Introduction
Atherosclerosis (AS) is a chronic inflammatory
arterial angiopathy and a common cause of cardiovascular diseases
(1). The common pathological
changes of AS include endothelial cell dysfunction, lipid
deposition and vascular wall thickening (2). Vascular endothelial cells
constitute the inner lining of arterial vessels, and their
morphological integrity and functional homeostasis play an
important role. Damaged integrity and dysfunction in vascular
endothelial cells can initiate AS and thus lead to the development
of severe clinical acute events, such as myocardial infarction and
stroke (3). Proprotein
convertase subtilisin/kexin type 9 (PCSK9) is an important risk
factor of AS that induces disordered lipid metabolism by cutting
low-density lipoprotein receptor (4), and participates in the emergence
and progression of AS through the non-lipid metabolic pathway,
resulting in vascular endothelial cell death and vascular
inflammation (5). However, the
underlying mechanisms remain unclear.
Programmed cell death, such as autophagy, apoptosis
and pyroptosis, occurs in pathological and disease progression.
Cell pyroptosis is accompanied by the release of inflammatory
mediators, such as interleukin (IL)-1β and IL-18 (6,7),
which can induce cell dysfunction and plaque formation (8). Oxidized low-density lipoprotein
(oxLDL), a risk factor for the development of AS, can cause
mitochondrial dysfunction and increase reactive oxygen species
(ROS) levels in human umbilical vein endothelial cells (HUVECs),
thereby inducing the pyroptosis of endothelial cells and the
release of inflammatory mediators (9). However, the involvement of PCSK9 in
the pyroptosis of vascular endothelial cells remains unclear.
As the energy supplying organelle, the mitochondria
regulate cell functions and adjust homeostasis by releasing related
signal molecules (10).
Homeostasis disruption causes mitochondrial dysfunction, which
reduces mitochondrial membrane potential, decreases ATP level, and
increases ROS production, leading to cell dysfunction. oxLDL can
damage mitochondrial function, induce ROS release and cause
endothelial cell pyroptosis (9).
ROS are important mediators in the signaling pathway of pyroptosis.
Abnormal shear stress can promote PCSK9 expression in endothelial
cells (11), and PCSK9
upregulation increases ROS levels (12). Therefore, PCSK9 possibly causes
pyroptosis. The present study aimed to determine whether PCSK9
regulates the pyroptosis of vascular endothelial cells through the
ROS pathway.
Materials and methods
Cell lines and cell culture
HUVECs (Science Cell Research Laboratories) were
cultured in endothelial cell medium (Science Cell Research
Laboratories) at 37°C with 5% CO2. The small-interfering
RNA (siRNA) kit was purchased from Guangzhou RiboBio Co., Ltd. In
brief, HUVECs at a density of 30-50% were transfected with siRNA
(RiboBio Transfection reagent, 50 µM) (siPCSK9, 3′-GAG GTG
TAT CTC CTA GAC A-5′; siUQCRC1, 3′-GTG CAC ACT TTG TCT GTG A-5′) or
lentivirus (LV)-PCSK9 (Shanghai GeneChem Co., Ltd.; with Genechem
HitransG, MOI=50) in the presence or absence of PCSK9 and then
incubated with oxLDL (100 µg/ml; Yiyuan Biotechnology Co.,
Ltd.) for 24 h in an incubator.
Detection of cell pyroptosis
Lactate dehydrogenase (LDH) release was measured
with an LDH kit (Jinagcheng bioengineering institute, Nanjing,
China) and Hoechst 33342/propidium iodide (PI) staining were used
to evaluate pyroptosis. For LDH release detection, 25 µl of
cell supernatant and 25 µl of substrate were incubated at
37°C for 15 min. Subsequently, 2,4-dinitrophenylhydrazine (25
µl) and 0.4 mol/l NaOH solution (250 µl) (Nanjing
Jinagcheng Bioengineering Institute) were added. Following
incubation at room temperature for 5 min, the absorbance was
determined using a spectrophotometric microplate reader (iMark-03;
Bio-Rad Laboratories, Inc.).
For Hoechst 33342/PI staining, the HUVECs were first
washed thrice with phosphate-buffered saline (PBS), followed by the
addition of 10 µl of Hoechst 33342 solution, and incubation
at 37°C in the dark for 10 min. After removing the dye, the cells
were washed times with PBS, added with 5 µl of PI dye, and
then incubated at 37°C in the dark for 10 min. After being washed
with PBS 3 times, the cells were observed under a fluorescence
microscope (IX3; Olympus Corporation).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
TRIzol reagent (Roche Diagnostics) was used to
extract total RNA, which was then reverse transcribed into cDNA
using the RT kit (FSQ-101, Toyobo Co., Ltd.). The product was
processed with SYBR-Green Premix Dimer Eraser (Takara Biotechnology
Co., Ltd.), and the level of mRNA was then measured using the ABI
7000 sequence detection system (ABI PRISM® 7000, Thermo
Fisher Scientific, Inc.) (1 cycle, 95°C, 15 min; 40 cycles, 95°C,
10 sec, 60°C, 32 sec). GAPDH was used as the housekeeping gene for
normalization. The primers and sequences used were as follows:
IL-1β forward, 5′-CGA TCA CTG AAC TGC ACG CT-3′ and reverse, 5′-AGA
ACA CCA CTT GTT GCT CCA-3′; GAPDH forward, 5′-AAGATC AAG ATC ATT
GCT CCT CCT G-3′ and reverse, 5′-GCC GGA CTC GTC ATA CTC CT-3′; and
PCSK9 forward, 5′-ACG ATG CCT GCC TCT ACT CC-3 and reverse, 5′-GCC
TGT GAT GTC CCA CTC TGT-3′.
Enzyme-linked immunosorbent assay
(ELISA)
ELISA kit (NeoBioscience, HongKong) was employed to
measure the concentration of IL-1β secreted by HUVECs. Cells were
first seeded into 24-well plates. Following the different
treatments (oxLDL with/without PCSK9 siRNA, LV-PCSK9), the
supernatant (100 µl) was collected for ELISA in accordance
with the manufacturer's protocol (NeoBioscience Technology
Company).
Western blot analysis
Cells were first washed thrice with cold PBS and
then lysed with radioimmunoprecipitation assay buffer and
phenylmethanesulfonyl fluoride (v/v, 94:6) on ice for 30 min. Lysed
samples were centrifuged at 9,500 × g for 15 min at 4°C. The
concentration of total proteins was determined using the BCA kit
(Beijing CWBio). Protein samples (20 µg/well) were loaded
for 8-12% sodium dodecyl sulfate polyacrylamide gel electrophoresis
and then transferred onto a nitrocellulose filter membrane, which
was blocked with 5% skimmed milk for 2 h at room temperature.
Subsequently, the membrane was incubated with caspase-1 antibody
(1:500; 22915-1-AP; ProteinTech Group, Inc.), IL-1β antibody
(1:500; 16806-1-AP; ProteinTech Group, Inc.), IL-18 antibody
(1:1,000; 10663-1-AP; ProteinTech Group, Inc.), NLRP3 antibody
(1:500; A5652; ABclonal Biotech Co., Ltd.), GAPDH antibody
(1:1,000; 10494-1-AP; ProteinTech Group, Inc.), PCSK9 antibody
(1:500; 55206-1-AP; ProteinTech Group, Inc.), gasdermin D (GSDMD-N;
1:500; ab253350, Abcam), cleaved caspase-1 (1:1,000; ab207802;
Abcam) and ubiquinol-cytochrome c reductase core protein 1
(UQCRC1) antibody (1:2,000; 21705-1-AP; ProteinTech Group, Inc.)
diluted by the mixture of Tris-buffered saline (containing
Tween-20) and 2.5% skimmed milk buffer at 4°C overnight. After
washing 5 times with PBS containing Tween-20, the membrane was
incubated with fluorescence-conjugated antirabbit IgG secondary
antibody (1:2,000, SA00001-2; ProteinTech Group, Inc.) at room
temperature for 2 h. After washing 5 times with Tris-buffered
saline with Tween-20 (TBST; #9997; Cell Signaling Technology,
Inc.), the optical density of the related bands was examined using
Quantity One software V4.6.6 (Bio-Rad Laboratories, Inc.) with
GAPDH as the internal reference.
Immunofluorescence
Following stimulation with various concentrations of
oxLDL (50 and 100 µg/ml) for 24 h, the cells were fixed with
4% formaldehyde for 30 min. Following incubation with 0.1% Triton
X-100 at room temperature for 30 min, the cells were blocked with
goat serum for 1 h at room temperature. After being washed thrice
with PBS, the cells were incubated with diluted PCSK9 antibody
(1:100; #85813, Cell Signaling Technology, Inc.) at 4°C overnight.
After being washed 5 times with PBS, the cells were incubated with
Alexa Fluor-conjugated secondary antibody (1:50, A32731,
Invitrogen; Thermo Fisher Scientific, Inc.) in the dark for 1 h at
room temperature. Finally, the nuclei were stained with
4′,6-diamidino-2-phenylindole (Beyotime Institute of Biotechnology,
Inc.) for 5 min at room temperature. After being washed 5 times
with PBS, the cells were observed under a fluorescence microscope
(IX3; Olympus Corporation).
Determination of ATP concentration in
cells
Cells (1.0×106/well) were washed with PBS
twice and then lysed with 150 ml of lysis buffer. After being
centrifuged at 1,500 × g for 5 min at 4°C, ATP detection reagent
(Nanjing Jinagcheng Bioengineering Institute) was added, followed
by examination using a microplate reader (iMark-03; Bio-Rad
Laboratories, Inc.). ATP concentration was calculated using the
standard curve.
Detection of ROS
The mitochondrial ROS probe (DCFH-DA; Nanjing KeyGen
Biotech Co. Ltd.) was used to detect ROS production by HUVECs. Upon
reaching 30-50% confluence, the cells were transfected with siRNA
against PCSK9 and siUQCRC1. The transfected cells were incubated
with oxLDL (100 µg/ml) for 24 h at room temperature. After
discarding the medium, the cells were washed with cold PBS (pH 7.4)
and then incubated with DCFH-DA at room temperature for 30 min.
After being washed thrice with PBS, the cells were observed under a
fluorescence microscope (IX3; Olympus Corporation).
Detection of mitochondrial membrane
potential
The cells were first washed with PBS and then
centrifuged at 1,500 × g for 5 min at room temperature. After
discarding the supernatant, the cells were resuspended with 500
µl of JC-1 and then incubated at 37°C and 5% CO2
for 15 min. Following centrifugation at 1,500 × g for 5 min at room
temperature, the supernatant was removed and the cells were
resuspended with 500 µl of 1X incubation buffer. The cells
were then observed under a fluorescence microscope (IX3; Olympus
Corporation) and examined by flow cytometry (Ex=488 nm; Em=530 nm)
(Sysmex-Partec CyFlow® Cube 6, Sysmex Partec). The ratio
of green fluorescence intensity/red fluorescence intensity directly
reflected mitochondrial membrane potential and mitochondrial
function.
Statistical analysis
All data are presented as the means value with
standard deviation. Differences between groups were determined by
one-way ANOVA accompanied by the Tukey's or Bonferroni post hoc
test (GraphPad Prism version 5.0; GraphPad, Inc.). A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
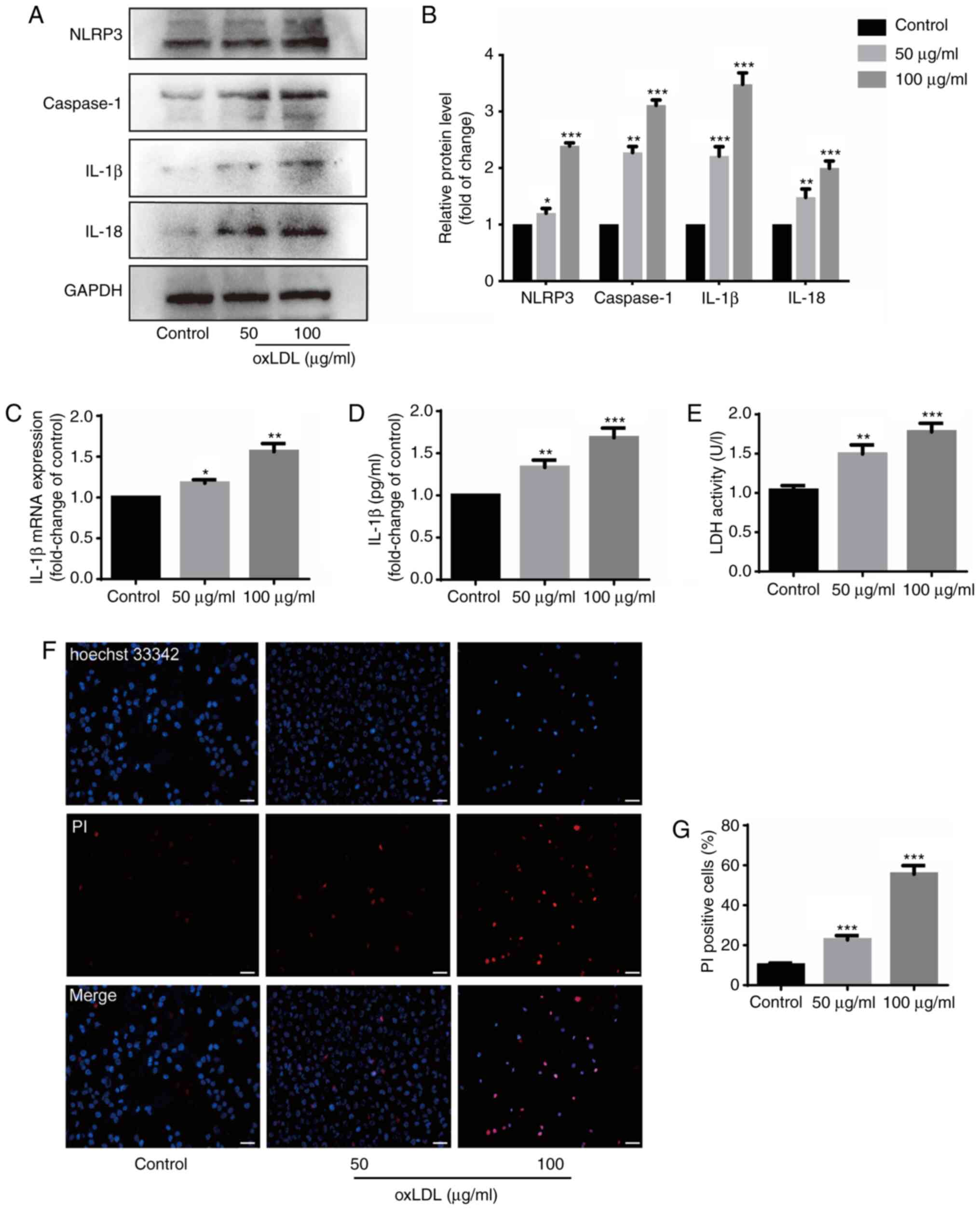
oxLDL induces HUVEC injury, pyroptosis
and the release of inflammatory factors
HUVECs were incubated with oxLDL (50 and 100
µg/ml) for 24 h prior to detecting the expression levels of
related molecules to explore whether oxLDL can cause damage to and
the pyroptosis of these cells. oxLDL upregulated the protein
expression levels of NLRP3, caspase-1 and IL-18 (Fig. 1A and B), and also increased the
protein and mRNA expression of IL-1β (Fig. 1A-C). GSDMD-N expression was also
increased by oxLDL stimulation (Fig. S1). The results of ELISA revealed
that the IL-1β protein content increased in the supernatant of the
HUVECs stimulated with oxLDL (Fig.
1D). The LDH level was also measured to determine the damage
caused to the HUVECs by oxLDL. The data indicated that oxLDL
promoted the release of LDH in a dose-dependent manner (Fig. 1E). Double staining with Hoechst
33342 and PI revealed that the ratio of PI-positive cells increased
with the increasing oxLDL concentration (Fig. 1F and G). These results suggest
that oxLDL induces injury to, and the pyroptosis of HUVECs, as well
as the release of inflammatory factors from HUVECs.
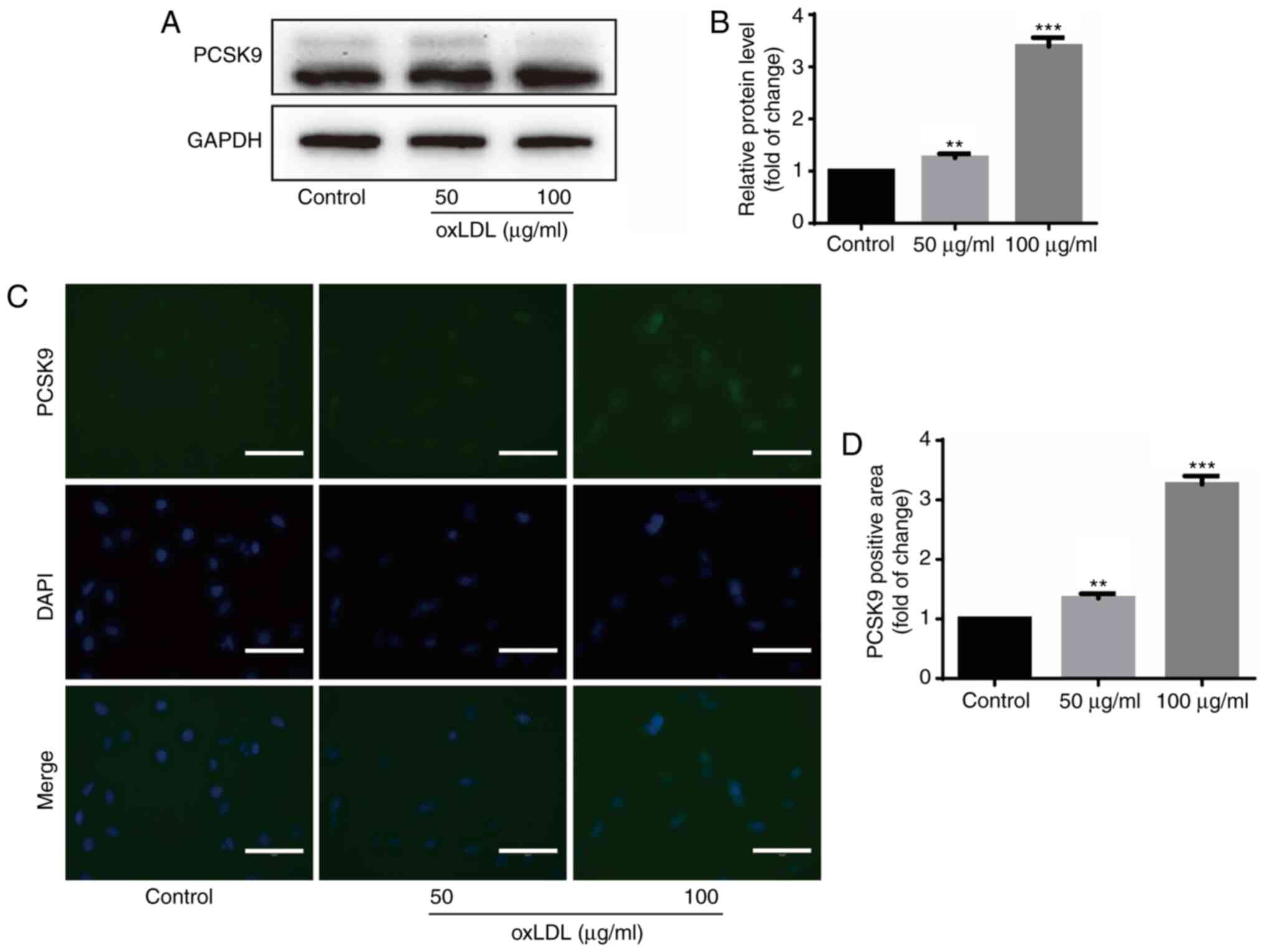
oxLDL upregulates PCSK9 protein
expression in HUVECs
PCSK9 expression was examined following stimulation
with oxLDL to examine the association between oxLDL and PCSK9.
Western blot analysis revealed that oxLDL stimulation upregulated
PCSK9 expression in HUVECs (Fig. 2A
and B). Immunofluorescence also revealed similar findings, in
which the PCSK9 fluorescence intensity increased following
stimulation with oxLDL (Fig. 2C and
D). These results indicate that oxLDL upregulates PCSK9 protein
expression in HUVECs.
siPCSK9 suppresses the damage, release of
inflammatory factors and pyroptosis of HUVECs induced by oxLDL
It has been demonstrated that PCSK9 inhibition
prevents inflammation and alleviates cell death (13). In the present study, the level of
pyroptosis in the HUVECs was detected following stimulation with
oxLDL with or without siPCSK9 transfection (50 µM siRNA).
The siRNA efficiency of PCSK9 was determined (Fig. 3A). The expression levels of
NLRP3, caspase-1 and IL-18 in the oxLDL group were significantly
higher than those in the control group and siPCSK9 group (Fig. 3B and C). In addition,
transfection with siPCSK9 decreased the protein and mRNA expression
levels of IL-1β in the oxLDL group (Fig. 3B-D). The results of ELISA and LDH
assay revealed that PCSK9 interference attenuated the upregulating
effect of oxLDL on IL-1β expression (Fig. 3E) and reduced the damage to
HUVECs caused by oxLDL, evidenced by the decreased activity of LDH
(Fig. 3F). The PI staining data
revealed that the pyroptosis of HUVECs was reduced by PCSK9
interference (Fig. 3G and H).
These results suggest that PCSK9 interference suppresses the
oxLDL-induced damage, the release of inflammatory substances and
the pyroptosis of HUVECs.
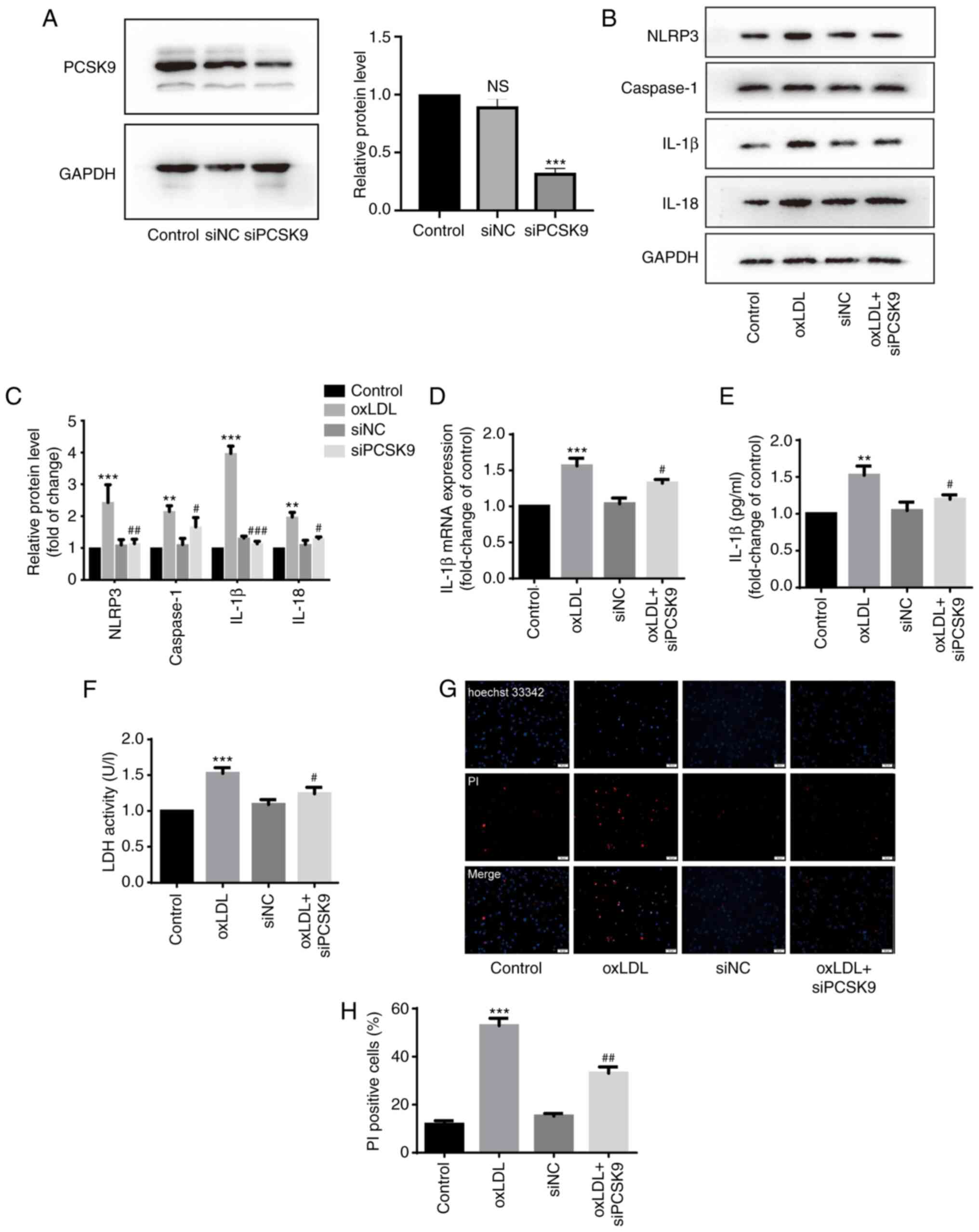 | Figure 3Interference with PCSK9 inhibits the
oxLDL-induced pyroptosis of HUVECs. (A) Expression of PCSK9 protein
in HUVECs following interference with PCSK9 (50 nM, 24 h) Western
blot analysis was used to measure protein expression.
***P<0.001 compared with the control group; NS, not
significant. (B and C) Expression of NLRP3, caspase-1, IL-1β and
IL-18 protein in HUVECs following interference with PCSK9 following
stimulation with 100 µg/ml oxLDL for 24 h. Western blot
analysis was used to measure protein expression.
**P<0.01 and ***P<0.001 compared with
the control group; #P<0.05, ##P<0.01
and ###P<0.001 compared with the oxLDL group. (D) Expression of
IL-1β mRNA. ***P<0.001 compared with the control
group; #P<0.05 compared with the oxLDL group. (E)
Concentration of IL-1β in supernatant of HUVECs.
**P<0.01 compared with the control group;
#P<0.05 compared with the oxLDL group. (F) LDH levels
in supernatant of HUVECs. ***P<0.001 compared with
the control group; #P<0.05 compared with the oxLDL
group. (G and H) Ratio of PI-positive HUVECs. Original
magnification, ×100; scale bar, 100 µm.
***P<0.001 compared with the control group;
##P<0.01 compared with the oxLDL group. All results
are expressed as the means ± SD of 3 independent experiments.
oxLDL, oxidized low-density lipoprotein; HUVECs, human umbilical
cord endothelial cells; PCSK9, proprotein convertase
subtilisin/kexin type 9. |
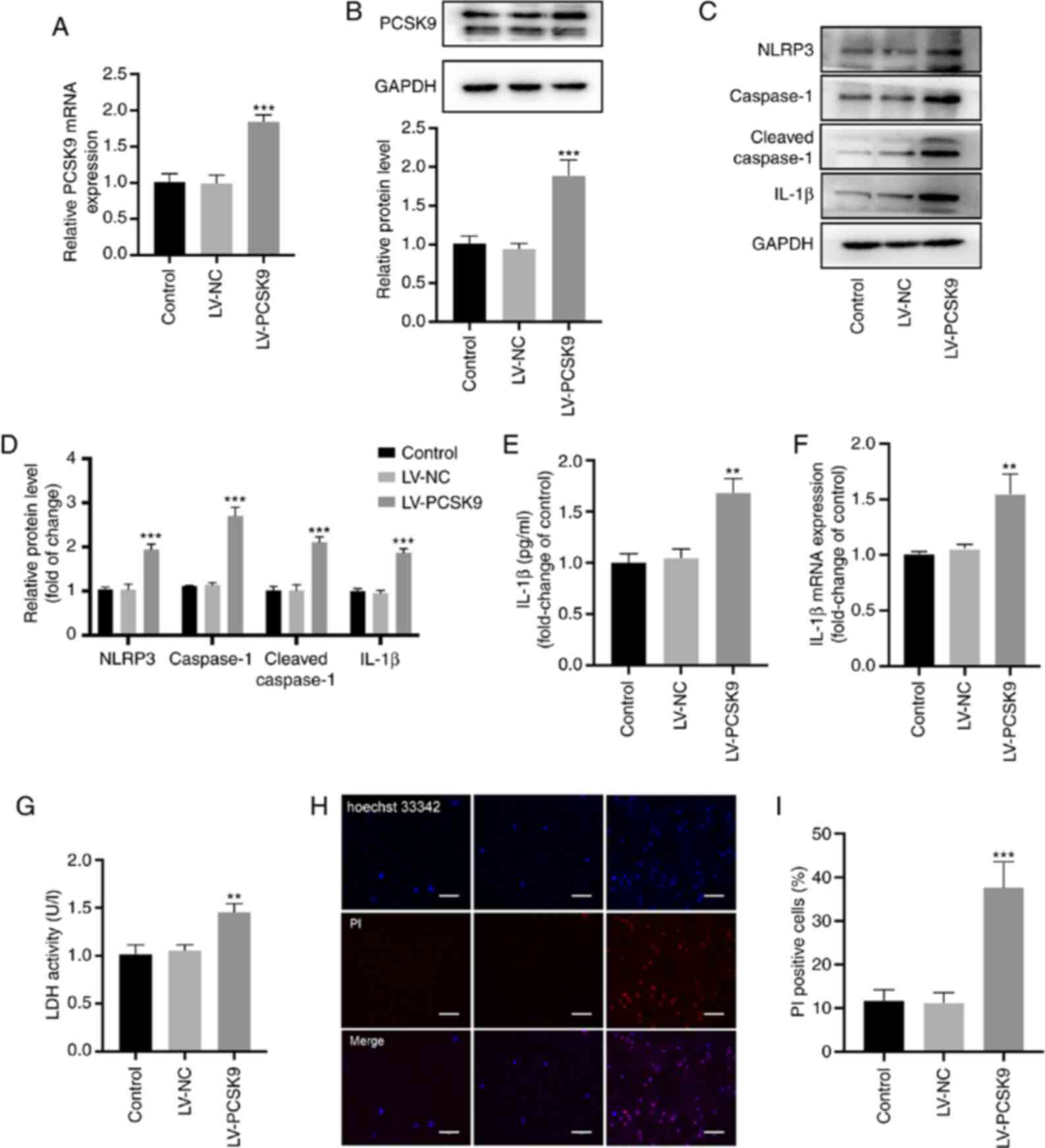
Overexpression of PCSK9 induces the
pyroptosis of HUVECs
HUVECs were transfected with LV-PCSK9 to explore the
association between PCSK9 and pyroptosis. The mRNA and protein
levels of PCSK9 significantly increased following transfection with
LV-PCSK9 (Fig. 4A and B). In
addition, NLRP3, caspase-1 cleaved caspase-1 and IL-1β protein
expression increased following the overexpression of PCSK9
(Fig. 4C and D). The protein and
mRNA expression of IL-1β (Fig. 4E
and F) and the LDH level (Fig.
4G) in the supernatant increased following the overexpression
of PCSK9. In addition, the ratio of PI-positive cells significantly
increased following the overexpression of PCSK9 (Fig. 4H and I). These results indicate
that PCSK9 overexpression induces the pyroptosis of HUVECs.
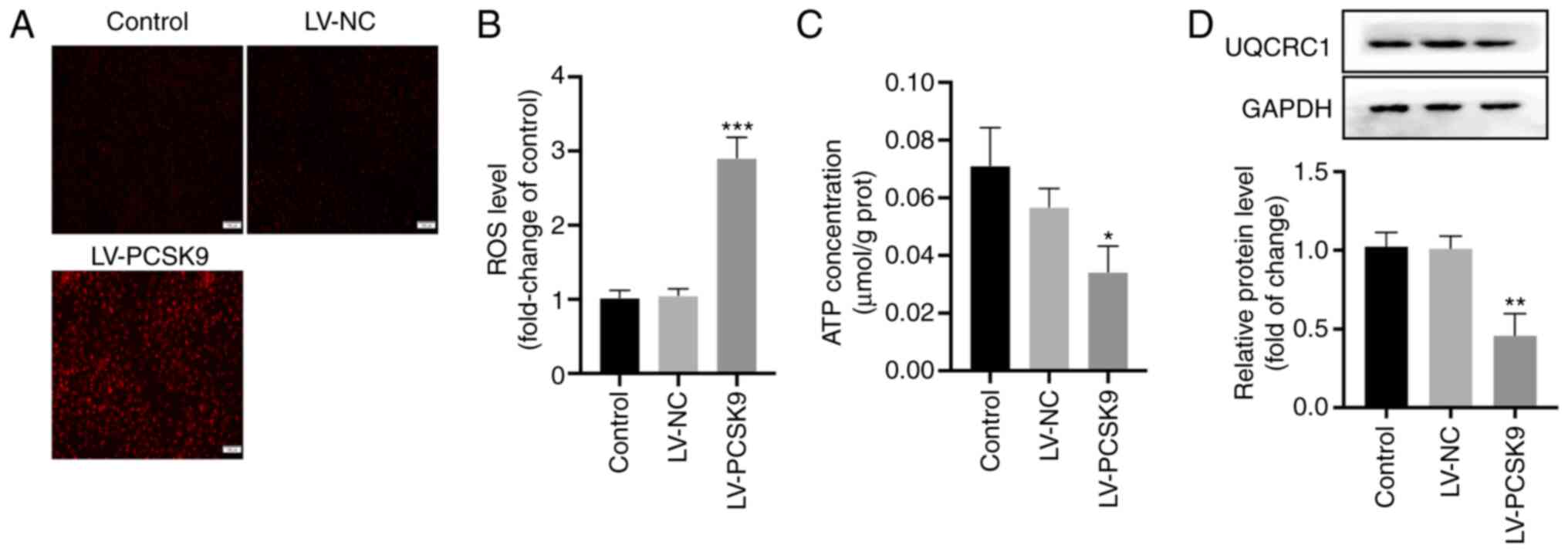
PCSK9 overexpression increases ROS
generation and induces mitochondrial dysfunction by inhibiting
UQCRC1
Excessive ROS production caused by mitochondrial
dysfunction can promote the pyroptosis of endothelial cells
(9) and reduce ATP production
(14). In the present study, the
intracellular ROS levels were detected following transfection with
LV-PCSK9 to determine whether PCSK9 promotes the pyroptosis of
endothelial cells through the ROS pathway. PCSK9 overexpression
elevated the ROS levels (Fig. 5A and
B) and inhibited ATP production (Fig. 5C). UQCRC1 is a subunit of
mitochondrial respiratory chain complex III and maintains
mitochondrial function (14).
UQCRC1 expression was thus examined following the overexpression of
PCSK9 in order to determine the association between PCSK9 and
UQCRC1. The data indicated that PCSK9 overexpression inhibited the
protein expression of UQCRC1 (Fig.
5D). These results suggest that PCSK9 increases ROS production
and causes mitochondrial disorder by inhibiting UQCRC1.
siPCSK9 reverses mitochondrial
dysfunction induced by oxLDL by recovering UQCRC1 expression
HUVECs were incubated with siPCSK9 for 24 h and then
stimulated with oxLDL (100 µg/ml) to investigate whether
oxLDL inhibits UQCRC1 expression through PCSK9. The data indicated
that UQCRC1 expression was significantly increased following
transfection with siPCSK9 compared with that after oxLDL
stimulation alone (Fig. 6A). The
ROS level significantly decreased following transfection with
siPCSK9, but was significantly increased following transfection
with siPCSK9 and siUQCRC1 (the transfection efficiency is shown in
Fig. 6B and C) compared with
that following transfection with siPCSK9 only (Fig. 6D and E). The cell membrane
potential level was also examined by JC-1 staining. The green
fluorescence was higher in the oxLDL group than in the control
group and recovered compared with that in the siPCSK9-transfected
group. However, transfection with siPCSK9 and siUQCRC1 enhanced the
intensity of green fluorescence (Fig. 6F). Flow cytometry revealed that
the area in the Q2 quadrant decreased, whereas the areas in the Q3
and Q4 quadrants increased following oxLDL stimulation. PCSK9
interference increased the area in Q2 quadrant. The simultaneous
interference of PCSK9 and UQCRC1 decreased the area in the Q2
quadrant, but increased the areas in the Q3 and Q4 quadrants
(Fig. 6F and G). These results
indicate that oxLDL inhibits UQCRC1 expression, promotes
mitochondrial membrane potential collapse, and damages
mitochondrial function, and these processes can be reversed PCSK9
interference.
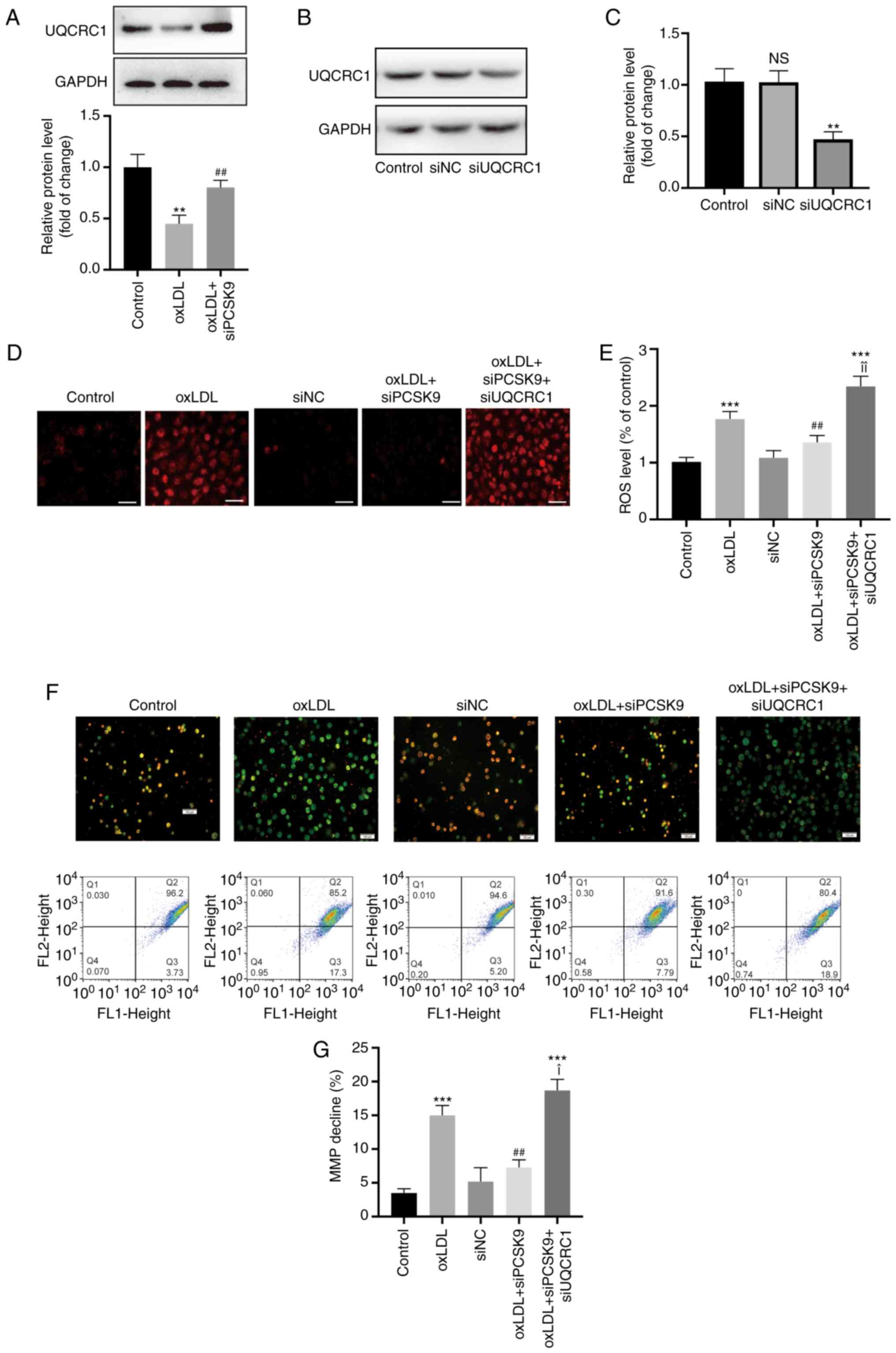 | Figure 6Interference with PCSK9 attenuates
excessive production of ROS. (A) Expression of UQCRC1 following
stimulation with oxLDL (100 µg/ml), and the effect of
interference with PCSK9 expression. **P<0.01 compared
with the control group; ##P<0.01 compared with the
oxLDL group. (B and C) Expression of UQCRC1 protein in HUVECs
following interference with UQCRC1 (50 nM, 24 h). Western blot
analysis was used to measure protein expression.
**P<0.01 compared with the control group. (D and E)
Mitochondrial ROS level following stimulation with oxLDL (100
µg/ml), and the effect of interference with PCSK9 expression
and UQCRC1 expression. Original magnification, ×200; scale bar, 20
µm. ***P<0.001 compared with the control
group; ##P<0.01 compared with the oxLDL group;
¥¥P<0.01 compared with the oxLDL + siPCSK9 group. (F)
Changes in mitochondrial membrane potential as observed by
fluorescence microscopy. Original magnification, ×100; scale bar,
100 µm. (G) Mitochondrial membrane potential levels measured
by flow cytometry. ***P<0.001 compared with the
control group; ##P<0.01 compared with the oxLDL
group; ¥P<0.05 compared with the oxLDL + siPCSK9
group. All results are expressed as the means ± SD of 3 independent
experiments. oxLDL, oxidized low-density lipoprotein; HUVECs, human
umbilical cord endothelial cells; PCSK9, proprotein convertase
subtilisin/kexin type 9; UQCRC1, ubiquinol-cytochrome c
reductase core protein 1. |
Discussion
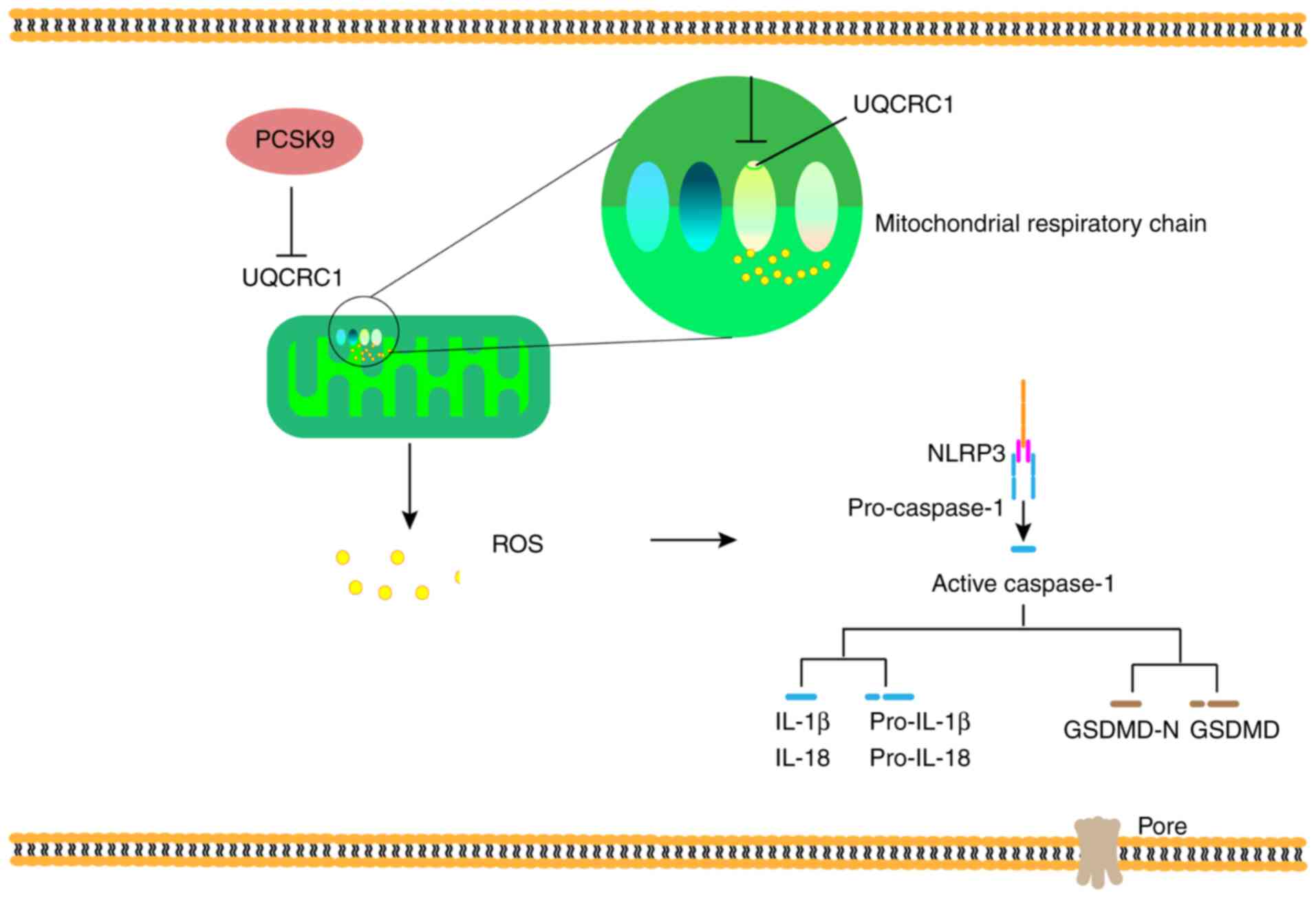
The present study demonstrated that PCSK9
interference attenuated HUVEC pyroptosis induced by oxLDL. On the
one hand, oxLDL induced the pyroptosis of HUVECs in a
concentration-dependent manner and upregulated PCSK9 expression.
PCSK9 interference relieved the pyroptosis induced by oxLDL. On the
other hand, PCSK9 overexpression promoted pyroptosis, UQCRC1
downregulation and ROS generation. siPCSK9 can alleviate the
inhibition of UQCRC1 expression by oxLDL, ROS production, and the
disorder of mitochondrial membrane potential (Fig. 7).
AS is a chronic inflammatory disease accompanied by
endothelial dysfunction and lipid deposition (1). Endothelial dysfunction is the
initial cause of AS. In AS lesions, endothelial cells are often
accompanied by pyroptosis (15),
which is associated with the activation of related inflammatory
bodies (NLRP3 and caspase-1) and the release of inflammatory
factors (IL-1β and IL-18) and cell dysfunction to stimulate the
recruitment of immune cells and then promote AS (16). However, the mechanisms underlying
pyroptosis remain unclear. NLRP3 and pyrin in cells can recruit
inflammatory bodies to form mature bodies, thus leading to the
maturation of inflammatory factors and the activation of gasdermin
family (17). ROS can activate
NLRP3 and mediate pyroptosis (18). In the present study, PCSK9
overexpression induced pyroptosis and increased ROS production.
PCSK9 is involved in the pathogenesis of AS by
regulating lipid metabolism (19,20) as confirmed by the clinical
lipid-lowering effect of PCSK9 inhibitors (21). This enzyme binds to LDLR,
promotes its intracellular degradation, and then reduces LDLR level
and LDL-C metabolism (19).
PCSK9 is also involved in cell death and inflammation. Low shear
stress can promote the expression of lectin-like oxidized
low-density lipoprotein receptor-1 (LOX-1) by activating ROS and
NF-κB, and the upregulation of LOX-1 can upregulate PCSK9
expression and vice versa (11).
In the present study, PCSK9 overexpression promoted the pyroptosis
of endothelial cells, and its interference alleviated the
pyroptosis induced by oxLDL. These results suggest that PCSK9 is an
important mediating factor in pyroptosis and ROS production.
However, the mechanisms through which PCSK9 upregulates ROS
expression have been rarely reported, at least to the best of our
knowledge.
The mitochondria can regulate cell function by
releasing signal molecules (10). Excessive inflammatory stimulation
and oxidative stress can promote the disorder of mitochondrial
regulation (22). oxLDL inhibits
mitochondrial gene expression (14), induces mitochondrial dysfunction,
and promotes AS (23). Moreover,
PCSK9 is closely related to mitochondrial function and oxidative
damage (24,25), promotes the mitochondrial DNA
damage of endothelial cells, and facilitates the occurrence of
apoptosis (26). However, the
association between PCSK9 and mitochondrial damage remains unclear.
UQCRC1 is a subunit of mitochondrial complex III, and its
interference in endothelial cells can lead to excess ROS production
(14). The present study
revealed that PCSK9 inhibited the function of mitochondrial complex
III by downregulating UQCRC1, leading to ROS elevation. This
finding also suggests that PCSK9 is an important mediator of the
oxidative damage mediated by oxLDL through mitochondrial-related
DNA. Elevated ROS levels have a pathological potential in leading
mtDNA mutation and oxidative damage to the respiratory chain
(27), which also increased
following the overexpression of PCSK9 in the present study.
Therefore, the inhibition of UQCRC1 may result from the ROS level
regulated by PCSK9. However, ROS elevation also upregulates PCSK9
expression and damages mitochondrial genes (11). Therefore, ROS and PCSK9 can be
regulated by mitochondrial genes (such as UQCRC1) and then
participate in the occurrence and development of AS.
The present study has some limitations. First, the
extra-cellular level of PCSK9 was not detected under the effect of
oxLDL. Second, the mechanism through which PCSK9 inhibits the
expression of UQCRC1 was not elucidated. Third, the effect of
UQCRC1 defect was only observed in vitro, and in vivo
experiments were not conducted. These limitations also provide
directions for future studies.
In conclusion, the present study demonstrates that
oxLDL promotes the pyroptosis of HUVECs and simultaneously
upregulates PSCK9 expression, leading to mitochondrial dysfunction
and ROS production resulting from UQCRC1 inhibition. However, PCSK9
interference blocks oxLDL-induced UQCRC1 inhibition, pyroptosis and
mitochondrial ROS production in HUVECs. These results indicate that
UQCRC1 plays a potential role in the antioxidant response and AS
treatment.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL and ZW contributed to the conception and design
of the study. JZ, JT and LX performed the experiments, collected
the data and analyzed the data and assisted in the interpretation
of the results. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Moss JW and Ramji DP: Cytokines: Roles in
atherosclerosis disease progression and potential therapeutic
targets. Future Med Chem. 8:1317–1330. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Libby P, Ridker PM and Hansson GK:
Progress and challenges in translating the biology of
atherosclerosis. Nature. 473:317–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Glass CK and Witztum JL: Atherosclerosis.
Cell. 104:503–516. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Horton JD, Cohen JC and Hobbs HH:
Molecular biology of PCSK9: Its role in LDL metabolism. Trends
Biochem Sci. 32:71–77. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hedrick JA: Targeting PCSK9 for the
treatment of hypercholesterolemia. Curr Opin Invest Drugs.
10:938–946. 2009.
|
|
6
|
Xu YJ, Zheng L, Hu YW and Wang Q:
Pyroptosis and its relationship to atherosclerosis. Clin Chim Acta.
476:28–37. 2018. View Article : Google Scholar
|
|
7
|
Hoseini Z, Sepahvand F, Rashidi B,
Sahebkar A, Masoudifar A and Mirzaei H: NLRP3 inflammasome: Its
regulation and involvement in atherosclerosis. J Cell Physiol.
233:2116–2132. 2018. View Article : Google Scholar
|
|
8
|
Shah PK and Lecis D: Inflammation in
atherosclerotic cardiovascular disease. F1000Res. 8:F10002019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhaolin Z, Jiaojiao C, Peng W, Yami L,
Tingting Z, Jun T, Shiyuan W, Jinyan X, Dangheng W, Zhisheng J and
Zuo W: OxLDL induces vascular endothelial cell pyroptosis through
miR-125a-5p/TET2 pathway. J Cell Physiol. 234:7475–7491. 2019.
|
|
10
|
Mottis A, Herzig S and Auwerx J:
Mitocellular communication: Shaping health and disease. Science.
366:827–832. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ding Z, Liu S, Wang X, Deng X, Fan Y, Sun
C, Wang Y and Mehta JL: Hemodynamic shear stress via ROS modulates
PCSK9 expression in human vascular endothelial and smooth muscle
cells and along the mouse aorta. Antioxid Redox Signal. 22:760–771.
2015. View Article : Google Scholar :
|
|
12
|
Bao HL, Liao FJ, Fang L, Zhong F, Liu W
and Li JQ: Effect and mechanism of PCSK9 on lectin-like oxidized
low-density lipoprotein receptor-1 mediated oxidized low-density
lipoprotein uptake by THP-1 derived macrophages. Zhonghua Xin Xue
Guan Bing Za Zhi. 47:367–373. 2019.In Chinese. PubMed/NCBI
|
|
13
|
Wu CY, Tang ZH, Jiang L, Li XF, Jiang ZS
and Liu LS: PCSK9 siRNA inhibits HUVEC apoptosis induced by ox-LDL
via bcl/Bax-caspase9-caspase3 pathway. Mol Cell Biochem.
359:347–358. 2012. View Article : Google Scholar
|
|
14
|
Chen JJ, Tao J, Zhang XL, Xia LZ, Zeng JF,
Zhang H, Wei DH, Lv YC, Li GH and Wang Z: Inhibition of the
ox-LDL-induced pyroptosis by FGF21 of human umbilical vein
endothelial cells through the TET2-UQCRC1-ROS pathway. DNA Cell
Biol. 39:661–670. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davies PF: Hemodynamic shear stress and
the endothelium in cardiovascular pathophysiology. Nat Clin Pract
Cardiovasc Med. 6:16–26. 2009. View Article : Google Scholar
|
|
16
|
Zhang L, Yuan M, Zhang L, Wu B and Sun X:
Adiponectin alleviates NLRP3-inflammasome-mediated pyroptosis of
aortic endothelial cells by inhibiting FoxO4 in arteriosclerosis.
Biochem Biophys Res Commun. 514:266–272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang W, Lin J, Dong J and Li D:
Pyroptosis: An inflammatory cell death implicates in
atherosclerosis. Med Hypotheses. 81:484–486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou R, Yazdi AS, Menu P and Tschopp J: A
role for mitochondria in NLRP3 inflammasome activation. Nature.
469:221–225. 2011. View Article : Google Scholar
|
|
19
|
Lin XL, Xiao LL, Tang ZH, Jiang ZS and Liu
MH: Role of PCSK9 in lipid metabolism and atherosclerosis. Biomed
Pharmacother. 104:36–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vozenilek AE, Vetkoetter M, Green JM, Shen
X, Traylor JG, Klein RL, Orr AW and Woolard MD: Absence of
nicotinamide nucleotide transhydrogenase in C57BL/6J mice
exacerbates experimental atherosclerosis. J Vasc Res. 55:98–110.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ruscica M, Ferri N, Corsini A and Sirtori
CR: PCSK9 antagonists and inflammation. Atherosclerosis.
268:235–236. 2018. View Article : Google Scholar
|
|
22
|
Aarreberg LD, Esser-Nobis K, Driscoll C,
Shuvarikov A, Roby JA and Gale M Jr: Interleukin-1β induces mtDNA
release to activate innate immune signaling via cGAS-STING. Mol
Cell. 74:801–815 e806. 2019. View Article : Google Scholar
|
|
23
|
He X, Wang L, Chen XF, Liang Q, Wang WQ,
Lin AQ, Yi L, Wang Y and Gao Q: Metformin improved oxidized
low-density lipoprotein-impaired mitochondrial function and
increased glucose uptake involving akt-AS160 pathway in raw264.7
macro-phages. Chin Med J (Engl). 132:1713–1722. 2019. View Article : Google Scholar
|
|
24
|
Thonusin C, Apaijai N, Jaiwongkam T,
Kerdphoo S, Arunsak B, Amput P, Palee S, Pratchayasakul W,
Chattipakorn N and Chattipakorn SC: The comparative effects of high
dose atorvastatin and proprotein convertase subtilisin/kexin type 9
inhibitor on the mitochondria of oxidative muscle fibers in
obese-insulin resistant female rats. Toxicol Appl Pharmacol.
382:1147412019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ding Z, Pothineni NVK, Goel A, Luscher TF
and Mehta JL: PCSK9 and inflammation: Role of shear stress,
pro-inflammatory cytokines, and LOX-1. Cardiovasc Res. 116:908–915.
2020. View Article : Google Scholar
|
|
26
|
Ding Z, Liu S, Wang X, Mathur P, Dai Y,
Theus S, Deng X, Fan Y and Mehta JL: Cross-Talk between PCSK9 and
damaged mtDNA in vascular smooth muscle cells: Role in apoptosis.
Antioxid Redox Signal. 25:997–1008. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Massaad CA and Klann E: Reactive oxygen
species in the regulation of synaptic plasticity and memory.
Antioxid Redox Signal. 14:2013–2054. 2011. View Article : Google Scholar :
|