Introduction
Esophageal carcinoma is one of the most aggressive
gastrointestinal malignancies and accounts for the sixth leading
cause of cancer-related mortality worldwide; there were 572,000
estimated new cases and 509,000 estimated deaths in 2018 worldwide
(1). Esophageal squamous cell
carcinoma (ESCC) is the major histological type, which accounts for
approximately 90% of all esophageal carcinoma cases (2). Due to the difficulty in the early
diagnosis of ESCC, the majority of patients with ESCC are not
suitable for radical surgical resection (3). The prognoses of patients with ESCC
are extremely poor (4). Further
revealing the critical molecular mechanisms underlying the
initiation and development of ESCC would promote the findings of
novel therapeutic targets and prognostic biomarkers for ESCC.
Genome and transcriptome sequencings have found that
the majority of the human genome is transcribed; however, only
approximately 2% of the human genome encodes for proteins (5). Non-coding RNAs comprise >90% of
the human transcriptome (6).
Long non-coding RNAs (lncRNAs) and microRNAs (miRNAs or miRs) are
two classes of important regulatory non-coding RNAs, which have
been revealed to play critical roles in various pathophysiological
statuses (6,7). lncRNAs are a class of RNAs with
>200 nucleotides in length and limited protein-coding potential
(8). Accumulating evidence has
indicated that a number of lncRNAs are dysregulated and play
important roles in various diseases, including cancers (9-17). lncRNAs have been revealed to
regulate various biological functions of cancer cells, such as
growth, cycle, apoptosis, migration, invasion, senescence, drug
resistance and others (18-25). The mechanisms of action of
lncRNAs are diverse. lncRNAs can directly bind proteins, mRNAs,
miRNAs, or DNAs, and further modulate the location, expression and
function of the interacted partners (26-29).
miRNAs are another class of regulatory non-coding
RNAs with 19-25 nucleotides in length (30). Consistent with lncRNAs, a number
of miRNAs have been revealed to be dysregulated and to play
important roles in various diseases (31-35). miRNAs negatively regulate the
expression of their targets by binding to the 3′-untranslated
regions (3′UTRs) of target mRNAs, and further causing translational
inhibition and/or target mRNAs degradation (36-39).
lncRNA LincIN (long intergenic non-coding RNA
between ITGB1 and NRP1) was recently identified to be overexpressed
in human breast tumors and to be associated with a poor prognosis
of patients with breast cancer (40). Furthermore, LincIN was revealed
to promote breast tumor cell migration and invasion by binding
nuclear factor 90 (NF90) (40).
However, the expression, clinical significance, biological role and
mechanisms of action of LincIN in ESCC remain unknown.
In the present study, LincIN expression was measured
in ESCC by RT-qPCR, and the roles of LincIN in ESCC were detected
using cell growth, migration and invasion assays. In addition, and
the mechanisms of action of LincIN were investigated by RNA
immunoprecipitation assay, RT-qPCR, dual-luciferase reporter assay
and western blot analysis.
Materials and methods
Clinical specimens
A total of 56 pairs of ESCC tissues and adjacent
noncancerous tissue specimens were acquired from patients with ESCC
who received surgical resection at Nanfang Hospital, Southern
Medical University (Guangzhou, China) between February, 2016 and
December, 2017. The clinical specimens were immediately frozen in
liquid nitrogen and stored at −80°C after surgery. All clinical
specimens were diagnosed by histopathological detection. Written
informed consent was acquired from all participants. The use of
clinical specimens was reviewed and approved by the Nanfang
Hospital Institutional Review Board (Guangzhou, China).
Cells and cell culture
The human immortalized normal esophageal epithelial
cell line, Het-1A, was acquired from the American Type Culture
Collection (ATCC, cat. no. CRL-2692). The ESCC cell lines, TE-1
(cat. no. TCHu 89), KYSE150 (cat. no. TCHu 236) and Eca-109 (cat.
no. TCHu 69), were acquired from the Institute of Biochemistry and
Cell Biology of the Chinese Academy of Sciences (Shanghai, China).
Het-1A cells were maintained in BEBM (Lonza Group, Ltd.). ESCC cell
lines were maintained in RPMI-1640 medium (Invitrogen; Thermo
Fisher Scientific, Inc.). All cells were cultured in medium
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) at a humidified incubator with 5% CO2
at 37°C.
Plasmid construction
LincIN full-length sequences were PCR-amplified
using the Phusion Flash High-Fidelity PCR Master Mix (Thermo Fisher
Scientific, Inc.) and the primers, 5′-CCC AAG CTT ACT CTG TAG TCA
CCC AGG CT-3′ (sense) and 5′-GCT CTA GAT TCT GTA AAT TAA GTT TAA
TGC TG-3′ (antisense). Subsequently, the PCR products were
sub-cloned into the HindIII and XbaI sites of the
pcDNA™3.1(+) plasmid (Invitrogen; Thermo Fisher Scientific, Inc.).
In total, 2 independent oligonucleotides inhibiting LincIN
expression were synthesized and inserted into the SuperSilencing
shRNA (short hairpin RNA) expression plasmid pGPU6/Neo (GenePharma,
Inc.), named LincIN-shRNA1 and LincIN-shRNA2. The targeted
sequences of the LincIN shRNAs were as follows: 5′-GAC ATT ATG CAA
GGA GAT GGC A-3′ (LincIN-shRNA1) and 5′-CAG TTG GTC ACT CTA CTC
AGT-3′ (LincIN-shRNA2) (40).
The HOXB13 3′UTR containing miR-7 target sites were PCR-amplified
using the Phusion Flash High-Fidelity PCR Master Mix (Thermo Fisher
Scientific, Inc.) and the primers, 5′-CGA GCT CCC CTT CCA TTA CAC
CTC TCA C-3′ (sense) and 5′-GCT CTA GAT CCT CCT CCT CGT CCT CTT-3′
(antisense). The PCR products were then sub-cloned into the
SacI and XbaI sites of pmirGLO plasmid (Promega
Corporation).
Transfection of plasmids and miRNAs
miR-7 mimics and miRNAs negative control (NC) were
purchased from GenePharma, Inc. HOXB13 specific shRNA
(HOXB13-shRNA) and scrambled shRNA which was used as a negative
control were purchased from GeneCopoeia, Inc. The transfections of
plasmids (2.5 µg) and miRNAs (10 pmol) were carried out
using Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the protocol. At 48 h following transfection,
subsequent experimentation was performed.
Construction of stable cell lines
The LincIN overexpression plasmid and control
pcDNA3.1 plasmid were transfected into TE-1 cells. TE-1 cells were
established from a well-differentiated human squamous cell
carcinoma of the esophagus and had a male karyotype (41). TE-1 has a mutation at codon 272
of p53, leading to p53 activity impairment (42). At 48 h following transfection,
the cells were selected with neomycin for 4 weeks to obtain LincIN
stably overexpressed and control TE-1 cells. LincIN-specific shRNAs
(2.5 µg) and control shRNA (2.5 µg) were transfected
into the Eca-109 cells. The Eca-109 cells were derived from a human
oesophageal carcinoma in 1973 (43). Eca-109 harbors wild-type p53
(44). At 72 h following
transfection, the cells were selected with neomycin for 4 weeks to
obtain LincIN stably-depleted and control Eca-109 cells.
2×106 transducing units of miR-7 overexpression
lentivirus (GeneCopoeia, Inc.) was infected into LincIN stably
overexpressing TE-1 cells. At 96 h following infection, the cells
were selected with neomycin and puromycin for 4 weeks to obtain
LincIN and miR-7 simultaneously stably overexpressed TE-1 cells.
HOXB13-specific shRNAs (2.5 µg) were transfected into LincIN
stably overexpressing TE-1 cells. At 72 h following transfection,
the cells were selected with neomycin and puromycin for 4 weeks to
obtain HOXB13 stably depleted and simultaneously LincIN stably
overexpressing TE-1 cells.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's instructions. The RNA was then used to carry out
reverse transcription using the M-MLV Reverse Transcriptase
(Invitrogen; Thermo Fisher Scientific, Inc.). The first-stand cDNA
generated from reverse transcription was used to carry out
quantitative polymerase chain reaction (qPCR) with SYBR Premix Ex
Taq™ II (Takara Biotechnology Co., Ltd.) on an ABI7500 System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) following the
standard SYBR-Green PCR protocols. The qPCR cycling conditions were
as follows: 95°C for 30 sec, then 40 cycles at the conditions of
95°C for 5 sec, 60°C for 20 sec, and 72°C for 20 sec. The RT-qPCR
primers were as follows: LincIN, 5′-AAG AGG GGA GTG CGG AAC A-3′
(forward) and 5′-TCA CCA GGG ACA CGA GAT G-3′ (reverse); HOXB13,
5′-TGA CTC CCT GTT GCC TGT G-3′ (forward) and 5′-GAA CTT GTT AGC
CGC ATA CTC-3′ (reverse); β-actin, 5′-GGG AAA TCG TGC GTG ACA TTA
AG-3′ (forward) and 5′-TGT GTT GGC GTA CAG GTC TTT G-3′ (reverse).
For the quantification of the expression of miR-7 and pri-miR-7,
RT-qPCR was carried out using the TaqMan microRNA assays (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and TaqMan pri-miRNA
assays (Applied Biosystems; Thermo Fisher Scientific, Inc.)
respectively following the manufacturer's protocols. β-actin was
used as a reference gene for the quantification of LincIN, HOXB13
and pri-miR-7. U6 was used as a reference gene for the
quantification of miR-7. The quantification of RNA expression was
calculated using the 2−ΔΔCq method (45).
Western blot analysis
Total proteins were extracted from the TE-1 and
Eca-109 cells using RIPA buffer (Beyotime Institute of
Biotechnology) supplemented with protease inhibitors (Beyotime
Institute of Biotechnology). Protein concentrations were determined
by bicinchoninic acid (BCA) assay with the BCA Protein Assay kit
(Beyotime Institute of Biotechnology) following the manufacturer's
protocol. Equal amounts of proteins (10 µg) were separated
by 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (EMD Millipore).
After blocking with 5% non-fat milk in Tris-buffered saline with
0.1% Tween-20 (TBST) at room temperature for 2 h, the membranes
were incubated with primary antibodies against HOXB13 (1:1,000,
cat. no. 90944, Cell Signaling Technology, Inc.) or β-actin
(1:10,000, cat T0022, Affinity Biosciences) overnight at 4°C. After
washing with TBST buffer, the membranes were incubated with IRDye
680RD goat anti-mouse IgG secondary antibody (1:10,000, cat. no.
925-68070, LI-COR Biosciences) or IRDye 800CW goat anti-rabbit IgG
secondary antibody (1:10,000, cat. no. 926-32211, LI-COR
Biosciences) at room temperature for 1 h, followed by detection on
an Odyssey infrared scanner (LI-COR Biosciences). β-actin was used
as a loading control.
Cell growth assays
Cell growth was assessed using a Cell Counting kit-8
(CCK-8) and 5-ethynyl-2-deoxyuridine (EdU) incorporation assays
(27). For CCK-8 assay, the TE-1
and Eca-109 cells were seeded at 2×103 cells/well into
96-well plates. After being incubated for 24, 48, or 72 at 37°C, 10
µl CCK-8 solution (Dojindo Laboratories, Inc.) were added to
each well. The optical density values at 450 nm were measured with
an enzyme-linked immune detector to indicate cell viability. EdU
incorporation assay was performed using the Cell-Light™ EdU
Apollo® 643 In Vitro Imaging kit (Guangzhou RiboBio Co.,
Ltd.) following the manufacturer's instructions. The results were
detected using a Zeiss photomicroscope (Carl Zeiss AG) and counting
was performed based on at least 10 random visual fields.
Transwell cell migration and invasion
assays
Cell migration and invasion were assessed using
Transwell assays (46). The TE-1
and Eca-109 cells suspended in serum-free medium were plated in the
upper chamber of Transwell inserts (8 µm-pore size, EMD
Millipore) with Matrigel (for invasion assay) or without Matrigel
(for migration assay). Medium containing 20% fetal bovine serum was
plated into the lower chamber. Following incubation for 48 h at
37°C, cells remaining on the upper chamber were wiped away using a
cotton swab. Cells migrating into the lower surfaces of the
Transwell inserts were fixed in methanol and stained with 0.5%
crystal violet (Beyotime Institute of Biotechnology) at room
temperature for 30 min. The results were detected using a Zeiss
photomicroscope and counted based on at least 10 random visual
fields.
RNA immunoprecipitation (RIP) assay
RIP assays were carried out in the TE-1 and Eca-109
cells using the EZ-Magna RIP™ RNA Binding Protein
Immunoprecipitation kit (EMD Millipore) and primary antibody
against NF90 (5 µg, ab131004, Abcam). The enriched RNA
(LincIN and pri-miR-7) was quantified by RT-qPCR as described
above.
Dual-luciferase reporter assay
Luciferase reporter plasmid pmirGLO containing
HOXB13 3′UTR (50 ng) was co-transfected with LincIN overexpression
plasmid (50 ng) and miR-7 mimics (1 pmol) into TE-1 cells.
Luciferase reporter plasmid pmirGLO containing HOXB13 3′UTR (50 ng)
was co-transfected with LincIN specific shRNAs (50 ng) into Eca-109
cells. Following incubation for 48 h at 37°C, the Firefly
luciferase and Renilla luciferase activities were measured
using the Dual-Luciferase Reporter Assay System (Promega
Corporation).
Statistical analysis
GraphPad Prism software (version 5.0) was employed
to carry out statistical analyses. For comparisons, the Wilcoxon
signed-rank test, Pearson's Chi-squared test, Log-rank test,
one-way ANOVA followed by Dunnett's multiple comparison test, the
Student's t-test, and Pearson's correlation analysis were performed
as indicated. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
LincIN expression is increased in ESCC
and is associated with a poor prognosis of patients with ESCC
To confirm the expression pattern of LincIN in ESCC,
the expression of LincIN was examined in 56 pairs of ESCC tissues
and adjacent non-cancerous tissues by RT-qPCR. The results revealed
that the expression of LincIN was significantly increased in ESCC
tissues compared with adjacent non-cancerous tissues (Fig. 1A). The analyses of the
clinicopathological characteristics of these 56 ESCC cases revealed
that the increased expression of LincIN was positively associated
with an advanced tumor invasion depth, lymph node metastasis and
TNM stage (Table I). Patients
with ESCC with a higher expression of LincIN had a shorter survival
time than those with a lower expression of LincIN (Fig. 1B). In addition, the expression of
LincIN was significantly increased in the ESCC cell lines, TE-1,
KYSE150 and Eca-109, compared with the normal esophageal epithelial
cell line, Het-1A (Fig. 1C).
Collectively, these results suggested that LincIN exprewssion was
increased in ESCC tissues and cell lines, and was associated with
an advanced clinical stage and a poor prognosis of patients with
ESCC.
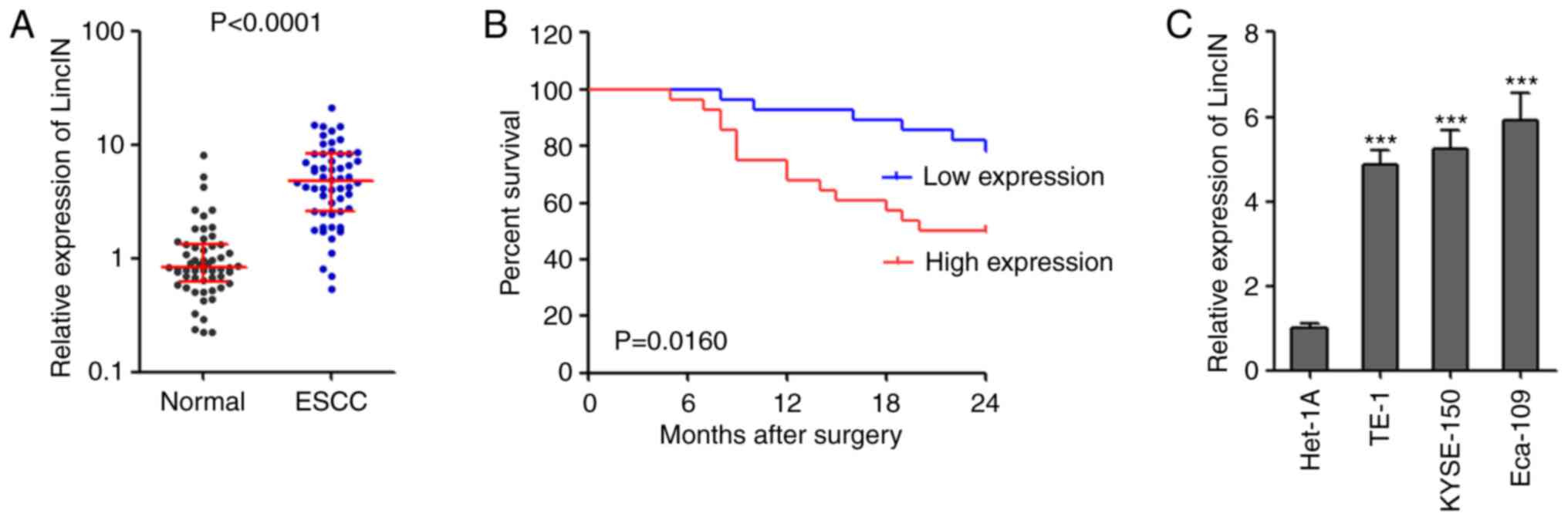 | Figure 1LincIN expression is increased in
ESCC and is associated with a poor prognosis of patients with ESCC.
(A) The expression of LincIN in 56 pairs of ESCC tissues and
adjacent noncancerous tissues was detected by RT-qPCR. P<0.0001,
determined by Wilcoxon signed-rank test. (B) Kaplan-Meier survival
analysis of the association between LincIN expression level and the
overall survival of 56 patients with ESCC. P=0.0160, determined by
the log-rank test. (C) The expression of LincIN in immortalized
normal esophageal epithelial cell line, Het-1A, and the ESCC cell
lines, TE-1, KYSE150 and Eca-109, was detected by RT-qPCR. Results
are shown as the means ± SD from 3 separate experiments.
***P<0.001, vs. control Het-1A cells, determined by
one-way ANOVA followed by Dunnett's multiple comparison test vs.
Het-1A group. ESCC, esophageal squamous cell carcinoma. |
 | Table IAssociation between LincIN expression
and the clinicopathological characteristics of patients with ESCC
(n=56). |
Table I
Association between LincIN expression
and the clinicopathological characteristics of patients with ESCC
(n=56).
|
Characteristics | Total | LincIN expression
| P-value |
|---|
| Low | High |
|---|
| Sex | | | | 1.000 |
| Male | 30 | 15 | 15 | |
| Female | 26 | 13 | 13 | |
| Age (years) | | | | 0.788 |
| ≤60 | 25 | 13 | 12 | |
| >60 | 31 | 15 | 16 | |
|
Differentiation | | | | 0.158 |
| Well or
moderate | 37 | 21 | 16 | |
| Poor | 19 | 7 | 12 | |
| Tumor invasion
depth (T) | | | | 0.032 |
| T1/T2 | 30 | 19 | 11 | |
| T3/T4 | 26 | 9 | 17 | |
| Lymph node
metastasis (N) | | | | 0.029 |
| N0 | 34 | 21 | 13 | |
| N1-N3 | 22 | 7 | 15 | |
| TNM stage | | | | 0.043 |
| I | 26 | 17 | 9 | |
| II | 17 | 8 | 9 | |
| III | 13 | 3 | 10 | |
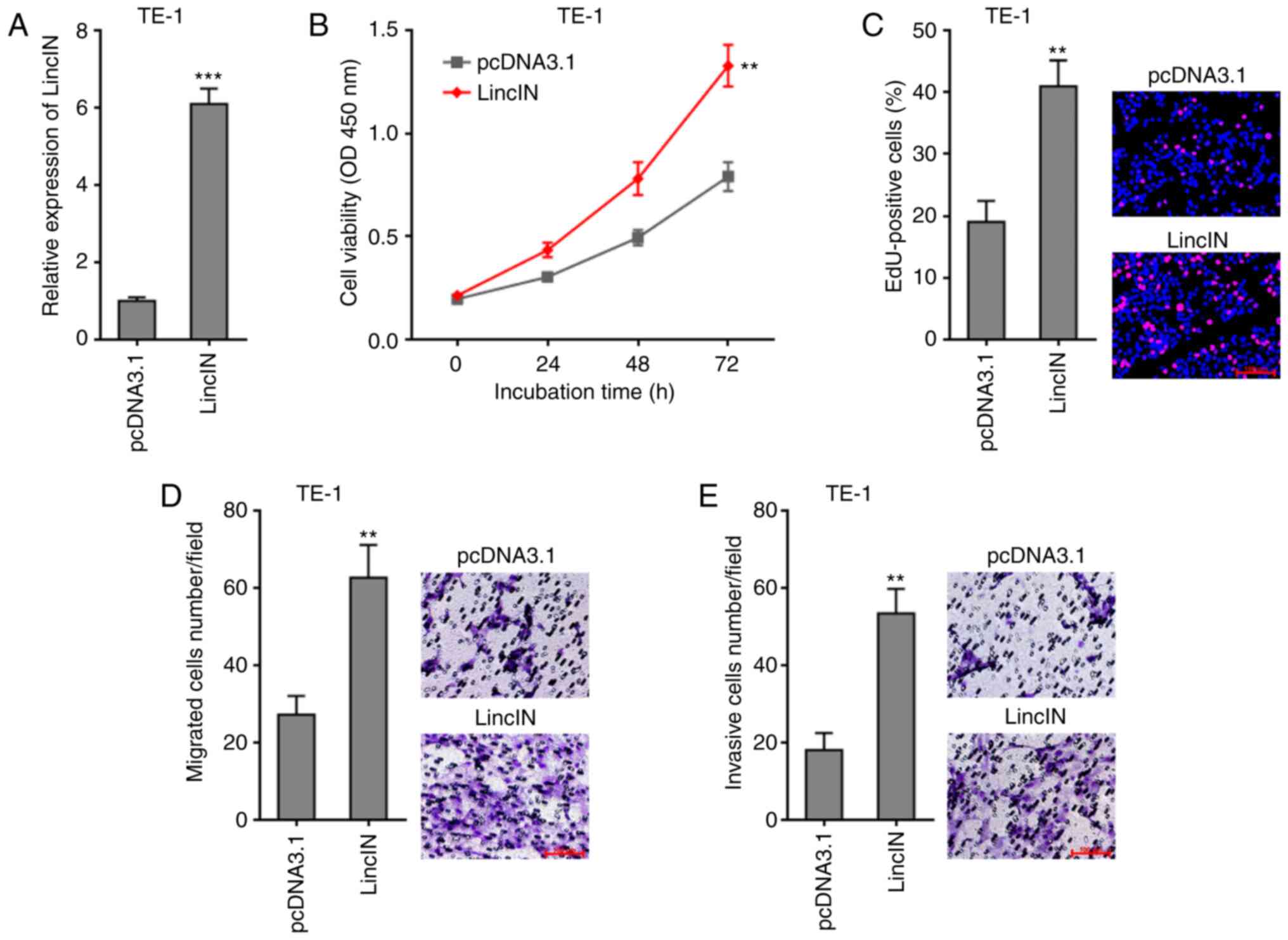
Overexpression of LincIN promotes ESCC
cell growth, migration and invasion
To explore the biological roles of LincIN in ESCC,
LincIN was stably overexpressed in the TE-1 cells, which exhibited
the lowest LincIN expression out of the 3 ESCC lines (Fig. 1C), by transfection with LincIN
overexpression and control plasmids (Fig. 2A). CCK-8 assays revealed that the
overexpression of LincIN promoted ESCC cell growth (Fig. 2B). EdU incorporation assays
further verified the roles of LincIN overexpression in promoting
ESCC cell growth (Fig. 2C).
Transwell migration and invasion assays also revealed that the
overexpression of LincIN promoted ESCC cell migration and invasion
(Fig. 2D and E). Collectively,
these results suggested that the overexpression of LincIN promoted
ESCC cell growth, migration and invasion.
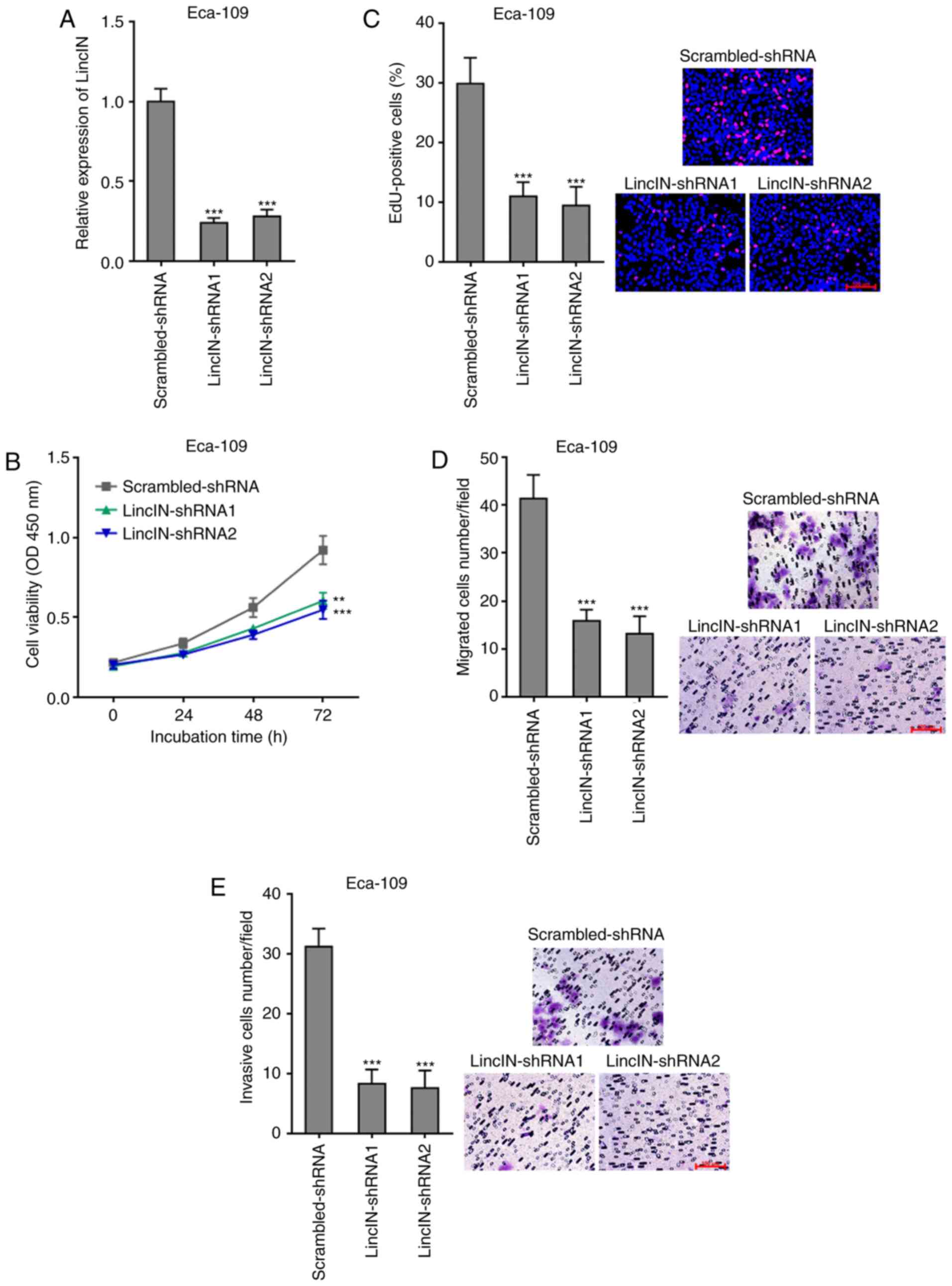
Knockdown of LincIN inhibited ESCC cell
growth, migration, and invasion
To further explore the biological roles of LincIN
knockdown in ESCC, LincIN expression was stably depleted in the
Eca-109 cells, which exhibited the highest expression of LincIN out
of the 3 ESCC cell lines, by transfection with 2 independent
LincIN-specific shRNAs (Fig.
3A). CCK-8 and EdU incorporation assays revealed that the
knockdown of LincIN inhibited ESCC cell growth (Fig. 3B and C). Transwell migration and
invasion assays also revealed that the knockdown of LincIN
inhibited ESCC cell migration and invasion (Fig. 3D and E). Collectively, these
results suggested that the knockdown of LincIN inhibited ESCC cell
growth, migration and invasion.
LincIN enhances the suppressive effects
of NF90 on miR-7 biogenesis
LincIN was previously identified as a NF90-binding
lncRNA in breast cancer (40).
Therefore, the present study further investigated whether LincIN
also bound NF90 in ESCC. RIP assays revealed that LincIN was
specifically enriched in the NF90 antibody group, suggesting the
binding between LincIN and NF90 (Fig. 4A). NF90 has been reported to
suppress miR-7 biogenesis by binding primary miR-7 (pri-miR-7)
(47). miR-7 is a well-known
tumor suppressor in a number of types of cancer, including ESCC
(46,48,49). Therefore, the present study
further explored the effects of LincIN on NF90/miR-7. RIP assays
revealed that the overexpression of LincIN promoted, while the
knockdown of LincIN suppressed the binding between NF90 and
pri-miR-7 (Fig. 4B and C),
suggesting that LincIN enhanced the binding between NF90 and
pri-miR-7. Subsequently, the expression levels of pri-miR-7 and
mature miR-7 in ESCC cells in which LincIN was stably overexpressed
or depleted were measured by RT-qPCR. The results displayed that
the overexpression of LincIN induced the accumulation of pri-miR-7
and the downregulation of mature miR-7 in ESCC cells (Fig. 4D and E). The knockdown of LincIN
downregulated the accumulation of pri-miR-7 and upregulated mature
miR-7 in ESCC cells (Fig. 4F and
G). Collectively, these results suggested that LincIN enhanced
the suppressive roles of NF90 on miR-7 biogenesis and therefore,
downregulated the mature miR-7 level.
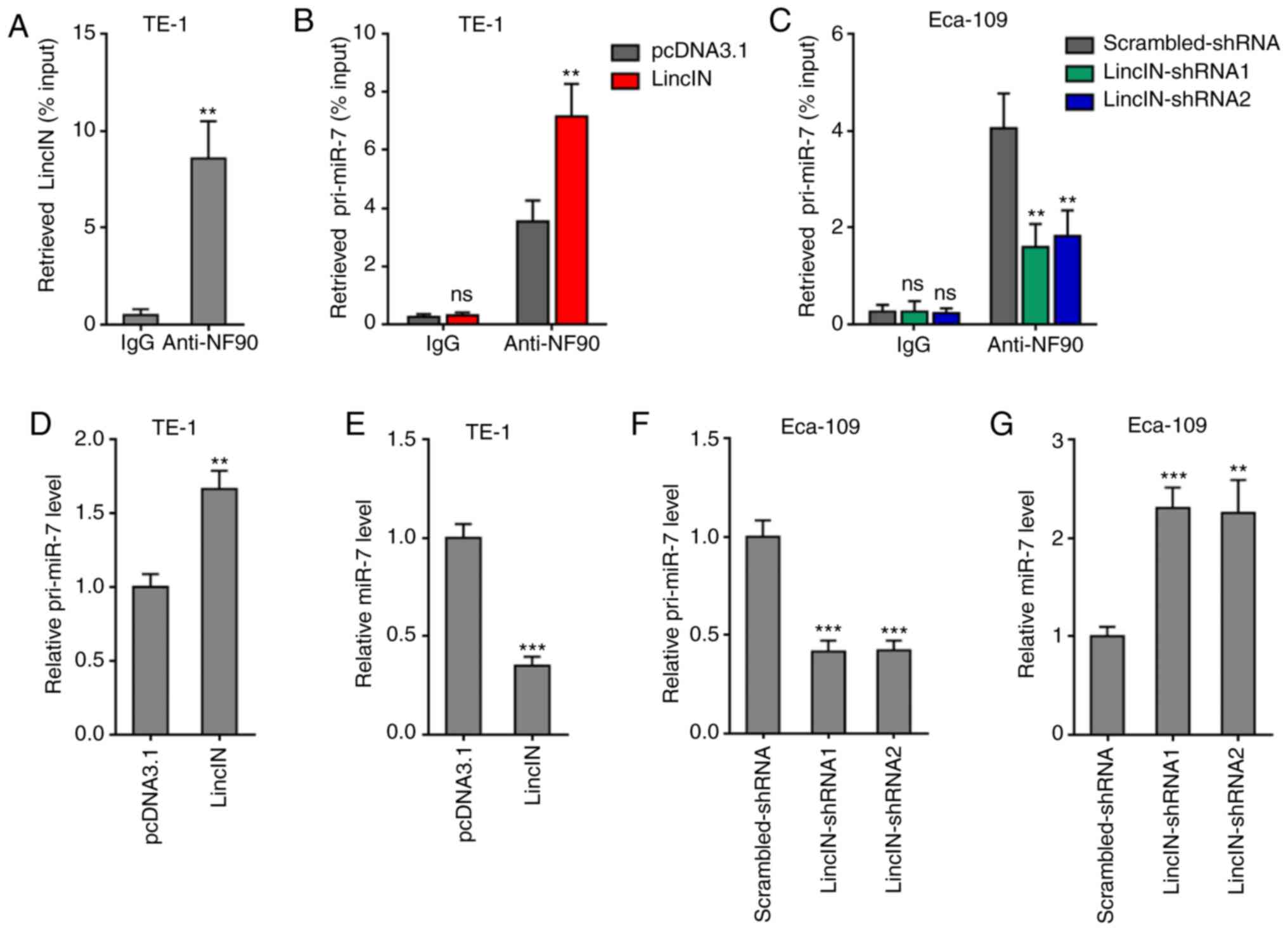 | Figure 4LincIN enhances the suppressive
effects of NF90 on miR-7 biogenesis. (A) RIP assays were performed
using the TE-1 cells, followed by RT-qPCR to detect LincIN
expression associated with NF90. (B) RIP assays were performed in
TE-1 cells transiently overexpressing LincIN or the control,
followed by RT-qPCR to detect pri-miR-7 associated with NF90. (C)
RIP assays were performed in Eca-109 cells transiently depleting
LincIN or control, followed by RT-qPCR to detect pri-miR-7
expression associated with NF90. (D) Expression of pri-miR-7 in
LincIN stably overexpressing and control TE-1 cells detected by
RT-qPCR. (E) Expression of miR-7 in LincIN stably overexpressing
and control TE-1 cells detected by RT-qPCR. (F) Expression of
pri-miR-7 in LincIN stably depleted and control Eca-109 cells
detected by RT-qPCR. (G) Expression of miR-7 in LincIN stably
depleted and control Eca-109 cells detected by RT-qPCR. Results are
shown as the means ± SD from 3 separated experiments.
**P<0.01, ***P<0.001, vs. vs. IgG,
control, or control-shRNA group; ns, not significant, determined by
the Student's t-test (A, B, D and E) or one-way ANOVA followed by
Dunnett's multiple comparisons test (C, F and G). ESCC, esophageal
squamous cell carcinoma. |
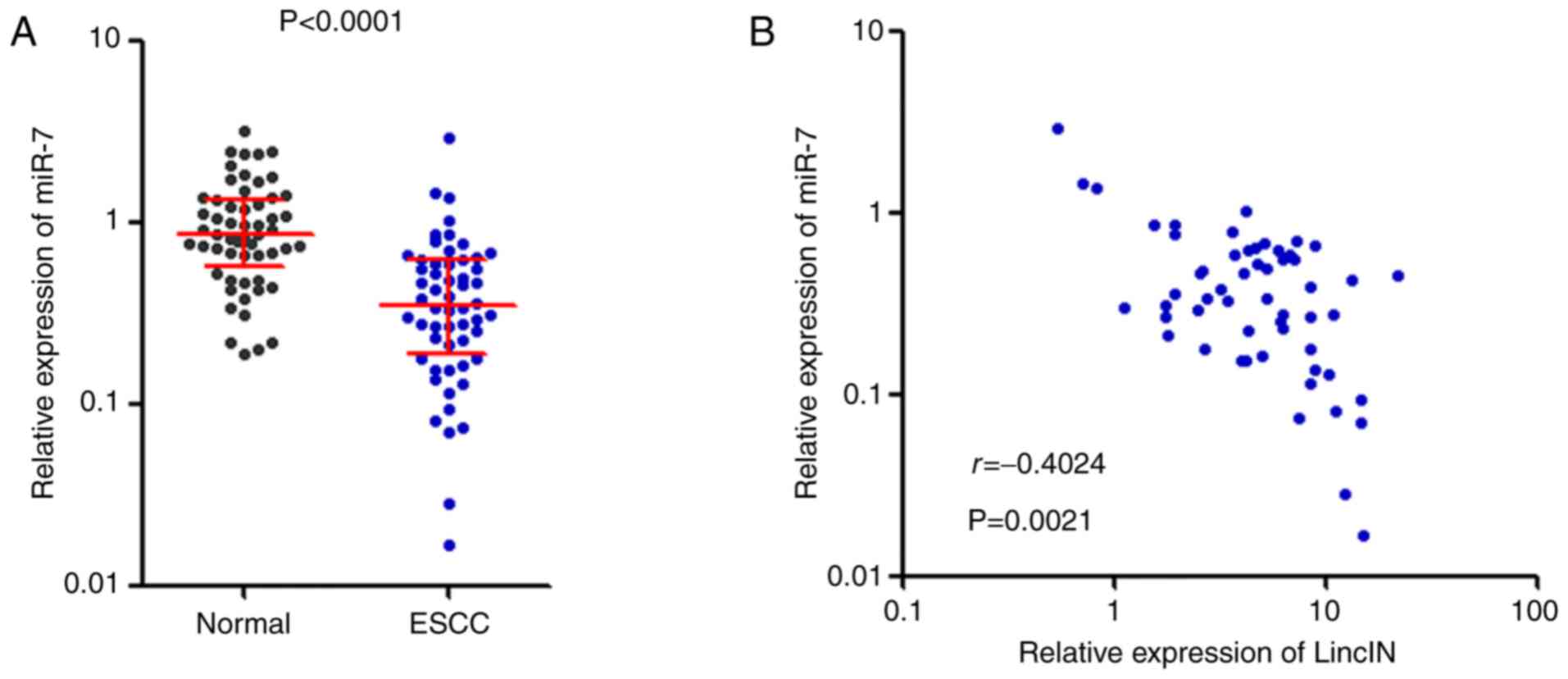
miR-7 expression is decreased and
inversely correlates with LincIN expression in ESCC tissues
To determine whether the downregulation of miR-7 by
LincIN exists in vivo, the expression of miR-7 was measured
in the same 56 pairs of ESCC tissues and adjacent non-cancerous
tissues used in Fig. 1.
Conversely to the expression pattern of LincIN, miR-7 expression
was significantly decreased in ESCC tissues compared with that in
adjacent non-cancerous tissues (Fig.
5A). Furthermore, a statistically significant inverse
correlation between miR-7 and LincIN expression levels was found in
these 56 ESCC tissues (r=−0.4024, P=0.0021, Fig. 5B).
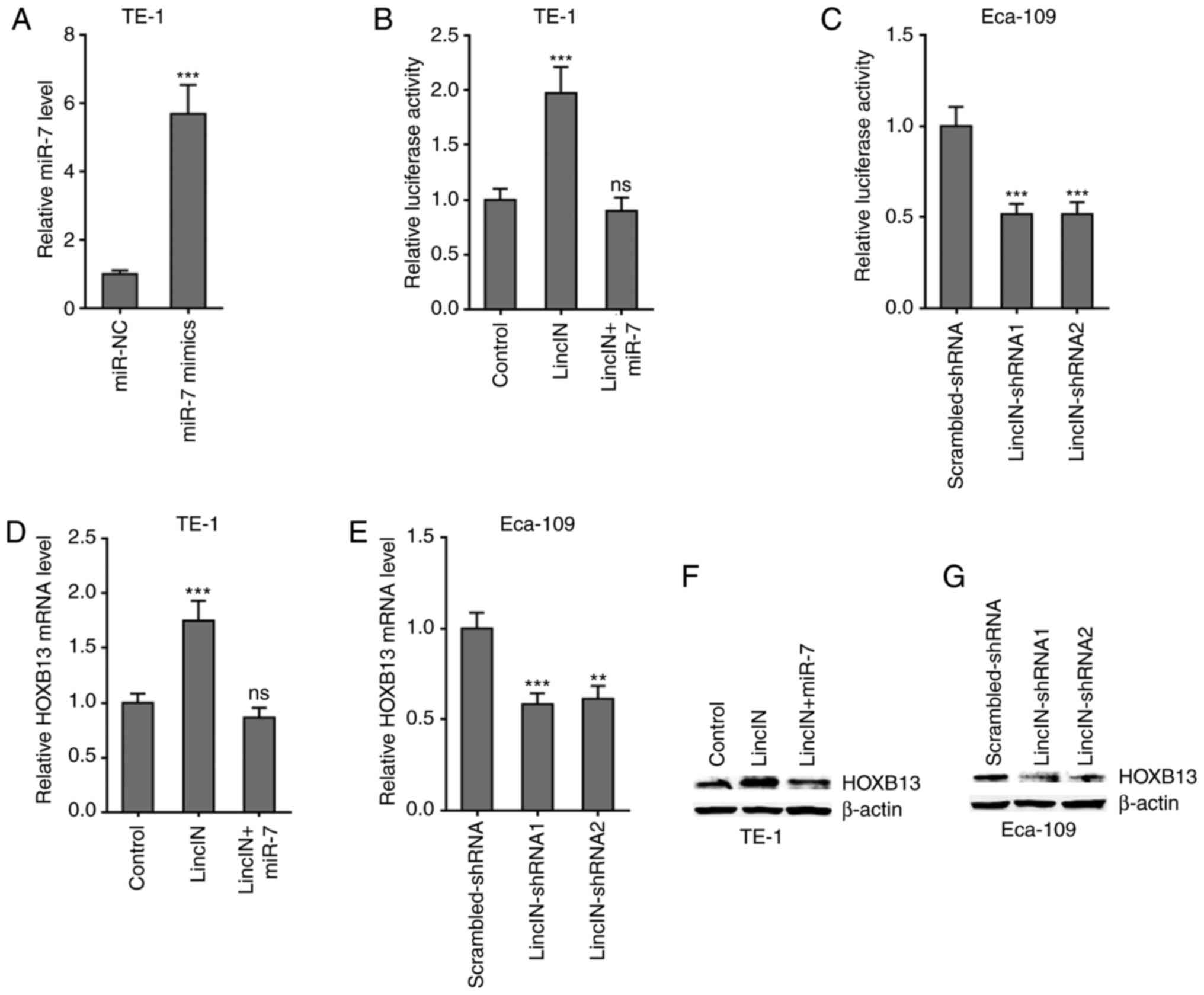
LincIN upregulates HOXB13 expression by
suppressing miR-7
HOXB13 was previously identified as a critical
target of miR-7 and it was found to mediate the tumor suppressive
roles of miR-7 (46,50). Therefore, the present study
further examined the effects of LincIN on HOXB13. Dual-luciferase
reporter assays revealed that the overexpression of LincIN
significantly upregulated the 3′UTR activity of HOXB13, which was
reversed by the concurrent overexpression of miR-7 (Fig. 6A and B). Reciprocally, the
knockdown of LincIN significantly reduced the 3′UTR activity of
HOXB13 (Fig. 6C). RT-qPCR assays
revealed that the overexpression of LincIN significantly
upregulated the HOXB13 mRNA level, which was reversed by the
concurrent overexpression of miR-7 (Fig. 6D). Reciprocally, the knockdown of
LincIN significantly reduced the HOXB13 mRNA level (Fig. 6E). Western blot assays revealed
that the overexpression of LincIN markedly upregulated the HOXB13
protein level, which was reversed by the concurrent overexpression
of miR-7 (Fig. 6F).
Reciprocally, the knockdown of LincIN markedly reduced the HOXB13
protein level (Fig. 6G).
Collectively, these results suggested that LincIN upregulated
HOXB13 by suppressing miR-7.
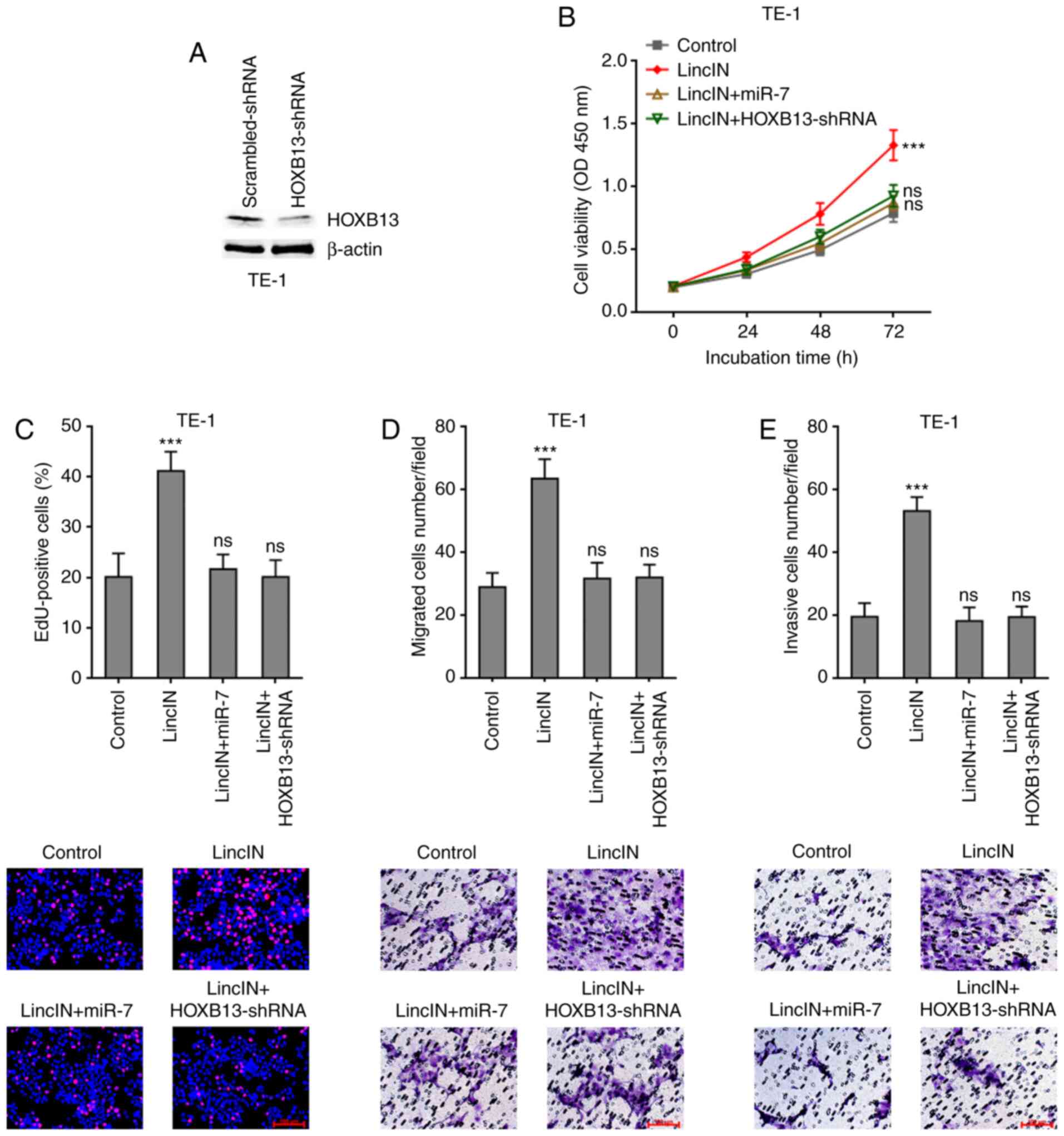
Overexpression of miR-7 or depletion of
HOXB13 attenuates the promoting effects of LincIN in ESCC cell
growth, migration and invasion
To examine whether miR-7/HOXB13 mediate the roles of
LincIN in promoting ESCC cell growth, migration and invasion, miR-7
was stably overexpressed or HOXB13 was depleted in TE-1 cells
stably overexpressing LincIN (Fig.
7A). CCK-8 and EdU incorporation assays revealed that both
miR-7 overexpression and HOXB13 depletion attenuated the promoting
effects of LincIN on ESCC cell growth (Fig. 7B and C). Transwell migration and
invasion assays also revealed that both miR-7 overexpression and
HOXB13 depletion attenuated the promoting effects of LincIN on ESCC
cell migration and invasion (Fig. 7D
and E). Collectively, these results suggested that the
overexpression of miR-7 or depletion of HOXB13 both attenuated the
promoting effects of LincIN on ESCC cell growth, migration and
invasion.
Discussion
As one of the most lethal types of cancer, ESCC
involves complex molecular mechanisms which remain unclear.
Whole-exome sequencings have identified a number of mutations in
ESCC, including TP53, CCND1, CDKN2A,
NFE2L2, RB1, KMT2D, KMT2C,
EP300, FAT1, NOTCH1 and others (51). Furthermore, transcriptome
sequencing analyses have identified a number of differentially
expressed mRNAs and lncRNAs in ESCC (52). The expression and roles of
several lncRNAs in ESCC have also been investigated (50). lncRNA CCAT1 has been shown to be
upregulated in ESCC tissues and to be associated with a poor
survival (50). CCAT1 promotes
ESCC cell proliferation and migration by downregulating SPRY4 and
upregulating HOXB13 (50). CCAT1
binds and recruits EZH2 and SUV39H1 to epigenetically silence SPRY4
(45). Furthermore, CCAT1
upregulates HOXB13 by competitively binding miR-7 (45). lncRNA TTN-AS1 is highly expressed
in ESCC, and promotes ESCC cell proliferation and metastasis via
sponging miR-133b and inducing Snail1 and FSCN1 expression
(53). lncRNA NMR promotes ESCC
progression via NSUN2 and BPTF (54). Other lncRNAs have also been
reported to have oncogenic or tumor suppressive roles in ESCC, such
as PART1, CASC9, HOTTIP, LUCAT1, HNF1A-AS1, AFAP1-AS1, POU3F3 and
others (55-61).
Transcriptome sequencing has identified >58,000
lncRNAs in human cells (62).
Due to the large number of lncRNAs, other lncRNAs may also play
biological roles in human ESCC. lncRNA LincIN, whose encoding gene
is located on chromosome 10p11-12, was first identified in breast
tumors (40). LincIN is
overexpressed in human breast tumors, and is associated with a poor
prognosis of patients with breast cancer, and is involved in the
metastasis of breast cancer (40). However, the expression, clinical
significance and role of LincIN in other diseases remain unknown.
In the present study, it was found that LincIN expression was also
significantly increased in ESCC tissues and cell lines, and was
associated with an advanced clinical stage and a poor prognosis of
patients with ESCC. Functional experiments revealed that the
overexpression of LincIN promoted ESCC cell growth, migration and
invasion. The knockdown of LincIN inhibited ESCC cell growth,
migration and invasion. Thus, these data suggested LincIN as a
promising prognostic biomarker and therapeutic target for ESCC.
The mechanisms of action of lncRNAs are complex. One
of the main mechanisms is to bind proteins (13). In breast tumors, LincIN was
revealed to bind NF90 (40).
NF90, also known as ILF3, is an RNA binding protein (63). By binding double-stranded RNA,
NF90 regulates gene expression and/or mRNA stabilities (64). In addition, by binding
pri-miRNAs, NF90 modulates miRNAs biogenesis (47). The biogenesis of miR-7, which
plays tumor suppressive roles in a number of types of cancer, has
been reported to be suppressed by NF90 (46,47). Therefore, the present study
further investigated the effects of LincIN on NF90/miR-7. It was
verified that LincIN directly bound NF90 in ESCC cells. Moreover,
LincIN promoted the binding between NF90 and pri-miR-7, thereby
enhancing the suppressive roles of NF90 on miR-7 biogenesis. It was
then verified that LincIN downregulated mature miR-7. Conversely to
LincIN, miR-7 expression was significantly decreased in ESCC
tissues. The expression of mature miR-7 was significantly and
inversely associated with that of LincIN in ESCC tissues,
supporting the negative regulation of miR-7 by LincIN. HOXB13 is a
direct target of miR-7 and plays oncogenic roles in several types
of cancer, including ESCC (46,50). By repressing miR-7, LincIN was
verified to upregulate HOXB13 expression in ESCC cells. Functional
rescue assays demonstrated that the overexpression of miR-7 or
depletion of HOXB13 attenuated the promoting effects of LincIN on
ESCC cell growth, migration and invasion, which further suggested
that miR-7/HOXB13 are critical mediators of the oncogenic roles of
LincIN in ESCC. Similar roles of lncRNAs in miRNAs biogenesis have
also been reported in other types of cancer, such as the promotion
of miR-145 biogenesis by lncRNA-ATB in bladder cancer and the
disruption of miR-125b biogenesis by LINC01578 in breast cancer
(63,65). The detailed mechanisms underlying
the inverse effects of lncRNAs on the biogenesis of different
miRNAs warrant further investigation.
Apart from miR-7, NF90 has also been reported to
suppress p21 translation, upregulate hypoxia inducible factor-1α
(HIF-1α) and VEGF-A, stabilize PARP1 mRNA, and facilitate DICER
expression (40,64,66,67). Thus, whether LincIN modulates
p21, HIF-1α, VEGF-A, PARP1 and DICER via interacting with NF90 in
ESCC cells needs to be further investigated. Nevertheless, the
present study identified a novel mechanism of lncRNA in ESCC and
provides a novel target against ESCC.
In conclusion, the present study demonstrates that
lncRNA LincIN is highly expressed in ESCC, and is associated with
an advanced clinical stage and a poor prognosis of patients with
ESCC. LincIN promotes ESCC cell growth, migration and invasion by
binding NF90, suppressing the biogenesis of miR-7 and upregulating
HOXB13. The data presented herein suggest that LincIN may be a
promising prognostic biomarker and therapeutic target for ESCC.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
DW and ZT designed the study. ZT, WS, PZ, YW, ZZ, ZG
and MS performed the experiments. DW, ZT and YX performed the
statistical analysis. DW and ZT wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The use of clinical specimens was reviewed and
approved by the Nanfang Hospital Institutional Review Board
(Guangzhou, China). Written informed consent was acquired from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Mr. Peng Jiang for
providing assistance with language editing.
Funding
The present study was supported by the National Nature Science
Foundation of China (grant nos. 81672756, 81872399, 81772631 and
81572849), the Natural Science Foundation of Guangdong Province
(grant no. 2018A030313530), the Chinese Scholarship Council (grant
no. 201908440010), the Shenzhen Healthcare Research Project (grant
no. SZFZ2017105), the Seedling Program of Shenzhen Hospital,
Southern Medical University (grant no. 2016MM06) and the Sanming
Project of Medicine in Shenzhen (grant no. SZSM201612041).
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rustgi AK and El-Serag HB: Esophageal
carcinoma. N Engl J Med. 371:2499–2509. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sjoquist KM, Burmeister BH, Smithers BM,
Zalcberg JR, Simes RJ, Barbour A and Gebski V; Australasian
Gastro-Intestinal Trials Group: Survival after neoadjuvant
chemotherapy or chemoradiotherapy for resectable oesophageal
carcinoma: An updated meta-analysis. Lancet Oncol. 12:681–692.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carninci P, Kasukawa T, Katayama S, Gough
J, Frith MC, Maeda N, Oyama R, Ravasi T, Lenhard B, Wells C, et al:
The transcriptional landscape of the mammalian genome. Science.
309:1559–1563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Berger AC, Korkut A, Kanchi RS, Hegde AM,
Lenoir W, Liu W, Liu Y, Fan H, Shen H, Ravikumar V, et al: A
comprehensive pan-cancer molecular study of gynecologic and breast
cancers. Cancer Cell. 33:690–705. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan JH, Yang F, Wang F, Ma Jz, Guo Yj,
Tao Qf, Liu F, Pan W, Wang Tt, Zhou Cc, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang C, Yuan J, Hu H, Chen W, Liu M,
Zhang J, Sun S and Guo Z: Long non-coding RNA CHCHD4P4 promotes
epithelial-mesenchymal transition and inhibits cell proliferation
in calcium oxalate-induced kidney damage. Braz J Med Biol Res.
51:e65362017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yin D, Lu X, Su J, He X, De W, Yang J, Li
W, Han L and Zhang E: Long noncoding RNA AFAP1-AS1 predicts a poor
prognosis and regulates non-small cell lung cancer cell
proliferation by epigenetically repressing p21 expression. Mol
Cancer. 17:922018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu WL, Jin L, Xu A, Wang YF, Thorne RF,
Zhang XD and Wu M: GUARDIN is a p53-responsive long non-coding RNA
that is essential for genomic stability. Nat Cell Biol. 20:492–502.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li JK, Cheng C, Liu JY, Shi JZ, Liu SP,
Liu B, Wu DS, Fang ZY, Bao Y, Jiang MM, et al: Long noncoding RNA
MRCCAT1 promotes metastasis of clear cell renal cell carcinoma via
inhibiting NPR3 and activating p38-MAPK signaling. Mol Cancer.
16:1112017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Zheng X, Wang N, Zhao W, Zhang X,
Teng S, Zhang Y and Lu Z: Long noncoding RNA DANCR, working as a
competitive endogenous RNA, promotes ROCK1-mediated proliferation
and metastasis via decoying of miR-335-5p and miR-1972 in
osteosarcoma. Mol Cancer. 17:892018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu H, Zheng T, Yu J, Zhou L and Wang L:
LncRNA XIST accelerates cervical cancer progression via
upregulating fus through competitively binding with miR-200a.
Biomed Pharmacother. 105:789–797. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z, Yang B, Zhang M, Guo W, Wu Z, Wang
Y, Jia L, Li S; Cancer Genome Atlas Research Network; Xie W and
Yang D: lncRNA epigenetic landscape analysis identifies EPIC1 as an
oncogenic lncRNA that interacts with MYC and promotes cell-cycle
progression in cancer. Cancer Cell. 33:706–720. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mondal T, Juvvuna PK, Kirkeby A, Mitra S,
Kosalai ST, Traxler L, Hertwig F, Wernig-Zorc S, Miranda C, Deland
L, et al: Sense-Antisense lncRNA pair encoded by locus 6p223
determines neuroblastoma susceptibility via the USP36-CHD7-SOX9
regulatory axis. Cancer Cell. 33:417–434. 2018. View Article : Google Scholar
|
|
20
|
Lin A, Hu Q, Li C, Xing Z, Ma G, Wang C,
Li J, Ye Y, Yao J, Liang K, et al: The LINK-A lncRNA interacts with
PtdIns(3,4,5) P3 to hyperactivate AKT and confer resistance to AKT
inhibitors. Nat Cell Biol. 19:238–251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grelet S, Link LA, Howley B, Obellianne C,
Palanisamy V, Gangaraju VK, Diehl JA and Howe PH: A regulated PNUTS
mRNA to lncRNA splice switch mediates EMT and tumour progression.
Nat Cell Biol. 19:1105–1115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gilot D, Migault M, Bachelot L, Journé F,
Rogiers A, Donnou-Fournet E, Mogha A, Mouchet N, Pinel-Marie ML,
Mari B, et al: A non-coding function of TYRP1 mRNA promotes
melanoma growth. Nat Cell Biol. 19:1348–1357. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu XT, Yuan JH, Zhu TT, Li YY and Cheng
XY: Long noncoding RNA glypican 3 (GPC3) antisense transcript 1
promotes hepatocellular carcinoma progression via epigenetically
activating GPC3. FEBS J. 283:3739–3754. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tsai KW, Lo YH, Liu H, Yeh CY, Chen YZ,
Hsu CW, Chen WS and Wang JH: Linc00659, a long noncoding RNA, acts
as novel oncogene in regulating cancer cell growth in colorectal
cancer. Mol Cancer. 17:722018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Wang W, Liu G, Xie S, Li Q, Li Y
and Lin Z: Long non-coding RNA HOTTIP promotes hypoxia-induced
epithelial-mesenchymal transition of malignant glioma by regulating
the miR-101/ZEB1 axis. Biomed Pharmacother. 95:711–720. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Michelini F, Pitchiaya S, Vitelli V,
Sharma S, Gioia U, Pessina F, Cabrini M, Wang Y, Capozzo I,
Iannelli F, et al: Damage-Induced lncRNAs control the DNA damage
response through interaction with DDRNAs at individual
double-strand breaks. Nat Cell Biol. 19:1400–1411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuan JH, Liu XN, Wang TT, Pan W, Tao QF,
Zhou WP, Wang F and Sun SH: The MBNL3 splicing factor promotes
hepatocellular carcinoma by increasing PXN expression through the
alternative splicing of lncRNA-PXN-AS1. Nat Cell Biol. 19:820–832.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su R, Cao S, Ma J, Liu Y, Liu X, Zheng J,
Chen J, Liu L, Cai H, Li Z, et al: Knockdown of SOX2OT inhibits the
malignant biological behaviors of glioblastoma stem cells via
up-regulating the expression of miR-194-5p and miR-122. Mol Cancer.
16:1712017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao J, Zhang L, Zheng L, Hong Y and Zhao
L: LncRNATCF7 promotes the growth and self-renewal of glioma cells
via suppressing the miR-200c-EpCAM axis. Biomed Pharmacother.
97:203–208. 2018. View Article : Google Scholar
|
|
30
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mohr S, Doebele C, Comoglio F, Berg T,
Beck J, Bohnenberger H, Alexe G, Corso J, Ströbel P, Wachter A, et
al: Hoxa9 and meis1 cooperatively induce addiction to syk signaling
by suppressing miR-146a in acute myeloid leukemia. Cancer Cell.
31:549–562. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bu Y, Yoshida A, Chitnis N, Altman BJ,
Tameire F, Oran A, Gennaro V, Armeson KE, McMahon SB, Wertheim GB,
et al: A PERK-miR-211 axis suppresses circadian regulators and
protein synthesis to promote cancer cell survival. Nat Cell Biol.
20:104–115. 2018. View Article : Google Scholar
|
|
33
|
Yan W, Wu X, Zhou W, Fong MY, Cao M, Liu
J, Liu X, Chen CH, Fadare O, Pizzo DP, et al: Cancer-Cell-Secreted
exosomal miR-105 promotes tumour growth through the MYC-dependent
metabolic reprogramming of stromal cells. Nat Cell Biol.
20:597–609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chernyy V, Pustylnyak V, Kozlov V and
Gulyaeva L: Increased expression of miR-155 and miR-222 is
associated with lymph node positive status. J Cancer. 9:135–140.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Leite KR, Reis ST, Viana N, Morais DR,
Moura CM, Silva IA, Pontes J Jr, Katz B and Srougi M: Controlling
RECK miR21 promotes tumor cell invasion and is related to
biochemical recurrence in prostate cancer. J Cancer. 6:292–301.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Celia-Terrassa T, Liu DD, Choudhury A,
Hang X, Wei Y, Zamalloa J, Alfaro-Aco R, Chakrabarti R, Jiang YZ,
Koh BI, et al: Normal and cancerous mammary stem cells evade
interferon-induced constraint through the miR-199a-LCOR axis. Nat
Cell Biol. 19:711–723. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu MZ, Cheng WC, Chen SF, Nieh S, O'Connor
C, Liu CL, Tsai WW, Wu CJ, Martin L, Lin YS, et al: MiR-25/93
mediates hypoxia-induced immunosuppression by repressing cGAS. Nat
Cell Biol. 19:1286–1296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yuan JH, Yang F, Chen BF, Lu Z, Huo Xs,
Zhou Wp, Wang F and Sun Sh: The histone deacetylase
4/SP1/microrna-200a regulatory network contributes to aberrant
histone acetylation in hepatocellular carcinoma. Hepatology.
54:2025–2035. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ho CS, Noor SM and Nagoor NH: MiR-378 and
MiR-1827 regulate tumor invasion, migration and angiogenesis in
human lung adenocarcinoma by targeting RBX1 and CRKL, respectively.
J Cancer. 9:331–345. 2018. View Article : Google Scholar :
|
|
40
|
Jiang Z, Slater CM, Zhou Y, Devarajan K,
Ruth KJ, Li Y, Cai KQ, Daly M and Chen X: LincIN, a novel
NF90-binding long non-coding RNA, is overexpressed in advanced
breast tumors and involved in metastasis. Breast Cancer Res.
19:622017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nishihira T, Kasai M, Mori S, Watanabe T,
Kuriya Y, Suda M, Kitamura M, Hirayama K, Akaishi T and Sasaki T:
Characteristics of two cell lines (TE-1 and TE-2) derived from
human squamous cell carcinoma of the esophagus. Gan. 70:575–584.
1979.PubMed/NCBI
|
|
42
|
Barnas C, Martel-Planche G, Furukawa Y,
Hollstein M, Montesano R and Hainaut P: Inactivation of the p53
protein in cell lines derived from human esophageal cancers. Int J
Cancer. 71:79–87. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gao J, Xue KX, Li BG, Dong HY, Wei LS,
Zhang ZH, Tian S and Xu ZG: Site-Dependence of invasiveness of
ECA109 human oesophageal carcinoma cells in nude mice. Clin Exp
Metastasis. 2:205–212. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ye Z, Fang J, Dai S, Wang Y, Fu Z, Feng W,
Wei Q and Huang P: MicroRNA-34a induces a senescence-like change
via the down-regulation of SIRT1 and up-regulation of p53 protein
in human esophageal squamous cancer cells with a wild-type p53 gene
background. Cancer Lett. 370:216–221. 2016. View Article : Google Scholar
|
|
45
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
46
|
Li RC, Ke S, Meng FK, Lu J, Zou XJ, He ZG,
Wang WF and Fang MH: CiRS-7 promotes growth and metastasis of
esophageal squamous cell carcinoma via regulation of miR-7/HOXB13.
Cell Death Dis. 9:8382018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Higuchi T, Todaka H, Sugiyama Y, Ono M,
Tamaki N, Hatano E, Takezaki Y, Hanazaki K, Miwa T, Lai S, et al:
Suppression of microRNA-7 (miR-7) biogenesis by nuclear factor
90-nuclear factor 45 complex (NF90-NF45) controls cell
proliferation in hepatocellular carcinoma. J Biol Chem.
291:21074–21084. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Su C, Han Y, Zhang H, Li Y, Yi L, Wang X,
Zhou S, Yu D, Song X, Xiao N, et al: CiRS-7 targeting miR-7
modulates the progression of non-small cell lung cancer in a manner
dependent on NF-κB signalling. J Cell Mol Med. 22:3097–3107. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu B, Yang T, Wang Z, Zhang Y, Liu S and
Shen M: CircRNA CDR1as/miR-7 signals promote tumor growth of
osteosarcoma with a potential therapeutic and diagnostic value.
Cancer Manag Res. 10:4871–4880. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang E, Han L, Yin D, He X, Hong L, Si X,
Qiu M, Xu T, De W, Xu L, et al: H3K27 acetylation activated-long
non-coding RNA CCAT1 affects cell proliferation and migration by
regulating SPRY4 and HOXB13 expression in esophageal squamous cell
carcinoma. Nucleic Acids Res. 45:3086–3101. 2017. View Article : Google Scholar :
|
|
51
|
Gao YB, Chen ZL, Li JG, Hu XD, Shi XJ, Sun
ZM, Zhang F, Zhao ZR, Li ZT, Liu ZY, et al: Genetic landscape of
esophageal squamous cell carcinoma. Nat Genet. 46:1097–1102. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li CQ, Huang GW, Wu ZY, Xu YJ, Li XC, Xue
YJ, Zhu Y, Zhao JM, Li M, Zhang J, et al: Integrative analyses of
transcriptome sequencing identify novel functional lncRNAs in
esophageal squamous cell carcinoma. Oncogenesis. 6:e2972017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lin C, Zhang S, Wang Y, Wang Y, Nice E,
Guo C, Zhang E, Yu L, Li M, Liu C, et al: Functional role of a
novel long noncoding RNA TTN-AS1 in esophageal squamous cell
carcinoma progression and metastasis. Clin Cancer Res. 24:486–498.
2018. View Article : Google Scholar
|
|
54
|
Li Y, Li J, Luo M, Zhou C, Shi X, Yang W,
Lu Z, Chen Z, Sun N and He J: Novel long noncoding RNA NMR promotes
tumor progression via NSUN2 and BPTF in esophageal squamous cell
carcinoma. Cancer Lett. 430:57–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kang M, Ren M, Li Y, Fu Y, Deng M and Li
C: Exosome-mediated transfer of lncRNA PART1 induces gefitinib
resistance in esophageal squamous cell carcinoma via functioning as
a competing endogenous RNA. J Exp Clin Cancer Res. 37:1712018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wu Y, Hu L, Liang Y, Li J, Wang K, Chen X,
Meng H, Guan X, Yang K and Bai Y: Up-Regulation of lncRNA CASC9
promotes esophageal squamous cell carcinoma growth by negatively
regulating PDCD4 expression through EZH2. Mol Cancer. 16:1502017.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin C, Wang Y, Wang Y, Zhang S, Yu L, Guo
C and Xu H: Transcriptional and posttranscriptional regulation of
HOXA13 by lncRNA HOTTIP facilitates tumorigenesis and metastasis in
esophageal squamous carcinoma cells. Oncogene. 36:5392–5406. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yoon JH, You BH, Park CH, Kim YJ, Nam JW
and Lee SK: The long noncoding RNA LUCAT1 promotes tumorigenesis by
controlling ubiquitination and stability of DNA methyltransferase 1
in esophageal squamous cell carcinoma. Cancer Lett. 417:47–57.
2018. View Article : Google Scholar
|
|
59
|
Yang X, Song JH, Cheng Y, Wu W, Bhagat T,
Yu Y, Abraham JM, Ibrahim S, Ravich W, Roland BC, et al: Long
non-coding RNA HNF1A-AS1 regulates proliferation and migration in
oesophageal adenocarcinoma cells. Gut. 63:881–890. 2014. View Article : Google Scholar
|
|
60
|
Luo HL, Huang MD, Guo JN, Fan RH, Xia XT,
He JD and Chen XF: AFAP1-AS1 is upregulated and promotes esophageal
squamous cell carcinoma cell proliferation and inhibits cell
apoptosis. Cancer Med. 5:2879–2885. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li W, Zheng J, Deng J, You Y, Wu H, Li N,
Lu J and Zhou Y: Increased levels of the long intergenic
non-protein coding RNAPOU3F3 promote DNA methylation in esophageal
squamous cell carcinoma cells. Gastroenterology. 146:1714–1726
e1715. 2014. View Article : Google Scholar
|
|
62
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhuang J, Shen L, Yang L, Huang X, Lu Q,
Cui Y, Zheng X, Zhao X, Zhang D, Huang R, et al: TGFβ1 promotes
gemcitabine resistance through regulating the
LncRNA-LET/NF90/miR-145 signaling axis in bladder cancer.
Theranostics. 7:3053–3067. 2017. View Article : Google Scholar :
|
|
64
|
Zhang W, Xiong Z, Wei T, Li Q, Tan Y, Ling
L and Feng X: Nuclear factor 90 promotes angiogenesis by regulating
HIF-1α/VEGF-A expression through the PI3K/akt signaling pathway in
human cervical cancer. Cell Death Dis. 9:2762018. View Article : Google Scholar
|
|
65
|
Liu J, Zhan Y, Wang J, Wang J, Guo J and
Kong D: Long noncoding RNA LINC01578 drives colon cancer metastasis
through a positive feedback loop with the NF-κB/YY1 axis. Mol
Oncol. 14:3211–3233. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Song D, Huang H, Wang J, Zhao Y, Hu X, He
F, Yu L and Wu J: NF90 regulates PARP1 mRNA stability in
hepatocellular carcinoma. Biochem Biophys Res Commun. 488:211–217.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Barbier J, Chen X, Sanchez G, Cai M,
Helsmoortel M, Higuchi T, Giraud P, Contreras X, Yuan G, Feng Z, et
al: An NF90/NF110-mediated feedback amplification loop regulates
dicer expression and controls ovarian carcinoma progression. Cell
Res. 28:556–571. 2018. View Article : Google Scholar : PubMed/NCBI
|