Introduction
Lung cancer is the main cause of cancer-related
mortality worldwide, and non-small cell cancer accounts for
approximately 85% of all lung cancer cases (1,2).
It is mainly characterized by a poor prognosis and a high risk of
recurrence. The current treatment for lung cancer includes
radiotherapy and chemotherapy. Despite the rapid development of
modern technology and treatments, the mortality rate of patients
with lung cancer remains high, and a tendency for relapse has been
observed (3-5). Therefore, there is an urgent need
to explore new drugs and treatment methods for lung cancer.
Ceramide is a biologically active sphingolipid that
is the precursor of all major sphingomyelins. It plays an important
role in the cell cycle, differentiation, senescence and apoptosis
(6,7). In addition, ceramide is considered
to be an important secondary messenger in the apoptosis and
necrosis pathways in normal and cancer cells (8-11). Being in the center of
sphingolipid metabolism, ceramide synthesis mainly occurs via de
novo, salvage and sphingomyelinase synthesis pathways. In
ceramide degradation, mainly 3 ceramidases, namely alkaline,
neutral and acid ceramidase, are known to be involved (12,13). In previous studies, ceramide has
been shown to exhibit significant antitumor activity, and its
mediated antitumor activity has been reported in multiple cancer
types, such as breast and prostate cancers (14-16). Inhibiting the activity of
ceramidases and increasing the level of ceramide can effectively
prevent resistance to chemotherapeutic drugs and induce apoptosis.
Enzymes of the ceramide and ceramide synthase family play an
important role in cancer (17,18). However, the mechanisms through
which ceramide exerts its antitumor activity remain unclear.
The thioredoxin-interacting protein (Txnip), also
known as thioredoxin-binding protein 2 (TBP-2) (19), in several cases, regulates a
variety of oxidative stresses together with glutathione; it can
also directly inhibit the exchange between the disulfide bonds of
thioredoxin (Trx)1 to inhibit the reducing activity of Trx
(19-22). Trx is an important redox
regulator and a sensor of energy and glucose metabolism. It is
involved in various physiological processes, such as DNA damage
repair, inflammation and apoptosis. It is divided into two
different types, depending on the subcellular localization: Trx1 in
the cytoplasm and Trx2 in the mitochondria (23-25). Previous research has demonstrated
that Trx mimetic peptides inhibit the expression of p38 MAPK in
vitro, thereby protecting the rat brain from apoptosis and
inflammation (26). Furthermore,
regulating the Txnip/Trx1 complex can inhibit the activation of the
NLR family pyrin domain containing 3 (NLRP3) inflammasome, thus
reducing cerebral ischemia-reperfusion injury in rat (27).
Therefore, it was hypothesized that ceramide
regulates the apoptosis of non-small cell lung cancer cells,
induced by the Txnip/Trx1 complex. The present study aimed to
examine this hypothesis by stimulating the apoptosis of non-small
lung cancer cells with ceramide in vitro. In addition, the
expression levels of Txnip, Trx1, p38 and other proteins in the
cells detecting, and the association analyzing between Txnip and
Trx1 proteins was investigated. The importance of the Txnip/Trx1
complex in ceramide-induced apoptosis of lung cancer cells was
further investigated at the cellular and molecular level.
Materials and methods
Cells, reagents and antibodies
The A549 and PC9 cell lines (human lung
adenocarcinoma cells) were provided by the School of Life Sciences,
University of Science and Technology of China. All cells were
cultured in Dulbecco's modified Eagle's medium (DMEM) (containing
10% fetal bovine serum, 50 U/ml penicillin, and 0.05 mg/ml
streptomycin) at 37°C in a 5% CO2 incubator. C2-Ceramide
and verapamil were purchased from Sigma-Aldrich; Merck KGaA.
Anti-β-actin (no. ab8224), anti-p38 (no. ab170099), anti-caspase-3
(no. ab32351) and anti-cleaved caspase-3 (no. ab2302) antibodies
were purchased from Abcam. Anti-Txnip (no. 18243-1-AP) antibody was
purchased from ProteinTech Group. Anti-Trx (no. NBP2-52983)
antibody was purchased from Novus Biologicals. The Cell Counting
kit-8 (CCK-8) and apoptosis detection kits were provided by
Beyotime Biotechnology, and TRIzol reagent was provided by Takara
Bio, Inc.
Cell line viability determination
The CCK-8 method was used to detect the cytotoxicity
induced by C2-ceramide application. The A549 and PC9 cells were
seeded in 96-well plates and treated with various ceramide
concentration gradients of 0, 20, 50, 100 and 200 µmol/l.
These were incubated in 37°C for 12, 24 and 36 h. A microplate
reader (Thermo Fisher Scientific, Inc.) was then used to detect the
absorbance in each well at 450 nm. Cell viability was calculated
based on the absorbance.
Western blot analysis
Western blot analysis was used to detect the
expression of Txnip, Trx1, p38, caspase-3 and cleaved-caspase-3
proteins. Cells were treated with 50 µmol/l C2-ceramide, 100
µmol/l verapamil and 100 µmol/l verapamil + 50
µmol/l C2-ceramide for 24 h to allow for the induction of
apoptosis. The cells were then collected, lysed cells with RIPA
Lysis Buffer (no. P0013K; Beyotime Institute of Biotechnology) on
ice, and centrifuged at 12,000 × g at 4°C for 30 min. After
measuring the protein concentration using the BCA method, the same
amount of protein (25 µg per lane for each sample) was
dissolved in the buffer and boiled for 10 min to induce
denaturation. Following separation by electrophoresis on 10%
SDS-PAGE, the proteins were transferred to a PVDF membrane,
incubated with 5% skim milk for 1.5 h, and then washed with water.
The primary antibodies [Txnip (18243-1-AP, 1:1,000; ProteinTech
Group, Inc.), Trx (NBP2-52983, 1:1,000; Novus Biologicals), p38
(ab170099, 1:2,000; Abcam), caspase-3 (ab32351, 1:2,000; Abcam) and
cleaved caspase-3 (ab2302, 1:2,000; Abcam)] were applied to the
membranes at room temperature for 1 h and the membranes were then
incubated overnight at 4°C. After washing with TBST, the
corresponding secondary antibody (ab6721,1:5000; Abcam) from the
same species was applied at room temperature for 1 h and the
membranes were washed with TBST for 30 min. Subsequently, the ECL
detection kit (P0018FS; Beyotime Institute of Biotechnology) for
used for signal development. Densitometry was performed by using
the ImageJ software (version 1.48; National Institutes of
Health).
Apoptosis detection
To further detect cell apoptosis following ceramide
treatment, the cells were seeded in a 6-well plate and treated with
DMEM, 50 µmol/l C2-ceramide, 50 µmol/l C2-ceramide +
100 µmol/l verapamil and 100 µmol/l verapamil for 24
h. They were then stained with Hoechst 33258 (Beyotime Institute of
Biotechnology) at room temperature for 5 min and observed under a
fluorescence microscope (Olympus Corporation). Apoptotic cells were
densely stained showing strong fluorescent signal in their nuclei,
while non-apoptotic cells exhibited a weak fluorescence signal. In
addition, cells treated in a similar manner, were labeled using
fluorescein fluorescein-isothiocyanate-labeled Annexin V (C1062M;
Beyotime Institute of Biotechnology) and incubated in the dark for
15 min. The cells were then stained with propidium iodide (C1062M;
Beyotime Institute of Biotechnology) for 5 min in the dark. The
apoptotic rates of the cells were determined immediately after
using flow cytometry (BD Falcon; BD Biosciences).
Caspase-3 activity detection
The treated cells were collected and washed with PBS
after centrifugation. Lysis buffer (Beyotime Biotechnology) was
added followed by shaking the samples for 15 sec every 5 min,
repeated 3 times. Following centrifugation at 4°C and 12,000 g for
15 min, the supernatant was aspirated for BCA quantification, and
an appropriate amount of detection buffer was then added together
with 10 µl caspase-3 substrate, and the samples were
incubated at 37°C for 1-4 h. A microplate reader (Thermo Fisher
Scientific, Inc.) was used to detect the absorbance in each well at
A405 nm wavelength.
Immunofluorescence
The A549 and PC9 cells were seeded on a cover glass
and treated with DMEM, 50 µmol/l C2-ceramide, 50
µmol/l C2-ceramide +100 µmol/l verapamil, and 100
µmol/l verapamil for 24 h. The cells were then fixed in 4%
paraformaldehyde for 15 min, permeabilized in 0.5% Triton X-100 for
20 min, and then blocked in bovine serum albumin at room
temperature for 30 min. The cells were then incubated with primary
antibodies to anti-Txnip (1:200; 18243-1-AP, ProteinTech Group,
Inc.) and anti-Trx (1:200; NBP2-52983, Novus Biologicals) from
different species at a 1:200 dilution at 4°C overnight. After
washing with TBST, then stained with Alexa Fluor 488-labeled goat
anti-mouse (1:500; SA00013-1, ProteinTech Group, Inc.) and
FITC-labeled goat anti-rabbit secondary antibodies (1:500;
SA00003-2, ProteinTech Group, Inc.) were added to the cells and
incubated at 37°C in the dark for 1 h. The cell nuclei were
counterstained with DAPI (Beyotime Institute of Biotechnology) for
5 min at room temperature. Images were observed using a fluorescent
confocal microscope (Olympus IX71, Olympus Corporation).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total cell RNA was extracted using TRIzol reagent,
and the RNA concentration was measured using a NanoDrop instrument
(Thermo Fisher Scientific, Inc.). cDNA synthesis was performed
using a reverse transcriptase kit (PrimeScript™ RT Reagent kit
-Perfect Real-Time; Takara Biotechnology, Inc.). qPCR was performed
using the SYBR-Green Master Mix kit (Takara Biotechnology Co.,
Ltd.) in a 10 µl system on a real-time PCR system. The
thermocycling conditions were conducted as follows: Initial
denaturation 95°C for 5 min, followed by 40 cycles of denaturation
at 95°C for 15 sec, annealing at 60°C for 30 sec and extension at
72°C for 30 sec, and a final extension at 72°C for 5 min. The
sequence of the primers used in the present study were as follows:
Txnip forward, 5′-GCC ACA CTT ACC TTG CCA AT-3′ and reverse, 5′-TGT
TGC AGC CCA GGA TAG AA-3′; Trx1 forward, 5′-GTG AAG CAG ATC GAG AGC
AAG-3′ and reverse, 5′-CGT GGC TGA GAA GTC AAC TAC TA-3′; caspase-3
forward, 5′-ATC GGA CTG TGG CAT TGA GA-3′ and reverse, 5′-ATA ACC
AGG TGC TGT GGA GT-3′; and β-actin forward, 5′-CCC TGG AGA AGA GCT
ACG AG-3′ and reverse, 5′-GGA AGG AAG GCT GGA AGA GT-3′. The
expression of each target gene was statistically compared with the
RNA level using the 2−ΔΔCq method (28), and β-actin was used as an
internal reference.
Statistical analysis
All experiments were repeated 3 times. Statistical
analysis was performed using SPSS 21.0 (IBM Corp). All data are
expressed as the means + SD, and the data were analyzed using the
Student's t-test or one-way ANOVA followed by post hoc comparisons
with Student-Newman-Keuls (S) and Tukey's post hoc tests. P<0.05
was considered to indicate a statistically significant
difference.
Results
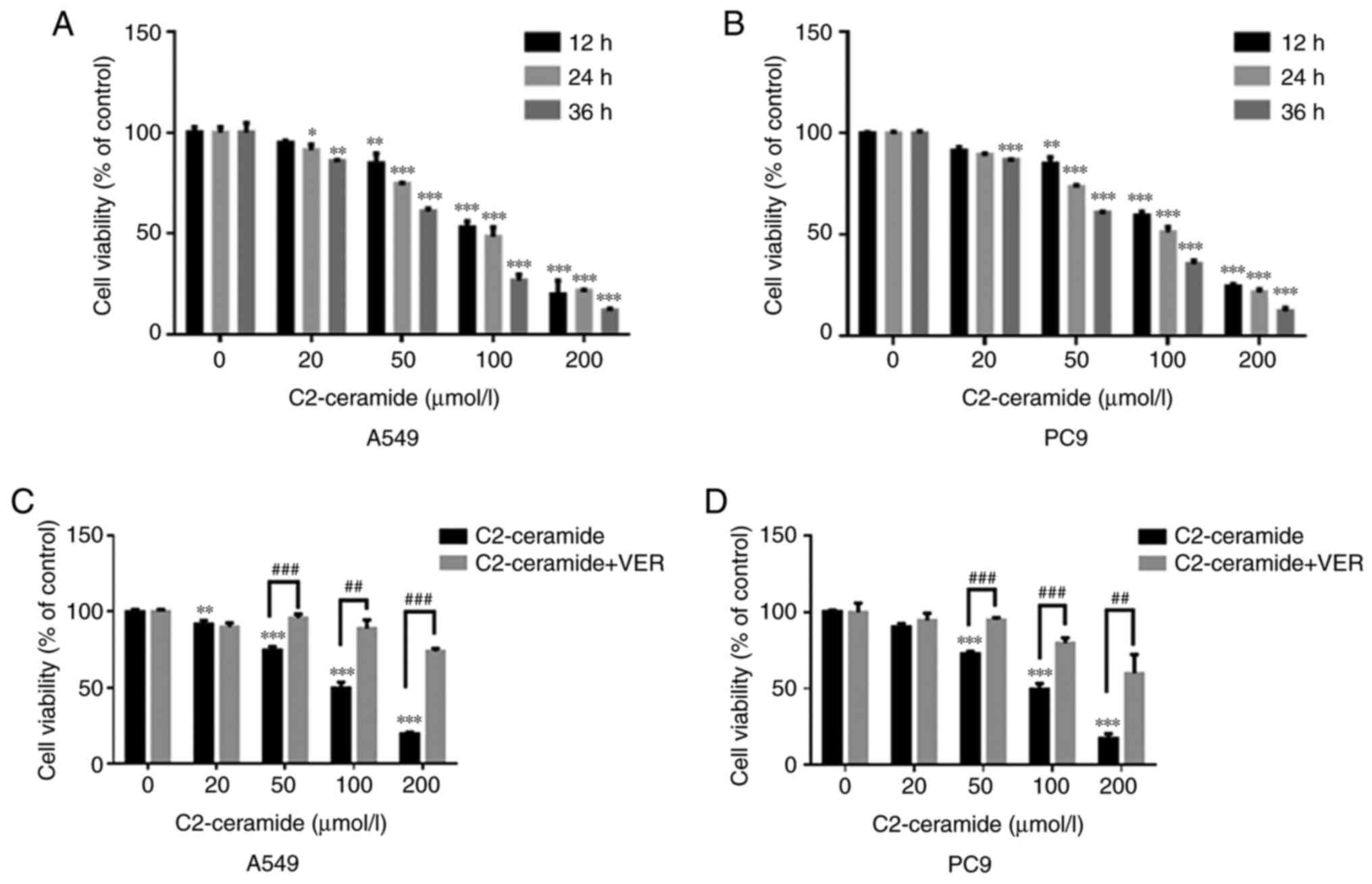
Effect of ceramide on cell viability
As shown in Fig.
1A (A549 cells) and B (PC9 cells), ceramide reduced cell
viability in a time- and concentration-dependent manner. The cell
survival rate in the treatment group was significantly lower than
that in the control group when the ceramide concentration was 50,
100 and 200 µmol/l (P<0.05). When the concentration of
the treatment group was 50 µmol/l and the treatment duration
was 24 h, the cell viability in the 2 treatment groups was
approximately 70%. However, when verapamil was incubated with
C2-ceramide, the cell viability was increased (Fig. 1C and D).
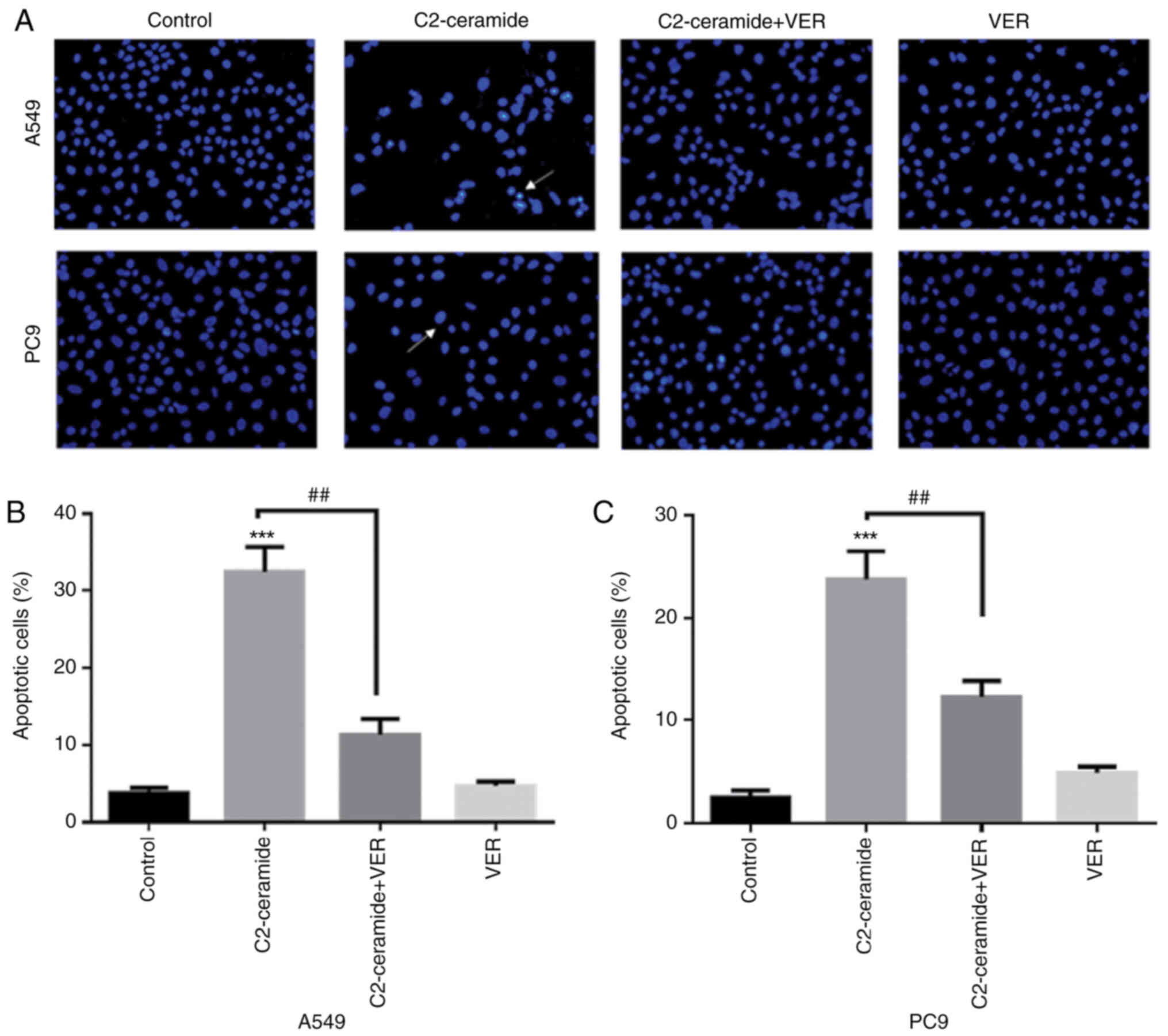
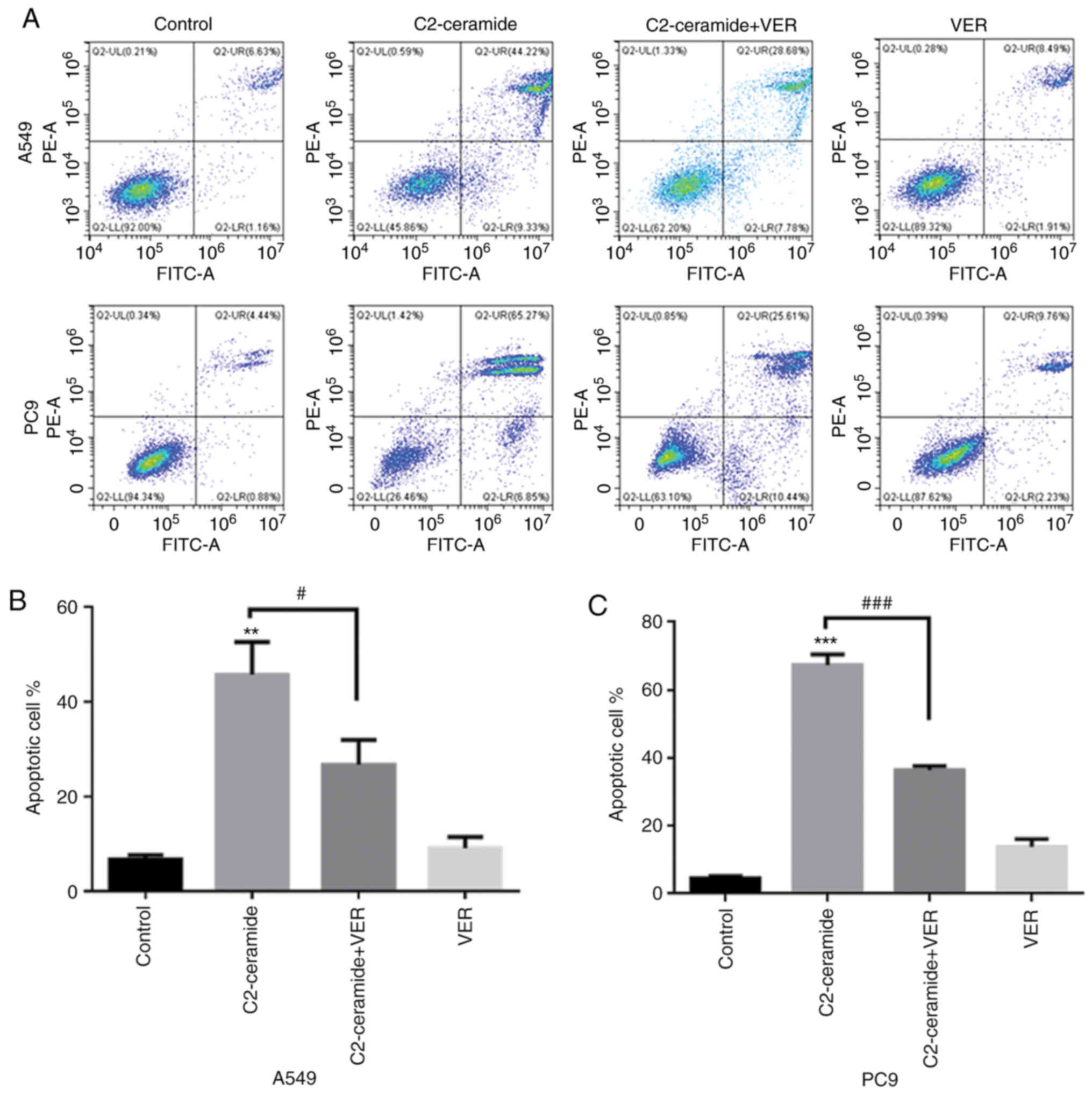
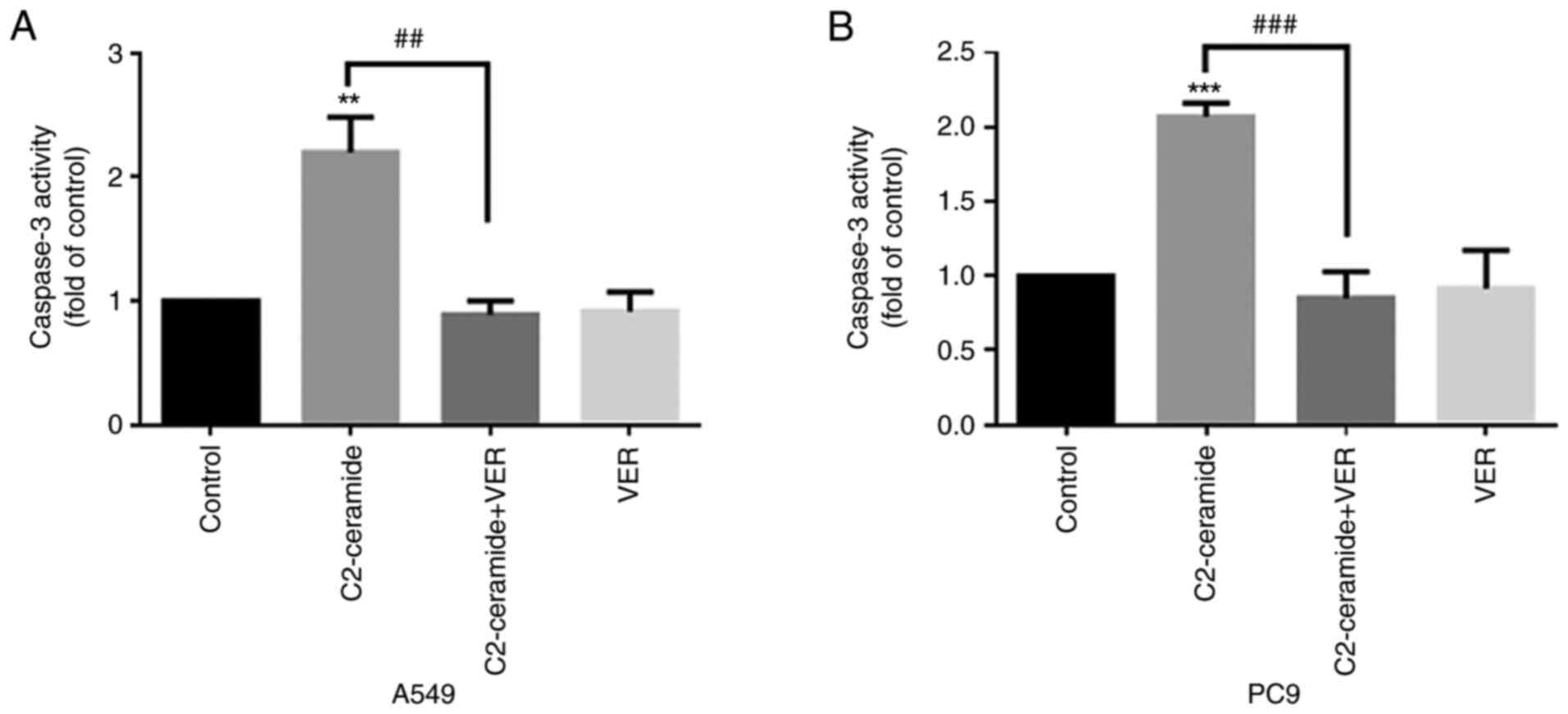
Apoptosis promoted by ceramide is
affected by verapamil
As shown in Fig.
2, the results of Hoechst 33258 staining revealed that the
apoptotic rates of the two cell lines were significantly higher in
the ceramide-treated group than in the control group. Moreover, the
use of verapamil significantly reduced the number of cells
undergoing apoptosis (P<0.05). The results of flow cytometry
revealed shown in Fig. 3 also
demonstrated similar trends (P<0.05). Furthermore, caspase-3
activity was enhanced in the ceramide group, and co-incubation with
verapamil rescued the cells from apoptosis (P<0.05), as shown in
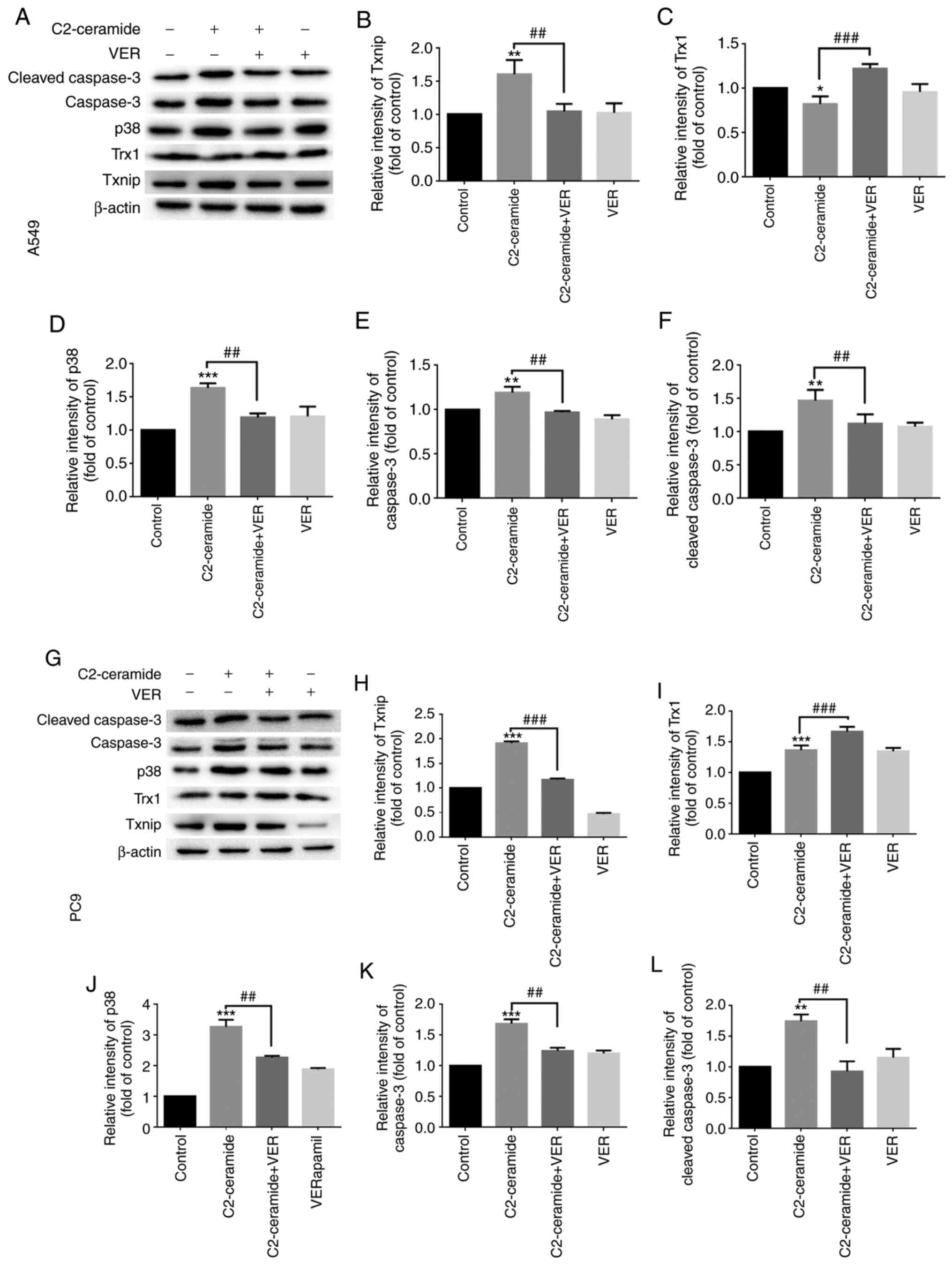
Fig. 4. The expression of
apoptosis-related proteins also exhibited a similar tendency
(Fig. 5).
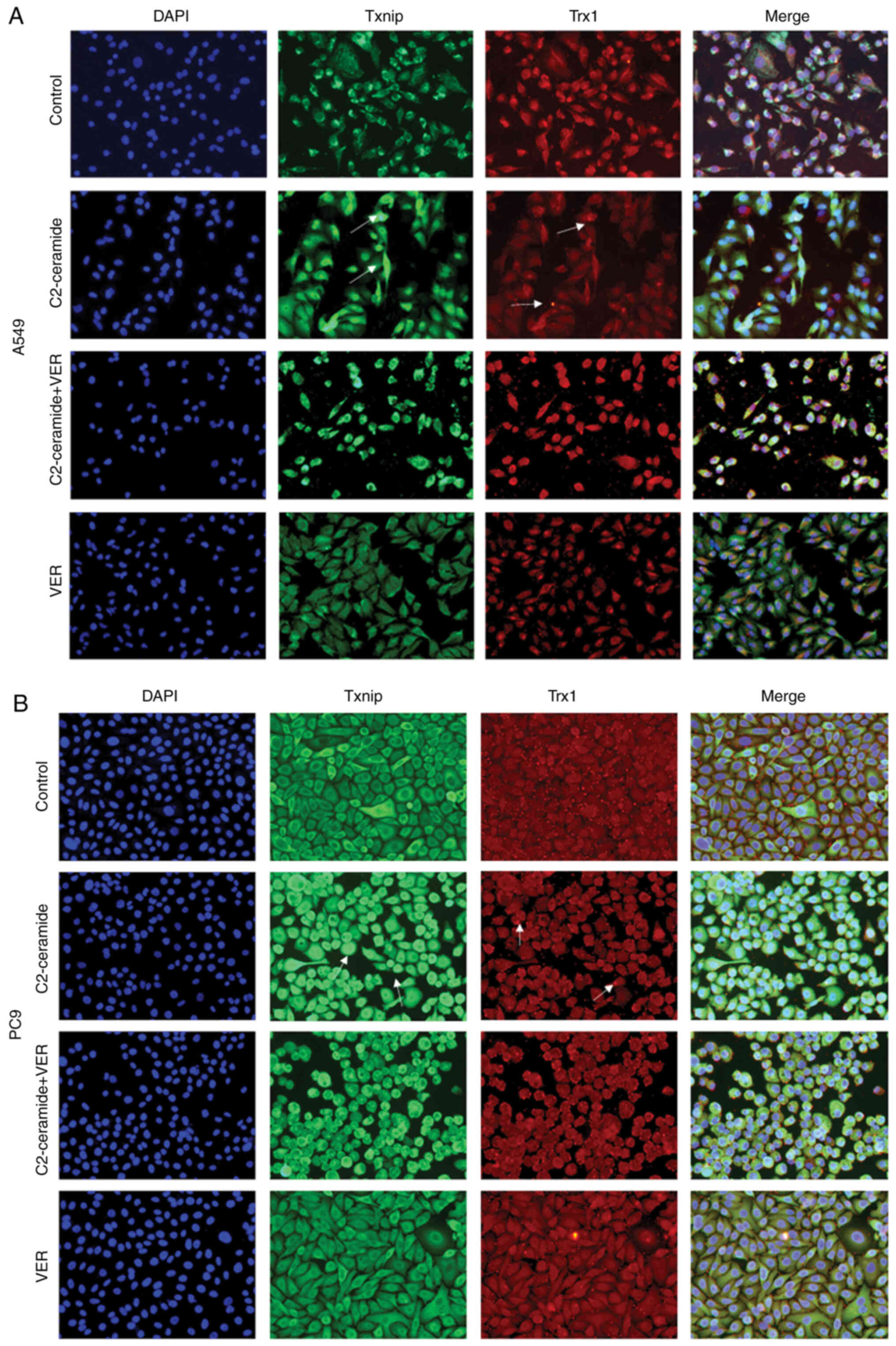
Effect of ceramide on Txnip and Trx1 on
the promotion of apoptosis
The results of immunofluorescence staining shown in
Fig. 6A (A549 cells) and B (PC9
cells) revealed that compared to the control group, the two cell
lines exhibited a significantly increased fluorescence signal
corresponding to the expression of Txnip in the cytoplasm. Txnip is
typically located in the nucleus, and Txnip translocation occurred
in response to ceramide treatment. This indicates that Txnip reacts
through the cell membrane during apoptosis. Following co-incubation
with verapamil, the expression of Txnip in the cytoplasm decreased.
This trend was inversely related to the expression of Trx1. The
protein expression level also exhibited similar results, as shown
in Fig. 5.
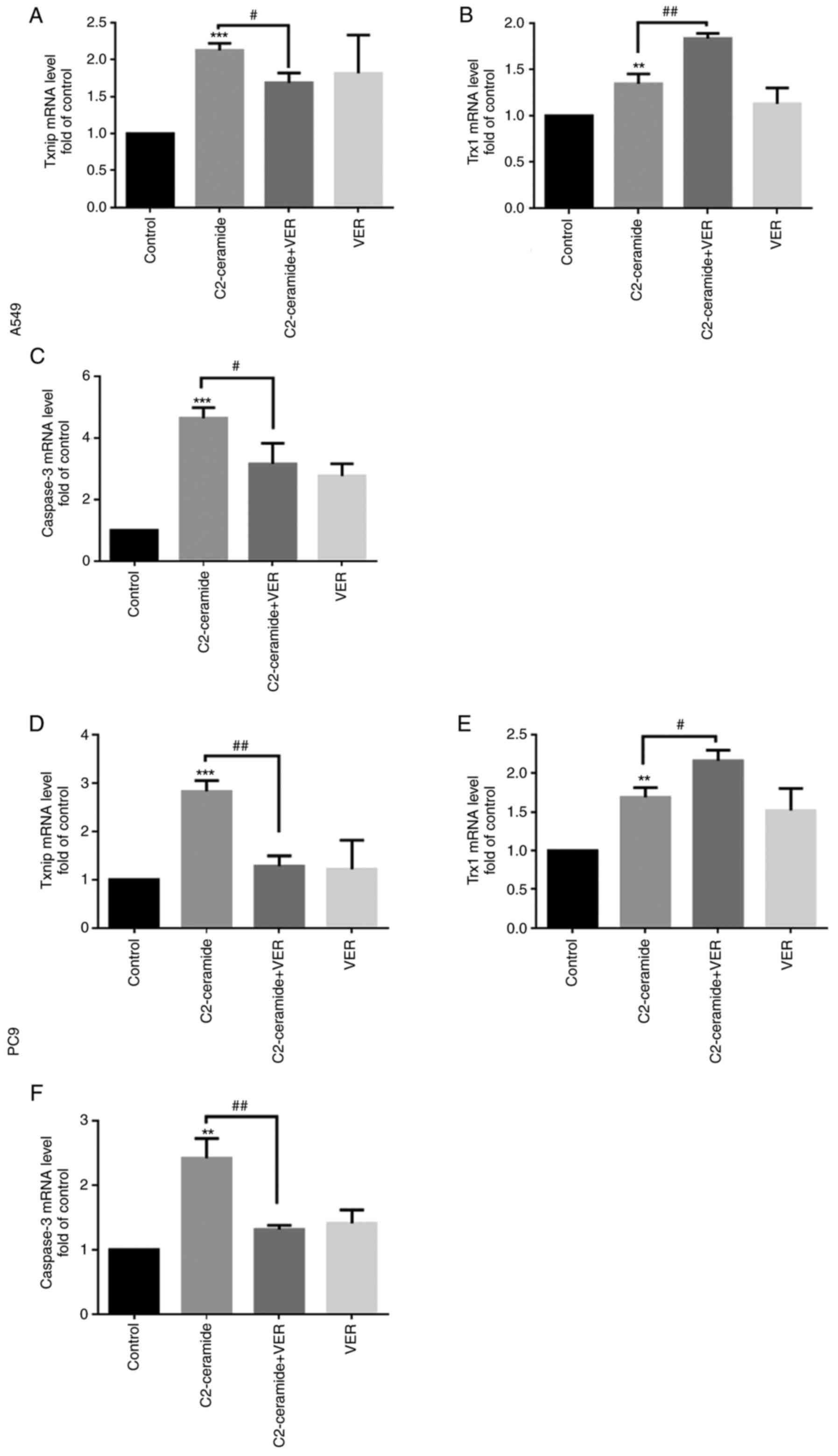
Expression of Txnip, Trx1 and caspase-3
at the mRNA level
As shown in Fig.
7, compared to the control group, the Txnip expression level
(Fig. 7A and D) was
significantly increased after the two cell lines were treated with
50 µmol/l C2-ceramide, and the Trx1 level (Fig. 7B and E) was significantly
decreased (P<0.05). Compared to the control group, the
expression levels of apoptotic proteins were also increased
(Fig. 7C and F). Following
co-incubation with 100 µmol/l verapamil, this pattern of
expression was significantly reversed (P<0.05).
Discussion
Studies have demonstrated that ceramide is closely
related to apoptosis and cancer. Ceramide inducers can enhance the
sensitivity to anticancer drugs (7,12,29). In previous studies (30,31), the authors demonstrated that
ceramide was significantly increased in in mice with
lipopolysaccharide-induced ALI, and the survival rate in mice was
reduced. The use of the aSmase inhibitor, imipramine, reduced lung
injury. However, the specific pathway through which ceramide
induces lung cancer cell apoptosis is not yet fully understood. In
the pre-experiment before this research (data not shown), the
H1299, H1975, A549 and PC9 cell lines were tested and it was found
that the A549 and PC9 cells had the most significant drug response
to ceramide treatment. Therefore, in the present study, the A549
and PC9 cell lines were used in the experiments and it was proven
that ceramide induced non-small cell lung cancer apoptosis through
the Txnip/Trx1 complex.
Lung cancer is the main cause of cancer-related
mortality worldwide, and the average 5-year survival rate is only
19% (32,33). Although chemotherapy is the main
treatment method, its side-effects pose a significant challenge.
Therefore, alternative chemotherapeutic drugs and those that reduce
the side-effects of chemotherapy are required (34). Studies have demonstrated that
sphingolipid metabolism-targeted therapy may have broad prospects
in oncological clinical application. The regulatory effects of
ceramide on chemotherapeutics have become the focus of research in
recent years (35,36). The human lung adenocarcinoma cell
lines, A549 and PC9, are non-small cell lung cancer cell lines. The
present study revealed that ceramide reduced cell viability in a
time- and concentration-dependent manner. Following treatment with
50 µmol/l ceramide, caspase-3 activity increased, and the
levels of apoptosis-related proteins, such as Txnip and Trx1 were
also altered accordingly. This indicated that Txnip and Trx1 were
downstream of ceramide during the induction of cell apoptosis.
Txnip is a tumor suppressor gene and an endogenous
inhibitor of Trx. Usually, Txnip binds to Trx1 in a redox-dependent
manner and participates in various redox reactions (27,37,38). Previous research has demonstrated
that the activation of the Txnip/Trx1 complex increases oxidative
stress and inflammation, thereby aggravating the symptoms of
diabetes; this finding proved that the upregulation of Txnip or
activation of the Txnip/Trx1 complex plays an important role in
aggravating diabetic myocardial damage (39). The present study also
demonstrated, by immunofluorescence staining, that Txnip induced by
C2-ceramide co-localized with Trx1. Txnip is mainly localized in
the nucleus, while Trx1 is mainly distributed in the cytoplasm. The
fluorescent expression of Txnip was significantly increased, while
Trx1 expression was reduced. Verapamil treatment reversed this
expression-dose association. The decreased Trx1 activity was mainly
caused by the increase in Txnip; however, other factors that can
influence this process may be involved as well (19). It has been demonstrated that
radiation, hydrogen peroxide, ultraviolet light and growth
inhibition stimulates Txnip production (40-42). p38 MAPK is a protein kinase that
responds to stress stimuli inside and outside the cell. It plays an
important role in regulating cell differentiation, proliferation,
survival and death. Studies have proven the central role of p38
MAPK in inflammation (43,44). In the present study, when
verapamil was used to inhibit ceramide-induced Txnip expression,
the expression of p38 protein was decreased. It can thus be
inferred that the change in p38 protein expression was affected by
Txnip protein, which acts upstream of p38. However, further
experiments are required to confirm the accuracy of this
interpretation.
In conclusion, the present study investigated the
anticancer mechanisms of sphingolipid ceramide action. It was found
that it induced lung cancer cell apoptosis through the Txnip/Trx1
complex, which plays an important role in biological activities,
such as oxidative stress. Ceramide has great potential and is
expected to become a prospective candidate in the development of
therapies for cancer. In addition, research on various targets in
the Txnip/Trx1 complex may provide new possibilities for cancer
treatment. However, the present study also had limitations. First,
the present study was limited to the experiments performed at the
cellular level in vitro. The human body is more complex and
numerous factors are involved. Thus, the same effect of ceramide on
the human body as that observed in cell culture cannot be
guaranteed. Second, the present study selected only one
representative type of ceramide. Differences across the other types
of ceramide are unclear. Furthermore, only used two types of lung
adenocarcinoma cells were used. The effect of ceramide on other
types of lung cancer cells thus warrant further investigation.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YS and YJ designed the study, performed the
immunofluorescence assay and drafted the manuscript. JJ and FL
designed the study, performed the cell viability assays and
analyzed the data. YS was responsible for cell culture. YS and YJ
performed the western blot analysis. YL, JY and JC were involved in
the study design, quality control, manuscript drafting and
coordination. YS and YJ confirm the authenticity of all the raw
data. All authors have read and approved the final draft.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 81400058) and the Anhui Province
Science and Technology Tackling Plan Project (no. 1401045016).
Abbreviations:
|
Txnip
|
thioredoxin-interacting protein
|
|
Trx
|
thioredoxin
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
CCK-8
|
Cell Counting kit-8
|
References
|
1
|
Remark R, Becker C, Gomez JE, Damotte D,
Dieu-Nosjean MC, Sautès-Fridman C, Fridman WH, Powell CA, Altorki
NK, Merad M and Gnjatic S: The non-small cell lung cancer immune
contexture. A major determinant of tumor characteristics and
patient outcome. Am J Respir Crit Care Med. 191:377–390. 2015.
View Article : Google Scholar
|
|
2
|
Skřičková J, Kadlec B, Venclíček O and
Merta Z: Lung cancer. Cas Lek Cesk. 157:226–236. 2018.
|
|
3
|
Misra P and Singh S: Role of cytokines in
combinatorial immunotherapeutics of non-small cell lung cancer
through systems perspective. Cancer Med. 8:1976–1995. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Naylor EC, Desani JK and Chung PK:
Targeted therapy and immunotherapy for lung cancer. Surg Oncol Clin
N Am. 25:601–609. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Owen DH, Williams TM, Bertino EM, Mo X,
Webb A, Schweitzer C, Liu T, Roychowdhury S, Timmers CD and
Otterson GA: Homologous recombination and DNA repair mutations in
patients treated with carboplatin and nab-paclitaxel for metastatic
non-small cell lung cancer. Lung Cancer. 134:167–173. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuş G, Özkurt M, Öztopcu Vatan P, Erkasap
N, Uyar R and Kabadere S: Comparison of a ceramidase inhibitor
(ceranib-2) with C2 ceramide and cisplatin on cytotoxicity and
apoptosis of glioma cells. Turk J Biol. 42:259–265. 2018.
|
|
7
|
Ordoñez R, Fernández A, Prieto-Domínguez
N, Martínez L, García-Ruiz C, Fernández-Checa JC, Mauriz JL and
González-Gallego J: Ceramide metabolism regulates autophagy and
apoptotic cell death induced by melatonin in liver cancer cells. J
Pineal Res. 59:178–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kolesnick R and Fuks Z: Radiation and
ceramide-induced apoptosis. Oncogene. 22:5897–5906. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brodowicz J, Przegaliński E, Müller CP and
Filip M: Ceramide and its related neurochemical networks as targets
for some brain disorder therapies. Neurotox Res. 33:474–484. 2018.
View Article : Google Scholar :
|
|
10
|
Chang YC, Fong Y, Tsai EM, Chang YG, Chou
HL, Wu CY, Teng YN, Liu TC, Yuan SS and Chiu CC: Exogenous
C8-ceramide induces apoptosis by overproduction of ROS
and the switch of superoxide dismutases SOD1 to SOD2 in human lung
cancer cells. Int J Mol Sci. 19:30102018. View Article : Google Scholar
|
|
11
|
Ma JQ, Liu CM and Yang W: Protective
effect of rutin against carbon tetrachloride-induced oxidative
stress, inflammation and apoptosis in mouse kidney associated with
the ceramide, MAPKs, p53 and calpain activities. Chem Biol
Interact. 286:26–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maeng HJ, Song JH, Kim GT, Song YJ, Lee K,
Kim JY and Park TS: Celecoxib-mediated activation of endoplasmic
reticulum stress induces de novo ceramide biosynthesis and
apoptosis in hepatoma HepG2 cells mobilization. BMB Rep.
50:144–149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang SW, Hojabrpour P, Zhang P, Kolesnick
RN, Steinbrecher UP, Gómez-Muñoz A and Duronio V: Regulation of
ceramide generation during macrophage apoptosis by ASMase and de
novo synthesis. Biochim Biophys Acta. 1851:1482–1489. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai L, Smith CD, Foroozesh M, Miele L and
Qin Z: The sphingosine kinase 2 inhibitor ABC294640 displays
anti-non-small cell lung cancer activities in vitro and in vivo.
Int J Cancer. 142:2153–2162. 2018. View Article : Google Scholar :
|
|
15
|
Chang WT, Wu CY, Lin YC, Wu MT, Su KL,
Yuan SS, Wang HD, Fong Y, Lin YH and Chiu CC: C(2)-ceramide-induced
Rb-dominant senescence-like phenotype leads to human breast cancer
MCF-7 escape from p53-dependent cell death. Int J Mol Sci.
20:42922019. View Article : Google Scholar
|
|
16
|
Rahman A, Pallichankandy S, Thayyullathil
F and Galadari S: Critical role of H(2)O(2) in mediating
sanguinarine-induced apoptosis in prostate cancer cells via
facilitating ceramide generation, ERK1/2 phosphorylation, and Par-4
cleavage. Free Radic Biol Med. 134:527–544. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yildiz-Ozer M, Oztopcu-Vatan P and Kus G:
The investigation of ceranib-2 on apoptosis and drug interaction
with carboplatin in human non small cell lung cancer cells in
vitro. Cytotechnology. 70:387–396. 2018. View Article : Google Scholar :
|
|
18
|
Suzuki M, Cao K, Kato S, Komizu Y,
Mizutani N, Tanaka K, Arima C, Tai MC, Yanagisawa K, Togawa N, et
al: Targeting ceramide synthase 6-dependent metastasis-prone
phenotype in lung cancer cells. J Clin Invest. 126:254–265. 2016.
View Article : Google Scholar
|
|
19
|
Xu L, Lin X, Guan M, Zeng Y and Liu Y:
Verapamil attenuated prediabetic neuropathy in high-fat diet-fed
mice through inhibiting TXNIP-mediated apoptosis and inflammation.
Oxid Med Cell Longev. 2019:18960412019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kizhakkayil J, Thayyullathil F, Chathoth
S, Hago A, Patel M and Galadari S: Glutathione regulates
caspase-dependent ceramide production and curcumin-induced
apoptosis in human leukemic cells. Free Radic Biol Med.
52:1854–1864. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu W, Wang L, Li J, Cai Y and Xue Y: TXNIP
mediated the oxidative stress response in glomerular mesangial
cells partially through AMPK pathway. Biomed Pharmacother.
107:785–792. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lv H, Zhu C, Wei W, Lv X, Yu Q, Deng X and
Ci X: Enhanced Keap1-Nrf2/Trx-1 axis by daphnetin protects against
oxidative stress-driven hepatotoxicity via inhibiting ASK1/JNK and
Txnip/NLRP3 inflammasome activation. Phytomedicine. 71:1532412020.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu Y, Xing K, Badamas R, Kuszynski CA, Wu
H and Lou MF: Overexpression of thioredoxin-binding protein 2
increases oxidation sensitivity and apoptosis in human lens
epithelial cells. Free Radic Biol Med. 57:92–104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Muri J, Heer S, Matsushita M, Pohlmeier L,
Tortola L, Fuhrer T, Conrad M, Zamboni N, Kisielow J and Kopf M:
The thioredoxin-1 system is essential for fueling DNA synthesis
during T-cell metabolic reprogramming and proliferation. Nat
Commun. 9:18512018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ungerstedt J, Du Y, Zhang H, Nair D and
Holmgren A: In vivo redox state of human thioredoxin and redox
shift by the histone deacetylase inhibitor suberoylanilide
hydroxamic acid (SAHA). Free Radic Biol Med. 53:2002–2007. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cohen-Kutner M, Khomsky L, Trus M,
Ben-Yehuda H, Lenhard JM, Liang Y, Martin T and Atlas D:
Thioredoxin-mimetic peptide CB3 lowers MAPKinase activity in the
Zucker rat brain. Redox Biol. 2:447–456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hou Y, Wang Y, He Q, Li L, Xie H, Zhao Y
and Zhao J: Nrf2 inhibits NLRP3 inflammasome activation through
regulating Trx1/TXNIP complex in cerebral ischemia reperfusion
injury. Behav Brain Res. 336:32–39. 2018. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Morad SA, Davis TS, MacDougall MR, Tan SF,
Feith DJ, Desai DH, Amin SG, Kester M, Loughran TP Jr and Cabot MC:
Role of P-glycoprotein inhibitors in ceramide-based therapeutics
for treatment of cancer. Biochem Pharmacol. 130:21–33. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu H, Yang J, Wang Y, Wei Y, Cao J and Lu
Y: C8-ceramide induces apoptosis of alveolar type II epithelial
cells. Chinese J Tuberculosis Respir Zhonghua Jie He He Hu Xi Za
Zhi. 38:445–450. 2015.In Chinese.
|
|
31
|
Yang J, Wang Y, Liu H, Bi J and Lu Y:
C2-ceramide influences alveolar epithelial barrier function by
downregulating Zo-1, occludin and claudin-4 expression. Toxicol
Mech Methods. 27:293–297. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mattiuzzi C and Lippi G: Current cancer
epidemiology. J Epidemiol Glob Health. 9:217–222. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Du A, Jiang Y and Fan C: NDRG1
downregulates ATF3 and inhibits cisplatin-induced cytotoxicity in
lung cancer A549 cells. Int J Med Sci. 15:1502–1507. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ogretmen B: Sphingolipid metabolism in
cancer signalling and therapy. Nat Rev Cancer. 18:33–50. 2018.
View Article : Google Scholar :
|
|
36
|
Che J, Huang Y, Xu C and Zhang P:
Increased ceramide production sensitizes breast cancer cell
response to chemotherapy. Cancer Chemother Pharmacol. 79:933–941.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
World C, Spindel ON and Berk BC:
Thioredoxin-interacting protein mediates TRX1 translocation to the
plasma membrane in response to tumor necrosis factor-α: A key
mechanism for vascular endothelial growth factor receptor-2
transactivation by reactive oxygen species. Arterioscler Thromb
Vasc Biol. 31:1890–1897. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bharti V, Tan H, Zhou H and Wang JF: Txnip
mediates glucocorticoid-activated NLRP3 inflammatory signaling in
mouse microglia. Neurochem Int. 131:1045642019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hou R, Shen M, Wang R, Liu H, Gao C, Xu J,
Tao L, Yin Z and Yin T: Thioredoxin1 inactivation mediates the
impairment of ischemia-induced angiogenesis and further injury in
diabetic myocardium. J Vasc Res. 57:76–85. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yoshihara E: TXNIP/TBP-2: A master
regulator for glucose homeostasis. Antioxidants (Basel). 9:7652020.
View Article : Google Scholar
|
|
41
|
Nishiyama A, Matsui M, Iwata S, Hirota K,
Masutani H, Nakamura H, Takagi Y, Sono H, Gon Y and Yodoi J:
Identification of thioredoxin-binding protein-2/vitamin D(3)
up-regulated protein 1 as a negative regulator of thioredoxin
function and expression. J Biol Chem. 274:21645–21650. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, De Keulenaer GW and Lee RT:
Vitamin D(3)-up-regulated protein-1 is a stress-responsive gene
that regulates cardiomyocyte viability through interaction with
thioredoxin. J Biol Chem. 277:26496–26500. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li D, Ren W, Jiang Z and Zhu L: Regulation
of the NLRP3 inflammasome and macrophage pyroptosis by the p38 MAPK
signaling pathway in a mouse model of acute lung injury. Mol Med
Rep. 18:4399–4409. 2018.PubMed/NCBI
|
|
44
|
Song W, Wei L, Du Y, Wang Y and Jiang S:
Protective effect of ginsenoside metabolite compound K against
diabetic nephropathy by inhibiting NLRP3 inflammasome activation
and NF-κB/p38 signaling pathway in high-fat
diet/streptozotocin-induced diabetic mice. Int Immunopharmacol.
63:227–238. 2018. View Article : Google Scholar : PubMed/NCBI
|