Introduction
Keloids are benign fibroproliferative tumors that
commonly occur in response to tissue injury. These pathological
scars are characterized by hyperproliferation of fibroblasts and
abnormal deposition of collagen fibers (1,2).
Although keloids are benign hyperplasias, they induce esthetic
deformity and dermatological dysfunction by invading adjacent
normal tissue (3,4). In addition, frequent
keloid-associated episodes of pain and itching also result in
psychological and physical distress. No satisfactory treatment
strategy has been developed, primarily because of the high
recurrence rate of keloids, which can reach up to 70-80%
post-excision (5,6). Extensive research has indicated
that keloid development is complex and involves alterations in
non-coding RNAs, DNA methylation and histone modification (7-9);
however, the precise pathological mechanisms underlying how keloid
formation and progression are initiated and regulated remain
unknown, which has hindered the development of novel therapeutic
strategies.
Accumulating evidence has indicated that circular
(circ) RNAs produced from precursor mRNA backsplicing are
aberrantly expressed in multiple diseases and are associated with
cell proliferation, apoptosis and metastasis (10,11). For example, circ-low density
lipoprotein receptor class A domain containing 3, a
well-characterized circRNA, regulates the proliferation, migration
and invasion of pancreatic cancer cells via the microRNA (miRNA or
miR)-137-3p/PTEN axis (12).
Additionally, circ-collagen type III α 1 chain-859267 has been
shown to regulate the expression levels of type I collagen by
sponging miR-29c in human dermal fibroblasts (13). Shi et al (14) investigated circRNA expression
profiles in keloids via microarray analysis and found that a number
were abnormally expressed, including 24 that were significantly
downregulated and 52 markedly upregulated in keloids compared with
normal skin tissue. Therefore, it was hypothesized that circRNAs
may be involved in the regulation of keloid progression.
circ-nuclear receptor interacting protein 1 (NRIP1) originates from
the NRIP1 gene located on chromosome 21 and is the product
of the head-to-tail splicing of exons 2 and 3 (15). Several studies have investigated
the expression profile and function of circNRIP1 in various types
of cancer (16,17). One study (15) showed that circNRIP1 promotes the
proliferation, migration and invasion of gastric cancer cells via
the AKT1/mTOR pathway, while another study (18) reported that silencing of
circNRIP1 inhibits proliferation and induces apoptosis in breast
cancer. Although the contribution of circNRIP1 to tumor growth and
metastasis has been widely reported (16,17), pronounced knowledge gaps remain
regarding the biology of keloid progression. Recent studies have
shown that dysregulation of circRNAs can affect gene expression
profiles by creating complex regulatory networks through
interactions with proteins and RNA (19-21). Du et al (22) revealed that circ-Foxo3 promotes
mouse double minute 2 homolog-induced p53 ubiquitination and
degradation, causing an overall decrease in the levels of p53.
circβ-catenin stabilizes β-catenin by impeding GSK3β-mediated
β-catenin phosphorylation and degradation, leading to activation of
the Wnt pathway (23). It was
hypothesized that circNRIP1 may harbor a putative binding site for
the RNA-binding protein Fragile-X mental retardation 1 (FXR1) and
that circNRIP1 may be involved in keloid progression via regulating
FXR1.
The aim of the present study was to investigate the
putative role of circNRIP1 in keloid progression and the underlying
regulatory mechanisms by performing loss-of-function, RNA pulldown
and RNA immunoprecipitation assays to provide novel insight into
keloid progression and potential therapeutic options.
Materials and methods
Patient tissue samples
Between January 2016 and August 2018, 50 keloid and
50 matched normal skin tissue samples were obtained from 50
patients (21 males and 29 females) in Zaozhuang Municipal Hospital
(Zaozhuang, China). Diagnosis of hypertrophic scar was confirmed
via routine pathological examination with clinical symptoms such as
pain, growth, hyperemia and pruritus. Following excision of keloid
tissues at the Department of Plastic Surgery, skin samples were
immediately stored in liquid nitrogen for further processing. A
total of 50 patients was enrolled (age, 31.86±6.37 years) who
exhibited non-pedunculated keloid on the chest (n=18), back (n=17)
and upper arm (n=15) for ≥1 year. The etiology of hypertrophic
scarring included acne (n=18), blister (n=13), post-operative
(n=14) and unknown (n=5). Furthermore, none of the patients had
received systemic and topical therapy for >60 days before
pathological examination. The present study was approved by the
Clinical Research Ethics Committee of Zaozhuang Municipal Hospital.
Additionally, written informed consent was obtained from all
patients.
Cell culture
Primary cultures of keloid-derived fibroblasts and
normal dermal fibroblasts were established as previously described
(2). Briefly, keloid and matched
normal skin tissue were excised and dissected into 1-mm3 pieces.
Subsequently, the pieces were digested with 0.25% trypsin at 37°C
for 3 min. After washing and mechanical dissociation, the
fibroblasts were filtered, centrifuged (500 × g for 5 min at 25°C)
and resuspended in DMEM supplemented with 10% fetal bovine serum
(both Thermo Fisher Scientific, Inc.) at 37°C in a tissue culture
incubator with 5% CO2. The medium was changed every 3
days. Fibroblasts from generation 2 to 5 were used in subsequent
experiments.
RNA extraction and reverse
transcription-quantitative (RT-q) PCR
Total RNA from fibroblasts and tissue was extracted
using the RNAeasy Gamma Animal RNA Extraction kit (Beyotime
Institute of Biotechnology). Following RNase-R treatment, the M-MLV
universal RT-PCR kit (Beijing Solarbio Science & Technology
Co., Ltd.) was used for cDNA synthesis, according to the
manufacturer's instructions. The expression levels of circRNAs and
miRNAs was evaluated by qPCR using the SuperScript IV RT-PCR kit
(Thermo Fisher Scientific, Inc.). The reaction conditions for PCR
(5 µl cDNA) were as follows: 95°C for 10 min, followed by 40
cycles of 93°C for 15 sec and 54°C for 60 sec. GAPDH and
U6 were used as controls for circRNA and miRNA,
respectively. The 2−ΔΔCq method was employed to
calculate relative circRNA and miRNA levels (24). The primers were designed and
synthesized by Sangon Biotech Co., Ltd. The primer sequences were
as follows: circNRIP1, forward, 5′-AGTTGCTCCAATGACAGAGTTACC-3′ and
reverse, 5′-CCTCCTTCAGTCAAGTGTGCATC-3′; pre-miR-503, forward,
5′-TGCCCTAGCAGCGGGAAC-3′ and reverse, 5′-ACCCTGGCAGCGGAAACAATA-3′;
miR-503-3p, forward, 5′-GGGGTATTGTTTCCGCTGCCAGG-3′, reverse,
5′-CCTGGCAGCGGAAACAATACCCC-3′ and RT,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCTGGC-3′;
miR-503-5p, forward, 5′-CGTAGCAGCGGGAACAGTT-3′, reverse,
5′-AGTGCAGGGTCCGAGGTATT-3′ and RT,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGCAG-3′; GAPDH,
forward, 5′-GTCAACGGATTTGGTCGTATTG-3′ and reverse,
5′-CCGTTCTCAGCCATGTAGTT-3′; and U6, forward,
5′-TCGGCAGCACATATACTAAAAT-3′, reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′ and RT,
5′-CGCTTCACGAATTTGCGTGTCAT-3′.
Cell transfection
Keloid-derived fibroblasts (1×105 per
well) were seeded in a 6-well plate and grown overnight at 37°C.
Small interfering (si)RNA targeting NRIP1 or FXR1 was
cloned into the pENTR H1 vector. Plasmid vectors (pcDNA- circNRIP1:
siNRIP1#1, siNRIP1#2; pcDNA-FXR1: siFXR1#1, siFXR1#2) were designed
and synthesized by Sangon Biotech Co., Ltd. A mock vector with no
siRNA sequence was used as a control. miR-503 precursor pGCMV
vectors were provided by Shanghai GenePharma Co., Ltd. Front and
back circular frames were synthesized by Sangon Biotech Co., Ltd.
and inserted into the pcDNA3.1 (+) circRNA mini vector for
transcript circularization. The expression vectors for Flag-tagged
MS2 coat protein were provided by Weizhen Biotech Co., Ltd.
Flag-tagged expression vectors for full-length Fbxo4 and HA-tagged
expression vectors for full-length FXR1 were provided by Weizhen
Biotechnology. Keloid-derived fibroblasts were transfected with the
corresponding plasmids (25 nM) for 2 days at 37°C using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Overexpression and
knockdown efficiency were monitored by RT-qPCR and western blotting
after transfection for 24 h. The targeted sequences were as
follows: siNRIP1#1, 5′-GACAGACGGAAGTGTTTGGAT-3′; siNRIP1#2,
5′-GAATCTGAAGACTCCGGAT-3′; siFXR1#1, 5′-GGCAAAGTGATCGGAAAGA-3′ and
siFXR1#2, 5′-CGAGCTGAGTGATTGGTCA-3′. The inhibitor sequences were
as follows: miR-503-3p, 5′-CCTGGCAGCGGAAACAATACCCC-3′; miR-503-5p,
5′-CTGCAGAACTGTTCCCGCTGCTA-3′ and control (ctrl),
5′-TGACTGTACTGACTCGACTG-3′.
Cell proliferation and colony formation
assay
For the Cell Counting Kit (CCK)-8 assay,
keloid-derived fibroblasts (3,000 per well) were plated in 96-well
plates. Following incubation for different durations (1, 2 and 3
days), cell viability was determined by adding 10 µl CCK-8
reagent to each well, according to the manufacturer's instructions
(Abmole Bioscience, Inc.). The absorbance (450 nm) was determined
using a microplate reader (Shenzhen Mindray Bio-Medical Electronics
Co., Ltd.). For the colony formation assay, keloid-derived
fibroblasts (3,000 per well) were plated in 6-well plates. After
incubation at 37°C for two weeks, keloid-derived fibroblasts were
fixed in 4% paraformaldehyde for 20 min at 25°C and then stained
with 0.1% crystal violet (Beyotime Institute of Biotechnology) for
20 min at 25°C. Visible colonies were observed using a WMS-1033
Digital light biomicroscope at 200× magnification (Shanghai Wumox
Optical Instrument Co., Ltd.). Images were captured using an EOS
M50 digital camera (Canon, Inc.) and quantified by three
observers.
Flow cytometric assays
For cell apoptosis, keloid-derived fibroblasts
underwent double staining with 20 ng/l FITC-Annexin V (10
µl, 30 min, 25°C) then 50 ng/l propidium iodide (5
µl, 5 min, 25°C) using the Annexin V-FITC cell apoptosis
detection kit (Beyotime Institute of Biotechnology) according to
the manufacturer's instructions. Cell apoptosis was analyzed using
a BD FACSCelesta flow cytometer (BD Biosciences). For cell cycle
analysis, keloid-derived fibroblasts were stained with propidium
iodide using the DNA Content Quantitation Assay kit (Beijing
Solarbio Science & Technology Co., Ltd.) following the
manufacturer's protocol and then analyzed by flow cytometry. The
percentage of fibroblasts in the G0/G1, S and
G2/M phases were counted using FlowJo10.4 software
(Becton, Dickinson & Company).
Western blot analysis
Total protein from fibroblasts and tissue was
extracted on ice using the Total Extraction Sample kit
(Sigma-Aldrich; Merck KGaA). The protein concentration was
determined via BCA assay (Beyotime Institute of Biotechnology). 50
µg protein were separated by 12 or 8% SDS-PAGE and then
electrophoretically transferred onto PVDF membranes. Following
incubation with 5% non-fat milk for 2 h at 25°C, the membranes were
incubated with rabbit anti-FXR1 (1:1,000; cat. no. ab155124), mouse
anti-fibronectin (1:500; cat. no. ab6328), anti-collagen I
(1:1,000; cat. no. ab6308), anti-collagen III (1:1,000; cat. no.
ab6310), rabbit anti-SMN/Gemin 1 (1,1000; cat. no. ab108531),
anti-FBXO4 (1:2,000; cat. no. ab153803) and mouse anti-β actin
antibody (1:25,000; cat. no. ab49900; all Abcam) for 12 h at 4°C
and then with horseradish peroxidase-conjugated goat anti-rabbit or
anti-mouse IgG H&L secondary antibody (both 1:20,000; cat. nos.
ab205718 and ab6789, respectively; both Abcam) for 2 h at 25°C.
Protein bands were visualized using Cobas E
601electrochemiluminescence (Roche Diagnostics) and X-ray film.
ImageJ 2.1 software (National Institutes of Health) was used for
densitometric analysis. GAPDH (1:10,000; cat. no. ab181602; Abcam)
was used as an internal control.
Database analysis
The targets of circNRIP1 were acquired using the RNA
Interactome Database (rna-society.org/rnainter/home.html). The binding sites
of FXR1 om miR-503 were acquired through starBase (starbase.sysu.edu.cn/).
RNA pulldown and RNA immunoprecipitation
assay (RIPA)
For the RNA pulldown assay, a biotin-labeled RNA
probe complementary to circNRIP1 was designed and synthesized by
Sangon Biotech Co., Ltd. Then, 293T cells (1×107) and
keloid-derived fibroblasts were harvested, washed with ice-cold
PBS, lysed with RIPA Lysis Buffer (500 µl, Beyotime
Institute of Biotechnology) and incubated with biotinylated probes
(3 µg) for 2 h at 25°C. Streptavidin C1 magnetic beads (50
µl; Thermo Fisher Scientific, Inc.) were added to each
binding reaction followed by incubation at room temperature for 60
min. After briefly washing the beads with co-IP buffer, the bound
proteins were detected using western blotting as aforementioned.
For RIPA, 1×107 293T cells or keloid-derived fibroblasts
were harvested, washed with ice-cold PBS, lysed with RIPA Lysis
Buffer (500 µl; Beyotime Institute of Biotechnology) and
isolated using the RNAeasy Gamma Animal RNA Extraction kit
(TransGen Biotech Co., Ltd.). The cells were incubated with 5
µg primary antibody from Abcam [anti-FXR1 (1:1,000; cat. no.
ab155124), anti-Dicer (1:1,000; cat. no. ab227518), anti-GAPDH
(1:10,000; cat. no. ab181602; Abcam); anti-Flag (1:1,000; cat. no.
ab236777)] for 2 h at 4°C. Subsequently, each sample was
supplemented with 50% protein A-Sepharose slurry (50 µl) and
incubated for 4 h at 4°C. Following centrifugation (1,000 × g, 4°C
for 3 min) and three PBS washes, samples were resuspended in 0.5 ml
Tris reagent (Sigma-Aldrich; Merck KGaA) followed by elution. The
eluted, co-precipitated RNA was subjected to RT-qPCR as
aforementioned to measure the levels of the circRNA of interest.
Telomerase RNA component (TERC) was used as the positive
control.
Co-immunoprecipitation (CO-IP) assay
Cell lysate was produced using RIPA Lysis Buffer
(Beyotime Institute of Biotechnology) and supernatant were
collected. A total of 200 µl A/G beads (Thermo Fisher
Scientific, Inc.) were washed with pre-cooled PBS. Then, 2
µg antibody from Abcam [anti-Flag (1:30; cat. no. ab236777),
anti-HA (1:30; cat. no. ab236632)] was added, followed by
incubation for 3 h at 4°C. Following centrifugation (4,000 × g,
4°C, 60 sec) and washing (3 times), preprepared cell extracts (1
ml) were added and incubated overnight at 4°C in the presence of
phenylmethylsulfonyl fluoride (0.5 mM) and protease inhibitors (10
µl). Following centrifugation (4,000 × g, 4°C, 60 sec) and
washing (3 times) with bead wash solution, the precipitates were
boiled in loading buffer (60 µl) and incubated for 300-600
sec (100°C). Western blot analysis was conducted as aforementioned
to evaluate the co-IP results.
Northern blot assay
Northern blotting was conducted using a High
Sensitive miRNA Northern Blot Assay kit according to the
manufacturer's instructions (Signosis, Inc.). Total RNA (10
µg) was denatured with glyoxal loading dye at 50°C for 30
min, loaded onto agarose gels (1.2%) and electrophoresed for 150
min at 60 V. RNA was transferred by capillarity onto a Hybond N+
membrane (GE Healthcare) overnight in 10X saline sodium citrate
buffer at 4°C. After crosslinking using UV light (1,200 × 100
mJ/cm2), the membrane was washed with Tris for 20 min.
Subsequently, prehybridization (62°C, 2 h) and hybridization (62°C,
16 h) were performed in Northern Max buffer (Sigma-Aldrich; Merck
KGaA). After washing, the blots were visualized using two-color
infrared laser imaging (Gene Company, Ltd.). Then bands were
analyzed by ImageJ 1.8 software (National Institutes of
Health).
Statistical analysis
All statistical analysis was performed using
GraphPad Prism v.8 (GraphPad Software, Inc.). All data are
presented as the mean ± SEM of three independent repeats. Paired
Student's t-test was used to compare differences between two
groups. Significance between multiple groups was analyzed by
one-way ANOVA followed by Tukey's post hoc test. Correlations were
analyzed via Pearson's correlation analysis. P<0.05 and
P<0.01 were considered to indicate a statistically significant
difference.
Results
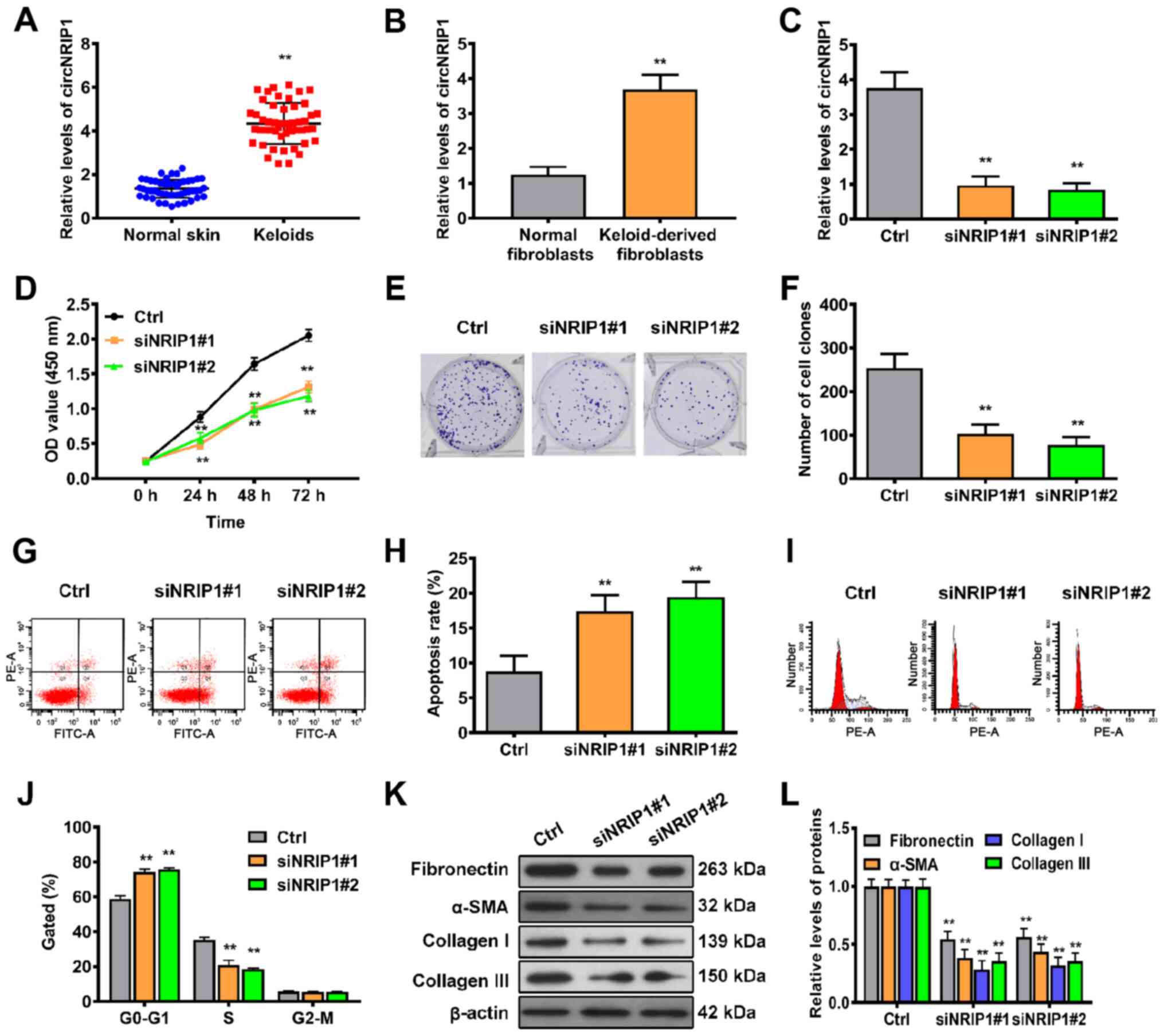
circNRIP1 is overexpressed in human
keloid tissue
In order to identify whether circNRIP1 is expressed
in human keloid tissue, 50 keloid and adjacent normal skin tissue
samples were collected. RT-qPCR showed that the levels of circNRIP1
in keloid tissue were 3.24-fold higher than those of adjacent
normal skin tissue (Fig. 1A).
Furthermore, the expression levels of circNRIP1 in keloid-derived
and human normal fibroblasts were compared. circNRIP1 expression
was significantly increased in keloid-derived fibroblasts compared
with that in normal human fibroblasts (Fig. 1B).
circNRIP1 knockdown suppresses
proliferation and extracellular matrix accumulation but promotes
cell apoptosis in keloid-derived fibroblasts
In order to confirm whether circ-NRIP1 participates
in the regulation of cell proliferation and apoptosis in
fibroblasts, expression of circNRIP1 was knocked down using siRNA
(Fig. 1C). Cell viability curves
constructed at different time points using CCK-8 assay revealed
that siRNA-mediated silencing of circNRIP1 significantly inhibited
cell viability compared with the control (Fig. 1D). In addition, colony formation
assay demonstrated that cell survival was significantly decreased
following inhibition of circNRIP1 (Fig. 1E and F). Subsequently, flow
cytometric analysis was used to evaluate whether circNRIP1
influenced the proliferative ability of keloid-derived fibroblasts
via effects on cell apoptosis and cycle progression. The rate of
apoptosis was significantly higher in circNRIP1-silenced
keloid-derived fibroblasts than in control cells (Fig. 1G and H). Additionally, circNRIP1
knockdown resulted in a marked increase in the
G0/G1 phase and decrease in the S phase of
the cell cycle compared with control fibroblasts (Fig. 1I and J). In order to assess
whether circNRIP1 has a role in extracellular matrix deposition,
the accumulation of extracellular matrix components was assessed by
western blotting. Expression of fibronectin, α-smooth muscle actin
(SMA), collagen I and III was decreased in circNRIP1-silenced
fibroblasts compared with that in control fibroblasts (Fig. 1K and L). Collectively, these
results indicated that circNRIP1 facilitated keloid
progression.
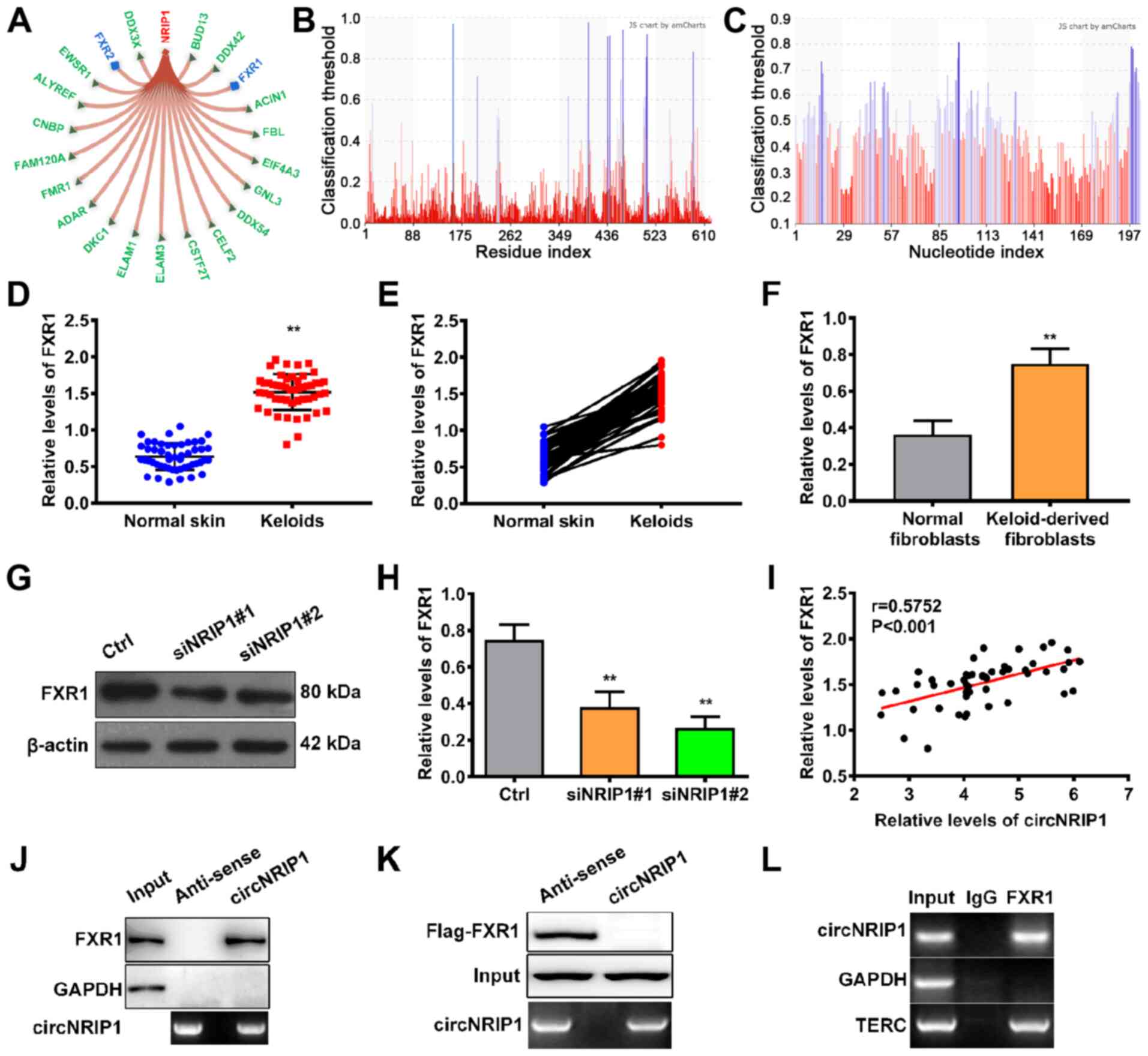
circNRIP1 stabilizes FXR1 by impeding
Fbxo4-mediated FXR1 ubiquitination
Next, the mechanism by which circNRIP1 facilitates
keloid-derived fibroblast proliferation and suppresses apoptosis
was investigated. Using the bioinformatic database RNAInter, 21
putative downstream targets of circNRIP1 were identified (Fig. 2A). Among these downstream
targets, members of the fragile X protein family (FXR1, FXR2 and
FMR1) were identified as binding partners. RNA binding protein FXR1
(score, 0.6116) is of particular interest due to the high score and
FXR1 biological function. The binding sites in FXR1 and circ- NRIP1
are shown in Fig. 2B and C,
respectively. Subsequently, the expression profile of FXR1 in
keloid tissue was assessed. Expression levels of FXR1 in keloid
tissue were 2.38-fold higher than in adjacent normal skin tissue
(Fig. 2D and E). Moreover, FXR1
expression was also higher in keloid-derived fibroblasts relative
to normal human fibroblasts (Fig.
2F), suggesting that FXR1 ws positively associated with
circNRIP1 expression in keloids. Furthermore, knockdown of
circNRIP1 led to a significant decrease in expression of FXR1
(Fig. 2G and H). Pearson's
correlation analysis also confirmed that circNRIP1 levels were
positively associated with the expression of FXR1 in keloid tissues
(Fig. 2I). These results
suggested that circNRIP1 was involved in maintaining FXR1
stability.
RNA pulldown assays were used to investigate the
regulatory mechanism involved in circNRIP1-mediated maintenance of
FXR1 stability. The results showed that circNRIP1 was pulled down
with endogenous FXR1 protein, but not with GAPDH (negative control;
Fig. 2J). RNA pulldown assays
using 293T cells transfected with Flag-FXR1 and circNRIP1-MS2 or
its antisense variant demonstrated that circNRIP1 was pulled down
with Flag-FXR1, but not Flag-GAPDH (Fig. 2K). RIPA was performed to validate
the interaction between circNRIP1 and FXR1. Compared with the
positive control (TERC), circNRIP1 was enriched in FXR1 binding
(Fig. 2L). These results
confirmed that circNRIP1 specifically interacted with the
RNA-binding protein FXR1 in keloid-derived fibroblasts.
In order to clarify how circNRIP1 regulates FXR1
stability, circNRIP1-depleted keloid-derived fibroblasts were
treated with proteasome inhibitor MG132. FXR1 expression was
significantly increased following circNRIP1 depletion in the
presence of MG132 compared with circNRIP1 depletion-alone (Fig. 3A and B). This suggested that the
role of circNRIP1 in maintaining FXR1 stability was associated with
proteasomal degradation. Knockdown of endogenous circNRIP1 in
fibroblasts notably enhanced the levels of endogenous FXR1
ubiquitination (Fig. 3C),
whereas FXR1 ubiquitination was markedly diminished by circNRIP1
overexpression in 293T cells (Fig.
3D). Combined, these data indicated that circNRIP1 stabilized
FXR1 protein by impeding its ubiquitination and proteasomal
degradation in keloid-derived fibroblasts. Subsequently, E3
ubiquitin ligase Fbxo4 was overexpressed, which led to a
significant increase in the expression levels of Fbxo4 (Fig. 3E and F). Moreover, circNRIP1
overexpression rescued Fbxo4-induced FXR1 degradation (Fig. 3E and G). circNRIP1 overexpression
also inhibited the interaction between FXR1 and Fbxo4: FXR1
co-immunoprecipitated with less Fbxo4, while Fbxo4 also
co-immunoprecipitated with less FXR1 (Fig. 3H and I). These results
collectively demonstrated that circNRIP1 blocked Fbxo4-mediated
FXR1 ubiquitination by inhibiting the interaction between FXR1 and
Fbxo4, thereby preventing FXR1 proteasomal degradation.
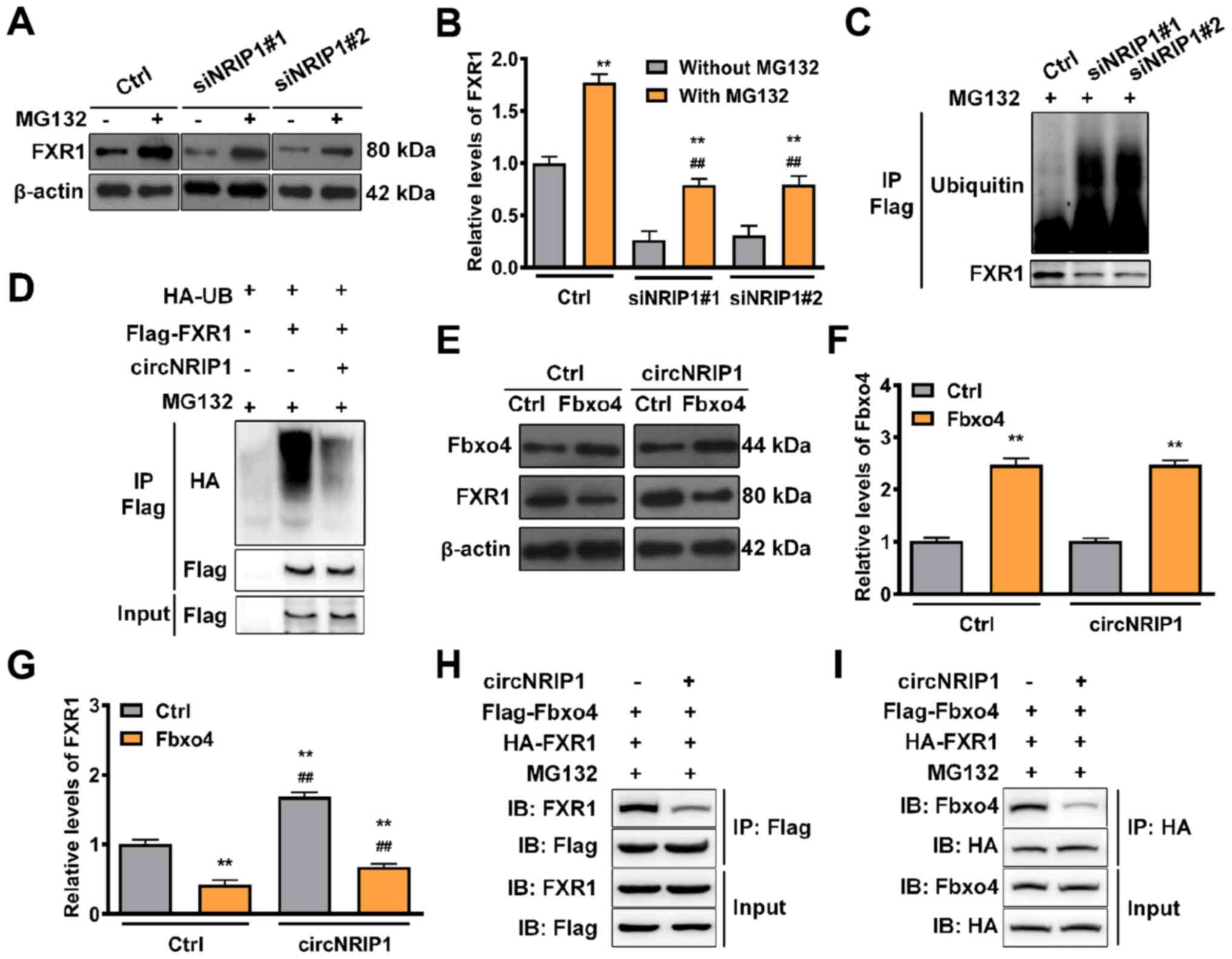 | Figure 3circNRIP1 protects FXR1 from
Fbxo4-mediated ubiquitination. (A and B) Expression of FXR1 in
keloid-derived fibroblasts in response to MG132 exposure (20
µmol/l) for 6 h was examined by western blot assay. (C)
Keloid-derived fibroblasts with circNRIP1 knockdown were
immunoprecipitated with anti-Flag antibody and immunoblotted with
an anti-HA-ubiquitin antibody following MG132 exposure for 6 h. (D)
293T cells were transfected with HA-UB, Flag-FXR1, or circNRIP1.
Whole-cell lysates were immunoprecipitated with the anti-Flag
antibody and then immunoblotted with anti-HA-ubiquitin antibody to
detect the interaction between FXR1 and Ubiquitin following MG132
exposure (20 µmol/l). (E) Expression of (F) Fbxo4 and (G)
FXR1 in keloid-derived fibroblasts transfected with Fbxo4 was
investigated by western blot analysis following circNRIP1
overexpression. 293T cells were transfected with Flag-Fbxo4,
HA-FXR1 and circNRIP1, followed by MG132 treatment (20
µmol/l, 6 h); cell lysates were immunoprecipitated with (H)
anti-Flag or (I) anti-HA antibody. The precipitates and inputs were
analyzed by western blot analysis. **P<0.01 vs. Ctrl
or MG132 treatment group. ##P<0.01 vs. MG132
treatment or Fbox4 overexpression group. circNRIP1, circular
nuclear receptor interacting protein 1; FXR1, Fbxo4-mediated FMR1
autosomal homolog 1; Ctrl, control; si, small interfering; IP,
immunoprecipitation; IB, immunoblot. |
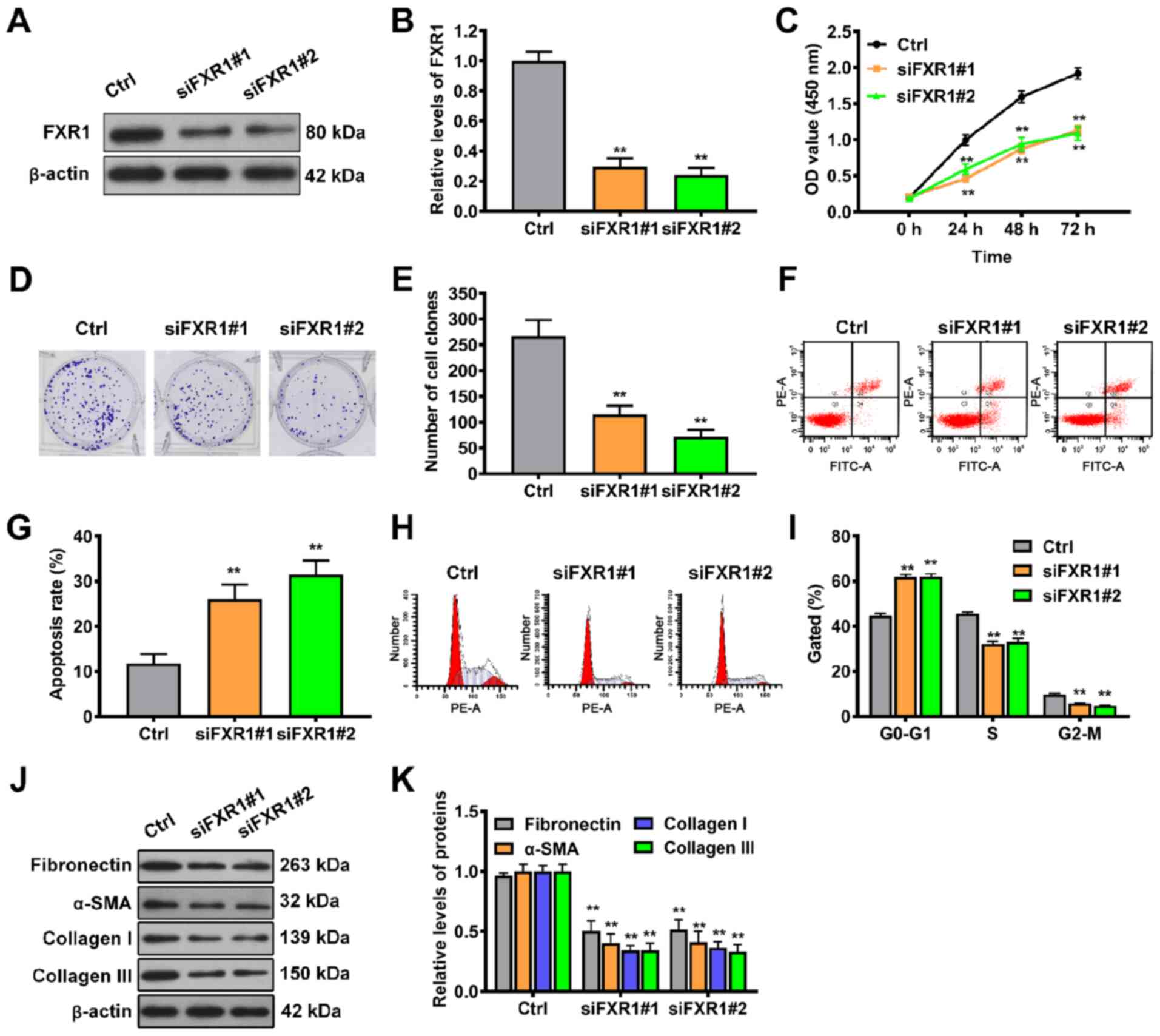
FXR1 drives keloid-derived fibroblast
proliferation and accumulation of extracellular matrix but inhibits
apoptosis
In order to evaluate whether FXR1 is functionally
involved in keloid progression, FXR1 expression was knocked down in
keloid-derived fibroblasts by siRNA transfection (Fig. 4A and B). Silencing FXR1 by
transfection of siRNA#1 and siRNA#2 significantly decreased cell
viability over 3 days (Fig. 4C).
Consistent with the cell viability data, FXR1 knockdown by either
of the two siRNAs also significantly decreased the colony-forming
ability of fibroblasts (Fig. 4D and
E). Next, the effect of FXR1 on cell apoptosis and cell cycle
progression in keloid-derived fibroblasts was assessed. The results
of flow cytometry showed that silencing FXR1 by siRNA#1 or siRNA#2
transfection significantly increased the rate of apoptosis of
keloid-derived fibroblasts compared with control cells (Fig. 4F and G). Additionally, there was
a significant increase in the number of FXR1-silenced fibroblasts
arrested at the G1/G0 phase compared with
that in control cells (Fig. 4H and
I). Expression levels of extracellular matrix components were
detected by western blot assay. Silencing FXR1 significantly
decreased the expression levels of fibronectin, α-SMA, collagen I
and III in circNRIP1-silenced fibroblasts compared with control
fibroblasts (Fig. 4J and K).
These data revealed that FXR1 served a vital role in keloid
progression.
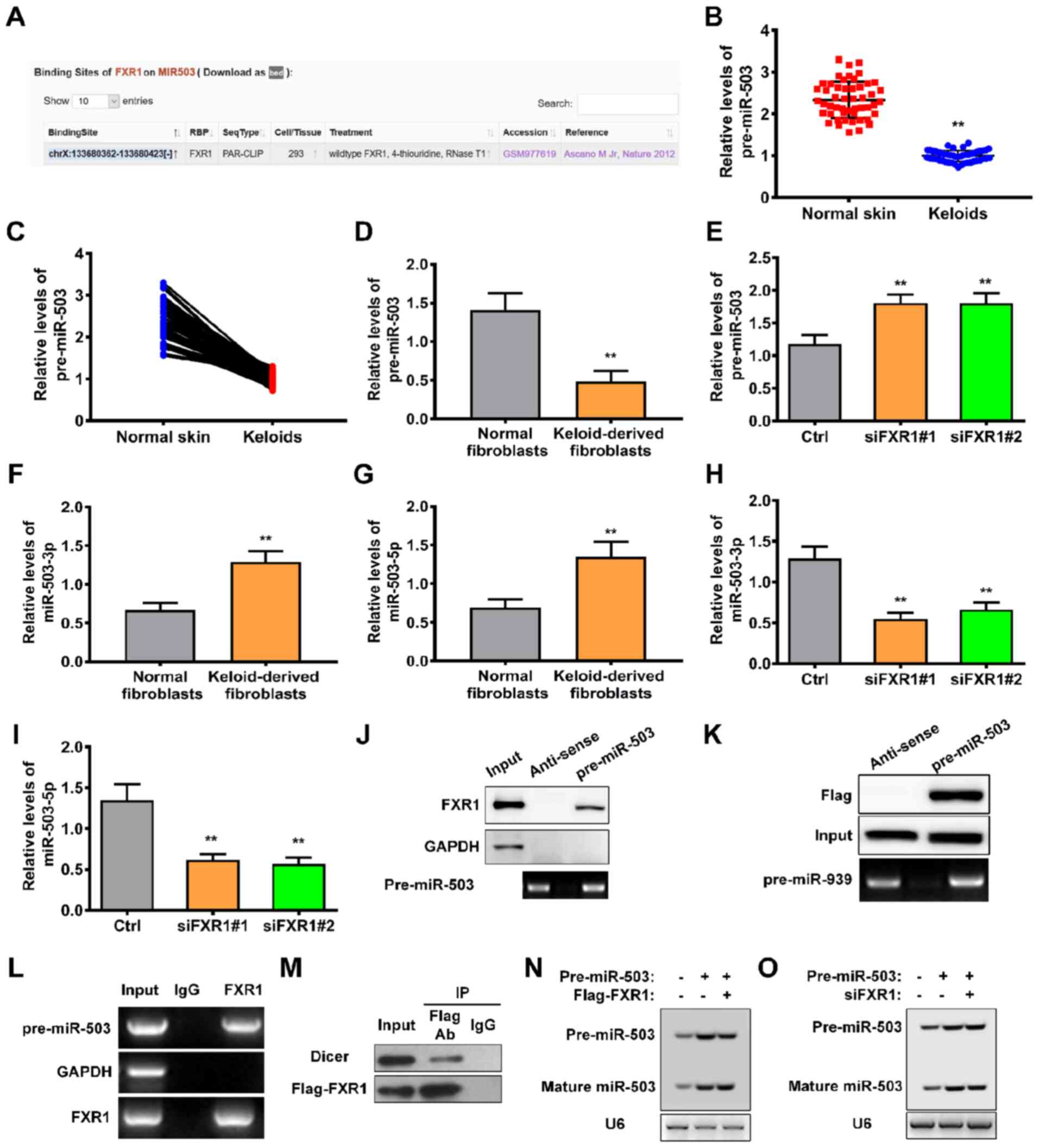
FXR1 modulates the maturation of
pre-miR-503
In order to elucidate the mechanism by which FXR1
regulates keloid progression, the potential interacting sites
between FXR1 and miRNAs were predicted using starBase. miR-503 of
82 predicted miRNAs were identified; binding sites in pre-miR-503
are shown in Fig. 5A. RT-qPCR
was performed to determine the levels of pre-miR-503 in keloid and
adjacent normal skin tissue. Levels of pre-miR-503 were
significantly decreased in keloid compared with adjacent normal
skin tissue (Fig. 5B and C).
Similar results were obtained in keloid-derived fibroblasts
(Fig. 5D). Moreover, the levels
of pre-miR-503 were significantly upregulated following knockdown
of FXR1 using either siRNA#1 or siRNA#2 (Fig. 5E). Subsequently, the expression
profiles of mature miR-503, miR-503-3p and miR-503-5p were
investigated. The levels of miR-503-3p and miR-503-5p were
significantly increased in keloid-derived fibroblasts (Fig. 5F and G). However, silencing of
FXR1 significantly decreased levels of miR-503-3p and miR-503-5p
(Fig. 5H and I). These results
indicated that FXR1 contributed to the maturation of miR-503.
In order to confirm this possibility, the
interaction between FXR1 and pre-miR-503 was assessed. RNA pulldown
assays demonstrated that pre-miR-503 specifically pulled down with
endogenous FXR1 protein but not the negative control GAPDH
(Fig. 5J). Consistent with this,
pre-miR-503 was able to pull down Flag-FXR1 in 293T cells (Fig. 5K). The interaction between
pre-miR-503 and FXR1 was also validated by RIPA, in which
pre-miR-503 was enriched for FXR1 binding (Fig. 5L). Furthermore, FXR1 formed
complexes with Dicer, a double-stranded RNA nuclease essential for
the biogenesis of numerous miRNAs, such as miRNA-103 (25) (Fig. 5M). In order to demonstrate a
direct role for FXR1 in pre-miR-503 processing, northern blot
analysis was performed using 293T cells transfected with both
siFXR1 and pre-miR-503. Upregulation of Flag-tagged FXR1 in 293T
cells increased the level of mature miR-503 (Fig. 5N); however, processing of
pre-miR-503 into mature miR-503 was impaired in the absence of FXR1
(Fig. 5O). Together, these
results demonstrated that FXR1 increased the levels of mature
miR-503 via regulating the processing of pre-miR-503.
Mature miR-503 promotes keloid-derived
fibroblast proliferation and accumulation of extracellular matrix
but inhibits apoptosis
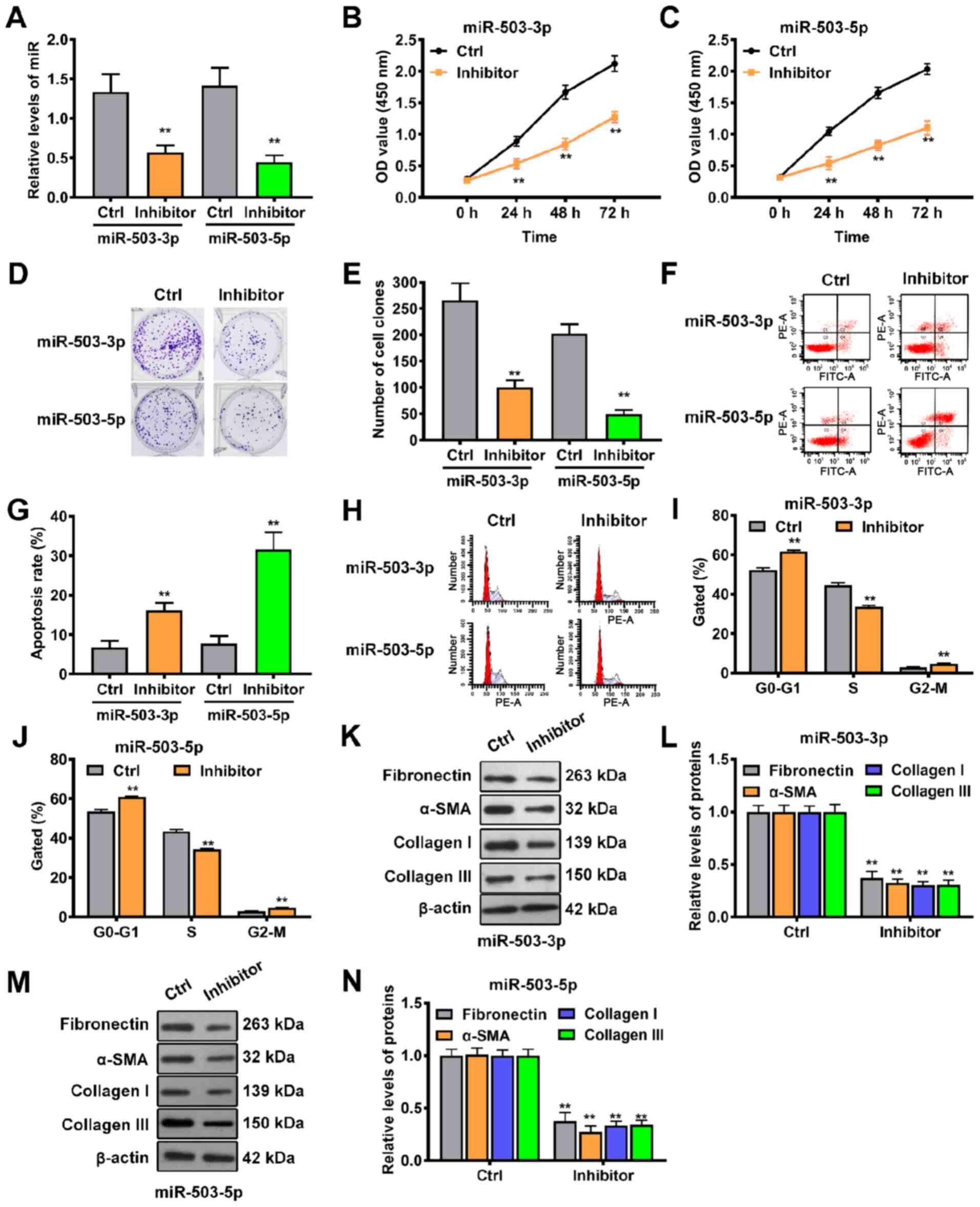
Next, the biological functions of miR-503-3p and
miR-503-5p in keloid progression were investigated through
loss-of-function experiments. miR-503-3p or miR-503-5p-specific
inhibitors were infected into keloid-derived fibroblasts, resulting
in knockdown of miR-503-3p or miR-503-5p (Fig. 6A). CCK-8 assay showed that cell
proliferation was suppressed by miR-503-3p or miR-503-5p knockdown
at different times (Fig. 6B and
C), while colony formation assay revealed that knockdown of
miR-503-3p or miR-503-5p significantly inhibited the proliferative
ability of keloid-derived fibroblasts (Fig. 6D and E). Cell cycle progression
and apoptosis were also investigated via flow cytometry following
miR-503-3p or miR-503-5p knockdown. Annexin V/PI double staining
showed that the rates of apoptosis were significantly higher in
fibroblasts in the inhibitor group compared with the control group
(Fig. 6F and G). Additionally,
cell cycle analysis demonstrated that more fibroblasts were
distributed in the G0/G1 phase (Fig. 6H and I) and less in the S phase
(Fig. 6H and J) following
miR-503-3p or miR-503-5p knockdown. In addition, western blotting
results showed that downregulation of miR-503-3p significantly
decreased the expression levels of fibronectin, α-SMA, collagen I
and III, indicating that extracellular matrix accumulation was
reduced (Fig. 6K and L).
Consistent with the miR-503-3p results, knockdown of miR-503-5p
also led to a significant decrease in the expression levels of
fibronectin, α-SMA, collagen I and III (Fig. 6M and N). These results indicated
that both miR-503-3p and miR-503-5p contributed to keloid
progression.
Discussion
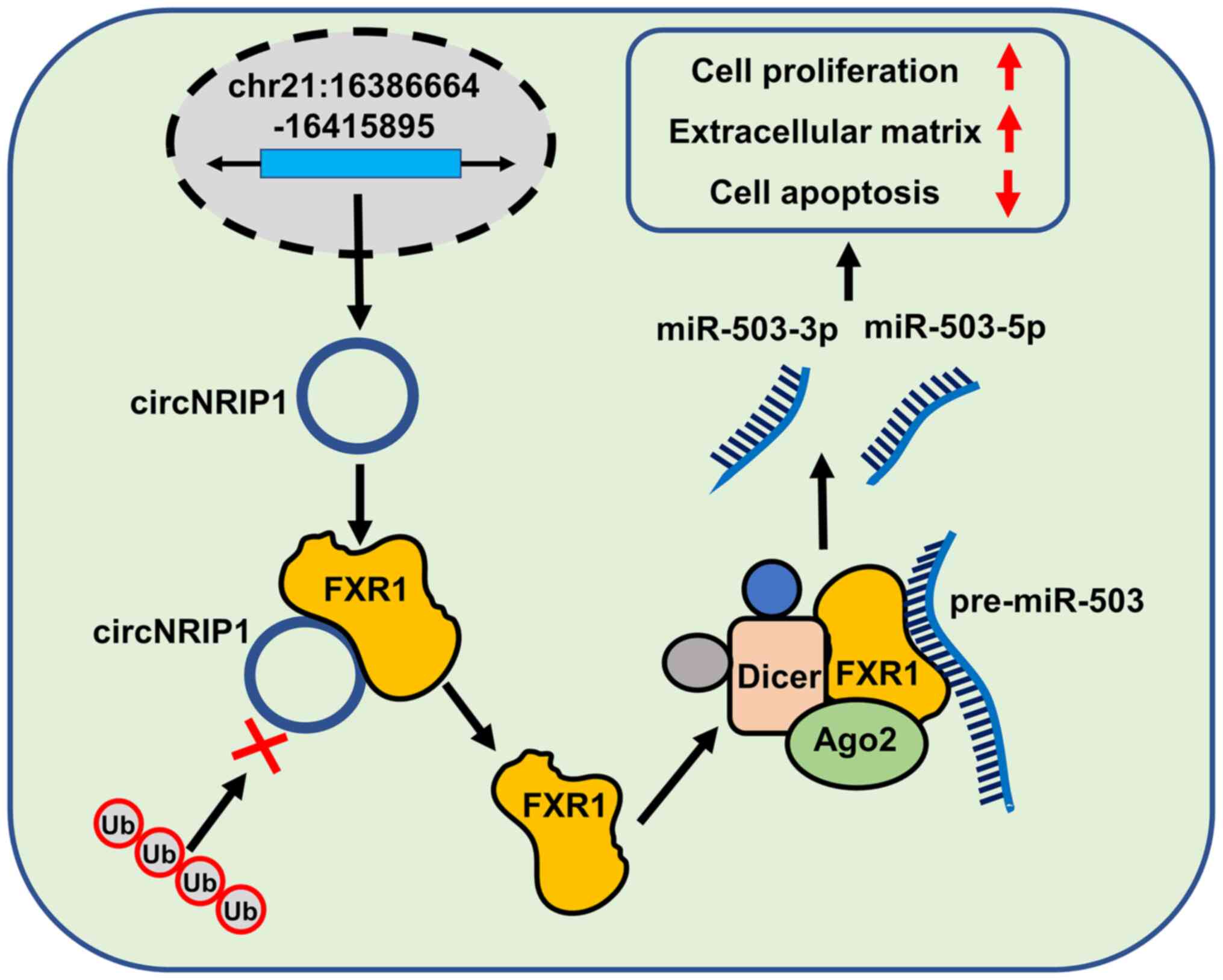
To the best of our knowledge, the present study is
the first analysis of circNRIP1-guided keloid progression.
circNRIP1 was markedly upregulated in keloid tissue and
keloid-derived fibroblasts. Loss-of-function of circNRIP1 promoted
cell apoptosis and inhibited proliferation and extracellular matrix
accumulation of keloid-derived fibroblasts. Mechanistic studies
confirmed that circNRIP1 exerted its keloid-promoting effects via
upregulation of FXR1 expression by interacting with FXR1 and
protecting it from Fbxo4-mediated ubiquitination and degradation.
Consistent with the circNRIP1 results, FXR1 deficiency exhibited
similar effects on keloid progression. FXR1 interaction was
required for the maturation of pre-miR-503, a precursor of both
miR-503-3p and miR-503-5p. Additionally, miR-503-3p and miR-503-5p
also facilitated keloid progression. These results provided
mechanistic and functional insights into keloid progression and
suggested that circNRIP1 may be a novel therapeutic target for
keloid treatment (Fig. 7).
The discovery of circRNAs altered understanding of
the complex biology of keloids (26-28). Increasing evidence has indicated
that dysregulated circRNA expression affects a multitude of
cellular processes, including cell proliferation, apoptosis and
migration (29,30). For example, Zhang et al
(28) analyzed the expression
profiles of circRNAs and found that 411 were differentially
expressed in keloid dermal fibroblasts, 206 of which were
upregulated and 205 downregulated. Additionally, overexpression of
hsa_circRNA_0008259 suppressed type I and III collagen expression.
circNRIP1, which originates from the NRIP1 gene, serves a
role in cancer progression: circNRIP1 deficiency significantly
inhibits the proliferation, migration and invasion abilities of
gastric cancer cells (15).
These observations suggested that circNRIP1 may be of
pathobiological significance in keloid progression. The present
study investigated the biological role of circNRIP1 in keloid
progression and the underlying regulatory mechanisms. To the best
of our knowledge, the present study is the first to profile the
expression of circNRIP1, which was highly expressed in keloid
tissue and keloid-derived fibroblasts. This was consistent with a
previous report showing that circNRIP1 is also highly expressed in
gastric cancer (15). Although
these findings lack clinical evidence, such as an association
between circ-NRIP1 levels and clinicopathological characteristics,
they nevertheless suggested that circNRIP1 may be involved in
keloid progression.
Keloids are pathological scars characterized by
excessive proliferation and invasive growth of dermal fibroblasts
and abnormal deposition of collagen fibers, and exhibit cancer-like
properties (31,32). For example, Jin et al
(33) found abnormal arrangement
and hyperplasia of fibers in keloid tissue, along with increased
ColI levels. The mechanisms underlying keloid progression are
poorly understood and effective prevention and treatment are
lacking. To the best of our knowledge, no study has investigated
the biological role of circNRIP1 either in keloids or in other
types of pathological scar. Loss-of-function analysis here
demonstrated that knockdown of circNRIP1 impaired cell
proliferation and accumulation of extracellular matrix, and induced
cell apoptosis in keloid-derived fibroblasts. To the best of our
knowledge, the present study is the first report on the role of
circNRIP1 in keloid progression, and the results indicate that
silencing circNRIP1 may represent a promising therapeutic strategy
for the treatment of keloids. However, in vivo experiments
are required to confirm the function of circNRIP1 in keloid
progression.
Increasing evidence has shown that RNA-binding
proteins act in concert with circRNAs in disease progression
(34,35). For example, circADD3 has been
shown to facilitate enhancer of zeste 2 polycomb repressive complex
2 subunit degradation via CDK1-mediated ubiquitination (36). The present study predicted the
downstream targets of circNRIP1 using RNAInter and found that FXR1
interacted with circNRIP1. Several studies have demonstrated that
upregulation of FXR1 promotes cell proliferation, migration and
invasion (37), which suggested
that FXR1 may play a crucial role in keloid progression. Validation
experiments confirmed that circNRIP1 maintained FXR1 stability via
direct interaction. Accumulating evidence has shown that circRNAs
directly bind to RNA-binding proteins and regulate their expression
or activity at the post-translational level (38,39). For example, Qie et al
(40) reported that the E3
ubiquitin ligase Fbxo4 interacts with, and promotes the
ubiquitination and degradation of, FXR1. Consequently, it was
speculated that circNRIP1 may inhibit FXR1 ubiquitination by
blocking the interaction between FXR1 and Fbxo4 in keloid tissue.
RNA pulldown and RIP assays verified that circNRIP1 blocked the
Fbxo4-mediated ubiquitination of FXR1. Several studies have shown
that FXR1 facilitates the malignant behavior of cancer cells
(41-43). Inhibition of FXR1 selectively
blocks proliferation in human cancer cells (42). Here, in vitro
loss-of-function experiments demonstrated that FXR1 deficiency
markedly decreased cell proliferation and extracellular matrix
accumulation but induced cell apoptosis in keloid-derived
fibroblasts, further confirming the aforementioned hypothesis. To
the best of our knowledge, the present study is the first to
demonstrate that circNRIP1 promotes keloid progression via
stabilizing FXR1.
Although FXR1 is not the primary regulatory molecule
in pre-miRNA processing, studies have shown that FXR1 is involved
in the efficient processing of pre-miRNA and forms a complex with
pre-miRNA, Dicer and argonaute RISC catalytic component 2 (44,45). Additionally, Majumder and
Palanisamy (43) proposed that
FXR1 controls the expression of a subset of mature miRNAs,
including miR-301a-3p, which is highly expressed in oral cancer
cells. In the present study, prediction results obtained from the
starBase database indicated that FXR1 interacted with pre-miR-503.
Zhong et al (46)
performed miRNA expression profile analysis and found that both
miR-503-3p and miR-503-5p are highly expressed in keloids.
Therefore, the present study aimed to characterize the mechanisms
underlying FXR1-mediated regulation of pre-miR-503 maturation.
Expression profiling showed that the levels of both miR-503-3p and
miR-503-5p were significantly upregulated in keloid tissue.
However, pre-miR-503 showed the opposite expression profile. These
results were consistent with those of Zhong et al (46) for microarray data. Moreover, the
present findings provided evidence that FXR1 directly interacts
with pre-miR-503. FXR1 overexpression led to an increase in the
levels of mature miR-503 and a decrease in those of pre-miR-503
in vitro. These data highlight a novel molecular mechanism
for FXR1 in keloid progression. At present, the roles of miR-503-3p
and miR-503-5p in regulating cell proliferation, apoptosis and
extracellular matrix accumulation remain contradictory (47,48). Here, knockdown of miR-503-3p or
miR-503-5p impaired cell proliferation and excessive extracellular
matrix accumulation and promoted cell apoptosis. Consistent with
these findings, inhibition of miR-503-5p is reported to suppress
chondrocyte proliferation (49),
while miR-503-3p enhances the proliferation of breast cancer cells
(47). These findings further
confirmed the hypothesis that circNRIP1 facilitates keloid
progression via FXR1-mediated pre-miR-503 maturation. However,
contradictory results have been reported for the roles of
miR-503-3p and miR-503-5p in cell proliferation and apoptosis
(50,51). For example, Sun et al
(52) found that miR-503-3p
suppresses viability and promotes apoptosis of lung cancer cells.
Fu et al (50) showed
that increasing miR-503-5p expression markedly inhibited the
proliferation and colony-forming ability of cervical cancer cells.
These discrepancies may be due to different experimental
conditions, such as the use of different cell types. Further
studies are needed to confirm the role of miR-503-3p and miR-503-5p
in keloids. Additionally, the combination of miR-503-3p and
miR-503-5p, as well as their downstream targets need to be further
studied in the future.
In conclusion, the present study is the first to
show that circNRIP1 is highly expressed in keloid tissue, which
triggers cell proliferation and accumulation of extracellular
matrix, but inhibits cell apoptosis. Mechanistically, circNRIP1
contributes to pre-miR-503 maturation by antagonizing
Fbxo4-mediated FXR1 ubiquitination and degradation. These results
provide a novel theoretical and experimental basis for keloid
pathogenesis and identify potential therapeutic targets for the
treatment of keloids.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
TW and HY conceptualized the study. BW designed the
methodology and collected the data. HZ operated the software and
visualized the data. BW validated the data and drafted the
manuscript. HZ and TW analyzed the data. HY performed the
experiments and supervised the study. TW acquired resources and
funding, reviewed and edited the manuscript and was responsible for
project administration. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Clinical
Research Ethics Committee of Zaozhuang Municipal Hospital (approval
no. 2018ZMHE015). Additionally, written informed consent was
obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
1
|
Liu T, Ma X, Ouyang T, Chen H, Xiao Y,
Huang Y, Liu J and Xu M: Efficacy of 5-aminolevulinic acid-based
photodynamic therapy against keloid compromised by downregulation
of SIRT1-SIRT3-SOD2-mROS dependent autophagy pathway. Redox Biol.
20:195–203. 2019. View Article : Google Scholar
|
|
2
|
Shi K, Qiu X, Zheng W, Yan D and Peng W:
MiR-203 regulates keloid fibroblast proliferation, invasion, and
extracellular matrix expression by targeting EGR1 and FGF2. Biomed
Pharmacother. 108:1282–1288. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Har-Shai Y, Brown W, Labbé D, Dompmartin
A, Goldine I, Gil T, Mettanes I and Pallua N: Intralesional
cryosurgery for the treatment of hypertrophic scars and keloids
following aesthetic surgery: The results of a prospective
observational study. Int J Low Extrem Wounds. 7:169–175. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Berman B, Maderal A and Raphael B: Keloids
and hypertrophic scars: Pathophysiology, classification, and
treatment. Dermatol Surg. 43(Suppl 1): S3–S18. 2017. View Article : Google Scholar
|
|
5
|
Yang JY and Yang SY: Are auricular keloids
and persistent hypertrophic scars resectable? The role of intrascar
excision. Ann Plast Surg. 69:637–642. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shin JY, Yun SK, Roh SG, Lee NH and Yang
KM: Efficacy of 2 representative topical agents to prevent keloid
recurrence after surgical excision. J Oral Maxillofac Surg.
75:401.e1–401.e6. 2017. View Article : Google Scholar
|
|
7
|
Nong Q, Li S, Wu Y and Liu D: LncRNA
COL1A2-AS1 inhibits the scar fibroblasts proliferation via
regulating miR-21/Smad7 pathway. Biochem Biophys Res Commun.
495:319–324. 2018. View Article : Google Scholar
|
|
8
|
Yan L, Wang LZ, Xiao R, Cao R, Pan B, Lv
XY, Jiao H, Zhuang Q, Sun XJ and Liu YB: Inhibition of
microRNA-21-5p reduces keloid fibroblast autophagy and migration by
targeting PTEN after electron beam irradiation. Lab Invest.
100:387–399. 2020. View Article : Google Scholar
|
|
9
|
Zhang G, Guan Q, Chen G, Qian F and Liang
J: DNA methylation of the CDC2L1 gene promoter region decreases the
expression of the CDK11p58 protein and reduces apoptosis in keloid
fibroblasts. Arch Dermatol Res. 310:107–115. 2018. View Article : Google Scholar
|
|
10
|
Kristensen LS, Andersen MS, Stagsted LVW,
Ebbesen KK, Hansen TB and Kjems J: The biogenesis, biology and
characterization of circular RNAs. Nat Rev Genet. 20:675–691. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu J, Xu QG, Wang ZG, Yang Y, Zhang L, Ma
JZ, Sun SH, Yang F and Zhou WP: Circular RNA cSMARCA5 inhibits
growth and metastasis in hepatocellular carcinoma. J Hepatol.
68:1214–1227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yao J, Zhang C, Chen Y and Gao S:
Downregulation of circular RNA circ-LDLRAD3 suppresses pancreatic
cancer progression through miR-137-3p/PTN axis. Life Sci.
239:1168712019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Peng Y, Song X, Zheng Y, Cheng H and Lai
W: circCOL3A1- 859267 regulates type I collagen expression by
sponging miR-29c in human dermal fibroblasts. Eur J Dermatol.
28:613–620. 2018.PubMed/NCBI
|
|
14
|
Shi J, Yao S, Chen P, Yang Y, Qian M, Han
Y, Wang N, Zhao Y, He Y, Lyu L and Lu D: The integrative regulatory
network of circRNA and microRNA in keloid scarring. Mol Biol Rep.
47:201–209. 2020. View Article : Google Scholar
|
|
15
|
Zhang X, Wang S, Wang H, Cao J, Huang X,
Chen Z, Xu P, Sun G, Xu J, Lv J and Xu Z: Circular RNA circNRIP1
acts as a microRNA-149-5p sponge to promote gastric cancer
progression via the AKT1/mTOR pathway. Mol Cancer. 18:202019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li M, Cai J, Han X and Ren Y:
Downregulation of circNRIP1 suppresses the paclitaxel resistance of
ovarian cancer via regulating the miR-211-5p/HOXC8 axis. Cancer
Manag Res. 12:9159–9171. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu G, Li M, Wu J, Qin C, Tao Y and He H:
Circular RNA circ- NRIP1 sponges microRNA-138-5p to maintain
hypoxia-induced resistance to 5-fluorouracil through
HIF-1α-dependent glucose metabolism in gastric carcinoma. Cancer
Manag Res. 12:2789–2802. 2020. View Article : Google Scholar :
|
|
18
|
Xie R, Tang J, Zhu X and Jiang H:
Silencing of hsa_circ_0004771 inhibits proliferation and induces
apoptosis in breast cancer through activation of miR-653 by
targeting ZEB2 signaling pathway. Biosci Rep. May 17–2019.Epub
ahead of print. View Article : Google Scholar
|
|
19
|
Wang L, Tong X, Zhou Z, Wang S, Lei Z,
Zhang T, Liu Z, Zeng Y, Li C, Zhao J, et al: Circular RNA
hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced
epithelial-mesenchymal transition and metastasis by controlling
TIF1γ in non-small cell lung cancer. Mol Cancer. 17:1402018.
View Article : Google Scholar
|
|
20
|
Zheng X, Chen L, Zhou Y, Wang Q, Zheng Z,
Xu B, Wu C, Zhou Q, Hu W, Wu C and Jiang J: A novel protein encoded
by a circular RNA circPPP1R12A promotes tumor pathogenesis and
metastasis of colon cancer via Hippo-YAP signaling. Mol Cancer.
18:472019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang X, He M, Huang S, Lin R, Zhan M,
Yang D, Shen H, Xu S, Cheng W, Yu J, et al: Circular RNA circERBB2
promotes gallbladder cancer progression by regulating
PA2G4-dependent rDNA transcription. Mol Cancer. 18:1662019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar :
|
|
23
|
Liang WC, Wong CW, Liang PP, Shi M, Cao Y,
Rao ST, Tsui SK, Waye MM, Zhang Q, Fu WM and Zhang JF: Translation
of the circular RNA circβ-catenin promotes liver cancer cell growth
through activation of the Wnt pathway. Genome Biol. 20:842019.
View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Hartmann P, Zhou Z, Natarelli L, Wei Y,
Nazari-Jahantigh M, Zhu M, Grommes J, Steffens S, Weber C and
Schober A: Endothelial Dicer promotes atherosclerosis and vascular
inflammation by miRNA-103-mediated suppression of KLF4. Nat Commun.
7:105212016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Patop IL, Wüst S and Kadener S: Past,
present, and future of circRNAs. EMBO J. 38:e1008362019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu H, Wu S, Zhu Y, Ye M, Shen J, Liu Y,
Zhang Y and Bu S: Hsa_circRNA_0054633 is highly expressed in
gestational diabetes mellitus and closely related to glycosylation
index. Clin Epigenetics. 11:222019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Z, Yu K, Liu O, Xiong Y, Yang X,
Wang S, Zhang S, Feng Y and Peng Y: Expression profile and
bioinformatics analyses of circular RNAs in keloid and normal
dermal fibroblasts. Exp Cell Res. 388:1117992020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu J, Yang M, Zhou B, Luo J, Zhang Z,
Zhang W and Yan Z: CircRNA-104718 acts as competing endogenous RNA
and promotes hepatocellular carcinoma progression through
microRNA-218-5p/TXNDC5 signaling pathway. Clin Sci (Lond).
133:1487–1503. 2019. View Article : Google Scholar
|
|
30
|
Chen X, Ouyang Z, Shen Y, Liu B, Zhang Q,
Wan L, Yin Z, Zhu W, Li S and Peng D: CircRNA_28313/miR-195a/CSF1
axis modulates osteoclast differentiation to affect OVX-induced
bone absorption in mice. RNA Biol. 16:1249–1262. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Ren J, Su L, Cheng S, Zhou J, Ye X,
Dong Y, Sun S, Qi F, Liu Z, et al: Human adipose tissue-derived
stem cells inhibit the activity of keloid fibroblasts and fibrosis
in a keloid model by paracrine signaling. Burns. 44:370–385. 2018.
View Article : Google Scholar
|
|
32
|
Andrews JP, Marttala J, Macarak E,
Rosenbloom J and Uitto J: Keloids: The paradigm of skin
fibrosis-pathomechanisms and treatment. Matrix Biol. 51:37–46.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin J, Zhai HF, Jia ZH and Luo XH: Long
non-coding RNA HOXA11-AS induces type I collagen synthesis to
stimulate keloid formation via sponging miR-124-3p and activation
of Smad5 signaling. Am J Physiol Cell Physiol. 317:C1001–C1010.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dong W, Dai ZH, Liu FC, Guo XG, Ge CM,
Ding J, Liu H and Yang F: The RNA-binding protein RBM3 promotes
cell proliferation in hepatocellular carcinoma by regulating
circular RNA SCD-circRNA 2 production. EBioMedicine. 45:155–167.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang L, Long H, Zheng Q, Bo X, Xiao X and
Li B: Circular RNA circRHOT1 promotes hepatocellular carcinoma
progression by initiation of NR2F6 expression. Mol Cancer.
18:1192019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun S, Wang W, Luo X, Li Y, Liu B and Li
X, Zhang B, Han S and Li X: Circular RNA circ-ADD3 inhibits
hepatocellular carcinoma metastasis through facilitating EZH2
degradation via CDK1-mediated ubiquitination. Am J Cancer Res.
9:1695–1707. 2019.PubMed/NCBI
|
|
37
|
Cao S, Zheng J, Liu X, Liu Y, Ruan X, Ma
J, Liu L, Wang D, Yang C, Cai H, et al: FXR1 promotes the malignant
biological behavior of glioma cells via stabilizing MIR17HG. J Exp
Clin Cancer Res. 38:372019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang S, Li X, Zheng H, Si X, Li B, Wei G,
Li C, Chen Y, Chen Y, Liao W, et al: Loss of
super-enhancer-regulated circRNA Nfix induces cardiac regeneration
after myocardial infarction in adult mice. Circulation.
139:2857–2876. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zang J, Lu D and Xu A: The interaction of
circRNAs and RNA binding proteins: An important part of circRNA
maintenance and function. J Neurosci Res. 98:87–97. 2020.
View Article : Google Scholar
|
|
40
|
Qie S, Majumder M, Mackiewicz K, Howley
BV, Peterson YK, Howe PH, Palanisamy V and Diehl JA: Fbxo4-mediated
degradation of Fxr1 suppresses tumorigenesis in head and neck
squamous cell carcinoma. Nat Commun. 8:15342017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fan Y, Yue J, Xiao M, Han-Zhang H, Wang
YV, Ma C, Deng Z, Li Y, Yu Y, Wang X, et al: FXR1 regulates
transcription and is required for growth of human cancer cells with
TP53/FXR2 homozygous deletion. Elife. 6:e261292017. View Article : Google Scholar :
|
|
42
|
Cao H, Gao R, Yu C, Chen L and Feng Y: The
RNA-binding protein FXR1 modulates prostate cancer progression by
regulating FBXO4. Funct Integr Genomics. 19:487–496. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Majumder M and Palanisamy V: RNA binding
protein FXR1-miR301a-3p axis contributes to p21WAF1 degradation in
oral cancer. PLoS Genet. 16:e10085802020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu XL, Zong R, Li Z, Biswas MH, Fang Z,
Nelson DL and Gao FB: FXR1P but not FMRP regulates the levels of
mammalian brain-specific microRNA-9 and microRNA-124. J Neurosci.
31:13705–13709. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vasudevan S and Steitz JA:
AU-rich-element-mediated upregulation of translation by FXR1 and
Argonaute 2. Cell. 128:1105–1118. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhong L, Bian L, Lyu J, Jin H, Liu Z, Lyu
L and Lu D: Identification and integrated analysis of microRNA
expression profiles in keloid. J Cosmet Dermatol. 17:917–924. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhao Z, Fan X, Jiang L, Xu Z, Xue L, Zhan
Q and Song Y: miR-503-3p promotes epithelial-mesenchymal transition
in breast cancer by directly targeting SMAD2 and E-cadherin. J
Genet Genomics. 44:75–84. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jiang SP and Li ZR: MiR-503-5p regulates
cell epithelial-to-mesenchymal transition, metastasis and prognosis
of hepatocellular carcinoma through inhibiting WEE1. Eur Rev Med
Pharmacol Sci. 23:2028–2037. 2019.PubMed/NCBI
|
|
49
|
Jee YH, Wang J, Yue S, Jennings M, Clokie
SJ, Nilsson O, Lui JC and Baron J: mir-374-5p, mir-379-5p, and
mir-503-5p regulate proliferation and hypertrophic differentiation
of growth plate chondrocytes in male rats. Endocrinology.
159:1469–1478. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fu Y, Meng Y, Gu X, Tian S, Hou X and Ji
M: miR-503 expression is downregulated in cervical cancer and
suppresses tumor growth by targeting AKT2. J Cell Biochem. Jan
29–2019.Epub ahead of print. View Article : Google Scholar
|
|
51
|
Cai X, Nie J, Chen L and Yu F:
Circ_0000267 promotes gastric cancer progression via sponging
MiR-503-5p and regulating HMGA2 expression. Mol Genet Genomic Med.
8:e10932020. View Article : Google Scholar
|
|
52
|
Sun Y, Li L, Xing S, Pan Y, Shi Y, Zhang L
and Shen Q: miR-503-3p induces apoptosis of lung cancer cells by
regulating p21 and CDK4 expression. Cancer Biomark. 20:597–608.
2017. View Article : Google Scholar : PubMed/NCBI
|