Introduction
Esophageal carcinoma (EC) is one of the most common
gastrointestinal tumors with two main histological subtypes:
Esophageal squamous cell carcinoma (ESCC) and esophageal
adenocarcinoma (EAC). In East Asia, the incidence of EC is known to
be the highest worldwide (1).
Although much progress has been made recently towards EC treatment
with respect to targeted therapy, surgery and neoadjuvant
chemotherapy, the 5-year survival rate of patients with locally
advanced tumors remains <55% (2). Thus, optimal and effective
therapeutic strategies are required.
MicroRNAs (miRNAs or miRs) are highly conserved
non-coding small RNAs that are capable of regulating gene
expression via the translational repression and degradation
induction of mRNAs in cells (3),
and have been shown to be involved in the major signaling pathways
of histogenesis and cell apoptosis (4). Moreover, miRNAs can serve as
biomarkers for tumor staging and prognosis. miR-93-5p is a paralog
(miR-106b-25) derived from the miR-17-92 cluster. It is implicated
in the occurrence and development of various human solid cancers,
including breast cancer, colorectal cancer, liver cancer, lung
cancer, ovarian cancer and pancreatic cancer, etc. (5). Some studies have demonstrated the
abnormally high expression of miR-93-5p in liver (6), breast (7) and lung cancer (8), and it is able to promote cell
proliferation and migration by binding to various target genes.
Moreover, it has been reported that miR-93-5p may be a potential
biomarker for the detection of the presence of cancer (9).
The transforming growth factor-β (TGF-β) pathway is
a pivotal player in cell carcinogenesis and metastasis. TGF-β
receptor 2 (TGFβR2) is a key molecule that regulates the TGF-β
pathway and its expression is often downregulated or lost in
several cancer (10). The
decrease in TGFβR2 expression can result in a variety changes in
tumor behavior, such as poor tumor differentiation, higher tumor
staging and an increase in the lymph node metastasis rate (11). In tumor cells, TGFβR2 signaling
can regulate a variety of activities, such as
epithelial-mesenchymal transition (EMT), cell migration and
invasion, angiogenesis, immune regulation and cytokine secretion
(12). The present study explored
the regulatory effects of miR-93-5p on TGFβR2 in EC cells, with an
aim to provide novel targeted diagnostic and prognostic approaches
for EC.
Materials and methods
Bioinformatics analysis
miRNA expression profiles, including 13 normal
samples and 176 EC tissue samples, and mRNA expression profiles,
including 11 normal samples and 160 EC tissue samples, were
obtained from the TCGA-ESCA database (https://portal.gdc.cancer.gov/). The R package 'edgeR'
was employed to identify the differentially expressed miRNAs
(DEmiRNAs) with the criteria of |logFC|>2 and adj. P-value
<0.05. Target genes for miR-93-5p were predicted through
bioinformatics analysis and 3 target prediction databases, miRDB
(http://mirdb.org/miRDB/index.html),
miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/index.php) and
TargetScan (http://www.targetscan.org/vert_71/). Survival analysis
was then conducted to identify the mRNA of interest. Based on the
downloaded data, the t-test was used to determine the significance
of the expression of these genes in normal tissues and tumor
tissues. The samples were divided into the high and low expression
groups based on the median expression of each gene, and survival
analysis was performed using the 'survival' package, respectively,
and genes that were significantly related to the prognosis and
targeted downregulation were selected as the target gene. Finally,
the targeted binding sites between miR-93-5p and its target gene
were predicted.
Cells and cell culture
The human normal esophageal cell line, Het-1A
(BNCC337688), and EC cell lines [TE-1 (BNCC100151), Eca-109
(BNCC337687) and EC9706 (BNCC339892)] were obtained from Bena
Culture Collection (BNCC, Beijing, China). The Het-1A, Eca-109 and
EC9706 cell lines were grown in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.) containing 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin. The TE-1 cell lines were cultured in RPMI-1640 medium
containing 10% FBS. All of the cells were placed in a wet incubator
with 5% CO2 at 37°C.
Cell transfection and vector
construction
The EC cells in logarithmic growth phase were
collected for transfection with miR-93-5p mimic, miR-93-5p
inhibitor and miR-93-5p mimic + oe-TGFβR2 as well as their
corresponding controls (obtained from Guangzhou RiboBio Co., Ltd.)
using Lipofectamine® 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), respectively. The transfection
concentration was 50 nM. Following 6 h of incubation with 5%
CO2 at 37°C, the cells were then continuously cultured
in fresh medium for 48 h for use in subsequent experiments. The
lentiviral vector was used to construct the TGFβR2 overexpression
vector: TGFβR2 cDNA with restriction enzyme sites of KpnI
and XhoI, as well as a corresponding control sequence
(designed by BLOCK-iT™ RNAi Deshgner website) was synthesized, and
then ligated into the lentiviral expression vector pLVX-IRES-neo
(Clontech Laboratories, Inc.) by T4 ligase. Following 24 h of
culture at 37°C, the expression vector was extracted and sequenced.
Finally, the packaged vector and viral particle were used to infect
EC cells (2×104 cells) cultured in 5 µg/ml
Polybrene cultured in a 12-well plate with a multiplicity of
infection (MOI) of 50. At 24 h following transfection, cells were
harvested for analysis.
RT-qPCR
Total RNA was isolated from the cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). miR-93-5p
cDNA was synthesized using the qScript microRNA cDNA synthesis kit
(Quantabio), and TGFβR2 cDNA was synthesized using cDNA synthesis
kit (Thermo Fisher Scientific, Inc.). qPCR was performed using the
miScript SYBR-Green PCR kit (Qiagen GmbH) under the following
thermal cycling conditions: 95°C for 2 min; 95°C for 5 sec and 60°C
for 30 sec, with a total of 40 cycles. U6 and GAPDH were used as
loading controls for miR-93-5p and TGFβR2, respectively. The primer
sequences are listed in Table I.
The quantitative expression values were calculated using the
2−ΔΔCq method (13).
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Gene | Forward | Reverse |
|---|
| U6 |
CAGCACATATACTAAAATTGGAACG |
ACGAATTTGCGTGTCATCC |
| hsa-miR-93-5p |
GCCGCCAAAGTGCTGTTC |
CAGAGCAGGGTCCGAGGTA |
| TGFβR2 |
GTAGCTCTGATGAGTGCAATGAC |
GGGGTCATTGATGGCAACAATA |
| GAPDH |
AAGGTGAAGGTCGGAGTCAAC |
GGGGTCATTGATGGCAACAATA |
Western blot analysis
Cells (following 48 h of transfection) of each group
were lysed on ice for 10 min with RIPA lysis buffer (Sigma-Aldrich;
Merck KGaA). The BCA protein assay kit (Thermo Fisher Scientific,
Inc.) was employed to detect the concentration of the protein
samples. The protein samples (30 mg per lane) were then separated
using sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) with
10% gel, and then transferred onto nitrocellulose membranes
(ZY-160FP, Zeye Bio Co., Ltd.). The membranes were then blocked
with 5% BSA/TBST for 2 h at room temperature and washed 3 times
with 1X TBST. The membranes were incubated with primary antibodies,
including rabbit polyclonal antibody TGFβR2 (ab184948; 1:1,000) and
rabbit polyclonal antibody GAPDH (ab181602; 1:2,500) at 4°C
overnight. The membranes were then washed with 1X TBST and
incubated with horseradish peroxidase-labeled goat anti-rabbit IgG
(ab205718; 1:1,000) for hybridization at room temperature for 2 h.
Finally, protein bands were visualized using
electrochemiluminescence (ECL) solution and then observed and
analyzed. All the antibodies mentioned above were purchased from
Abcam.
Detection of cell proliferation by WST-1
assay
The WST-1 kit (Roche Diagnostics) was used to
determine cell proliferation. EC cells were plated into 96-well
plates at a density of 2.5×105 cell/well, and incubated
with WST-1 at 37°C for 3 h. The absorbance at 450 nm was measured
using a microplate reader (SpectraMax i3, Molecular Devices, LLC)
to determine the cell proliferation rate.
Detection of cell apoptosis by flow
cytometry
Cell apoptosis was detected by flow cytometry using
an Apoptosis kit with Annexin V FITC/propidium iodide (PI) (V13242,
Thermo Fisher Scientific, Inc.) in accordance with the
manufacturer's guidelines. Cells were harvested and rinsed
following 48 h of incubation at 37°C. The cell suspension was mixed
with 10 µl of Annexin V and 5 µl of PI at room
temperature for 10 min. Subsequently, fluorescence activated cell
sorting (FACS) flow cytometry (FACSCalibur, 342976, BD Biosciences)
was used to detect cell apoptosis.
Wound healing assay
After 24 h of cell transfection, when the cells
reached 70-80% confluency, a wound was scraped into the cells using
a tip of 200 µl pipette across the hole center and the cells
were then washed with PBS twice to remove the floating cells.
Subsequently, cells were cultured in fresh serum-free DMEM for a
further 24 h at 37°C. The relative distance of the scratches was
observed under a microscope (Axioskop 40, Carl Zeiss AG). The
photographs were analyzed using an image analysis system (Wound
Healing ACAS, ibidi). For each field of view, 2 straight lines were
drawn on the front of both sides of the gap, and the average
distance was calculated as the average of the distances of the 2
straight lines at the left, center and right points of the
view.
Transwell assay
For the invasion assay, 24-well Transwell chambers
(8 µm pores, BD Biosciences) were used. Approximately
2×104 cells were seeded into the upper chamber coated
with Matrigel (Corning, Inc.), and DMEM medium containing 10% FBS
was added to the lower chamber. The cells in the upper chamber were
wiped off with a cotton swab following incubation at 37°C for 24 h,
while the cells invading the lower chamber were stained with
crystal violet (Sigma-Aldrich; Merck KGaA) at room temperature for
20 min. Finally, cells were counted in randomly selected fields
under a microscope (Axioskop 40, Carl Zeiss AG,) at ×100
magnification.
Dual-luciferase reporter assay
Dual-luciferase reporter gene assay was utilized to
validate whether TGFβR2 is the direct target gene of miR-93-5p. DNA
fragments of TGFβR2 3′-UTR containing the putative binding sites
with miR-93-5p (or mutated binding sites) were amplified by PCR,
digested with XhoI and BamHI, and then ligated into
the Firefly luciferase vector pGL3 (Promega Corporation). The
constructs were named wild-type (TGFβR2-wt) and mutated-type
(TGFβR2-mut) TGFβR2, respectively. TGFβR2-wt or TGFβR2-mut reporter
was transfected into cells along with miR-93-5p mimic or mimic NC.
Following 48 h of transfection, cells were harvested and lysed.
Luciferase activity was determined using a dual luciferase assay
system (Promega Corporation). Relative Firefly luciferase activity
was normalized to Renilla luciferase activity as a control
for transfection efficiency.
Statistical analysis
All data were processed using SPSS 22.0 software
(IBM, Corp.). Measurement data are expressed as the means ±
standard deviation. All experiments were repeated at least 3 times.
A Student's t-test was used for comparisons between 2 groups, while
one-way ANOVA and Tukey's post hoc test were adopted in the case of
comparisons among ≥3 groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
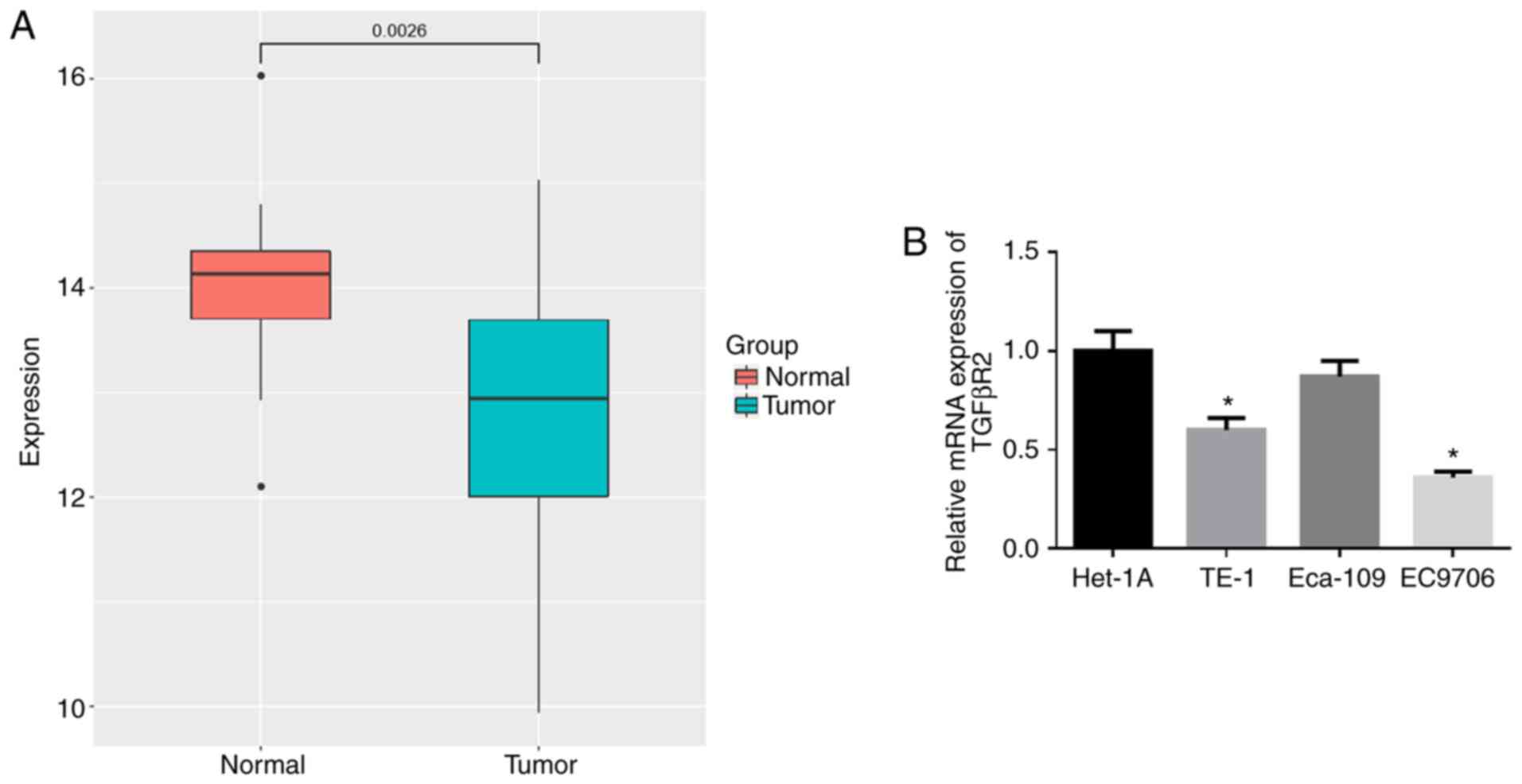
miR-93-5p is highly expressed in EC
tissues and cells
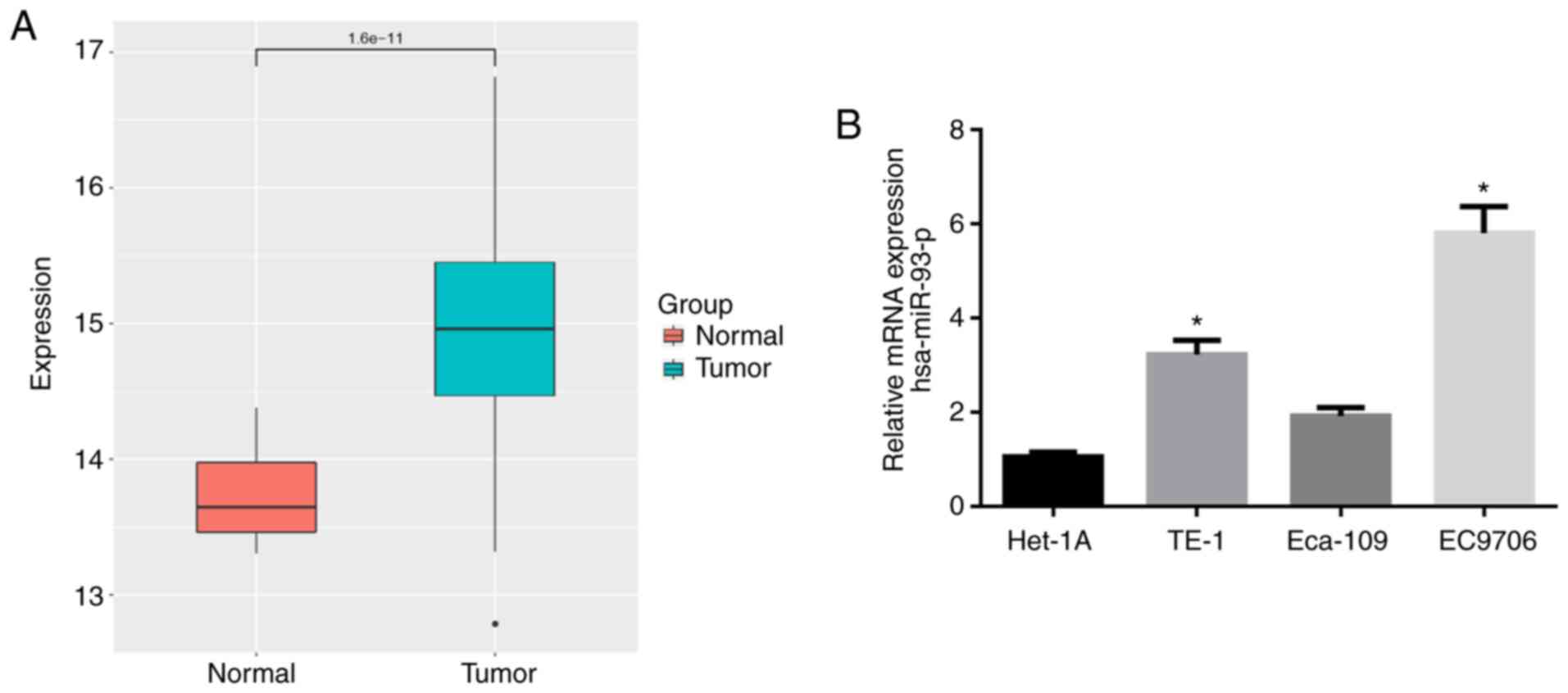
The expression data of miR-93-5p in 176 EC tissue
samples and 13 normal samples were retrieved from the TCGA
database. As shown in Fig. 1A,
the expression of miR-93-5p was significantly higher in EC tissues
compared with normal tissues. RT-qPCR was then performed to verify
that miR-93-5p was highly expressed in the EC cell lines (TE-1,
Eca-109 and EC9706) compared with that in the human normal
esophageal cell line Het-1A (Fig.
1B). The EC9706 cell line with the most differential expression
of miR-93-5p (P<0.05) was thereby selected for use in subsequent
experiments.
Overexpression of miR-93-5p facilitates
the proliferation, migration and invasion, and inhibits the
apoptosis of EC cells
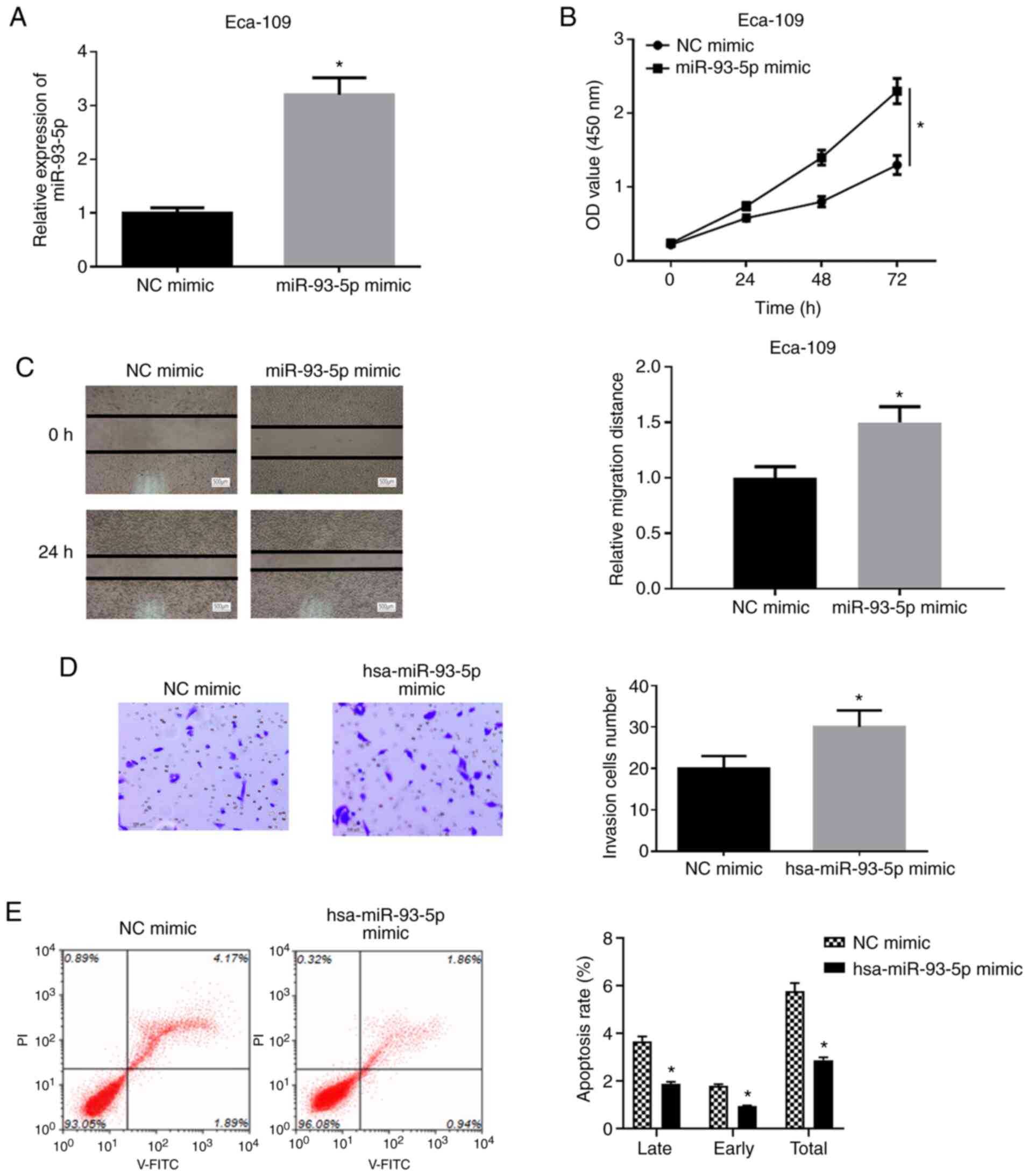
To determine the effects of miR-93-5p on the
biological function of EC cells, hsa-miR-93-5p mimic or NC mimic
were transfected into EC9706 cells and it was found that the
expression of hsa-miR-93-5p was significantly upregulated in the
cells transfected with hsa-miR-93-5p mimic (Fig. 2A). A series of experiments were
then performed using the EC9706 cell line. WST-1 assay was applied
to examine the viability of the cells in the NC mimic group and
miR-93-5p mimic group. As shown in Fig. 2B, the viability of the EC9706
cells in the miR-93-5p mimic group was significantly increased by
comparison with that in the NC mimic group (P<0.05). Wound
healing assay was implemented to detect the migratory ability of
the EC9706 cells (Fig. 2C). The
results revealed that the migration rate of the EC9706 cells in
miR-93-5p mimic group was significantly increased relative to that
in the NC mimic group (P<0.05). Moreover, Transwell invasion
assay was conducted to measure the cell invasive ability (Fig. 2D), and it was found that the
invasive ability of the cells was increased in the miR-93-5p mimic
group (P<0.05). Flow cytometry was also performed for the
detection of cell apoptosis following 48 h of transfection
(Fig. 2E). The apoptotic rate of
the cells in the miR-93-5p mimic group was significantly decreased
relative to that in the NC mimic group (P<0.05). The
above-mentioned results indicated that the overexpression of
miR-93-5p facilitated the viability, migration and invasion, and
decreased the apoptosis of EC cells.
Suppression of miR-93-5p inhibits the
proliferation, migration and invasion, and promotes the apoptosis
of EC cells
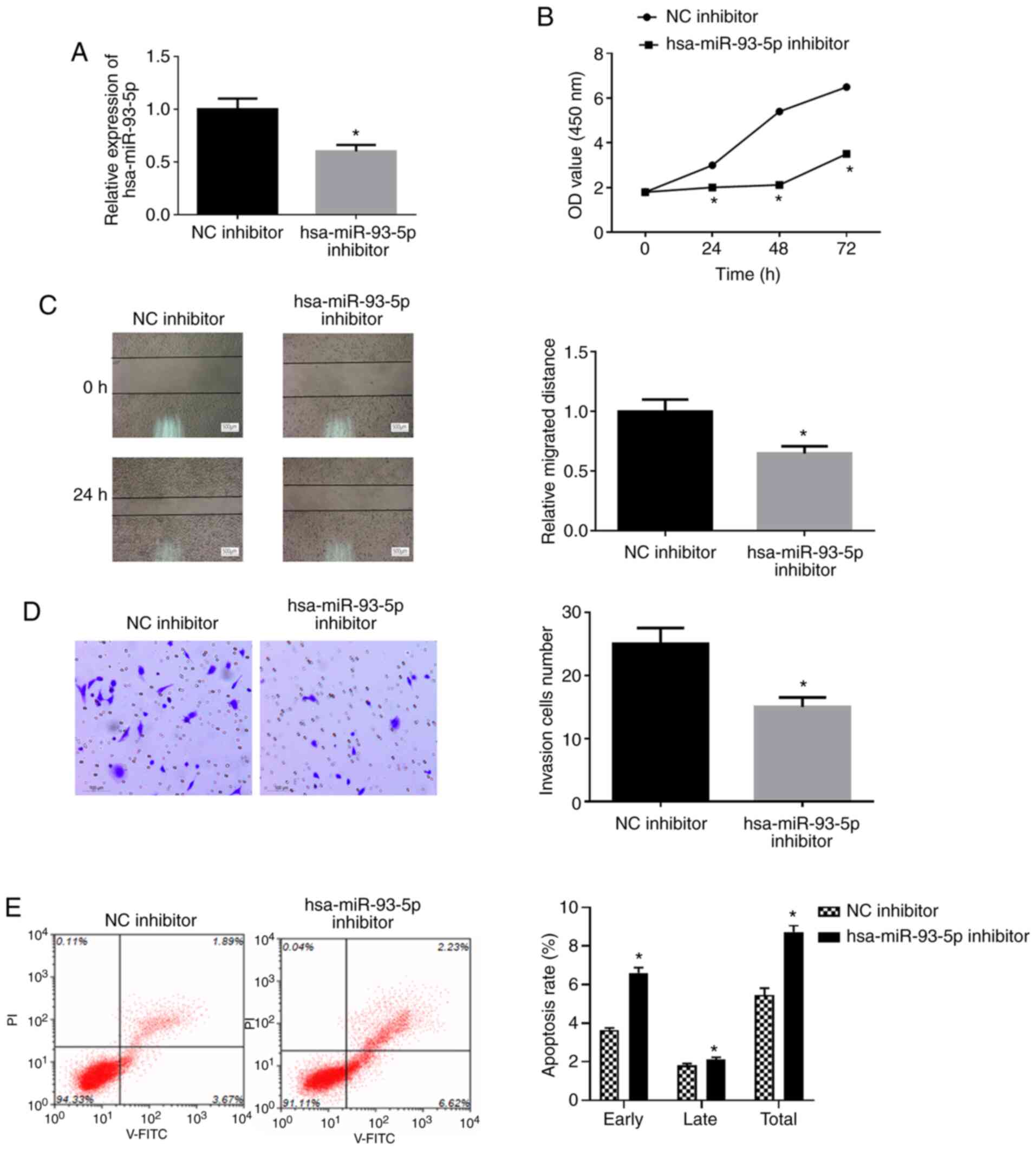
To further examine the effects of miR-93-5p on EC,
EC9706 cells were transfected with miR-93-5p inhibitor or NC
inhibitor (Fig. 3A). WST-1 assay,
wound healing assay and Transwell assay revealed that the viability
(Fig. 3B), migration (Fig. 3C) and invasion (Fig. 3D) of the EC9706 transfected with
miR-93-5p inhibitor were significantly decreased compared with
those in the control group. Flow cytometry was employed for
examination of cell apoptosis. As shown in Fig. 3E, the apoptotic rate of the cells
was significantly increased in the miR-93-5p inhibitor group
compared with the NC inhibitor group (P<0.05). Overall, these
findings indicate that the suppression of miR-93-5p inhibits the
proliferation, migration and invasion, and promotes the apoptosis
in EC cells.
miR-93-5p targets and downregulates
TGFβR2 expression
miRDB (http://mirdb.org/), miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/index.php)
and TargetScan (http://www.targetscan.org/vert_72/) databases were
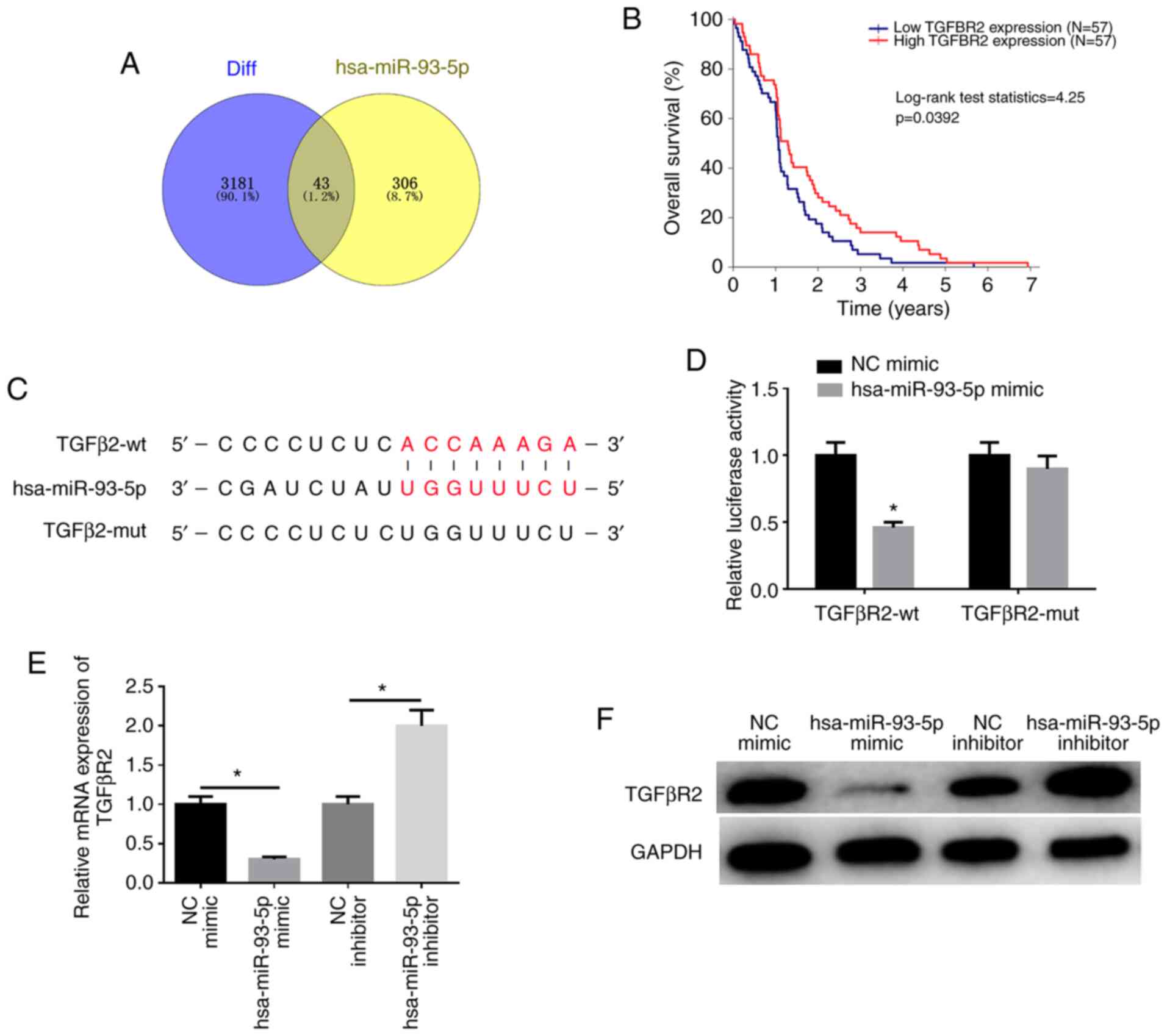
employed to predict the potential mRNAs binding to miR-93-5p and it
was found that there were 43 overlapping mRNAs between the
predicted mRNAs and downregulated DEmRNAs (Fig. 4A). Among these genes, TGFβR2 was
found to be significantly associated with EC prognosis (Fig. 4B). The biological website,
TargetScan (http://www.targetscan.org/vert_72/), was used and it
was found that there was a potential binding site of miR-93-5p on
the TGFβR2 3′-UTR (Fig. 4C),
which was further verified by dual-luciferase reporter gene assay
(Fig. 4D). Compared with the
control group, the luciferase activity of TGFβR2-wt was suppressed
by miR-93-5p overexpression, while that of TGFβR2-mut was
unaffected (P<0.05). These findings demonstrate that miR-93-5p
can target TGFβR2. In addition, the results of western blot
analysis and RT-qPCR revealed that the expression of TGFβR2 was
markedly downregulated in the miR-93-5p mimic group compared with
the NC mimic group, while the expression of TGFβR2 was considerably
upregulated in the miR-93-5p inhibitor group than that in the NC
inhibitor group (P<0.05; Fig. 4E
and F). These results indicate that hsa-miR-93-5p targets
TGFβR2 expression.
TGFβR2 is expressed in low levels in EC
tissues and cells
In addition, the expression of TGFβR2 in EC tissue
samples in TCGA-ESCA was significantly downregulated (Fig. 5A). RT-qPCR was used to examine the
expression of TGFβR2 in the human normal esophageal cell line,
Het-1A, and in the EC cell lines TE-1, Eca-109 and EC9706. As
illustrated in Fig. 5B, TGFβR2
was markedly downregulated in the TE-1 and EC9706 cells by
comparison with that in the Het-1A cell line (P<0.05), with the
lowest expression observed in the EC9706 cells (P<0.05).
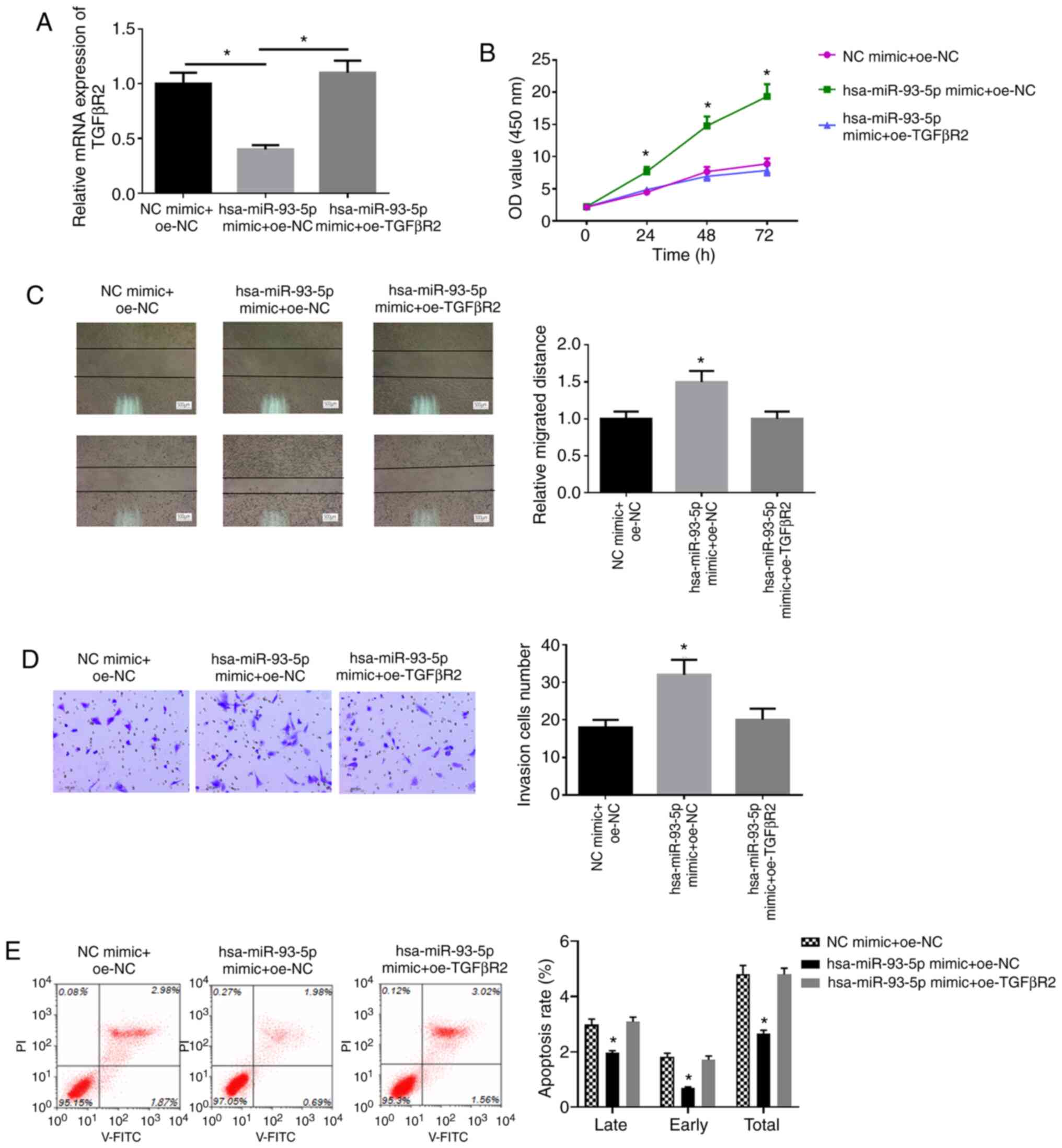
miR-93-5p promotes the proliferation,
migration and invasion, and inhibits the apoptosis of EC cells by
downregulating TGFβR2 expression
To confirm whether TGFβR2 can reverse the effects of
miR-93-5p on the proliferation, migration and invasion of EC cells,
EC9706 cells were transfected with miR-93-5p mimic + oe-NC, NC
mimic + oe-NC, miR-93-5p mimic + oe-TGFβR2, respectively. The
expression of TGFβR2 following transfection was detected by
RT-qPCR. It was found that the mRNA and protein expression of
TGFβR2 was successfully increased following transfection with
TGFβR2 overexpression vector (Fig.
S1). The results also revealed that the overexpression of
hsa-miR-93-5p inhibited TGFβR2 mRNA expression, and oe-TGFβR2
reversed the inhibitory effects of hsa-miR-93-5p mimic on TGFβR2
mRNA expression (Fig. 6A). WST-1
assay was used to examine the viability of the cells in each group.
As shown in Fig. 6B, the cells in
the miR-93-5p mimic + oe-NC group had a higher viability compared
with those in the NC mimic + oe-NC group (P<0.05), and no
significant differences were observed between the NC mimic + oe-NC
group and miR-93-5p mimic + oe-TGFβR2 group (P>0.05). Wound
healing assay and Transwell assay were applied to detect the cell
migration and invasion ability. The cells in the miR-93-5p mimic +
oe-NC group also exhibited higher migratory and invasive abilities
relative to those in the NC mimic + oe-NC group (P<0.05;
Fig. 6C and D), and there was no
marked differences between the NC mimic + oe-NC group and miR-93-5p
mimic + oe-TGFβR2 group (P>0.05). Flow cytometry was performed
to detect cell apoptosis. The cell apoptotic ability was
significantly decreased in the miR-93-5p mimic + oe-NC group by
comparison with that in the NC mimic + oe-NC group (P<0.05;
Fig. 6E), while there were no
obvious differences between the NC mimic + oe-NC group and
miR-93-5p mimic + oe-TGFβR2 group (P>0.05). These findings
confirm that overexpressing miR-93-5p can promote proliferation,
migration, invasion and inhibit cell apoptosis of EC cells upon
TGFβR2 suppression.
Discussion
miRNAs are types of small RNAs with a length of
approximately 20-24 bp, and they serve as a tumor promoter or
suppressor via regulating the expression of their specific target
genes (14). Therefore, the
regulation of miRNA expression can be used as a novel method for
cancer diagnosis and treatment (15). miR-93-5p is located in the intron
of the MCM7 gene and is a part of the cluster containing two other
miRNAs (miR-25 and miR-106b) (8).
The present study found that miR-93-5p was highly expressed in EC
cells, which was consistent with previous findings on breast cancer
(7), gastric cancer (16), prostate cancer (17) and colorectal cancer (6). Studies have reported that tje
knockdown of miR-93-5p inhibits cell proliferation, migration and
invasion in gastric cancer tissues (18). The present study also revealed
that tje inhibition of miR-93-5p expression inhibited the
proliferation, migration and invasion of EC cells, and promoted
apoptosis.
During tumorigenesis and development, TGFβR2, a
receptor serine/threonine kinase, initiates downstream TGF-β
signaling (17), and thje loss or
decrease of TGFβR2 expression can inhibit the TGF-β signaling,
which is beneficial for early tumor growth (19). In the present study, it was
predicted that TGFβR2 was expressed in low levels in EC tissues by
bioinformatics analysis, and western blot analysis was performed to
verify this prediction and further explore the association between
TGFβR2 and the occurrence and development of EC. RT-qPCR then
revealed that TGFβR2 mRNA expression was decreased in EC cell lines
relative to that in normal esophageal cell line. It has been
demonstrated that TGFβR2 is the major target of miR-93-5p in
nasopharyngeal carcinoma invasion (20). In the present study, target
prediction websites were used, and it was found that miR-93-5p
bound to the 3′-UTR of TGFβR2, which was further verified by
performing dual-luciferase reporter gene assay. It was then
demonstrated that the overexpression of miR-93-5p inhibited the
expression of TGFβR2, thereby inhibiting apoptosis and promoting
the proliferation, migration and invasion of EC cells. However,
miR-93-5p inhibitor was used to downregulate miR-93-5p expression,
and opposite results were observed with the inhibition of miR-93-5p
expression. When miR-93-5p and TGFβR2 were simultaneously
overexpressed in the cells, no marked changes were observed in the
proliferation, migration and invasion of the cells. All the
findings described above indicate that miR-93-5p targets and
downregulates the expression of TGFβR2, ultimately promoting the
occurrence and development of EC cells.
In conclusion, the present study identified the
regulatory effects of miR-93-5p on TGFβR2 in EC cells and clarified
its role in biological cell behaviors. The results provide a
potential target by inhibiting miR-93-5p or promoting TGFβR2
expression, which may provide additional insight into the
mechanisms underlying the occurrence and development of EC and may
lead to the development of novel strategies for EC diagnosis and
treatment.
Supplementary Data
Funding
No funding was received.
Availability of data and materials
The data used to support the findings of this study
are included in the current article or are available from the
corresponding author upon request.
Authors' contributions
YC, WR, JD and KZ contributed to the study design.
NW, JW and HZ conducted the literature search. NM acquired the
data. YC, WR and KZ wrote the article. NM, GW, WS and YL performed
the data analysis. All authors gave the final approval of the
version to be submitted. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nishimura T, Tamaoki M, Komatsuzaki R, Oue
N, Taniguchi H, Komatsu M, Aoyagi K, Minashi K, Chiwaki F,
Shinohara H, et al: SIX1 maintains tumor basal cells via
transforming growth factor-β pathway and associates with poor
prognosis in esophageal cancer. Cancer Sci. 108:216–225. 2017.
View Article : Google Scholar :
|
|
3
|
Wu K, He J, Pu W and Peng Y: The role of
exportin-5 in MicroRNA biogenesis and cancer. Genomics Proteomics
Bioinformatics. 16:120–126. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen S, Chen X, Sun KX, Xiu YL, Liu BL,
Feng MX, Sang XB and Zhao Y: MicroRNA-93 promotes
epithelial-mesenchymal transition of endometrial carcinoma cells.
PLoS One. 11:e01657762016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paul S, Lakatos P, Hartmann A,
Schneider-Stock R and Vera J: Identification of miRNA-mRNA modules
in colorectal cancer using rough hypercuboid based supervised
clustering. Sci Rep. 7:428092017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li N, Miao Y, Shan Y, Liu B, Li Y, Zhao L
and Jia L: MiR-106b and miR-93 regulate cell progression by
suppression of PTEN via PI3K/Akt pathway in breast cancer. Cell
Death Dis. 8:e27962017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du L, Zhao Z, Ma X, Hsiao TH, Chen Y,
Young E, Suraokar M, Wistuba I, Minna JD and Pertsemlidis A:
miR-93-directed downregulation of DAB2 defines a novel oncogenic
pathway in lung cancer. Oncogene. 33:4307–4315. 2014. View Article : Google Scholar
|
|
9
|
Zhu W, He J, Chen D, Zhang B, Xu L, Ma H,
Liu X, Zhang Y and Le H: Expression of miR-29c, miR-93, and miR-429
as potential biomarkers for detection of early stage non-small lung
cancer. PLoS One. 9:e877802014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kucuksayan H, Akgun S, Ozes ON, Alikanoglu
AS, Yildiz M, Dal E and Akca H: TGF-β-SMAD-miR-520e axis regulates
NSCLC metastasis through a TGFBR2-mediated negative-feedback loop.
Carcinogenesis. 40:695–705. 2019. View Article : Google Scholar
|
|
11
|
Malkoski SP, Haeger SM, Cleaver TG,
Rodriguez KJ, Li H, Lu SL, Feser WJ, Barón AE, Merrick D, Lighthall
JG, et al: Loss of transforming growth factor beta type II receptor
increases aggressive tumor behavior and reduces survival in lung
adenocarcinoma and squamous cell carcinoma. Clin Cancer Res.
18:2173–2183. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fricke F, Lee J, Michalak M, Warnken U,
Hausser I, Suarez-Carmona M, Halama N, Schnölzer M, Kopitz J and
Gebert J: TGFBR2-dependent alterations of exosomal cargo and
functions in DNA mismatch repair-deficient HCT116 colorectal cancer
cells. Cell Commun Signal. 15:142017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Liu JJ, Zhang X and Wu XH: miR-93 promotes
the growth and invasion of prostate cancer by upregulating its
target genes TGFBR2, ITGB8, and LATS2. Mol Ther Oncolytics.
11:14–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu YF, Yu JR, Yang Z, Zhu GX, Gao P, Wang
H, Chen SY, Zhang J, Liu MY, Niu Y, et al: Promoter hypomethylation
mediated upregulation of MicroRNA-10b-3p targets FOXO3 to promote
the progression of esophageal squamous cell carcinoma (ESCC). J Exp
Clin Cancer Res. 37:3012018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma DH, Li BS, Liu JJ, Xiao YF, Yong X,
Wang SM, Wu YY, Zhu HB, Wang DX and Yang SM: miR-93-5p/IFNAR1 axis
promotes gastric cancer metastasis through activating the STAT3
signaling pathway. Cancer Lett. 408:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou H, Wu G, Ma X, Xiao J, Yu G, Yang C,
Xu N, Zhang B, Zhou J, Ye Z and Wang Z: Attenuation of TGFBR2
expression and tumour progression in prostate cancer involve
diverse hypoxia-regulated pathways. J Exp Clin Cancer Res.
37:892018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schumacher K, Dagres N, Hindricks G,
Husser D, Bollmann A and Kornej J: Characteristics of PR interval
as predictor for atrial fibrillation: Association with biomarkers
and outcomes. Clin Res Cardiol. 106:767–775. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mishra S, Deng JJ, Gowda PS, Rao MK, Lin
CL, Chen CL, Huang T and Sun LZ: Androgen receptor and microRNA-21
axis downregulates transforming growth factor beta receptor II
(TGFBR2) expression in prostate cancer. Oncogene. 33:4097–4106.
2014. View Article : Google Scholar
|
|
20
|
Lyu X, Fang W, Cai L, Zheng H, Ye Y, Zhang
L, Li J, Peng H, Cho WC, Wang E, et al: TGFβR2 is a major target of
miR-93 in nasopharyngeal carcinoma aggressiveness. Mol Cancer.
13:512014. View Article : Google Scholar
|