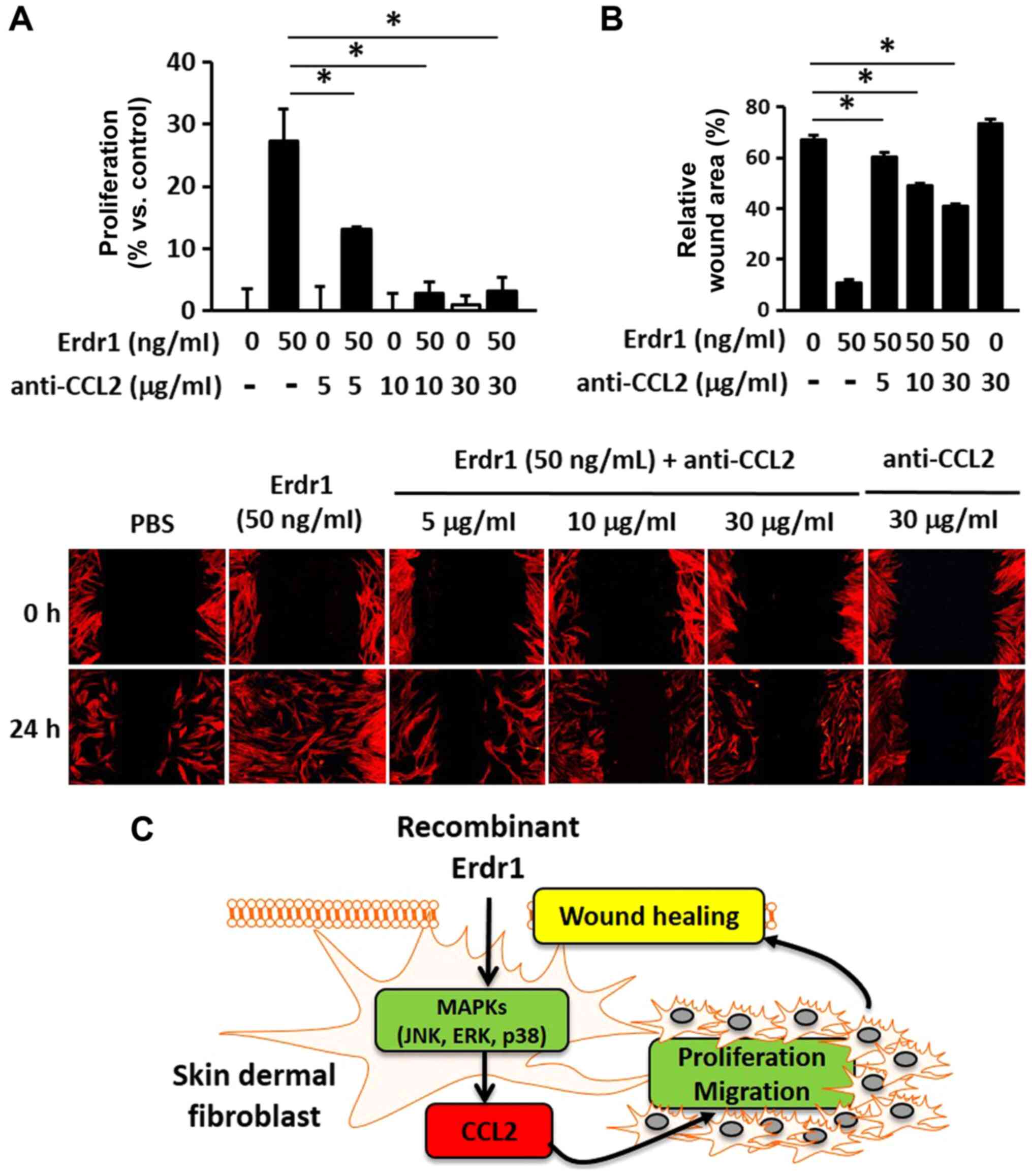
Involvement of CCL2 in Erdr1-mediated
wound healing
To determine the role of the Erdr1-induced
production of CCL2 in wound healing, HDFs were treated with a
CCL2-neutralizing antibody. As presented in Fig. 5A, the CCL2-neutralizing antibody
inhibited the proliferation of HDFs in the Erdr1-treated group.
Furthermore, the migration of the cells treated with only
CCL2-neutralizing antibody did not increase compared with that in
the negative control (PBs-treated) group (Fig. 5B). Taken together, these results
suggested that Erdr1 promoted wound healing by inducing the
production of CCL2 via the activation of the ERK1/2, p38 and JNK1/2
MAP kinase signaling pathways (Fig.
5C).
Discussion
Wound healing is an intricate, interactive
biological process (34,35). Successful wound healing involves
inflammation, tissue formation and remodeling (2,5,36).
The various behaviors of skin cells, including the migration and
proliferation of fibroblasts, are crucial events for wound
contraction (2,5,36,37). Additionally, dermal fibroblasts
serve an important role in process of wound healing via
extracellular modulation (2,5,36,37). The migration and proliferation of
dermal fibroblasts serve an essential role in tissue regeneration
and repair; fibroblasts and keratinocytes are involved in the
proliferation, migration and deposition of the extracellular matrix
as well as the formation of granulation tissue (2,6,8).
Erdr1 is expressed in various tissues and cells, including the
normal human epidermis, CD4+ T cells, skin tumors and
human keratinocytes (18-20,38). Our previous study demonstrated
that the sequence of human Erdr1 identified from the cDNA from
human tissues exhibited 100% homology with the sequence of Erdr1
from murine tissues (21).
However, the sequence of human Erdr1 was not identified in the
present study; the sequence of human Erdr1 will be analyzed in
future studies. Our recent study reported that Erdr1 is a potential
therapeutic agent for inflammatory skin diseases including
psoriasis (25), rosacea
(26) and hair loss disorders
(39); therefore, we hypothesized
in the present study that Erdr1 may also promote wound healing.
In the present study, topical administration of
Erdr1-HG enhanced the healing of skin wounds and induced complete
wound closure after 10 days of treatment in vivo. It was
observer that tissue repair was more rapid in the wounds that were
treated with Erdr1 compared with that in the wounds treated with
saline, and the wound healing effects of Erdr1 were comparable to
those of EGF-HG in vivo. Notably, in the present study, the
skin wounds in vivo were treated with saline, Erdr1 or EGF
in HG at a final concentration of 20% (w/v); HG was used in these
experiments as it has been used in several studies for delivering
therapeutic agents to wound sites (40-42). However, HG was unable to penetrate
the wound site owing to the formation of a fibrin clot, which
inhibited the penetration of the compounds when the skin wounds
were treated with saline, Erdr1 or EGF after 4 days of inflicting
the wounds. Therefore, HG was not used for delivering the compounds
to the wound sites after 4 days. It is necessary to modify the
composition of the HG to improve the penetrative efficiency and
targeted delivery of the compounds to the wound site in future
studies.
It has been demonstrated that the role of the MAP
kinase pathway in inflammatory skin diseases is mediated by
inflammatory cytokines, such as IL-22 and IL-23, and several
cellular functions, such as fibroblast and keratinocyte migration
(5,31). The results of the present study
demonstrated that the ERK, p38 and JNK pathways served an important
role in promoting the migration and proliferation of HDFs, and were
consistent with the results of several previous studies (5,28,31). Therefore, the role of the MAP
kinase pathway in the migration, proliferation and CCL2 production
was evaluated in the present study. The results demonstrated that
Erdr1 served an important role in the migration and proliferation
of HDFs via the activation of the ERK1/2, p38 and JNK1/2 signaling
pathways. Although the levels of p-ERK1/2 and p-p38 began to
decrease or disappeared after 5 min of EGF and Erdr1 treatments,
this did not represent a reduction in CCL2 production. In addition,
certain MAPK pathway inhibitors suppressed Erdr1-induced cell
proliferation, CCL2 production and migration at 24 h.
Chemokines are chemoattractants that recruit
monocytes, neutrophils and lymphocytes by binding to
G-protein-coupled receptors (6,14,17). CCL2 is one of the key chemokines
that regulate the migration and infiltration of monocytes and
macrophages (43,44). Recent studies have demonstrated
that several cytokines are expressed at the site of the wound and
serve an essential role in the recruitment of inflammatory cells to
the wound site (6,11,14). Furthermore, the expression of
chemokine receptors on the resident cells suggests that chemokines
contribute to re-epithelialization, tissue remodeling, and
angiogenesis (10). In
particular, MCP-1/CCL2 is a major chemoattractant that participates
in the infiltration of inflammatory cells, including monocytes and
macrophages, to the wound site (9-11,13,14,16,17,44). These studies suggest that CCL2 is
directly involved in wound healing. The production of CCL2 is
induced by oxidative stress, cytokines or growth factors in a
variety of cell types, including endothelial, epithelial, smooth
muscle and mesangial cells, astrocytes, monocytes, microglial cells
and fibroblasts (45). It has
been demonstrated that CCL2 is involved in wound healing and
various diseases, including multiple sclerosis, rheumatoid
arthritis, atherosclerosis, insulin-resistant diabetes (46-48).
A previous study has reported that treatment with a
CCL-2-neutralizing antibody suppresses the number of macrophages at
the wound site (15). In the
present study, treatment with the CCL2-neutralizing antibody
decreased in the migration and proliferation of HDFs. Furthermore,
Erdr1 significantly induced the production of CCL2 by activating
ERK1/2, p38 and JNK1/2 in HDFs.
The results of the present study have several
clinical limitations. First, Erdr1 is a novel factor for wound
healing that significantly increased the CCL2 production in HDFs.
Thus, we suggest that Erdr1 may be used as a therapeutic agent with
other growth factors such as EGF for synergistic improvement of
wound repair. Second, undefined receptors of Erdr1 should be
investigated for increasing the therapeutic potential of Erdr1 to
promote tissue repair. Third, it would be interesting to assess the
effect of topical application of Erdr1 in chronic/diabetic wounds
in future studies.
In conclusion, the results of the present study
suggested that Erdr1 accelerated wound healing by increasing the
production of CCL2, which induced the migration and proliferation
of fibroblasts via the activation of the ERK1/2, p38 and JNK1/2
signaling pathways in vitro. Therefore, Erdr1 may serve as a
potential therapeutic target for wound healing and for the
development of therapeutic agents for topical treatment of
wounds.
Acknowledgments
No applicable.
Funding
This study was supported by the Creative Materials
Discovery Program through the National Research Foundation of Korea
(grant no. 2016M3D1A1021387) and the National Research Foundation
of Korea (grant no. NRF-2018R1A2B6008434), and a grant from Kine
Sciences.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
BCL conceived and designed the experiments. BCL, JS
and AL performed the experiments. BCL and TSK wrote the manuscript.
BCL, DC and TSK analyzed data. BCL and TSK were responsible for
overall study design and supervised the project. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were performed following the ethical
guidelines of the Korea university Institutional Animal Care and
use Committee (Seoul, Korea; approval no. KUIACUC-2018-0045).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Singer AJ and Clark RA: Cutaneous wound
healing. N Engl J Med. 341:738–746. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Darby IA, Laverdet B, Bonte F and
Desmouliere A: Fibroblasts and myofibroblasts in wound healing.
Clin Cosmet Investig Dermatol. 7:301–311. 2014.PubMed/NCBI
|
|
3
|
Walraven M, Gouverneur M, Middelkoop E,
Beelen RH and Ulrich MM: Altered TGF-ß signaling in fetal
fibroblasts: What is known about the underlying mechanisms? Wound
Repair Regen. 22:3–13. 2014. View Article : Google Scholar
|
|
4
|
Lee BC, Song J, Lee A, Cho D and Kim TS:
Visfatin promotes wound healing through the activation of ERK1/2
and JNK1/2 pathway. Int J Mol Sci. 19:36422018. View Article : Google Scholar :
|
|
5
|
Kanazawa S, Fujiwara T, Matsuzaki S,
Shingaki K, Taniguchi M, Miyata S, Tohyama M, Sakai Y, Yano K,
Hosokawa K and Kubo T: bFGF regulates PI3-kinase-Rac1-JNK pathway
and promotes fibroblast migration in wound healing. PLoS One.
5:e122282010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barrientos S, Stojadinovic O, Golinko MS,
Brem H and Tomic-Canic M: Growth factors and cytokines in wound
healing. Wound Repair Regen. 16:585–601. 2008. View Article : Google Scholar
|
|
7
|
Lichtman MK, Otero-Vinas M and Falanga V:
Transforming growth factor beta (TGF-ß) isoforms in wound healing
and fibrosis. Wound Repair Regen. 24:215–222. 2016. View Article : Google Scholar
|
|
8
|
Raja, Sivamani K, Garcia MS and Isseroff
RR: Wound re-epithelialization: Modulating keratinocyte migration
in wound healing. Front Biosci. 12:2849–2868. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding J and Tredget EE: The role of
chemokines in fibrotic wound healing. Adv Wound Care (New
Rochelle). 4:673–686. 2015. View Article : Google Scholar
|
|
10
|
Rees PA, Greaves NS, Baguneid M and Bayat
A: Chemokines in wound healing and as potential therapeutic targets
for reducing cutaneous scarring. Adv Wound Care (New Rochelle).
4:687–703. 2015. View Article : Google Scholar
|
|
11
|
DiPietro LA, Polverini PJ, Rahbe SM and
Kovacs EJ: Modulation of JE/MCP-1 expression in dermal wound
repair. Am J Pathol. 146:868–875. 1995.PubMed/NCBI
|
|
12
|
Wetzler C, Kampfer H, Stallmeyer B,
Pfeilschifter J and Frank S: Large and sustained induction of
chemokines during impaired wound healing in the genetically
diabetic mouse: Prolonged persistence of neutrophils and
macrophages during the late phase of repair. J Invest Dermatol.
115:245–253. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Low QE, Drugea I A, Duffner LA, Quinn DG,
Cook DN, Rollins BJ, Kovacs EJ and DiPietro LA: Wound healing in
MIP-1alpha(-/-) and MCP-1(-/-) mice. Am J Pathol. 159:457–463.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Engelhardt E, Toksoy A, Goebeler M, Debus
S, Brocker EB and Gillitzer R: Chemokines IL-8, GROalpha, mcp-1,
IP-10, and Mig are sequentially and differentially expressed during
phase-specific infiltration of leukocyte subsets in human wound
healing. Am J Pathol. 153:1849–1860. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dipietro LA, Reintjes MG, Low QE, Levi B
and Gamelli RL: Modulation of macrophage recruitment into wounds by
monocyte chemoattractant protein-1. Wound Repair Regen. 9:28–33.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wood S, Jayaraman V, Huelsmann EJ, Bonish
B, Burgad D, Sivaramakrishnan G, Qin S, DiPietro LA, Zloza A, Zhang
C and Shafikhani SH: Pro-inflammatory chemokine CCL2 (MCP-1)
promotes healing in diabetic wounds by restoring the macrophage
response. PLoS One. 9:e915742014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin Q, Fang D, Fang J, Ren X, Yang X, Wen
F and Su SB: Impaired wound healing with defective expression of
chemokines and recruitment of myeloid cells in TLR3-deficient mice.
J Immunol. 186:3710–3717. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dormer P, Spitzer E, Frankenberger M and
Kremmer E: Erythroid differentiation regulator (EDR), a novel,
highly conserved factor I. Induction of haemoglobin synthesis in
erythroleukaemic cells. Cytokine. 26:231–242. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jung MK, Houh YK, Ha S, Yang Y, Kim D, Kim
TS, Yoon SR, Bang SI, Cho BJ, Lee WJ, et al: Recombinant Erdr1
suppresses the migration and invasion ability of human gastric
cancer cells, SNU-216, through the JNK pathway. Immunol Lett.
150:145–151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee YB, Kim HJ, Jung HY, Park YG, Kim SY,
Cho BK, Cho D and Park HJ: Downregulation of erythroid
differentiation regulator 1 as a novel marker of skin tumors. Int J
Dermatol. 53:723–730. 2014. View Article : Google Scholar
|
|
21
|
Jung MK, Park Y, Song SB, Cheon SY, Park
S, Houh Y, Ha S, Kim HJ, Park JM, Kim TS, et al: Erythroid
differentiation regulator 1, an interleukin 18-regulated gene, acts
as a metastasis suppressor in melanoma. J Invest Dermatol.
131:2096–2104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Houh YK, Kim KE, Park HJ and Cho D: Roles
of erythroid differentiation regulator 1 (Erdrl) on inflammatory
skin diseases. Int J Mol Sci. 17:20592016. View Article : Google Scholar
|
|
23
|
Kim KE, Kim S, Park S, Houh Y, Yang Y,
Park SB, Kim S, Kim D, Hur DY, Kim S, et al: Therapeutic effect of
erythroid differentiation regulator 1 (Erdrl) on collagen-induced
arthritis in DBA/1J mouse. Oncotarget. 7:76354–76361. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim KE, Houh Y, Lee J, Kim S, Cho D and
Park HJ: Downregulation of erythroid differentiation regulator 1
(Erdrl) plays a critical role in psoriasis pathogenesis. Exp
Dermatol. 25:570–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim KE, Houh Y, Park HJ and Cho D:
Therapeutic effects of erythroid differentiation regulator 1 on
imiquimod-induced psoriasis-like skin inflammation. Int J Mol Sci.
17:2442016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim M, Kim KE, Jung HY, Jo H, Jeong SW,
Lee J, Kim CH, Kim H, Cho D and Park HJ: Recombinant erythroid
differentiation regulator 1 inhibits both inflammation and
angiogenesis in a mouse model of rosacea. Exp Dermatol. 24:680–685.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Asai J, Takenaka H, Hirakawa S, Sakabe JI,
Hagura A, Kishimoto S, Maruyama K, Kajiya K, Kinoshita S, Tokura Y
and Katoh N: Topical simvastatin accelerates wound healing in
diabetes by enhancing angiogenesis and lymphangiogenesis. Am J
Pathol. 181:2217–2224. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matsubayashi Y, Ebisuya M, Honjoh S and
Nishida E: ERK activation propagates in epithelial cell sheets and
regulates their migration during wound healing. Curr Biol.
14:731–735. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Coutelle O, Hornig-Do HT, Witt A, Andree
M, Schiffmann LM, Piekarek M, Brinkmann K, Seeger JM, Liwschitz M,
Miwa S, et al: Embelin inhibits endothelial mitochondrial
respiration and impairs neoangiogenesis during tumor growth and
wound healing. EMBO Mol Med. 6:624–639. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Satish L, Babu M, Tran KT, Hebda PA and
Wells A: Keloid fibroblast responsiveness to epidermal growth
factor and activation of downstream intracellular signaling
pathways. Wound Repair Regen. 12:183–192. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wahedi HM, Park YU, Moon EY and Kim SY:
Juglone ameliorates skin wound healing by promoting skin cell
migration through Rac1/Cdc42/PAK pathway. Wound Repair Regen.
24:786–794. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chrissouli S, Pratsinis H, Velissariou V,
Anastasiou A and Kletsas D: Human amniotic fluid stimulates the
proliferation of human fetal and adult skin fibroblasts: The roles
of bFGF and PDGF and of the ERK and Akt signaling pathways. Wound
Repair Regen. 18:643–654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang WG, Ye L, Patel G and Harding KG:
Expression of WAVEs, the WASP (Wiskott-Aldrich syndrome protein)
family of verprolin homologous proteins in human wound tissues and
the biological influence on human keratinocytes. Wound Repair
Regen. 18:594–604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Aya KL and Stern R: Hyaluronan in wound
healing: Rediscovering a major player. Wound Repair Regen.
22:579–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Eming SA, Martin P and Tomic-Canic M:
Wound repair and regeneration: Mechanisms, signaling, and
translation. Sci Transl Med. 6:265sr2662014. View Article : Google Scholar
|
|
36
|
Rosinczuk J, Taradaj J, Dymarek R and
Sopel M: Mechanoregulation of wound healing and skin homeostasis.
Biomed Res Int. 2016:39434812016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rosique RG, Rosique MJ and Farina Junior
JA: Curbing inflammation in skin wound healing: A review. Int J
Inflam. 2015:3162352015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Soto R, Petersen C, Novis CL, Kubinak JL,
Bell R, Stephens WZ, Lane TE, Fujinami RS, Bosque A, O'Connell RM
and Round JL: Microbiota promotes systemic T-cell survival through
suppression of an apoptotic factor. Proc Natl Acad Sci USA.
114:5497–5502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Woo YR, Hwang S, Jeong SW, Cho DH and Park
HJ: Erythroid differentiation regulator 1 as a novel biomarker for
hair loss disorders. Int J Mol Sci. 18:3162017. View Article : Google Scholar :
|
|
40
|
Zhang M and Zhao X: Alginate hydrogel
dressings for advanced wound management. Int J Biol Macromol.
162:1414–1428. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Francesko A, Petkova P and Tzanov T:
Hydrogel dressings for advanced wound management. Curr Med Chem.
25:5782–5797. 2018. View Article : Google Scholar
|
|
42
|
Dhaliwal K and Lopez N: Hydrogel dressings
and their application in burn wound care. Br J Community Nurs.
23(Suppl 9): S24–S27. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu B, Rutledge BJ, Gu L, Fiorillo J,
Lukacs NW, Kunkel SL, North R, Gerard C and Rollins BJ:
Abnormalities in monocyte recruitment and cytokine expression in
monocyte chemoattractant protein 1-deficient mice. J Exp Med.
187:601–608. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Werner S and Grose R: Regulation of wound
healing by growth factors and cytokines. Physiol Rev. 83:835–870.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Craig MJ and Loberg RD: CCL2 (Monocyte
Chemoattractant Protein-1) in cancer bone metastases. Cancer
Metastasis Rev. 25:611–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Standiford TJ, Kunkel SL, Phan SH, Rollins
BJ and Strieter RM: Alveolar macrophage-derived cytokines induce
monocyte chemoattractant protein-1 expression from human pulmonary
type II-like epithelial cells. J Biol Chem. 266:9912–9918.
1991.PubMed/NCBI
|
|
47
|
Kusano KF, Nakamura K, Kusano H, Nishii N,
Banba K, Ikeda T, Hashimoto K, Yamamoto M, Fujio H, Miura A, et al:
Significance of the level of monocyte chemoattractant protein-1 in
human atherosclerosis. Circ J. 68:671–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sartipy P and Loskutoff DJ: Monocyte
chemoattractant protein 1 in obesity and insulin resistance. Proc
Natl Acad Sci USA. 100:7265–7270. 2003. View Article : Google Scholar : PubMed/NCBI
|