The stem cell population in dental pulp possesses
multilineage differentiation potential and the pulp vascular system
participates in the reaction of dental pulp tissue to external
stimulus, the initiation of dental pulp inflammation and pulp
tissue repair (1). When sound
dentin suffers damage, such as tooth wear, fracture, or caries,
bacterial infection and the subsequent inflammatory response lead
to damage of pulp tissue and impaired periapical tissue through
blood circulation (2).
Therefore, endodontic therapy has become a necessary option to
preserve teeth by removing microorganisms, their by-products and
residual necrotic tissue (3).
However, the success rate of traditional endodontic therapy was
only 70-80% over the past decade globally (4-6).
The apical seal is important to improve the success rate in
endodontic therapy; an excellent apical seal by root-end filling
prevents the spread of dental pulp inflammation to the periapical
tissue (7). In order to achieve
a higher success rate in endodontic therapy, an ideal root-end
filling material is required that possesses excellent root-end
sealing capacity, good biocompatibility with surrounding
cells/tissue, superior antibacterial properties and ability to
promote tissue regeneration.
Mineral trioxide aggregate (MTA) has been used in
root-end filling as calcium silicate-based bioceramic and displays
better root apical sealing ability and higher success rates
compared with conventional root-end filling materials, such as
amalgam and intermediate restorative material (8-10). Furthermore, given its clinical
effect in root-end filling in endodontic therapy, MTA has also been
used in other endodontic application, such as pulp capping and
regeneration and root perforation repair, and is currently
considered as the gold standard in endodontics. As ProRoot MTA
(Dentsply Sirona) has shown good clinical performance in
endodontics, other calcium silicate-based bioceramics have been
developed, including Bioaggregate (Innovative Bioceramix, Inc.),
Biodentin (Septodont Holding) and iRoot BP/FS/SP (Innovative
Bioceramix, Inc.) (11,12).
The chemical constituents of these novel calcium
silicate-based bioceramics are similar to that of MTA, but
Bioaggregate, Biodentine and iRoot BP/BP Plus display better color
stability than MTA because bismuth oxide is replaced by tantalum or
zirconium oxide as a radiopacifier. Bioaggregate exhibits superior
stable bond strength but inferior mechanical properties and bond
strength in comparison with MTA. Biodentine shows increased
mechanical strength and longer setting time as it does not contain
calcium aluminate and calcium sulfate, which are present in MTA
(13). iRoot BP/BP Plus, novel
calcium silicate-based bioceramics applied in permanent root canal
repair and filling, exhibit easier manipulation and faster setting
time compared with MTA (14).
It is essential to clarify the effect and mechanism
how these bioceramics influence the surrounding cells/tissue when
used in endodontics. Numerous studies have investigated the
biocompatibility and bioactivity of bioceramics in endodontics.
Materials with good biocompatibility should not induce notable and
continuous toxic effects on surrounding cells and tissue (15,16). Biocompatibility can be defined as
the interaction between implanted biomaterials and the associated
tissue (17). Bioactive
materials also induce apatite layer formation and biomineralization
(18). Increased deposition of
hydroxyapatite over time is observed when calcium silicate-based
bioceramics are exposed to PBS, which suggests that these materials
are bioactive (19-22). Bioceramics have been demonstrated
to have excellent biocompatibility and lasting bioactivity during
and after setting by the secretion of molecules. When calcium
silicate-based bioceramics are applied in endodontics, the
interaction between the materials and cells affect the biological
behavior, such as proliferation, differentiation, migration and
apoptosis (23,24). Various types of cell are involved
in changes of biological behavior when bioceramics are used in
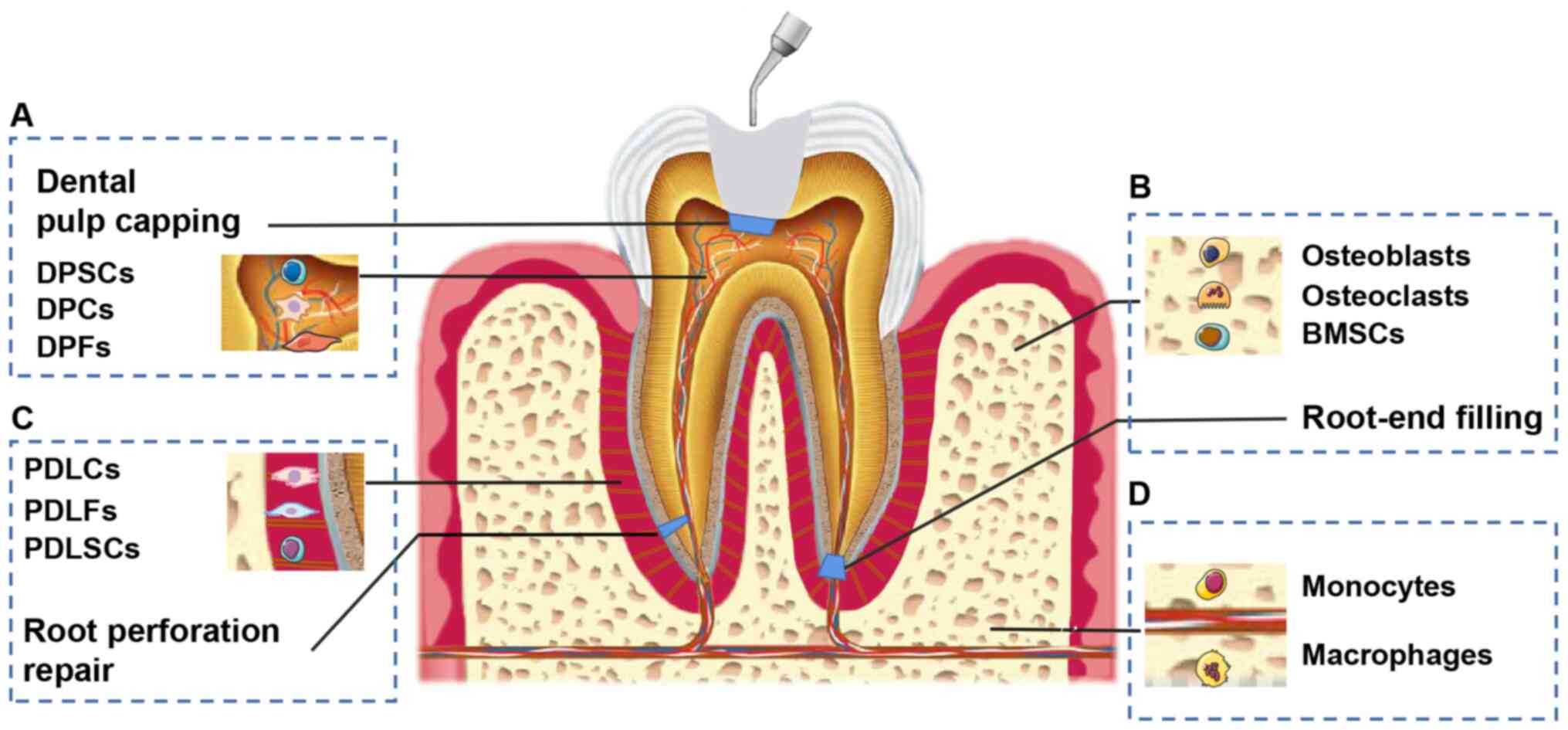
endodontics (Fig. 1). For
example, calcium silicate-based bioceramics affect the biological
behavior of dental pulp stem cells (DPSCs) in dental pulp capping,
whereas osteoblasts/osteoclasts are influenced when bioceramics are
applied as root-end filling material (25,26). Despite the weakness of in
vitro studies in mimicking the human body reaction to
environmental stimuli and providing accurate results compared with
animal or human studies, it is essential to investigate
biocompatibility and bioactivity in vitro to clarify the
mechanism underlying how calcium silicate-based bioceramics
influence cell behavior. The present review focuses on in
vitro biocompatibility and bioactivity when calcium
silicate-based bioceramics are applied in endodontics. All
information is summarized in Table
I.
Mesenchymal stem cells (MSCs) derived from dental
tissue originate from teeth and surrounding support tissue, possess
similar biological characteristics to bone marrow-derived MSCs and
differentiate into osteoblasts, adipocytes, chondrocytes and neural
cells (27,28). MSCs derived from dental tissue
are capable of dentinogenesis or angiogenesis and secretion of
growth factors, which influence behavior, such as proliferation,
differentiation and mineralization (29). MSCs derived from dental tissue
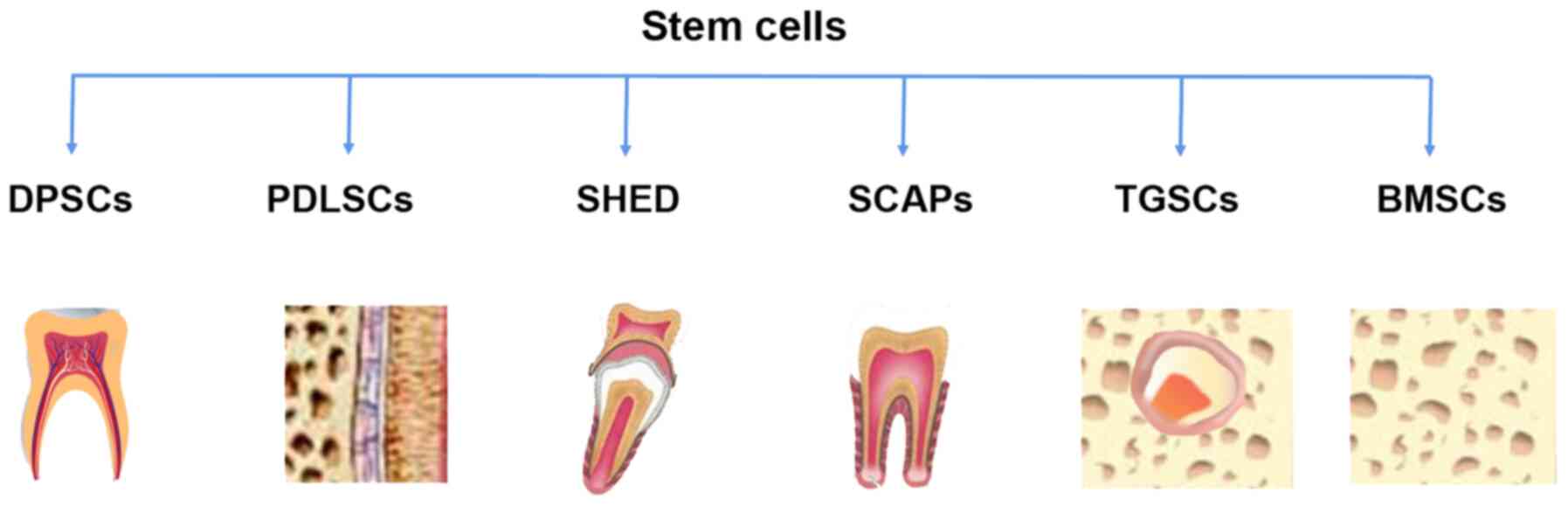
include DPSCs, periodontal ligament stem cells (PDLSCs), stem cells
from human exfoliated deciduous teeth (SHED) and stem cells from
apical papilla (SCAPs) that are involved in renewal and
regeneration of dental tissue via the repair of injured dentin,
root structure and the pulp-dentin complex (26,30,31) (Fig. 2). Calcium silicate-based
bioceramics significantly promote attachment and survival of stem
cells derived from dental tissue but their effects on biological
behavior appear to be cell type-dependent (25). Several common markers are used to
test the osteo/odontogenic and angiogenic potential of stem cells
in the presence of calcium silicate-based bioceramics. For example,
alkaline phosphatase (Alp) is a marker protein of mineralization
and is associated with early osteo/odontogenic differentiation
(32,33). Collagen type I (COL1),
osteocalcin (Ocn) and osteopontin (OPN) are expressed in the
extracellular matrix and serve an important role in osteoblastic
mineralization (34).
Runt-related transcription factor 2 (Runx2) acts as a marker of
osteogenesis in the early stage (35,36), while Ocn functions in the late
stage of osteogenic differentiation (37,38). Regarding dentinogenesis, Runx2
and its downstream molecule osterix (Osx), dentin sialoprotein and
dentin sialophosphoprotein (DSPP) and its downstream molecule
dentin matrix protein 1 (DMP1) (39,40) are considered as protein markers
closely associated with the formation and mineralization of
odontoblasts.
MTA promotes the proliferation and survival of human
DPSCs, bone marrow stromal/stem cells (BMSCs) and PDLSCs via the
ERK signaling pathway (25), and
also exhibit a positive effect on viability of human DPSCs
(41). MTA at high
concentrations (20 and 10 mg/ml) is toxic to human DPSCs but MTA at
low concentrations (2.0, 1.0, 0.2 and 0.1 mg/ml) enhances viability
of human DPSCs but has no effect lower concentrations (0.020 and
0.002 mg/ml) (42). Similarly,
undiluted MTA extract slightly increase survival of human DPSCs,
while 1/2 and 1/4 dilutions of MTA extract have no effect on cell
viability (43). In addition,
MTA at high concentrations (20 mg/ml) decreases proliferation of
DPSCs under inflammatory conditions but has no effect at low
concentrations (0.020 and 0.002 mg/ml) (44). Moreover, various commercial MTA
extracts, such as Angelus MTA (Angelus Dental Products Industry)
and Root MTA (University of Tabriz, Iran), show similar effects on
human DPSCs but were more biocompatible when applied at lower
concentration (1:2) and longer exposure times compared with MTA.
These results suggested that the biocompatibility of MTA is
dependent on not only dosage but also exposure time (45). In terms of time-dependent
biocompatibility of MTA, the cytotoxicity of MTA decreased over
time and the viability and proliferation of human DPSCs increased
following two aging cycles, which further supported the
aforementioned time- and concentration-dependent effects of MTA
(46). Moreover, the
proliferation and viability of DPSCs decreased significantly when
in direct contact with MTA for the first day but subsequently
raised after three days (47).
The initial cytotoxicity of MTA to growth and viability of DPSCs
(47-49) may be partly ascribed to the
relatively rough surface of biomaterials (50) or leakage of components such as
bismuth oxide (51) and Al
(52) and Si ions (53). Higher levels of Si ion
concentrations from the SiO2 phase of materials may lead
to hyperosmoticity and subsequently stimulates production of
inflammatory cytokines (54).
Furthermore, production of MTA during the hydration reaction and
its exposure concentration may result in early slight cytotoxicity
of MTA. Calcium hydroxide is produced when calcium silicate
contained in MTA reacts with water and increases pH of the culture
media (55,56). Basic pH environment and release
of inorganic salts induced by high concentrations of MTA
significantly decreases cell proliferation (57). Furthermore, 24-h set MTA promotes
viability of human DPSCs, whereas 1-h set MTA exhibits an
inhibitory effect after 7 days, which suggests that incompletely
solidified MTA is cytotoxic (58). The subsequent rise in cell
viability may be caused by hydroxyapatite layer formation on the
hydrated bioceramic surface (59) controlled continuous production of
calcium, silica and phosphate ions from bioceramics along with
alkaline pH in the later stage (60,61). Regarding differentiation of human
DPSCs, in the first week following induction with MTA in
odontogenic differentiation medium, DPSCs began to change from
spindle to rounded shape upon reaching confluency and then migrated
to form clusters (62). DPSCs
began to differentiate at day 7 (63) and mineralization was observed by
Alizarin Red staining until day 21. Combination of MTA and
odontogenic differentiation medium enhanced odontogenic
differentiation of DPSCs but MTA extract-alone did not induce this
(62). Treatment with MTA for 1
day affected more genes in uninduced DPSCs than in DPSCs induced by
odontogenic differentiation medium, which suggested that MTA
exhibits a greater stimulative effect on odontogenic
differentiation of uninduced DPSCs compared with induced DPSCs
(64). In addition, MTA at a
concentration of 0.2 mg/ml displayed the strongest capacity to
induce odontoblastic differentiation of human DPSCs via the p42/44
ERK signaling pathway (42).
MTA-conditioned medium at the same concentration enhances the
odonto/osteogenic capacity of DSPCs from inflammatory sites by
activating the NF-κB pathway, as shown by significantly upregulated
odonto/osteoblastic gene expression levels, such as ALP, RUNX2,
OSX, OCN and DSPP (44). Consistent with the changes in
genes associated with osteo/dentinogenic differentiation, MTA
promotes mineralized nodule formation of human DPSCs (65,66). Increased secretion of angiogenic
factor VEGF has been detected in human DPSCs induced by MTA
(63), which in turn affects
viability and function of DPSCs (67). MTA contributes to dentin bridge
formation in endodontics (68,69). In addition, MTA is applied in
pulp capping because of its excellent bioactivity, which has been
confirmed by the elongated shape of DPSCs, formation of collagen
fibers and calcified deposition in the presence of MTA in a model
simulating indirect pulp capping (70). In previous studies, accelerants,
including 5% CaCl2 and 2.5%
Na2HPO4, have been mixed with MTA to shorten
setting time. Compared with MTA in the presence of distilled water,
MTA in the presence of 5% CaCl2 and 2.5%
Na2HPO4 is more biocompatible and exhibits
greater ability to promote odontoblastic differentiation of DPSC
niches (48,71). Propolis, a natural alternative
endodontic material produced by honeybees from tree resin, also
enhances the ability of MTA to promote odontogenic differentiation
and mineralization of DPSCs via the ERK pathway (72).
Compared with human PDLSCs and tooth germ stem cells
(TGSCs), human DPSCs exhibit better viability in the presence of
both Biodentine and MTA (73).
Moreover, Biodentine displays a superior ability to promote
viability, adhesion and migration of human DPSCs compared with MTA.
Human DPSCs spread on the surface of Biodentine show a spindle,
polygonal and flattened morphology (74). Similar to MTA, high
concentrations of Biodentine extract (20 mg/ml) exhibit slight
cytotoxicity, whereas 0.2 mg/ml Biodentine enhances biological
behaviors of human DPSCs, including cell proliferation, viability,
migration, adhesion and mineralization formation. In addition, low
concentrations of Biodentine (0.2 mg/ml) promote odontoblast
differentiation and biomineralization of human DPSCs by activating
ERK1/2 and JNK and attenuating the NF-κB pathway (75,76). Increased Alp activity and dentin
matrix protein expression levels have been observed in human DPSCs
stimulated with Biodentine (75-78). Furthermore, Biodentine
significantly increases calcium deposition (79) and enhances the production of Ocn
and Runx2 in human DPSCs stimulated with lipopolysaccharide (LPS)
although there is no change in ALP expression levels
(53). In terms of the
inflammatory response, Biodentine does not affect high expression
of IL-6 and IL-8 in DPSCs induced by LPS stimulation but decreases
levels of the anti-inflammatory cytokine TGF-β1 (53). Although Biodentine medium is
biocompatible with DPSCs, when in direct contact with DPSCs,
Biodentine exhibits a slight toxic effect and delays closure of
wound edges, which implies that direct contact between Biodentine
and DPSCs leads to cell death or decreased proliferation (80,81). However, unaltered expression
levels of actin, tubulin and vimentin indicate that Biodentine
neither induces apoptosis, inflammation and genotoxicity nor
impairs cellular architecture. The inhibitory effect of Biodentine
on cell proliferation and migration may result from the decreased
space for cell growth in a direct culture model and prolonged cell
doubling time (80). Similarly,
other studies have observed that the viability of human DPSCs in
direct contact with Biodentine is initially decreased (49,77), which may be due to substantial
calcium ion released from Biodentine in the first 3 h (82). Relatively large amounts of Ca and
Si ions and the absence of Sr, Al and S in Biodentine extract
contribute to the biocompatibility of Biodentine (74). Considering the increased release
of Ca and Si ions (83) and
microstructural (84,85) changes induced by acidic
conditions, human DPSCs cultured in Biodentine stored in acidic
environment display more spindle-shaped formation and higher
adherent cell density compared with that in Biodentine stored in
saline (58) Similar to MTA,
Biodentine also increases gene expression levels of osteogenic and
odontogenic markers, such as OPN and DSPP in human
DPSCs when in direct contact with DPSCs (49,77). However, Biodentine promotes
odontogenic differentiation of DPSCs more significantly than
osteogenic differentiation, as indicated by detection of expression
levels of odontoblastic marker DSPP and osteogenic gene
markers ALP, COL1A1 and OPN (80). Moreover, Biodentine promotes
biomineralization and secretion of extracellular mineral matrix in
human DPSCs cultured with osteogenic medium for 21 days (77) and induces more mineralized
nodules in the osteogenic medium compared with MTA, suggesting that
increased calcium ion release, along with a neutral pH, promotes
differentiation and mineralization of DPSCs and subsequently
generates a greater number of structured dentin bridges (82). Similarly, when DPSCs are cultured
with three-dimensional models, Biodentine induces higher viability
compared with MTA. Furthermore, expression levels of COL1A1,
ALP and DSPP in DPSCs on MTA and Biodentine are
initially upregulated significantly and then decrease gradually
until day 21; however, expression of RUNX2 in
three-dimensional cultures remains lower than that in control group
(86). The reason for this may
be that COL1A1, ALP and DSPP are associated with
initiation of dentinogenesis and mineralization. Low expression
levels of RUNX2 contribute to odontoblast differentiation
and cell maturation, whereas increased expression is observed
during terminal odontoblast differentiation (87,88). MTA and Biodentine stimulated
angiogenesis by improving the expression levels of VEGF in
human DPSCs on day 14 (49).
VEGF enhances the proliferation, migration, and
tubulogenesis of endothelial cells close to microvessels, which
regulates both vascularization and angiogenesis. Angiogenesis is a
key factor for wound healing and tissue regeneration of damaged
dental pulp (89-91). The pro-angiogenic capacity of
bioceramics depends on dissolution products, such as Si, Mg and Ca
ions, which induce secretion of angiogenic factors from cells.
Dissolution of calcium ions and subsequent precipitation reactions
on the surface of bioceramics leads to vascular penetration and
osteoblastic differentiation. Interconnections and pore size of the
scaffold also influence the size and amount of the blood vessels,
which are necessary for the vascularization of bioceramic material
(92).
iRoot BP Plus promotes proliferation, attachment,
migration and mineralization of DPSCs compared with MTA (93). Furthermore, iRoot BP Plus
releases more Si ions than MTA, which may explain why iRoot BP Plus
induces greater apatite formation. iRoot BP Plus and MTA promote
stretched and highly organized stress fibre assembly of DPSCs,
which is indicative of reorganization of the actin cytoskeleton.
Moreover, iRoot BP Plus and MTA enhance phosphorylation of both
Paxillin and focal adhesion kinase (FAK) and increase protein
expression levels of Vinculin, FAK and Paxillin in human DPSCs
(94); this is associated with
the formation of focal adhesions (95). Cytoskeleton reorganization and
focal adhesion formation is essential for cell adhesion and
migration (96). These results
confirmed that both iRoot BP Plus and MTA promote attachment and
migration of human DPSCs. Likewise, iRoot FS enhances
proliferation, migration, and osteoblastic differentiation of human
DPSCs (97). Additionally, iRoot
FS displays superior ability than Biodentine to promote
proliferation and migration of human DPSCs on day 7. However, iRoot
FS showed no significant effect on osteogenic differentiation on
day 7 (98), which implied that
iRoot FS affects proliferation and migration of human DPSCs and
later influenced osteoblastic differentiation. A longer
experimental observation period should be used to investigate the
bioactivity of calcium silicate-based bioceramics on human
DPSCs.
MTA exhibits concentration-dependent cellular
compatibility with human PDLSCs; MTA at higher dilution exhibits
better biocompatibility with human PDLSCs (99). Due to the biological
characteristics of MTA, other MTA-based endodontic materials have
been developed, including Endoseal MTA (ES; Maruchi Co., Ltd.),
Nanoceramic Sealer (NCS; B&L Biotech USA, Inc.), Bioroot BC
Sealer (BR; Septodont Holding) and MTA Fillapex (Angelus Dental
Products Industry). Although human PDLSCs in the presence of these
MTA-based bioceramics maintain high expression levels of MSC
markers, including CD105, CD73 and CD90, the capacity of human
PDLSCs to migrate, adhere and grow is higher when treated with BR
at different concentrations compared with ES and NCS. ES and MTA
Fillapex show cytotoxicity to human PDLSCs at 24, 48 and 72 h and
give rise to worse cell attachment and spread, which may be caused
by tungsten contained in MTA Fillapex (100-102). By contrast, 2 mg/ml MTA extract
is the optimal concentration to markedly increase calcified nodule
formation, Alp activity and odonto/osteogenic differentiation in
human PDLSCs; these effects are mediated by activating NF-κB and
MAPK signaling pathways (103).
Likewise, 2 mg/ml Biodentine is the most biocompatible
concentration to promote migration, attachment, and mineralization
of human PDLSCs. Biodentine at low concentrations (2.00, 0.20 and
0.02 mg/ml) significantly enhances viability of human PDLSCs, while
Biodentine at higher concentrations (20 mg/ml) exhibits
cytotoxicity, which may be associated with high pH (104). High pH of Biodentine results in
an increased concentration of iron and calcium ions in the
extracellular environment (105). The inhibitory effect of
Biodentine at high concentration (20 mg/ml) on the viability of
human PDLSCs may be explained by increased or unbalanced ions
levels, which generate a toxic effect on cells (106).
SHEDs originate from deciduous teeth and regenerate
bone and dentin, but not dentin/pulp-like complexes as human DPSCs
do (124). Due to the good
porous microstructures in MTA or Biodentine, SHEDs attach and
spread well on the surface of MTA and Biodentine, which helps SHEDs
maintain mesenchymal properties in the presence of MTA or
Biodentine with positive expression of CD105, CD90 and CD73
(125). Moreover, the capacity
of SHEDs to adhere and proliferate is enhanced by MTA or Biodentine
after 48 h (125). MTA and
Biodentine exhibit a comparable ability to promote migration of
SHEDs. In addition, the viability and proliferation of SHEDs
cultured with 1 mg/ml MTA- or Biodentine-conditioned medium is
similar to that of negative control during the whole incubation
period, which implies that both MTA and Biodentine at 1 mg/ml are
non-toxic to SHEDs (126).
However, MTA shows greater potential to promote odontogenic
differentiation compared with Biodentine (126), whereas Biodentine has better
capability to promote proliferation and calcified matrix deposition
in SHEDs than MTA (125,127).
Furthermore, Biodentine affects behavior of SHEDs in a
concentration-dependent manner. Biodentine at low concentrations
(2.00, 0.20 and 0.02 mg/ml) stimulates proliferation, viability and
migration of SHEDs, whereas high concentrations of Biodentine (20
mg/ml) exhibit slight cytotoxicity to SHEDs. Changes in the
concentration of Biodentine have no impact on the adhesion ability
of SHEDs (128). Similarly,
Biodentine at higher dilutions (1:16 and 1:32) is more effective in
promoting proliferation, odontogenic differentiation and
biomineralization of SHEDs, which may be because Sr and Si are
gradually released from Biodentine as the concentration of
Biodentine decreases over time (129). By contrast, fresh mixed MTA
impairs the viability and migration of SHEDs and enhance apoptosis
over 7 days. Furthermore, the cytotoxicity of MTA to SHEDs is more
apparent when SHEDs directly contact MTA (130). The different results may be
associated with the preparation of MTA. For example, freshly mixed
MTA is frequently used in endodontics whereas bioceramic eluate or
aged bioceramics were used in the aforementioned in vitro
studies (86). However, the
cytotoxicity of these bioceramics generally decreased as the
bioceramic set and pH changed of medium over time. Freshly mixed
MTA caused severe damage to cells due to the initially high
concentration of calcium hydroxide and subsequent raise in pH to
12.5 after mixing for 3 h (131,132). Although iRoot BP Plus possesses
a similar capacity to MTA in terms of SHEDs proliferation, it
displays more prominent capacity to enhance adhesion, migration and
osteogenesis of SHEDs compared with MTA (93).
Both MTA and Biodentine have been shown to promote
the proliferation, odontoblastic differentiation and
biomineralization of SCAPs over 14 days (133,134). However, Schneider et al
(135) found MTA induces early
short-term proliferation of SCAPs over 5 days and promotes the
migration of SCAPs after 6 h. Saberi et al (136) discovered that the cytotoxicity
of both complete set MTA and Biodentine to SCAPs decreased over
time. By contrast, Miller et al (137) revealed that incompletely set
MTA inhibits proliferation of SCAPs, whereas Biodentine promotes
proliferation of SCAPs. The difference in results may be due to the
method of cytotoxicity assessment, contact between cells and
material, concentration of material and assessment time points. MTA
affects survival of SCAPs in concentration-dependent manner. MTA at
lower concentrations (0.02, 0.20 and 2.00 mg/ml) exhibits excellent
biocompatibility with SCAPs; however, proliferation of SCAPs is
inhibited and normal morphological cells disappeared when treated
with MTA at higher concentrations (10 and 20 mg/ml) (138). Low concentrations of MTA or
Biodentine (2.00, 0.20 and 0.02 mg/ml) enhance Alp activity and
osteoblastic/odontoblastic differentiation in SCAPs, while high
concentrations of MTA or Biodentine (20 mg/ml) exhibit a negative
effect (138,139). MTA extract enhances the ability
of osteogenic medium to induce mineralization and increase
expression of mineralization-associated genes, such as Ocn
(140). In comparison with MTA,
SCAPs treated with Biodentine display greater odontoblastic
differentiation, as demonstrated by positive alizarin red staining
and expression of genes encoding DMP-1, DSPP, OCN and matrix
extracellular phosphoglycoprotein (133,134). Biodentine enhances expression
of odontoblast specific marker DSPP, while MTA promotes
osteoblastic differentiation of SCAPs by increasing expression of
the osteoblastic marker integrin-binding sialoprotein (137). Both MTA and Biodentine enhance
the secretion of pro-inflammatory cytokines, such as IL-1α, IL-1β
and IL-6 (139), and MTA
activates NF-κB signaling pathway, which affects the
odonto/osteogenic differentiation of SCAPs (141,142). In addition, p38 and ERK
signaling pathways serve an essential role in
odontoblastic/osteoblastic differentiation of SCAPs stimulated by
MTA (138). MTA and Biodentine
enhance the angiogenic potential of SCAPs; these bioceramics
promote the expression of angiogenic genes in human SCAPs, such as
VEGFA and c-fos induced growth factor (FIGF)
(134), which induce
endothelial cell proliferation, migration and differentiation, and
promote formation of endothelial tubules (143-146).
Compared with MTA, iRoot FS exhibits similar
biocompatibility with human SCAPs but possesses markedly stronger
capacity to enhance migration and osteo/odontogenesis
differentiation of human SCAPs, and mineralized nodule formation
via the Wnt/β-catenin signaling pathway (147). iRoot FM at low concentrations
(0.5 mg/ml) increases proliferation and osteo/odontoblastic
differentiation of SCAPs, whereas there is no marked effect on
SCAPs stimulated with iRoot FM at high concentrations (1.0 and 2.5
mg/ml). Moreover, mineralized nodule formation and expression of
DMP-1 and ALP are enhanced by iRoot FM compared with
Ca(OH)2. However, iRoot FM at different concentrations
has no impact on morphology of SCAPs (148).
TGSCs, a popularized stem cell source derived from
wisdom teeth, display MSC properties and can differentiate into
endothelial or epithelial cells in dental tissue regeneration
(149,150). Consistent with human DPSCs and
PLSCs, MTA and Biodentine exhibit no cytotoxicity to TGSCs
(73). Nonetheless, viability
and odontogenic differentiation of TGSCs are inhibited slightly
when in direct contact with MTA, which has been confirmed by
decreased numbers of attached cells and Alp activity (151). MTA and Biodentine induce
angiogenesis of TGSCs by promoting the release of angiogenic growth
factors (platelet-derived growth factor, fibroblast growth factor-2
and VEGF) and enhancing tube formation of human umbilical vein
endothelial cells (73).
Furthermore, iRoot SP exhibits good biocompatibility with human
TGSCs and promote their attachment (152). iRoot SP possesses an inferior
capacity to MTA in terms of inducing odontogenic differentiation of
human TGSCs and hard tissue deposition; human TGSCs in the presence
of MTA exhibit higher Alp activity and enhanced odontoblastic
differentiation compared with those in the presence of iRoot SP
(153).
Restoration of bone tissue around teeth with
lesions relies on the amount of, and balance between, osteoblasts
and osteoclasts (154). When
calcium silicate-based bioceramics are used in perforation repair,
apical plugs in necrotic teeth or root-end filling in endodontics,
the interaction between the bioceramics and osteoblasts in
periapical tissue is key to inflammation control and wound repair
of (155). The biological
influence of calcium silicate-based bioceramics on
osteoblasts/osteoclasts must be characterized.
Compared with MTA, Biodentine displays good
cytocompatibility with primary human osteoblasts, indicated by
enhancement of cell viability, attachment and proliferation
(164,181). Similar biocompatibility of MTA
and Biodentine has been observed with human osteoblast-like cell
line MG63; both enhance viability, adhesion and proliferation of
MG63 cells, which may be because Biodentine and MTA have similar
surface roughness, heterogeneous morphology and particle size
(182). Biodentine and MTA both
exhibit positive effects on viability and calcification of MC3T3-E1
cells (183). Biodentine and
MTA show dose-dependent effects on viability of Saos-2 cells.
Biodentine or MTA at lower concentrations (1:4 and 1:8) result in
higher viability of Saos-2 cells. Furthermore, Biodentine
stimulates proliferation and migration of Saos-2 cells and induces
expression of ALP and mineralization (184).
Compared with MTA, iRoot BP Plus induces greater
cytotoxicity to primary human osteoblasts but is still considered
as biocompatible because cell viability in the presence of iRoot BP
Plus remained >70% compared with that in the control group
(185). When MTA or iRoot BP
Plus is applied in acidic pH conditions, the secretion of Ca and Si
ions is enhanced and apatite formation is decreased. However, cell
attachment of MC3T3-E1 on these bioceramics is not affected
significantly in an acidic environment. Furthermore, MTA decreases
viability, whereas iRoot BP Plus increases survival of MC3T3-E1
cells, which suggests that, compared with MTA, iRoot BP Plus may be
more suitable as root-end filling material under inflammatory
acidic conditions when used in endodontics (186). By contrast, iRoot FS exhibits
better biocompatibility with human osteoblast-like MC3T3-E1 and
MG63 cells by promoting their attachment and proliferation
(187,186). iRoot FS shows better
biocompatibility than MTA or iRoot BP Plus because certain toxic
metal substances, such as bismuth (189), aluminium and manganese, are
excluded in iRoot FS to enhance its compatibility. On the other
hand, smaller particle size on the surface of iRoot FS results in
higher cell attachment and subsequent proliferation (188). iRoot SP is non-cytotoxic to
MG63 cells and enhances osteoblastic differentiation, which is
beneficial to healing inflammatory periapical tissue (190).
The migration and fusion of osteoclast precursors
is key to osteoclast formation. MTA and Bioaggregate inhibit bone
resorption and osteoclast differentiation via preventing the
migration and fusion of osteoclast precursors, including mouse bone
marrow macrophages (191) and
RAW264.7 macrophages (192-194). In addition, MTA inhibits
osteoclastogenesis in a dose-dependent manner in the co-culture of
mouse bone marrow cells with primary osteoblast cells. Furthermore,
MTA suppresses expression of osteoprotegerin in primary osteoblast
cells without affecting receptor activator of NF-κB ligand (RANKL)
expression levels (195,196).
Moreover, MTA solution at low concentration (20%) impairs
phosphorylation of c-Src, decreases expression levels of genes
encoding MMP-9 and cathepsin K and disrupt formation of actin
rings. MTA solution at high concentration (50%) upregulate
expression levels of Bim to increase apoptosis of osteoclasts
(196). The mechanism
underlying MTA-induced inhibition of osteoclastogenesis is
associated with attenuation of the autophagic pathway, as
demonstrated by decrease in autophagic vacuole and expression
levels of autophagic genes and proteins (194). Bioaggregate inhibits
osteoclastogenesis via the NF-κB/RANK signaling pathway by
decreasing expression levels of Rank, TNF receptor-associated
factor 6, NF-κB and nuclear factor of activated T cells 1 (192,193). Bioaggregate and MTA possess
comparable ability to decrease osteoclast numbers and attenuate
bone resorption (191).
Bioaggregate or MTA inhibit osteoclast differentiation and bone
resorption due to activation of autophagy in osteoclast
differentiation; MTA inhibits osteoclast differentiation via
inhibition of the autophagic pathway (194). Compared with MTA, Biodentine
exhibits a lower inhibitory effect on osteoclast differentiation
and activity of murine bone marrow macrophages by inhibiting ERK1/2
and NF-κB signaling pathways (197). Moreover, the inhibitory effect
on osteoclast differentiation and activity of both MTA and
Biodentine is similar to that of alendronate, which has been
reported to prevent root resorption by inhibiting macrophages
(197), which suggests the
application of calcium silicate-based bioceramics as treatment to
prevent root resorption in endodontics (198,199). iRoot SP shows more cytotoxicity
to RAW264.7 cells than MTA but possesses a similar ability to
inhibit osteoclastogenesis (200). The aforementioned studies
suggested that calcium silicate-based bioceramics attenuate
osteoclast differentiation and the primary mechanism is associated
with their bioactive elements. Bioactive elements contained in
these bioceramics, such as Ca, Mg, Si and Sr, enhance osteoblastic
differentiation and suppress RANKL-induced osteoclastogenesis
(201-204). A significant increase in Si and
Sr ions has been observed in extracts of calcium silicate-based
bioceramics (192,205). Si ions enhance the viability,
adhesion, differentiation, mineralization and angiogenesis of
osteoblasts via the Wnt/β-catenin and MAPK signaling pathways
(206-210). Meanwhile, the effect of Si ion
on surface roughness is characterized by increased adhesion and
proliferation of human osteoblast cell lines (211). Furthermore, Si and Sr ions
suppress RANKL-mediated osteoclastic differentiation and bone
resorption by inhibiting expression levels of cathepsin K,
tartrate-resistant acid phosphatase and c-Fos (205,212,213). In addition, Si and Sr ions
create alkaline conditions, which neutralize lactic acid from
osteoclasts and promotes accumulation of mineralized components of
teeth (214,215). Therefore, the bioactive
elements exhibit synergistic effects on osteogenesis,
osteoclastogenesis and angiogenesis of associated cells in
endodontics (216).
Dental pulp or PDLC/Fs are associated with wound
healing and tissue regeneration of dental or periapical tissue,
respectively (217). When
calcium silicate-based bioceramics are used in pulp capping,
cells/fibroblasts from dental pulp are involved in interactions
between cells and bioceramics. Cells/fibroblasts from periodontal
ligament are affected by bioceramics applied in perforation repair
or root-end filling. Numerous studies have investigated the effect
on biocompatibility and bioactivity of cells/fibroblasts from
dental pulp or periodontal ligament.
Compared with SuperEBA and Vitrebond, MTA exhibits
decreased suppression of mitochondrial activity in the rat DPC
RPC-C2A cell line (218).
Furthermore, MTA significantly promotes proliferation, odontogenic
differentiation and mineralization of human DPCs but inhibits
secretion of lL-1β and IL-6 (219,220). In order to improve the
bioactivity potential of MTA, the growth factor FGF-2 has been
added to MTA to enhance its effect on proliferation and osteogenic
differentiation of human DPCs (221). Set and fresh MTA display
similar biocompatibility with human DPCs. In addition, MTA
increases expression of the angiogenic factors VEGF and angiogenin
(222). In comparison with MTA,
Bioaggregate and Biodentine possess equal biocompatibility with
human DPCs. Moreover, Bioaggregate, Biodentine and MTA enhance mRNA
expression levels of ostogenic/odontogenic genes, such as ALP, OPN,
OCN, DSPP and DMP-1, increase Alp activity and promote
mineralization nodule formation due to activation of the MAPK
signaling pathway induced by these calcium silicate-based
bioceramics (223,224). In addition, Bioaggregate
exhibits superior capacity to MTA in terms of adhesion, attachment
and migration of human DPCs (225). Moreover, compared with MTA,
Bioaggregate induces enhanced mineralization and odontoblastic
differentiation in human DPCs (226). In terms of osteogenic
differentiation of human DPCs, Bioaggregate displays stronger
potential than MTA (223). Both
Biodentine and MTA promote mineralization by increasing secretion
of TGF-β1 from human DPCs, which mediates mineralization-associated
cellular activity and subsequent dentin bridge formation (227,228). Biodentine enhances cell
proliferation, viability, migration, adhesion, odontoblastic
differentiation and biomineralization of the immortalized murine
DPC OD-21 cell line (229).
iRoot BP Plus exhibits a higher proliferation rate of human DPCs
compared with MTA during the whole culture period (230). Moreover, iRoot BP Plus promotes
migration and upregulates the expression of focal adhesion
molecules in human DPCs via the ERK 1/2, JNK and Akt signaling
pathways (231). In addition,
iRoot BP Plus possesses stronger potential than MTA to enhance the
mineralization and odontoblastic differentiation of human DPCs
(226).
Human PDLFs exhibit decreased proliferation rate
when cultured on the surface of MTA compared with culturing on the
surface of a coverslip (250);
PDLFs on the surface of MTA have a rounded morphology with blunted
extensions, while PDLFs on the surface of glass coverslips show
good attachment and spreading (251). Balto investigated the effect of
MTA surface characteristics on attachment of human PDLFs; human
PDLFs did not attach to fresh MTA and the surface of cells appeared
less smooth and exhibited more vacuoles. By contrast, human PDLFs
on the surface of set MTA were round and flattened with smooth
surfaces and attached well to MTA (252). Similarly, Bonson et al
reported that human PDLFs exposed to washed MTA possess greater
proliferation capacity those exposed to fresh MTA. Moreover,
compared with fresh MTA, washed MTA exhibits stronger potential to
induce osteogenic differentiation of human PDLFs (253). Compared with other endodontic
materials, such as Diaket (ESPE; 3M), Super-EBA (Harry J Bosworth
Company) and amalgam, MTA also displays better biocompatibility
with human PDLFs and does not induce apoptosis and necrosis of
human PDLFs (254-257). Compared with these other
materials in root perforation models in vitro, MTA also
results in higher viability in human PDLFs and induces mRNA
expression levels of COL1 and RUNX2 in human PDLFs,
which suggests that MTA has potential to induce osteogenic
differentiation of PDLFs, which is key for periodontal regeneration
(105). Likewise,
MTA-conditioned medium at low concentrations (0.5, 5.0 and 50.0
µg/ml) possesses superior capacity to formocresol and ferric
sulphate in maintaining the viability of human PDLFs, whereas MTA
at higher concentrations (5,000 µg/ml) shows slight
cytotoxicity to human PDLFs (258,259). Bioaggregate displays
biocompatibility with human PDLFs comparable to that of MTA,
whereas viability of human PDLFs in the presence of Biodentine is
slightly decreased compared with MTA (260-262). By contrast, Akbulut et
al (263) reported that
Biodentine possesses better biocompatibility with human PDLFs. This
discrepancy may be associated with chemical composition of the
material, assessment time point and surface characteristics. In
terms of chemical composition, calcium chloride used in the liquid
of Biodentine as an accelerator decreases the setting time
(264) and results in early
production of calcium hydroxide, which contributes to relatively
decreased cell survival after 24 h in the presence of Biodentine
(265). Moreover, zirconium
oxide in Biodentine is non-toxic to murine PDLFs, but bismuth oxide
in MTA has no impact on cell growth (266). MTA supports higher cell
viability during the first 24 h but decreases cell viability to 80%
later (251,260). Biodentine maintains lower cell
viability during the first 24 h but viability increases gradually
over time (261). Human PDLFs
attach well to the surfaces of both MTA and Biodentine and maintain
their original morphology. Nevertheless, more cell aggregates have
been observed on the surface of Biodentine, whereas human PDLFs
tend to show greater spread and elongation on MTA (267). Moreover, expression levels of
Integrin β1 and Vinculin, which are associated with focal contacts
between human PDLFs and bioceramics, are higher in human PDLFs
treated with Biodentine than in those treated with MTA, which
suggests that the surface characteristics of Biodentine promote the
adhesion and survival of human PDLFs more strongly compared with
MTA (268).
Immune cells, such as monocytes and macrophages,
respond immediately when biomaterials are placed into tissue, which
causes the initial inflammatory response and tissue healing.
Macrophages release pro-inflammatory cytokines in the beginning of
an acute inflammatory response, such as TNF-α, IL-1 and IL-12, but
release anti-inflammatory cytokines during regeneration and healing
of tissue, such as IL-4, which contributes to the production of
fibronectin (269-271). MTA displays detectable, but not
statistically significant, cytotoxicity to human monocytic cell
line THP1 and alters secretion of inflammatory cytokines (272). In addition, macrophages and
mast cells participate in leukocyte recruitment and extravasation
via secretion of inflammatory cytokines that regulate inflammation
control and tissue healing in pulpitis and apical periodontitis
(273,274). Moreover, neutrophil chemotactic
factor is induced from macrophages and mast cells by MTA; the
upregulation of these neutrophil chemotactic factor substances
participates in migration and accumulation of neutrophils,
monocytes and lymphocytes (275-277). Similarly, Cavalcanti et
al (278) found that MTA
increases secretion of IL-8 and IL-1β, which supports the migration
of human neutrophils. Chang et al (279) discovered that MTA enhances
migration of immune cells, which is regulated by calcium-sensing
receptors and the PI3K pathway for chemotaxis, as well as the
Ca2+-calmodulin-dependent MLCK pathway for
chemokinesis.
M1/M2 macrophage polarization is associated with
the inflammatory response and subsequent tissue regeneration
following biomaterial implantation (270,280,281). Tu et al found that iRoot
SP induces greater cytotoxicity to RAW264.7 macrophages than MTA.
Furthermore, both MTA and iRoot SP induce expression of
pro-inflammatory cytokines without inducing osteoclastogenesis in
RAW264.7 macrophages. In addition, MTA primarily induces M2
macrophage polarization, whereas iRoot SP induces M1 macrophage
polarization (200). Both MTA
and iRoot SP are non-toxic to RAW264.7 cells (282). Moreover, MTA does not affect
the viability and adherence of M1 and M2 macrophages isolated from
mice (283,284). MTA and iRoot SP reinforce
expression of inflammatory cytokines in RAW264.7 cells.
Furthermore, MTA and iRoot SP possess equal capacity to stimulate
M1/M2 macrophage polarization but greater M2 macrophage
polarization is induced, which implies that calcium silicate-based
bioceramics shift M1/M2 polarization balance to M2 macrophage
polarization under inflammatory conditions (282,285). Yeh et al reported that
MTA induces THP-1 cells toward M2 polarization by activating the
Axl/Akt/NF-κB signaling pathway. MTA promotes tissue regeneration
and wound healing via M2 macrophage polarization (286). MTA does not induce DNA breakage
of human peripheral lymphocytes, which is the first step in
carcinogenesis. This suggests that MTA exhibits no potential
carcinogenic risk when used in endodontics (287).
Compared with other pulp capping materials, such as
TheraCal and Xeno III, Biodentine decreases migration and adhesion
of THP-1 cells to endothelial cells and inhibits their activation
to macrophages in vitro (240). In addition, Biodentine does not
stimulate expression of inflammation-associated enzymes in
vitro, such as prostaglandin E2 and thromboxane (288). Compared with MTA, Biodentine
displays a more notable inhibitory effect on mRNA and protein
expression levels of inflammatory cytokines in RAW264.7 macrophages
(183). Biodentine is
biocompatible with immune cells, which is consistent with another
study that demonstrated that Biodentine exhibits similar
biocompatibility with human monocytes compared with MTA (289).
Not applicable.
WS conceived and wrote the manuscript. WS and SL
contributed to data acquisition, analysis and interpretation. QT
made substantial contributions to conception and design. LC and ZY
critically revised the manuscript. Data sharing is not applicable.
All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare they have no competing
interests.
Not applicable.
The present review was supported by grants from the National Key
R&D Program of China (grant no. 2017YFC1104301) and National
Natural Science Foundation of China (grant no. 81700955).
|
1
|
Liu H, Gronthos S and Shi S: Dental pulp
stem cells. Methods Enzymol. 419:99–113. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heyeraas KJ and Kvinnsland I: Tissue
pressure and blood flow in pulpal inflammation. Proc Finn Dent Soc.
88(Suppl 1): S393–S401. 1992.
|
|
3
|
Mohammadi Z and Dummer PMH: Properties and
applications of calcium hydroxide in endodontics and dental
traumatology. Int Endod J. 44:697–730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siew K, Lee AH and Cheung GS: Treatment
outcome of repaired root perforation: A systematic review and
meta-analysis. J Endod. 41:1795–1804. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Juneja P and Kulkarni S: Clinical and
radiographic comparison of biodentine, mineral trioxide aggregate
and formocresol as pulpotomy agents in primary molars. Eur Arch
Paediatr Dent. 18:271–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suhag K, Duhan J, Tewari S and Sangwan P:
Success of direct pulp capping using mineral trioxide aggregate and
calcium hydroxide in mature permanent molars with pulps exposed
during carious tissue removal: 1-year follow-up. J Endod.
45:840–847. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gutmann JL and Pitt Ford TR: Management of
the resected root end: A clinical review. Int Endod J. 26:273–283.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Abusrewil SM, McLean W and Scott JA: The
use of Bioceramics as root-end filling materials in periradicular
surgery: A literature review. Saudi Dent J. 30:273–282. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Torabinejad M, Watson TF and Pitt Ford TR:
Sealing ability of a mineral trioxide aggregate when used as a root
end filling material. J Endod. 19:591–595. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Asgary S, Eghbal MJ and Parirokh M:
Sealing ability of a novel endodontic cement as a root-end filling
material. J Biomed Mater Res A. 87:706–709. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parirokh M, Torabinejad M and Dummer PMH:
Mineral trioxide aggregate and other bioactive endodontic cements:
An updated overview-part I: Vital pulp therapy. Int Endod J.
51:177–205. 2018. View Article : Google Scholar
|
|
12
|
Torabinejad M, Parirokh M and Dummer P:
Mineral trioxide aggregate and other bioactive endodontic cements:
An updated overview part II: Other clinical applications and
complications. Int Endod J. 51:284–317. 2018. View Article : Google Scholar
|
|
13
|
Majeed A and AlShwaimi E: Push-out bond
strength and surface microhardness of calcium silicate-based
biomaterials: An in vitro study. Med Princ Pract. 26:139–145. 2017.
View Article : Google Scholar :
|
|
14
|
Song W, Sun W, Chen L and Yuan Z: In vivo
biocompatibility and bioactivity of calcium silicate-based
bioceramics in endodontics. Front Bioeng Biotechnol. 8:5809542020.
View Article : Google Scholar :
|
|
15
|
Rodriguez-Lozano FJ, Lopez-Garcia S,
Garcia-Bernal D, Sanz JL, Lozano A, Pecci-Lloret MP, Melo M,
Lopez-Gines C and Forner L: Cytocompatibility and bioactive
properties of the new dual-curing resin-modified calcium
silicate-based material for vital pulp therapy. Clin Oral Investig.
Feb 27–2021.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garrido M, Morales D, Saldias MP,
Fernandez C, Villalobos V, Cerda O and Caceres M: Cellular response
of human apical papilla cells to calcium hydroxide and tricalcium
silicate-based cements. BMC Oral Health. 21:1062021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lemons JE: Ceramics: Past, present, and
future. Bone. 19(Suppl 1): 121S–128S. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hench LL and Wilson J: Surface-active
biomaterials. Science. 226:630–636. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Grech L, Mallia B and Camilleri J:
Investigation of the physical properties of tricalcium silicate
cement-based root-end filling materials. Dent Mater. 29:e20–e28.
2013. View Article : Google Scholar
|
|
20
|
Shokouhinejad N, Nekoofar MH, Razmi H,
Sajadi S, Davies TE, Saghiri MA, Gorjestani H and Dummer PM:
Bioactivity of EndoSequence root repair material and bioaggregate.
Int Endod J. 45:1127–1134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han L, Okiji T and Okawa S: Morphological
and chemical analysis of different precipitates on mineral trioxide
aggregate immersed in different fluids. Dent Mater J. 29:512–517.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Camilleri J: Characterization and
hydration kinetics of tricalcium silicate cement for use as a
dental biomaterial. Dent Mater. 27:836–844. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao W, Chang J and Zhai W: Self-setting
properties and in vitro bioactivity of
Ca3SiO5/CaSO4.1/2H2O
composite cement. J Biomed Mater Res A. 85:336–344. 2008.
View Article : Google Scholar
|
|
24
|
Zhao W, Wang J, Zhai W, Wang Z and Chang
J: The self-setting properties and in vitro bioactivity of
tricalcium silicate. Biomaterials. 26:6113–6121. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen I, Salhab I, Setzer FC, Kim S and Nah
HD: A new calcium silicate-based bioceramic material promotes human
osteo- and odontogenic stem cell proliferation and survival via the
extracellular signal-regulated kinase signaling pathway. J Endod.
42:480–486. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Morsczeck C and Reichert TE: Dental stem
cells in tooth regeneration and repair in the future. Expert Opin
Biol Ther. 18:187–196. 2018. View Article : Google Scholar
|
|
27
|
Aydin S and Sahin F: Stem cells derived
from dental tissues. Adv Exp Med Biol. 1144:123–132. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rodriguez-Lozano FJ, Bueno C, Insausti CL,
Meseguer L, Ramirez MC, Blanquer M, Marin N, Martinez S and
Moraleda JM: Mesenchymal stem cells derived from dental tissues.
Int Endod J. 44:800–806. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rombouts C, Giraud T, Jeanneau C and About
I: Pulp Vascularization during tooth development, regeneration, and
therapy. J Dent Res. 96:137–144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Murray PE, Garcia-Godoy F and Hargreaves
KM: Regenerative endodontics: A review of current status and a call
for action. J Endodont. 33:377–390. 2007. View Article : Google Scholar
|
|
31
|
Orti V, Collart-Dutilleul PY, Piglionico
S, Pall O, Cuisinier F and Panayotov I: Pulp regeneration concepts
for nonvital teeth: From tissue engineering to clinical approaches.
Tissue Eng Part B Rev. 24:419–442. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sugawara Y, Suzuki K, Koshikawa M, Ando M
and Iida J: Necessity of enzymatic activity of alkaline phosphatase
for mineralization of osteoblastic cells. Jpn J Pharmacol.
88:262–269. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cormier C: Markers of bone metabolism.
Curr Opin Rheumatol. 7:243–248. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rodan GA and Noda M: Gene expression in
osteoblastic cells. Crit Rev Eukaryot Gene Expr. 1:85–98.
1991.PubMed/NCBI
|
|
35
|
Camilleri S and McDonald F: Runx2 and
dental development. Eur J Oral Sci. 114:361–373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rathinam E, Rajasekharan S, Chitturi RT,
Martens L and De Coster P: Gene expression profiling and molecular
signaling of dental pulp cells in response to tricalcium silicate
cements: A systematic review. J Endod. 41:1805–1817. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bai Y, Bai Y, Matsuzaka K, Hashimoto S,
Kokubu E, Wang X and Inoue T: Formation of bone-like tissue by
dental follicle cells co-cultured with dental papilla cells. Cell
Tissue Res. 342:221–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gibson MP, Zhu Q, Wang S, Liu Q, Liu Y,
Wang X, Yuan B, Ruest LB, Feng JQ, D'Souza RN, et al: The rescue of
dentin matrix protein 1 (DMP1)-deficient tooth defects by the
transgenic expression of dentin sialophosphoprotein (DSPP)
indicates that DSPP is a downstream effector molecule of DMP1 in
dentinogenesis. J Biol Chem. 288:7204–7214. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee SK, Lee KE, Jeon D, Lee G, Lee H, Shin
CU, Jung YJ, Lee SH, Hahn SH and Kim JW: A novel mutation in the
DSPP gene associated with dentinogenesis imperfecta type II. J Dent
Res. 88:51–55. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chang CC, Yeh CL, Chang HH, Kuo YF, Huang
PY and Lin CP: Effect of different zinc concentrations on
partially-stabilized cement for vital pulp therapy. J Formos Med
Assoc. 118:1610–1615. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao X, He W, Song Z, Tong Z, Li S and Ni
L: Mineral trioxide aggregate promotes odontoblastic
differentiation via mitogen-activated protein kinase pathway in
human dental pulp stem cells. Mol Biol Rep. 39:215–220. 2012.
View Article : Google Scholar
|
|
43
|
Tomas-Catala CJ, Collado-Gonzalez M,
Garcia-Bernal D, Onate-Sanchez RE, Forner L, Llena C, Lozano A,
Castelo-Baz P, Moraleda JM and Rodriguez-Lozano FJ: Comparative
analysis of the biological effects of the endodontic bioactive
cements MTA-Angelus, MTA Repair HP and NeoMTA Plus on human dental
pulp stem cells. Int Endod J. 50(Suppl 2): e63–e72. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang Y, Yan M, Fan Z, Ma L, Yu Y and Yu J:
Mineral trioxide aggregate enhances the odonto/osteogenic capacity
of stem cells from inflammatory dental pulps via NF-κB pathway.
Oral Dis. 20:650–658. 2014. View Article : Google Scholar
|
|
45
|
Jaberiansari Z, Naderi S and Tabatabaei
FS: Cytotoxic effects of various mineral trioxide aggregate
formulations, calcium-enriched mixture and a new cement on human
pulp stem cells. Iran Endod J. 9:271–276. 2014.PubMed/NCBI
|
|
46
|
Niu LN, Watson D, Thames K, Primus CM,
Bergeron BE, Jiao K, Bortoluzzi EA, Cutler CW, Chen JH, Pashley DH
and Tay FR: Effects of a discoloration-resistant calcium
aluminosilicate cement on the viability and proliferation of
undifferentiated human dental pulp stem cells. Sci Rep.
5:171772015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mohamed DA, Abdelfattah MI and Aboulezz
EH: The effect of three different biomaterials on proliferation and
viability of human dental pulp stem cells (In-vitro Study). Open
Access Maced J Med Sci. 5:657–663. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kulan P, Karabiyik O, Kose GT and Kargul
B: The effect of accelerated mineral trioxide aggregate on
odontoblastic differentiation in dental pulp stem cell niches. Int
Endod J. 51:758–766. 2018. View Article : Google Scholar
|
|
49
|
Youssef AR, Emara R, Taher MM, Al-Allaf
FA, Almalki M, Almasri MA and Siddiqui SS: Effects of mineral
trioxide aggregate, calcium hydroxide, biodentine and Emdogain on
osteogenesis, Odontogenesis, angiogenesis and cell viability of
dental pulp stem cells. BMC Oral Health. 19:1332019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Minyong W, He L, Shenglin L and Man Q:
Effects of mineral trioxide aggregate on the proliferation and
differentiation of human pulp cells from primary and permanent
teeth. Hua Xi Kou Qiang Yi Xue Za Zhi. 33:75–79. 2015.In Chinese.
PubMed/NCBI
|
|
51
|
Camilleri J, Montesin FE, Papaioannou S,
McDonald F and Pitt Ford TR: Biocompatibility of two commercial
forms of mineral trioxide aggregate. Int Endod J. 37:699–704. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Demirkaya K, Can Demirdöğen B, Öncel Torun
Z, Erdem O, Cetinkaya S and Akay C: In vivo evaluation of the
effects of hydraulic calcium silicate dental cements on plasma and
liver aluminium levels in rats. Eur J Oral Sci. 124:75–81. 2016.
View Article : Google Scholar
|
|
53
|
Chung M, Lee S, Chen D, Kim U, Kim Y, Kim
S and Kim E: Effects of different calcium silicate cements on the
inflammatory response and odontogenic differentiation of
lipopolysaccharide-stimulated human dental pulp stem cells.
Materials (Basel). 12:12592019. View Article : Google Scholar
|
|
54
|
Chen M, Hu DN, Pan Z, Lu CW, Xue CY and
Aass I: Curcumin protects against hyperosmoticity-induced IL-1beta
elevation in human corneal epithelial cell via MAPK pathways. Exp
Eye Res. 90:437–443. 2010. View Article : Google Scholar
|
|
55
|
Tang JJ, Shen ZS, Qin W and Lin Z: A
comparison of the sealing abilities between Biodentine and MTA as
root-end filling materials and their effects on bone healing in
dogs after periradicular surgery. J Appl Oral Sci.
27:e201806932019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang W and Peng B: Tissue reactions after
subcutaneous and intraosseous implantation of iRoot SP, MTA and AH
Plus. Dent Mater J. 34:774–780. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Masuda-Murakami Y, Kobayashi M, Wang X,
Yamada Y, Kimura Y, Hossain M and Matsumoto K: Effects of mineral
trioxide aggregate on the differentiation of rat dental pulp cells.
Acta Histochem. 112:452–458. 2010. View Article : Google Scholar
|
|
58
|
Agrafioti A, Taraslia V, Chrepa V, Lymperi
S, Panopoulos P, Anastasiadou E and Kontakiotis EG: Interaction of
dental pulp stem cells with Biodentine and MTA after exposure to
different environments. J Appl Oral Sci. 24:481–486. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Utneja S, Nawal RR, Talwar S and Verma M:
Current perspectives of bio-ceramic technology in endodontics:
Calcium enriched mixture cement-review of its composition,
properties and applications. Restor Dent Endod. 40:1–13. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bin CV, Valera MC, Camargo SE, Rabelo SB,
Silva GO, Balducci I and Camargo CH: Cytotoxicity and genotoxicity
of root canal sealers based on mineral trioxide aggregate. J Endod.
38:495–500. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu X, Zhao M, Lu J, Ma J, Wei J and Wei
S: Cell responses to two kinds of nanohydroxyapatite with different
sizes and crystallinities. Int J Nanomedicine. 7:1239–1250. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hanafy AK, Shinaishin SF, Eldeen GN and
Aly RM: Nano Hydroxyapatite & mineral trioxide aggregate
efficiently promote odontogenic differentiation of dental pulp stem
cells. Open Access Maced J Med Sci. 6:1727–1731. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Paranjpe A, Zhang H and Johnson JD:
Effects of mineral trioxide aggregate on human dental pulp cells
after pulp-capping procedures. J Endod. 36:1042–1047. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Seo MS, Hwang KG, Lee J, Kim H and Baek
SH: The effect of mineral trioxide aggregate on odontogenic
differentiation in dental pulp stem cells. J Endod. 39:242–248.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Okamoto M, Ali M, Komichi S, Watanabe M,
Huang H, Ito Y, Miura J, Hirose Y, Mizuhira M, Takahashi Y, et al:
Surface pre-reacted glass filler contributes to tertiary dentin
formation through a mechanism different than that of hydraulic
calcium-silicate cement. J Clin Med. 8:14402019. View Article : Google Scholar :
|
|
66
|
Asgary S, Nazarian H, Khojasteh A and
Shokouhinejad N: Gene expression and cytokine release during
odontogenic differentiation of human dental pulp stem cells induced
by 2 endodontic biomaterials. J Endod. 40:387–392. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Paranjpe A, Cacalano NA, Hume WR and
Jewett A: N-acetylcysteine protects dental pulp stromal cells from
HEMA-induced apoptosis by inducing differentiation of the cells.
Free Radic Biol Med. 43:1394–1408. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Caicedo R, Abbott PV, Alongi DJ and
Alarcon MY: Clinical, radiographic and histological analysis of the
effects of mineral trioxide aggregate used in direct pulp capping
and pulpotomies of primary teeth. Aust Dent J. 51:297–305. 2006.
View Article : Google Scholar
|
|
69
|
Maroto M, Barberia E, Planells P and
García Godoy F: Dentin bridge formation after mineral trioxide
aggregate (MTA) pulpotomies in primary teeth. Am J Dent.
18:151–154. 2005.PubMed/NCBI
|
|
70
|
Javid B, Panahandeh N, Torabzadeh H,
Nazarian H, Parhizkar A and Asgary S: Bioactivity of endodontic
biomaterials on dental pulp stem cells through dentin. Restor Dent
Endod. 45:e32019. View Article : Google Scholar
|
|
71
|
Kulan P, Karabiyik O, Kose GT and Kargul
B: Biocompatibility of accelerated mineral trioxide aggregate on
stem cells derived from human dental pulp. J Endod. 42:276–279.
2016. View Article : Google Scholar
|
|
72
|
Kim JH, Kim SY, Woo SM, Jeong HN, Jung JY,
Kim SM and Lim HS: Combination of mineral trioxide aggregate and
propolis promotes odontoblastic differentiation of human dental
pulp stem cells through ERK signaling pathway. Food Sci Biotechnol.
28:1801–1809. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Olcay K, Tasli PN, Guven EP, Ulker G, Ogut
EE, Ciftcioglu E, Kiratli B and Sahin F: Effect of a novel
bioceramic root canal sealer on the angiogenesis-enhancing
potential of assorted human odontogenic stem cells compared with
principal tricalcium silicate-based cements. J Appl Oral Sci.
28:e201902152020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tomas-Catala CJ, Collado-Gonzalez M,
Garcia-Bernal D, Onate-Sanchez RE, Forner L, Llena C, Lozano A,
Moraleda JM and Rodriguez-Lozano FJ: Biocompatibility of New
Pulp-capping Materials NeoMTA Plus, MTA Repair HP, and biodentine
on human dental pulp stem cells. J Endod. 44:126–132. 2018.
View Article : Google Scholar
|
|
75
|
Luo Z, Li D, Kohli MR, Yu Q, Kim S and He
WX: Effect of Biodentine™ on the proliferation, migration and
adhesion of human dental pulp stem cells. J Dent. 42:490–497. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Luo Z, Kohli MR, Yu Q, Kim S, Qu T and He
WX: Biodentine induces human dental pulp stem cell differentiation
through mitogen-activated protein kinase and
calcium-/calmodulin-dependent protein kinase II pathways. J Endod.
40:937–942. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Bortoluzzi EA, Niu LN, Palani CD, El-Awady
AR, Hammond BD, Pei DD, Tian FC, Cutler CW, Pashley DH and Tay FR:
Cytotoxicity and osteogenic potential of silicate calcium cements
as potential protective materials for pulpal revascularization.
Dent Mater. 31:1510–1522. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Jeanneau C, Laurent P, Rombouts C, Giraud
T and About I: Light-cured tricalcium silicate toxicity to the
dental pulp. J Endod. 43:2074–2080. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Weekate K, Chuenjitkuntaworn B, Chuveera
P, Vaseenon S, Chompu-Inwai P, Ittichaicharoen J, Chattipakorn S
and Srisuwan T: Alterations of mitochondrial dynamics, inflammation
and mineralization potential of lipopolysaccharide-induced human
dental pulp cells after exposure to N-acetyl cysteine, Biodentine
or ProRoot MTA. Int Endod J. Jan 27–2021.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Loison-Robert LS, Tassin M, Bonte E,
Berbar T, Isaac J, Berdal A, Simon S and Fournier BPJ: In vitro
effects of two silicate-based materials, Biodentine and BioRoot
RCS, on dental pulp stem cells in models of reactionary and
reparative dentinogenesis. PLoS One. 13:e1900142018. View Article : Google Scholar
|
|
81
|
Kuru S, Sepet E, Irez T, Aktas E, Yazir Y,
Duruksu G, Osmanoglu Akyol E and Erguven M: Effects of different
pulp-capping materials on cell death signaling pathways of
lipoteichoic acid-stimulated human dental pulp stem cells.
Odontology. 109:547–559. 2021. View Article : Google Scholar
|
|
82
|
Petta TM, Pedroni ACF, Saavedra DF, Faial
KDCF, Marques MM and Couto RSD: The effect of three different pulp
capping cements on mineralization of dental pulp stem cells. Dent
Mater J. 39:222–228. 2020. View Article : Google Scholar
|
|
83
|
Tsujimoto M, Ookubo A, Wada Y, Matsunaga
T, Tsujimoto Y and Hayashi Y: Surface changes of mineral trioxide
aggregate after the application of bleaching agents: Electron
microscopy and an energy-dispersive X-ray microanalysis. J Endod.
37:231–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Namazikhah MS, Nekoofar MH, Sheykhrezae
MS, Salariyeh S, Hayes SJ, Bryant ST, Mohammadi MM and Dummer PM:
The effect of pH on surface hardness and microstructure of mineral
trioxide aggregate. Int Endod J. 41:108–116. 2008.
|
|
85
|
Shie MY, Huang TH, Kao CT, Huang CH and
Ding SJ: The effect of a physiologic solution pH on properties of
white mineral trioxide aggregate. J Endod. 35:98–101. 2009.
View Article : Google Scholar
|
|
86
|
Widbiller M, Lindner SR, Buchalla W, Eidt
A, Hiller KA, Schmalz G and Galler KM: Three-dimensional culture of
dental pulp stem cells in direct contact to tricalcium silicate
cements. Clin Oral Investig. 20:237–246. 2016. View Article : Google Scholar
|
|
87
|
Chen S, Gluhak-Heinrich J, Wang YH, Wu YM,
Chuang HH, Chen L, Yuan GH, Dong J, Gay I and MacDougall M: Runx2,
osx, and dspp in tooth development. J Dent Res. 88:904–909. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Li S, Kong H, Yao N, Yu Q, Wang P, Lin Y,
Wang J, Kuang R, Zhao X, Xu J, et al: The role of runt-related
transcription factor 2 (Runx2) in the late stage of odontoblast
differentiation and dentin formation. Biochem Biophys Res Commun.
410:698–704. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lambrichts I, Driesen RB, Dillen Y,
Gervois P, Ratajczak J, Vangansewinkel T, Wolfs E, Bronckaers A and
Hilkens P: Dental pulp stem cells: Their potential in reinnervation
and angiogenesis by using Scaffolds. J Endod. 43:S12–S16. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Brown LF, Detmar M, Claffey K, Nagy JA,
Feng D, Dvorak AM and Dvorak HF: Vascular permeability
factor/vascular endothelial growth factor: A multifunctional
angiogenic cytokine. EXS. 79:233–269. 1997.PubMed/NCBI
|
|
91
|
Folkman J and Shing Y: Angiogenesis. J
Biol Chem. 267:10931–10934. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bai F, Wang Z, Lu J, Liu J, Chen G, Lv R,
Wang J, Lin K, Zhang J and Huang X: The correlation between the
internal structure and vascularization of controllable porous
bioceramic materials in vivo: A quantitative study. Tissue Eng Part
A. 16:3791–3803. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wang J, Fangteng JZ and Liu H: Effect of
iRoot BP Plus on biological behavior of deciduous tooth pulp stem
cells and human pulp stem cells. Shanghai Kou Qiang Yi Xue.
28:251–258. 2019.In Chinese. PubMed/NCBI
|
|
94
|
Zhu L, Yang J, Zhang J, Lei D, Xiao L,
Cheng X, Lin Y and Peng B: In vitro and in vivo evaluation of a
nanoparticulate bioceramic paste for dental pulp repair. Acta
Biomater. 10:5156–5168. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kuo JC: Mechanotransduction at focal
adhesions: Integrating cytoskeletal mechanics in migrating cells. J
Cell Mol Med. 17:704–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Plotnikov SV and Waterman CM: Guiding cell
migration by tugging. Curr Opin Cell Biol. 25:619–626. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Sun Y, Luo T, Shen Y, Haapasalo M, Zou L
and Liu J: Effect of iRoot fast set root repair material on the
proliferation, migration and differentiation of human dental pulp
stem cells in vitro. PLoS One. 12:e01868482017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Sun Y, Liu J, Luo T, Shen Y and Zou L:
Effects of two fast-setting pulp-capping materials on cell
viability and osteogenic differentiation in human dental pulp stem
cells: An in vitro study. Arch Oral Biol. 100:100–105. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Collado-Gonzalez M, Lopez-Garcia S,
Garcia-Bernal D, Onate-Sanchez RE, Tomas-Catala CJ, Moraleda JM,
Lozano A, Forner L and Rodriguez-Lozano FJ: Biological effects of
acid-eroded MTA Repair HP and ProRoot MTA on human periodontal
ligament stem cells. Clin Oral Investig. 23:3915–3924. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Collado-Gonzalez M, Garcia-Bernal D,
Onate-Sanchez RE, Ortolani-Seltenerich PS, Lozano A, Forner L,
Llena C and Rodriguez-Lozano FJ: Biocompatibility of three new
calcium silicate-based endodontic sealers on human periodontal
ligament stem cells. Int Endod J. 50:875–884. 2017. View Article : Google Scholar
|
|
101
|
Collado-Gonzalez M, Tomas-Catala CJ,
Onate-Sanchez RE, Moraleda JM and Rodriguez-Lozano FJ: Cytotoxicity
of GuttaFlow Bioseal, GuttaFlow2, MTA Fillapex, and AH Plus on
human periodontal ligament stem cells. J Endod. 43:816–822. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Rodriguez-Lozano FJ, Garcia-Bernal D,
Onate-Sanchez RE, Ortolani-Seltenerich PS, Forner L and Moraleda
JM: Evaluation of cytocompatibility of calcium silicate-based
endodontic sealers and their effects on the biological responses of
mesenchymal dental stem cells. Int Endod J. 50:67–76. 2017.
View Article : Google Scholar
|
|
103
|
Wang Y, Zhou Y, Jin L, Pang X, Lu Y, Wang
Z, Yu Y and Yu J: Mineral trioxide aggregate enhances the
osteogenic capacity of periodontal ligament stem cells via NF-κB
and MAPK signaling pathways. J Cell Physiol. 233:2386–2397. 2018.
View Article : Google Scholar
|
|
104
|
Abuarqoub D, Aslam N, Jafar H, Abu Harfil
Z and Awidi A: Biocompatibility of Biodentine™(R) with periodontal
ligament stem cells: In vitro study. Dent J (Basel). 8:172020.
View Article : Google Scholar
|
|
105
|
Hakki SS, Bozkurt SB, Ozcopur B, Purali N
and Belli S: Periodontal ligament fibroblast response to root
perforations restored with different materials: A laboratory study.
Int Endod J. 45:240–248. 2012. View Article : Google Scholar
|
|
106
|
Lee MN, Hwang HS, Oh SH, Roshanzadeh A,
Kim JW, Song JH, Kim ES and Koh JT: Elevated extracellular calcium
ions promote proliferation and migration of mesenchymal stem cells
via increasing osteopontin expression. Exp Mol Med. 50:1–16. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
D'Anto V, Di Caprio MP, Ametrano G,
Simeone M, Rengo S and Spagnuolo G: Effect of mineral trioxide
aggregate on mesenchymal stem cells. J Endod. 36:1839–1843. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Eid AA, Hussein KA, Niu LN, Li GH,
Watanabe I, Al-Shabrawey M, Pashley DH and Tay FR: Effects of
tricalcium silicate cements on osteogenic differentiation of human
bone marrow-derived mesenchymal stem cells in vitro. Acta Biomater.
10:3327–3334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Edrees HY, Abu Zeid ST, Atta HM and
AlQriqri MA: Induction of osteogenic differentiation of mesenchymal
stem cells by bioceramic root repair material. Materials (Basel).
12:23112019. View Article : Google Scholar
|
|
110
|
Wang Y, Li J, Song W and Yu J: Mineral
trioxide aggregate upregulates odonto/osteogenic capacity of bone
marrow stromal cells from craniofacial bones via JNK and ERK MAPK
signalling pathways. Cell Prolif. 47:241–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Vidovic Zdrilic I, de Azevedo Queiroz IO,
Matthews BG, Gomes-Filho JE, Mina M and Kalajzic I: Mineral
trioxide aggregate improves healing response of periodontal tissue
to injury in mice. J Periodontal Res. 52:1058–1067. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Moon HJ, Lee JH, Kim JH, Knowles JC, Cho
YB, Shin DH, Lee HH and Kim HW: Reformulated mineral trioxide
aggregate components and the assessments for use as future dental
regenerative cements. J Tissue Eng. 9:20417314188073962018.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Gandolfi MG, Ciapetti G, Perut F, Taddei
P, Modena E, Rossi PL and Prati C: Biomimetic calcium-silicate
cements aged in simulated body solutions. Osteoblast response and
analyses of apatite coating. J Appl Biomater Biomech. 7:160–170.
2009.
|
|
114
|
Lu J, Li Z, Wu X, Chen Y, Yan M, Ge X and
Yu J: iRoot BP Plus promotes osteo/odontogenic differentiation of
bone marrow mesenchymal stem cells via MAPK pathways and autophagy.
Stem Cell Res Ther. 10:2222019. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
De-Deus G, Canabarro A, Alves G, Linhares
A, Senne MI and Granjeiro JM: Optimal cytocompatibility of a
bioceramic nanoparticulate cement in primary human mesenchymal
cells. J Endod. 35:1387–1390. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Fathy SM, Abd El-Aziz AM and Labah DA:
Cellular interaction and antibacterial efficacy of two hydraulic
calcium silicate-based cements: Cell-dependent model. J Conserv
Dent. 22:17–22. 2019.PubMed/NCBI
|
|
117
|
Margunato S, Tasli PN, Aydin S, Karapınar
Kazandağ M and Sahin F: In Vitro evaluation of ProRoot MTA,
biodentine, and MM-MTA on human alveolar bone marrow stem cells in
terms of biocompatibility and mineralization. J Endod.
41:1646–1652. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Sultana N, Singh M, Nawal RR, Chaudhry S,
Yadav S, Mohanty S and Talwar S: Evaluation of biocompatibility and
osteogenic potential of tricalcium silicate-based cements using
human bone marrow-derived mesenchymal stem cells. J Endod.
44:446–451. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Costa F, Sousa Gomes P and Fernandes MH:
Osteogenic and angiogenic response to calcium silicate-based
endodontic sealers. J Endod. 42:113–119. 2016. View Article : Google Scholar
|
|
120
|
Ali MRW, Mustafa M, Bardsen A and Bletsa
A: Tricalcium silicate cements: Osteogenic and angiogenic responses
of human bone marrow stem cells. Eur J Oral Sci. 127:261–268. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Grech L, Mallia B and Camilleri J:
Characterization of set Intermediate Restorative Material,
Biodentine, Bioaggregate and a prototype calcium silicate cement
for use as root-end filling materials. Int Endod J. 46:632–641.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Kang JY, Lee BN, Son HJ, Koh JT, Kang SS,
Son HH, Chang HS, Hwang IN, Hwang YC and Oh WM: Biocompatibility of
mineral trioxide aggregate mixed with hydration accelerators. J
Endod. 39:497–500. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Silva EJ, Rosa TP, Herrera DR, Jacinto RC,
Gomes BP and Zaia AA: Evaluation of cytotoxicity and
physicochemical properties of calcium silicate-based endodontic
sealer MTA Fillapex. J Endod. 39:274–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Estrela C, Alencar AH, Kitten GT, Vencio
EF and Gava E: Mesenchymal stem cells in the dental tissues:
Perspectives for tissue regeneration. Braz Dent J. 22:91–98. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Collado-Gonzalez M, Garcia-Bernal D,
Onate-Sanchez RE, Ortolani-Seltenerich PS, Alvarez-Muro T, Lozano
A, Forner L, Llena C, Moraleda JM and Rodriguez-Lozano FJ:
Cytotoxicity and bioactivity of various pulpotomy materials on stem
cells from human exfoliated primary teeth. Int Endod J. 50(Suppl
2): e19–e30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Araujo LB, Cosme-Silva L, Fernandes AP,
Oliveira TM, Cavalcanti BDN, Gomes Filho JE and Sakai VT: Effects
of mineral trioxide aggregate, BiodentineTM and calcium hydroxide
on viability, proliferation, migration and differentiation of stem
cells from human exfoliated deciduous teeth. J Appl Oral Sci.
26:e201606292018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Dahake PT, Panpaliya NP, Kale YJ, Dadpe
MV, Kendre SB and Bogar C: Response of stem cells from human
exfoliated deciduous teeth (SHED) to three bioinductive
materials-An in vitro experimental study. Saudi Dent J. 32:43–51.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Hasweh N, Awidi A, Rajab L, Hiyasat A,
Jafar H, Islam N, Hasan M and Abuarqoub D: Characterization of the
biological effect of Biodentine(TM) on primary dental pulp stem
cells. Indian J Dent Res. 29:787–793. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Athanasiadou E, Paschalidou M,
Theocharidou A, Kontoudakis N, Arapostathis K and Bakopoulou A:
Biological interactions of a calcium silicate based cement
(Biodentine) with stem cells from human exfoliated deciduous teeth.
Dent Mater. 34:1797–1813. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Tsai CL, Ke MC, Chen YH, Kuo HK, Yu HJ,
Chen CT, Tseng YC, Chuang PC and Wu PC: Mineral trioxide aggregate
affects cell viability and induces apoptosis of stem cells from
human exfoliated deciduous teeth. BMC Pharmacol Toxicol. 19:212018.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Saidon J, He J, Zhu Q, Safavi K and
Spangberg LS: Cell and tissue reactions to mineral trioxide
aggregate and Portland cement. Oral Surg Oral Med Oral Pathol Oral
Radiol Endod. 95:483–489. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Torabinejad M, Hong CU, McDonald F and
Pitt Ford TR: Physical and chemical properties of a new root-end
filling material. J Endod. 21:349–353. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wongwatanasanti N, Jantarat J,
Sritanaudomchai H and Hargreaves KM: Effect of bioceramic materials
on proliferation and odontoblast differentiation of human stem
cells from the apical papilla. J Endod. 44:1270–1275. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Peters OA, Galicia J, Arias A, Tolar M, Ng
E and Shin SJ: Effects of two calcium silicate cements on cell
viability, angiogenic growth factor release and related gene
expression in stem cells from the apical papilla. Int Endod J.
49:1132–1140. 2016. View Article : Google Scholar
|
|
135
|
Schneider R, Holland GR, Chiego D Jr, Hu
JC, Nör JE and Botero TM: White mineral trioxide aggregate induces
migration and proliferation of stem cells from the apical papilla.
J Endod. 40:931–936. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Saberi EA, Karkehabadi H and Mollashahi
NF: Cytotoxicity of various endodontic materials on stem cells of
human apical papilla. Iran Endod J. 11:17–22. 2016.PubMed/NCBI
|
|
137
|
Miller AA, Takimoto K, Wealleans J and
Diogenes A: Effect of 3 bioceramic materials on stem cells of the
apical papilla proliferation and differentiation using a dentin
disk model. J Endod. 44:599–603. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Du J, Lu Y, Song M, Yang L, Liu J, Chen X,
Ma Y and Wang Y: Effects of ERK/p38 MAPKs signaling pathways on
MTA-mediated osteo/odontogenic differentiation of stem cells from
apical papilla: A vitro study. BMC Oral Health. 20:502020.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Saberi E, Farhad-Mollashahi N, Sargolzaei
Aval F and Saberi M: Proliferation, odontogenic/osteogenic
differentiation, and cytokine production by human stem cells of the
apical papilla induced by biomaterials: A comparative study. Clin
Cosmet Investig Dent. 11:181–193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Hajizadeh N, Madani ZS, Zabihi E, Golpour
M, Zahedpasha A and Mohammadnia M: Effect of MTA and CEM on
mineralization-associated gene expression in stem cells derived
from apical papilla. Iran Endod J. 13:94–101. 2018.PubMed/NCBI
|
|
141
|
Chang J, Liu F, Lee M, Wu B, Ting K, Zara
JN, Soo C, Al Hezaimi K, Zou W, Chen X, et al: NF-κB inhibits
osteogenic differentiation of mesenchymal stem cells by promoting
β-catenin degradation. Proc Natl Acad Sci USA. 110:9469–9474. 2013.
View Article : Google Scholar
|
|
142
|
Yan M, Wu J, Yu Y, Wang Y, Xie L, Zhang G,
Yu J and Zhang C: Mineral trioxide aggregate promotes the
odonto/osteogenic differentiation and dentinogenesis of stem cells
from apical papilla via nuclear factor kappa B signaling pathway. J
Endod. 40:640–647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Li H and Chang J: Bioactive silicate
materials stimulate angiogenesis in fibroblast and endothelial cell
co-culture system through paracrine effect. Acta Biomater.
9:6981–6991. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Day RM: Bioactive glass stimulates the
secretion of angiogenic growth factors and angiogenesis in vitro.
Tissue Eng. 11:768–777. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Leu A and Leach JK: Proangiogenic
potential of a collagen/bioactive glass substrate. Pharm Res.
25:1222–1229. 2008. View Article : Google Scholar
|
|
146
|
Wang G, Roohani-Esfahani SI, Zhang W, Lv
K, Yang G, Ding X, Zou D, Cui D, Zreiqat H and Jiang X: Effects of
Sr-HT-Gahnite on osteogenesis and angiogenesis by adipose derived
stem cells for critical-sized calvarial defect repair. Sci Rep.
7:411352017. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Liu Y, Liu XM, Bi J, Yu S, Yang N, Song B
and Chen X: Cell migration and osteo/odontogenesis stimulation of
iRoot FS as a potential apical barrier material in apexification.
Int Endod J. 53:467–477. 2020. View Article : Google Scholar
|
|
148
|
Bi J, Liu Y, Liu XM, Jiang LM and Chen X:
iRoot FM exerts an antibacterial effect on Porphyromonas
endodontalis and improves the properties of stem cells from the
apical papilla. Int Endod J. 51:1139–1148. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Dogan A, Yalvac ME, Sahin F, Kabanov AV,
Palotas A and Rizvanov AA: Differentiation of human stem cells is
promoted by amphiphilic pluronic block copolymers. Int J
Nanomedicine. 7:4849–4860. 2012.PubMed/NCBI
|
|
150
|
Dogan A, Demirci S and Sahin F: In vitro
differentiation of human tooth germ stem cells into endothelial-
and epithelial-like cells. Cell Biol Int. 39:94–103. 2015.
View Article : Google Scholar
|
|
151
|
Guven EP, Yalvac ME, Sahin F, Yazici MM,
Rizvanov AA and Bayirli G: Effect of dental materials calcium
hydroxidecontaining cement, mineral trioxide aggregate, and enamel
matrix derivative on proliferation and differentiation of human
tooth germ stem cells. J Endod. 37:650–656. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Guven EP, Yalvac ME, Kayahan MB, Sunay H,
Sahin F and Bayirli G: Human tooth germ stem cell response to
calcium-silicate based endodontic cements. J Appl Oral Sci.
21:351–357. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Güven EP, Taşlı PN, Yalvac ME, Sofiev N,
Kayahan MB and Sahin F: In vitro comparison of induction capacity
and biomineralization ability of mineral trioxide aggregate and a
bioceramic root canal sealer. Int Endod J. 46:1173–1182. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Jiang N, Guo W, Chen M, Zheng Y, Zhou J,
Kim SG, Embree MC, Songhee Song K, Marao HF and Mao JJ: Periodontal
ligament and alveolar bone in health and adaptation: Tooth
movement. Front Oral Biol. 18:1–8. 2016.
|
|
155
|
Zhou Y, Wu C and Xiao Y: Silicate-based
bioceramics for periodontal regeneration. J Mater Chem B.
2:3907–3910. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Proksch S, Brossart J, Vach K, Hellwig E,
Altenburger MJ and Karygianni L: Evaluation of the bioactivity of
fluoride-enriched mineral trioxide aggregate on osteoblasts. Int
Endod J. 51:912–923. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Nakayama A, Ogiso B, Tanabe N, Takeichi O,
Matsuzaka K and Inoue T: Behaviour of bone marrow osteoblast-like
cells on mineral trioxide aggregate: Morphology and expression of
type I collagen and bone-related protein mRNAs. Int Endod J.
38:203–210. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Castro-Raucci LM, Teixeira LN, Oliveira
IR, Raucci-Neto W, Jacobovitz M, Rosa AL and de Oliveira PT:
Osteogenic cell response to calcium aluminate-based cement. Int
Endod J. 50:771–779. 2017. View Article : Google Scholar
|
|
159
|
Castro-Raucci LM, Oliveira IR, Teixeira
LN, Rosa AL, Oliveira PT and Jacobovitz M: Effects of a novel
calcium aluminate cement on the early events of the progression of
osteogenic cell cultures. Braz Dent J. 22:99–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Orrenius S, Zhivotovsky B and Nicotera P:
Regulation of cell death: The calcium-apoptosis link. Nat Rev Mol
Cell Biol. 4:552–565. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Dvorak MM and Riccardi D: Ca2+
as an extracellular signal in bone. Cell Calcium. 35:249–255. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Oliveira IR, Andrade TL, Jacobovitz M and
Pandolfelli VC: Bioactivity of calcium aluminate endodontic cement.
J Endod. 39:774–778. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Pires-de-Souza FC, Moraes PC, Garcia Lda
F, Aguilar FG and Watanabe E: Evaluation of pH, calcium ion release
and antimicrobial activity of a new calcium aluminate cement. Braz
Oral Res. 27:324–330. 2013. View Article : Google Scholar
|
|
164
|
Scelza MZ, Nascimento JC, Silva LE,
Gameiro VS, DE Deus G and Alves G: Biodentine™ is cytocompatible
with human primary osteoblasts. Braz Oral Res. 31:e812017.
View Article : Google Scholar
|
|
165
|
Coelho MJ and Fernandes MH: Human bone
cell cultures in biocompatibility testing. Part II: Effect of
ascorbic acid, beta-glycerophosphate and dexamethasone on
osteoblastic differentiation. Biomaterials. 21:1095–1102. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Rifaey HS, Villa M, Zhu Q, Wang YH, Safavi
K and Chen IP: Comparison of the osteogenic potential of mineral
trioxide aggregate and endosequence root repair material in a
3-dimensional culture system. J Endod. 42:760–765. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Ciasca M, Aminoshariae A, Jin G,
Montagnese T and Mickel A: A comparison of the cytotoxicity and
proinflammatory cytokine production of EndoSequence root repair
material and ProRoot mineral trioxide aggregate in human osteoblast
cell culture using reverse-transcriptase polymerase chain reaction.
J Endod. 38:486–489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Tani-Ishii N, Hamada N, Watanabe K,
Tujimoto Y, Teranaka T and Umemoto T: Expression of bone
extracellular matrix proteins on osteoblast cells in the presence
of mineral trioxide. J Endod. 33:836–839. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Yoshimine Y, Ono M and Akamine A: In vitro
comparison of the biocompatibility of mineral trioxide aggregate,
4META/MMA-TBB resin, and intermediate restorative material as
root-end-filling materials. J Endod. 33:1066–1069. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Maeda T, Suzuki A, Yuzawa S, Baba Y,
Kimura Y and Kato Y: Mineral trioxide aggregate induces
osteoblastogenesis via Atf6. Bone Rep. 2:36–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Lee GW, Yoon JH, Jang JH, Chang HS, Hwang
YC, Hwang IN, Oh WM and Lee BN: Effects of newly-developed
retrograde filling material on osteoblastic differentiation in
vitro. Dent Mater J. 38:528–533. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Sarkar NK, Caicedo R, Ritwik P, Moiseyeva
R and Kawashima I: Physicochemical basis of the biologic properties
of mineral trioxide aggregate. J Endod. 31:97–100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Estrela C, Sydney GB, Pesce HF and Felippe
Júnior O: Dentinal diffusion of hydroxyl ions of various calcium
hydroxide pastes. Braz Dent J. 6:5–9. 1995.PubMed/NCBI
|
|
174
|
Ko H, Yang W, Park K and Kim M:
Cytotoxicity of mineral trioxide aggregate (MTA) and bone
morphogenetic protein 2 (BMP-2) and response of rat pulp to MTA and
BMP-2. Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
109:e103–e108. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Wozney JM: The bone morphogenetic protein
family: Multifunctional cellular regulators in the embryo and
adult. Eur J Oral Sci. 106(Suppl 1): S160–S166. 1998. View Article : Google Scholar
|
|
176
|
Zhu Q, Haglund R, Safavi KE and Spangberg
LS: Adhesion of human osteoblasts on root-end filling materials. J
Endod. 26:404–406. 2000. View Article : Google Scholar
|
|
177
|
Pelliccioni GA, Ciapetti G, Cenni E,
Granchi D, Nanni M, Pagani S and Giunti A: Evaluation of
osteoblast-like cell response to Proroot MTA (mineral trioxide
aggregate) cement. J Mater Sci Mater Med. 15:167–173. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Tanomaru-Filho M, Andrade AS, Rodrigues
EM, Viola KS, Faria G, Camilleri J and Guerreiro-Tanomaru JM:
Biocompatibility and mineralized nodule formation of Neo MTA Plus
and an experimental tricalcium silicate cement containing tantalum
oxide. Int Endod J. 50(Suppl 2): e31–e39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Modareszadeh MR, Di Fiore PM, Tipton DA
and Salamat N: Cytotoxicity and alkaline phosphatase activity
evaluation of endosequence root repair material. J Endod.
38:1101–1105. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Yuan Z, Peng B, Jiang H, Bian Z and Yan P:
Effect of bioaggregate on mineral-associated gene expression in
osteoblast cells. J Endod. 36:1145–1148. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Jung S, Mielert J, Kleinheinz J and
Dammaschke T: Human oral cells' response to different endodontic
restorative materials: An in vitro study. Head Face Med. 10:552014.
View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Attik GN, Villat C, Hallay F,
Pradelle-Plasse N, Bonnet H, Moreau K, Colon P and Grosgogeat B: In
vitro biocompatibility of a dentine substitute cement on human MG63
osteoblasts cells: Biodentine versus MTA((®)). Int Endod J.
47:1133–1141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Kim HS, Kim S, Ko H, Song M and Kim M:
Effects of the cathepsin K inhibitor with mineral trioxide
aggregate cements on osteoclastic activity. Restor Dent Endod.
44:e172019. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Gomes-Cornelio AL, Rodrigues EM, Salles
LP, Mestieri LB, Faria G, Guerreiro-Tanomaru JM and Tanomaru-Filho
M: Bioactivity of MTA Plus, Biodentine and an experimental calcium
silicate-based cement on human osteoblast-like cells. Int Endod J.
50:39–47. 2017. View Article : Google Scholar
|
|
185
|
De-Deus G, Canabarro A, Alves GG, Marins
JR, Linhares AB and Granjeiro JM: Cytocompatibility of the
ready-to-use bioceramic putty repair cement iRoot BP Plus with
primary human osteoblasts. Int Endod J. 45:508–513. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Tian J, Zhang Y, Lai Z, Li M, Huang Y,
Jiang H and Wei X: Ion release, microstructural, and biological
properties of iRoot BP Plus and ProRoot MTA exposed to an acidic
environment. J Endod. 43:163–168. 2017. View Article : Google Scholar
|
|
187
|
Lv F, Zhu L, Zhang J, Yu J, Cheng X and
Peng B: Evaluation of the in vitro biocompatibility of a new
fast-setting ready-to-use root filling and repair material. Int
Endod J. 50:540–548. 2017. View Article : Google Scholar
|
|
188
|
Jiang Y, Zheng Q, Zhou X, Gao Y and Huang
D: A comparative study on root canal repair materials: A
cytocompatibility assessment in L929 and MG63 cells.
ScientificWorldJournal. 2014:4638262014. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Levine BR, Sporer S, Poggie RA, Della
Valle CJ and Jacobs JJ: Experimental and clinical performance of
porous tantalum in orthopedic surgery. Biomaterials. 27:4671–4681.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Zhang W, Li Z and Peng B: Effects of iRoot
SP on mineralization-related genes expression in MG63 cells. J
Endod. 36:1978–1982. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Zhang J, Zhu L and Peng B: Effect of
BioAggregate on osteoclast differentiation and inflammatory bone
resorption in vivo. Int Endod J. 48:1077–1085. 2015. View Article : Google Scholar
|
|
192
|
Tian J, Qi W, Zhang Y, Glogauer M, Wang Y,
Lai Z and Jiang H: Bioaggregate inhibits osteoclast
differentiation, fusion, and bone resorption in vitro. J Endodont.
41:1500–1506. 2015. View Article : Google Scholar
|
|
193
|
Zhang J, Zhu L, Yan P and Peng B: Effect
of BioAggregate on receptor activator of nuclear factor-kappa B
ligand-induced osteoclastogenesis from murine macrophage cell line
in vitro. J Endod. 41:1265–1271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Cheng X, Zhu L, Zhang J, Yu J, Liu S, Lv
F, Lin Y, Liu G and Peng B: Anti-osteoclastogenesis of mineral
trioxide aggregate through inhibition of the autophagic pathway. J
Endodont. 43:766–773. 2017. View Article : Google Scholar
|
|
195
|
Hashiguchi D, Fukushima H, Nakamura M,
Morikawa K, Yasuda H, Udagawa N, Maki K and Jimi E: Mineral
trioxide aggregate solution inhibits osteoclast differentiation
through the maintenance of osteoprotegerin expression in
osteoblasts. J Biomed Mater Res A. 96:358–364. 2011. View Article : Google Scholar
|
|
196
|
Hashiguchi D, Fukushima H, Yasuda H,
Masuda W, Tomikawa M, Morikawa K, Maki K and Jimi E: Mineral
trioxide aggregate inhibits osteoclastic bone resorption. J Dent
Res. 90:912–917. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Kim M, Kim S, Ko H and Song M: Effect of
ProRoot MTA® and Biodentine® on osteoclastic
differentiation and activity of mouse bone marrow macrophages. J
Appl Oral Sci. 27:e201801502019. View Article : Google Scholar
|
|
198
|
Choi SC, Kwon YD, Kim KC and Kim GT: The
effects of topical application of bisphosphonates on replanted rat
molars. Dent Traumatol. 26:476–480. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Komatsu K, Shimada A, Shibata T, Shimoda
S, Oida S, Kawasaki K and Nifuji A: Long-term effects of local
pretreatment with alendronate on healing of replanted rat teeth. J
Periodontal Res. 43:194–200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Tu MG, Sun KT, Wang TH, He YZ, Hsia SM,
Tsai BH, Shih YH and Shieh TM: Effects of mineral trioxide
aggregate and bioceramics on macrophage differentiation and
polarization in vitro. J Formos Med Assoc. 118:1458–1465. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Camilleri J: Characterization of hydration
products of mineral trioxide aggregate. Int Endod J. 41:408–417.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Varanasi VG, Leong KK, Dominia LM, Jue SM,
Loomer PM and Marshall GW: Si and Ca individually and
combinatorially target enhanced MC3T3-E1 subclone 4 early
osteogenic marker expression. J Oral Implantol. 38:325–336. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Xia L, Yin Z, Mao L, Wang X, Liu J, Jiang
X, Zhang Z, Lin K, Chang J and Fang B: Akermanite bioceramics
promote osteogenesis, angiogenesis and suppress osteoclastogenesis
for osteoporotic bone regeneration. Sci Rep. 6:220052016.
View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Wu C, Chen Z, Yi D, Chang J and Xiao Y:
Multidirectional effects of Sr-, Mg-, and Si-containing bioceramic
coatings with high bonding strength on inflammation,
osteoclastogenesis, and osteogenesis. ACS Appl Mater Interfaces.
6:4264–4276. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Hung CJ, Kao CT, Chen YJ, Shie MY and
Huang TH: Antiosteoclastogenic activity of silicate-based materials
antagonizing receptor activator for nuclear factor kappaB
ligand-induced osteoclast differentiation of murine marcophages. J
Endod. 39:1557–1561. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Sun T, Wang M, Shao Y, Wang L and Zhu Y:
The effect and osteoblast signaling response of trace silicon
doping hydroxyapatite. Biol Trace Elem Res. 181:82–94. 2018.
View Article : Google Scholar
|
|
207
|
Wang S, Wang X, Draenert FG, Albert O,
Schroder HC, Mailander V, Mitov G and Muller WE: Bioactive and
biodegradable silica biomaterial for bone regeneration. Bone.
67:292–304. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Zhai W, Lu H, Wu C, Chen L, Lin X, Naoki
K, Chen G and Chang J: Stimulatory effects of the ionic products
from Ca-Mg-Si bioceramics on both osteogenesis and angiogenesis in
vitro. Acta Biomater. 9:8004–8014. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Gough JE, Notingher I and Hench LL:
Osteoblast attachment and mineralized nodule formation on rough and
smooth 45S5 bioactive glass monoliths. J Biomed Mater Res A.
68:640–650. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Gough JE, Clupper DC and Hench LL:
Osteoblast responses to tape-cast and sintered bioactive glass
ceramics. J Biomed Mater Res A. 69:621–628. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Ibrahim S, Sabudin S, Sahid S, Marzuke MA,
Hussin ZH, Kader Bashah NS and Jamuna-Thevi K: Bioactivity studies
and adhesion of human osteoblast (hFOB) on silicon-biphasic calcium
phosphate material. Saudi J Biol Sci. 23:S56–S63. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Caudrillier A, Hurtel-Lemaire AS, Wattel
A, Cournarie F, Godin C, Petit L, Petit JP, Terwilliger E, Kamel S,
Brown EM, et al: Strontium ranelate decreases receptor activator of
nuclear factor-κB ligand-induced osteoclastic differentiation in
vitro: Involvement of the calcium-sensing receptor. Mol Pharmacol.
78:569–576. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Mladenovic Z, Johansson A, Willman B,
Shahabi K, Bjorn E and Ransjo M: Soluble silica inhibits osteoclast
formation and bone resorption in vitro. Acta Biomater. 10:406–418.
2014. View Article : Google Scholar
|
|
214
|
Okabe T, Sakamoto M, Takeuchi H and
Matsushima K: Effects of pH on mineralization ability of human
dental pulp cells. J Endod. 32:198–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Stuart CH, Schwartz SA, Beeson TJ and
Owatz CB: Enterococcus faecalis: Its role in root canal treatment
failure and current concepts in retreatment. J Endod. 32:93–98.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Gao J, Wang M, Shi C, Wang L, Wang D and
Zhu Y: Synthesis of trace element Si and Sr codoping hydroxyapatite
with non-cytotoxicity and enhanced cell proliferation and
differentiation. Biol Trace Elem Res. 174:208–217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Lin LM and Rosenberg PA: Repair and
regeneration in endodontics. Int Endod J. 44:889–906. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Koulaouzidou EA, Economides N, Beltes P,
Geromichalos G and Papazisis K: In vitro evaluation of the
cytotoxicity of ProRoot MTA and MTA Angelus. J Oral Sci.
50:397–402. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Peng W, Liu W, Zhai W, Jiang L, Li L,
Chang J and Zhu Y: Effect of tricalcium silicate on the
proliferation and odontogenic differentiation of human dental pulp
cells. J Endod. 37:1240–1246. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Kim DH, Jang JH, Lee BN, Chang HS, Hwang
IN, Oh WM, Kim SH, Min KS, Koh JT and Hwang YC: Anti-inflammatory
and mineralization effects of ProRoot MTA and endocem MTA in
studies of human and rat dental pulps in vitro and in vivo. J
Endod. 44:1534–1541. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Liu CH, Huang TH, Hung CJ, Lai WY, Kao CT
and Shie MY: The synergistic effects of fibroblast growth factor-2
and mineral trioxide aggregate on an osteogenic accelerator in
vitro. Int Endod J. 47:843–853. 2014. View Article : Google Scholar
|
|
222
|
Chung CJ, Kim E, Song M, Park JW and Shin
SJ: Effects of two fast-setting calcium-silicate cements on cell
viability and angiogenic factor release in human pulp-derived
cells. Odontology. 104:143–151. 2016. View Article : Google Scholar
|
|
223
|
Chang SW, Lee SY, Kum KY and Kim EC:
Effects of ProRoot MTA, Bioaggregate, and Micromega MTA on
odontoblastic differentiation in human dental pulp cells. J Endod.
40:113–118. 2014. View Article : Google Scholar
|
|
224
|
Jung JY, Woo SM, Lee BN, Koh JT, Nor JE
and Hwang YC: Effect of Biodentine and Bioaggregate on
odontoblastic differentiation via mitogen-activated protein kinase
pathway in human dental pulp cells. Int Endod J. 48:177–184. 2015.
View Article : Google Scholar
|
|
225
|
Zhu L, Yang J, Zhang J and Peng B: A
comparative study of BioAggregate and ProRoot MTA on adhesion,
migration, and attachment of human dental pulp cells. J Endod.
40:1118–1123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Zhang S, Yang X and Fan M: BioAggregate
and iRoot BP Plus optimize the proliferation and mineralization
ability of human dental pulp cells. Int Endod J. 46:923–929. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Tomson PL, Grover LM, Lumley PJ, Sloan AJ,
Smith AJ and Cooper PR: Dissolution of bio-active dentine matrix
components by mineral trioxide aggregate. J Dent. 35:636–642. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Laurent P, Camps J and About I:
Biodentine(TM) induces TGF-β1 release from human pulp cells and
early dental pulp mineralization. Int Endod J. 45:439–448. 2012.
View Article : Google Scholar
|
|
229
|
Zanini M, Sautier JM, Berdal A and Simon
S: Biodentine induces immortalized murine pulp cell differentiation
into odontoblast-like cells and stimulates biomineralization. J
Endod. 38:1220–1226. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Liu S, Wang S and Dong Y: Evaluation of a
bioceramic as a pulp capping agent in vitro and in vivo. J Endod.
41:652–657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Zhang J, Zhu LX, Cheng X, Lin Y, Yan P and
Peng B: Promotion of dental pulp cell migration and pulp repair by
a bioceramic putty involving FGFR-mediated signaling pathways. J
Dent Res. 94:853–862. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Kierat A, Laszczynska M, Kowalska E and
Weyna E: Comparison of the influence of mineral trioxide aggregate
and calcium hydroxide on dental pulp of permanent teeth in
biological treatment and cell cultures. Ann Acad Med Stetin.
56:89–96. 2010.In Polish.
|
|
233
|
Zakerzadeh A, Esnaashari E and Dadfar S:
In Vitro comparison of cytotoxicity and genotoxicity of three vital
pulp capping materials. Iran Endod J. 12:419–425. 2017.PubMed/NCBI
|
|
234
|
Nikfarjam F, Beyer K, Konig A, Hofmann M,
Butting M, Valesky E, Kippenberger S, Kaufmann R, Heidemann D,
Bernd A and Zöller NN: Influence of Biodentine(R)-A dentine
substitute-on collagen type i synthesis in pulp fibroblasts in
vitro. PLoS One. 11:e1676332016. View Article : Google Scholar
|
|
235
|
Begue-Kirn C, Smith AJ, Ruch JV, Wozney
JM, Purchio A, Hartmann D and Lesot H: Effects of dentin proteins,
transforming growth factor beta 1 (TGF beta 1) and bone
morphogenetic protein 2 (BMP2) on the differentiation of
odontoblast in vitro. Int J Dev Biol. 36:491–503. 1992.PubMed/NCBI
|
|
236
|
Strong DD, Beachler AL, Wergedal JE and
Linkhart TA: Insulinlike growth factor II and transforming growth
factor beta regulate collagen expression in human osteoblastlike
cells in vitro. J Bone Miner Res. 6:15–23. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Helder MN, Bronckers AL and Woltgens JH:
Dissimilar expression patterns for the extracellular matrix
proteins osteopontin (OPN) and collagen type I in dental tissues
and alveolar bone of the neonatal rat. Matrix. 13:415–425. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Mathieu S, Jeanneau C, Sheibat-Othman N,
Kalaji N, Fessi H and About I: Usefulness of controlled release of
growth factors in investigating the early events of dentin-pulp
regeneration. J Endod. 39:228–235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Shimabukuro Y, Ueda M, Ozasa M, Anzai J,
Takedachi M, Yanagita M, Ito M, Hashikawa T, Yamada S and Murakami
S: Fibroblast growth factor-2 regulates the cell function of human
dental pulp cells. J Endod. 35:1529–1535. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Giraud T, Jeanneau C, Bergmann M, Laurent
P and About I: Tricalcium silicate capping materials modulate pulp
healing and inflammatory activity in vitro. J Endod. 44:1686–1691.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Guo RF and Ward PA: Role of C5a in
inflammatory responses. Annu Rev Immunol. 23:821–852. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Chmilewsky F, Jeanneau C, Dejou J and
About I: Sources of dentin-pulp regeneration signals and their
modulation by the local microenvironment. J Endod. 40(Suppl 4):
S19–S25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Giraud T, Rufas P, Chmilewsky F, Rombouts
C, Dejou J, Jeanneau C and About I: Complement activation by pulp
capping materials plays a significant role in both inflammatory and
pulp stem cells' recruitment. J Endod. 43:1104–1110. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
244
|
Karimjee CK, Koka S, Rallis DM and Gound
TG: Cellular toxicity of mineral trioxide aggregate mixed with an
alternative delivery vehicle. Oral Surg Oral Med Oral Pathol Oral
Radiol Endod. 102:e115–e120. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Maeda H, Nakano T, Tomokiyo A, Fujii S,
Wada N, Monnouchi S, Hori K and Akamine A: Mineral trioxide
aggregate induces bone morphogenetic protein-2 expression and
calcification in human periodontal ligament cells. J Endod.
36:647–652. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Kasaj A, Willershausen B, Reichert C,
Rohrig B, Smeets R and Schmidt M: Ability of nanocrystalline
hydroxyapatite paste to promote human periodontal ligament cell
proliferation. J Oral Sci. 50:279–285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Wang MC, Yeh LY, Shih WY, Li WC, Chang KW
and Lin SC: Portland cement induces human periodontal ligament
cells to differentiate by upregulating miR-146a. J Formos Med
Assoc. 117:308–315. 2018. View Article : Google Scholar
|
|
248
|
Luo T, Liu J, Sun Y, Shen Y and Zou L:
Cytocompatibility of Biodentine and iRoot FS with human periodontal
ligament cells: An in vitro study. Int Endod J. 51:779–788. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
249
|
Chang SW, Lee SY, Kang SK, Kum KY and Kim
EC: In vitro biocompatibility, inflammatory response, and
osteogenic potential of 4 root canal sealers: Sealapex, Sankin
apatite root sealer, MTA Fillapex, and iRoot SP root canal sealer.
J Endod. 40:1642–1648. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
250
|
Willershausen I, Wolf T, Kasaj A, Weyer V,
Willershausen B and Marroquin BB: Influence of a bioceramic root
end material and mineral trioxide aggregates on fibroblasts and
osteoblasts. Arch Oral Biol. 58:1232–1237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Gupta SK, Saxena P, Pant VA and Pant AB:
Adhesion and biologic behavior of human periodontal fibroblast
cells to resin ionomer Geristore: A comparative analysis. Dent
Traumatol. 29:389–393. 2013. View Article : Google Scholar
|
|
252
|
Balto HA: Attachment and morphological
behavior of human periodontal ligament fibroblasts to mineral
trioxide aggregate: A scanning electron microscope study. J Endod.
30:25–29. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
253
|
Bonson S, Jeansonne BG and Lallier TE:
Root-end filling materials alter fibroblast differentiation. J Dent
Res. 83:408–413. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
254
|
Gorduysus M, Avcu N, Gorduysus O, Pekel A,
Baran Y, Avcu F and Ural AU: Cytotoxic effects of four different
endodontic materials in human periodontal ligament fibroblasts. J
Endod. 33:1450–1454. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
255
|
Badr AE: Marginal adaptation and
cytotoxicity of bone cement compared with amalgam and mineral
trioxide aggregate as root-end filling materials. J Endod.
36:1056–1060. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
256
|
Yoshino P, Nishiyama CK, Modena KC, Santos
CF and Sipert CR: In vitro cytotoxicity of white MTA, MTA
Fillapex(R) and Portland cement on human periodontal
ligament fibroblasts. Braz Dent J. 24:111–116. 2013. View Article : Google Scholar
|
|
257
|
Keiser K, Johnson CC and Tipton DA:
Cytotoxicity of mineral trioxide aggregate using human periodontal
ligament fibroblasts. J Endod. 26:288–291. 2000. View Article : Google Scholar
|
|
258
|
Al-Haj Ali SN, Al-Jundi SH and Ditto DJ:
In vitro toxicity of formocresol, ferric sulphate, and grey MTA on
human periodontal ligament fibroblasts. Eur Arch Paediatr Dent.
16:51–55. 2015. View Article : Google Scholar
|
|
259
|
Al-Haj AS: In vitro toxicity of propolis
in comparison with other primary teeth pulpotomy agents on human
fibroblasts. J Investig Clin Dent. 7:308–313. 2016. View Article : Google Scholar
|
|
260
|
Samyuktha V, Ravikumar P, Nagesh B,
Ranganathan K, Jayaprakash T and Sayesh V: Cytotoxicity evaluation
of root repair materials in human-cultured periodontal ligament
fibroblasts. J Conserv Dent. 17:467–470. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
261
|
Kucukkaya S, Gorduysus MO, Zeybek ND and
Muftuoglu SF: In vitro cytotoxicity of calcium silicate-based
endodontic cement as root-end filling materials. Scientifica
(Cairo). 2016:92039322016.
|
|
262
|
Jang YE, Lee BN, Koh JT, Park YJ, Joo NE,
Chang HS, Hwang IN, Oh WM and Hwang YC: Cytotoxicity and physical
properties of tricalcium silicate-based endodontic materials.
Restor Dent Endod. 39:89–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
263
|
Akbulut MB, Arpaci PU and Eldeniz AU:
Effects of four novel root-end filling materials on the viability
of periodontal ligament fibroblasts. Restor Dent Endod. 43:e242018.
View Article : Google Scholar : PubMed/NCBI
|
|
264
|
Camilleri J, Sorrentino F and Damidot D:
Investigation of the hydration and bioactivity of radiopacified
tricalcium silicate cement, Biodentine and MTA Angelus. Dent Mater.
29:580–593. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
265
|
Kim JR, Nosrat A and Fouad AF: Interfacial
characteristics of Biodentine and MTA with dentine in simulated
body fluid. J Dent. 43:241–247. 2015. View Article : Google Scholar
|
|
266
|
Gomes Cornélio AL, Salles LP, Campos da
Paz M, Cirelli JA, Guerreiro-Tanomaru JM and Tanomaru Filho M:
Cytotoxicity of Portland cement with different radiopacifying
agents: A cell death study. J Endod. 37:203–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
267
|
Akbulut MB, Uyar Arpaci P and Unverdi
Eldeniz A: Effects of novel root repair materials on attachment and
morphological behaviour of periodontal ligament fibroblasts:
Scanning electron microscopy observation. Microsc Res Tech.
79:1214–1221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Escobar-Garcia DM, Aguirre-Lopez E,
Mendez-Gonzalez V and Pozos-Guillen A: Cytotoxicity and initial
biocompatibility of endodontic biomaterials (MTA and Biodentine™)
used as root-end filling materials. Biomed Res Int.
2016:79269612016. View Article : Google Scholar
|
|
269
|
Futami T, Fujii N, Ohnishi H, Taguchi N,
Kusakari H, Ohshima H and Maeda T: Tissue response to titanium
implants in the rat maxilla: Ultrastructural and histochemical
observations of the bone-titanium interface. J Periodontol.
71:287–298. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
270
|
Kou PM and Babensee JE: Macrophage and
dendritic cell phenotypic diversity in the context of biomaterials.
J Biomed Mater Res A. 96:239–260. 2011. View Article : Google Scholar
|
|
271
|
Gratchev A, Guillot P, Hakiy N, Politz O,
Orfanos CE, Schledzewski K and Goerdt S: Alternatively activated
macrophages differentially express fibronectin and its splice
variants and the extracellular matrix protein betaIG-H3. Scand J
Immunol. 53:386–392. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Brackett MG, Lewis JB, Messer RL, Lei L,
Lockwood PE and Wataha JC: Dysregulation of monocytic cytokine
secretion by endodontic sealers. J Biomed Mater Res B Appl
Biomater. 97:49–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Kabashima H, Nagata K, Maeda K and Iijima
T: Involvement of substance P, mast cells, TNF-alpha and ICAM-1 in
the infiltration of inflammatory cells in human periapical
granulomas. J Oral Pathol Med. 31:175–180. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
274
|
Ribeiro RA, Souza-Filho MV, Souza MH,
Oliveira SH, Costa CH, Cunha FQ and Ferreira HS: Role of resident
mast cells and macrophages in the neutrophil migration induced by
LTB4, fMLP and C5a des arg. Int Arch Allergy Immunol. 112:27–35.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
275
|
Gomes AC, Gomes-Filho JE and Oliveira SH:
Mineral trioxide aggregate stimulates macrophages and mast cells to
release neutrophil chemotactic factors: Role of IL-1beta, MIP-2 and
LTB(4). Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
109:e135–e142. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
276
|
Baggiolini M: Chemokines and leukocyte
traffic. Nature. 392:565–568. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
277
|
Gomes AC, Filho JE and de Oliveira SH:
MTA-induced neutrophil recruitment: A mechanism dependent on
IL-1beta, MIP-2, and LTB4. Oral Surg Oral Med Oral Pathol Oral
Radiol Endod. 106:450–456. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
278
|
Cavalcanti BN, Rode Sde M, Franca CM and
Marques MM: Pulp capping materials exert an effect on the secretion
of IL-1β and IL-8 by migrating human neutrophils. Braz Oral Res.
25:13–18. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
279
|
Chang F, Kim JM, Choi Y and Park K: MTA
promotes chemotaxis and chemokinesis of immune cells through
distinct calcium-sensing receptor signaling pathways. Biomaterials.
150:14–24. 2018. View Article : Google Scholar
|
|
280
|
Martinez FO and Gordon S: The M1 and M2
paradigm of macrophage activation: Time for reassessment. F000Prime
Rep. 6:132014.
|
|
281
|
Murray PJ, Allen JE, Biswas SK, Fisher EA,
Gilroy DW, Goerdt S, Gordon S, Hamilton JA, Ivashkiv LB, Lawrence
T, et al: Macrophage activation and polarization: Nomenclature and
experimental guidelines. Immunity. 41:14–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Zhu X, Yuan Z, Yan P, Li Y, Jiang H and
Huang S: Effect of iRoot SP and mineral trioxide aggregate (MTA) on
the viability and polarization of macrophages. Arch Oral Biol.
80:27–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
283
|
Braga JM, Oliveira RR, Martins RC and
Ribeiro Sobrinho AP: The effects of a mineral trioxide
aggregate-based sealer on the production of reactive oxygen
species, nitrogen species and cytokines by two macrophage subtypes.
Int Endod J. 47:909–919. 2014. View Article : Google Scholar
|
|
284
|
Rezende TM, Vieira LQ, Cardoso FP,
Oliveira RR, de Oliveira Mendes ST, Jorge ML and Ribeiro Sobrinho
AP: The effect of mineral trioxide aggregate on phagocytic activity
and production of reactive oxygen, nitrogen species and arginase
activity by M1 and M2 macrophages. Int Endod J. 40:603–611. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
285
|
Yuan Z, Zhu X, Li Y, Yan P and Jiang H:
Influence of iRoot SP and mineral trioxide aggregate on the
activation and polarization of macrophages induced by
lipopolysaccharide. BMC Oral Health. 18:562018. View Article : Google Scholar : PubMed/NCBI
|
|
286
|
Yeh HW, Chiang CF, Chen PH, Su CC, Wu YC,
Chou L, Huang RY, Liu SY and Shieh YS: Axl involved in mineral
trioxide aggregate induces macrophage polarization. J Endod.
44:1542–1548. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
287
|
da Silva GN, Braz MG, de Camargo EA,
Salvadori DM and Ribeiro DA: Genotoxicity in primary human
peripheral lymphocytes after exposure to regular and white mineral
trioxide aggregate. Oral Surg Oral Med Oral Pathol Oral Radiol
Endod. 102:e50–e54. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
288
|
Barczak K, Palczewska-Komsa M, Nowicka A,
Chlubek D and Buczkowska-Radlinska J: Analysis of the activity and
expression of cyclooxygenases COX1 and COX2 in THP-1 monocytes and
macrophages cultured with Biodentine™ Silicate cement. Int J Mol
Sci. 21:22372020. View Article : Google Scholar
|
|
289
|
Khedmat S, Dehghan S, Hadjati J, Masoumi
F, Nekoofar MH and Dummer PM: In vitro cytotoxicity of four calcium
silicate-based endodontic cements on human monocytes, a
colorimetric MTT assay. Restor Dent Endod. 39:149–154. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
290
|
Chiang YC, Chang HH, Wong CC, Wang YP,
Wang YL, Huang WH and Lin CP: Nanocrystalline calcium
sulfate/hydroxyapatite biphasic compound as a TGF-β1/VEGF reservoir
for vital pulp therapy. Dent Mater. 32:1197–1208. 2016. View Article : Google Scholar : PubMed/NCBI
|