Introduction
Lung cancer is the leading cause of cancer-related
mortality worldwide (1-3), and the morbidity and mortality
rates for the disease remain high. Over the past two decades,
significant advancements have been achieved in the diagnosis and
treatment of lung cancer, which has relied on the improved
understanding of the disease biology and the underlying mechanisms
of lung cancer progression and metastasis, as well as the
advancements in early detection methods and multimodal care
(1). However, despite this
progress, the prognosis of lung cancer remains unsatisfactory and
the underlying mechanisms of lung cancer remain poorly understood.
Therefore, further studies investigating the underlying mechanisms
of lung cancer and identifying novel targets to devise novel drugs,
as well as combination therapies, are required to improve the
outcomes of patients with lung cancer.
DnaJ heat shock protein family (HSP40) member C12
(DNAJC12) is a J domain-containing protein that belongs to the DnaJ
homology C (DNAJC) family, which is a subclass of heat shock
proteins (4,5). To the best of our knowledge, the
current physiological and pathological functions of DNAJC12 remain
unclear. During endoplasmic reticulum (ER) stress, the binding of
the cochaperone DNAJC12 and heat shock protein family A (HSP70)
member 8 is enhanced (6).
Several genetic studies have revealed that DNAJC12 participates in
the development of numerous types of human disease. For example,
biallelic mutations in the DNAJC12 gene in humans have been shown
to lead to dystonia and intellectual disability (7). In addition, DNAJC12 mutations
identified by whole-exome sequencing have been observed in patients
with mild hyperphenylalaninemia (8,9).
In the Chinese Han population, DNAJC12 mutations have also been
found to be associated with Parkinson's disease (10).
Over the past few years, the roles of DNAJC12 in
cancer biology have also been reported. For instance, in patients
with rectal cancer, upregulated expression levels of DNAJC12 have
been shown to predict a poor response to neoadjuvant concurrent
chemoradiotherapy (11). The
aggressive phenotype of gastric cancer has also been found to be
associated with the upregulation of DNAJC12 expression levels
(12). In addition, in breast
cancer, DNAJC12 expression has been found to be associated with the
estrogen receptor status (13).
However, the association between DNAJC12 and lung cancer remains
unknown. Moreover, although the association between DNAJC12 and
several types of cancer has been reported, to the best of our
knowledge, the biological functions of DNAJC12 in cancer
development and metastasis also remain unknown.
β-catenin is a 90 kDa multifunctional protein that
participates in cell development under normal physiological
conditions (14,15). β-catenin is a pivotal
transcriptional factor for Wnt signaling and plays an important
role in stem cell self-renewal and organ regeneration, in addition
to cancer development and drug resistance (16). The roles of β-catenin in lung
cancer have been well established. For example, β-catenin has been
discovered to promote tumorigenesis, cancer stem cell self-renewal,
drug resistance and metastasis in lung cancer cells by promoting
the activation of Wnt signaling and the expression of downstream
target genes (17,18). In addition, β-catenin expression
has been reported to be induced by epidermal growth factor receptor
(EGFR) mutations and to contribute to lung cancer development
(19). However, the mechanisms
through which β-catenin responds to upstream regulators remain only
partially understood and upstream regulators of β-catenin also
remain to be identified.
The present study thus aimed to investigate the
roles of DNAJC12 in lung cancer. The findings presented herein
demonstrate that DNAJC12 promotes lung cancer cell tumorigenesis by
regulating the expression and activation of β-catenin.
Materials and methods
Patient samples
A total of 15 pairs of lung cancer and adjacent
non-cancer tissues were obtained from patients with lung cancer
(age range, 45-76 years; sex, 9 males and 6 females) between
January, 2019 and February, 2020 from the Shandong Provincial
Hospital (Jinan, China). The samples were stored at −80°C until
required for subsequent experimentation. Written informed consent
was obtained from each patient prior to participation and the
clinical study was approved by the Ethics Committee of Clinical
Research of Shandong Provincial Hospital.
For the analysis using public data, 57 pairs of lung
cancer and adjacent non-cancer tissues were analyzed using a
dataset from The Cancer Genome Atlas (TCGA) and the Genotype-Tissue
Expression (GTEx) databases. In addition, the expression levels of
DNAJC12 were also analyzed in 54 control and 498 cancer unpaired
tissues.
Cell lines and culture
Lung cancer cell lines (A549, NCI-H1299, NCI-H1975
and 95D) and 293T cells were purchased from the American Type
Culture Collection (ATCC). Mycoplasma testing was performed for all
cell lines and all cell lines were authenticated using STR
profiling. Cells were cultured in DMEM (HyClone; Cytiva)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin (Thermo Fisher Scientific, Inc.).
All cells were maintained at 37°C in a humidified atmosphere
containing 5% CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cancer tissues and
cells using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA (1 μg) was reverse transcribed
into cDNA using a cDNA synthesis kit (cat. no. 6130, Takara Bio,
Inc.). qPCR was subsequently performed using an SYBR-Green II
reagent kit (cat. no. RR820A; Takara Bio, Inc.) as previously
described (20). The following
thermocycling conditions were used for the qPCR: Initial
denaturation at 95°C for 5 min, followed by 40 cycles at 95°C for
15 sec, 60°C for 30 sec and 70°C for 10 sec. The following primer
pairs were used for the qPCR: DNAJC12 forward,
5′-AATGGTTGGCACCTTCGTTTC-3′ and reverse,
5′-GTTGGCAGCATAGGGGACAG-3′; CTNNB1 forward,
5′-AGCTTCCAGACACGCTATCAT-3′ and reverse,
5′-CGGTACAACGAGCTGTTTCTAC-3′; VIM forward,
5′-AGTCCACTGAGTACCGGAGAC-3′ and reverse,
5′-CATTTCACGCATCTGGCGTTC-3′; RELA forward,
5′-GTGGGGACTACGACCTGAATG-3′ and reverse,
5′-GGGGCACGATTGTCAAAGATG-3′; and GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′. The mRNA expression levels were
quantified using the 2−ΔΔCq method as previously
described (21).
Lentiviral transfection
Short hairpin RNA (shRNA/sh) targeting DNAJC12
(shDNAJC12; 5′-GGATGTGATGAACTATCTT-3′) and control shRNA (shCtrl;
5′-TTCTCCGAACGTGTCACGT-3′) were cloned into GV115 plasmids
(Shanghai GeneChem Co., Ltd.). For overexpression, the coding
sequences of CTNNB1 (NM_001904.4), p65 (NM_021975.4), VIM
(NM_003380.5) were cloned into GV610 vectors (Shanghai GeneChem
Co., Ltd.). The GV115 or GV610 plasmids (20 μg) were
subsequently co-transfected into 293T cells alongside pHelper1.0
(15 μg) and pHelper2.0 (10 μg) packaging vectors
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) as previously described (22). Following transfection with the
lentiviruses for 48 h (MOI=10), the transduced cells were selected
with 5 μg/ml puromycin (Sigma-Aldrich; Merck KGaA).
High-content screening (HCS) assay
A HCS assay was performed to determine the cell
number by counting the GFP-expressing cells, as previously
described (22), following the
culture of 2×103 A549 cells in 96-well plates.
ArrayScan™ HCS software (Cellomics Inc.) was used to analyze the
cell proliferation of each well every 24 h.
MTT proliferation assay
The proliferation of the A549 and NCI-H1975 cells
seeded into 96-well plates at a density of 2×103
cells/well was analyzed using an MTT assay kit (Gen-view
Scientific, Inc.). MTT solution was added to each well and
incubated for 4 h at 37°C. Subsequently, the solution was replaced
with 100 μl/well DMSO, and the plates were agitated for 5
min. The optical density of each well was read at 490 nm using a
microplate spectrophotometer (Tecan infinite, M2009PR; Tecan Group,
Ltd.).
Colony formation assay
A total of 1×103 A549 and NIC-H1975 cells
with or without DNAJC12 knockdown were plated into 6-well plates
and cultured for 2 weeks. Colonies were subsequently stained with
crystal violet (Sangon Biotech Co., Ltd.) and stained colonies were
visualized, followed by evaluation of colony number and size.
Wound healing assay
A549 and NCI-H1975 cell migration was determined in
6-well plates (105 cells/well) using a wound healing
assay, as previously described (23). Briefly, the cells were used in
the wound healing assay when the confluency of the cells was closed
to 100%. After the cells were scratched using 10 μl pipette
tips, the medium was replaced with FBS-free medium. The images were
captured at 0, 24 and 48 h using an inverted fluorescence
microscope (Olympus Corporation; magnification, ×100).
Cell migration and invasion assay
A Transwell migration and Matrigel invasion assay
was used to determine the migration and invasive ability of the
A549 and NCI-H1975 cells. For migration, 100 μl FBS free
medium were added onto the upper surface of the migration chambers
(cat. no. 3422; Corning, Inc.) and incubated at 37°C for 30 min.
Subsequently, the FBS-free medium was discarded and a total of
1×105 cells in 100 μl FBS-free medium was added
onto the upper surface of the migration chambers. The lower
chambers were filled with 30% FBS medium. For invasion assay, a
total of 500 μl of the mixture was added onto the upper
surface of the invasion chambers (cat. no. 354480; Corning, Inc.),
while the lower chambers were filled with 30% FBS medium. Following
incubation for 24 or 48 h at 37°C, the culture medium and the cells
attached on the upper surface were removed. Cells attached on the
lower surface were fixed with 4% paraformaldehyde for 30 min at
room temperature and stained with Giemsa stain (cat. no. 32884;
Sigma-Aldrich; Merck KGaA) for 30 min at room temperature. The
images of migratory and invasive cells were collected using a light
microscope (Olympus Corporation; magnification, ×100).
Apoptosis assay
The apoptosis of the lung cancer cells was analyzed
using an Annexin V-allophycocyanin (APC) apoptosis detection kit
(eBioscience; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Briefly, the A549 and NCI-H1975 cells were
infected with lentiviruses expressing shCtrl or shDNAJC12 for 24 h.
The cells were then harvested (1,000 × g, 5 min, room temperature)
and washed with PBS. The cells were subsequently resuspended in a
staining buffer to a final density of 1×106 cells/ml.
Finally, 100 μl cell suspension was incubated with 5
μl Annexin V-APC at room temperature for 15 min. The stained
cells were subjected to flow cytometry using a FACSCalibur flow
cytometer (BD Biosciences) and data were analyzed with FlowJo
software (version 7.6.1; FlowJo LLC).
In vivo tumor growth experiment
Female BALB/c athymic nude mice (age, 4 weeks;
weight, 15-17 g) were obtained from Charles River Laboratories,
Inc. and were randomly divided into the following two groups (10
mice/group): i) ShCtrl; and ii) shDNAJC12. Mice were healthy and
housed under SPF-grade conditions. A total of 1×107
shCtrl- or shDNAJC12-transfected A549 cells were subcutaneously
implanted into the right flanks of the nude mice, according to the
protocol as described previously (24-26). The tumor volume was measured
every 3 days starting at 30 days following transplantation and the
mice were sacrificed by cervical dislocation at day 42. The animal
experiments were performed according to the indicated protocols and
were approved by the Animal Research Ethics Committee of Shandong
Provincial Hospital.
Western blot analysis
Total protein was extracted from the lung cancer
tissues and A549 and NCI-H1975 cells using RIPA lysis buffer
(Beyotime Institute of Biotechnology) supplemented with protease
inhibitor cocktail. The protein concentration was determined using
a BCA assay kit (Thermo Fisher Scientific, Inc.) and 40 μg
protein/lane was separated via 12% SDS-PAGE. The separated proteins
were subsequently transferred onto PDVF membranes and blocked with
5% fat-free milk in TBS-Tween-20 (TBST) for 1 h at room
temperature. The membranes were then washed with TBST and incubated
with the following primary antibodies at 4°C overnight:
Anti-DNAJC12 (Abcam; cat. no. ab167425), anti-GAPDH (Santa Cruz
Biotechnology, Inc.; cat. no. sc-32233), anti-p65 (Cell Signaling
Technology, Inc.; cat. no. 8242), anti-phosphorylated (p)-p65 (Cell
Signaling Technology, Inc.; cat. no. 3033), anti-β-catenin (Cell
Signaling Technology, Inc.; cat. no. 8480), anti-p-β-catenin (Cell
Signaling Technology, Inc.; cat. no. 2009) and anti-vimentin (Cell
Signaling Technology, Inc.; cat. no. 3932). Following the primary
antibody incubation, the membranes were washed with TBST and
incubated with HRP-conjugated anti-mouse or anti-rabbit secondary
antibodies (Cell Signaling Technology, Inc.; cat. nos. 7076 and
7074) at room temperature for 2 h. The membranes were subsequently
washed with TBST and protein bands were visualized using Pierce™
ECL Western Blotting substrate (Thermo Fisher Scientific,
Inc.).
Statistical analysis
All experiments were performed at least in
triplicate and data are presented as the mean ± SEM. Statistical
differences between two groups were determined using a Student's
t-test, while statistical differences between ≥3 groups were
determined using a one-way ANOVA followed by a Tukey's post hoc
test. Categorical data were analyzed using a χ2 test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
DNAJC12 expression levels are upregulated
in lung cancer tissues
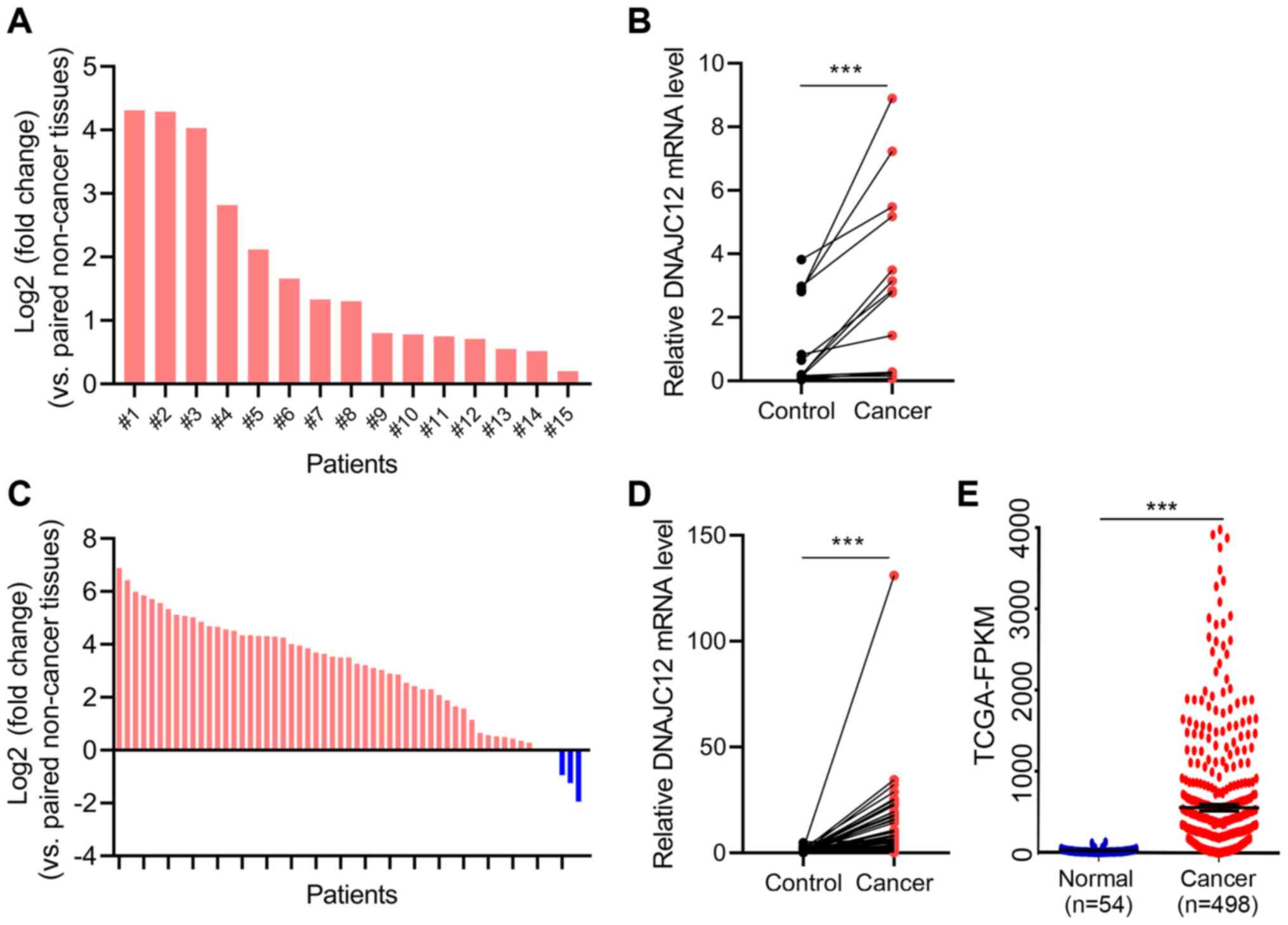
To determine the potential roles of DNAJC12 in lung
cancer, the expression levels of DNAJC12 in lung cancer and
adjacent non-cancer tissues were analyzed by RT-qPCR. The
expression levels of DNAJC12 were significantly upregulated in the
lung cancer tissues compared with the adjacent non-cancer tissues
(Fig. 1A and B). In addition,
the expression levels of DNAJC12 in 57 pairs of lung cancer and
control tissues from TCGA database were analyzed. The results also
revealed that the expression levels of DNAJC12 were upregulated in
lung cancer tissues compared with non-cancer control lung tissues
(Fig. 1C and D). Furthermore,
analysis of a large unpaired TCGA dataset (54 control and 498
cancer tissues) also demonstrated that the expression levels of
DNAJC12 were upregulated in lung cancer tissues (Fig. 1E). In addition, the association
between patient clinicopathological features and DNAJC12 expression
in 498 lung cancer tissues from TCGA database is presented in
Table I. The expression of
DNAJC12 was not associated with age, sex, T stage, N metastasis and
M metastasis. Therefore, these findings suggested that DNAJC12 may
be involved in the development of lung cancer.
 | Table IAssociation between patient
clinicopathological features and DNAJC12 expression in 498 lung
cancer tissues from TCGA database. |
Table I
Association between patient
clinicopathological features and DNAJC12 expression in 498 lung
cancer tissues from TCGA database.
|
Characteristics | DNAJC 12
| Total | P-value |
|---|
| Low | High |
|---|
| Sex | | | | 0.151 |
| Male | 107 | 123 | 230 | |
| Female | 142 | 126 | 268 | |
| Total | 249 | 249 | 498 | |
| Age, years | | | | 0.744 |
| ≤65 | 119 | 113 | 232 | |
| >65 | 123 | 124 | 247 | |
| Total | 242 | 237 | 479 | |
| T stage | | | | 0.852 |
| T1/2 | 214 | 218 | 432 | |
| T3/4 | 32 | 31 | 63 | |
| Total | 246 | 249 | 495 | |
| N metastasis | | | | 0.876 |
| N0 | 160 | 161 | 321 | |
| N1/2/3 | 82 | 85 | 167 | |
| Total | 242 | 246 | 488 | |
| M metastasis | | | | 0.555 |
| M0 | 166 | 167 | 333 | |
| M1 | 10 | 13 | 23 | |
| Total | 176 | 180 | 356 | |
Knockdown of DNAJC12 suppresses lung
cancer cell proliferation in vitro
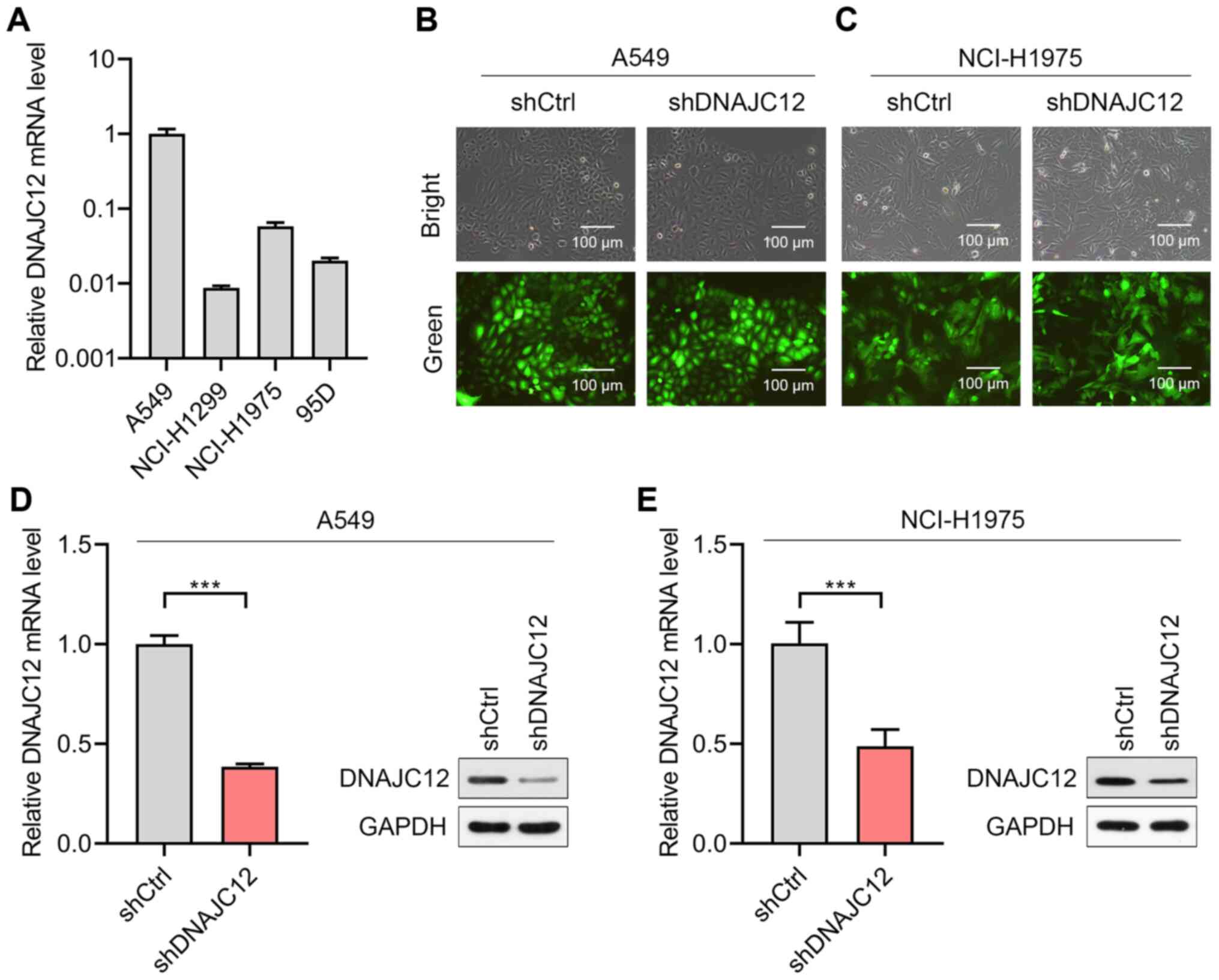
To determine the potential roles of DNAJC12 in lung
cancer cells, the expression levels of DNAJC12 were first analyzed
in four lung cancer cell lines. The results revealed that the A549
and NCI-H1975 cells expressed higher expression levels of DNAJC12
compared with the NCI-H1299 and 95D cells (Fig. 2A). Therefore, the A549 and
NCI-H1975 cells were selected for use in further experiments.
Lentiviruses expressing shRNA targeting DNAJC12 (shDNAJC12) and
control shRNA (shCtrl) were subsequently successfully infected into
A549 and NCI-H1975 cells (Fig. 2B
and C). The results of RT-qPCR and western blot analysis
demonstrated that the lentivirus-mediated knockdown of DNAJC12
significantly downregulated the expression levels of DNAJC12 in the
A549 and NCI-H1975 lung cancer cells (Fig. 2D and E). Thus, A549 and NCI-H1975
cells with/without stable DNAJC12 knockdown were successfully
generated for use in further experiments.
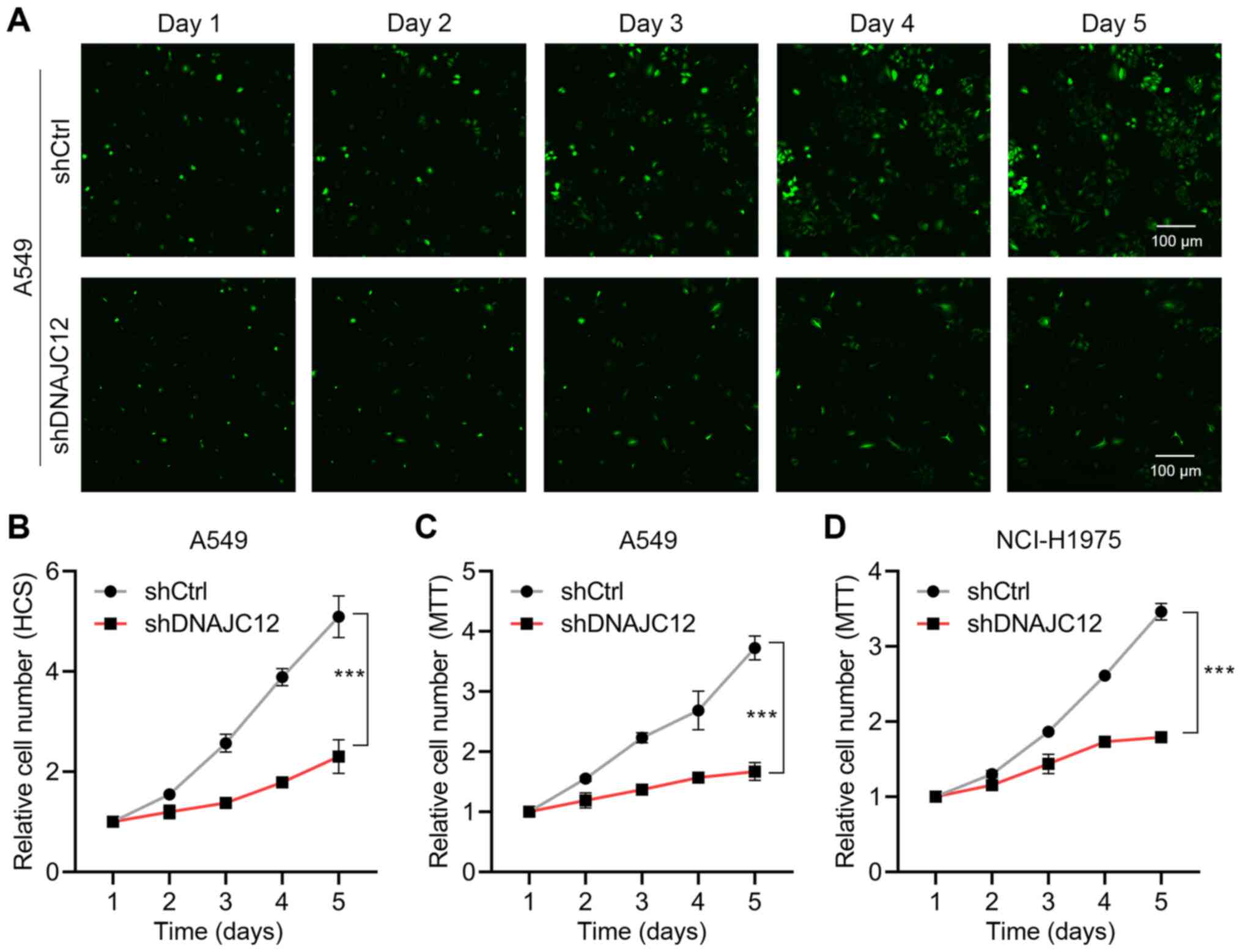
The present study first determined the effects of
the knockdown of DNAJC12 on the proliferation rate of A549 and
NCI-H1975 lung cancer cells using HCS and MTT assays. The results
determined that the knockdown of DNAJC12 significantly reduced the
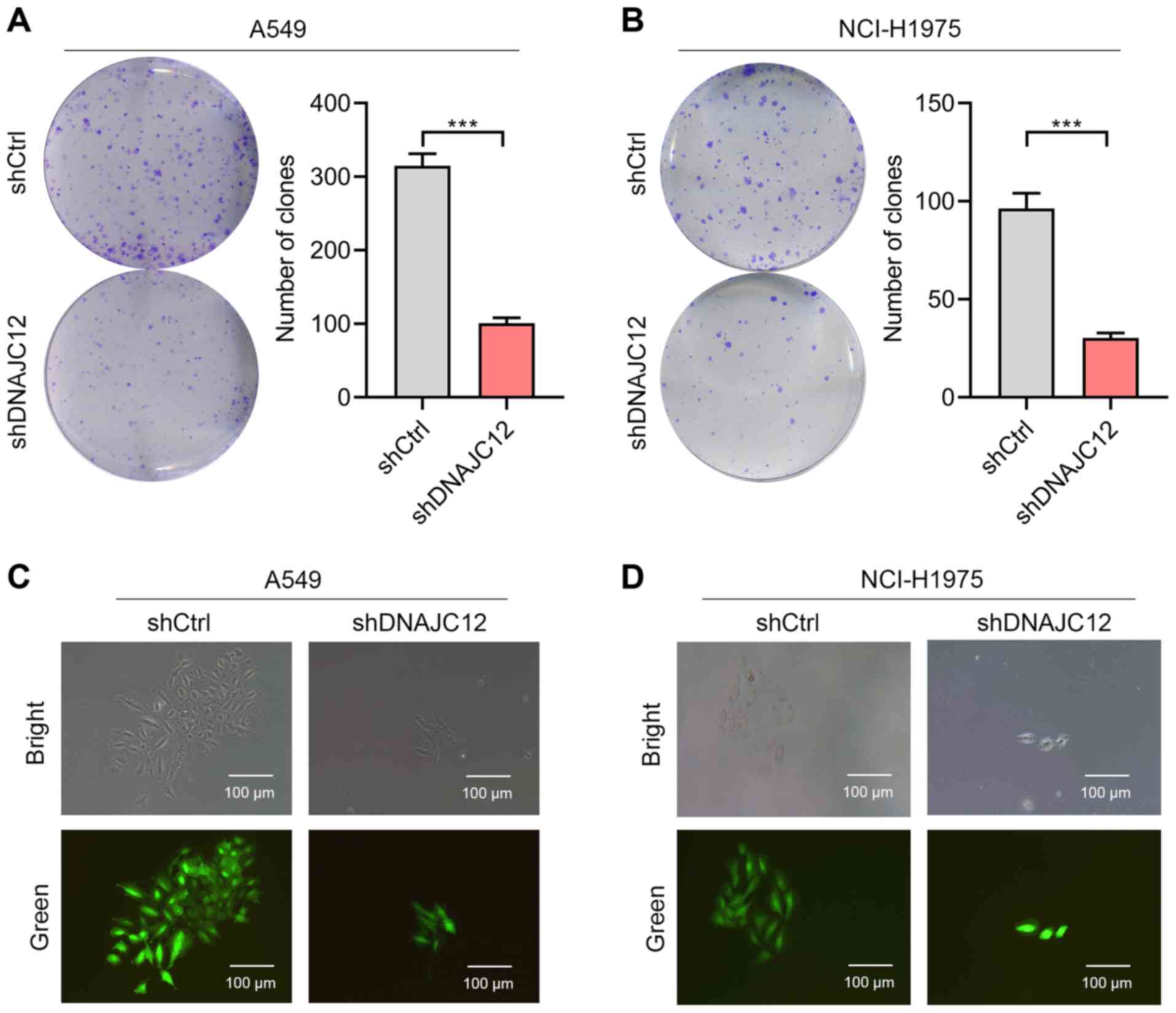
proliferation rate of the A549 and NCI-H1975 cells (Fig. 3). Colony formation is a key
feature of cancer cells (27);
therefore, the present study also analyzed the effects of DNAJC12
on the colony formation of lung cancer cells. The results revealed
that DNAJC12 knockdown reduced the number and size of colonies
formed from the A549 and NCI-H1975 lung cancer cells (Fig. 4). Taken together, these findings
suggest that the knockdown of DNAJC12 may suppress the
proliferation of lung cancer cells.
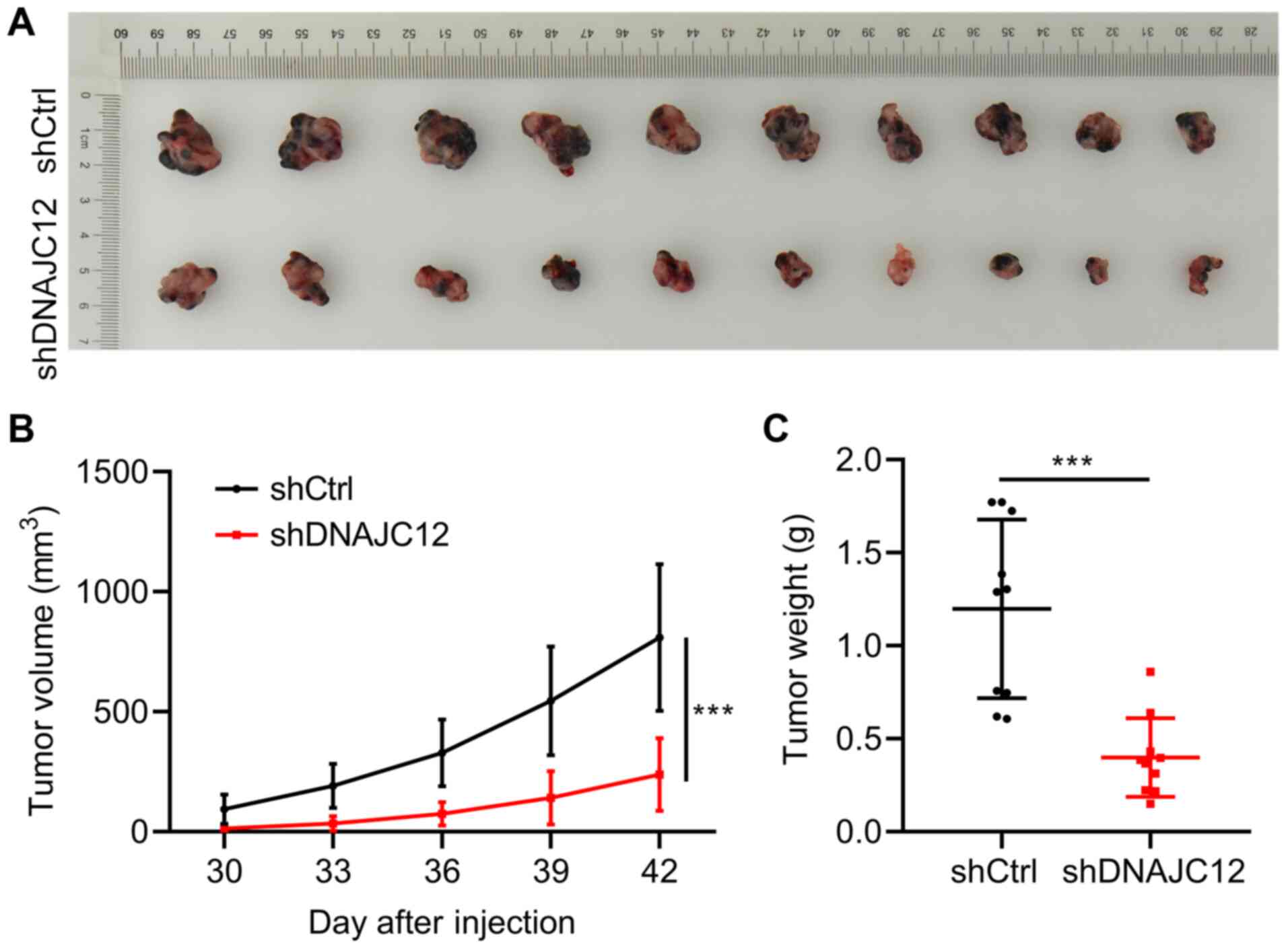
Knockdown of DNAJC12 suppresses lung
cancer cell growth in vivo
To determine whether DNAJC12 regulates lung cancer
cell growth in vivo, xenograft experiments were performed.
A549 cells expressing shCtrl and shDNAJC12 lentiviruses were
subcutaneously transplanted into the right flanks of 4-week-old
female nude mice. Tumor growth was monitored for 42 days following
implantation. The results revealed that the knockdown of DNAJC12
reduced the growth rate, size and weight of the lung cancer tumors
(Fig. 5). Thus, these results
indicate that DNAJC12 may contribute to lung cancer growth in
vivo.
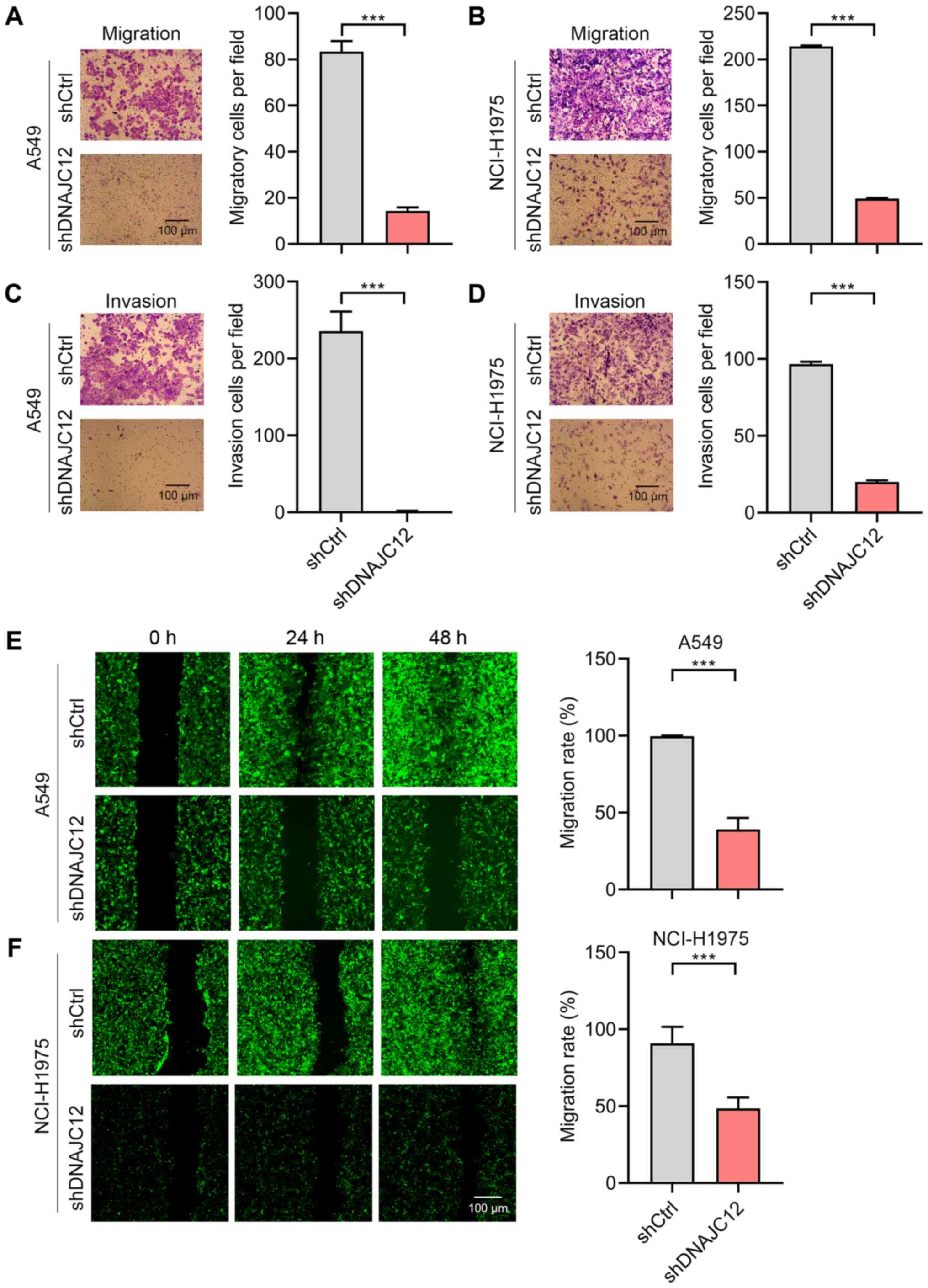
Knockdown of DNAJC12 suppresses lung
cancer migration and invasion
Metastasis is a feature of human lung cancer cells
(27). To determine whether
DNAJC12 regulates the invasion of lung cancer cells, the Transwell
assay and Transwell Matrigel assay was performed using the A549 and
NCI-H1975 cells. The results revealed that A549 and NCI-H1975 cell
invasion was significantly suppressed following the knockdown of
DNAJC12 (Fig. 6A and B).
Matrigel invasion assay also demonstrated that DNAJC12 knockdown
suppressed the invasion of A549 and NCI-H1975 cells (Fig. 6C and D). The present study
subsequently analyzed the effects of DNAJC12 on the migration of
lung cancer cells by performing wound healing assays. The results
revealed that the migration of the A549 and NCI-H1975 cells, which
was observed within 48 h, was inhibited by DNAJC12 knockdown
(Fig. 6E and F). Taken together,
these findings suggest that DNAJC12 regulates the invasion and
migration of lung cancer cells.
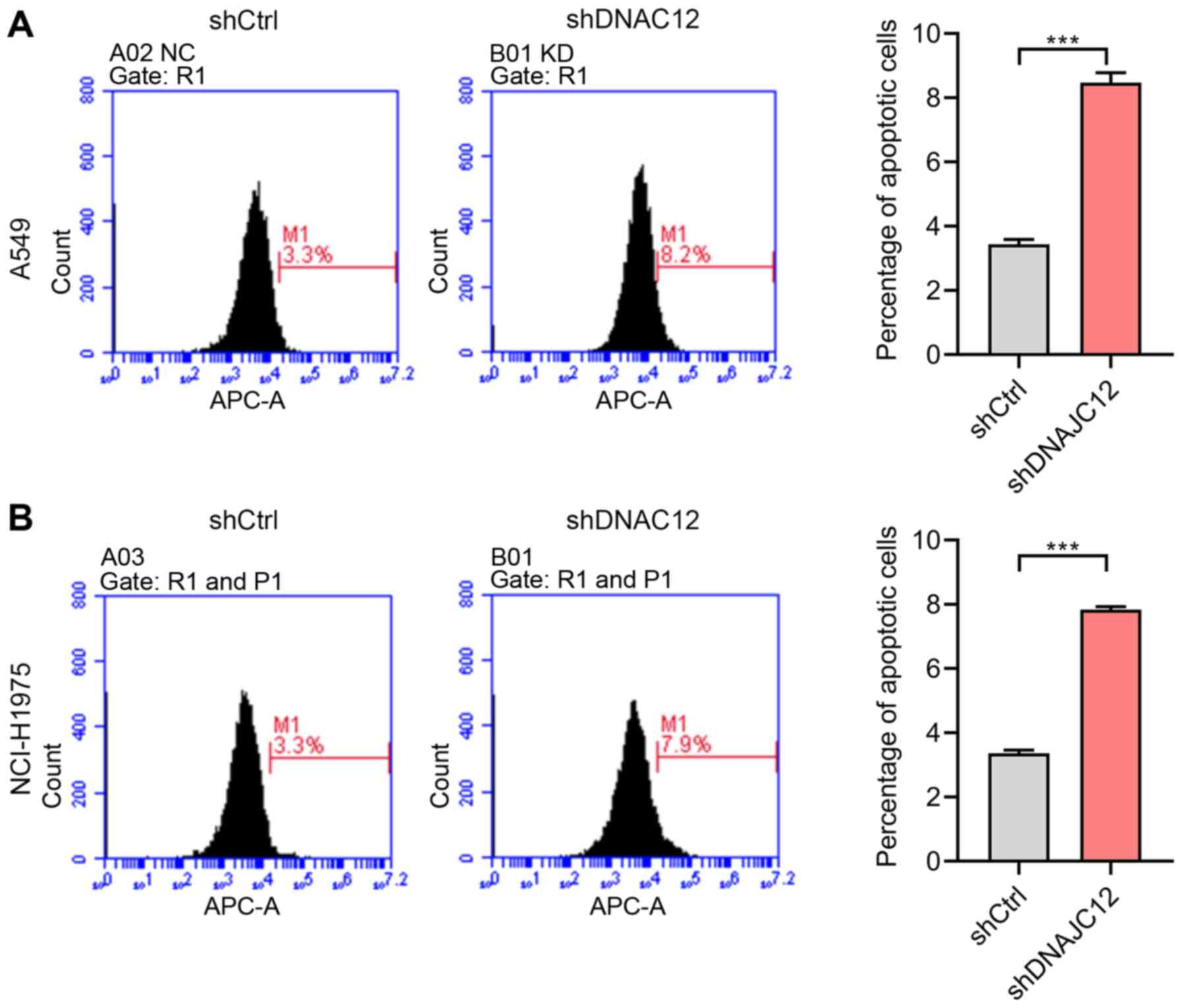
Knockdown of DNAJC12 induces the
apoptosis of lung cancer cells
The suppression of cancer growth and metastasis may
result from cancer cell apoptosis induced by clinical drug
treatment (27). Thus, the
present study aimed to determine whether the DNAJC12-induced
effects on the proliferation, migration and invasion of lung cancer
cells were associated with the apoptosis of lung cancer cells. Cell
apoptosis was analyzed by flow cytometry in the A549 and NCI-H1975
cells. The results revealed that the knockdown of DNAJC12 induced
the apoptosis of A549 and NCI-H1975 cells (Fig. 7), which may be associated with
the effects of DNAJC12 on the proliferation, migration and invasion
of lung cancer cells.
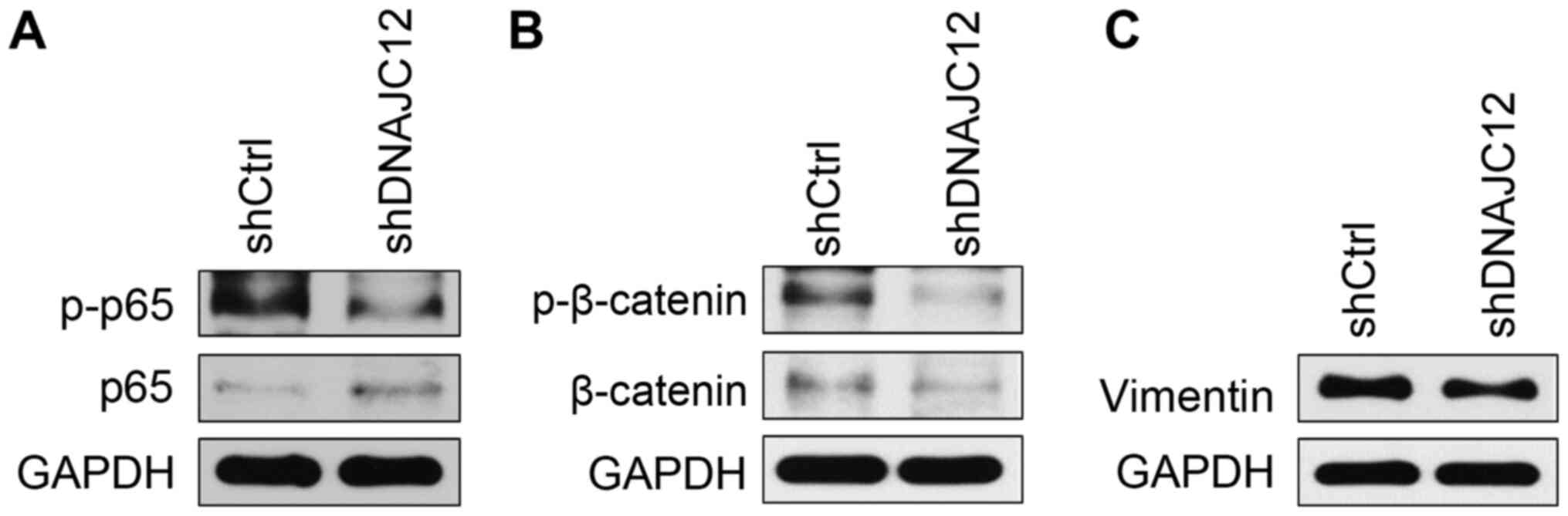
DNAJC12 regulates NF-κB, β-catenin and
vimentin signaling
The biological pathways regulated by DNAJC12 were
subsequently investigated. The growth and metastasis of lung cancer
cells are regulated by pivotal intracellular transcriptional
regulators, such as NF-κB and β-catenin, as well as key
extracellular matrix regulators, such as matrix metalloproteinase
(MMP) and vimentin (14,28,29). The present study analyzed the
effects of DNAJC12 on the expression levels of these proteins by
western blotting. The knockdown of DNAJC12 reduced the
phosphorylation of NF-κB p65 and downregulated the expression
levels and activation of β-catenin in A549 cells (Fig. 8A and B). The knockdown of DNAJC12
also downregulated the expression levels of vimentin (Fig. 8C), which is a factor of the
extracellular matrix. Therefore, these findings suggest that
DNAJC12 may target key regulators of lung cancer growth and
metastasis.
Overexpression of β-catenin blocks the
effects of DNAJC12 knockdown
To determine whether p65, β-catenin and vimentin are
involved in the effects of DNAJC12 on lung cancer cells, p65,
β-catenin and vimentin were overexpressed in A549 cells in which
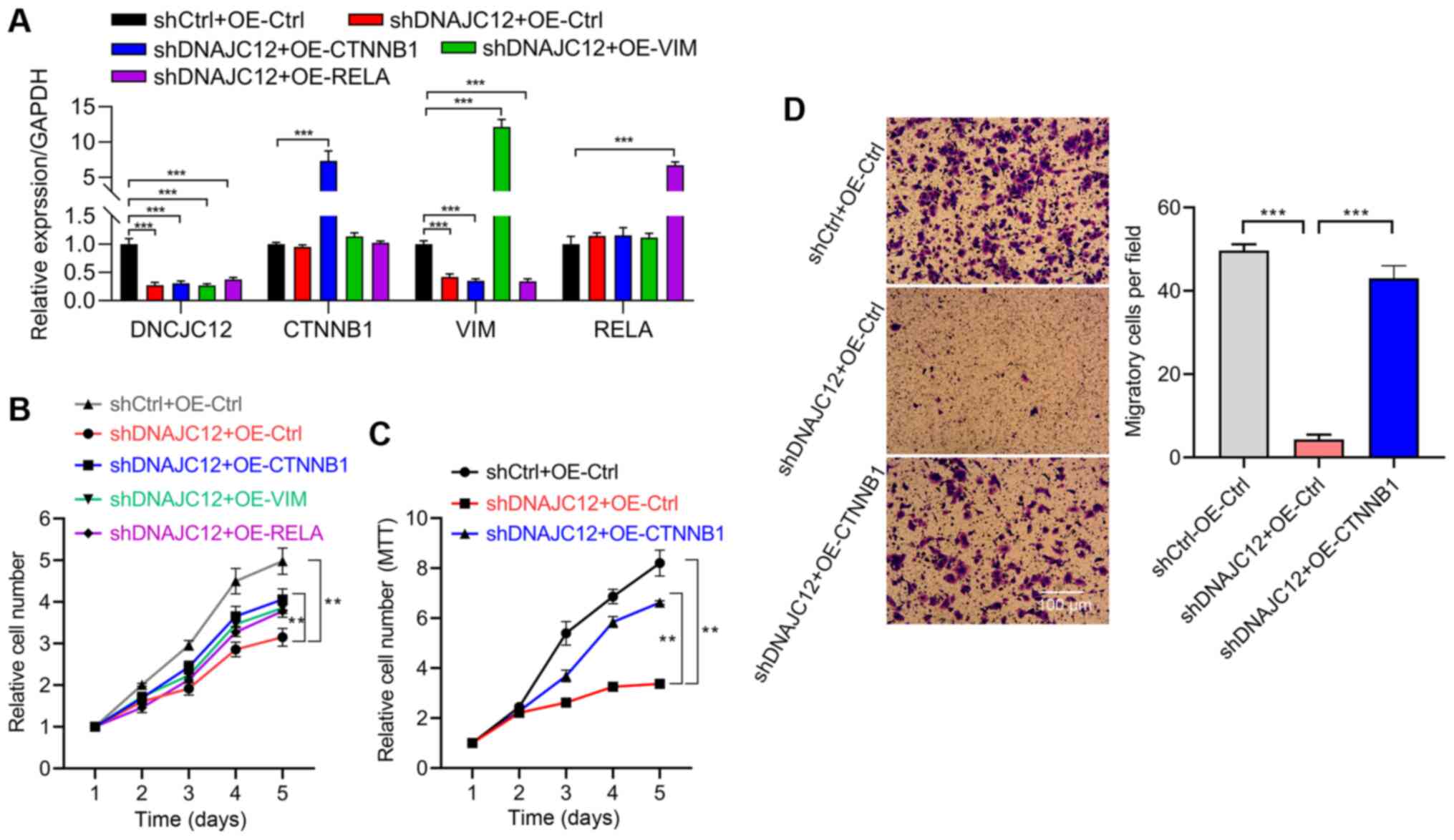
DNAJC12 expression was knocked down. RT-qPCR analysis indicated
that DNAJC12 was successfully knockdown, while p65, β-catenin and
vimentin were effectively overexpressed in the A549 cells (Fig. 9A). An HCS assay was subsequently
performed and the results revealed that the overexpression of
β-catenin, but not that of p65 or vimentin, reversed the
suppressive effects of DNAJC12 knockdown on the proliferation of
A549 cells (Fig. 9B). The
involvement of β-catenin in the regulatory role of DNAJC12 over
A549 cell proliferation was also validated using an MTT assay
(Fig. 9C). Furthermore, the
reduced invasive capacity of the A549 cells induced by DNAJC12
knockdown was also reversed by the overexpression of β-catenin
(Fig. 9D). Taken together, these
findings suggest that DNAJC12 may regulate the proliferation and
invasion of lung cancer cells in a β-catenin-dependent manner.
Discussion
The present study provides evidence to suggest that
DNAJC12 may promote lung cancer proliferation, invasion, migration
and survival, which may be partially mediated by β-catenin. Using
data generated in the present study and public data, the DNAJC12
expression levels were found to be upregulated in lung cancer
tissues compared with non-cancerours tissues. The
lentivirus-mediated knockdown of DNAJC12 demonstrated that DNAJC12
was important for lung cancer cell proliferation and growth in
vitro and in vivo, respectively. In addition, the
knockdown of DNAJC12 reduced the migration and invasion, and
induced the apoptosis in lung cancer cells. Signaling pathway
analysis revealed that DNAJC12 targeted NF-κB, β-catenin and
vimentin signaling. Finally, rescue experiments revealed that
β-catenin was essential for the function of DNAJC12 in regulating
the proliferation, migration and invasion of lung cancer cells.
DNAJC12 belongs to the DNAJC protein family of heat
shock proteins. The potential association between DNAJC12 and
cancer biology has been previously reported (11,12). However, to the best of our
knowledge, the role of DNAJC12 in lung cancer remains unknown.
Therefore, the present study investigated the potential roles of
DNAJC12 in lung cancer. Using patient lung cancer samples and
public data from the TCGA database, the present results
demonstrated that DNAJC12 mRNA expression levels were significantly
upregulated in lung cancer tissues compared with adjacent
non-cancer lung tissues. Furthermore, DNAJC12 expression was
significantly elevated in lung cancer patients with higher
stage.
Although previous studies have reported the
potential association between DNAJC12 and numerous types of cancer
(11-13), the role of DNAJC12 in cancer
biology remains unknown. The present study performed HCS and MTT
assays, which revealed that the knockdown of DNAJC12 reduced the
proliferative rate of lung cancer cells. Colony formation is a key
feature of cancer cells (27).
The present study also demonstrated that DNAJC12 not only regulated
the number of colonies formed, but it also altered the size of the
colonies formed from A549 and NCI-H1975 cells. Notably, the data
presented herein also demonstrated that DNAJC12 knockdown reduced
lung cancer cell growth in vivo. Taken together, these
findings suggested that DNAJC12 may be crucial for lung cancer cell
proliferation and growth in vitro and in vivo,
respectively.
The underlying mechanisms of lung cancer metastasis
remain poorly understood (30).
To determine the potential role of DNAJC12 in the metastasis of
lung cancer cells, the present study performed in vitro
Transwell and wound healing assays to assess cell invasion and
migration, respectively. The results revealed that the knockdown of
DNAJC12 significantly reduced the invasive and migratory capacities
of A549 and NCI-H1975 cells. These findings indicated that the
upregulated expression of DNAJC12 in lung cancer tissues might be
one of the underlying mechanisms of metastasis in lung cancer
cells.
Following clinical drug treatment, the suppression
of cancer growth and metastasis occurs as a result of cancer cell
apoptosis (27). Therefore, the
present study also aimed to determine whether the effects of
DNAJC12 on lung cancer proliferation, invasion and migration were
associated with cell apoptosis in A549 and NCI-H1975 cells using
flow cytometry. The present data demonstrated that DNAJC12
knockdown significantly induced the apoptosis of A549 and NCI-H1975
cells. These results suggested that DNAJC12 may regulate the
apoptosis of lung cancer cells to modulate cell proliferation,
migration and invasion.
NF-κB plays a crucial role in lung cancer by
regulating growth and apoptosis (29). The findings of the present study
demonstrated that the knockdown of DNAJC12 reduced the
phosphorylation levels of the NF-κB p65 subunit, suggesting that
DNAJC12 may promote the activation of NF-κB. β-catenin has been
reported to regulate several functions of lung cancer, including
growth, stemness, metastasis and drug resistance (resistance to
apoptosis) (17). It was also
found that DNAJC12 knockdown downregulated the expression levels
and phosphorylation of β-catenin. Vimentin is a mesenchymal marker
that participates in the remodeling of the extracellular matrix and
contributes to the metastasis of numerous types of cancer,
including lung cancer (28).
Thus, the present study also aimed to determine the effects of
DNAJC12 on the expression levels of vimentin. The knockdown of
DNAJC12 also downregulated the expression levels of vimentin. These
data suggested that DNAJC12 may regulate several key modulators of
proliferation, migration and invasion in lung cancer cells. To
investigate which of these factors was a key downstream target gene
of DNAJC12 in lung cancer, rescue experiments were performed by
overexpressing these factors in lung cancer cells following DNAJC12
knockdown. Further rescue experiments indicated that β-catenin, but
not NF-κB or vimentin, may be involved in DNAJC12 function in lung
cancer cell proliferation, invasion and migration. However, the
biological mechanisms through which DNAJC12 regulated β-catenin
expression and activation remain to be determined in further
studies. A previous study reported that DNAJC12 participated in ER
stress (6). ER stress is an
important regulator involved in the activation of β-catenin
(31), thus DNAJC12 may regulate
β-catenin via activating ER stress. Further study is required to
determine this hypothesis.
In conclusion, the findings of the present study
suggested that DNAJC12 may promote lung cancer development and
metastatic features, at least in part, by activating β-catenin.
Thus, DNAJC12 may serve as a potential prognostic factor and
therapeutic target in human lung cancer. However, several
limitations exist for the present study. First, the mechanism
through which DNAJC12 was regulated in lung cancer remains unknown.
Second, the potential effects of DNAJC12 on the prognosis of
patients with lung cancer were not determined. Finally, the
underlying mechanisms of DNAJC12 regulation over β-catenin remain
unknown. Further studies addressing these points would improve the
current understanding of the role and mechanism of DNAJC12 in lung
cancer.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL designed the study, and drafted and revised the
manuscript. ML and FJ performed the majority of the experiments. JL
and MC analyzed and interpreted the data. JY performed the animal
experiments. All authors have read and approved the final
manuscript. All authors confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The clinical study was approved by the Ethics
Committee of Clinical Research of Shandong Provincial Hospital and
written informed consent was obtained from each patient prior to
participation. The animal experiments were performed according to
the indicated protocols and were approved by the Animal Research
Ethics Committee of Shandong Provincial Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
1
|
Long K and Suresh K: Pulmonary toxicity of
systemic lung cancer therapy. Respirology. 25(Suppl 2): 72–79.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Minna JD, Roth JA and Gazdar AF: Focus on
lung cancer. Cancer Cell. 1:49–52. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tang X, Mo C, Wang Y, Wei D and Xiao H:
Anti-tumour strategies aiming to target tumour-associated
macrophages. Immunology. 138:93–104. 2013. View Article : Google Scholar :
|
|
4
|
Roosen DA, Blauwendraat C, Cookson MR and
Lewis PA: DNAJC proteins and pathways to parkinsonism. Febs J.
286:3080–3094. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zylicz M, Yamamoto T, McKittrick N, Sell S
and Georgopoulos C: Purification and properties of the dnaJ
replication protein of Escherichia coli. J Biol Chem.
260:7591–7598. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Choi J, Djebbar S, Fournier A and Labrie
C: The co-chaperone DNAJC12 binds to Hsc70 and is upregulated by
endoplasmic reticulum stress. Cell Stress Chaperones. 19:439–446.
2014. View Article : Google Scholar :
|
|
7
|
Anikster Y, Haack TB, Vilboux T,
Pode-Shakked B, Thöny B, Shen N, Guarani V, Meissner T, Mayatepek
E, Trefz FK, et al: Biallelic mutations in DNAJC12 cause
hyperphenylalaninemia, dystonia, and intellectual disability. Am J
Hum Genet. 100:257–266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li M, Yang Q, Yi S, Qin Z, Luo J and Fan
X: Two novel mutations in DNAJC12 identified by whole-exome
sequencing in a patient with mild hyperphenylalaninemia. Mol Genet
Genomic Med. 8:e13032020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gallego D, Leal F, Gámez A, Castro M,
Navarrete R, Sanchez- Lijarcio O, Vitoria I, Bueno-Delgado M,
Belanger-Quintana A, Morais A, et al: Pathogenic variants of
DNAJC12 and evaluation of the encoded cochaperone as a genetic
modifier of hyperphenylalaninemia. Human Mutation. 41:1329–1338.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fan Y, Yang Zh, Li F, Hu XC, Yue YW, Yang
J, Liu YT, Liu H, Wang YL, Shi CH and Xu YM: DNAJC12 mutation is
rare in Chinese Han population with Parkinson's disease. Neurobiol
Aging. 68:159.e1–159.e2. 2018. View Article : Google Scholar
|
|
11
|
He HL, Lee YE, Chen HP, Hsing CH, Chang
IW, Shiue YL, Lee SW, Hsu CT, Lin LC, Wu TF and Li CF:
Overexpression of DNAJC12 predicts poor response to neoadjuvant
concurrent chemoradiotherapy in patients with rectal cancer. Exp
Mol Pathol. 98:338–345. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Uno Y, Kanda M, Miwa T, Umeda S, Tanaka H,
Tanaka C, Kobayashi D, Suenaga M, Hattori N, Hayashi M, et al:
Increased expression of DNAJC12 is associated with aggressive
phenotype of gastric cancer. Ann Surg Oncol. 26:836–844. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
De Bessa SA, Salaorni S, Patrão DF, Neto
MM, Brentani MM and Nagai MA: JDP1 (DNAJC12/Hsp40) expression in
breast cancer and its association with estrogen receptor status.
Int J Mol Med. 17:363–367. 2006.PubMed/NCBI
|
|
14
|
Martin-Orozco E, Sanchez-Fernandez A,
Ortiz-Parra I and Ayala-San Nicolas M: WNT Signaling in tumors: The
way to evade drugs and immunity. Front Immunol. 10:28542019.
View Article : Google Scholar
|
|
15
|
Peifer M, McCrea PD, Green KJ, Wieschaus E
and Gumbiner BM: The vertebrate adhesive junction proteins
beta-catenin and plakoglobin and the Drosophila segment polarity
gene armadillo form a multigene family with similar properties. J
Cell Biol. 118:681–691. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Haseeb M, Pirzada RH, Ain QU and Choi S:
Wnt signaling in the regulation of immune cell and cancer
therapeutics. Cells. 8:13802019. View Article : Google Scholar
|
|
17
|
Stewart DJ: Wnt signaling pathway in
non-small cell lung cancer. J Natl Cancer Inst. 106:djt3562014.
View Article : Google Scholar
|
|
18
|
Krishnamurthy N and Kurzrock R: Targeting
the Wnt/beta-catenin pathway in cancer: Update on effectors and
inhibitors. Cancer Treat Rev. 62:50–60. 2018. View Article : Google Scholar
|
|
19
|
Nakayama S, Sng N, Carretero J, Welner R,
Hayashi Y, Yamamoto M, Tan AJ, Yamaguchi N, Yasuda H, Li D, et al:
β-catenin contributes to lung tumor development induced by EGFR
mutations. Cancer Res. 74:5891–5902. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang X, Ma H, Han L, Zheng W, Lu YB, Chen
XF, Liang ST, Wei GH, Zhang ZQ, Chen HZ and Liu DP: SIRT1
deacetylates the cardiac transcription factor Nkx2.5 and inhibits
its transcriptional activity. Sci Rep. 6:365762016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expres- sion data using real-time quantitative PCR
and the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Cui X, Song L, Bai Y, Wang Y, Wang B and
Wang W: Stromal interaction molecule 1 regulates growth, cell
cycle, and apoptosis of human tongue squamous carcinoma cells.
Biosci Rep. 37:BSR201605192017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu R, Zhao B, Ren X, Wu S, Liu M, Wang Z
and Liu W: MiR-27a-3p targeting GSK3β promotes triple-negative
breast cancer proliferation and migration through Wnt/β-catenin
pathway. Cancer Manag Res. 12:6241–6249. 2020. View Article : Google Scholar :
|
|
24
|
Kim JY, Kim HJ, Jung CW, Lee TS, Kim EH
and Park MJ: CXCR4 uses STAT3-mediated slug expression to maintain
radioresistance of non-small cell lung cancer cells: Emerges as a
potential prognostic biomarker for lung cancer. Cell Death Dis.
12:482021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sivaraman A, Kim D, Bhattarai D, Kim M,
Lee HY, Lim S, Kong J, Goo JI, Shim S, Lee S, et al: Synthesis and
structure-activity relationships of arylsulfonamides as AIMP2-DX2
inhibitors for the development of a novel anticancer therapy. J Med
Chem. 63:5139–5158. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liang G, Meng W, Huang X, Zhu W, Yin C,
Wang C, Fassan M, Yu Y, Kudo M, Xiao S, et al: miR-196b-5p-mediated
downregulation of TSPAN12 and GATA6 promotes tumor progression in
non-small cell lung cancer. Proc Natl Acad Sci USA. 117:4347–4357.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Paolillo M and Schinelli S: Extracellular
matrix alterations in metastatic processes. Int J Mol Sci.
20:49472019. View Article : Google Scholar :
|
|
29
|
Wong KK, Jacks T and Dranoff G: NF-kappaB
fans the flames of lung carcinogenesis. Cancer Prev Res (Phila).
3:403–405. 2010. View Article : Google Scholar
|
|
30
|
Popper HH: Progression and metastasis of
lung cancer. Cancer Metastasis Rev. 35:75–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xia Z, Wu S, Wei X, Liao Y, Yi P, Liu Y
and Liu J and Liu J: Hypoxic ER stress suppresses β-catenin
expression and promotes cooperation between the transcription
factors XBP1 and HIF1α for cell survival. J Biol Chem.
294:13811–13821. 2019. View Article : Google Scholar : PubMed/NCBI
|