Introduction
Atherosclerosis (AS) is a common type of
cardiovascular disease, characterized by progressive inflammation
and lesions on the arterial wall (1). Lipid deposition in the arterial
wall is the main cause of AS, and lipoproteins and lipid-related
factors are implicated in the pathogenesis of AS (2). Furthermore, the dysfunctions of
endothelial cells substantially contribute to the development of
AS, as endothelial cells are important components that protect the
histological layers of the arterial wall from damage (3,4).
Thus, it is a feasible strategy to study the induction factors and
molecular mechanisms of endothelial cell injury and dysfunction for
the treatment of AS.
Oxidized low-density lipoprotein (ox-LDL) evokes
cellular oxidative stress, inflammatory responses and cell
apoptosis, leading to endothelial cell dysfunction, and ox-LDL is
thus regarded as a crucial element in the pathogenesis of AS
(5,6). Human umbilical vein endothelial
cells (HUVECs) exposed to ox-LDL are widely used to as an in
vitro cell model of AS (7,8).
However, the mechanisms through which ox-LDL induces HUVEC damage
and dysfunction are complex and variable, and have not yet been
fully elucidated. RNA sequencing technology provides a precise
approach for the identification of circRNAs that are differently
regulated in human diseases (9).
CircRNAs derive from precursor mRNAs in a 'back-splicing' manner
(10). With the increase in
circRNA research, circRNAs as indicators or tools of disease
diagnosis, treatment and prognosis have been widely recognized
(11). Pan et al obtained
circRNA expression profiles using RNA sequencing and identified 66
specific circRNAs in coronary artery AS (12). Li et al utilized circRNA
expression profiles to analyze significantly dysregulated circRNAs
in ox-LDL-induced HUVECs (13),
and circ_0003204 was found as one of the differentially expressed
circRNAs with an upregulated expression in ox-LDL-induced HUVECs
(13). Circbase (http://www.circbase.org/) indicates that circ_0003204
is derived from precursor mRNA USP36, thus terming it as circUSP36.
However, the function of circUSP36 in ox-LDL-induced HUVECs is
largely unclear.
MicroRNAs (miRNAs or miRs) are characterized by RNAs
with 18-24 nucleotides in length that govern extensive biological
functions (14). Accumulating
evidence has suggested that miRNAs can be used as diagnostic and
therapeutic markers for AS and that they regulate the pathologies
of vascular diseases (15,16). Previous studies have demonstrated
that miR-20a-5p is involved in the development of cardiovascular
diseases (17,18). Research into the role of
miR-20a-5p in ox-LDL-induced HUVECs is not limited. Rho-associated
coiled-coil kinase 2 (ROCK2) is one of the isoforms of the ROCK
family (19). It has been
demonstrated that ROCKs are substantially implicated in
cardiovascular diseases, including AS, and the dysregulation of
ROCKS regulates various cardiovascular pathological processes, such
as abnormal apoptosis, cardiac hypertrophy and cardiac fibrosis
(19,20). The functional role of ROCK2 in AS
is largely clear, and the mechanisms of ROCK2 action need to be
further explored, particularly in association with circUSP36 and
miR-20a-5p.
In the present study, the expression of circUSP36
was verified in ox-LDL-exposed HUVECs, and a loss-of-function assay
was performed to explore the function of circUSP36 in HUVECs.
Moreover, the interaction between miR-20a-5p and circUSP36 or ROCK2
was validated. The present study provides a novel mechanism (at
least to the best of our knowledge) to explain the functional mode
of circUSP36 in ox-LDL-induced HUVEC dysfunction.
Materials and methods
HUVEC treatment
HUVECs were obtained from Procell Life Science &
Technology Co., Ltd. (no. CL-0122) and maintained in HUVEC specific
medium [Ham's F-12K+10% fetal bovine serum (FBS); Procell Life
Science & Technology] at 37°C conditions with 5%
CO2. HUVECs were exposed to various concentrations of
ox-LDL (40, 60, 80 and 100 µg/ml) and incubated for
different periods of time (6, 12, 24 and 48 h) at 37°C conditions
with 5% CO2. The cell model of AS was constructed using
HUVECs exposed to 60 µg/ml ox-LDL with a 24-h incubation at
37°C conditions with 5% CO2.
Cell viability detection
Cell counting kit-8 (CCK-8) assay was implemented to
detect cell viability. HUVECs exposed to various concentrations of
ox-LDL were supplemented with 10 µl CCK-8 reagent (Beyotime
Institute of Biotechnology) at 24 h post-treatment and mixed for a
further 2 h. Moreover, HUVECs exposed to 60 µg/ml ox-LDL
were reacted with 10 µl CCK-8 reagent at 6, 12, 24 and 48 h
post-treatment and mixed for a further 2 h. The absorbance at 450
nm was then monitored using a microplate reader (Thermo Fisher
Scientific, Inc.) to observe cell viability.
Cell cycle analysis
HUVECs were collected and resuspended in fresh
culture medium containing 10% FBS. Cells were collected and gently
resuspended in 70% ice-cold ethanol for fixing at -20°C overnight.
Subsequently, the cells were collected, washed and resuspended in
fresh culture medium containing 10% FBS. The cells were then
collected and resuspended in 200 µl Cell Cycle Solution
[containing propidium iodide (PI), RNase A, and Triton X-100;
Invitrogen; Thermo Fisher Scientific, Inc.] at a density of
2×105 cells. Cells were then incubated at room
temperature for 30 min without light and finally analyzed using an
EPICS XL-MCL FACScan flow cytometer (Beckman Coulter, Inc.).
Cell apoptosis analysis
Cell apoptosis was monitored using the Annexin
V-FITC Apoptosis Detection kit (Beyotime Institute of
Biotechnology). Briefly, a total of 5×104 HUVECs were
collected at 48 h post-transfection and resuspended in cold
phosphate-buffered saline (PBS). Cells were then collected and
resuspended in 195 µl Annexin V-FITC binding buffer followed
by the addition of 5 µl Annexin V-FITC and 10 µl PI.
Cells were incubated at room temperature for 20 min in the dark and
subsequently analyzed using an EPICS XL-MCL FACScan flow cytometer
(Beckman Coulter, Inc.).
Cell migration and invasion assay
Transwell migration and invasion assays were
implemented using Transwell chambers (Corning, Inc.). Chambers were
pre-coated with or without Matrigel (Corning, Inc.) at 4°C
overnight. HUVECs were collected at 24 h post-transfection,
resuspended in fresh culture medium with serum depletion, and
transferred into the top of chambers (5×104 cells)
coated with or without Matrigel for invasion or migration analysis,
respectively. The bottom chambers were filled with fresh culture
medium containing 10% FBS to induce cell migration and invasion.
Allowing for migration and invasion for 24 h at 37°C, the cells
that had migrated or invaded into the lower surface were fixed with
formaldehyde, stained with 0.1% crystal violet (Beyotime Institute
of Biotechnology) for 20 min at 37°C and then observed under a
CKX41 light microscope (Olympus Corporation) at a magnification of
×100.
Western blot analysis
Total proteins were extracted from the HUVECs and
quantified using the Enhanced BCA Protein Assay kit (Beyotime
Institute of Biotechnology). Equal proteins (20 µg per lane)
were then separated by 12% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis (SDS-PAGE) and loaded onto polyvinylidene
fluoride (PVDF) membranes (Beyotime Institute of Biotechnology).
The membranes were subjected to blocking buffer (5% skim milk) at
room temperature for 1 h and probed with the primary antibodies,
including anti-interleukin (IL)-6 (ab6672; 1/1,000), anti-IL-1β
(ab226918; 1/1,000), anti-β-actin (ab8227; 1/2,000) and anti-ROCK2
(ab71598; 1/1,000) (all from Abcam) at 4°C overnight. The membranes
were then probed with the secondary antibodies (ab205718; 1/5,000;
Abcam) at room temperature for 2 h. Finally, the enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology) was
adopted to visualize the protein blots, and the blot bands were
then quantified using ImageJ software (version 1.46; National
Institutes of Health).
Enzyme-linked immunosorbent assay
(ELISA)
The release of IL-6 and IL-1β in the culture medium
was detected using ELISA, with the application of the Human IL-6
ELISA kit (cat. no. PI330; Beyotime Institute of Biotechnology) and
the human IL-1β ELISA kit (cat. no. PI305; Beyotime Institute of
Biotechnology). The assay was performed in line with the guidelines
from the manufacturer's protocols.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using a Total RNA Extraction
kit (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
product manual. Complementary DNA (cDNA) for circUSP36 and ROCK2
was synthesized using a Universal RT-PCR kit (cat. no. RP1100;
Beijing Solarbio Science & Technology Co., Ltd.), and cDNA for
miR-20a-5p was synthesized using a miRNA cDNA Synthesis kit (cat.
no. 638315; Takara Biotechnology, Inc.). Subsequently, SYBR Master
Mix (cat. no. SR1110; Beijing Solarbio Science & Technology
Co., Ltd.) was used for qPCR analysis on a CFX96 Touch real-time
PCR system (Bio-Rad Laboratories, Inc.). All experimental
procedures were conducted in line with the manufacturers'
protocols. β-actin or U6 was set as the housekeeping gene for
circUSP36 and ROCK2 or miR-20a-5p, respectively. Relative
expression was calculated using the 2−ΔΔCq method
(21). The primer sequences are
listed as follows: circUSP36 forward, 5′-GGC AGT AGA AGA GGA TGG
GC-3′ and reverse, 5′-GGA GCA GGT GAC ACA GCC-3′; miR-20a-5p
forward, 5′-GCC CGC TAA AGT GCT TAT AGT G-3′ and reverse, 5′-GCT
GTC AAC GAT ACG CTA CGT-3′; ROCK2 forward, 5′-GAG AGC TTG CTG GAT
GGC TT-3′ and reverse, 5′-CGA ACC AAC TGC ACT TCA CC-3′; U6
forward, 5′-ATT GGA ACG ATA CAG AGA AGA TT-3′ and reverse, 5′-GGA
ACG CTT CAC GAA TTT G-3′; and β-actin forward, 5′-AGA TCA AGA TCA
TTG CTC CTC CTG A-3′ and reverse, 5′-ATA CTC CTG CTT GCT GAT CCA
CAT C-3′.
Cell transfection
Small interfering RNA (siRNA) targeting circUSP36
(si-circUSP36: 5′-GGG GAC CGC ATG GGG CTG TGT-3′) or ROCK2
(si-ROCK2: 5′-CAG AAG TGC AAG TCT ATT A-3′) for circUSP36 knockdown
or ROCK2 knockdown and their corresponding controls (si-NC;
scramble) were synthesized by Guangzhou RiboBio Co., Ltd. The mimic
for miR-20a-5p for the restoration of miR-20a-5p expression
(miR-20a-5p mimic), the inhibitor of miR-20a-5p for the inhibition
of miR-20a-5p expression (miR-20a-5p inhibitor), and their
corresponding controls (miR-NC mimic or miR-NC inhibitor; scramble)
were purchased from Guangzhou RiboBio Co., Ltd. CircUSP36 was
assembled into the pCD-ciR vector for circUSP36 overexpression
(oe-circUSP36) (GenePharma, Inc.), and pCD-ciR empty vector
(Vector) was used as its control. Lipofectamine 3000 kit
(Invitrogen; Thermo Fisher Scientific, Inc.) was applied to perform
cell transfection (siRNAs, 30 nM; miRNA mimic or inhibitor, 40 nM;
vector, 1.5 µg) according to the manufacturer's
instructions.
Bioinformatics analysis
For the prediction of putative targets of circUSP36,
the online tool starBase (http://starbase.sysu.edu.cn/) was adopted. For the
prediction of putative targets of miR-20a-5p, the online tool
Targetscan (http://www.targetscan.org/vert_72/) was utilized.
Dual-luciferase reporter assay
According to the wild-type sequence of circUSP36 or
ROCK2 3′UTR containing miR-20a-5p binding sites, the mutant
sequence of circUSP36 or ROCK2 3′UTR (binding sites mutation) was
designed. These sequences (wild-type and mutant-type) were
amplified and separately inserted into pGL4 reporter plasmid
(Promega Corporation) to generate luciferase reporter plasmids,
which were named as WT-circUSP36, MUT-circUSP36, WT-ROCK2 and
MUT-ROCK2, respectively. WT-circUSP36 (2 µg), MUT-circUSP36
(2 µg), WT-ROCK2 (2 µg) or MUT-ROCK2 (2 µg)
together with miR-20a-5p mimic (40 nM) or miR-NC mimic (40 nM) were
co-transfected into the HUVECs using Lipofectamine 3000 kit
(Invitrogen; Thermo Fisher Scientific, Inc.). Transfected cells
were then incubated for 48 h at 37°C, and the luciferase activities
in these cells were investigated using the dual-luciferase reporter
system (Promega Corporation) according to the manufacturer's
protocol. Relative luciferase activity was expressed as
normalization of Renilla luciferase activity to Firefly
luciferase activity.
RNA binding protein immunoprecipitation
(RIP) assay
HUVECs were transfected with miR-20a-5p mimic (60
nM) or miR-NC mimic (60 nM) using Lipofectamine 3000 kit
(Invitrogen; Thermo Fisher Scientific, Inc.) and lysed in RIP lysis
buffer from a Magna RIP kit (cat. no. 17-700; EMD Millipore). Cell
lysates were reacted with magnetic beads conjugated with Argonaute
2 (Ago2) antibody (cat. no. ab186733; 1/1,000; Abcam) or
immunoglobulin G (IgG; control) antibody (cat. no. ab190475;
1/1,000; Abcam) at 4°C overnight. The unbound materials were washed
off, and complexes were eluted to extract RNAs. RT-qPCR analysis
was then carried out to examine the expression of circUSP36 and
ROCK2 as described above.
Statistical analysis
All experiments contained 3 independent repeats.
Statistical analysis was processed using GraphPad Prism 5.0
(GraphPad Software, Inc.). The comparison of the differences
between 2 groups was evaluated by unpaired Student's t-test, and
the comparison of differences among ≥3 groups was distinguished by
one- and two-way analyses of variance (ANOVA) with Tukey's post hoc
test. Data are presented as the means ± standard deviation. A
P-value <0.05 was considered to indicate a statistically
significant difference.
Results
CircUSP36 is highly expressed in
ox-LDL-exposed HUVECs
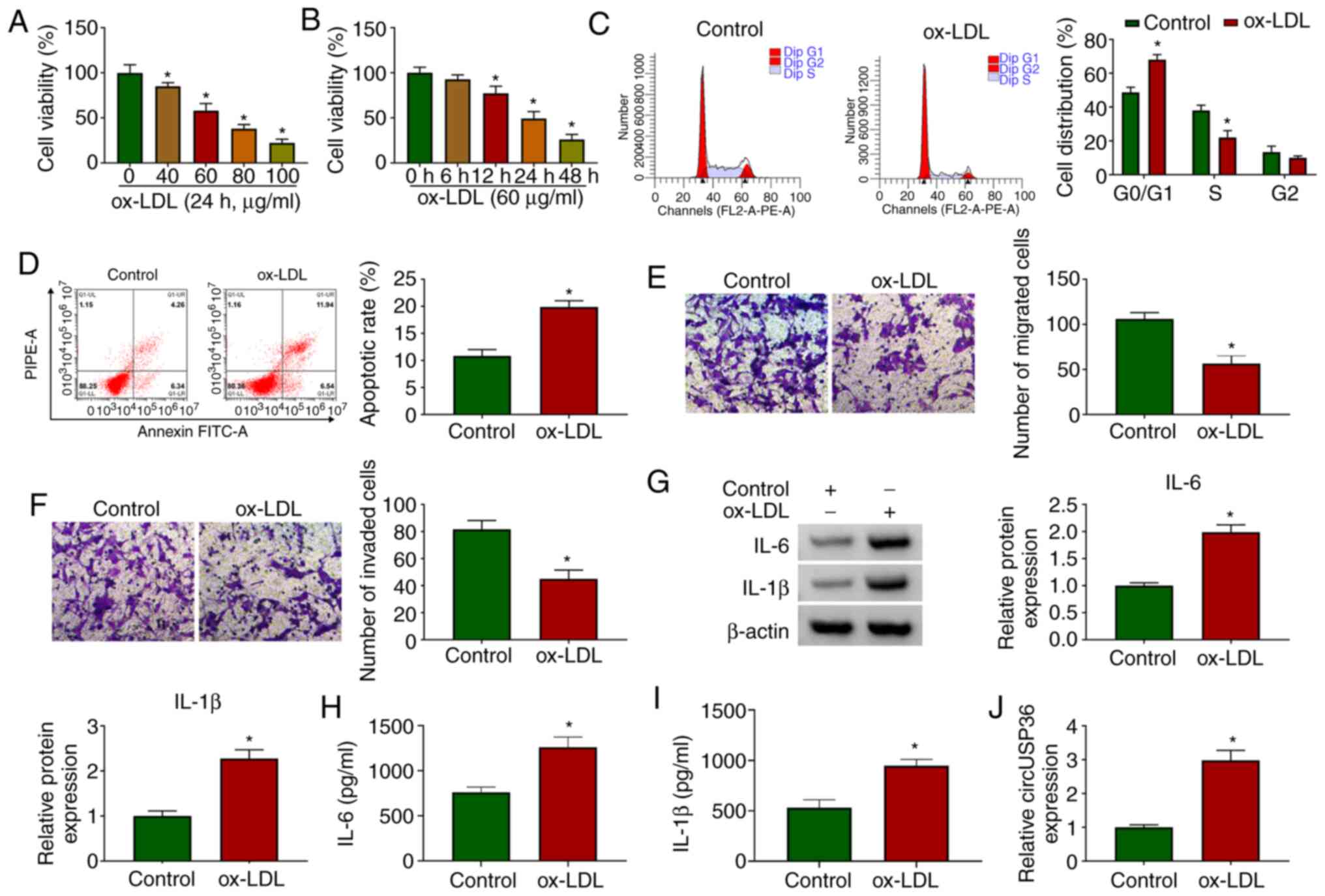
HUVECs were exposed to ox-LDL to examine the effects
of ox-LDL on HUVECs. As shown by CCK-8 assay, the viability of the
HUVECs was significantly decreased with the addition of ox-LDL (24
h) in a dose-dependent manner (Fig.
1A). Moreover, the viability of the HUVECs was also
significantly decreased with the addition of ox-LDL (60
µg/ml) in a time-dependent manner (Fig. 1B). For the HUVECs exposed to 60
µg/ml ox-LDL for 24 h, flow cytometric assay revealed that
the cell cycle was notably arrested (Fig. 1C), and cell apoptosis was notably
promoted compared to the control (Fig. 1D). In addition, the addition of
ox-LDL visibly blocked HUVEC migration and invasion, as shown by
Transwell assay (Fig. 1E and F).
The protein levels of pro-inflammatory factors (IL-6 and IL-1β)
were significantly elevated in the HUVECs exposed to ox-LDL
(Fig. 1G). The release of IL-6
and IL-1β was significantly enhanced in the culture medium of
HUVECs exposed to ox-LDL (Fig. 1H
and I). All these data suggested that the in vitro model
of AS was successfully established, and that ox-LDL impaired HUVEC
survival and metastasis, but induced apoptosis and inflammatory
responses. Furthermore, it was observed that the expression of
circUSP36 was significantly increased in the ox-LDL-exposed HUVECs
(Fig. 1J), in a dose- and
time-dependent manner (Fig.
S1), suggesting that circUSP36 plays a role in ox-LDL-induced
HUVEC injury.
CircUSP36 inhibition attenuates
ox-LDL-induced cell cycle arrest, apoptosis and inflammatory
responses, but enhances the impaired migration and invasion of
HUVECs induced by ox-LDL
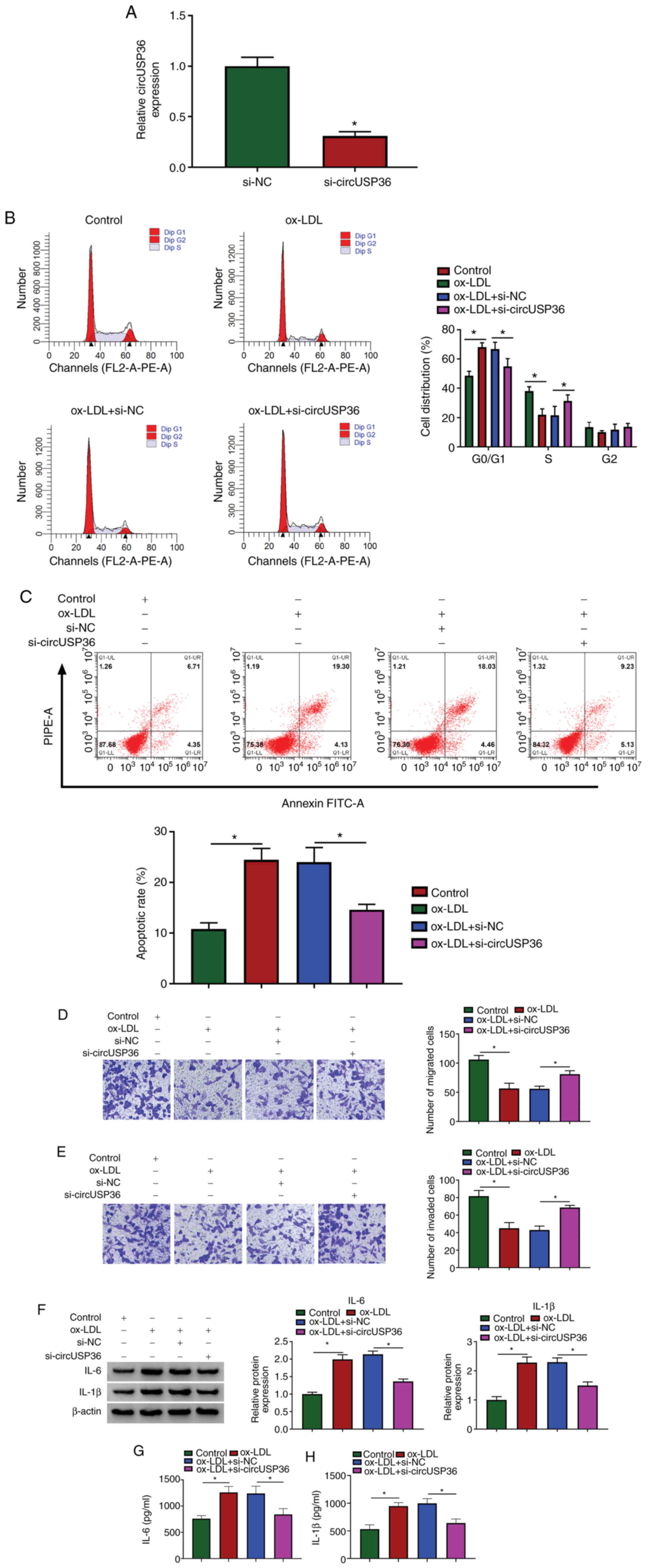
Given that the expression of circUSP36 was elevated
in the ox-LDL-exposed HUVECs, the endogenous level of circUSP36 was
decreased to explore its potential function. The expression of
circUSP36 was significantly decreased in the HUVECs following
transfection with si-circUSP36 (Fig.
2A). Flow cytometric assay revealed that si-circUSP36
transfection substantially diminished the cell cycle arrest and
apoptosis which were induced by ox-LDL (Fig. 2B and C). Transwell assay revealed
that the number of migrated and invaded cells which were blocked by
ox-LDL exposure was largely recovered with the knockdown of
circUSP36 (Fig. 2D and E). In
addition, the protein levels of IL-6 and IL-1β in the HUVECs
stimulated by ox-LDL were blocked by the knockdown of circUSP36
(Fig. 2F), and the release of
IL-6 and IL-1β in culture medium was increased by ox-LDL, but
weakened by transfection with si-circUSP36 (Fig. 2G and H). These data indicated
that ox-LDL-induced HUVEC dysfunctions and inflammatory responses
were alleviated by circUSP36 inhibition.
CircUSP36 targets miR-20a-5p and
negatively regulates miR-20a-5p expression
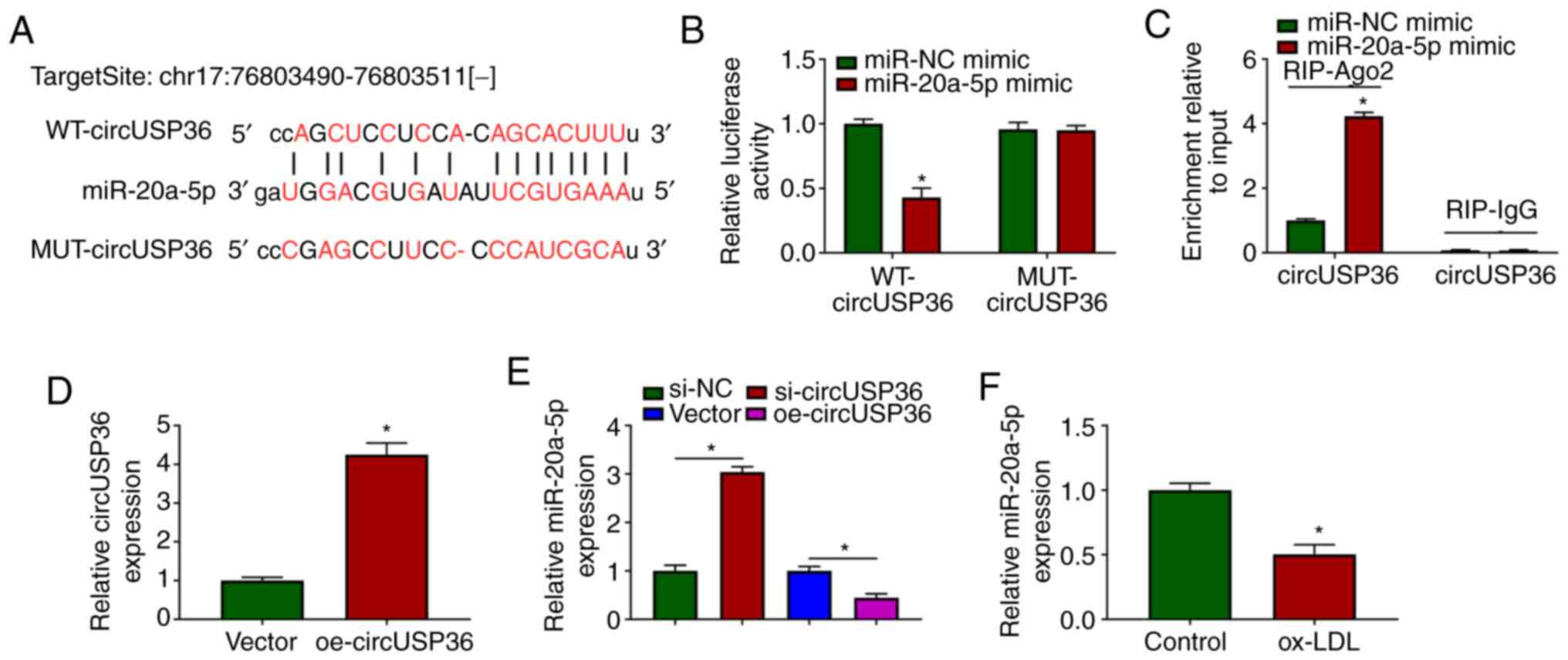
To ascertain whether circUSP36 serves as a molecular
sponge to mediate the expression of downstream miRNAs, the
potential target miRNAs of circUSP36 were identified. The targeting
sites between circUSP36 and miR-20a-5p were analyzed using the
bioinformatics tool, starBase, which revealed that miR-20a-5p may
be a target of circUSP36 (Fig.
3A). Subsequently, dual-luciferase reporter assay demonstrated
that transfection with miR-20a-5p mimic significantly suppressed
the luciferase activities in HUVECs transfected with WT-circUSP36
reporter plasmid, but not with the MUT-circUSP36 reporter plasmid
(Fig. 3B). In addition, RIP
assay revealed that transfection with miR-20a-5p mimic notably
enriched circUSP36 in the RIP-Ago group compared with miR-NC mimic,
while circUSP36 in the RIP-IgG group was hardly enriched (Fig. 3C). A circUSP36 overexpression
plasmid was constructed, and transfection with oe-circUSP36
markedly enhanced the expression of circUSP36 (Fig. 3D). It was noted that the
expression of miR-20a-5p was markedly increased in the HUVECs in
which circUSP36 was knocked down, while it was decreased in the
HUVECs in which circUSP36 was overexpressed (Fig. 3E). Notably, the expression of
miR-20a-5p was markedly decreased in the ox-LDL-exposed HUVECs
(Fig. 3F). On the whole, these
findings demonstrated that miR-20a-5p was a target of circUSP36,
and that circUSP36 negatively regulated the expression of
miR-20a-5p.
CircUSP36 downregulation mediates
ox-LDL-induced HUVEC dysfunction and inflammation by upregulating
miR-20a-5p
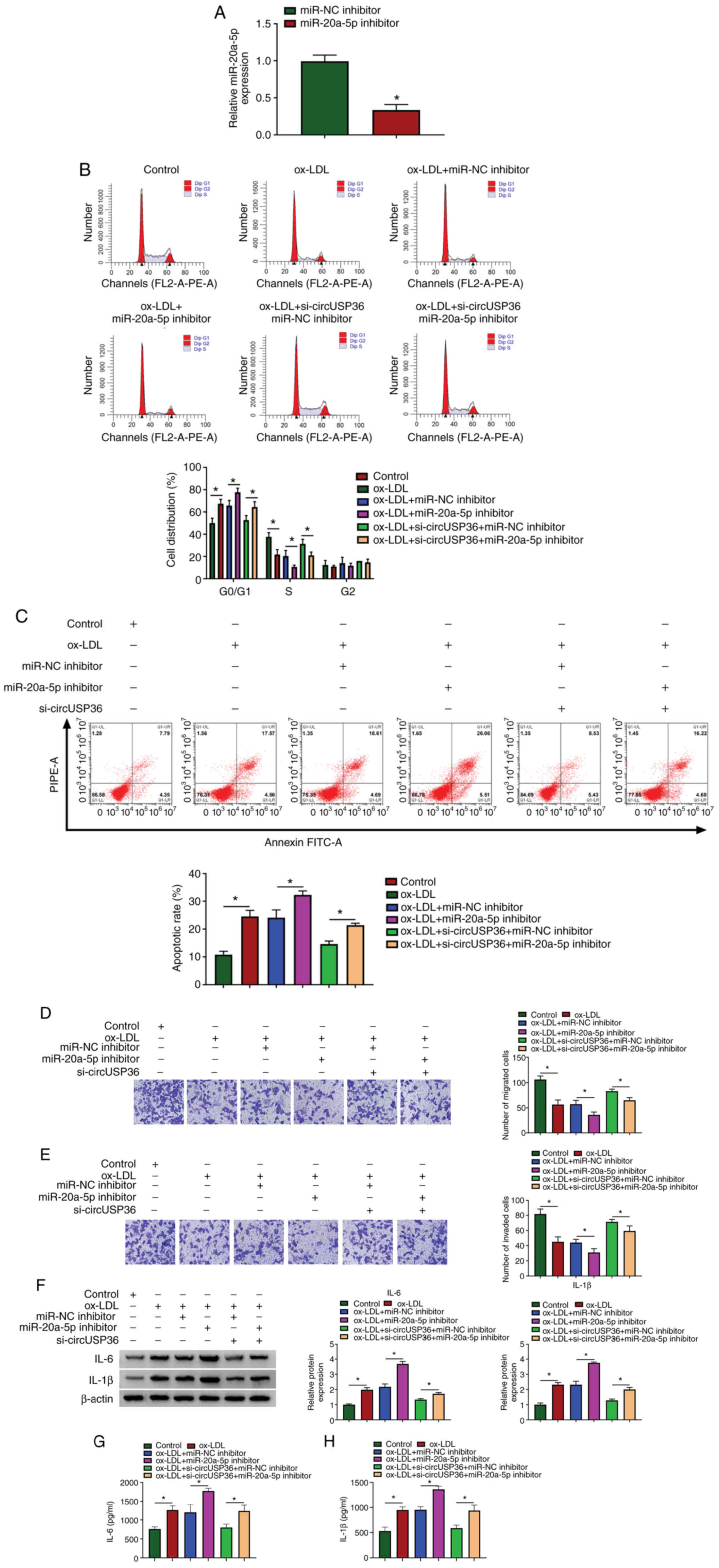
To determine whether circUSP36 targets miR-20a-5p to
regulate ox-LDL-induced apoptosis and inflammatory responses,
circUSP36 and miR-20a-5p were knocked down alone or in combination
in the ox-LDL-exposed HUVECs. The expression of miR-20a-5p was
predominantly decreased in the HUVECs following transfection with
miR-20a-5p inhibitor compared to miR-NC inhibitor (Fig. 4A). Functionally, miR-20a-5p
inhibition aggravated ox-LDL-induced cell cycle arrest and
apoptosis, and the effects of circUSP36 knockdown were partly
abolished by miR-20a-5p inhibition, leading to the promotion of
cell cycle arrest and apoptosis (Fig. 4B and C). Moreover, miR-20a-5p
inhibition further suppressed HUVEC migration and invasion which
were blocked by ox-LDL exposure, and cell migration/invasion which
were partly attenuated by circUSP36 knockdown was suppressed by
miR-20a-5p inhibition (Fig. 4D and
E). Moreover, miR-20a-5p inhibition increased the protein
levels of IL-6 and IL-1β in the HUVECs, which were stimulated by
ox-LDL exposure, and miR-20a-5p inhibition also impaired the
effects of circUSP36 knockdown, increasing the levels of IL-6 and
IL-1β (Fig. 4F). The release of
IL-6 and IL-1β in the culture medium stimulated by ox-LDL was
further promoted by transfection with miR-20a-5p inhibitor, and the
release of IL-6 and IL-1β, which was suppressed by si-circUSP36
transfection was partly enhanced by the reintroduction of
miR-20a-5p inhibitor (Fig. 4G and
H). Combined with the above-mentioned data, it was thus
concluded that circUSP36 exerts its functions in ox-LDL-exposed
HUVECs by targeting miR-20a-5p.
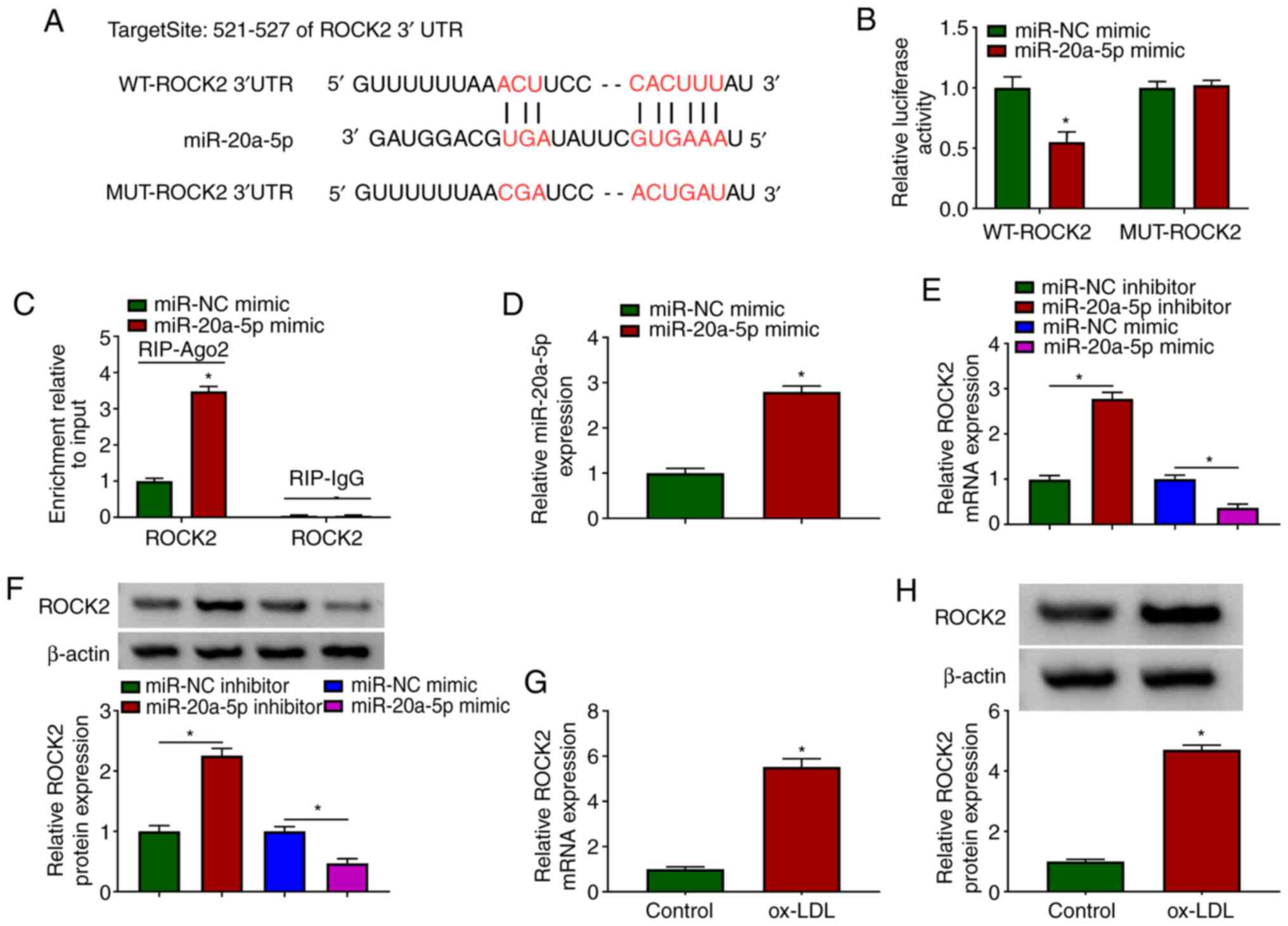
miR-20a-5p binds to ROCK2 3′UTR and
negatively regulates ROCK2 expression
To further address the functional mechanism of
circUSP36, mRNAs targeted by miR-20a-5p were identified. The
bioinformatics tool, TargetScan revealed that miR-20a-5p bound to
ROCK2 through several targeting sites (Fig. 5A). Likewise, their interaction
was validated by dual-luciferase reporter assay and RIP assay. The
data indicated that the reintroduction of miR-20a-5p mimic
significantly diminished the luciferase activity in HUVECs
transfected with WT-ROCK2, but did not affect the luciferase
activity in the HUVECs transfected with MUT-ROCK2 (Fig. 5B). In addition, transfection with
miR-20a-5p mimic markedly enriched ROCK2 in the RIP-Ago2 group, but
not in the RIP-IgG group (Fig.
5C). In the HUVECs transfected with miR-20a-5p mimic, it was
found that the expression of miR-20a-5p was notably increased
(Fig. 5D). It was then found
that the expression of ROCK2 in the HUVECs transfected with
miR-20a-5p inhibitor was substantially enhanced, while it was
markedly decreased in the HUVECs transfected with miR-20a-5p mimic
(Fig. 5E and F), suggesting that
miR-20a-5p negatively regulated ROCK2. Moreover, the expression of
ROCK2 was aberrantly elevated in the ox-LDL-exposed HUVECs
(Fig. 5G and H). Collectively,
ROCK2 was found to be a target of miR-20a-5p, and its expression
was negatively regulated by miR-20a-5p.
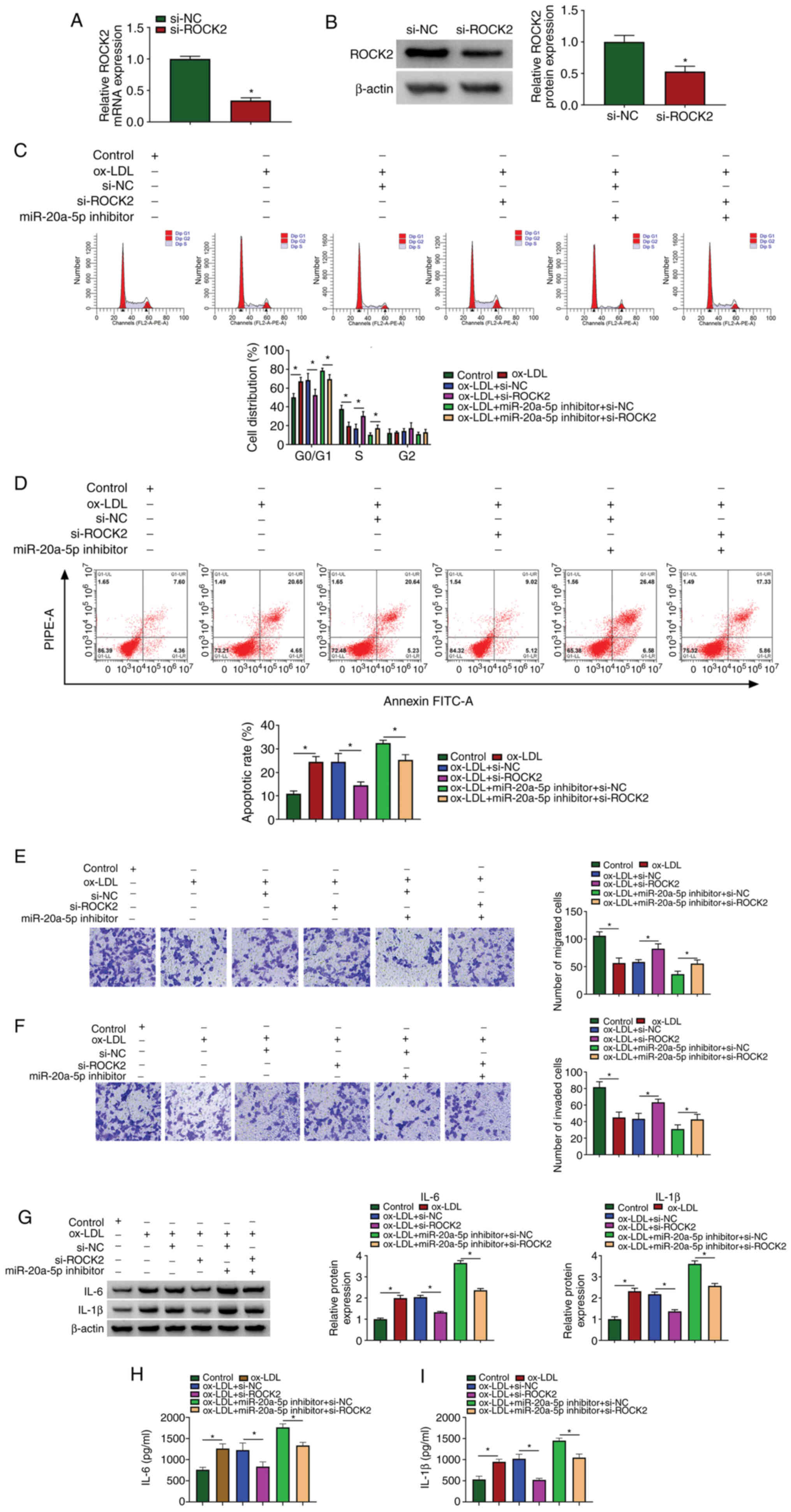
miR-20a-5p inhibition aggravates
ox-LDL-induced HIVEC injury and inflammation by increasing the
expression of ROCK2
The roles of miR-20a-5p and ROCK2 in ox-LDL-induced
HUVEC injury and were then investigated. siRNA targeting ROCK2 was
used to downregulate the expression of ROCK2, and the data from
RT-qPCR and western blot analysis revealed that the expression of
ROCK2 was notably decreased in the HUVECs following transfection
with si-ROCK2 (Fig. 6A and B).
Cell cycle arrest and cell apoptosis induced by ox-LDL exposure
were attenuated by transfection with si-ROCK2. In addition, cell
cycle arrest and cell apoptosis, which were aggravated in the
ox-LDL-exposed HUVECs transfected with miR-20a-5p inhibitor were
partly suppressed by the reintroduction of si-ROCK2 (Fig. 6C and D). Cell migration and cell
invasion, which were blocked by ox-LDL exposure, were recovered by
ROCK2 downregulation, and ROCK2 downregulation also partly reversed
the effects of miR-20a-5p inhibition, promoting cell migration and
invasion (Fig. 6E and F). The
protein levels of IL-6 and IL-1β stimulated in the ox-LDL-exposed
HUVECs were blocked by transfection with si-ROCK2, and transfection
with si-ROCK2 also reversed the effects of miR-20a-5p inhibitor,
and partly reduced the levels of IL-6 and IL-1β (Fig. 6G). The release of IL-6 and IL-1β
in the culture medium was also impaired by ROCK2 knockdown, and the
release of IL-6 and IL-1β induced by miR-20a-5p inhibition was
largely suppressed by ROCK2 knockdown (Fig. 6H and I). These data indicated
that ROCK2 knockdown attenuated ox-LDL-induced HUVEC injury and
inflammation, and ROCK2 knockdown also reversed the effects of
miR-20a-5p inhibition.
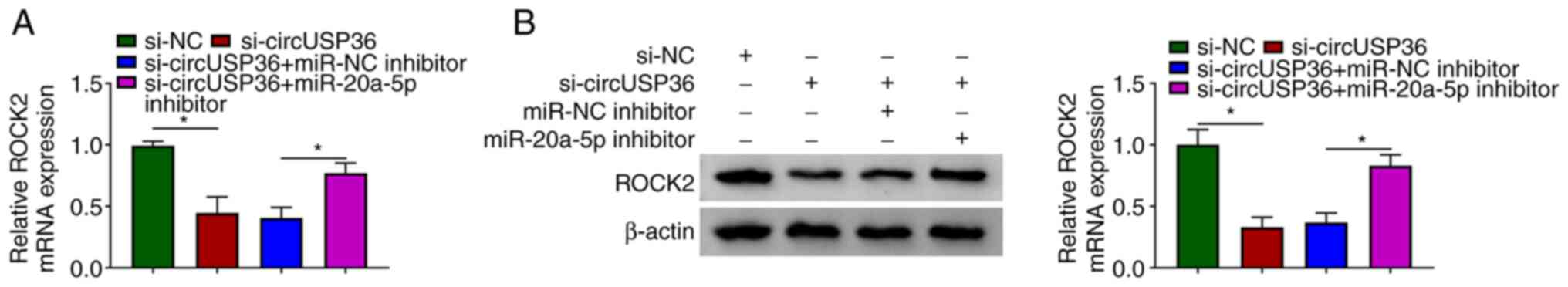
CircUSP36 indirectly regulates ROCK2
expression by targeting miR-20a-5p
Notably, it was found that the expression of ROCK2
was markedly decreased in the HUVECs following transfection with
si-circUSP36 compared to si-NC, but was partly recovered in the
HUVECs following transfection with si-circUSP36 + miR-20a-5p
inhibitor compared to si-circUSP36 + miR-NC inhibitor at both the
mRNA and protein level (Fig. 7).
These data illustrate that circUSP36 functions as miR-20a-5p sponge
to modulate the expression of ROCK2.
Discussion
The pathogenesis of AS is complex, and the
characteristics of AS include the uncontrolled proliferation of
smooth muscle cells (SMCs), necrosis and fibrosis, local
inflammatory responses and endothelial damage (22). The present study focused on the
molecular mechanisms of endothelial cell injury. The findings
mainly demonstrated that circUSP36 was dysregulated in
ox-LDL-exposed HUVECs, exhibiting a high expression. The silencing
of circUSP36 attenuated ox-LDL-induced HUVEC injury and
dysfunction, resulting in the inhibition of cell cycle arrest, cell
apoptosis and inflammatory responses, and in the activation of cell
migration and invasion. Mechanistically, circUSP36 functioned in
the HUVECs by targeting the miR-20a-5p/ROCK2 axis.
Several circRNAs have been functionally identified
in ox-LDL-exposed cell models (13,23). For example, circ_0003575 has been
found to be highly expressed in ox-LDL-stimulated HUVECs, and its
downregulation restores the ability of HUVEC proliferation and
angiogenesis (13). The reduced
expression of circANRIL has been shown to block mouse vascular
endothelial cell apoptosis and inflammatory factor expression, thus
preventing the progression of coronary AS (24). As for circUSP36, it was
identified to be highly expressed in ox-LDL-exposed HUVECs in a
previous study (13), and Zhang
et al and Liu et al reported that circUSP36
(circ_0003204) reintroduction suppressed the proliferation,
migration, angiogenesis and tube formation of human aortic
endothelial cells (HAECs) or HUVECs exposed to ox-LDL (25,26). Consistent with these findings,
the results of the present study demonstrated that circUSP36
knockdown attenuated ox-LDL-induced cell cycle arrest and
apoptosis, recovered ox-LDL-blocked cell migration and invasion,
and reduced ox-LDL-induced inflammatory responses. All these
functional effects suggested that circUSP36 aggravates the
progression of AS.
Cytoplasmic circRNAs harbor binding sites with
miRNAs and serve as competing endogenous RNAs (ceRNAs) to sponge
miRNAs (27). CircUSP36 has been
demonstrated to be mainly distributed in the cytoplasm (25). The present study thus screened
and validated the target miRNAs of circUSP36. Through the current
analyses, miR-20a-5p was identified as a target of circUSP36 by
dual-luciferase reporter assay and RIP assay. Moreover, a previous
study documented the role and mechanisms of miR-20a-5p in AS in
vitro, and the data indicated that miR-20a-5p expression was
suppressed by the exposure of HAECs to ox-LDL, and miR-20a-5p
restoration attenuated ox-LDL-induced inflammatory injury (28). Another study reported that
miR-20a-5p was weakly expressed in coronary artery disease, and
that miR-20a-5p agomir suppressed hydrogen peroxide-induced
apoptosis of HUVECs (29).
Consistently, miR-20a-5p in the present study was downregulated in
ox-LDL-exposed HUVECs, and its inhibition aggravated
ox-LDL-triggered HUVEC dysfunction and inflammatory injury.
The circRNA-miRNA-mRNA network has been found to be
an important mode involving in the pathogenesis of human diseases
(11). In the presen study, to
construct the same network of circUSP36, the potential target mRNAs
of miR-20a-5p were obtained. The current analysis indicated that
miR-20a-5p directly bound to ROCK2 3′UTR to degrade the expression
of ROCK2. Reports pf the role of ROCK2 in AS are abundant. Zhou
et al demonstrated that ROCK2 regulated the formation of
foam cells and contributed to the development of AS (30). Mattaliano et al indicated
that ox-LDL exposure catalyzed the dynamic combination of ROCK2 and
ox-LDL receptor-1 (LOX-1) and stimulated the activity of ROCK2, and
ROCK2 suppression impaired ox-LDL-induced inflammation in
endothelial cells (31). Huang
et al demonstrated that ROCK2 knockdown blocked apoptosis
and increased cytoskeleton stability of HUVECs, thus ameliorating
HUVEC dysfunction (32). In
brief, ROCK2 contributes to HUVEC dysfunction and AS progression
through multiple mechanisms. The data of the present study
demonstrated that ROCK2 silencing attenuated ox-LDL-induced HUVEC
cycle arrest, apoptosis and inflammation, but promoted HUVEC
migration and invasion which had been blocked by ox-LDL. Moreover,
ROCK2 expression was activated by circUSP36 through miR-20a-5p,
thereby participating in ox-LDL-induced HUVEC dysfunction.
However, the present study is only a preliminary
study of the role of circUSP36 in ox-LDL-induced HUVEC dysfunction.
Some effects of circUSP36, such as oxidative stress, were not
investigated in the present study. Moreover, the similar role of
circUSP36 in the progression of AS was not examined in vivo.
Therefore, further extensive in-depth studies are required,
including oxidative stress detection and animal experiments.
In conclusion, the present study identified that
circUSP36 regulated ox-LDL-induced HUVEC dysfunction and provided
the circUSP36/miR-20a-5p/ROCK2 network as a novel mechanism for
circUSP36 involvement in these processes. The present study
provides a molecular basis for the involvement of circUSP36 in the
pathogenesis of AS, and the targeting of the
circUSP36/miR-20a-5p/ROCK2 axis may prove to be a novel strategy
against AS.
Supplementary Data
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
JM and BW designed the study. RS and YW conducted
the reviewing and editing of the manuscript and were involved in
the design of the study. All the authors had full access to all the
data in the study and took responsibility for the integrity of the
data and the accuracy of the data analysis. JM supervised the
study. JM, BW and YW wrote the manuscript. RS revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Libby P and Hansson GK: Inflammation and
immunity in diseases of the arterial tree: Players and layers. Circ
Res. 116:307–311. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li B, Li W, Li X and Zhou H: Inflammation:
A novel therapeutic target/direction in atherosclerosis. Curr Pharm
Des. 23:1216–1227. 2017. View Article : Google Scholar
|
|
3
|
Gimbrone MA Jr and García-Cardeña G:
Endothelial cell dysfunction and the pathobiology of
atherosclerosis. Circ Res. 118:620–636. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang J, Wang Y, Li Y, Wang S and Zhu X:
Platelet-rich plasma protects HUVECs against oX-LDL-induced injury.
Open Med (Wars). 13:41–52. 2018. View Article : Google Scholar
|
|
5
|
Kattoor AJ, Goel A and Mehta JL: LOX-1:
Regulation, signaling and its role in atherosclerosis. Antioxidants
(Basel). 8:2182019. View Article : Google Scholar
|
|
6
|
Mitra S, Deshmukh A, Sachdeva R, Lu J and
Mehta JL: Oxidized low-density lipoprotein and atherosclerosis
implications in anti-oxidant therapy. Am J Med Sci. 342:135–142.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin J, Hou X and Yang S: microRNA-338-3p
promotes ox-LDL-induced endothelial cell injury through targeting
BAMBI and activating TGF-β/Smad pathway. J Cell Physiol.
234:11577–11586. 2019. View Article : Google Scholar
|
|
8
|
Peng J, Tang ZH, Ren Z, He B, Zeng Y, Liu
LS, Wang Z, Wei DH, Zheng XL and Jiang ZS: TET2 protects against
oxLDL-Induced HUVEC dysfunction by upregulating the CSE/H2S system.
Front Pharmacol. 8:4862017. View Article : Google Scholar :
|
|
9
|
Zeng X, Lin W, Guo M and Zou Q: A
comprehensive overview and evaluation of circular RNA detection
tools. PLoS Comput Biol. 13:e10054202017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen I, Chen CY and Chuang TJ: Biogenesis,
identification, and function of exonic circular RNAs. Wiley
Interdiscip Rev RNA. 6:563–579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bayoumi AS, Aonuma T, Teoh JP, Tang YL and
Kim IM: Circular noncoding RNAs as potential therapies and
circulating biomarkers for cardiovascular diseases. Acta Pharmacol
Sin. 39:1100–1109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pan RY, Zhao CH, Yuan JX, Zhang YJ, Jin
JL, Gu MF, Mao ZY, Sun HJ, Jia QW, Ji MY, et al: Circular RNA
profile in coronary artery disease. Am J Transl Res. 11:7115–7125.
2019.PubMed/NCBI
|
|
13
|
Li CY, Ma L and Yu B: Circular RNA
hsa_circ_0003575 regulates oxLDL induced vascular endothelial cells
proliferation and angiogenesis. Biomed Pharmacother. 95:1514–1519.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feinberg MW and Moore KJ: MicroRNA
regulation of atherosclerosis. Circ Res. 118:703–720. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dlouhá D and Hubáček JA: Regulatory RNAs
and cardiovascular disease- with a special focus on circulating
microRNAs. Physiol Res. 66(Suppl 1): S21–S38. 2017. View Article : Google Scholar
|
|
16
|
Wojciechowska A, Braniewska A and
Kozar-Kamińska K: MicroRNA in cardiovascular biology and disease.
Adv Clin Exp Med. 26:865–874. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao G, Chen W, Liu M, Yan X and Yang P:
Circulating MicroRNAs as novel potential biomarkers for left
ventricular remodeling in postinfarction heart failure. Dis
Markers. 2019:50938032019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Muendlein A, Geiger K, Leiherer A, Saely
CH, Fraunberger P and Drexel H: Evaluation of the associations
between circulating microRNAs and kidney function in coronary
angiography patients. Am J Physiol Renal Physiol. 318:F315–F321.
2020. View Article : Google Scholar
|
|
19
|
Shahbazi R, Baradaran B, Khordadmehr M,
Safaei S, Baghbanzadeh A, Jigari F and Ezzati H: Targeting ROCK
signaling in health, malignant and non-malignant diseases. Immunol
Lett. 219:15–26. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okamoto R, Li Y, Noma K, Hiroi Y, Liu PY,
Taniguchi M, Ito M and Liao JK: FHL2 prevents cardiac hypertrophy
in mice with cardiac-specific deletion of ROCK2. FASEB J.
27:1439–1449. 2013. View Article : Google Scholar :
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Singh RB, Mengi SA, Xu YJ, Arneja AS and
Dhalla NS: Pathogenesis of atherosclerosis: A multifactorial
process. Exp Clin Cardiol. 7:40–53. 2002.PubMed/NCBI
|
|
23
|
Yang L, Yang F, Zhao H, Wang M and Zhang
Y: Circular RNA circCHFR facilitates the proliferation and
migration of vascular smooth muscle via miR-370/FOXO1/Cyclin D1
pathway. Mol Ther Nucleic Acids. 16:434–441. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song CL, Wang JP, Xue X, Liu N, Zhang XH,
Zhao Z, Liu JG, Zhang CP, Piao ZH, Liu Y and Yang YB: Effect of
circular ANRIL on the inflammatory response of vascular endothelial
cells in a rat model of coronary atherosclerosis. Cell Physiol
Biochem. 42:1202–1212. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Song G, Yuan J, Qiao S, Xu S, Si
Z, Yang Y, Xu X and Wang A: Circular RNA circ_0003204 inhibits
proliferation, migration and tube formation of endothelial cell in
atherosclerosis via miR-370-3p/TGFbetaR2/phosph-SMAD3 axis. J
Biomed Sci. 27:112020. View Article : Google Scholar
|
|
26
|
Liu H, Ma X, Mao Z, Shen M, Zhu J and Chen
F: Circular RNA has_circ_0003204 inhibits oxLDL-induced vascular
endothelial cell proliferation and angiogenesis. Cell Signal.
70:1095952020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen M, Li W, Zhang Y and Yang J:
MicroRNA-20a protects human aortic endothelial cells from
Ox-LDL-induced inflammation through targeting TLR4 and TXNIP
signaling. Biomed Pharmacother. 103:191–197. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang D, Wang Y, Ma J, Wang W, Sun B, Zheng
T, Wei M and Sun Y: MicroRNA-20a participates in the aerobic
exercise-based prevention of coronary artery disease by targeting
PTEN. Biomed Pharmacother. 95:756–763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou Q, Mei Y, Shoji T, Han X, Kaminski K,
Oh GT, Ongusaha PP, Zhang K, Schmitt H, Moser M, et al:
Rho-associated coiled-coil-containing kinase 2 deficiency in bone
marrow-derived cells leads to increased cholesterol efflux and
decreased atherosclerosis. Circulation. 126:2236–2247. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mattaliano MD, Wooters J, Shih HH and
Paulsen JE: ROCK2 associates with lectin-like oxidized LDL
receptor-1 and mediates oxidized LDL-induced IL-8 production. Am J
Physiol Cell Physiol. 298:C1180–1187. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang L, Dai F, Tang L, Bao X, Liu Z,
Huang C, Zhang T and Yao W: Distinct roles for ROCK1 and ROCK2 in
the regulation of oxldl-mediated endothelial dysfunction. Cell
Physiol Biochem. 49:565–577. 2018. View Article : Google Scholar : PubMed/NCBI
|