Introduction
Air pollution is one of the most severe threats to
human health worldwide, particularly lung health (1). Fine particulate matter
(PM2.5) with an average aerodynamic diameter of <2.5
µm can directly access the alveoli of the lungs and induce
airway inflammation (2). The
airway epithelium acts as a mechanical and immunologic barrier and
is the first point of contact for air pollution in the lungs
(3). Recent studies have
suggested that the pro-inflammatory effects of PM2.5 on
airway epithelium are associated with the disruption of oxidation
reduction homeostasis, such as by inducing reactive oxygen species
(ROS) generation (4-6).
Exposure to particulate matter allows ROS to
accumulate in airway epithelial cells, and excess ROS triggers
mitogen-activated protein kinase signaling cascades and the
activation of redox-sensitive nuclear factor erythroid 2-related
factor 2 (NRF2) and nuclear factor-κB. Under normal conditions,
NRF2 is anchored in the cytoplasm through its interaction with
Kelch-like ECH-associated protein 1 (KEAP1). Oxidants interfere
with this interaction, resulting in the nuclear localization of
NRF2, which then promotes the transcription of several antioxidant
and detoxifying enzymes, such as heme oxygenase-1 (HO-1) and
NAD(P)H:quinone oxidoreductase l (NQO-1) (7). Previous studies reported that the
exposure of cells to a low concentration of PM2.5 (2
µg/cm2) induced the activity of NRF2 and the
transcription of target genes. However, the induction of NRF2 genes
was reduced in cells exposed to a high concentration of
PM2.5 (10 µg/cm2) (2,5).
In the early stages of inflammation-mediated tissue injury, the
activation of NRF2/antioxidant response element (ARE) inhibits the
production of inflammatory factors, including cytokines, chemokines
and cellular adhesion molecules (8). Thus, NRF2 plays important roles in
particulate matter-induced cell and tissue injury by regulating the
expression of target genes. However, the mechanisms of NRF2 in
PM2.5-induced tissue injury remain unknown.
Curcumin, a natural polyphenolic compound derived
from the rhizomes of Curcuma longa (turmeric), has
demonstrated anti-inflammatory and antioxidant properties in a
number of diseases (9). In
cancer, curcumin reduces cisplatin-related ototoxic adverse effects
by targeting the p-STAT3 and NRF2 signaling pathways in vivo
(10). Furthermore, curcumin
induces the translocation of NRF2 and promotes the expression of
ARE-related genes to mediate the antioxidant response (10). Thus, it was hypothesized that
curcumin may prevent PM2.5-induced oxidative
stress-related injury by upregulating the NRF2/ARE pathways.
To validate this hypothesis, the present study
evaluated the potential mechanisms of action of NRF2 in
PM2.5-induced tissue injury by investigating the
proliferation and apoptosis of BEAS-2B cells exposed to
PM2.5. In addition, the expression of NRF2 and
inflammatory factors was analyzed. Furthermore, cells exposed to
PM2.5 were treated with curcumin to determine whether
curcumin could alleviate PM2.5-induced oxidative stress.
The findings of the present study provide new insight into the
development of treatments against tissue injury caused by air
pollution-derived PM2.5.
Materials and methods
Reagents
The BEAS-2B cell line was purchased from the China
Center for Type Culture Collection. Dulbecco's modified Eagle's
medium (DMEM) and fetal bovine serum (FBS) were purchased from
Gibco; Thermo Fisher Scientific, Inc. PM2.5 was
purchased from Wuxi NEST Biotechnology Co., Ltd. Curcumin was
purchased from Shanghai Aladdin Biochemical Technology Co., Ltd.
and dissolved in dimethyl sulfoxide (DMSO). Trypsin-EDTA (0.25%),
the cell counting kit-8 (CCK-8), Hoechst 33258, kits to measure the
levels of ROS, the Annexin V-FITC/PI apoptosis kit, BCA protein
quantification kit, goat anti-rabbit IgG, and enzyme-linked
immunosorbent assay (ELISA) kits for interleukin (IL)-25, IL-33,
IL-9, interferon (IFN)-γ, IL-6, tumor necrosis factor (TNF)-α,
vascular endothelial growth factor (VEGF)-A, IL-5 and IL-13 were
purchased from Bioswamp Biotechnology Co., Ltd. Lipofectamine 2000
and TRIzol reagent were purchased from Invitrogen; Thermo Fisher
Scientific, Inc. The SYBR Green PCR kit was purchased from KAPA
Biosystems. The reverse transcription reagent kit was purchased
from Takara Biotechnology Co., Ltd. All primary antibodies were
purchased from Abcam. Polyvinylidene difluoride (PVDF) membranes
and enhanced chemiluminescence (ECL) reagents were obtained from
EMD Millipore.
Cells, cell culture, PM2.5 and
curcumin treatment
BEAS-2B cells were maintained in DMEM with 10% FBS
at 37°C in a humidified atmosphere containing 5% CO2.
Cells were treated with PM2.5 at concentrations of 0,
12.5, 25, 50 and 100 mg/ml for 24 h to analyze the intracellular
response. Cells were treated with curcumin at concentrations of 0,
1, 2.5, 5, 7.5, 10, 20 and 50 µM for 48 h for CCK-8 assay
(11,12). To evaluate the effects of
curcumin, the cells were incubated in DMEM without or with curcumin
(10 µM) for 1 h (10) and
treated with PM2.5 at 25 and 50 µg/ml for 24
h.
Cell viability assessment
Cell viability was evaluated by CCK-8 assay
according to the manufacturer's protocol. BEAS-2B cells were seeded
into 96-well plates (5×103 cells/well) and maintained in
regular growth medium overnight. The cells were then treated with
PM2.5 or curcumin as described above. CCK-8 solution (10
µl) was added to each well, and after 4 h, the absorbance
was measured at 450 nm using a Multiskan FC microplate photometer
(Thermo Fisher Scientific, Inc.). The experiment was performed in
triplicate.
Apoptosis assessment
The apoptosis of BEAS-2B cells was quantified by
flow cytometric analysis using the Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI) apoptosis kit. Treated
cells were washed with cold phosphate-buffered saline (PBS),
trypsinized and harvested following centrifugation at 1,000 × g for
5 min at 4°C. The cells were resuspended in cold PBS, washed,
diluted with cold PBS (1×105 cells/ml), and harvested
following centrifugation at 1,000 × g for 5 min at 4°C. The cells
were re-suspended in 100 µl of binding buffer and stained
with 5 µl of Annexin V-FITC and 5 µl of PI for 30 min
at 4°C in the dark. The fluorescence intensity of the stained cells
was measured using an FC500 MCL flow cytometer (Beckman Coulter,
Inc.). The data were analyzed using CXP analysis software (CXP
Analysis 2.0, Beckman Coulter, Inc.). The experiment was performed
in triplicate.
Hoechst staining
After treatment, the cells were fixed in 4%
formaldehyde for 10 min. After 2 washes in PBS, Hoechst 33258
solution was dropped onto the cells and incubated for 5 min in the
dark at 4°C. The cells were then observed and photographed under a
TS100-F microscope (Nikon Corporation).
Measurement of intracellular ROS
production
Intracellular ROS production was measured using the
fluorogenic dye 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA).
Following treatment, the cells were washed and incubated with
DCFH2-DA (10 µmol/l) for 20 min at 37°C in the dark. The
fluorescence intensity of the stained cells was analyzed using an
FC500 MCL flow cytometer, and the data were analyzed using CXP
analysis software (CXP Analysis 2.0). The experiment was performed
in triplicate.
Luciferase reporter assay
pmirGLO (Addgene, Inc.) containing ARE (5′-TCA CAG
TGA CTC AGC AAA ATT-3′) reporter plasmids were constructed as
previously described (13).
BEAS-2B cells were seeded into 24-well plates and incubated for 24
h. The pmirGLO/ARE plasmids were transfected into the cells using
Lipofectamine 2000 according to the manufacturer's instructions.
pmirGLO plasmids (without ARE) transferred into BEAS-2B cells were
considered as the control group. Following 24 h of transfection,
the transfection efficiency was validated by RT-qPCR, and cells
were treated with PM2.5 or curcumin for 24 h. The activities of
Firefly and Renilla luciferase were detected using the
Dual-luciferase Reporter Assay System (GeneCopoeia) and evaluated
using a SynergyH multiscan spectrum spectrophotometer (BioTek
Instruments, Inc.).
Western blot analysis
Following treatment, the cells were lysed in
radioimmunoprecipitation assay lysis buffer supplemented with a
protease and phosphatase inhibitor cocktail. Protein concentrations
were determined using a bicinchoninic acid protein quantification
kit. The proteins (20 µg) were subjected to sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto
polyvinylidene fluoride membranes. The membranes were blocked in 5%
dry milk for 2 h and incubated overnight at 4°C with primary
antibodies (Table I). Following
3 washes with PBS/Tween (PBST), the membranes were incubated with
goat anti-rabbit IgG secondary antibodies (1:10,000, PAB150011,
Bioswamp Biotechnology Co., Ltd.) for 1 h at 4°C. The blots were
washed 3 times with PBST. The resulting immunoreactive protein
complexes were detected using an ECL reagent according to the
manufacturer's instructions and imaged using a Tanon-5200 auto
chemiluminescence analyzer (Tanon Science & Technology Co.,
Ltd.) and analyzed using Tanon Gis image analysis software (Version
4.2, Tanon Science & Technology Co., Ltd.).
 | Table IInformation of primary antibodies
used for western blot analysis. |
Table I
Information of primary antibodies
used for western blot analysis.
| Name | Species | Dilution rate | Size (kDa) | Catalogue nos. | Supplier |
|---|
| NRF2 | Rabbit | 1:2,000 | 68 | ab62352 | Abcam |
| HO-1 | Rabbit | 1:2,000 | 33 | ab13248 | Abcam |
| NQO-1 | Rabbit | 1:1,000 | 31 | ab34173 | Abcam |
| KEAP1 | Rabbit | 1:1,000 | 70 | ab139729 | Abcam |
| GAPDH | Rabbit | 1:1,000 | 37 | 2118 | Cell Signaling
Technology |
| β-actin | Rabbit | 1:1,000 | 42 | ab8227 | Abcam |
| Histone H3 | Rabbit | 1:1,000 | 17 | ab8580 | Abcam |
ELISA
BEAS-2B cells were seeded into 24-well plates and
cultured for 24 h. Following 24 h of treatment, the supernatants
were collected and the release of cytokines (IL-25, IL-33, IL-9,
IFN-γ, IL-6, TNF-α, VEGF-A, IL-5, IL-10 and IL-13) was analyzed
using ELISA kits according to the manufacturer's instructions. The
absorbance of the wells was measured using a Multiskan MS apparatus
(Thermo Fisher Scientific, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using Trizol reagent, and 1
µg of total RNA was reverse transcribed using the
PrimeScript™ RT reagent kit (KM4101, KAPA Biosystems) with gDNA
Eraser according to the manufacturer's instructions (KAPA
Biosystems). qPCR was conducted using the SYBR-Green PCR kit in a
real-time system (Bio-Rad Laboratories, Inc.). The primers used are
listed in Table II.
Amplification was conducted at 95°C for 3 min, followed by 40
cycles at 95°C for 5 sec, 56°C for 10 sec, and 72°C for 25 sec; and
65°C for 5 sec, 95°C for 50 sec, and 40°C for 60 sec. The data were
analyzed using the 2−ΔΔCq method (14). GADPH was used as an
internal reference. The experiment was performed in triplicate.
| Table IISequences of primers used for
RT-qPCR. |
Table II
Sequences of primers used for
RT-qPCR.
| Name | Sequence
(5′-3′) | Size (bp) |
|---|
| IL-25-F |
GGAGATATGAGTTGGA | 144 |
| IL-25-R |
TCTGGTTGTGGTAGAG | |
| IL-33-F |
AGAGAAACCACCAAAA | 114 |
| IL-33-R |
TGATATACCAAAGGCA | |
| IL-9-F |
AGTTGTCTCTGTTTGGGC | 104 |
| IL-9-R |
AGTGGGTATCTTGTTTGC | |
| IFN-γ-F |
CTCTTTTCTTAGGCATTT | 192 |
| IFN-γ-R |
CATCTCGTTTCTTTTTGT | |
| IL-6-F |
TGGTCTTTTGGAGTTTGA | 130 |
| IL-6-R |
ATTCTTTGCCTTTTTCTG | |
| TNF-α-F |
TACTCCTCACCCACACCA | 152 |
| TNF-α-R |
GAAGACCCCTCCCAGATA | |
| VEGFA-F |
AAGACAAGAAAATCCCTG | 134 |
| VEGFA-R |
GTTCGTTTAACTCAAGCT | |
| IL-5-R |
CATAAAAATCACCAACTG | 160 |
| IL-5-R |
GTCTTTCTTCTCCACACT | |
| IL-13-F |
CCACGGTCATTGCTCTCA | 150 |
| IL-13-R |
TGCTCCATACCATGCTGC | |
| GAPDH-F |
CCACTCCTCCACCTTTG | 106 |
| GAPDH-R |
CACCACCCTGTTGCTGT | |
Statistical analysis
The data are presented as the means ± standard
deviation (SD). The significant difference between 2 groups was
analyzed by an unpaired Student's t-test and those between multiple
groups by one-way analysis of variance (ANOVA) followed by Tukey's
post-hoc test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
PM2.5 exposure-induced
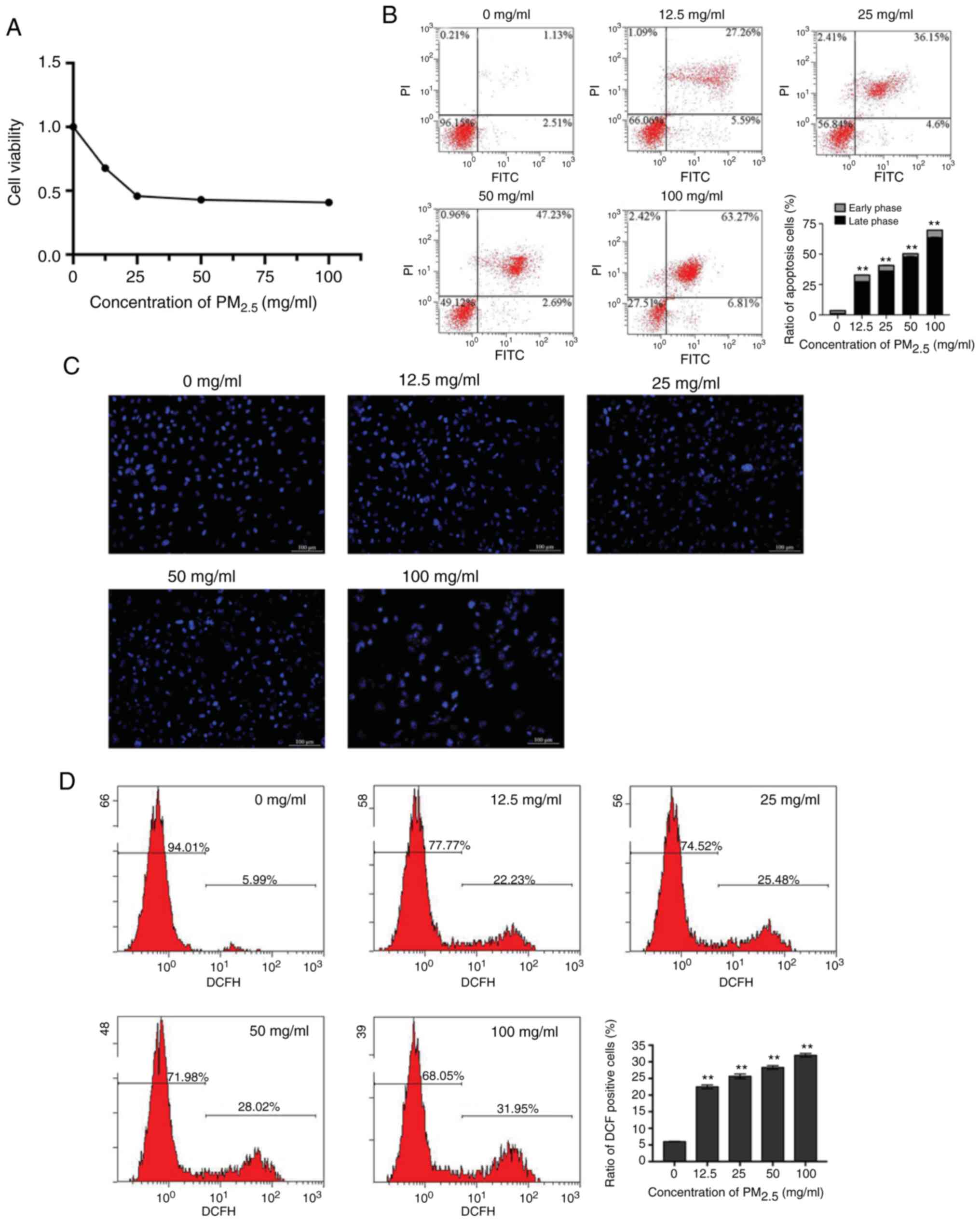
oxidative damage inhibits the viability of BEAS-2B cells
To determine the effects of PM2.5 on the
viability of lung epithelial cells, BEAS-2B cells were exposed to
PM2.5 at 0, 12.5, 25, 50 and 100 mg/ml. The results of
CCK-8 assay revealed that the proliferation of the BEAS-2B cells
was inhibited by PM2.5 in a concentration-dependent
manner (Fig. 1A). The apoptosis
of the BEAS-2B cells was examined by flow cytometry and Hoechst
33258 staining. The percentage of apoptotic cells was significantly
elevated by PM2.5 (Fig.
1B). Microscopic observations revealed that chromatin
condensation and marginalization appeared in the
PM2.5-exposed cells in a concentration-dependent manner
(Fig. 1C). The results suggested
that PM2.5 exposure induced the apoptosis of lung
epithelial cells, and these effects were more evident at a higher
concentration of PM2.5 compared with the lower
concentrations. To determine the association between the cellular
redox level and the inhibitory effects of PM2.5 on the
growth of BEAS-2B cells, the levels of intracellular ROS were
measured. The results revealed that intracellular ROS production
was increased by PM2.5 in a concentration-dependent
manner (Fig. 1D).
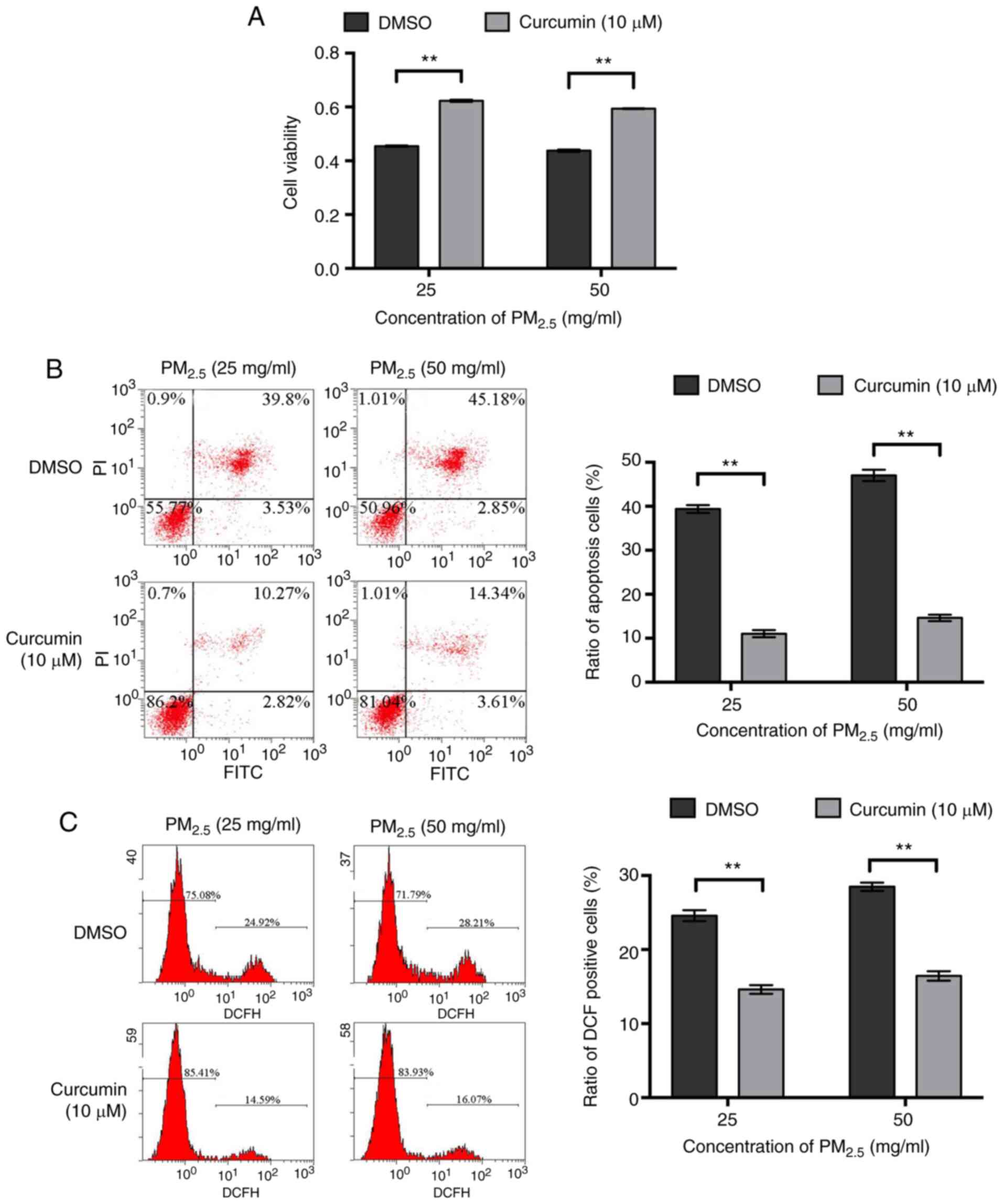
Curcumin promotes the survival and
reduces the oxidative stress of PM2.5-exposed BEAS-2B
cells
The effects of curcumin on the viability of BEAS-2B
cells were then detected and it was found that within a certain
concentration range (0-10 µM), curcumin promoted cell
proliferation in a concentration-dependent manner (Fig. S1A). Hoechst 33258 staining
revealed that curcumin did not affect cell apoptosis (Fig. S1B). The viability of the BEAS-2B
cells pre-treated with curcumin was significantly higher than that
of the cells pre-treated with DMSO following exposure to the same
concentration of PM2.5 (Fig. 2A). Flow cytometry also revealed
that following exposure to the same concentration of
PM2.5, the percentage of apoptosis was lower when the
cells were pre-treated with curcumin (Fig. 2B). In addition, following
exposure to the same concentration of PM2.5, the levels
of intracellular ROS in the cells pre-treated with curcumin were
significantly lower than those in the cells pre-treated with DMSO
(Fig. 2C). These results suggest
that pre-treatment with curcumin protected the BEAS-2B cells from
undergoing apoptosis induced by PM2.5 and promoted cell
survival by attenuating oxidative stress in
PM2.5-exposed cells.
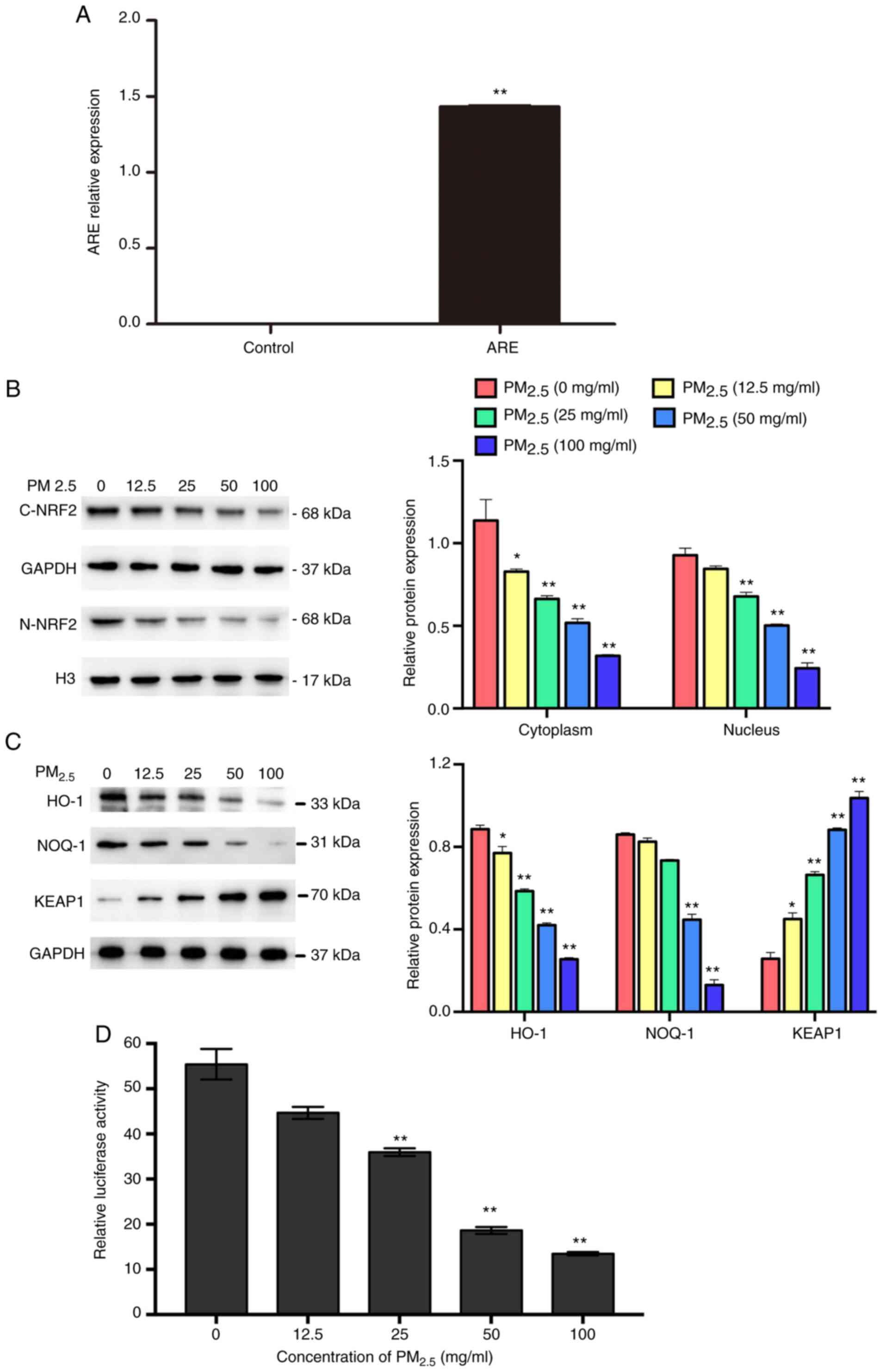
Curcumin activates NRF2/ARE pathways
As shown in Fig.
3A, the expression of ARE was significantly higher in the
ARE-transfected group compared with the control group, suggesting
that the ARE luciferase reporter vector was successfully
transfected into the BEAS-2B cells. The results of western blot
analysis revealed that PM2.5 exposure downregulated the
expression of NRF2 (Fig. 3B),
suppressed the activation of ARE and other downstream proteins
(HO-1 and NQO-1), and increased the expression of KEAP1 (Fig. 3C and D). To further examine
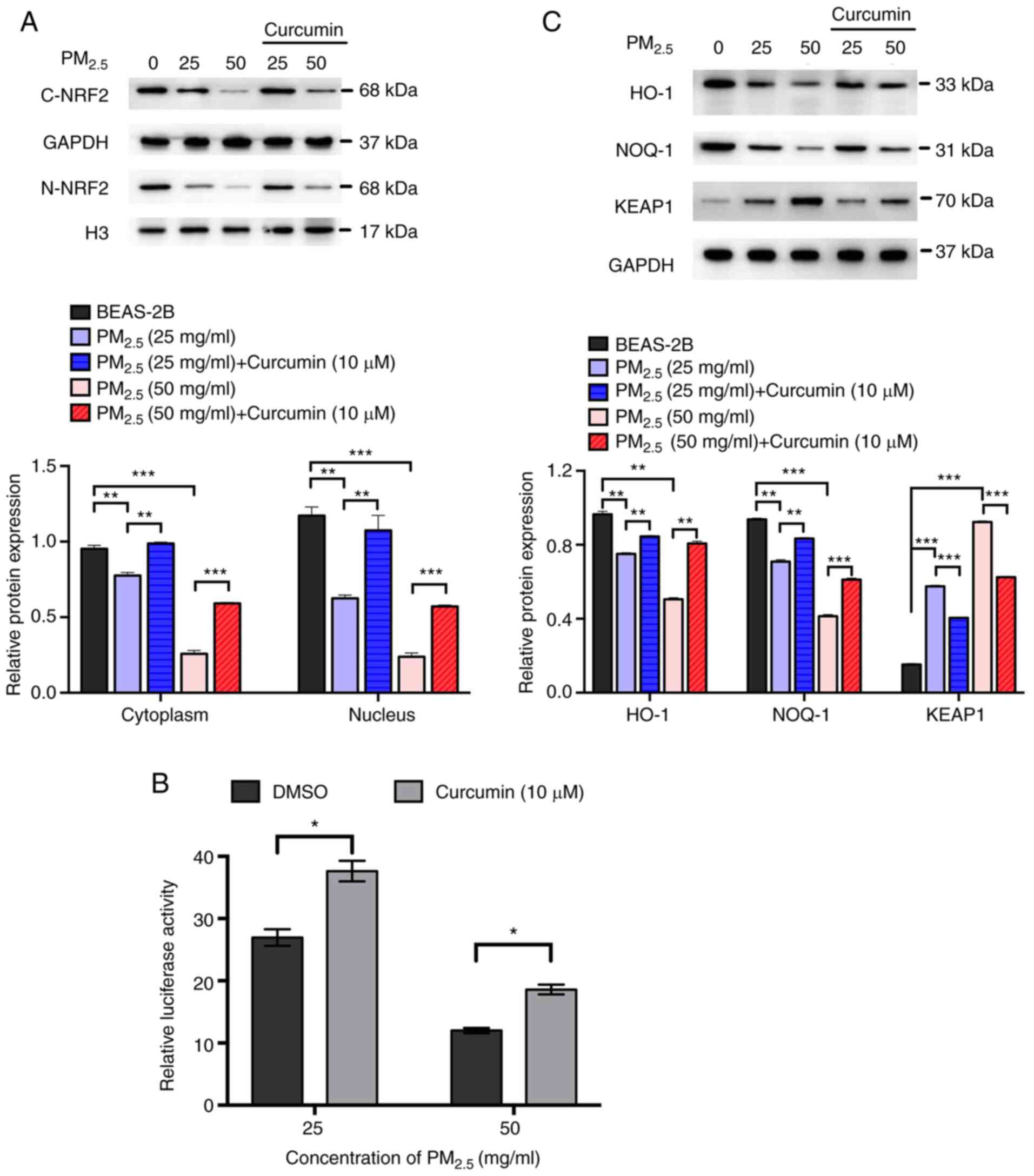
whether curcumin reduces oxidative stress by regulating the
activation of NRF2/ARE pathways, the expression of NRF2 and
downstream proteins in cells pre-treated with curcumin was
analyzed. The results of western blot analysis revealed that
curcumin pre-treatment significantly increased the expression of
NRF2 in the cell cytoplasm and nucleus (Fig. 4A). The luciferase activity assay
revealed that ARE activation was significantly increased by
curcumin pre-treatment (Fig.
4B). Furthermore, the expression of HO-1 and NQO-1 was
significantly elevated, whereas that of KEAP1 was reduced by
curcumin pre-treatment (Fig.
4C). These results suggested that curcumin attenuated oxidative
stress in PM2.5-exposed cells by activating the NRF2/ARE
pathways.
Curcumin inhibits inflammatory factor
production induced by PM2.5 exposure in BEAS-2B
cells
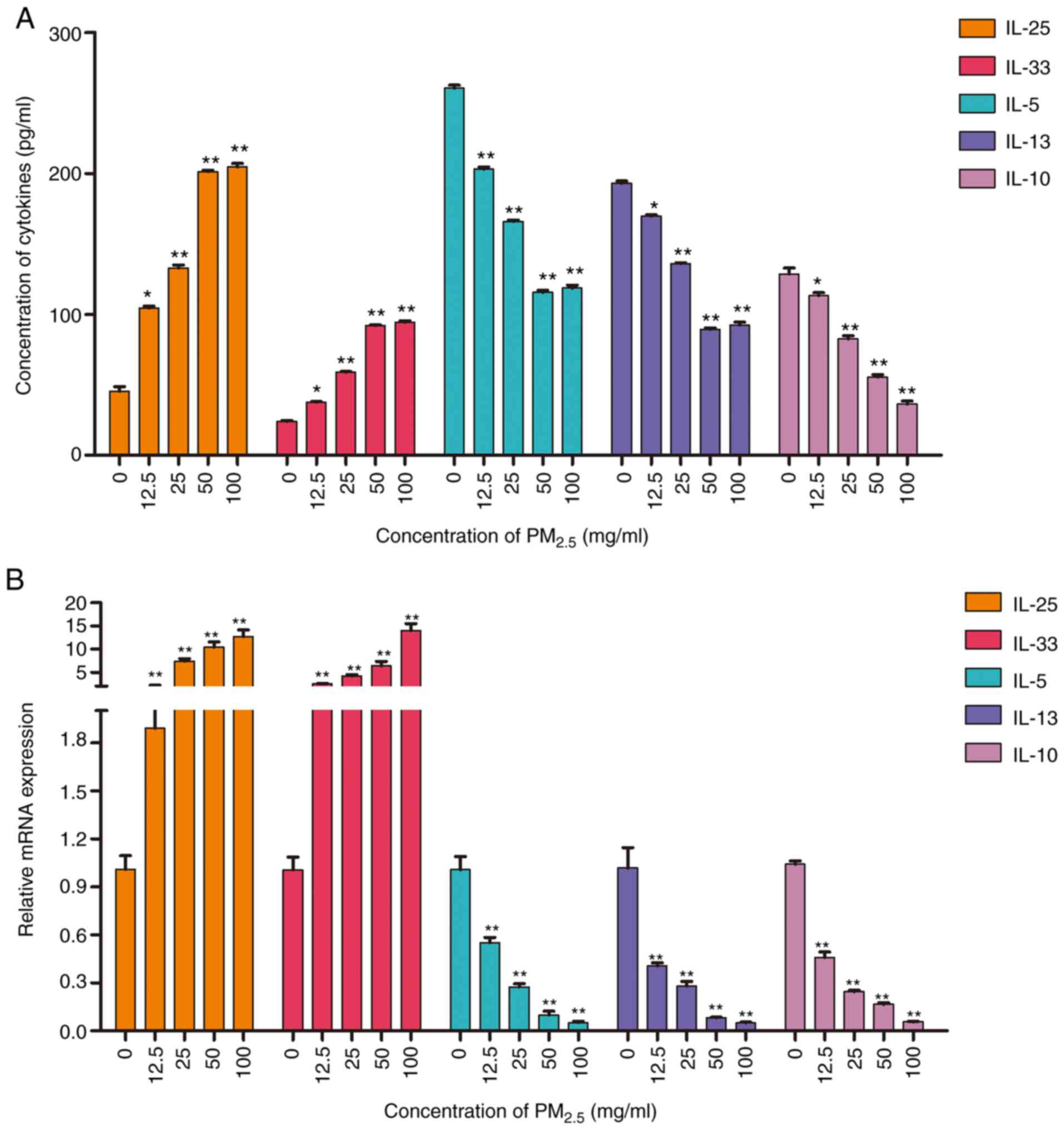
To assess the pro-inflammatory effects of
PM2.5, the production and expression of inflammatory
factors in PM2.5-exposed BEAS-2B cells was evaluated by
ELISA and RT-qPCR, respectively. Both assays revealed that
PM2.5 exposure induced the upregulation of the
pro-inflammatory factors IL-25, IL-33, IL-9, IFN-γ, IL-6, TNF-α,
and VEGF-A (Figs. 5 and S2). Moreover, the expression of the
anti-inflammatory factors, IL-5, IL-10 and IL-13, was decreased by
PM2.5 exposure (Fig.
5). The results suggested that long-term exposure to
PM2.5 (for >24 h) induced a type II inflammatory
reaction in lung epithelial cells. The production and expression of
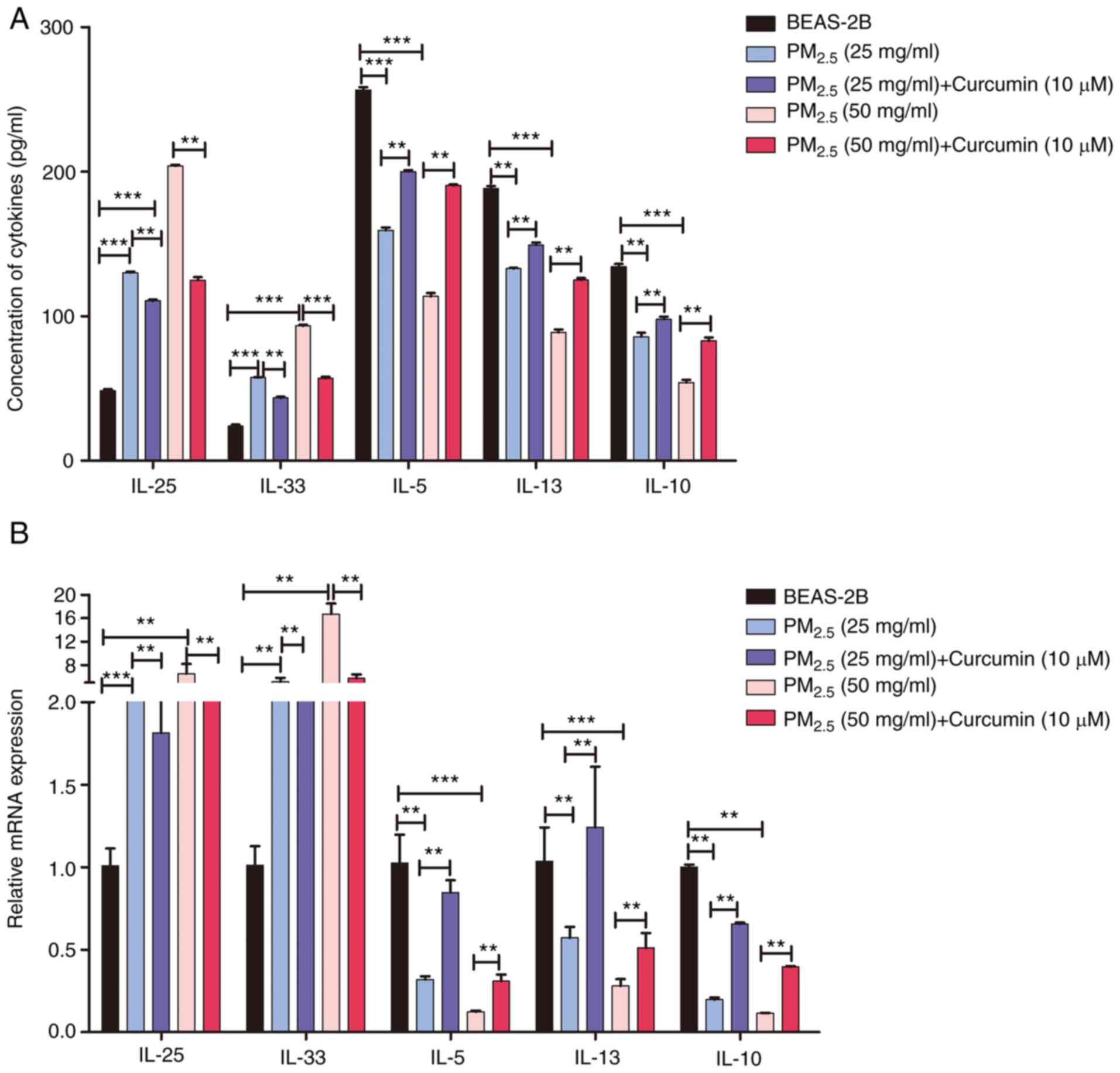
inflammatory factors in cells pre-treated with curcumin were also
analyzed. The results of RT-qPCR and ELISA revealed that curcumin
pre-treatment inhibited the expression and production of
pro-inflammatory factors (Figs.
6 and S3), and increased
the levels of anti-inflammatory factors (Fig. 6). The results suggested that
curcumin inhibited the type II inflammatory reaction induced by
PM2.5.
Discussion
Air pollution, particularly from PM2.5,
has severely threatened human health in recent decades. Previous
studies have demonstrated that oxidative stress is an important
factor in PM2.5-induced cell apoptosis (6,15). Thus, it is necessary to identify
appropriate antioxidants which may be used to protect human
bronchial epithelial cells from PM2.5 exposure. In the present
study, it was found that curcumin promoted the survival and reduced
the oxidative stress of PM2.5-exposed BEAS-2B cells by
activating NRF2/ARE pathways, subsequently inhibiting the type II
inflammatory reaction.
Mitochondria are the main target of ROS, the
over-production of which, together with cell internalization,
induces mitochondrial permeability transition pore opening and
results in mitochondrial dysfunction (7). Previous studies have indicated that
oxidative stress induced by PM2.5 disrupts the
antioxidant system and subsequently triggers
mitochondrial-dependent apoptosis by activating caspase-3 (3,16). The present study demonstrated
that PM2.5 exposure significantly increased the levels
of intracellular ROS and promoted BEAS-2B cell apoptosis,
suggesting that PM2.5-suppressed BEAS-2B cell viability
was associated with oxidative stress.
To better understand the mechanisms of ROS
accumulation in cells and the related mitochondrial dysfunction
following PM2.5 exposure, the activation of NRF2
pathways was investigated. NRF2 is a key regulator of cell redox
homeostasis that counterbalances ROS production and maintains redox
balance (17). Under normal
conditions, NRF2 is located in the cytoplasm by binding to KEAP1.
In response to oxidative stress, activated NRF2 is translocated
into the nucleus, where it binds to the ARE of the target genes of
NRF2, including HO-1 and NQO-1, and induces their expression
(18). HO-1 and NQO-1 are
important intracellular, antioxidant, anti-inflammatory and
anti-apoptotic enzymes. In the present study, it was found that
PM2.5 exposure suppressed the expression of NRF2 and
increased that of KEAP1, which inhibits the activity of NRF2
pathways. The results also revealed that the activity of ARE and
the expression of HO-1 and NQO-1 were inhibited by PM2.5
exposure. These results are in agreement with those of previous
studies (3,19). In NRF2-deficient mice, the
expression of NQO-1 and HO-1 has been shown to be absent (20). These findings indicate that
PM2.5 exposure increases the levels of intracellular ROS
by suppressing the activation of NRF2 and decreasing the expression
of downstream genes. However, the effects of NRF2 pathways on
PM2.5-exposed BEAS-2B cells remain unclear.
Inflammation is the main outcome of
PM2.5-induced cell injury (21). Airway epithelial cells secrete
various cytokines and participate in immune response after
stimulation by allergens or pathogenic microorganisms. IL-33 is
expressed in epithelial and smooth muscle cells, and its expression
level is positively related to the severity of asthma (22,23). IL-25, also known as IL-17,
belongs to the IL-17 cytokine family and induces a Th2-type immune
response (24). The
overexpression of IL-25 induces eosinophilia and B-lymphocyte
hyperplasia, and alters antibody production in mice (25). A previous study demonstrated that
PM2.5 exposure induced an innate immune cellular
response through the expression of IL-25 and IL-33, which was
reduced by Nac, a ROS-quenching agent (26). NRF2 has been reported to be an
anti-inflammatory factor as it directly inhibits the transcription
of pro-inflammatory cytokine genes (27). In the present study, following
PM2.5 exposure, the levels of pro-inflammatory factors
were significantly upregulated, whereas those of anti-inflammatory
factors were significantly downregulated. Taken together, the
results indicated that PM2.5-induced inflammation was
associated with a redox imbalance via the suppression of NRF2
pathway activation, which subsequently induced cell apoptosis.
Curcumin has been shown to have antioxidant and
anti-inflammatory properties (28,29) and has been used in the treatment
of allergies and asthma in Asia for a number of years (30). It has been proven to increase the
activity of NRF2/ARE pathways, as well as the expression of
downstream genes (HO-1 and NOQ-1) in different types of cells in
recent studies (31). In the
present study, BEAS-2B cells pre-treated with curcumin exhibited
higher levels of NRF2/ARE activation and lower levels of
intercellular ROS than untreated cells. This effect rendered
BEAS-2B cells resistant to PM2.5-induced oxidative
damage. Moreover, curcumin pre-treatment reduced the expression and
production of PM2.5-induced inflammatory factors. Thus,
the results of the present study suggest that curcumin
pre-treatment can protect BEAS-2B cells from apoptosis in response
to PM2.5-induced oxidative stress. However, elucidating
the mechanisms of NRF2 activation by curcumin in
PM2.5-exposed human bronchial epithelial cells requires
further experiments.
As mentioned above, curcumin has demonstrated to
exhibit anti-inflammatory and antioxidant properties in a number of
diseases (9), which is widely
used in the treatment of respiratory diseases, such as lung tissue
injury (32-34). These previous studies focused on
the specific mechanisms of action of curcumin in the treatment of
severe lung injury rather than its efficacy. Therefore, the present
study did not use a positive control group, which is one of the
limitations of the present study; thus, the current experiments may
not be optimal, but may be sufficient to draw a conclusion in that
curcumin protects BEAS-2B cells against PM2.5-induced oxidative
damage and inflammation, and prevents cell apoptosis by increasing
the activation of NRF2-related pathways. The effects of curcumin on
PM2.5-induced oxidative stress and inflammation were also not
detected in in vivo experiments using animals. Thus, the
authors aim to verify the present findings in in vivo
experiments and using a positive control in future studies.
In conclusion, the present study confirmed that
exposure to PM2.5 induced the apoptosis of BEAS-2B human
bronchial epithelial cells through redox imbalance and inflammation
via the suppression of NRF2 activation. Moreover, curcumin
pre-treatment protected the cells against PM2.5-induced
oxidative damage and inflammation and prevented cell apoptosis by
promoting the activation of NRF2 pathways. These findings provide a
potential treatment scheme against diseases induced by
PM2.5 exposure.
Supplementary Data
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81300008) and the Hubei
Provincial Natural Science Fund (grant no. 2013CFB372).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
SY, XLH and LNM were involved in the
conceptualization of the study. JC was involved in the study
methodology. LNM provided software. XL was involved in data
validation and provided resources. WSY was involved in formal
analysis. XJW was involved in the investigative aspects of the
study. XLH was involved in data curation. WSY, XL and GWL were
involved in the writing and preparation of the original draft and
visualization, as well as in project design and administration. SY
was involved in the writing, reviewing and editing of the
manuscript, and in study supervision, and also in funding
acquisition. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Huff RD, Carlsten C and Hirota JA: An
update on immunologic mechanisms in the respiratory mucosa in
response to air pollutants. J Allergy Clin Immunol. 143:1989–2001.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li N, Alam J, Venkatesan MI, Eiguren FA,
Schmitz D, Di SE, Slaughter N, Killeen E, Wang X, Huang A, et al:
Nrf2 is a key transcription factor that regulates antioxidant
defense in macrophages and epithelial cells: Protecting against the
proinflammatory and oxidizing effects of diesel exhaust chemicals.
J Immunol. 173:3467–3481. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Georas SN and Rezaee F: Epithelial barrier
function: At the front line of asthma immunology and allergic
airway inflammation. J Allergy Clin Immunol. 134:509–520. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guan L, Rui W, Bai R, Zhang W, Zhang F and
Ding W: Effects of size-fractionated particulate matter on cellular
oxidant radical generation in human bronchial epithelial BEAS-2B
cells. Int J Environ Res Public Health. 13:4832016. View Article : Google Scholar :
|
|
5
|
Leclercq B, Kluza J, Antherieu S, Sotty J,
Alleman LY, Perdrix E, Loyens A, Coddeville P, Lo Guidice JM,
Marchetti P and Garçon G: Air pollution-derived PM2.5
impairs mitochondrial function in healthy and chronic obstructive
pulmonary diseased human bronchial epithelial cells. Environ
Pollut. 243:1434–1449. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raudoniute J, Stasiulaitiene I,
Kulvinskiene I, Bagdonas E, Garbaras A, Krugly E, Martuzevicius D,
Bironaite D and Aldonyte R: Pro-inflammatory effects of extracted
urban fine particulate matter on human bronchial epithelial cells
BEAS-2B. Environ Sci Pollut Res Int. 25:32277–32291. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gong P, Hu B, Stewart D, Ellerbe M,
Figueroa YG, Blank V, Beckman BS and Alam J: Cobalt induces heme
oxygenase-1 expression by a hypoxia-inducible factor-independent
mechanism in Chinese hamster ovary cells: Regulation by Nrf2 and
MafG transcription factors. J Biol Chem. 276:27018–27025. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu MX, Zhu YF, Chang HF and Liang Y:
Nanoceria restrains PM2.5-induced metabolic disorder and
hypothalamus inflammation by inhibition of astrocytes activation
related NF-κB pathway in Nrf2 deficient mice. Free Radic Biol Med.
99:259–272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Motterlini R, Foresti R, Bassi R and Green
CJ: Curcumin, an anti-oxidant and anti-inflammatory agent, induces
heme oxygenase-1 and protects endothelial cells against oxidative
stress. Free Radic Biol Med. 28:1303–1312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fetoni AR, Paciello F, Mezzogori D, Rolesi
R, Eramo SL, Paludetti G and Troiani D: Molecular targets for
anticancer redox chemotherapy and cisplatin-induced ototoxicity:
The role of curcumin on pSTAT3 and Nrf-2 signalling. Br J Cancer.
113:1434–1444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qi Ni, Zeng SE, Tan N, Huang LZ, Cheng QY
and Zhang MY: Effect of curcumin on proliferation of hepatocellular
carcinoma Hep1 cells and expression of HIF-1α mRNA in normal oxygen
conditions. Liaoning Tra Chin Med. 8:21–23. 2010.
|
|
12
|
Song XJ, Zhou HY, Sun YX and Huang HC:
Inhibitory effects of curcumin on H2O2-induced cell damage and APP
expression and processing in SH-SY5Y cells transfected with APP
gene with Swedish mutation. Mol Biol Rep. 47:2047–2059. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Woods CG, Fu J, Xue P, Hou Y, Pluta LJ,
Yang L, Zhang Q, Thomas RS, Andersen ME and Pi J: Dose-dependent
transitions in Nrf2-mediated adaptive response and related stress
responses to hypochlorous acid in mouse macrophages. Toxicol Appl
Pharmacol. 238:27–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Suo D, Zeng S, Zhang J, Meng L and Weng L:
PM2.5 induces apoptosis, oxidative stress injury and
melanin metabolic disorder in human melanocytes. Exp Ther Med.
19:3227–3238. 2020.PubMed/NCBI
|
|
16
|
Li X, Ding Z, Zhang C, Zhang X, Meng Q, Wu
S, Wang S, Yin L, Pu Y and Chen R: MicroRNA-1228(*) inhibit
apoptosis in A549 cells exposed to fine particulate matter. Environ
Sci Pollut Res Int. 23:10103–10113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li N and Nel AE: Role of the Nrf2-mediated
signaling pathway as a negative regulator of inflammation:
Implications for the impact of particulate pollutants on asthma.
Antioxid Redox Signal. 8:88–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang LF, Su SW, Wang L, Zhang GQ, Zhang R,
Niu YJ, Guo YS, Li CY, Jiang WB, Liu Y and Guo HC:
Tert-butylhydroquinone ameliorates doxorubicin-induced
cardiotoxicity by activating Nrf2 and inducing the expression of
its target genes. Am J Transl Res. 7:1724–35. 2015.PubMed/NCBI
|
|
19
|
Block ML and Calderón-Garcidueñas L: Air
pollution: Mechanisms of neuroinflammation and CNS disease. Trends
Neurosci. 32:506–516. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang S, Jiao Y, Li C, Liang X, Jia H, Nie
Z and Zhang Y: Dimethyl itaconate alleviates the inflammatory
responses of macrophages in sepsis. Inflammation. Oct 7–2020.Epub
ahead of print. View Article : Google Scholar
|
|
21
|
Jia H, Liu Y, Guo D, He W, Zhao L and Xia
S: PM2.5-induced pulmonary inflammation via activating
of the NLRP3/caspase-1 signaling pathway. Environ Toxicol. Sep
30–2020.Epub ahead of print.
|
|
22
|
Ravanetti L, Dijkhuis A, Dekker T, Sabogal
Pineros YS, Ravi A, Dierdorp BS, Erjefält JS, Mori M, Pavlidis S,
Adcock IM, et al: IL-33 drives influenza-induced asthma
exacerbations by halting innate and adaptive antiviral immunity. J
Allergy Clin Immunol. 143:1355–1370.e16. 2019. View Article : Google Scholar
|
|
23
|
Gorska K, Nejman-Gryz P, Paplinska-Goryca
M, Korczynski P, Prochorec-Sobieszek M and Krenke R: Comparative
study of IL-33 and IL-6 levels in different respiratory samples in
mild-to-moderate asthma and COPD. COPD. 15:36–45. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang SH, Reynolds JM, Pappu BP, Chen G,
Martinez GJ and Dong C: Interleukin-17C promotes Th17 cell
responses and autoimmune disease via interleukin-17 receptor E.
Immunity. 35:611–621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Swaidani S, Bulek K, Kang Z, Liu C, Lu Y,
Yin W, Aronica M and Li X: The critical role of epithelial-derived
Act1 in IL-17- and IL-25-mediated pulmonary inflammation. J
Immunol. 182:1631–1640. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bao ZJ, Fan YM, Cui YF, Sheng YF and Zhu
M: Effect of PM2.5 mediated oxidative stress on the
innate immune cellular response of Der p1 treated human bronchial
epithelial cells. Eur Rev Med Pharmacol Sci. 21:2907–2912.
2017.PubMed/NCBI
|
|
27
|
Keleku LN, Suzuki M and Yamamoto M: An
overview of the advantages of KEAP1-NRF2 system activation during
inflammatory disease treatment. Antioxid Redox Signal.
29:1746–1755. 2018. View Article : Google Scholar
|
|
28
|
Menon VP and Sudheer AR: Antioxidant and
anti-inflammatory properties of curcumin. Adv Exp Med Biol.
595:105–125. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Merrell JG, McLaughlin SW, Tie L,
Laurencin CT, Chen AF and Nair LS: Curcumin-loaded
poly(epsilon-caprolactone) nanofibres: Diabetic wound dressing with
anti-oxidant and anti-inflammatory properties. Clin Exp Pharmacol
Physiol. 36:1149–1156. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang X, Lv JN, Li H, Jiao B, Zhang QH,
Zhang Y, Zhang J, Liu YQ, Zhang M, Shan H, et al: Curcumin reduces
lung inflammation via Wnt/β-catenin signaling in mouse model of
asthma. J Asthma. 54:335–340. 2017. View Article : Google Scholar
|
|
31
|
Lin X, Bai D, Wei Z, Zhang Y, Huang Y,
Deng H and Huang X: Curcumin attenuates oxidative stress in
RAW264.7 cells by increasing the activity of antioxidant enzymes
and activating the Nrf2-Keap1 pathway. PLoS One. 14:e02167112019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng LZ, Chen LW, Hu XY, Lian J, Zhao GJ,
Hong GL, Lu ZQ and Qiu QM: Curcumin upregitates mitochondrial
fusion protein 2 to relieve acute lung injury in sepsis mice. Chin
J Burns. 29:58–64. 2020.
|
|
33
|
Qin K and Guo XJ: Research progress on the
mechanism of curcumin in the treatment of chronic obstructive
pulmonary disease. Chin J Respirat Cri Care. 19:99–103. 2020.
|
|
34
|
Qin Z, Wang B, Tan ZX and Luo SH: Curcumin
inhibits the TLR4/HMGB1 pathway to protect lipopolysaccharide from
inducing acute lung injury. Chin J Clin Thor Cardiovasc Sur.
27:89–92. 2020.
|