Introduction
Cardiovascular disorders pose a major health burden
and are associated with high morbidity and mortality rates
worldwide (1). Hypoxia/ischemia
is a major and crucial feature of various cardiac pathologies,
including congenital heart disease, myocardial infarction and
ischemic heart disease (2-4).
Therefore, effective prevention and advances in myocardial
protective therapy are necessary for the improvement of myocardial
ischemic disease. Naringin is a non-toxic natural bioflavonoid
found in citrus fruit peel with multiple pharmacological
properties, including anticancer, antioxidant, anti-apoptotic,
anti-inflammatory, anti-diabetic, neuroprotective and
cardioprotective functions (5-8).
Recently, the cardioprotective properties of naringin in various
cardiovascular disease models have attracted increasing attention
(9,10). Naringin has been shown to protect
H9C2 myocardial cells against doxorubicin-induced cardiotoxicity
and apoptosis (11). Naringin
has also been shown to attenuate anoxia/reoxygenation-induced
injury by inhibiting apoptosis and oxidative stress, which is
dependent on the Nrf2 signaling pathway (12). However, the cardioprotective
effects of naringin on ischemic injury and the underlying
mechanisms are not yet fully understood.
Autophagy, a lysosome-mediated degradation pathway,
is involved in major areas of cardiovascular disorders; however, it
is still contested whether autophagy exerts a protective effect on
cell survival or whether it results in cell death (13-15). Recent studies have found that the
dysfunction of autophagy plays a vital role in the development and
progression of myocardial damage (16,17). Autophagy is a vital degradation
process required to maintain intracellular homeostasis under
conditions of hypoxic/ischemic injury (18,19). It has been confirmed that hypoxia
enhances the flux of autophagosome formation, and restoring
autophagy function is thus a novel promising therapeutic option for
managing ischemic injury (13,20). Hypoxia-inducible factor-1α
(HIF-1α) is the main regulator of the cellular response to hypoxia
(21). HIF-1α is closely
associated with the regulation of myocardial survival by activating
the downstream protein, BCL2 interacting protein 3 (BNIP3)
(22). Notably, studies have
demonstrated that BNIP3 is involve in regulating autophagy
(23,24). Recently, studies have revealed
that the hypoxic/ischemic condition promotes HIF-1α expression, and
the HIF-1α/BNIP3 signaling pathway also plays essential roles in
hypoxia-induced autophagy (23-25). In addition, studies have
confirmed that naringin exerts beneficial effects by regulating
autophagy (14,26). Hence, the present study aimed to
further investigate whether naringin attenuates ischemic injury via
modulating autophagy and the HIF-1α/BNIP3 signaling pathway.
Cobalt chloride (CoCl2), a type of
chemical hypoxia mimetic agent (27), is widely used to mimic
hypoxic/ischemic conditions, triggering cell damage and stabilizing
intracellular HIF-1α expression, and is a key regulator in the
adaptation to a low concentration of oxygen and cellular survival
(28). H9C2 embryonic rat
cardiac cells are derived from rat embryonic hearts and are widely
used as in vitro models for exploring the molecular
mechanisms or protective strategies underlying cardiovascular
disorders (29). Therefore,
CoCl2-treated H9C2 cells were used as a hypoxic model in
in vitro in the present study. The present study explored
the protective effects of naringin against CoCl2 injury
and further investigated the underlying mechanisms, focusing on the
autophagic flux and HIF-1α/BNIP3 signaling pathway in H9C2 cells.
The findings of the present study may provide novel scientific
evidence in favor of the use of naringin for the prevention of
hypoxic/ischemic injury, and may highlight potentially novel
targets for the management of hypoxic/ischemic injury.
Materials and methods
Cell culture and treatment
Rat H9C2 cardiomyocytes were obtained from the
American Tissue Culture Collection (ATCC). Cells were maintained in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
(v/v) FBS (Gibco; Thermo Fisher Scientific, Inc.) and 100 U/ml
penicillin-streptomycin (Beyotime Institute of Biotechnology) in an
incubator (Thermo Fisher Scientific Inc.) with 5% CO2 at
37°C.
To mimic hypoxic conditions, H9C2 cells were treated
with various concentrations of CoCl2 (200, 400, 600, 800
or 1,000 µmol/l; Sigma-Aldrich; Merck KGaA) for different
periods of time (0, 1.5, 3, 6, 12 and 24 h). To investigate the
protective effects of naringin (Sigma-Aldrich; Merck KGaA) on
CoCl2 injury, H9C2 cells were pre-treated with naringin
(5, 10, 20 and 40 µmol/l) for 1 h prior to treatment with
CoCl2 (400 µmol/l) for 12 h. Cells incubated in
normoxic medium (DMEM, 10% FBS, and 1% penicillin-streptomycin
solution) in an incubator with 5% CO2 at 37°C served as
the control group.
Treatment with 3-methyladenine (3-MA),
bafilomycin A1 (Baf A1) or
3-(5′-hydroxymethyl-2′-furyl)-1-benzylindazole (YC-1)
3-MA and Baf A1 were obtained from Santa Cruz
Biotechnology, Inc. YC-1 was purchased from Cayman Chemical Co. All
reagents were dissolved in DMEM containing 1% DMSO. To inhibit
autophagy, H9C2 cells were incubated with 3-MA (5 mM) for 1 h prior
to treatment with CoCl2 (400 µmol/l) for 12 h in
the presence or absence of naringin (20 µmol/l). To block
autophagosome fusion with lysosomes, H9C2 cells were treated with
Baf A1 (10 nmol/l) and CoCl2 (400 µmol/l) for 12
h in the presence or absence of naringin (20 µmol/l). To
inhibit the HIF-1α/BNIP3 signaling pathway, H9C2 cells were treated
with YC-1 (10 µM) for 1 h prior to treatment with naringin
(20 µmol/l) for 2 h followed by co-treatment with
CoCl2.
Cell Counting kit-8 (CCK-8) assay
Cell viability was assessed using a CCK-8 kit
(Beijing Solarbio Science & Technology Co., Ltd.) according to
the manufacturer's protocol. Briefly, H9C2 cells were added to
96-well plates (5×103 cells/well) and cultured.
Following treatment as described above, the media from each group
were replaced with serum-free DMEM (90 µl) plus CCK-8 regent
(10 µl), and incubated for 3 h at 37°C. The absorbance was
measured at 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.). Cell viability as a percentage was calculated by
comparing the value of the treated group to that of the control
group.
Total lactate dehydrogenase (LDH) release
assay
Cell injury was assessed based on the amount of
total LDH released into the culture supernatant, measured using an
LDH kit (Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. Following centrifugation at 1,000 × g, the
supernatant was collected and plated in 96-well plates. The
absorbance was measured using a microplate reader (Thermo Fisher
Scientific, Inc.) at 450 nm.
Annexin V-FITC/PI double staining
assay
The apoptosis of H9C2 cells was measured using an
Annexin V-FITC/PI apoptosis detection kit (Nanjing Keygen Biotech
Co., Ltd.) according to the manufacturer's protocol. H9C2 cells
were seeded in 6-well plates (1×106 cells/well), and
treated as described above. After washing with PBS twice, the cells
were re-suspended in Annexin V-binding buffer (500 µl) and
stained with FITC-labelled Annexin V (5 µl) and PI (10
µl) for 15 min in the dark at 37°C. The rate of apoptosis
was quantified using a flow cytometer (BD LSRFortessa X-20; BD
Biosciences).
Caspase-3/caspase-9 activities assay
The H9C2 cells were seeded in 6-well plates at a
density of 1×106 per well. Following treatment as
described above, the cells were sonicated, centrifuged at 12,000 ×
g for 10 min at 4°C, and the supernatant was retained. The
concentration of proteins was determined using a bicinchoninic acid
kit (Thermo Fisher Scientific, Inc.). The activities of caspase-3
and caspase-9 was determined using a Caspase-3 Activity Assay kit
(Beyotime Institute of Biotechnology) and Caspase-9 Activity Assay
kit (Beyotime Institute of Biotechnology) in accordance with the
manufacturer's protocol. Caspase-3 and caspase-9 activities were
determined by cleavage of the Ac-DEVD-pNA and Ac-LEHD-pNA
substrates, respectively, and the absorbance was measured at 405
nm. The data are presented as fold increases over the pre-treatment
levels.
Western blot analysis
The treated H9C2 cells were lysed using RIPA lysis
buffer (Beyotime Institute of Biotechnology) containing 1 mM PMSF
(Beyotime Institute of Biotechnology) and centrifuged at 12,000 × g
for 10 min at 4°C. The concentration of proteins was determined
using a BCA kit. Equivalent quantities of proteins (40 µg)
were loaded on a 10% SDS-gel, resolved using SDS-PAGE and
transferred to PVDF membranes (EMD Millipore). After blocking with
5% non-fat milk in TBS containing 0.01% Tween-20 (TBST) overnight
at 4°C, the membranes were incubated with the following primary
antibodies all at a dilution of 1:2,000: Rabbit monoclonal
anti-Beclin1 (#3495), rabbit polyclonal LC3B-II/I (#3868), rabbit
polyclonal anti-p62 (#39749), rabbit polyclonal anti-HIF-1α
(#14179), rabbit polyclonal anti-BNIP3 (#44060) and rabbit
polyclonal anti-tubulin (#2128) at 4°C overnight. All antibodies
were purchased from Cell Signaling Technology, Inc. After washing
twice with TBST, the membranes were incubated with HRP-conjugated
goat anti-rabbit secondary antibody (1:5,000; #7074; Cell Signaling
Technology, Inc.) at room temperature for 2 h. Finally, the bands
were visualized using enhanced chemiluminescence reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Densitometric
analysis was performed using the Bio-Rad Microscopic Imaging system
and Odyssey Image software (LICOR).
Statistical analysis
All data were analyzed using GraphPad Prism version
6 (GraphPad Software Inc.). Data are expressed as the means ±
standard deviation. Differences between multiple groups were
compared using a one-way ANOVA followed by a post-hoc Tukey's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Naringin inhibits
CoCl2-induced injury to H9C2 cells
To investigate the toxic effects of CoCl2
on H9C2 cells, the cells were incubated with various concentrations
of CoCl2 (200, 400, 600, 800 and 1,000 µmol/l)
for different periods of time (1.5, 3, 6, 12 and 24 h). The results
of CCK-8 assay revealed that CoCl2 significantly reduced
cell viability in a concentration- and time-dependent manner
(Fig. 1A). CoCl2 (400
µmol/l) treatment for 12 h significantly decreased cell
viability (P<0.01) to 40% of the control; thus, this
concentration and treatment duration were used in subsequent
experiments. Subsequently, the protective effects of naringin on
CoCl2-induced toxicity to H9C2 cells were assessed.
CCK-8 assay revealed that treatment with various concentrations of
naringin (5, 10, 20 or 40 µmol/l) alone for 24 h did not
exert any cytotoxic effects on the cells (Fig. 1B), and naringin (10, 20 and 40
µmol/l) pre-treatment reversed the CoCl2-induced
decrease in cell viability (Fig.
1C). Pre-treatment with naringin at 20 µmol/l
significantly reversed the CoCl2-induced decrease in
cell viability (P<0.01), and thus this concentration was used in
subsequent experiments to explore the protective effects of
naringin against CoCl2 injury. These results indicated
that naringin protected the H9C2 cells against
CoCl2-induced injury.
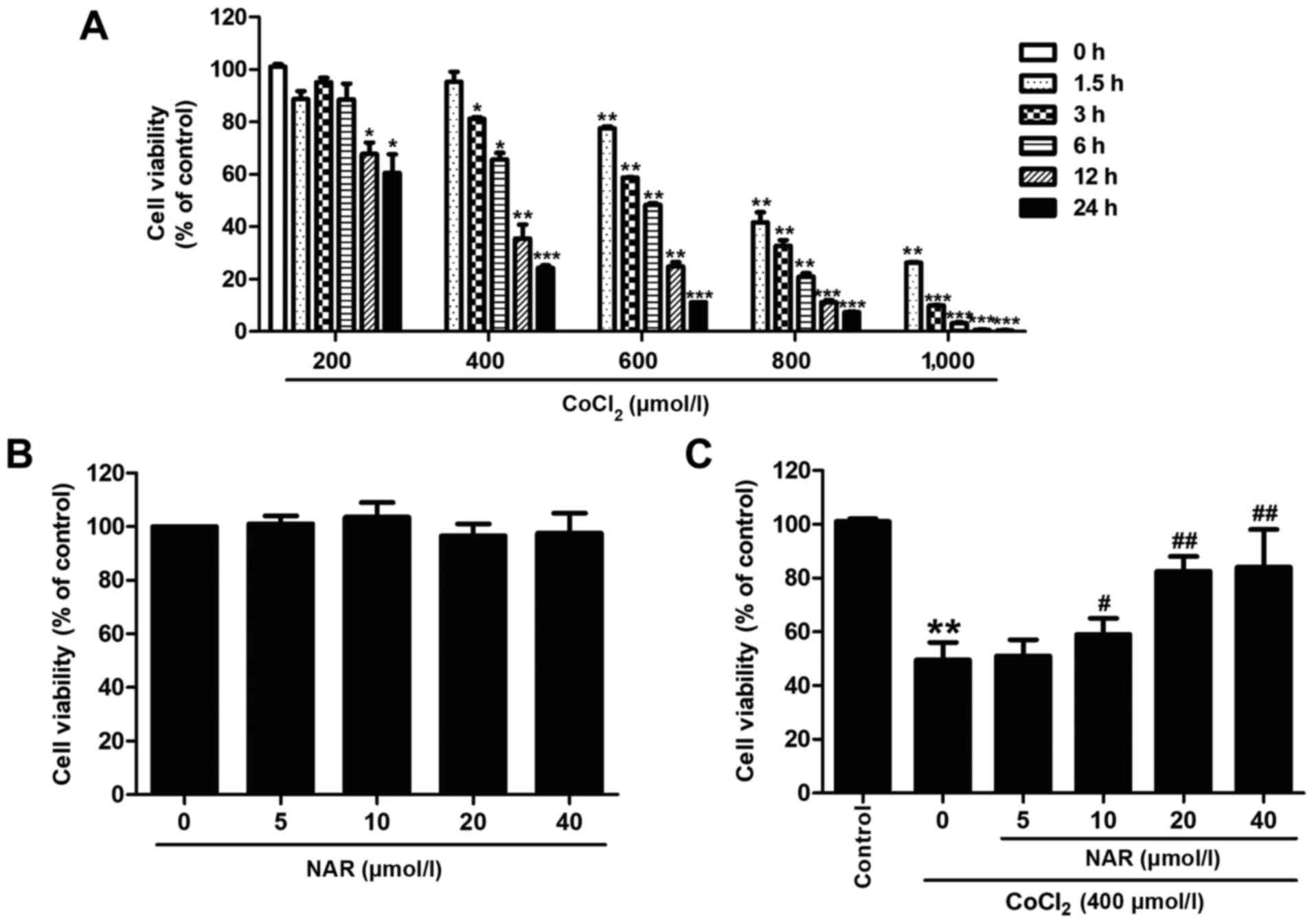 | Figure 1Naringin attenuates
CoCl2-induced cytotoxicity. H9C2 cells were incubated
with various concentrations of CoCl2 (200, 400, 600, 800
or 1,000 µmol/l) for different periods of time (1.5, 3, 6,
12 or 24 h). (A) Cell viability was determined using a CCK-8 assay.
*P<0.05, **P<0.01,
***P<0.001 vs. control group. (B) H9C2 cells were
incubated with various concentrations of naringin (5, 10, 20 or 40
µmol/l) and cell viability was measured by CCK-8 assay. H9C2
cells were pre-treated with 20 µmol/l naringin for 2 h
followed by co-treatment with 400 µM CoCl2 for 12
h. (C) Cell viability was assessed by CCK-8 assay. Results are
presented as the means ± standard deviation; n=3.
**P<0.01 vs. control group; #P<0.05,
##P<0.01 vs. CoCl2 group. CCK-8, Cell
Counting kit-8; CoCl2, cobalt chloride; NAR,
naringin. |
Naringin reduces the
CoCl2-induced apoptosis of H9C2 cells
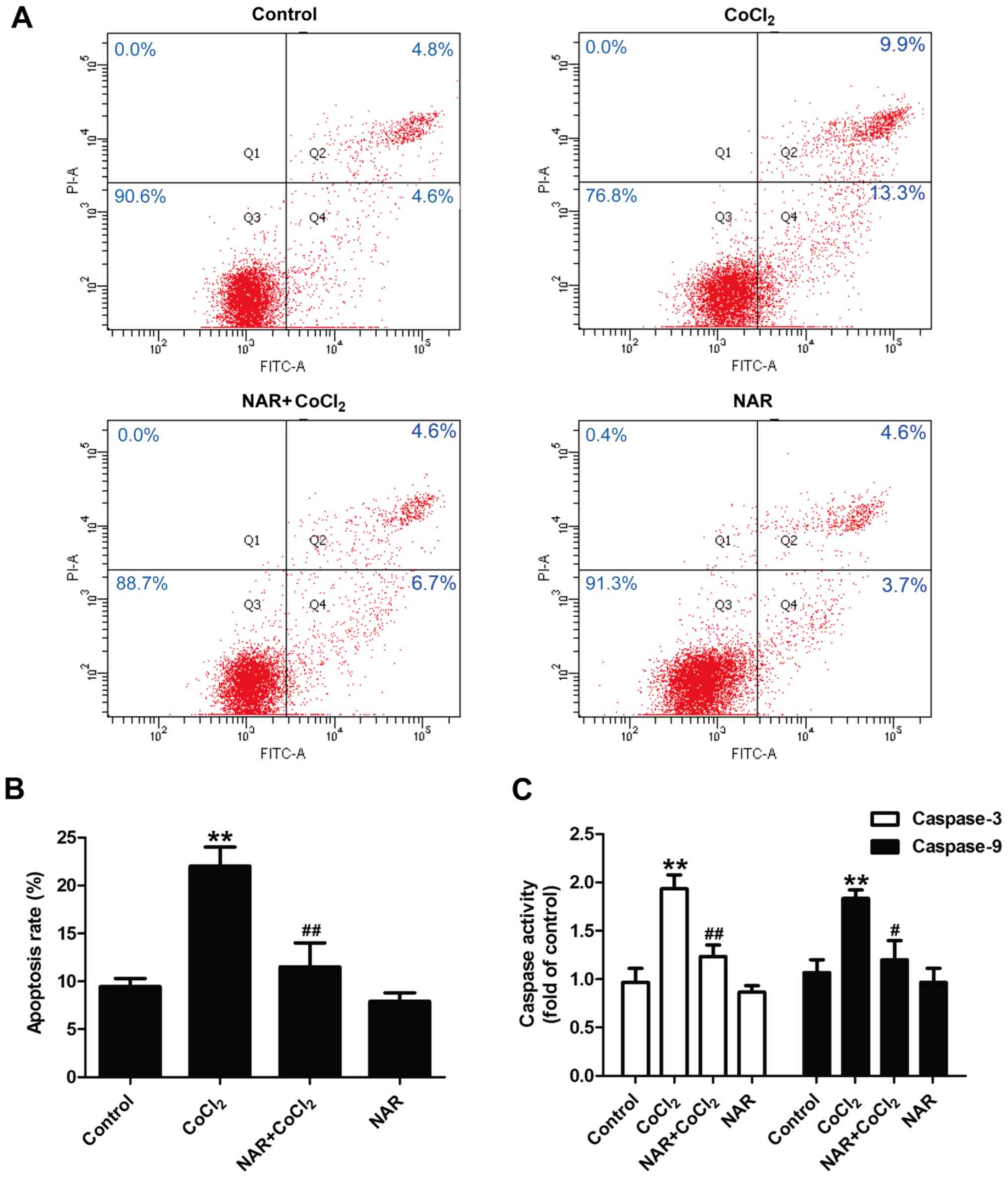
Subsequently, the effect of naringin on the
apoptosis of CoCl2-treated H9C2 cells was assessed. Flow
cytometric analysis (Fig. 2A and
B) revealed that treatment with CoCl2 resulted in a
notable increase in the apoptotic rate (P<0.01), and this was
prevented by naringin pre-treatment (P<0.01). Caspase-3 and
caspase-9 are essential for apoptosis during myocardial damage
(30). As shown in Fig. 2, the activities of caspase-3 and
caspase-9 (Fig. 2C) were
increased by CoCl2 (P<0.01), and these effects were
significantly reversed by naringin (P<0.05). Naringin treatment
alone had no effect on apoptosis and caspase-3/caspase-9 activity.
These results demonstrate that naringin mitigates the
CoCl2-induced apoptosis of H9C2 cells.
CoCl2 induces the blockage of
the autophagic flux and leads to myocardial cell injury
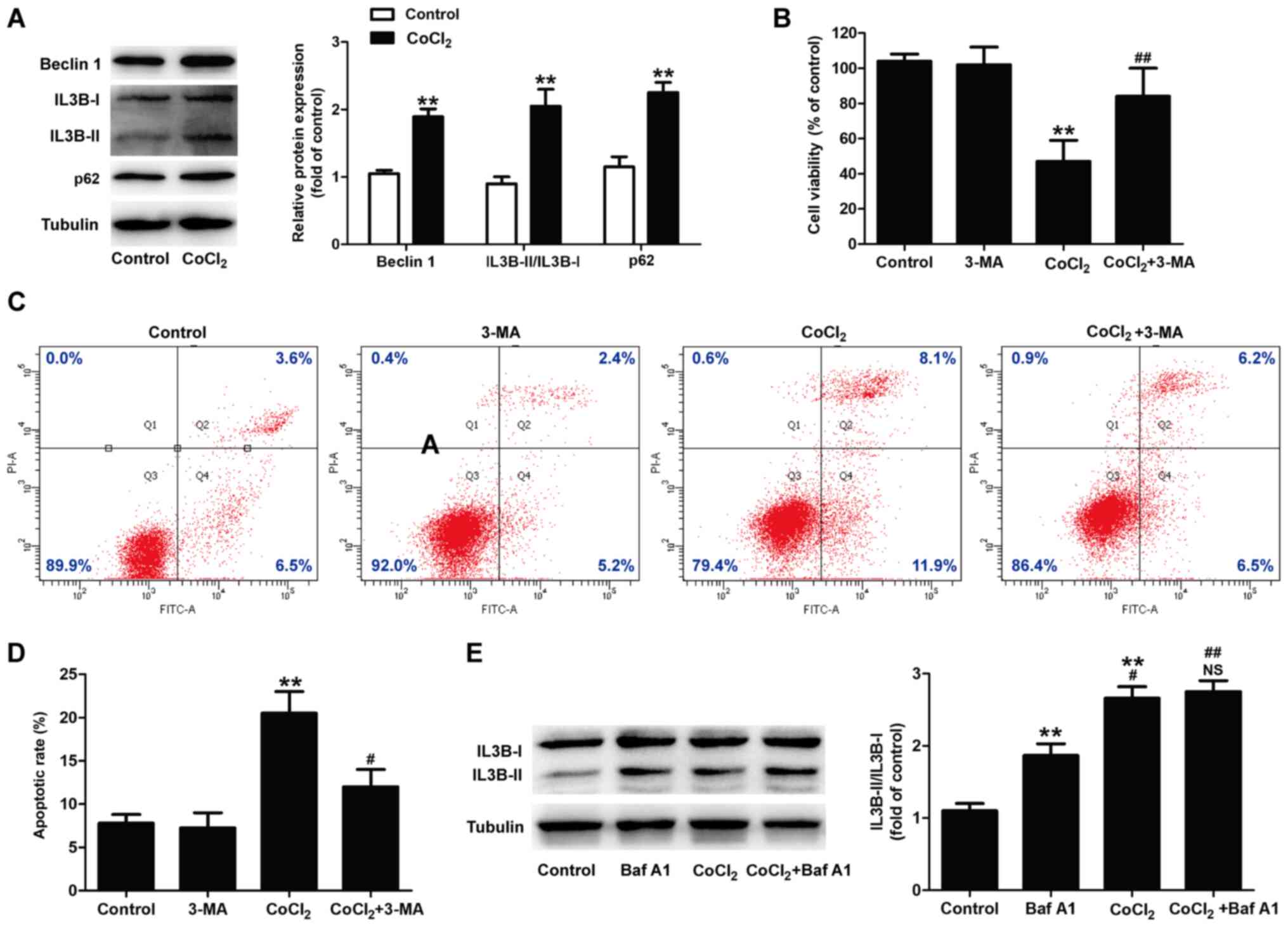
Autophagy is closely associated with cell death in
the hypoxic-ischemic process (31,32). To confirm the role of autophagy
in CoCl2-induced injury to H9C2 cells, the expression of
autophagy-related markers was determined. Western blot analysis
revealed that following treatment with CoCl2, the levels
of Beclin 1 expression (P<0.01) and the LC3B-II/LC3B-I ratio
(P<0.01) were both significantly upregulated compared with the
control group (Fig. 3A). p62 is
a marker of the autophagic flux that is degraded in autophagosomes
(33). In the present study,
CoCl2 treatment increased the expression of p62 in H9C2
cells (Fig. 3A).
To determine whether the autophagic response
following CoCl2 treatment resulted in damage to the H9C2
cells, the CoCl2-treated cells were co-treated with
3-MA, a selective inhibitor of autophagic initiation. The results
of CCK-8 assay revealed that cell viability was significantly
increased in the 3-MA and CoCl2 co-treatment groups
compared with CoCl2 treatment alone (Fig. 3B). 3-MA also attenuated the
CoCl2-induced increase in apoptosis (Fig. 3C and D). 3-MA treatment alone did
not affect cell viability and apoptosis compared with the control
group. The increases in the expression levels of pro-autophagic
proteins may be due to either enhanced autophagosome formation or
reduced autophagosome degradation. Furthermore, western blot
analysis revealed that treatment with Baf A1 (an inhibitor of the
vacuolar H+ ATPase of lysosomes) resulted in the
upregulation of the LC3II/LC3I ratio compared with the control
group (Fig. 3E), indicating a
block on the basal lysosomal-dependent degradation of autophagosome
cargo. Notably, the LC3II/LC3I ratio in the CoCl2 group
was upregulated compared with the Baf A1 group, and there was no
significant difference in the LC3II/LC3I ratio between
CoCl2 treatment alone and co-treatment with Baf A1 and
CoCl2, indicating that CoCl2 reduced the
autophagic flux. Taken together, these data suggest that
CoCl2-induced H9C2 cell injury results in the blockage
of autophagic clearance rather than in autophagosome formation.
Naringin protects H9C2 cells against
CoCl2-induced injury by restoring the impaired
autophagic flux
To further investigate the role of the autophagic
flux in the protective effects of naringin against
CoCl2-induced H9C2 cell injury, the effects of naringin
on the levels of autophagy-related markers were detected. As shown
in Fig. 4, western blot analysis
revealed that naringin treatment further increased Beclin 1
expression (Fig. 4A and B) and
the LC3II/LC3I ratio (Fig. 4A and
C) compared with CoCl2 treatment. Naringin reversed
the CoCl2-induced upregulation of p62 expression in H9C2
cells (Fig. 4A and D). Compared
with the control group, naringin treatment alone had no effect on
the expression of these autophagy-related proteins. These results
suggested an increase in the autophagic flux induced by naringin
pre-treatment. In addition, the inhibitors of autophagy (3-MA and
Baf A1) both attenuated the naringin-induced upregulation of cell
viability (Fig. 4E) and the
downregulation of LDH release (Fig.
4F) in CoCl2-treated H9C2 cells. Flow cytometric
analysis also revealed that 3-MA and Baf A1 both mitigated the
naringin-induced decrease in the apoptotic rate (Fig. 4G and H), as well as caspase-3
activity (Fig. 4I) and caspase-9
activity (Fig. 4J) in
CoCl2-treated H9C2 cells. These results suggest that
naringin enhances the autophagic flux, including autophagosome
formation and autophagic clearance, and mitigates CoCl2
injury in H9C2 cells.
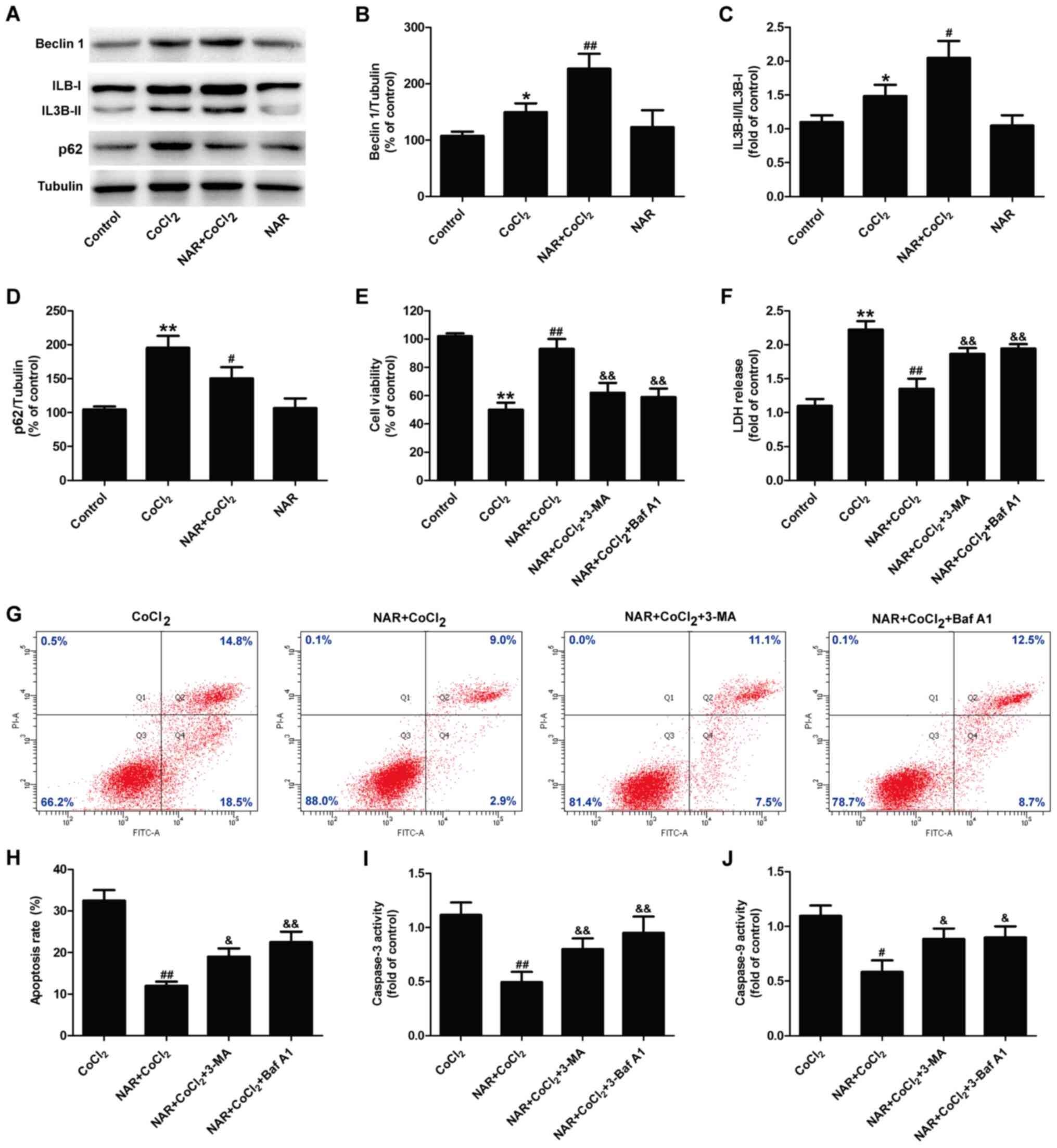 | Figure 4Enhancement of the autophagic flux
mediates the protective effects of naringin on
CoCl2-induced injury in H9C2 cells. H9C2 cells were
pre-treated with 20 µmol/l naringin for 2 h followed by
co-treatment with 400 µmol/l CoCl2 for 12 h. (A)
Expression of autophagy-related proteins. Quantitative analysis of
(B) Beclin 1 expression, (C) the IL3B-II/IL3B-I ratio and (D) p62
expression. *P<0.05, **P<0.01 vs.
control group; #P<0.05, ##P<0.01 vs.
CoCl2 group. H9C2 cells were pre-treated with 5 mM 3-MA
for 1 h, followed by treatment with 20 µmol/l naringin for 2
h and then co-treatment with 400 µmol/l CoCl2 for
12 h, or H9C2 cells were treated with naringin for 2 h prior to
co-treatment with 10 nmol/l Baf A1 and 400 µmol/l
CoCl2 for 12 h. (E) Cell viability was measured by CCK-8
assay. (F) LDH release was measured using a LDH assay kit. (G and
H) Apoptosis was analyzed by Annexin V-FITC/PI double staining and
the apoptotic rate was measured and quantified by flow cytometry.
(I) Caspase-3 activity was determined using a Caspase-3 Activity
Assay kit. (J) Caspase-9 activity was determined using a Caspase-9
Activity Assay kit. Results are presented as the means ± standard
deviation n=3. **P<0.01 vs. control group;
#P<0.05, ##P<0.01 vs. CoCl2
group; &P<0.05, &&P<0.01
vs. NAR + CoCl2 group. CCK-8, Cell Counting Kit-8;
CoCl2, cobalt chloride; NAR, naringin; Baf A1,
bafilomycin A1; LDH, lactate dehydrogenase. |
Naringin attenuates
CoCl2-induced injury and promotes the autophagic flux by
promoting the activation of the HIF-1α/BNIP3 signaling pathway in
H9C2 cells
The HIF-1α/BNIP3 signaling pathway has been shown to
play an essential role in hypoxia-induced autophagy (28,29). In the present study, the effects
of naringin on the HIF-1α/BNIP3 signaling pathway in
CoCl2-treated H9C2 cells was then assessed. The results
of western blot analysis revealed that naringin pre-treatment
further increased the expression of HIF-1α (Fig. 5A and B) and BNIP3 (Fig. 5A and C) in
CoCl2-treated H9C2 cells, indicating that naringin
promoted the activity of the HIF-1α/BNIP3 signaling pathway in the
CoCl2-treated H9C2 cells. Furthermore, YC-1, an
inhibitor of HIF-1α, blocked the naringin-induced increase in the
expression of HIF-1α (Fig. 5D and
E) and BNIP3 (Fig. 5D and F)
in CoCl2-treated H9C2 cells. On these basis, YC-1 also
attenuated the naringin-induced upregulation of Beclin 1 expression
(Fig. 5G and H) and the ratio of
IL3B-II to IL3B-I (Fig. 5G and
I), as well as the downregulation of p62 expression (Fig. 5G and J) in
CoCl2-treated H9C2 cells. This suggested that
HIF-1α/BNIP3 mediated the naringin-induced increase in the
autophagic flux. Moreover, treatment with YC-1 eliminated the
naringin-induced increase in cell viability (Fig. 5K) and the decrease in LDH release
(Fig. 5L). These results
indicated that naringin protects H9C2 cells against
CoCl2 injury by enhancing autophagic flux via activation
of the HIF-1α/BNIP3 signaling pathway.
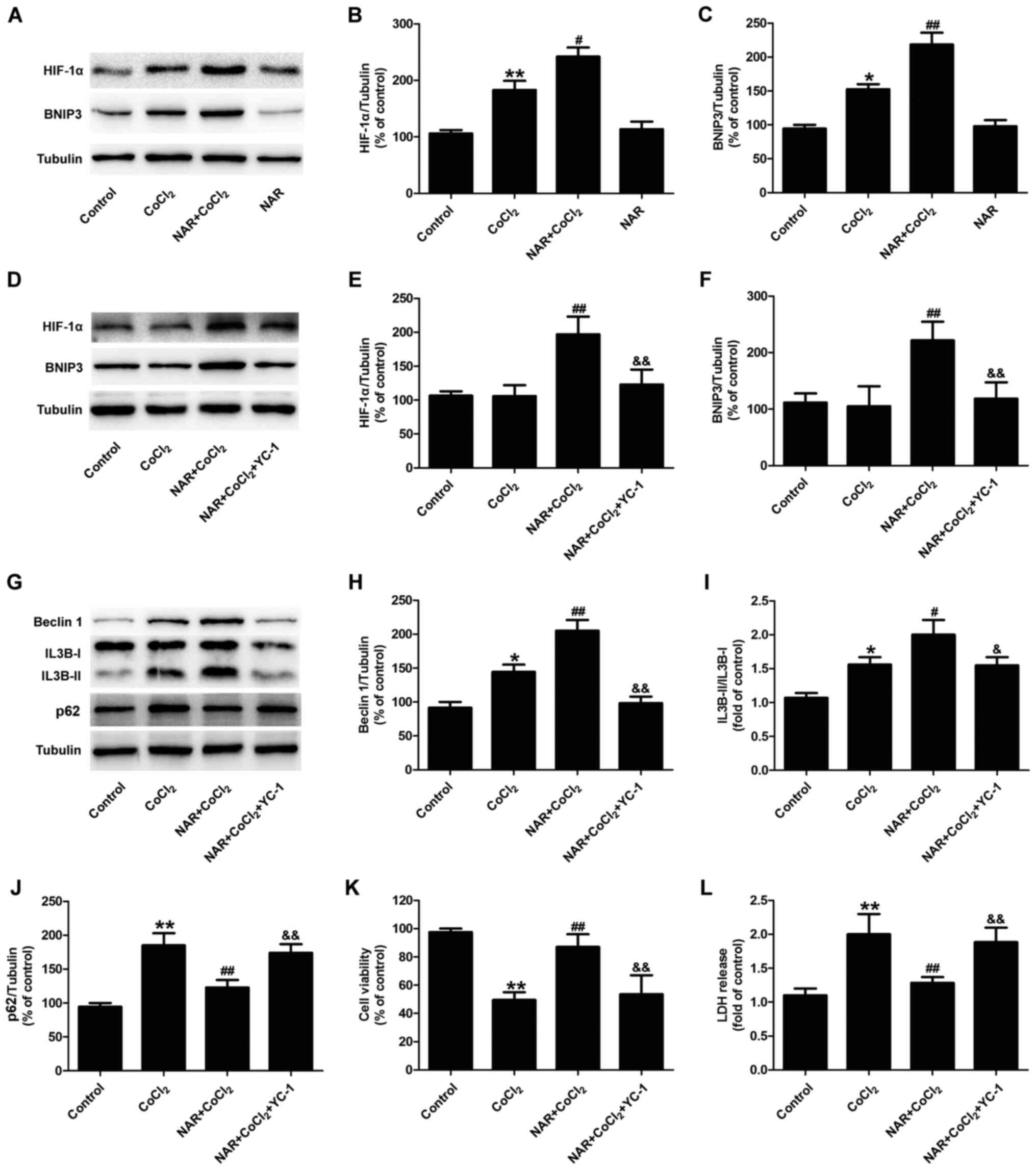 | Figure 5HIF-1α/BNIP3 contributes to the
naringin-induced enhancement of the autophagic flux and the
cardioprotective effects against CoCl2-induced injury to
H9C2 cells. H9C2 cells were treated with 20 µmol/l naringin
for 2 h followed by co-treatment with 400 µmol/l
CoCl2 for 12 h. (A) HIF-1α and BNIP3 protein expression
was determined by western blot analysis. Quantitative analysis of
(B) HIF-1α and (C) BNIP3 expression. Results are expressed as the
means ± SD; n=3. *P<0.05, **P<0.01 vs.
control group; #P<0.05 and ##P<0.01 vs.
CoCl2 group. H9C2 cells were pre-treated with 10
µM YC-1 for 1 h, then 20 µmol/l naringin for 2 h,
followed by co-treatment with 400 µmol/l CoCl2
for 12 h. (D) Expression of HIF-1α and BNIP3 were measured by
western blot analysis. Quantitative analysis of (E) HIF-1α and (F)
BNIP3 expression. (G) Expression of autophagy-associated proteins
was determined by western blot analysis. (H) Quantitative analysis
of Beclin 1 expression. (I) Quantitative analysis of IL3B-II/IL3B-I
ratio. (J) Quantitative analysis of p62 expression. (K) Cell
viability was measured by CCK-8 assay. (L) LDH release was measured
using an LDH assay kit. Results are presented as the means ±
standard deviation. n=3. *P<0.05,
**P<0.01 vs. control group; #P<0.05,
##P<0.01 vs. CoCl2 group;
&P<0.05, &&P<0.01 vs. NAR +
CoCl2 group. CCK-8, Cell Counting kit-8;
CoCl2, cobalt chloride; LDH, lactate dehydrogenase; NAR,
naringin. |
Discussion
Hypoxia/ischemia can induce sudden cardiomyocyte
death, and this may result in further cardiovascular diseases
(4,34). Naringin possesses anti-apoptotic,
antioxidant and cardioprotective properties in the cardiovascular
system (11,12). However, the effects of naringin
on hypoxia/ischemia injury remain undetermined. The results of the
present study demonstrated that naringin protected H9C2 cells
against hypoxia-induced injury by promoting the autophagic flux via
activation of the HIF-1α/BNIP3 signaling pathway.
CoCl2 (a chemical hypoxia-mimetic
agent)-induced H9C2 cell injury has been shown to be a suitable
in vitro model of hypoxia-associated damage in
cardiomyocytes (35,36), and was thus used to explore the
underlying mechanisms in the present study. The mechanisms
underlying naringin-induced cardioprotection against hypoxia injury
in the CoCl2-treated H9C2 cells were determined. The
results revealed that CoCl2 significantly reduced H9C2
cell viability in a concentration- and time-dependent manner,
consistent with the findings of previous studies (27,37). Recently, naringin, which is the
major active constituent of tomentose pummelo peel, has been found
to exert cardioprotective effects in animal and cell damage models
(12,38). Naringin has been shown to
attenuated the effects of several damaging stimuli, such as
doxorubicin (11), hyperglycemia
(39) and anoxia/reoxygenation
induced myocardial injury (13)
via the inhibition of apoptosis and oxidative stress in
vitro and in vivo. Consistent with the findings of these
studies, the results of the present study demonstrated that
naringin attenuated the CoCl2-induced decrease in cell
viability and the increase in apoptosis. These results indicate
that naringin protects H9C2 cells against CoCl2-induced
injury.
Autophagy is an evolutionarily controlled
lysosome-mediated process that eliminates damaged proteins and
intracellular organelle, involved in a variety of physiological
processes (40,41). An increasing number of studies
have demonstrated that autophagy dysfunction is increased in the
heart following hypoxia/ischemia, and restoring impaired autophagic
flux may prevent myocardial damage against hypoxia/ischemia
(42-44). Recent studies have also indicated
that CoCl2 increases the levels of markers associated
with the autophagic-lysosomal pathway, including Beclin1 (an
indicator of initiation of autophagy), the conversion of LC3B-I to
LC3B-II (a surrogate index of autophagosome formation) and p62 (a
commonly used indicator of autophagic flux inhibition) (13,45). Consistent with these studies, the
results of the present study also indicated that CoCl2
significantly increased Beclin 1 expression and the LC3B-II/LC3B-I
ratio, and decreased p62 expression in H9C2 cells. Thereafter, it
is was shown that 3-MA attenuated CoCl2-induced cell
damage and apoptosis in H9C2 cells, indicating that reducing
autophagosome formation contributed to CoCl2-induced
myocardial damage. In addition, previous studies have revealed that
autophagy also plays important roles in the regulation of cell
death, particularly in apoptosis-signaling pathways (46,47). Hence, it was hypothesized that
the impaired autophagic flux leads to an increase in the apoptosis
of CoCl2-treated H9C2 cells. Increases in the levels of
pro-autophagic proteins may be due to either an enhanced
autophagosome formation or reduced autophagosome degradation
(48,49). To distinguish between these two
possibilities, the H9C2 cells were treated with Baf A1 in the
present study. The results revealed that Baf A1 treatment alone
resulted in the upregulation of the LC3II/LC3I ratio, indicating a
block on the basal lysosomal-dependent degradation of autophagosome
cargo. Notably, the LC3B-II/LC3B-I ratio in the CoCl2
group was upregulated compared with the Baf A1 group, and there was
no significant difference in LC3II/LC3I ratio between
CoCl2 and Baf A1 co-treatment group with the
CoCl2 group, indicating that CoCl2 inhibited
the autophagic flux. Taken together, these data suggest that
CoCl2-induced H9C2 cell injury is the result of blockage
of autophagic flux, as evidenced by the enhancement of
autophagosome formation and the reduction of lysosome-mediated
autophagosome degradation.
Recently, naringin has been shown to regulate
autophagy in several types of diseases (38,50,51). Naringin has been shown to promote
the levels of autophagy markers, such as LC3B-II/I and Beclin-1,
and autophagy is involved in the protective effect of naringin on
oxidative stress-induced apoptosis in nucleus pulposus cells
(14). Feng et al
(50) demonstrated that naringin
reduced cerebral apoptotic cell death underlying
ischemia-reperfusion via the regulation of the activation of
mitophagy. However, the effects of naringin on autophagy or the
autophagic flux under hypoxia/ischemic conditions have not yet been
studied, at least to the best of our knowledge. To the best of our
knowledge, the present study was the first to demonstrate that
naringin pre-treatment further increased Beclin 1 expression and
the IL3B-II/IL3B-I ratio in CoCl2-treated H9C2 cells,
and that naringin pretreatment decreased p62 expression in
CoCl2-treated H9C2 cells, suggesting that naringin
restores the autophagic flux under hypoxic conditions. In addition,
Baf A1 attenuated the protective effects of naringin on cell injury
and apoptosis induced by CoCl2 in H9C2 cells. These
results suggest that naringin rescues the CoCl2-induced
impaired autophagic flux by promoting the initiation of autophagy
and degradation, and then protects H9C2 cells against
CoCl2-induced injury. Combined with the role of
autophagy in CoCl2 injury as discussed above, the
ability of naringin to promote lysosome-mediated autophagosome
degradation was more potent than the ability to promote
autophagosome formation, thus exerting a cardioprotective effect
against hypoxia injury.
HIF-1α is an important transcription factor in the
response to hypoxia, and BNIP3, a transcriptional target of HIF-1α,
is upregulated in response to hypoxia (21,52,53). Notably, previous studies have
demonstrated that the HIF-1α/BNIP3 signaling pathway is closely
associated with hypoxia-induced autophagy (25,54). Previous studies have also
indicated that CoCl2 can disrupt HIF-1α degradation, and
increase HIF-1α and BNIP3 expression, thus inducing apoptosis and
autophagic cell death (25,55). Consistent with these studies, the
expression levels of HIF-1α and BNIP3 in H9C2 cells were also
significantly increased following CoCl2 treatment in the
present study. Notably, in the present study, it was shown that
naringin further increased the HIF-1α and BNIP3 expression levels,
activating the HIF-1α/BNIP3 signaling pathway. The HIF-1α
inhibitor, YC-1, induced the inhibition of the HIF-1α/BNIP3
signaling pathway and reversed the naringin-induced increase in
Beclin 1 expression, in the IL3B-II/IL3B-1 ratio, and the decrease
in p62 expression, suggesting that the enhancement of the
autophagic flux induced by naringin was mediated by the
HIF-1α/BNIP3 signaling pathway. In addition, YC-1 also blocked the
protective effects of naringin against CoCl2-induced
injury. These results suggest that naringin protects the H9C2 cells
against CoCl2-induced injury by enhancing the autophagic
flux through the activation of the HIF-1α/BNIP3 signaling
pathway.
Limitations still exist in the present study. First,
the present study did not detect the change in the autophagic flux
with IL-3 fluorescence staining. In addition, drugs were simply
used to intervene with the HIF-1 signaling pathway without using
gene silencing. Second, as all our assays were performed using
cells, in future experiments, the authors aim to introduce tests
and to investigate the underlying mechanisms in in vitro
animal models, such as rats. Other studies have confirmed that
myocardial ischemia injury induces the inflammatory response
(56,57). Naringin also has been shown to
exhibit anti-inflammatory activity (58,59). Hence, it is meaningful to explore
the protective mechanism of naringin on myocardial ischemia injury
focusing on the inflammatory response in the future studies. In
addition, naringin has been shown to exhibit anti-inflammatory
activity. In addition, further identification and confirmation of
the precise mechanisms underlying naringin and HIF-1α/BNIP3
signaling pathway are required.
In conclusion, the present study demonstrated that
naringin attenuated CoCl2-induced injury in H9C2 cells
by restoring the autophagic flux via the activation of the
HIF-1α/BNIP3 signaling pathway. These findings reinforce the
cardioprotective effects of naringin against hypoxic injury and
indicate that the HIF-1α/BNIP3 signaling pathway may serve as a
novel therapeutic target for the management of hypoxia/ischemia
injury.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL and JJ designed the experiments and contributed
to the drafting of the manuscript. JF, CH and XL collated and
analyzed the data. WL and KW performed the experiments and wrote
the manuscript. JJ and CH wrote and revised the manuscript. SL and
KW confirm the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Guangdong Medical Science
And Technology Research Fund Project (grant nos. A2017494 and
A2016267) and the Project of Traditional Chinese Medicine Bureau of
Guangdong Province (grant no. 20181149).
References
|
1
|
GBD 2017 Causes of Death Collaborators:
Global, regional, and national age-sex-specific mortality for 282
causes of death in 195 countries and territories, 1980-2017: A
systematic analysis for the global burden of disease study 2017.
Lancet. 392:1736–1788. 2018. View Article : Google Scholar
|
|
2
|
Aouiss A, Anka Idrissi D, Kabine M and
Zaid Y: Update of inflammatory proliferative retinopathy: Ischemia,
hypoxia and angiogenesis. Curr Res Transl Med. 67:62–71. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cordova Martinez A, Pascual Fernandez J,
Fernandez Lazaro D and Alvarez Mon M: Muscular and heart
adaptations of execise in hypoxia. Is training in slow hypoxy
healthy? Med Clin (Barc). 148:469–474. 2017.In English,
Spanish.
|
|
4
|
Rodgers JL, Iyer D, Rodgers LE,
Vanthenapalli S and Panguluri SK: Impact of hyperoxia on cardiac
pathophysiology. J Cell Physiol. 234:12595–12603. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Den Hartogh DJ and Tsiani E: Antidiabetic
properties of naringenin: A citrus fruit polyphenol. Biomolecules.
9:992019. View Article : Google Scholar :
|
|
6
|
Kwatra M, Kumar V, Jangra A, Mishra M,
Ahmed S, Ghosh P, Vohora D and Khanam R: Ameliorative effect of
naringin against doxorubicin-induced acute cardiac toxicity in
rats. Pharm Biol. 54:637–647. 2016. View Article : Google Scholar
|
|
7
|
Ming H, Chuang Q, Jiashi W, Bin L,
Guangbin W and Xianglu J: Naringin targets Zeb1 to suppress
osteosarcoma cell proliferation and metastasis. Aging (Albany NY).
10:4141–4151. 2018. View Article : Google Scholar
|
|
8
|
Salehi B, Fokou PVT, Sharifi-Rad M, Zucca
P, Pezzani R, Martins N and Sharifi-Rad J: The therapeutic
potential of naringenin: A review of clinical trials.
Pharmaceuticals (Basel). 12:112019. View Article : Google Scholar
|
|
9
|
Adebiyi OA, Adebiyi OO and Owira PM:
Naringin reduces hyperglycemia-induced cardiac fibrosis by
relieving oxidative stress. PLoS One. 11:e01498902016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park JH, Ku HJ, Kim JK, Park JW and Lee
JH: Amelioration of high fructose-induced cardiac hypertrophy by
naringin. Sci Rep. 8:94642018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jian CY, Ouyang HB, Xiang XH, Chen JL, Li
YX, Zhou X, Wang JY, Yang Y, Zhong EY, Huang WH and Zhang HW:
Naringin protects myocardial cells from doxorubicin-induced
apoptosis partially by inhibiting the p38MAPK pathway. Mol Med Rep.
16:9457–9463. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen RC, Sun GB, Wang J, Zhang HJ and Sun
XB: Naringin protects against anoxia/reoxygenation-induced
apoptosis in H9c2 cells via the Nrf2 signaling pathway. Food Funct.
6:1331–1344. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cai CC, Zhu JH, Ye LX, Dai YY, Fang MC, Hu
YY, Pan SL, Chen S, Li PJ, Fu XQ and Lin ZL: Glycine protects
against hypoxic-ischemic brain injury by regulating
mitochondria-mediated autophagy via the AMPK pathway. Oxid Med Cell
Longev. 2019:42485292019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Z, Wang C, Lin J, Jin H, Wang K, Yan
Y, Wang J, Wu C, Nisar M, Tian N, et al: Therapeutic potential of
naringin for intervertebral disc degeneration: Involvement of
autophagy against oxidative stress-induced apoptosis in nucleus
pulposus cells. Am J Chin Med. Oct 4–2018.Epub ahead of print.
View Article : Google Scholar
|
|
15
|
Sousa Fialho MD, Abd Jamil AH, Stannard GA
and Heather LC: Hypoxia-inducible factor 1 signalling, metabolism
and its therapeutic potential in cardiovascular disease. Biochim
Biophys Acta Mol Basis Dis. 1865:831–843. 2019. View Article : Google Scholar
|
|
16
|
Bravo-San Pedro JM, Kroemer G and Galluzzi
L: Autophagy and mitophagy in cardiovascular disease. Circ Res.
120:1812–1824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sciarretta S, Maejima Y, Zablocki D and
Sadoshima J: The role of autophagy in the heart. Annu Rev Physiol.
80:1–26. 2018. View Article : Google Scholar
|
|
18
|
Qiao J, Huang J, Zhou M, Cao G and Shen H:
Inhibition of HIF-1α restrains fracture healing via regulation of
autophagy in a rat model. Exp Ther Med. 17:1884–1890.
2019.PubMed/NCBI
|
|
19
|
Zimmerman MA, Biggers CD and Li PA:
Rapamycin treatment increases hippocampal cell viability in an
mTOR-independent manner during exposure to hypoxia mimetic, cobalt
chloride. BMC Neurosci. 19:822018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gallo S, Gatti S, Sala V, Albano R,
Costelli P, Casanova E, Comoglio PM and Crepaldi T: Agonist
antibodies activating the Met receptor protect cardiomyoblasts from
cobalt chloride-induced apoptosis and autophagy. Cell Death Dis.
5:e11852014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee JW, Bae SH, Jeong JW, Kim SH and Kim
KW: Hypoxia-inducible factor (HIF-1)alpha: Its protein stability
and biological functions. Exp Mol Med. 36:1–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang H, Zhang D, Jia S, Huang S, Xiao L,
Ma L, Liu G, Gong K and Xu L: Effect of sustained hypoxia on
autophagy of genioglossus muscle-derived stem cells. Med Sci Monit.
24:2218–2224. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Liu D, Hu H, Zhang P, Xie R and
Cui W: HIF-1α/BNIP3 signaling pathway-induced-autophagy plays
protective role during myocardial ischemia-reperfusion injury.
Biomed Pharmacother. 120:1094642019. View Article : Google Scholar
|
|
24
|
Liu XW, Lu MK, Zhong HT, Wang LH and Fu
YP: Panax notoginseng saponins attenuate myocardial
ischemia-reperfusion injury through the HIF-1α/BNIP3 pathway of
autophagy. J Cardiovasc Pharmacol. 73:92–99. 2019. View Article : Google Scholar
|
|
25
|
Wu H, Huang S, Chen Z, Liu W, Zhou X and
Zhang D: Hypoxia-induced autophagy contributes to the invasion of
salivary adenoid cystic carcinoma through the HIF-1α/BNIP3
signaling pathway. Mol Med Rep. 12:6467–6474. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang K, Peng S, Xiong S, Niu A, Xia M,
Xiong X, Zeng G and Huang Q: Naringin inhibits autophagy mediated
by PI3K-Akt-mTOR pathway to ameliorate endothelial cell dysfunction
induced by high glucose/high fat stress. Eur J Pharmacol.
874:1730032020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Manu TM, Anand T, Pandareesh MD, Kumar PB
and Khanum F: Terminalia arjuna extract and arjunic acid mitigate
cobalt chloride-induced hypoxia stress-mediated apoptosis in H9c2
cells. Naunyn Schmiedebergs Arch Pharmacol. 392:1107–1119. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Warbrick I and Rabkin SW:
Hypoxia-inducible factor 1-alpha (HIF-1α) as a factor mediating the
relationship between obesity and heart failure with preserved
ejection fraction. Obes Rev. 20:701–712. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hescheler J, Meyer R, Plant S, Krautwurst
D, Rosenthal W and Schultz G: Morphological, biochemical, and
electrophysiological characterization of a clonal cell (H9c2) line
from rat heart. Circ Res. 69:1476–1486. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haider N, Narula N and Narula J: Apoptosis
in heart failure represents programmed cell survival, not death, of
cardiomyocytes and likelihood of reverse remodeling. J Card Fail.
8(6 Suppl): S512–S517. 2002. View Article : Google Scholar
|
|
31
|
Semenza GL: Hypoxia-inducible factor 1:
Regulator of mitochondrial metabolism and mediator of ischemic
preconditioning. Biochim Biophys Acta. 1813:1263–1268. 2011.
View Article : Google Scholar
|
|
32
|
Yadav AK, Yadav PK, Chaudhary GR, Tiwari
M, Gupta A, Sharma A, Pandey AN, Pandey AK and Chaube SK: Autophagy
in hypoxic ovary. Cell Mol Life Sci. 76:3311–3322. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lamark T, Svenning S and Johansen T:
Regulation of selective autophagy: The p62/SQSTM1 paradigm. Essays
Biochem. 61:609–624. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mallet RT, Manukhina EB, Ruelas SS,
Caffrey JL and Downey HF: Cardioprotection by intermittent hypoxia
conditioning: Evidence, mechanisms, and therapeutic potential. Am J
Physiol Heart Circ Physiol. 315:H216–H232. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng CI, Lee YH, Chen PH, Lin YC, Chou MH
and Kao YH: Cobalt chloride induces RhoA/ROCK activation and
remodeling effect in H9c2 cardiomyoblasts: Involvement of PI3K/Akt
and MAPK pathways. Cell Signal. 36:25–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Munoz-Sanchez J and Chanez-Cardenas ME:
The use of cobalt chloride as a chemical hypoxia model. J Appl
Toxicol. 39:556–570. 2019. View Article : Google Scholar
|
|
37
|
Shi YN, Zhang XQ, Hu ZY, Zhang CJ, Liao
DF, Huang HL and Qin L: Genistein protects H9c2 cardiomyocytes
against chemical hypoxia-induced injury via inhibition of
apoptosis. Pharmacology. 103:282–290. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Caglayan C: The effects of naringin on
different cyclophosphamide-induced organ toxicities in rats:
Investigation of changes in some metabolic enzyme activities.
Environ Sci Pollut Res Int. 26:26664–26673. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
You Q, Wu Z, Wu B, Liu C, Huang R, Yang L,
Guo R, Wu K and Chen J: Naringin protects cardiomyocytes against
hyperglycemia-induced injuries in vitro and in vivo. J Endocrinol.
230:197–214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Moosavi MA and Djavaheri-Mergny M:
Autophagy: New insights into mechanisms of action and resistance of
treatment in acute promyelocytic leukemia. Int J Mol Sci.
20:35592019. View Article : Google Scholar :
|
|
41
|
Ravanan P, Srikumar IF and Talwar P:
Autophagy: The spotlight for cellular stress responses. Life Sci.
188:53–67. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gao C, Wang R, Li B, Guo Y, Yin T, Xia Y,
Zhang F, Lian K, Liu Y, Wang H, et al: TXNIP/Redd1 signaling and
excessive autophagy: A novel mechanism of myocardial
ischemia/reperfusion injury in mice. Cardiovasc Res. 116:645–657.
2019. View Article : Google Scholar
|
|
43
|
McCormick J, Knight RA, Barry SP,
Scarabelli TM, Abounit K, Latchman DS and Stephanou A: Autophagy in
the stress-induced myocardium. Front Biosci (Elite Ed).
4:2131–2141. 2012. View
Article : Google Scholar
|
|
44
|
Ryter SW, Lee SJ, Smith A and Choi AM:
Autophagy in vascular disease. Proc Am Thorac Soc. 7:40–47. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen Y, Yan Q, Xu Y, Ye F, Sun X, Zhu H
and Wang H: BNIP3-mediated autophagy induced inflammatory response
and inhibited VEGF expression in cultured retinal pigment
epithelium cells under hypoxia. Curr Mol Med. 19:395–404. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Glick D, Barth S and Macleod KF:
Autophagy: Cellular and molecular mechanisms. J Pathol. 221:3–12.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kaizuka T, Morishita H, Hama Y, Tsukamoto
S, Matsui T, Toyota Y, Kodama A, Ishihara T, Mizushima T and
Mizushima N: An autophagic flux probe that releases an internal
control. Mol Cell. 64:835–849. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu J and Lipinski MM: Autophagy in
neurotrauma: Good, bad, or dysregulated. Cells. 8:6932019.
View Article : Google Scholar :
|
|
50
|
Feng J, Chen X, Lu S, Li W, Yang D, Su W,
Wang X and Shen J: Naringin attenuates cerebral
ischemia-reperfusion injury through inhibiting
peroxynitrite-mediated mitophagy activation. Mol Neurobiol.
55:9029–9042. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Raha S, Yumnam S, Hong GE, Lee HJ,
Saralamma VV, Park HS, Heo JD, Lee SJ, Kim EH, Kim JA and Kim GS:
Naringin induces autophagy-mediated growth inhibition by
downregulating the PI3K/Akt/mTOR cascade via activation of MAPK
pathways in AGS cancer cells. Int J Oncol. 47:1061–1069. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Adams JM, Difazio LT, Rolandelli RH, Luján
JJ, Haskó G, Csóka B, Selmeczy Z and Németh ZH: HIF-1: A key
mediator in hypoxia. Acta Physiol Hung. 96:19–28. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chinnadurai G, Vijayalingam S and Gibson
SB: BNIP3 subfamily BH3-only proteins: Mitochondrial stress sensors
in normal and pathological functions. Oncogene. 27(Suppl 1):
S114–S127. 2008. View Article : Google Scholar
|
|
54
|
Lu N, Li X, Tan R, An J, Cai Z, Hu X, Wang
F, Wang H, Lu C and Lu H: HIF-1α/beclin1-mediated autophagy is
involved in neuroprotection induced by hypoxic preconditioning. J
Mol Neurosci. 66:238–250. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen R, Jiang T, She Y, Xu J, Li C, Zhou
S, Shen H, Shi H and Liu S: Effects of cobalt chloride, a
hypoxia-mimetic agent, on autophagy and atrophy in skeletal C2C12
myotubes. Biomed Res Int. 2017:70975802017.PubMed/NCBI
|
|
56
|
Qian X, Zhu M, Qian W and Song J: Vitamin
D attenuates myocardial ischemia-reperfusion injury by inhibiting
inflammation via suppressing the RhoA/ROCK/NF-κB pathway.
Biotechnol Appl Biochem. 66:850–857. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Vinten-Johansen J, Jiang R, Reeves JG,
Mykytenko J, Deneve J and Jobe LJ: Inflammation, proinflammatory
mediators and myocardial ischemia-reperfusion Injury. Hematol Oncol
Clin North Am. 21:123–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhao H, Liu M, Liu H, Suo R and Lu C:
Naringin protects endothelial cells from apoptosis and inflammation
by regulating the hippo-YAP pathway. Biosci Rep.
40:BSR201934312020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sun LJ, Qiao W, Xiao YJ, Cui L, Wang X and
Ren WD: Naringin mitigates myocardial strain and the inflammatory
response in sepsis-induced myocardial dysfunction through
regulation of PI3K/AKT/NF-κB pathway. Int Immunopharmacol.
75:1057822019. View Article : Google Scholar
|