Introduction
Mushrooms have been widely used and consumed as part
of the human diet in numerous countries for centuries due to their
high nutritional values, since they contain high protein and fiber
contents with small amounts of fat (1). Additionally, mushrooms are sources
of minerals, such as calcium, zinc and magnesium, and vitamins
(2). Recently, the medicinal
properties of mushrooms have attracted increasing attention.
Polysaccharides are the main active ingredients of edible mushrooms
(3). Moreover, mushroom
polysaccharides, which have been isolated from the fruiting bodies,
mycelia and culture media, serve an important role in the growth
and development of fungal organisms and have abundant activities,
such as antioxidant, anti-inflammatory, anticancer, antidiabetic
and immunomodulatory effects (4).
Glucans are considered as one of the most essential
polysaccharides in mushrooms, based on their diversified chemical
structures and marked medicinal effects (5). Glucans have been discovered in
numerous Basidiomycetes in the form of α-1,3-D-glucans
(6), α-1,4-D-glucans (7), β-1,3-D-glucans (8) and β-1,6-D-glucans (9). The differences in structure among
these glucans may affect their biological properties. A recent
study has revealed that α-1,6-D-glucans and β-1,3-D-glucans
isolated from the basidiome and the mycelium of Pleurotus
albidus differentially inhibit lipid-induced inflammation and
pro-inflammatory lipid-laden macrophage (foam cell) formation in
macrophage-like cells (10).
Additionally, it has been demonstrated that linear β-(1→3)-D-glucan
from Cordyceps militaris exerts an anti-inflammatory effect
in vitro and in vivo (11). The linear (1→6)-β-D-glucan of
Agaricus bisporus has an effect on the expression levels of
pro-inflammatory genes and significantly inhibits the production of
inflammatory cytokines caused by lipopolysaccharide (LPS) (9). Furthermore, a branched
1,3-β-D-glucan, which is obtained from Sparassis crispa,
stimulates leukocytes of DBA/2 mice and secretes cytokines in
vitro, indicating its immunological activity (12). Previous studies have reported
that β-1,3-D-glucan with a branching degree (DB) between 0.20 and
0.33 present the most potent antitumor effects (13-15), while the β-1,3-D-glucan with a DB
that is too high or too low is not effective (16). The linear α-1,3-glucan and
branched β-1,3/1,6-glucan from the stems of Pleurotus
eryngii and Pleurotus ostreatus exhibit potential
prebiotic activity by regulating the growth of
Lactobacillus, Bifidobacterium and
Enterococcus (17).
Macrophages are innate immune cells present in every
tissue and are necessary for normal tissue development, homeostasis
and repair of damaged tissues (18). Activated macrophages can
phagocytose and neutralize cancer cells by secreting nitric oxide
(NO), TNF-α and IL-6 cytokines. Polysaccharides have been reported
to exert immune and antitumor activities by regulating macrophage
activities (19,20). For example, a polysaccharide from
Ganoderma sinense (GSP-2) specifically upregulates the
protein expression levels of Toll-like receptor 4 and activates the
MAPK signaling pathway in RAW246.7 macrophages. Moreover, GSP-2
exerts its immunomodulatory activity by inducing the secretion of
the cytokines TNF-α, IL-1β and IL-6 (21). Additionally, it has been reported
that a water-soluble β-D-glucan obtained from Hericium
erinaceus is able to induce the proliferation of lymphocytes
and improve the expression levels of inflammatory cytokines
produced by THP-1 macrophages (22).
Cantharellus cibarius is a common wild edible
mushroom belonging to the phylum Basidiomycota.
Polysaccharides are one of the main active ingredients of C.
cibarius (23). A linear
α-1,6-D-mannan and a branched α-1,6-D-mannan substituted with
mannan side chains can be isolated from the fruiting bodies of
C. cibarius (24-26). Our previous study indicated that
linear 3-O-methylated galactan isolated from C. cibarius
activated macrophages and modulated the antitumor immune response
by converting tumor-associated macrophages towards an M1-like
phenotype (27,28). In addition, C. cibarius
contains various types of glucans. α-1,6-D-glucan with
β-1,4-D-Glcp side chains is extracted using boiling water,
and β-1,3-D-glucan branched at O-6 and a β-1,6-D-glucan with single
and short side chains can be separated from the hot aqueous NaOH
fraction (26,29). The present study reported the
extraction and purification of a novel acidic β-glucan from the
fruiting bodies of C. cibarius and characterized its
structure. The immunomodulatory activity of the acidic β-glucan was
further investigated.
Materials and methods
Materials
Fruiting bodies of C. cibarius were purchased
from the Guilin Road Market (Changchun, China) and were identified
using rDNA-Internal Transcribed Spacer sequencing analysis
(27). Anion-exchange
chromatography (DEAE-cellulose) was purchased from Amersham
(Cytiva). Sepharose CL-6B was purchased from Cytiva, while LPS and
Polymyxin B sulfate (PMB) were obtained from Sigma-Aldrich (Merck
KGaA). TNF-α (cat. no. EK0527) and IL-6 ELISA kits (cat. no.
EK0411) were obtained from Boster Biological Technology. Nembutal
was acquired from Sinopharm Chemical Reagent Co., Ltd. SB203580
(cat. no. S1863), U0126 (cat. no. S1901) and SP600125 (cat. no.
S1876) were acquired from Beyotime Institute of Biotechnology. All
of the other reagents were of analytical grade or higher.
Extraction and purification of the
polysaccharides
Fruiting bodies of C. cibarius were first
extracted with distilled water at 100°C for 4 h (1:25 w/v),
followed by another extraction at 100°C for 2 h (1:20 w/v). The hot
water extracts were concentrated under a vacuum at 60°C and
precipitated using 4× volumes of 95% ethanol at room temperature
for 12 h. The precipitate was collected via centrifugation (3,200 ×
g for 15 min) at 25°C and re-dissolved in water, dialyzed and
lyophilized to obtain the polysaccharide named water-soluble C.
cibarius polysaccharide (WCCP). WCCP was dissolved in distilled
water, added to a DEAE-cellulose column (8.0×20 cm; Cl−)
pre-equilibrated with distilled water for anion-exchange
chromatography (27) and eluted
with distilled water to yield a neutral polysaccharide fraction
(WCCP-N) or 0.3 M NaCl to obtain an acidic polysaccharide fraction
(WCCP-A). WCCP-A was further purified via gel-permeation
chromatography (Sepharose CL-6B), the eluate was collected and the
absorbance was measured at 490 nm for total sugar to give a
homogeneous fraction (WCCP-A-b), as previously described by Yang
et al (27).
General methods
The total carbohydrate content was determined using
the phenol-sulfuric acid protocol with glucose as the standard
(30). Uronic acid content was
determined using the colorimetric method with glucuronic acid as
the standard (31). Protein
content was determined using the Bradford assay with BSA (VWR
International, LLC) as the standard (32).
Homogeneity and molecular weight
determination
Molecular weight distributions were determined using
high-performance gel permeation chromatography (HPGPC) with a
TSK-gel G-3000PWXL (7.8×300 mm; Tosoh Corporation) coupled to a
Shimadzu high-performance liquid chromatography (HPLC) system
(LC-10ATvp pump and refractive index RID-10A detector) as described
by Zhang et al (33). The
column was pre-calibrated using standard dextrans (50, 25, 12, 5
and 1 kDa) and linear regression.
Monosaccharide composition analysis
Monosaccharide composition was determined using HPLC
as described by Zhang et al (33). Briefly, polysaccharide samples (2
mg) were first hydrolyzed with 1 ml anhydrous methanol containing
2M HCl at 80°C for 16 h and then with 1 ml 2M trifluoroacetic acid
at 120°C for 1 h. Following derivatization with
1-phenyl-3-methyl-5-pyrazo-lone, the derivatives were analyzed
using a Shimadzu HPLC system.
Fourier transform infrared (FT-IR)
spectroscopy
Polysaccharides were ground with KBr powder and
turned into a pellet that was ~1 mm for subsequent FT-IR analysis,
as previously described (34).
FT-IR spectra were obtained with a Spectrum Two FT-IR spectrometer
in the range of 4,000-400 cm−1 (PerkinElmer, Inc.).
Methylation analysis
Methylation analysis was conducted according to the
method of Needs and Selvendran (35). Uronic acid was firstly reduced
into neutral sugars using NaBD4 prior to methylation
analysis. Subsequently, the reduced polysaccharide (5 mg) was
dissolved in DMSO (0.5 ml) and methylated with a suspension of
NaOH/DMSO (0.5 ml) and iodomethane (1.0 ml). The reaction mixture
was extracted with CH2Cl2, and then the
solvent was removed via vacuum evaporation. Complete methylation
was confirmed by the disappearance of the -OH band (3,200-3,400
cm−1) in the FT-IR spectrum. The per-O-methylated
polysaccharide was subsequently hydrolyzed using HCOOH (85%; 1 ml)
for 4 h at 100°C and then CF3COOH (2 M; 1 ml) for 6 h at
100°C. The partially methylated sugars in the hydrolysate were
reduced using NaBH4 and were then acetylated. The
resulting alditol acetates were analyzed using gas
chromatography-mass spectrometry (GC-MS; 7890B-5977B; Agilent
Technologies, Inc.) with a DB-35 ms capillary column (30 m × 0.32
mm × 0.25 mm), as previously described (27). The oven temperature was programed
from 120°C (hold for 1 min) to 210°C (hold for 2 min) at 3°C/min,
then up to 260°C (hold for 4 min) at 10°C/min. The temperature of
both the inlet and detector was 300°C. Helium was used as the
carrier gas. The mass scan range was 50.0-500.0 m/z.
Nuclear magnetic resonance (NMR)
analysis
1H, 13C, heteronuclear
singular quantum correlation (HSQC) and heteronuclear multiple bond
correlation (HMBC) NMR spectra were recorded at 20°C on a Bruker
Avance 600 MHz spectrometer (Bruker Corporation) with a Bruker 5-mm
broadband probe, operating at 600 MHz for 1H NMR and 150
MHz for 13C NMR. Polysaccharides (20.0 mg) were
dissolved in D2O (0.5 ml) and centrifuged at 10,462 × g
for 3 min at 25°C to remove any undissolved polysaccharide. Data
were analyzed using standard Bruker software (MestReNova v10.0;
Bruker Corporation).
Cell culture
RAW264.7 cells were purchased from the American Type
Culture Collection. Cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) high glucose medium supplemented with 100
U/ml penicillin, 100 μg/ml streptomycin and 10%
heat-inactivated FBS (Gibco; Thermo Fisher Scientific, Inc.).
RAW264.7 cells were maintained at 37°C in a 5% CO2
incubator.
Animals
Specific-pathogen-free female Balb/c mice (10 mice;
age, 6-8 weeks; weight, 16-18 g) were obtained from Beijing HFK
Bioscience Co., Ltd. Animal experiments were conducted in
compliance with the Animal Management Rules of the Ministry of
Health of the People's Republic of China and were approved by the
Animal Care and Use Committee of Northeast Normal University
(Changchun, China). All animals were kept at a constant temperature
of 21°C, relative humidity of 55% and under a 12-h light-dark
cycle. The animals were maintained under pathogen-free conditions
and allowed access to food and water ad libitum. Nembutal
(50 mg/kg) was injected intraperitoneally into Balb/c mice. After
being anesthetized, mice were sacrificed by cervical dislocation. A
single cell suspension of peritoneal macrophages and bone marrow
macrophages were prepared from Balb/c mice under aseptic conditions
by frosted slides in PBS. The suspension was centrifuged at 400 × g
for 3 min at 4°C to obtain the cell pellet. After two washes in
PBS, the cells were resuspended in complete DMEM for subsequent
examinations.
Cell viability assay
RAW264.7 cells were seeded at a density of
5×104 cells/well in a 96-well plate overnight at 37°C
and were then treated with various concentrations (0, 25, 50, 100
and 200 μg/ml) of WCCP-A-b or 1 μg/ml LPS at 37°C for
24 h. The medium was removed, and 100 μl/well of MTT
solution (0.5 mg/ml) was added. After a 4-h incubation at 37°C,
supernatants were discarded and the resulting formazan was
dissolved in 100 μl DMSO. The absorbance was measured at 570
nm using a microplate reader (BioTek Instruments, Inc.; Agilent
Technologies, Inc.). Cell proliferation was expressed as the
percentage of the control, which was set to 100%. To examine the
effect of endotoxin contamination, the polysaccharides were
pre-treated with PMB (30 μg/ml) for 24 h at 37°C and then
used for cell viability assays.
Measurement of NO
RAW264.7 cells, mouse peritoneal macrophages and
bone marrow macrophages were seeded at a density of
1×105 cells/well in a 48-well plate overnight at 37°C
and were then treated with various concentrations (0, 25, 50, 100
and 200 μg/ml) of WCCP-A-b or 1 μg/ml LPS at 37°C for
24 h. After incubation, supernatants were collected and reacted
with Griess reagent as previously described (36).
Measurement of TNF-α and IL-6
RAW264.7, mouse peritoneal macrophages and bone
marrow macrophages were seeded at a density of 1×105
cells/well in a 48-well plate overnight at 37°C and were then
treated with various concentrations of WCCP-A-b (0, 25, 50, 100 and
200 μg/ml) or 1 μg/ml LPS at 37°C for 24 h. The
concentrations of TNF-α and IL-6 in the supernatants were assessed
using the aforementioned TNF-α and IL-6 ELISA kits according to the
manufacturer's instructions.
Western blotting
RAW264.7 cells were seeded at a density of
8×105 cells/well in a 6-well plate, and were then
pretreated with or without specific inhibitors (25 μM
SB203580, 25 μM U0126 or 20 μM SP600125) for 2 h at
37°C, followed by incubation with WCCP-A-b (200 μg/ml) at
37°C for 24 h. Western blot analysis was performed as described by
Meng et al (36).
RAW264.7 cells were rinsed twice with cold PBS and lysed in lysis
buffer (50 mM Tris/acetate, pH 7.4, 1 mM EDTA, 0.5% Triton X-100,
150 mM sodium chloride, 0.1 mM PMSF and Roche incomplete protease
inhibitor cocktail). Protein concentration was measured using the
Bradford method. Equal amounts of protein (30 μg/lane) were
separated via 12% SDS-PAGE and transferred to a PVDF membrane. The
membrane was blocked in 3% BSA at room temperature for 1 h and then
blotted with specific antibodies, and proteins were detected using
an electrochemiluminescence reagent (Tanon Science and Technology
Co., Ltd.). Both the primary and secondary antibody incubations
were 1 h at room temperature. Primary antibodies (1:1,000) against
phosphorylated (p)-JNK (cat. no. 4668s), JNK (cat. no. 9252s),
p-ERK (cat. no. 9101s), ERK (cat. no. 9102s), p-p38 (cat. no.
9215s) and p38 (cat. no. 9212s) were obtained from Cell Signaling
Technology, Inc. The antibody against β-actin (cat. no. 612657) was
purchased from BD Biosciences. HRP-conjugated goat anti-rabbit IgG
(cat. no. AS014) and goat anti-mouse IgG (cat. no. AS003) secondary
antibodies (1:5,000) were obtained from ABclonal Biotech Co.,
Ltd.
Statistical analysis
Data are presented as the mean ± SD from ≥3
independent experiments. Data were analyzed using one-way ANOVA
followed by Dunnett's test for comparisons among multiple groups.
Statistical analysis was performed using GraphPad Prism 8.0
software (GraphPad Prism, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results and Discussion
Preparation of polysaccharide from C.
cibarius
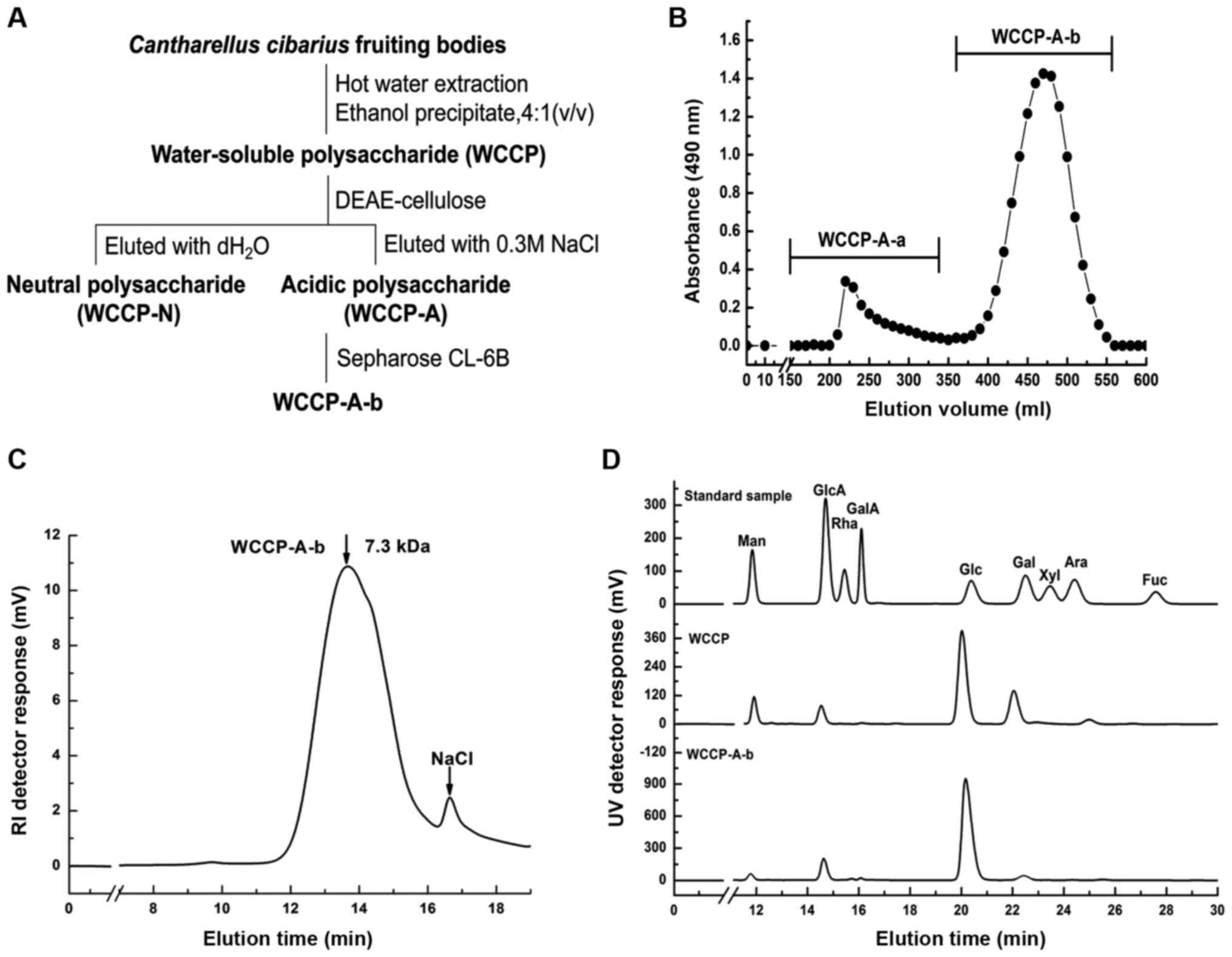
WCCP was extracted from fruiting bodies of C.
cibarius using boiling hot water, and the yield was 5.5%
relative to the dry weight of the material. WCCP contained 79.2% of
total sugar, 4.2% of uronic acids and 2.8% of protein (data not
shown). Moreover, it was composed of 56.0% glucose, 21.8%
galactose, 10.2% mannose, 8.6% glucuronic acid and minor
3-methyl-galactose (3.4%) (Fig.
1D). WCCP was separated into WCCP-N and WCCP-A using
anion-exchange chromatography (Fig.
1A). WCCP-A was further purified using gel-permeation
chromatography (Fig. 1B), and a
homogeneous fraction WCCP-A-b was obtained with the yield of 64.2%
relative to WCCP-A. The molecular weight of WCCP-A-b was ~7.3 kDa,
as determined via HPGPC (Fig.
1C). This contained glucose as the major sugar (89.7%),
followed by minor of glucuronic acid (8.8%) (Fig. 1D).
FT-IR spectrum analysis
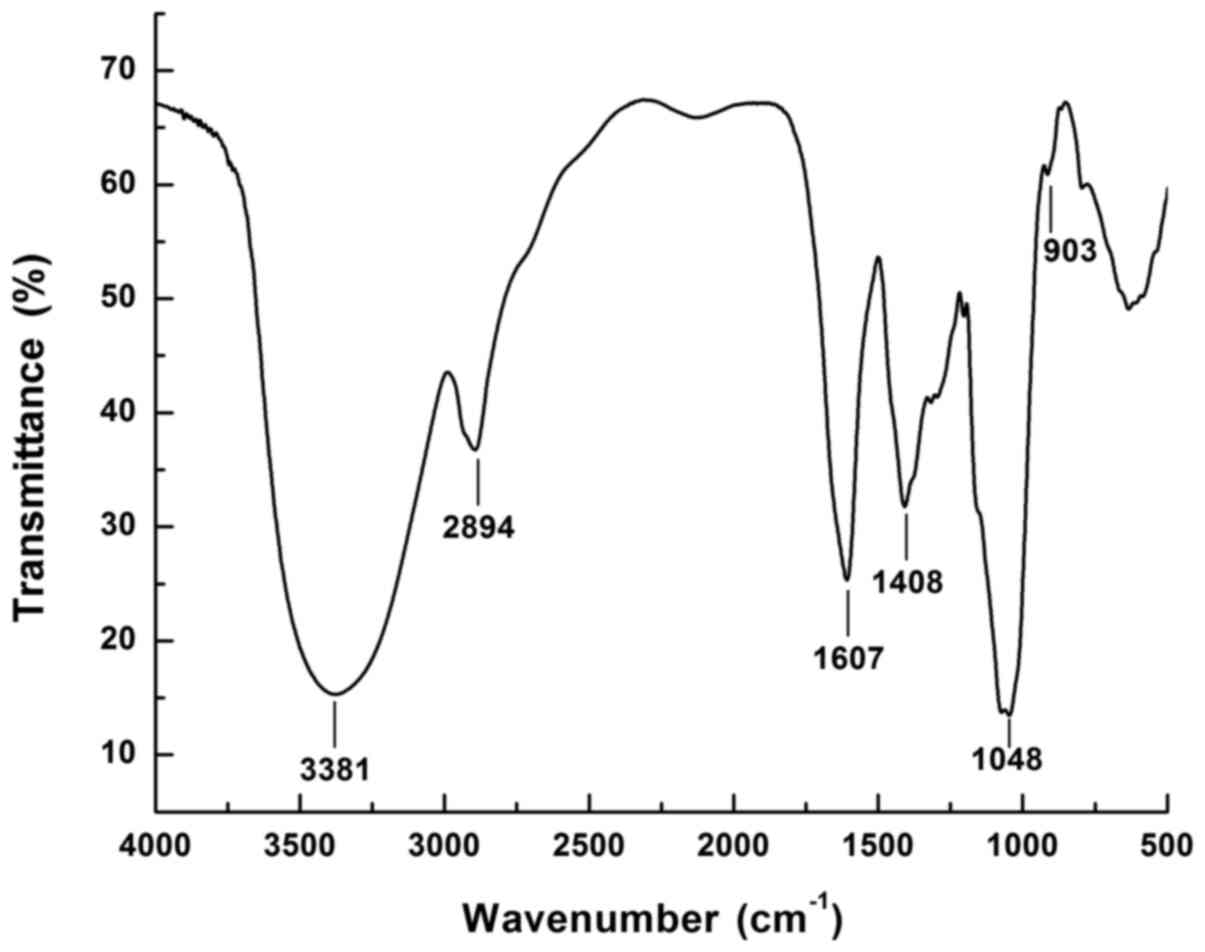
The FT-IR spectrum of WCCP-A-b is presented in
Fig. 2. The strong absorption
band at 3,381 cm−1 was attributed to the stretching
vibration of O-H. The weak band near 2,894 cm−1
indicated C-H stretching vibration. The stretching bands at ~1,607
cm−1 were observed as the bending vibration of O-H. The
band near 1,048 cm−1 suggested the presence of pyranose
ring and the weak bands at ~900 cm−1 was associated with
the presence of β-linked glycosyl residues (34).
Methylation analysis
To determine the glycosidic linkages in WCCP-A-b,
methylation analysis was performed. As WCCP-A-b contained minor
glucuronic acid, it was firstly reduced by NaBD4, then
methylated, hydrolyzed and acetylated. The partially methylated
alditol acetates were analyzed using GC-MS. As presented in
Table I, the glycosidic linkage
of glucose in WCCP-A-b was mainly in the form of 1,6-linked (57.4%)
and 1,3,6-linked (15.2%), suggesting that its backbone was
1,6-glucan, which was branched at O-3. The DB was ~20.9%. Terminal
glucose (17.3%) and 1,3-linked glucose (5.0%) were detected as side
chains (Table I). It was
indicated that terminal glucose may be linked to the backbone
through O-3 of 1,6-linked glucose or through short 1,3-linked
glucose. Moreover, 1,4-linked glucuronic acid (5.1%) residues were
detected in WCCP-A-b, which may be present in side chains (Table I).
 | Table IGlycosidic linkages of water-soluble
C. cibarius polysaccharide homogenous fraction (WCCP-A-b)
analyzed by GC-MS. |
Table I
Glycosidic linkages of water-soluble
C. cibarius polysaccharide homogenous fraction (WCCP-A-b)
analyzed by GC-MS.
| Methylated
sugars | Linkages | Molar % | Mass fragments.
m/z |
|---|
|
2,3,4-Me3-Glcp | 1,6- | 57.4 |
101,117,129,161,173,189,233 |
|
2,4-Me2-Glcp | 1,3,6- | 15.2 |
117,129,159,189,233,261,305 |
|
2,4,6-Me3-Glcp | 1,3- | 5.0 |
101,117,129,161,189,233,277 |
|
2,3,4,6-Me4-Glcp | 1- | 17.3 |
101,117,129,145,161,205 |
|
2,3,6-Me3-GlcApa | 1,4- | 5.1 |
101,117,129,161,235 |
Structure analysis by NMR spectra
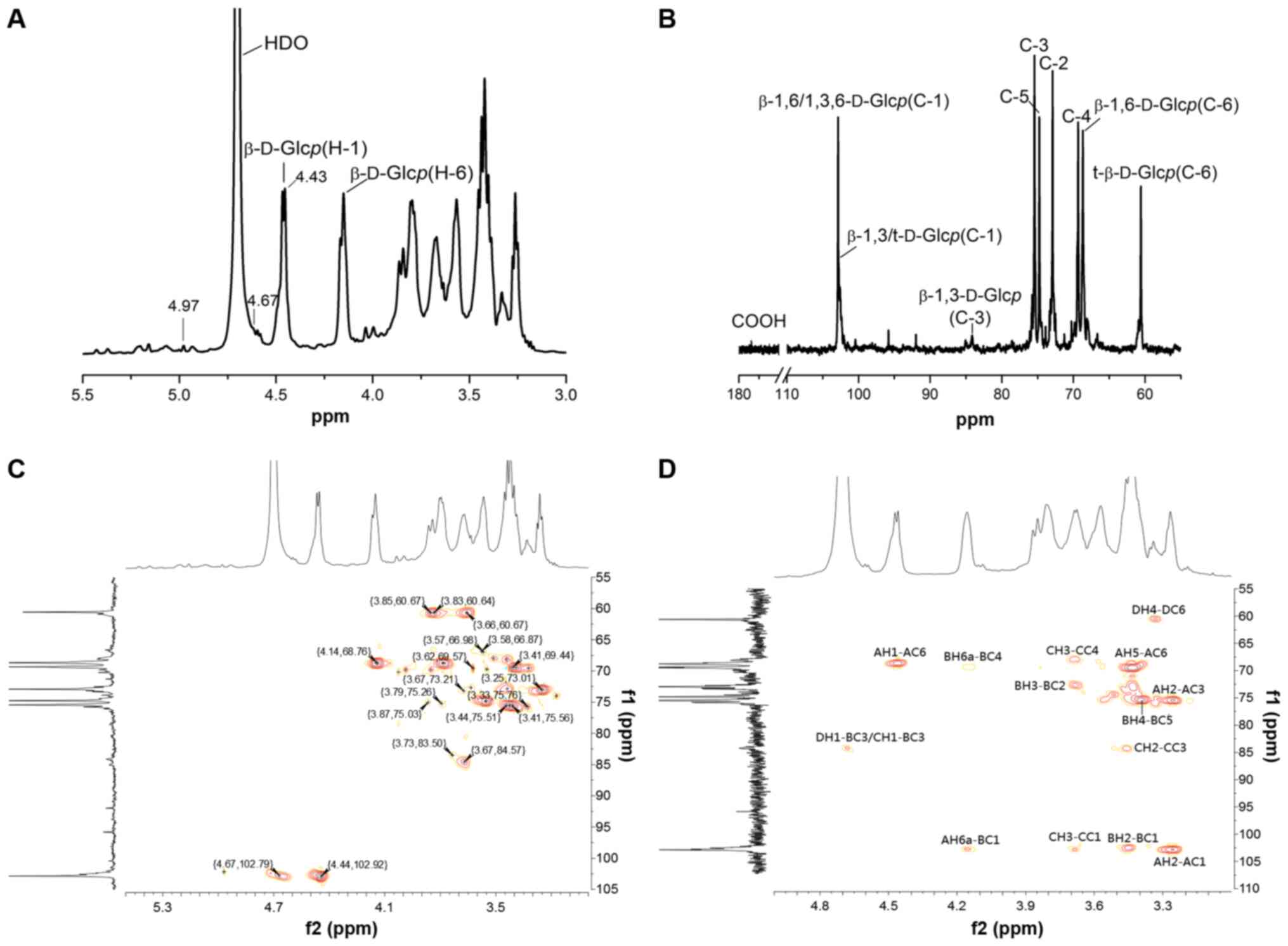
The 1D/2D NMR spectra of WCCP-A-b are presented in
Fig. 3 and the chemical shifts
are listed in Table II. In the
1H-NMR spectrum (Fig.
3A), there were three anomeric proton signals at 4.97, 4.67
(the signal peaks overlapped with HOD peaks) and 4.44 ppm, which
were assigned to anomeric protons of β-1,4-D-GlcAp (37),
t-β-D-Glcp/β-1,3-D-Glcp and
β-1,6-D-Glcp/β-1,3,6-D-Glcp (38), respectively. The proton chemical
shifts occurring in the 3.25-4.14 ppm region were H2-H6 of each
sugar residue. In the 13C-NMR spectrum (Fig. 3B), six obvious signals at
δ102.92, 73.01, 75.56, 69.44, 74.85 and 68.74 ppm arose from C-1,
C-2, C-3, C-4, C-5 and C-6 of β-1,6-D-Glcp residues
(39). The signals at 102.79 and
60.67 ppm were assigned to C-1 and C-6 of t-β-D-Glcp
(β-1,3-D-Glcp), respectively. The weak signal at 172.32 ppm
was assigned to carboxyl group of GlcpA.
| Table IIChemical shift assignments of H and C
signals for water-soluble C. cibarius polysaccharide
homogenous fraction (WCCP-A-b). |
Table II
Chemical shift assignments of H and C
signals for water-soluble C. cibarius polysaccharide
homogenous fraction (WCCP-A-b).
| Linkage type | 1 | 2 | 3 | 4 | 5 | 6 |
|---|
|
β-1,6-D-Glcp | | | | | | |
| H | 4.44 | 3.25 | 3.41 | 3.41 | 3.55 | 4.14;3.78 |
| C | 102.92 | 73.01 | 75.56 | 69.44 | 74.85 | 68.74 |
|
β-1,3,6-D-Glcp | | | | | | |
| H | 4.44 | 3.44 | 3.67 | 3.39 | 3.55 | 4.14;3.78 |
| C | 102.89 | 72.84 | 84.57 | 69.42 | 74.85 | 68.74 |
|
β-1,3-D-Glcp | | | | | | |
| H | 4.67 | 3.25 | 3.73 | 3.32 | 3.55 | 3.85;3.66 |
| C | 102.79 | 73.01 | 83.50 | 69.54 | 74.85 | 60.67 |
|
t-β-D-Glcp | | | | | | |
| H | 4.67 | 3.35 | 3.44 | 3.44 | 3.34 | 3.85;3.66 |
| C | 102.79 | 73.04 | 75.51 | 68.15 | 75.63 | 60.67 |
|
β-1,4-D-GlcAp | | | | | | |
| H | 4.97 | 3.67 | 3.79 | 3.66 | 3.87 | - |
| C | 102.18 | 73.21 | 75.26 | 80.55 | 75.03 | 172.32 |
Other proton and carbon signals of WCCP-A-b were
assigned according to the HSQC spectrum (Fig. 3C). The strong cross H1/C1 signal
at 4.44/102.92 ppm, H3/C3 signal at 3.41/75.56 ppm and H6/C6 signal
at 4.14;3.78/68.76 ppm arose from 1,6-linked β-D-Glcp. The
strong cross H1/C1 signal at 4.44/102.92 ppm, H3/C3 signal at
3.67/84.57 ppm and H6/C6 signal at 4.14;3.78/68.76 ppm were
attributed to 1,3,6-linked β-D-Glcp. The cross-peak at
4.67/102.79 and 3.85;3.66/60.67 ppm were from H1/C1 and H6a;b/C6 of
the terminal-β-D-Glcp, while the down-field shift at
3.73/83.50 ppm was from H3/C3 of 1,3- or 1,3,6-linked
β-D-Glcp (40,41). Furthermore, the weak signal peak
of H1/C1 (4.97/102.18 ppm) and down-field shift of H4/C4
(3.66/80.55 ppm) confirmed the existence of 1,4-linked
β-D-GlcAp (42,37).
In the HMBC spectrum (Fig. 3D), the cross peaks of both
anomeric protons and carbons of glycosyl residues AH1/AC6, AH2/AC1,
AH6a/BC1, BC3/CH1, BC3/DH1, CC1/CH3 and DC1/DH2 were observed. Due
to the low amount of GlcA in WCCP-A-b, no obvious cross peaks were
observed in HMBC for GlcA. Combined with the methylation analysis
results, the possible structure of WCCP-A-b was proposed as a
β-1,6-D-glucan, which was branched at O-3 of β-1,6-D-Glcp by
short β-1,3-D-Glcp oligosaccharides or single-unit terminal
β-Glcp residues. Small amounts of β-1,4-D-GlcpA may
also exist in side chains. According to the aforementioned
analysis, WCCP-A-b may be a novel β-1,6-D-glucan structure
containing β-1,4-D-GlcpA side chains, which to the best of
our knowledge has not been previously reported.
Acidic β-glucans have been previously reported
(37,43); however, their structures are
different from the WCCP-A-b that was identified in the present
study. A polysaccharide fraction (PSG-1-F0.2), isolated from
Ganoderma atrum, also mainly consisted of glucose and
glucuronic acid (37). The
backbone of PSG-1-F0.2 was identified to be composed of
β-(1→3)-glucose, which was different from WCCP-A-b. One glucan
fraction (WPOPA) obtained from Pleurotus ostreatus had a
similar main chain structure to WCCP-A-b, but most of the
GlcpA residues were in the form of t-β-D-GlcpA, which
was notably distinguished from WCCP-A-b (43).
WCCP-A-b promotes macrophage
activation
Macrophages can defend against pathogen invasion,
can kill tumor cells and can improve the immune capabilities by
releasing inflammatory mediators (27). NO, TNF-α and IL-6 serve an
important role in the immune process (34). In order to investigate the
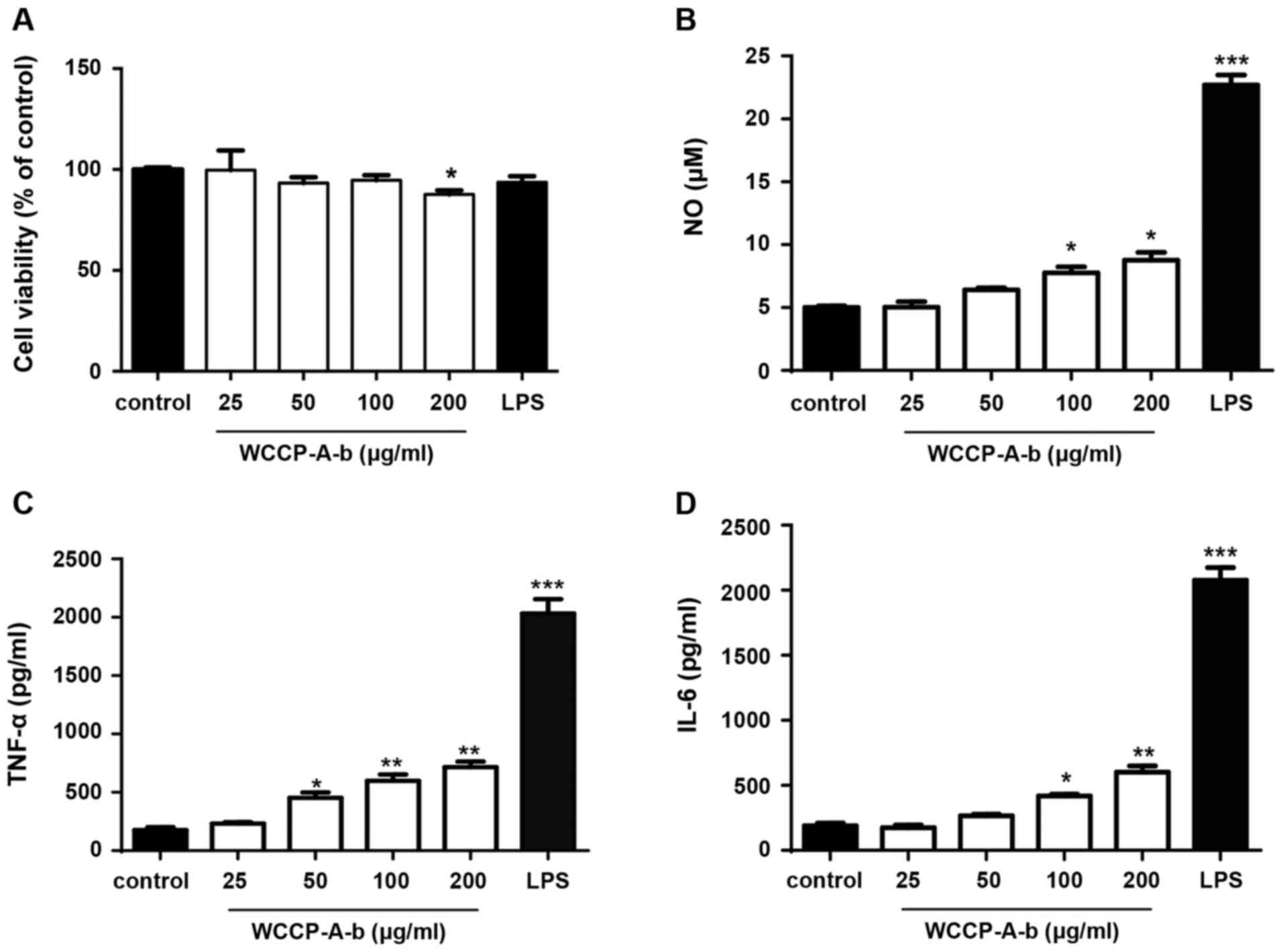
immunomodulatory activity of WCCP-A-b, RAW264.7 cells were treated
with WCCP-A-b. The MTT assay indicated that WCCP-A-b exhibited no
cytotoxicity towards RAW264.7 cells at concentrations of 0-100
μg/ml, but that there was a significant decrease in cell
viability at 200 μg/ml (Fig.
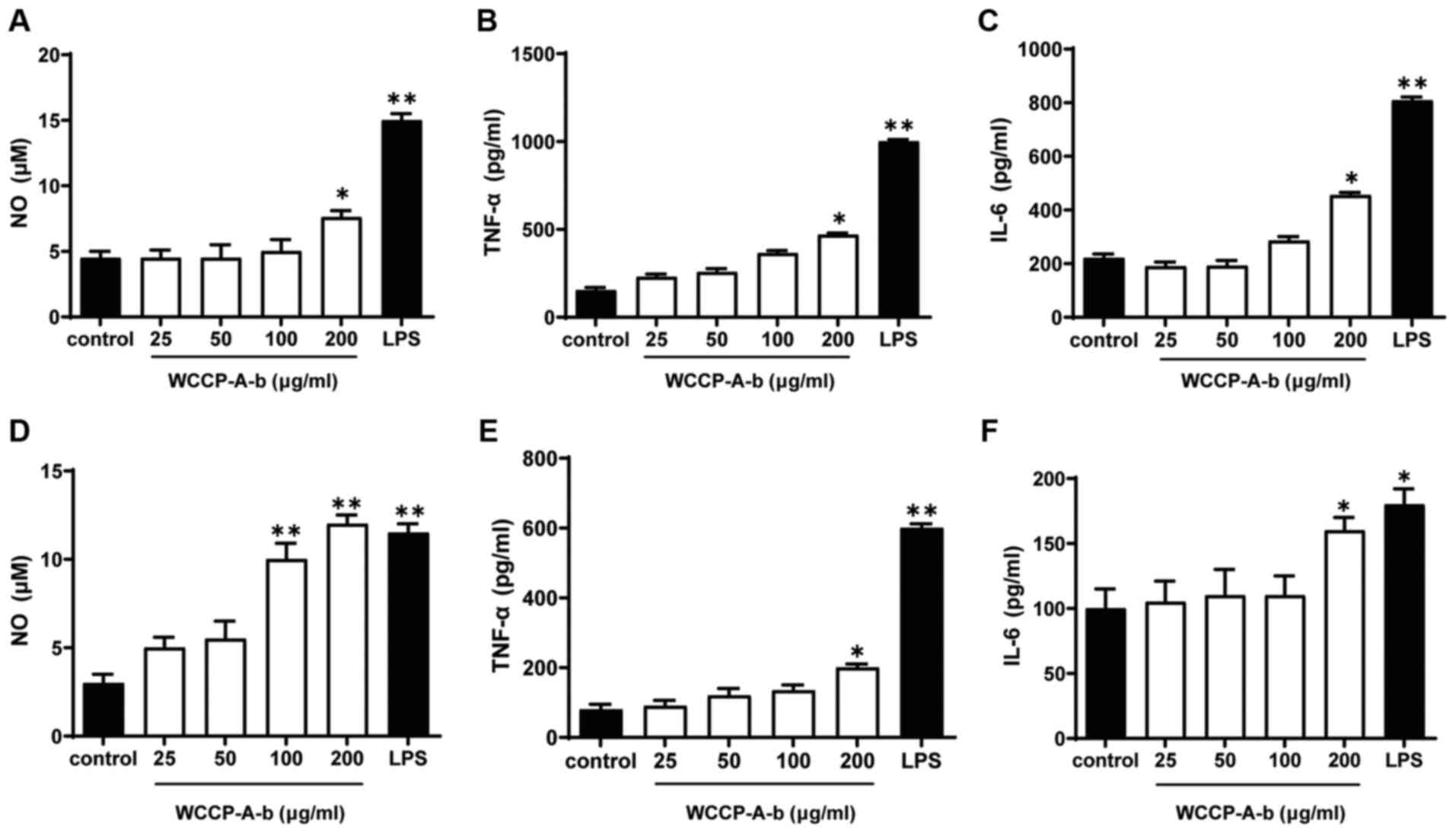
4A). ELISA assays and Griess reagent were used to quantify NO,
TNF-α and IL-6 concentrations in the conditioned medium of RAW264.7
cells. As the positive control, LPS significantly stimulated NO,
TNF-α and IL-6 production. Moreover, WCCP-A-b induced NO, TNF-α and
IL-6 production in RAW264.7 cells in a dose-dependent manner
(Fig. 4B-D), with a significant
difference for all at 100 and 200 μg/ml. To further confirm
these results, the aforementioned experiments were repeated using
mouse peritoneal macrophages and bone marrow macrophages (Fig. 5). The results demonstrated that
WCCP-A-b also induced NO, TNF-α and IL-6 production in mouse
peritoneal macrophages (Fig.
5A-C) and bone marrow macrophages (Fig. 5D-F), with significant increases
at 200 μg/ml, indicating that WCCP-A-b may promote
macrophage activation.
β-D-glucans obtained from mushrooms are effective
immunomodulators and are considered as modifiers of biological
responses (4). A neutral
branched β-glucan extracted from the fruiting bodies of
Amillariella mellea, containing β-D-(1→6)-linked Glcp
as its main chain, can promote macrophage phagocytosis and increase
production of NO, reactive oxygen species, TNF-α, IL-6 and IL-1β
(34). Another polysaccharide,
containing a higher content of (1→6)-linked β-glucan and lower
content of α-glucan, purified from Agaricus brasiliensis
increases the secretion of the pro-inflammatory cytokines IL-1β and
TNF-α in phorbol myristate acetate-differentiated THP-1 cells,
while it decreases pro-inflammatory effects caused by LPS,
indicating promising immune activity (44). However, a branched β-1,3-glucan,
derived from the fruiting bodies of Lentinus squarrosulus,
is able to activate macrophages, splenocytes and thymocytes at
certain concentrations (45).
Therefore, the novel β-1,6-D-glucan isolated from C.
cibarius in the present study may be used as a potential
natural immunostimulatory agent.
MAPK signaling pathway is involved in
macrophage activation
MAPKs, including ERKs, JNKs and p38-MAPKs, have been
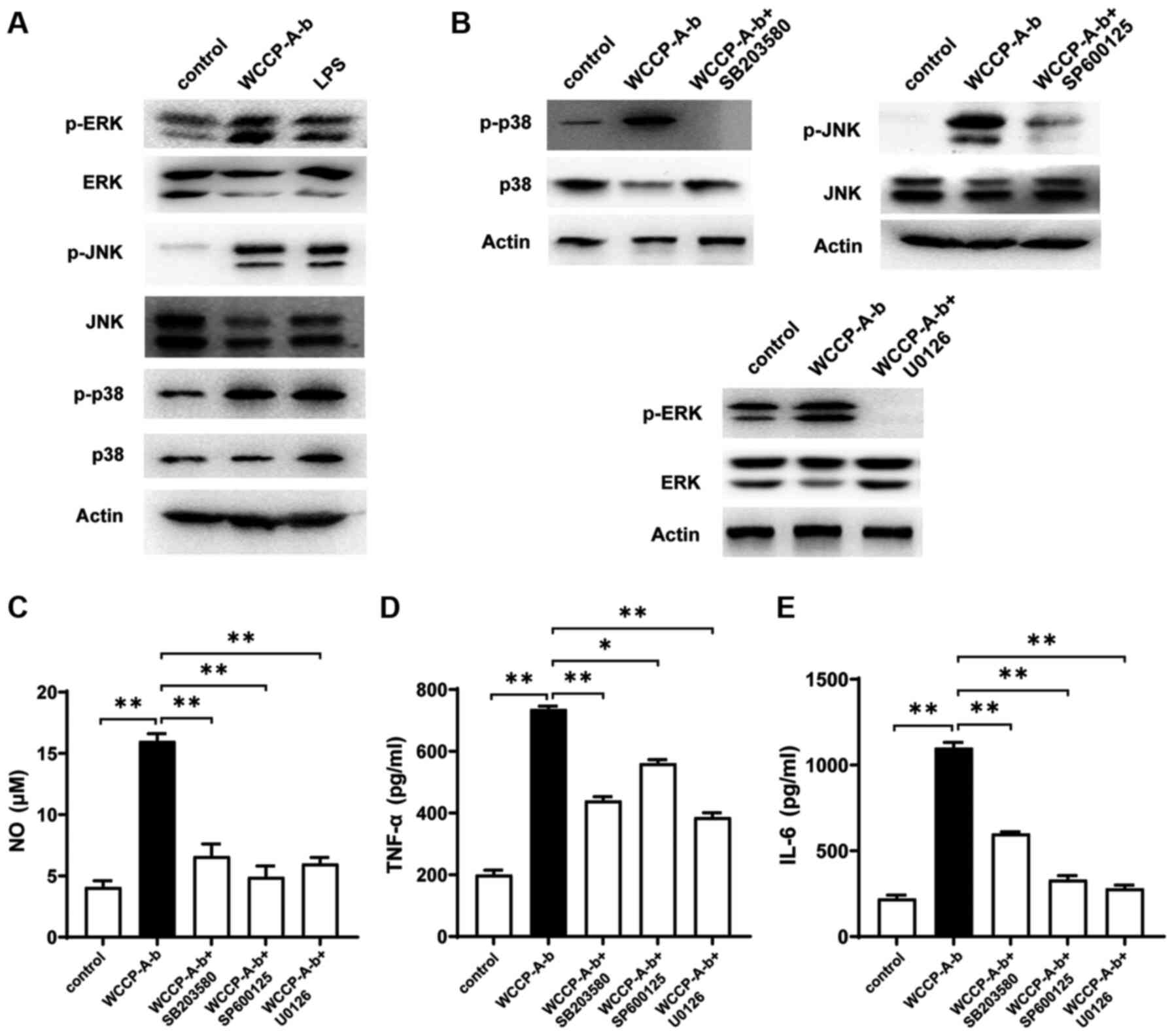
found to be associated with macrophage activation (36). The present study investigated
whether the MAPK signaling pathway was associated with
WCCP-A-b-induced macrophage activation. The results indicated that
WCCP-A-b increased the phosphorylation of ERK, JNK and p38
(Fig. 6A). Pre-treatment of
cells with inhibitors of JNK (SP600125), ERK (U0126) and p38
(SB203580) markedly decreased the phosphorylation of ERK, JNK and
p38, respectively (Fig. 6B). In
addition, the secretion of NO, TNF-α and IL-6 was significantly
suppressed following the addition of the aforementioned inhibitors
(Fig. 6C-E). Therefore, it was
suggested that the MAPK signaling pathway may be involved in the
macrophage activation by WCCP-A-b.
Conclusion
In the present study, an acidic β-glucan (WCCP-A-b)
was purified from hot water extracted polysaccharides from the
fruiting bodies of C. cibarius using anion exchange and
gel-permeation chromatography. The backbone of WCCP-A-b was a
β-D-1,6-glucan, which was branched at O-3 of Glcp by
β-1,3-D-Glcp short chains or single-unit of β-Glcp
residues. Furthermore, small amounts of β-1,4-D-GlcpA may be
present in the side chains. WCCP-A-b possessed a macrophage
activatory effect by promoting the secretion of NO, TNF-α and IL-6.
On a cellular mechanistic level, WCCP-A-b activated macrophages via
the MAPKs signaling pathway. However, the structure-activity
association of the glucan was not deeply discussed in the present
study. Therefore, more studies, such as in vivo animal
experiments, on the investigation of the immunomodulatory activity
should be further performed. The present data indicated that the
identified novel β-glucan may be used as a potent
immunomodulator.
Funding
The present study was supported by the Science &
Technology Major Project 'Key New Drug Creation and Manufacturing
Program' (grant no. 2019ZX09735001), the Fundamental Research Funds
for the Central Universities (grant no. 2412020FZ018), and the
Jilin Province Development and Reform Commission (grant no.
2019C018).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS and YZ conceived the study and revised the
manuscript. YQ purified and characterized the polysaccharides and
drafted the manuscript. XZ and HG performed the polysaccharide
extraction. YM and YW performed the macrophage activation
experiments and confirmed the authenticity of the data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee of Northeast Normal University
(approval no. AP20151009) and was conducted in accordance with the
National Standards of the People's Republic of China Laboratory
Animal-Guideline for Ethical Review of Animal Welfare.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Manzi P, Aguzzi A and Pizzoferrato L:
Nutritional value of mushrooms widely consumed in Italy. Food Chem.
73:321–325. 2001. View Article : Google Scholar
|
|
2
|
Sun Y, Zhang M and Fang Z: Efficient
physical extraction of active constituents from edible fungi and
their potential bioactivities: A review. Trends Food Sci Technol.
105:468–482. 2020. View Article : Google Scholar
|
|
3
|
Rathore H, Prasad S and Sharma S: Mushroom
nutraceuticals for improved nutrition and better human health: A
review. PharmaNutrition. 5:35–46. 2017. View Article : Google Scholar
|
|
4
|
Kothari D, Patel S and Kim SK: Anticancer
and other therapeutic relevance of mushroom polysaccharides: A
holistic appraisal. Biomed Pharmacother. 105:377–394. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ruthes AC, Smiderle FR and Iacomini M:
D-glucans from edible mushrooms: A review on the extraction,
purification and chemical characterization approaches. Carbohydr
Polym. 117:753–761. 2015. View Article : Google Scholar
|
|
6
|
Bhanja SK, Rout D, Patra P, Sena IK,
Nandan CK and Islam SS: Water-insoluble glucans from the edible
fungus Ramaria botrytis. Bioactive Carbohydrates and Dietary Fibre.
3:52–58. 2014. View Article : Google Scholar
|
|
7
|
Palacios I, García-Lafuente A, Guillamón E
and Villares A: Novel isolation of water-soluble polysaccharides
from the fruiting bodies of Pleurotus ostreatus mushrooms.
Carbohydr Res. 358:72–77. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chakraborty I, Mondal S, Rout D and Islam
SS: A Water-insoluble (1->3)-beta-D-glucan from the alkaline
extract of an edible mushroom Termitomyces eurhizus. Carbohydr Res.
341:2990–2993. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Smiderle FR, Alquini G, Tadra-Sfeir MZ,
Iacomini M, Wichers HJ and Van Griensven LJ: Agaricus bisporus and
Agaricus brasiliensis (1→6)-β-D-glucans show immunostimulatory
activity on human THP-1 derived macrophages. Carbohydr Polym.
94:91–99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Castro-Alves VC and Nascimento JROD: α-
and β-d-Glucans from the edible mushroom Pleurotus albidus
differentially regulate lipid-induced inflammation and foam cell
formation in human macrophage-like THP-1 cells. Int J Biol
Macromol. 111:1222–1228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smiderle FR, Baggio CH, Borato DG,
Santana-Filho AP, Sassaki GL, Iacomini M and Van Griensven LJ:
Anti-inflammatory properties of the medicinal mushroom Cordyceps
militaris might be related to its linear (1→3)-β-D-glucan. PLoS
One. 9:e1102662014. View Article : Google Scholar
|
|
12
|
Hida TH, Kawaminami H, Ishibashi K, Miura
NN, Adachi Y and Ohno N: Oral administration of soluble β-glucan
prepara- tion from the cauliflower mushroom, Sparassis crispa
(Higher Basidiomycetes) modulated cytokine production in mice. Int
J Med Mushrooms. 15:525–538. 2013. View Article : Google Scholar
|
|
13
|
Misaki A, Kakuta M, Sasaki T, Tanaka M and
Miyaji H: Studies on interrelation of structure and antitumor
effects of polysaccharides: Antitumor action of periodate-modified,
branched (1 goes to 3)-beta-d-glucan of auricularia auricula-judae,
and other polysaccharides containing (1 goes to 3)-glycosidic
linkages. Carbohydr Res. 92:115–129. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Misaki A, Kawaguchi K, Miyaji H, Nagae H,
Hokkoku S, Kakuta M and Sasaki T: Structure of pestalotan, a highly
branched (1-3)-beta-d-glucan elaborated by Pestalotia, sp. 815, and
the enhancement of its antitumor activity by polyol modification of
the side chains. Carbohydr Res. 129:209–227. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Konga H, Fang Y, Nishinari K and
Phillips GO: Schizophyllan: A review on its structure, properties,
bioactivities and recent developments. Bioactive Carbohydrate and
Dietary. Fiber. 1:53–71. 2013.
|
|
16
|
Lehtovaara BC and Gu FX: Pharmacological,
structural, and drug delivery properties and applications of
1,3-β-glucans. J Agric Food Chem. 59:6813–6828. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Synytsya A, Míčková K, Synytsya A,
Jablonský I, Spěváček J, Erban V, Kováříková E and Čopíková J:
Glucans from fruit bodies of cultivated mushrooms Pleurotus
ostreatus and Pleurotus eryngii: Structure and potential prebiotic
activity. Carbohydr Polym. 76:548–556. 2009. View Article : Google Scholar
|
|
18
|
Oishi Y and Manabe I: Macrophages in
inflammation, repair and regeneration. Int Immunol. 30:511–528.
2018.PubMed/NCBI
|
|
19
|
Martinez FO and Gordon S: The M1 and M2
paradigm of macrophage activation: Time for reassessment.
F1000Prime Rep. 6:132014. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu KS, Zhang C, Dong HL, Li KK, Han QB,
Wan Y, Chen R, Yang F, Li HL, Ko CH and Han XQ: GSP-2, a
polysaccharide extracted from Ganoderma sinense, is a novel
toll-like receptor 4 agonist. PLoS One. 14:e02216362019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu D, Tang C, Liu Y, Li Q, Wang W, Zhou S,
Zhang Z, Cui F and Yang Y: Structural elucidation and
immunomodulatory activity of a β-D-glucan prepared by
freeze-thawing from Hericium erinaceus. Carbohydr Polym.
222:1149962019. View Article : Google Scholar
|
|
23
|
Han XQ, Li WJ, Ko CH, Gao XM, Han CX and
Tu PF: Structure characterization and immunocompetence of a glucan
from the fruiting bodies of Cantharellus cibarius. J Asian Nat Prod
Res. 15:1204–1209. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nowacka-Jechalke N, Nowak R, Juda M, Malm
A, Lemieszek M, Rzeski W and Kaczyński Z: New biological activity
of the polysaccharide fraction from Cantharellus cibarius and its
structural characterization. Food Chem. 268:355–361. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lemieszek MK, Nunes FM, Marques G and
Rzeski W: Cantharellus cibarius branched mannans inhibits colon
cancer cells growth by interfering with signals transduction in
NF-ĸB pathway. Int J Biol Macromol. 134:770–780. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nyman AA, Aachmann FL, Rise F, Balance S
and Samuelsen AB: Structural characterization of a branched
(1→6)-α-mannan and β-glucans isolated from the fruiting bodies of
Cantharellus cibarius. Carbohydr Polym. 146:197–207. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang G, Qu Y, Meng Y, Wang Y, Song C,
Cheng H, Li X, Sun L and Zhou Y: A novel linear 3-O-methylated
galactan isolated from Cantharellus cibarius activates macrophages.
Carbohydr Polym. 214:34–43. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Meng Y, Qu Y, Wu W, Chen L, Sun L, Tai G,
Zhou Y and Cheng H: Galactan isolated from Cantharellus cibarius
modulates antitumor immune response by converting tumor-associated
macrophages toward M1-like phenotype. Carbohydr Polym.
226:1152952019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ana V, Ana GL, Eva G and Laura MV:
Separation and characterization of the structural features of
macromolecular carbohydrates from wild edible mushrooms. Bioactive
Carbohydrates and Dietary Fibre. 2:15–21. 2013. View Article : Google Scholar
|
|
30
|
Dubois M, Gilles K, Hamilton JK, Rebers PA
and Smith F: A colorimetric method for the determination of sugars.
Nature. 168:1671951. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Blumenkrantz N and Asboe-Hansen G: New
method for quantitative determination of uronic acids. Anal
Biochem. 54:484–489. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sedmak JJ and Grossberg SE: A rapid,
sensitive, and versatile assay for protein using Coomassie
brilliant blue G250. Anal Biochem. 79:544–552. 1977. View Article : Google Scholar
|
|
33
|
Zhang X, Yu L, Bi H, Li X, Ni W, Han H, Li
N, Wang B, Zhou Y and Tai G: Total fractionation and
characterization of the water-soluble polysaccharides isolated from
Panax ginseng C. A Meyer Carbohydr Polym. 77:544–552. 2009.
View Article : Google Scholar
|
|
34
|
Yan J, Han Z, Qu Y, Yao C, Shen D, Tai G,
Cheng H and Zhou Y: Structure elucidation and immunomodulatory
activity of a β-glucan derived from the fruiting bodies of
Amillariella mellea. Food Chem. 240:534–543. 2018. View Article : Google Scholar
|
|
35
|
Needs PW and Selvendran RR: Avoiding
oxidative degradation during sodium hydroxide/methyl
iodide-mediated carbohydrate methylation in dimethyl sulfoxide.
Carbohydr Res. 245:1–10. 1993. View Article : Google Scholar
|
|
36
|
Meng Y, Yan J, Yang G, Han Z, Tai G, Cheng
H and Zhou Y: Structural characterization and macrophage activation
of a hetero-galactan isolated from Flammulina velutipes. Carbohydr
Polym. 183:207–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang H, Nie S, Cui SW, Xu M, Ding H and
Xie M: Characterization of a bioactive polysaccharide from
Ganoderma atrum: Re-elucidation of the fine structure. Carbohydr
Polym. 158:58–67. 2017. View Article : Google Scholar
|
|
38
|
Han XQ, Yue GL, Yue RQ, Dong CX, Chan CL,
Ko CH, Cheung WS, Luo KW, Dai H, Wong CK, et al: Structure
elucidation and immunomodulatory activity of a beta glucan from the
fruiting bodies of Ganoderma sinense. PLoS One. 9:e1003802014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Siu KC, Xu L, Chen X and Wu JY: Molecular
properties and antioxidant activities of polysaccharides isolated
from alkaline extract of wild Armillaria ostoyae mushrooms.
Carbohydr Polym. 137:739–746. 2016. View Article : Google Scholar
|
|
40
|
Oliveira KS, Di Bastiani M, Cordeiro LM,
Costa MF, Toledo KA, Iacomini M, Babosa AM, Dekker RF and
Nascimento VM: (1→6)- and (1→3)(1→6)-β-glucans from Lasiodiplodia
theobromae MMBJ: Structural characterization and pro-inflammatory
activity. Carbohydrate Polymers. 133:539–546. 2015. View Article : Google Scholar
|
|
41
|
Ruthes AC, Carbonero ER, Córdova MM,
Baggio CH, Sassaki GL, Gorin PA, Santos AR and Iacomini M:
Fucomannogalactan and glucan from mushroom Amanita muscaria:
Structure and inflammatory pain inhibition. Carbohydr Polym.
98:761–769. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Du XJ, Zhang JS, Yang Y, Ye LB, Tang QJ,
Jia W, Liu Y, Zhou S, Hao R, Gong CY and Pan Y: Structural
elucidation and immuno-stimulating activity of an acidic
heteropolysaccharide (TAPA1) from Tremella aurantialba. Carbohydr
Res. 344:672–678. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yan J, Zhu L, Qu Y, Qu X, Mu M, Zhang M,
Muneer G, Zhou Y and Sun L: Analyses of active antioxidant
polysaccharides from four edible mushrooms. Int J Biol Macromol.
123:945–956. 2019. View Article : Google Scholar
|
|
44
|
Smiderle FR, Ruthes AC, van Arkel J,
Chanput W, Iacomini M, Wichers HJ and VanGriensven LJ:
Polysaccharides from Agaricus bisporus and Agaricus brasiliensis
show similarities in their structures and their immunomodulatory
effects on human monocytic THP-1 cells. BMC Complement Altern Med.
11:582011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bhunia SK, Dey B, Maity KK, Patra S,
Mandal S, Maiti S, Maiti TK, Sikdar SR and Islam SS: Isolation and
characterization of an immunoenhancing glucan from alkaline extract
of an edible mushroom, Lentinus squarrosulus (Mont.) Singer.
Carbohydr Res. 346:2039–2044. 2011. View Article : Google Scholar : PubMed/NCBI
|